Patent application title: Method for Linking Sequences of Interest
Inventors:
Martin B. Oleksiewicz (Vienna, AT)
Lars S. Nielsen (Nivaa, DK)
Peter S. Andersen (Vanloese, DK)
Margit H. Hansen (Copenhagen, DK)
Assignees:
SYMPHOGEN A/S
IPC8 Class: AA61K39395FI
USPC Class:
4241331
Class name: Drug, bio-affecting and body treating compositions immunoglobulin, antiserum, antibody, or antibody fragment, except conjugate or complex of the same with nonimmunoglobulin material structurally-modified antibody, immunoglobulin, or fragment thereof (e.g., chimeric, humanized, cdr-grafted, mutated, etc.)
Publication date: 2010-12-09
Patent application number: 20100310558
Claims:
1. A library of cognate pairs consisting of linked variable region
encoding sequences, where each individual cognate pair is inserted into a
vector capable of expressing said cognate pair.
2. The library according to claim 1, wherein said cognate pairs of variable region encoding sequences are obtained by the method comprising:a) providing a lymphocyte-containing cell fraction from a donor;b) optionally enriching for a particular lymphocyte population from said cell fraction;c) obtaining a population of isolated single cells, comprising distributing cells from said cell fraction individually into a plurality of vessels; andd) amplifying and effecting linkage of the variable region encoding sequences contained in said population of isolated single cells.
3. The library according to claim 1, wherein an individual member of said cognate pairs comprises an immunoglobulin light chain variable region encoding sequence associated with a heavy chain variable region encoding sequence.
4. The library according to claim 3, wherein said individual member encodes a full-length antibody selected from human immunoglobulin classes IgA1, IgA2, IgD, IgE, IgG1, IgG2, IgG3, IgG4 or IgM.
5. The library according to claim 1, wherein an individual member of said cognate pairs comprises a TcR alpha chain variable region encoding sequence associated with a beta chain variable region encoding sequence or a gamma chain variable region encoding sequence associated with delta chain variable region encoding sequence.
6. The library according claim 5, wherein said individual member encodes a full-length TcR.
7. The library of cognate pairs of linked variable region encoding sequences according to claim 1, wherein each individual member has been selected for its ability to encode for a protein exhibiting a desired binding specificity directed against a particular target.
8. The library of cognate pairs according to claim 3, where the antibodies expressible from said library are capable of reacting with, or binding to Tetanus Toxin.
9. A population of host cells comprising a library according to claim 1.
10. The population of host cells according to claim 9, wherein the cells are mammalian cells.
11. A recombinant polyclonal antibody expressed from host cells according to claim 9.
12. The recombinant polyclonal antibody according to claim 11, wherein the individual antibody members are full-length, and the constant heavy chain region is chosen from the human immunoglobulin classes IgG1, IgG2, IgG3, IgG4, IgM, IgA1, IgA2, IgD, or IgE, and the constant light chain region is chosen from human lambda or kappa chains.
13. A recombinant polyclonal immunoglobulin or fragments thereof capable of reacting with or binding to Tetanus Toxin.
14. The recombinant polyclonal immunoglobulin according to claim 13, wherein the individual members are cognate pairs.
15. The recombinant polyclonal immunoglobulin according to claim 13, comprising at least two individual cognate pairs comprising an immunoglobulin heavy chain variable region encoding sequence associated with a light chain variable region encoding sequence with at least 90% identity to one individual SEQ ID pair, selected from the group consisting of SEQ ID pairs 229:276, 262:309, 240:287, 245:292, 267:314, 246:293, 258:305, 242:289, 231:278, 263:310, 232:279, 237:284, 255:302, 260:307, 251:298, 233:280, 228:275, 244:291, 250:297, 252:299, 264:311, 272:319, 235:282 or 238:285.
16. A recombinant immunoglobulin or fragments thereof capable of reacting with or binding to Tetanus Toxin, wherein the complementarity determining regions (CDR1, CDR2 and CDR3) are derived from one or more of the SEQ ID pairs of claim 54.
17. A pharmaceutical composition comprising a recombinant polyclonal immunoglobulin according to claim 11 as active ingredient, combined with at least one pharmaceutically acceptable excipient.
18. A pharmaceutical composition comprising a recombinant polyclonal antibody capable of reacting with or binding to Tetanus Toxin as active ingredient, optionally combined with a pharmaceutically acceptable excipient.
19. A pharmaceutical composition according to claim 17 for use as a medicament.
20. A method of preventing or treating a patient at risk of developing tetanus by administering to a patient in need thereof a composition comprising a recombinant polyclonal antibody capable of reacting with or binding to Tetanus Toxin.
21. A method for producing a library of cognate pairs comprising linked variable region encoding nucleotide sequences, the method comprising:a) providing a lymphocyte-containing cell fraction from a donor;b) distributing cells from said cell fraction individually into a plurality of vessels to obtain a population of isolated single cells;c) amplifying, in a multiplex molecular amplification procedure, nucleotide sequences of interest comprising variable region encoding sequences using templates derived from said isolated single cells; andd) effecting linkage of the amplified nucleotide sequences of interest.
22. The method of claim 21, wherein the cell fraction is enriched for a particular lymphocyte population.
23. The method of claim 21, wherein individual isolated single cells in the population of single cells are expanded to populations of isogenic cells prior to performing amplification and linkage.
24. The method of claim 21, wherein the lymphocyte-containing cell fraction comprises whole blood, bone marrow, mononuclear cells or white blood cells.
25. The method of claim 22, wherein the lymphocyte-containing cell fraction is enriched for B lymphocytes.
26. The method of claim 22, wherein the lymphocyte-containing cell fraction is enriched for plasma cells.
27. The method of claim 22, wherein cells from the lymphocyte-containing cell fraction are enriched for antigen specificity.
28. The method of claim 22, wherein the lymphocyte-containing cell fraction is enriched for T lymphocytes.
29. The method of claim 21, wherein the nucleotide sequences of interest comprise immunoglobulin variable region encoding sequences and the linkage generates cognate pairs of a light chain variable region encoding sequence associated with a heavy chain variable region encoding sequence.
30. The method of claim 21, wherein the multiplex molecular amplification procedure is a multiplex RT-PCR amplification.
31. The method of claim 30, wherein the multiplex RT-PCR amplification is a two step process comprising a separate reverse transcription (RT) step prior to the multiplex PCR amplification.
32. The method of claim 31, wherein said linkage of the nucleotide sequences of interest is effected in association with the multiplex PCR amplification, utilizing a multiplex overlap-extension primer mix.
33. The method of claim 30, wherein the multiplex RT-PCR amplification is performed in a single step comprising initially adding all the components necessary to perform both reverse transcription (RT) and multiplex PCR amplification into a single vessel.
34. The method of claim 21, wherein said linkage of the nucleotide sequences of interest is performed in the same vessel as the multiplex molecular amplification.
35. The method of claim 21, further comprising creating a sub-library by selecting a subset of cognate pairs of linked variable region sequences that encode binding proteins with a desired target specificity, thereby generating a library of target-specific cognate pairs of variable region encoding sequences.
36. The method of claim 35, further comprising transferring said library of target-specific cognate pairs of variable region encoding sequences to a mammalian expression vector.
37. The method of claim 36, wherein the nucleotide sequences of interest comprise immunoglobulin variable region encoding sequences and the linkage generates a cognate pair of a light chain variable region encoding sequence associated with a heavy chain variable region encoding sequence.
38. The method of claim 37, wherein the mammalian expression vector encodes at least one constant domain selected from the group consisting of human immunoglobulin classes IgA1, IgA2, IgD, IgE, IgG1, IgG2, IgG3, IgG4, IgM, kappa light chain and lambda light chain.
39. The method of claim 21, further comprising inserting the library of cognate pairs into a vector.
40. The method of claim 39, wherein said vector is selected from the group consisting of cloning vectors, shuttle vectors, display vectors and expression vectors.
41. The method of claim 40, wherein individual members of the library of cognate pairs comprise an immunoglobulin heavy chain variable region encoding sequence associated with a light chain variable region encoding sequence, and said sequences are inserted in-frame into a vector containing sequences encoding at least one immunoglobulin constant domain or a fragment thereof.
42. The method of claim 41, wherein said vector is a mammalian expression vector.
43. The method of claim 39, further comprising the steps:a) introducing a vector comprising linked variable region encoding nucleotide sequences into a host cell;b) cultivating said host cell under conditions suitable for expression of the linked variable region sequences; andc) obtaining the protein product expressed from the vector inserted into said host cell.
44. The method of claim 43, wherein said protein product is a monoclonal antibody.
45. A library of cognate pairs of linked variable region encoding nucleotide sequences obtained by the method of claim 21.
46. The library of claim 45, wherein individual members of the library comprise a cognate pair of linked immunoglobulin variable region encoding sequences, each cognate pair comprising a light chain variable region encoding sequence associated with a heavy chain variable region encoding sequence.
47. The library of claim 46, wherein individual members of the library encode a full-length antibody selected from the group consisting of human immunoglobulin classes IgA1, IgA2, IgD, IgE, IgG1, IgG2, IgG3, IgG4 and IgM.
Description:
CROSS-REFERENCE TO RELATED APPLICATIONS
[0001]This application is a divisional of U.S. application Ser. No. 10/572,431 (allowed), with a 35 U.S.C. §371(c) date of Mar. 20, 2006, which is a National Stage of International Application No. PCT/DK2004/000633, filed Sep. 17, 2004 and published in the English language May 12, 2005, which claims the benefit of U.S. Provisional Application No. 60/504,589, filed Sep. 18, 2003, U.S. Provisional Application No. 60/504,455, filed Sep. 18, 2003, Danish Application No. PA 2003 01867, filed Dec. 17, 2003, and Danish Application No. PA 2004 00782, filed May 15, 2004. The entirety of each of these documents is fully incorporated by reference herein.
REFERENCE TO SEQUENCE LISTING SUBMITTED ELECTRONICALLY
[0002]The content of the electronically submitted sequence listing in ASCII text file (Name sequence_listing.ascii.txt; Size: 191,196 bytes; and Date of Creation: May 26, 2010) filed with the application is incorporated herein by reference in its entirety.
TECHNICAL FIELD OF THE INVENTION
[0003]The present invention relates to a multiplex molecular amplification procedure capable of linking nucleotide sequences of interest in connection with the amplification, in particular polymerase chain reaction (multiplex PCR). The method is particularly advantageous for the generation of cognate pair libraries as well as combinatorial libraries of variable region encoding sequences from immunoglobulins, T cell receptors or B cell receptors.
BACKGROUND OF THE INVENTION
[0004]Antigen binding proteins involved in the immune response are present in mammals as large polyclonal repertoires representing a broad diversity of binding specificities. This diversity is generated by rearrangement of gene sequences encoding variable regions of these binding proteins. Such variable region binding proteins include soluble and membrane-bound forms of the B cell receptor (also known as immunoglobulins or antibodies) and the membrane-bound T cell receptors (TcR). With respect to immunoglobulins, their affinity is enhanced subsequent to recognition of an antigen by a B cell antigen receptor, through a process termed affinity maturation which involves cycles of somatic hypermutation of these variable genes.
[0005]Notably, immunoglobulins or fragments thereof, such as Fab fragments, Fv fragments and single chain Fv (scFv) molecules have been subject to cloning and recombinant expression. However, all other variable region binding proteins can in principle also be cloned and expressed using the same concepts as for antibodies.
[0006]Known approaches for isolating antibodies with a desired binding specificity most often involves generation of hybridomas from immunized hosts followed by screening for specific clones or involves the generation of combinatorial expression libraries in E. coli composed of immunoglobulin variable domains, which are subsequently enriched using techniques such as, for example, phage display.
[0007]The main restriction in the use of the hybridoma technology for making therapeutic antibodies is the absence of a human lymphoma suitable as fusion partners for human B lymphocytes. Heterohybridomas (i.e., fusion of human B cells with mouse lymphomas) are notoriously unstable and thus rarely lead to suitable cell lines for production purposes. Human B cells immortalized through infection with Epstein-Barr virus exhibit similar challenges of instability. The lack of robust cellular methodology for making human antibodies for therapy can be compensated with more recent advantages in molecular biology.
[0008]The use of combinatorial libraries and phage display allows for generation of large repertoires of antibody clones with a potential diversity in excess of 1010. From this repertoire selection for binding to a specific target can be performed thereby generating a sub-library. This sub-library can be used to generate either polyclonal or monoclonal antibodies. The variable region encoding sequences (for example immunoglobulin heavy chain variable region and light chain variable region encoding sequences) which constitute the library can be amplified from lymphocytes, plasma cells, hybridomas or any other immunoglobulin expressing population of cells. Current technologies for generating combinatorial libraries involve separate isolation of the variable region encoding sequences from a population of cells. Thus, the original pairing of for example immunoglobulin heavy chain variable region and light chain variable region encoding sequences will be lost. Rather, in a combinatorial library said sequences are randomly paired and the original combinations of these variable sequences will only occur by chance. Thus, in order to isolate variable region encoding sequences responsible for a desired binding specificity, a considerable amount of screening is necessary. This is typically performed in combination with methods for enrichment of clones exhibiting a desired specificity, such as ribosome display or phage display. Even then, the diversity achieved might not be sufficiently large to isolate variable region encoding sequence pairs giving rise to binding proteins of similar high affinity as those found in the original cells. Further, the enrichment procedures normally used to screen combinatorial libraries introduce a strong bias e.g. for polypeptides of particular low toxicity in E. coli, efficient folding, slow off-rates, or other system dependent parameters, that reduce the diversity of the library even further. In addition, clones derived from such combinatorial libraries will be more prone to produce binding proteins with cross-reactivity against self-antigens because they as pairs, in contrast to original pairs (hereafter called cognate pairs), never have been through in vivo negative selection against self-antigens, such as it is the case for B and T lymphocyte receptors during particular stages of their development. Therefore, the cloning of original pairs of variable region encoding sequences is a desirable approach. Moreover, the frequency of clones exhibiting a desired binding specificity is expected to be considerably higher within a library of cognate pairs, than in a conventional combinatorial library, particularly if the starting material cells are derived from a donor with high frequency of cells encoding specific binding pairs e.g. immune or immunized donors. It follows that the size of a cognate pair library will not need to be as large as a combinatorial library: a cognate pair library size of 104 to 105 clones or even as small as 102 to 103 clones derived from a donor with a relevant ongoing immune response might very well suffice in order to obtain binding proteins representing a broad diversity of desired binding specificities.
[0009]In order to generate cognate pair libraries the linkage of the variable region encoding sequences derived from the same cell is required. At present, two different approaches which can achieve cognate pairing of variable region encoding sequences have been described.
[0010]In-cell PCR is an approach where a population of cells is fixed and permeabilized, followed by in-cell linkage of heavy chain variable region and light chain variable region encoding sequences from immunoglobulins. This linkage can be performed either by overlap-extension RT-PCR (WO 93/03151) or by recombination (Chapal, N. et al. 1997 BioTechniques 23, 518-524). The amplification process as described in these publications is a three or four step process consisting of i) reverse transcription utilizing constant region primers generating immunoglobulin cDNA, ii) PCR amplification of the heavy and light chain variable region encoding sequences utilizing primer sets containing either overlap-extension design or recombination sites, iii) linkage by recombination, if this approach is chosen, iv) nested PCR of the products generating restriction sites for cloning. Since the cells are permeabilized there is a considerable risk that amplification products might leak out of the cells, thereby introducing scrambling of the heavy chain variable region and light chain variable region encoding sequences, resulting in the loss of cognate pairing. Therefore, the procedure includes washing steps after each reaction which makes the process laborious and reduces the efficiency of the reactions.
[0011]More generally, the in-cell PCR is notoriously inefficient, resulting in low sensitivity. Accordingly, the in-cell PCR linkage technique has never found widespread usage, and the original study has in fact never been reliably repeated in a way which can be used to verify that the linkage actually occurs within the cell. This, however, is absolutely crucial to avoid scrambling of the heavy chain variable region and light chain variable region encoding sequences and thereby disrupting the cognate pairs.
[0012]A different in-cell approach is described in WO 01/92291. This approach is based on RNA trans-splicing, and achieves joining of VH and VL encoding mRNA within the cell. This approach requires the presence of a DNA construct driving the trans-splicing within the cells.
[0013]Single-cell PCR is a different approach to achieve cognate pairing of heavy chain variable region and light chain variable region encoding sequences (see, for example, Coronella, J. A. et al. 2000 Nucleic Acids Res. 28, E85; Wang, X., et al. 2000 J. Immunol. Methods 20, 217-225). In these publications a population of immunoglobulin expressing cells are distributed by diluting to a density of one cell per reaction, thereby eliminating scrambling of heavy chain variable region and light chain variable region encoding sequences during the cloning process. Basically, the process described is a three to four step procedure consisting of i) reverse transcription utilizing oligo-dT-, random hexamer- or constant region primers generating cDNA, ii) fractionating the cDNA product into several tubes and performing PCR amplification on the individual variable chain encoding sequences (in separate tubes) with primer sets containing restriction sites for cloning, iii) nested PCR of the products generating restriction sites for cloning (optionally) and iv) linking the heavy chain variable region and light chain variable region encoding sequences from the separate tubes by cloning them into an appropriate vector, which in itself is a multi-step process.
[0014]In humans there are two types of light chains: lambda (λ) and kappa (κ). This means that with the cDNA generated from every single cell at least three separate PCR reactions must be performed followed by analysis and cloning of the appropriate fragments into a single vector to achieve the cognate pairing. Thus, the single-cell PCR approach as described requires a large number of manipulations to generate a library of cognate pairs. Although, a cognate pair library does not need to be as large as a combinatorial library in order to obtain binding proteins representing a broad diversity of binding specificities it would still be a laborious task to generate a library of for example 104 to 105 clones by the described single-cell PCR approach. Further, the large number of manipulations highly increases the risk of contamination and human error.
[0015]In order to obtain high affinity binding proteins corresponding to the affinities normally observed during an immune response, cognate pairing of the variable region sequences in association with their amplification is highly advantageous. To generate a library of large diversity it is necessary to have a cloning technique that can be fitted to a high-throughput format and where the risk of contamination and scrambling is minimal.
[0016]A reduction of the number of cloning steps allowing the generation of combinatorial libraries to be fitted to a high-throughput format, is likewise desired.
DISCLOSURE OF CONTRIBUTION
[0017]The present invention provides an efficient method of linking two or more nucleotide sequences of interest, e.g. variable region encoding sequences, utilizing a multiplex molecular amplification procedure, such as multiplex overlap-extension RT-PCR or multiplex RT-PCR followed by linkage by ligation or recombination. The method is applicable on a single cell, thereby enabling cloning of cognate pairs in a high-throughput format.
DESCRIPTION OF THE FIGURES
[0018]FIG. 1 is a diagram illustrating the different types of overlap-extension tails. The bold line corresponds to a gene-specific part of the primer and the regular line corresponds to the overlapping tail. The vertical bars illustrate complementary regions. The primers facilitate the linkage of two nucleotide sequences of interest. FIG. 1 (I) illustrates two varieties of the type I overlap-extension tails where only the extension tails overlap, either completely or partially; FIG. 1 (II) illustrates the type II overlap-extension tails where some of the 5'-nucleotides of the first primer extension tail are complementary to the gene-specific part of the neighboring primer; FIG. 1 (III) illustrates the type III overlap-extension tails where the entire overlap-extension tails are complementary to the gene-specific region of the neighboring primer.
[0019]FIG. 2 is a diagram showing a schematic overview of a multiplex overlap-extension primer mix applicable in the linkage of immunoglobulin variable region encoding sequences. The cDNA encoding the light chain (LC) and the heavy chain variable region (VH) to be linked are illustrated as tubes with indication of their sense strand 5' and 3' ends as well as the expected size of the amplified product. The multiplex overlap-extension primer sets used to amplify the encoding sequence are illustrated by the arrows. The bended arrow tails with dashed 5' overhangs illustrate the cloning tails. The overlap-extension tails are in bold. The restriction sites present in the tails are named in connection with the tail. The total number of primers in the multiplex overlap-extension primer mix is sixteen, distributed on the outer primers comprising one C.sub.κ and one CH1 primer and the overlap extension primers comprising six VL and eight VH primers. The CH1 primer anneal at the 5' end of the heavy chain constant domain 1. The product resulting from the multiplex overlap-extension RT-PCR is expected to be approximately 1070 bp constituting the entire kappa light chain composed of the constant region, joining gene and variable gene (C.sub.κ+JL+VL) and the heavy chain variable region, composed of the variable gene, diversity segment and joining gene (VH+D+JH). 5' and 3' indicates the direction of the open reading frame. Only a small portion of the CH1 region encoding sequence is amplified by this multiplex overlap-extension primer mix, because the annealing position of the CH1 primer is close to the heavy chain J-region.
[0020]FIG. 3 is a series of diagrams illustrating the different linking direction of the products that can be obtained depending on which primers are equipped with the linkage tail. Solid black illustrates the overlap region. 5' and 3' indicates the direction of the open reading frame. FIG. 3A is a diagram illustrating a head-to-head orientation of the products. FIG. 3B is a diagram illustrating a tail-to-tail orientation. FIG. 3C is a diagram illustrating a head-to-tail orientation with the light chain encoding sequence first. FIG. 3D is a diagram illustrating a head-to-tail orientation with the heavy chain encoding sequence first.
[0021]FIG. 4 is a schematic diagram of the immunoglobulin expression vector pLL113, where the encoding sequences are in a head-to-tail orientation. The vector comprises the following elements: bla=promoter allowing expression of the ampicillin resistance gene. Amp=gene encoding for ampicillin resistance. pUC ori=pUC origin of replication. AdMLP=Adenovirus major late promoter. Human IgG1=Sequence encoding for immunoglobulin isotype G1 heavy chain. hGH pA=Human growth hormone poly A signal sequence. bGH polyA=Bovine Growth Hormone poly A sequence. Human kappa LC=Sequence encoding immunoglobulin kappa light chain. FRT=A Flp recognition target site. Hygromycin=gene encoding hygromycin resistance. SV40 poly A=Simian virus 40 poly A signal sequence.
[0022]FIG. 5A is an electrophoretic gel showing the results from a two-step multiplex overlap-extension RT-PCR followed by a semi-nested PCR. The amplification products are derived from cDNA isolated from single CHO Flp-In pLL113 cells. Lanes 1-12 are sample lanes and the arrows indicate correct nested multiplex-overlap-extension RT-PCR products of 1076 bp; M1 is a 100 bp ladder. W is water used as negative control template. C is a positive cDNA control template derived from cell-line HB-8501. In a separate panel the same lanes W, C and the 100 bp ladder have been depicted with less contrast in order to resolve the individual DNA fragments. FIG. 5B is a sketch of the gel shown in FIG. 5A, illustrating the relevant fragments from the gel.
[0023]FIG. 6 is a series of photographs and a graphical representation of electrophoretic gels showing the results from a single-step multiplex overlap-extension RT-PCR reaction without additional PCR amplification. In each panel, M1 is a 100 bp ladder and M2 is a 500 by ladder. FIG. 6A is an electrophoretic gel showing the amplification products derived from lysate corresponding to 100, 10, 1 or 0 cells. The arrow indicates the overlap-extension product. FIG. 6B is a sketch of the gel in FIG. 6A. FIG. 6C is an electrophoretic gel verifying the presence of overlap-extension product in the 100 and 1 cell lanes in FIG. 6A. FIG. 6D is an electrophoretic gel showing restriction enzyme cleavage with NheI and NcoI, respectively, of the overlap-extension product from the 1 cell lane in FIG. 6C.
[0024]FIG. 7 is an electrophoretic gel showing the results from a single-step multiplex overlap-extension RT-PCR followed by a semi-nested PCR amplification. M1 is a 100 bp ladder and M2 is a 500 bp ladder. Results from multiplex overlap-extension primer mixtures containing either CH1, CH2, CH3, CH4 or CH5 as the outer primer in the multiplex overlap-extension RT-PCR reaction. The reactions were performed on cell lysates corresponding to 100, 10, 1 or 0 cells. The size of the overlap-extension product is indicated with an arrow.
[0025]FIG. 8 is an electrophoretic gel showing the results from a single-step multiplex overlap-extension RT-PCR followed by a semi-nested PCR amplification using enriched human B lymphocytes as template. M1 is a 100 bp ladder. Lane 5 and 6 show bands of the size expected for the overlap-extension product.
[0026]FIG. 9A is a schematic diagram of the mammalian expression vectors (Em465/01P582/Em465/01P581) used for generation of IgG1-lambda expressing cell lines, where the encoding sequences are in a head-to-head orientation. The vectors comprise the following elements: Amp=gene encoding for ampicillin resistance. pUC ori=pUC origin of replication. AdMLP=Adenovirus major late promoter. EFP=Elongation factor promoter. AP leader=alkaline phosphatase leader sequence. VH=heavy chain variable region encoding sequence. IgG1 HC=Sequence encoding for immunoglobulin isotype G1 heavy chain constant region. rBG polyA=Rabbit beta-globin poly A signal sequence. bGH polyA=Bovine Growth Hormone poly A sequence. IgK leader=sequence encoding for murine kappa leader. IgL (1b or 1c)=Sequence encoding immunoglobulin lambda light chain family 1b or 1c. FRT=A Flp recognition target site. Hygromycin=gene encoding hygromycin resistance. SV40 poly A=Simian virus 40poly A signal sequence. FIGS. 9B and 9C are ethidium bromide stained agarose gels loaded with PCR products isolated form cell line CHO Flp-In/Em464/01P581 and CHO Flp-In/Em464/01P582, respectively. Lanes 1 to 4 correspond to total RNA template concentrations of 50 pg, 5 pg, 0.5 pg or 0 pg used for the multiplex overlap-extension RT-PCR reaction. M is a 100 bp ladder (New England Biolabs, New England, USA). The arrows indicate the overlap-extension PCR product.
[0027]FIG. 10 is a flow chart showing the steps applied in order to generate a cognate antibody expression library from which polyclonal or monoclonal antibodies can be expressed.
[0028]FIG. 11 is a schematic diagram of JSK301, an E. coli vector used to generate a library of Fab vectors by inserting the overlap-extension fragments comprising the cognate variable region encoding sequences into the vector at the indicated NotI/XhoI restriction sites. The vector comprises the following elements: Amp and Amp pro=ampicillin resistance gene and its promoter. pUC19 Ori=origin of replication. Human CH1=sequence encoding human immunoglobulin gamma 1 heavy chain domain 1. Stuffer=an irrelevant sequence insert which is cut out upon insertion of the overlap-extension fragments. tac P and lac Z=bacterial promoters which can be excised at the NheI and AscI restriction sites.
[0029]FIG. 12 is a schematic diagram illustrating the generation of a library of cognate Fab expression vectors. Step 1 illustrates the insertion of cognate pairs of variable region encoding sequences (VH1-VL1 to VHx-VLx) into E. coli vector JSK301 by XhoI-NotI digestion. Step II illustrates the insertion of a bacterial promoter and leader cassette (pelB leader-P tac-promoter driving the expression of VHx and P lac promoter-pelB leader driving the expression of VLx) by AscI-NheI digestion.
[0030]FIG. 13 illustrate the linkage of α-, β- and γ-subunits constituting a G-protein, utilizing single-step multiplex overlap-extension RT-PCR followed by an additional PCR amplification. The sizes of the individual coding regions are given as well as the size of the linked product. Restriction sites introduced by the primer tails during amplification are indicated for the final product.
[0031]FIG. 14 shows dot plots of an analytic FACS staining of (A) PBMC purified from donor blood; (B) the magnetically sorted non-labelled CD19 negative cell fraction and (C) magnetically sorted CD19+ cells fraction. A scatter plot, a CD19/CD38 plot, and a CD38/CD45 plot is shown for each fraction.
[0032]FIG. 15 shows the CD19+ fraction from FIG. 9 C, which had been stored in liquid nitrogen, thawed and stained with anti-CD19, anti-CD38 and anti-CD45. Dot plots corresponding to FIG. 9C are shown.
[0033]FIG. 16 shows gates used for sorting on the CD19+ cell fraction. A scatter gate and a fluorescence gate based on CD38 and CD45 were used for isolating the CD38high (CD38hi), CD45intermediate (CD45 in) cells.
[0034]FIG. 17 is an electrophoretic gel showing the successful multiplex overlap-extension RT-PCR reaction on donor TT03 (row A, well 1-12 from eight 96 well plates). The samples have been applied to the agarose gel in two rows (A and B) with 48 samples in each. The expected size of the overlap-extension fragment was approximately 1070 bp. The putative overlap-extension fragments are marked with arrows.
[0035]FIG. 18 shows ELISA analysis of periplasmic extracts from plate G060. The ELISA plate was coated with goat(gt)-anti-human Kappa, and captured Fab fragments were detected with a HRP-conjugated gt-anti-human Fab-specific antibody.
[0036]FIG. 19 shows ELISA analysis of periplasmic extracts from plate G060. The ELISA plate was coated with 10 μg/ml Ovalbumin (Sigma A-5503), and captured Fab fragments were detected with a HRP-conjugated gt-anti-human Fab-specific antibody.
[0037]FIG. 20 shows ELISA analysis of periplasmic extracts from plate G060. The ELISA plate was coated with Tetanus Toxoid, and captured Fab fragments were detected with a HRP-conjugated gt-anti-human Fab-specific antibody.
[0038]FIG. 21 shows a one step competition ELISA analysis of periplasmic extracts from plate G060. The ELISA plate was coated with Tetanus Toxoid (TT), and soluble TT was added to each well at 10-7 M in order to compete for the binding of Fab fragments from the bacterial supernatants, to immobilized TT. Captured Fab fragments were detected with a HRP-conjugated gt-anti-human Fab-specific antibody.
[0039]FIG. 22 shows alignment of variable heavy chain protein sequences from TT antigen-binding clones from plate G060. The degree of sequence homology was represented by different shadings; 100%, 80% and 60% were depicted with black, grey and light grey, respectively. CDR1 is located at alignment position 34 to 41. CDR2 is located at alignment position 55 to 73. CDR3 is located at alignment position 107 to 127. Premature stop codons were denoted by an asterisk. The alignment is divided into 8 separate Figures (a-h) distributed in two rows from left to right with FIG. 22a to d in the top row and FIG. 22e to h in the bottom row.
[0040]FIG. 23 shows alignment of variable light chain protein sequence from TT antigen-binding clones from plate G060. The degree of sequence homology was represented by different shadings; 100%, 80% and 60% were depicted with black, grey and light grey, respectively. CDR1 is located at alignment position 26 to 42. CDR2 is located at alignment position 58 to 64. CDR3 is located at alignment position 97 to 106. Premature stop codons were denoted by an asterisk. The alignment is divided into 8 separate Figures (a-h) distributed in two rows from left to right with FIG. 23a to d in the top row and FIG. 23e to h in the bottom row.
[0041]FIG. 24 shows a competition ELISA assay for determination of apparent affinities of selected clones from plate G060. Soluble TT dilutions in concentrations from 100 nM to 25 pM (four-fold dilutions) were added to the Fab fragments, thereby competing out the binding of the Fab-fragments to immobilized TT. The extents of the reactions are given as the ratio of the observed binding at a given soluble TT concentration to the binding found when no soluble TT was added to the reactions.
[0042]FIG. 25 shows a double phylogenetic dot matrix plot showing the intra- and intergenetic relationship between antibody heavy and light chain variable domain sequences. Phylogenetic threes of VH and VL sequences are paired in a dot matrix in order to indicate actual pairing of particular V genes. A) TT binding clones obtained from a combinatorial library using phage display. B) TT binding clones obtained from a library of cognate pairs using the present invention.
DESCRIPTION OF THE INVENTION
[0043]The present invention sets out to provide an amplification and linkage process of two or more non-contiguous nucleotide sequences of interest which enables the cloning of such sequences to be fitted to a high-throughput format. This is basically achieved by reducing the number of steps necessary to amplify and link sequences to be cloned.
[0044]One aspect of the invention is a method of linking a plurality of non-contiguous nucleotide sequences comprising amplifying, in a multiplex molecular amplification procedure, nucleotide sequences of interest using a template derived from an isolated single cell, a population of isogenic cells or a population of genetically diverse cells and effecting a subsequent linkage of the amplified sequences. If the template is an isolated single cell or a population of isogenic cells, the linkage results in a nucleic acid segment comprising nucleotide sequences of interest associated with each other in a cognate manner. If the template is a population of genetically diverse cells, the linkage results in a library of segments where each segment comprise nucleic acid sequences of interest associated in a is random manner, this is also termed a combinatorial library.
[0045]In one embodiment of the present invention this multiplex molecular amplification procedure is a multiplex PCR amplification, preferably preceded by a reverse transcription step. In a preferred embodiment reverse transcription, amplification and linkage are performed in a single step, using multiplex overlap-extension RT-PCR or alternatively in two steps using multiplex RT-PCR followed by linkage by ligation or recombination.
[0046]A further embodiment of the present invention relates to the generation of libraries of cognate pairs comprising linked variable region encoding sequences, in particular heavy chain variable region and light chain encoding sequences or T cell receptor (TcR) alpha chain encoding sequences and beta chain encoding sequences. The process involves obtaining a lymphocyte-containing cell fraction from at least one suitable donor and optionally enriching for a particular lymphocyte population from this fraction, for example B lymphocytes or T lymphocytes, depending on whether variable region encoding sequences from immunoglobulins or TcRs are desired. The lymphocyte-containing cell fraction or the enriched cell fraction is distributed into an array of vessels, obtaining one cell in each vessel. The array of single cells are subjected to a reverse transcription (RT) step or an alternative cDNA generating procedure, using the nucleic acids derived from the population of single cells as template. The RT step is followed by multiplex molecular amplification procedure and linkage of pairs of variable region encoding sequences generated from each cell according to one of the methods of the present invention.
[0047]The cloning techniques disclosed in the present invention omit laborious and inefficient cloning approaches and additionally reduces the risk of contamination and loss of diversity during multiple cloning steps.
[0048]Another aspect of the invention relates to libraries of cognate pairs produced by the multiplex molecular amplification and linkage process. The initial library of cognate pairs (parent library) generated by the method of the present invention can be subjected to screening, thereby generating a sub-library of cognate pairs encoding target-specific binding to protein variable domains or full-length binding proteins.
[0049]In a further embodiment of the invention, the libraries and sub-libraries of the present invention can be used in the expression of recombinant monoclonal or polyclonal proteins, where the original binding affinities and specificities present in the donor are preserved.
DEFINITIONS
[0050]The term "cognate pair" describes an original pair of non-contiguous nucleic acids of interest that are contained within or derived from a single cell. In preferred embodiments, a cognate pair comprises two variable region encoding sequences which together encode for a binding protein variable domain and which gene sequences are derived from the same cell. Thus, when expressed either as a complete binding protein or as a stable fragment thereof, they preserve the binding affinity and specificity of the binding protein originally expressed from this cell. A cognate pair can for example be comprised of an antibody variable heavy chain encoding sequence associated with a variable light chain encoding sequence from the same cell, or a T cell receptor a chain encoding sequence associated with a β chain encoding sequence from the same cell. A library of cognate pairs is a collection of such cognate pairs.
[0051]The term "hot-start polymerase" describes polymerases that are inactive or have very low activity at temperatures used for reverse transcription. Such polymerases need to be activated by high temperatures (90 to 95° C.) to become functional. This is for example an advantage in single-step RT-PCR procedures, since this prohibits interference of the polymerase with the reverse transcriptase reaction.
[0052]The term "isogenic population of cells" describes a population of genetically identical cells. In particular, an isogenic population of cells derived by clonal expansion of an isolated single cell is of interest in the present invention.
[0053]The term "isolated single cell" describes a cell that has been physically separated from a population of cells corresponding to "a single cell in a single vessel". When distributing a population of cells individually among a plurality of vessels, a population of isolated single cells is obtained. As specified in the section entitled "Template sources" the proportion of vessels with a single cell is not necessarily a 100% in order to call it a population of single cells.
[0054]Terms derived from "link" or "linkage" in relation to amplification describes the association of the amplified nucleic acid sequences encoding the nucleic acid sequences of interest into a single segment. In relation to cognate pairs a segment comprises nucleic acid sequences encoding a variable domain, e.g. an antibody heavy chain variable region associated with an antibody light chain variable region encoding sequence, derived from the same cell. The linkage can either be achieved simultaneously with the amplification or as an immediate step following the amplification. There are no requirements to the form or functionality of the segment, it may be linear, circular, single stranded or double stranded. Nor is the linkage necessarily permanent, one of the nucleic acid sequences of interest may be isolated from the segment if desired, one of the variable region encoding sequence may for example be isolated from a cognate pair segment. However, as long as the original variable regions constituting the cognate pair are not scrambled with other variable regions, they are still considered a cognate pair, although not linked together into a single segment. The linkage is preferably a nucleotide phosphodiester linkage. However, linkage can also be obtained by different chemical cross linking procedures.
[0055]The term "multiplex molecular amplification" describes the simultaneous amplification of two or more target sequences in the same reaction. Suitable amplification methods include the polymerase chain reaction (PCR) (U.S. Pat. No. 4,683,202), ligase chain reaction (LCR), (Wu and Wallace, 1989, Genomics 4, 560-9), strand displacement amplification (SDA) technique (Walker et al., 1992, Nucl. Acids Res. 20, 1691-6), self-sustained sequence replication (Guatelli et al., 1990, Proc. Nat. Acad. Sci. U.S.A., 87, 1874-8) and nucleic acid based sequence amplification (NASBA) (Compton J., 1991, Nature 350, 91-2). The latter two amplification methods involve isothermal reactions based on isothermal transcription, which produce both single stranded RNA (ssRNA) and double stranded DNA (dsDNA).
[0056]The term "multiplex PCR" describes a variant of PCR in which two or more target sequences are amplified simultaneously, by including more than one set of primers in the same reaction, e.g. one primer set adapted for amplification of the heavy chain variable region and one primer set adapted for amplification of the kappa chain variable region in the same PCR reaction. Additionally a primer set adapted for amplification of the lambda chain variable region may be combined with these primer sets.
[0057]The term "multiplex RT-PCR" describes a multiplex PCR reaction, which is preceded by a reverse transcription (RT) step. The multiplex RT-PCR, can either be performed as a two-step process with a separate RT step prior to the multiplex PCR, or as a single-step process where all components for both RT and multiplex PCR are combined in a single tube.
[0058]The terms "multiplex overlap-extension PCR" and "multiplex overlap-extension RT-PCR" implies that the multiplex PCR or multiplex RT-PCR is performed utilizing a multiplex overlap-extension primer mix to amplify the target sequences, thereby enabling simultaneous amplification and linkage of the target sequences.
[0059]The term "a plurality of vessels" describes any object (or collection of objects) which enables the physical separation of a single cell from a population of cells. This may be tubes, multiwell plates (e.g. 96-well, 384-well, microtitter plates or other multiwell plates), arrays, microarrays, microchips, gels, or a gel matrix. Preferably the object is applicable for PCR amplification.
[0060]The term "polyclonal protein" or "polyclonality" as used herein, refers to a protein composition comprising different, but homologous protein molecules, preferably selected from the immunoglobulin superfamily. Thus, each protein molecule is homologous to the other molecules of the composition, but also contains one or more stretches of variable polypeptide sequence, which is/are characterized by differences in the amino acid sequence between the individual members of the polyclonal protein. Known examples of such polyclonal proteins include antibody or immunoglobulin molecules, T cell receptors and B cell receptors. A polyclonal protein may consist of a defined subset of protein molecules, which has been defined by a common feature such as the shared binding activity towards a desired target, e.g. a polyclonal antibody exhibiting binding specificity towards a desired target antigen.
[0061]The term "a population of genetically diverse cells" as used herein, refers to a cell population where the individual cells in the population differ among each other on the genomic level. Such a population of genetically diverse cell is for example a population of cells derived from a donor, or a fraction of such cells, e.g. a B lymphocyte or a T lymphocyte containing cell fraction.
[0062]The term "primer set" is used interchangeably with the term "primer pair" and describes two or more primers which together are capable of priming the amplification of a nucleotide sequence of interest (i.e., one member of a cognate pair). A primer set of the present invention might be designed to prime a family of nucleotide sequences containing variable region encoding sequences. Examples of different families are antibody kappa light chains, lambda light chains, heavy chain variable regions, and α, β, γ, or δ T cell receptor variable regions. A primer set for the amplification of a family of nucleotide sequences containing variable region encoding sequences often constitutes a plurality of primers where several primers can be degenerate primers.
[0063]The term "sequence identity" is expressed as a percentage which indicates the degree of identity between to nucleic acid sequences over the length of the shortest of the two sequences. It can be calculated as (Nref-Ndif)×100/Nref, wherein Nref is the number of residues in the shorter of the sequences, and wherein Ndif is the total number of non-identical residues in an Nref long optimally aligned match between the two sequences. Hence, the DNA sequence AGTCAGTC will have a sequence identity of 75% with the sequence TAATCAATCGG (Ndif=2 and Nref=8) (underlining shows the optimal alignment, and bold indicates the two non-identical residues out of 8).
[0064]The terms "randomly" or "random" with respect to linkage refers to linkage of nucleotide sequences which are not derived from the same cell but are linked transversely among a population of genetically diverse cells. If the nucleotide sequences of interest are variable region encoding sequences, this will result in a combinatorial library of linked sequences. If, on the other hand, the nucleotide sequences of interest encode for a non-diverse heteromeric protein the randomly linked sequences will appear similar to sequences linked from a single cell.
[0065]The term "template derived from an isolated single cell," with regard to reverse transcription, relates to the nucleic acids within such an isolated cell. The nucleic acids can for example be in the form of RNA, mRNA, DNA or genomic DNA. The nucleic acids can either be isolated from the cell or still be with the remaining contents of the cell, where the cell is in an intact form or a lysed form.
The Amplification and Linkage Process
[0066]One feature of the present invention reduces the number of tubes necessary to amplify the nucleotide sequences of interest, utilizing a variant of PCR in which two or more target sequences are amplified simultaneously in the same tube, by including more than one set of primers, for example all the primers necessary to amplify variable region encoding sequences, in the same reaction. Generally this approach is known as multiplex polymerase chain reaction (multiplex PCR).
[0067]Multiplex PCR amplification and multiplex PCR preceded by reverse transcription (multiplex RT-PCR) are well known techniques within the diagnostic field, for example in the analysis of mutations, deletions and polymorphisms of DNA, for quantitative assays of mRNA levels and for identification of viruses, bacteria and parasites (reviewed in Markoulatos, P. et al. 2002. J. Clin. Lab. Anal. 16, 47-51). However, there are only very few examples where immunoglobulin light chain variable region encoding sequences has been amplified in the same vessel as immunoglobulin heavy chain variable region encoding sequences using a multiplex primer mix composed of more than four primers constituting a Vκ (and/or V.sub.λ, primer set together with a VH primer set (Chapal, N. et al. 1997. BioTechniques 23, 518-524; Liu, A. H. et al. 1992. Proc. Natl. Acad. Sci. U.S.A. 89, 7610-7614; Embleton, M. J. et al. 1992. Nucleic Acids Res. 20, 3831-3837). The reason for this, might be that primer sets adapted for the amplification of sequences encoding the variable domains of antigen binding proteins generally are constituted of a plurality of degenerate primers in order to capture the diversity of these variable region encoding sequences. Thus, the complexity of the PCR reaction is highly increased when performing multiplex PCR amplification on variable region encoding sequences.
[0068]A further feature of the present invention is that two or more target sequences amplified by multiplex PCR are linked in close proximity to the amplification process. In particular cognate pairs of variable region encoding sequences are linked by this process.
[0069]One embodiment of the present invention exploits that a multiplex primer mix can be designed to work in an overlap-extension PCR procedure, resulting in a simultaneous amplification and linkage of nucleotide sequences of interest. This multiplex overlap-extension PCR technique serves to reduce the number of reactions necessary to isolate and link nucleotide sequences of interest, in particular cognate pairs of linked variable regions.
[0070]Other embodiments of the present invention apply linkage by ligation or by recombination as an alternative to linkage by multiplex overlap-extension PCR. In these procedures, the linkage is not performed simultaneously with the multiplex PCR amplification, but as an immediate step following the amplification. However, linkage can still be performed in the same tube as the multiplex PCR was performed in.
[0071]In order to perform multiplex overlap-extension PCR, the presence of two or more primer sets (a multiplex primer mix), where at least one primer of each set is equipped with an overlap-extension tail is needed. The overlap-extension tails enable the linkage of the products generated by each of the primer sets during amplification. Such a primer mix is called a multiplex overlap-extension primer mix. The multiplex overlap-extension PCR, differ from conventional overlap-extension PCR in that the sequences to be linked are generated simultaneously in the same tube, thereby providing immediate linkage of the target sequences during amplification, without any intermediate purification. Further, conventional overlap-extension PCR requires a separate linking PCR reaction either with an outer primer set or a nested primer set in order to generate the linked product (Horton, R. M. et al. 1989. Gene 77, 61-68). Such an additional amplification step is optional in the multiplex overlap-extension PCR of the present invention.
[0072]A further feature of the present invention is a reverse transcription (RT) step preceding the multiplex PCR or multiplex overlap-extension PCR amplification, utilizing a is template derived from an isolated single cell or a population of isogenic cells.
[0073]A further feature of the present invention is the use of nucleotide sequences derived from an isolated single cell or a population of isogenic cells as template for the multiplex PCR amplification. Preferably, RNA from a single cell is reverse transcribed into cDNA prior to the multiplex PCR. For the amplification of some nucleic acid sequences of interest genomic DNA may be used as an alternative to mRNA. By using isolated single cells or a population of isogenic cells derived by clonal expansion of an isolated single cell as template source, it is possible to avoid scrambling of nucleotide sequences encoding a heteromeric protein of interest, with nucleotide sequences derived from different cells within a population of cells. This is of importance if one wishes to obtain the original composition of the sequences of interest. Especially for the generation of a cognate pair of variable region encoding sequences, is the use of an isolated single cell or a population of isogenic cells as template source an important feature.
[0074]Multiplex overlap-extension PCR is a rarely used technology. WO 99/16904 discloses the linkage of exons from a genomic sequence in a single reaction, thereby generating cDNA without utilizing reverse transcription. The process as described, utilized one primer set (constituted of two primers) per exon to be linked, thereby composing a multiplex overlap-extension primer mix. Each individual primer set was capable of overlapping with the adjacent primer set by complementary overlap-extension tails. The cDNA was generated from a template of genomic DNA by performing an overlap-extension PCR reaction utilizing the multiplex overlap-extension primer mix followed by a nested PCR, which was described as a necessary step.
[0075]The generation of cDNA from the exons of genomic DNA as described in WO 99/16904 is a different field than the cloning of sequences encoding for heteromeric proteins. First of all, heteromeric proteins are generally expressed from different genes, whereas the exon linkage as described in WO 99/16904 relates to the linkage of exons from a single gene. Additionally, the present invention facilitates the generation of libraries of linked nucleic acid sequences of interest, in particular combinatorial libraries and libraries of cognate pairs of variable regions; a completely different situation than linking a series of exons from a single gene, resulting in a single non-variable cDNA. Further, the present invention utilizes nucleic acids derived from single cells, preferably in the foul'. of RNA that does not need to be isolated from the remaining cell contents before it can be utilized as template.
[0076]There are few publications, where multiplex overlap-extension RT-PCR has been described in relation to linkage of variable region encoding sequences.
[0077]The most simple form of multiplex overlap-extension RT-PCR was described for the isolation of a scFv encoding sequence from a hybridoma cell line (Thirion, S. et al. 1996. Eur. J. Cancer Prev. 5, 507-511 and Mullinax, R. L et al. 1992. BioTechniques 12, 864-869). The methods described by Thirion and Mullinax utilized reverse transcription of mRNA with oligo-dT primers on total RNA extracted from the hybridoma cell line followed by a separate linkage step. The linkage step was performed with a total of four primers, constituting two primer pairs for amplification of heavy chain variable region and light chain variable region encoding sequences, respectively. The VL forward primer and the VH or CH reverse primer contained complementary overlap-extension tails, thereby enabling simultaneous amplification and linkage of the heavy chain variable region and light chain variable region encoding sequences. These methods did not use a nested PCR to increase the sensitivity of the linkage method.
[0078]The other example of multiplex overlap-extension RT-PCR in relation to linkage of variable region encoding sequences was described in the previously mentioned WO 93/03151, providing a method of cloning heavy chain variable region and light chain variable region encoding sequences originating from the same cell without having to isolate single cells prior to the cloning. The method described in WO 93/03151 required washing between the RT step and the multiplex overlap-extension PCR step. Further, one of the specific aims brought about by WO 93/03151, was to solve the problem of having to isolate single cells in order to obtain cognate pairs of variable region encoding sequences.
[0079]None of these known multiplex overlap-extension RT-PCR techniques were developed to function on template derived from an isolated single cell. Neither were any of the methods able to pedant), as single-step RT-PCR reactions.
[0080]One embodiment of the present invention encompasses the linkage of a plurality of non-contiguous nucleotide sequences of interest. The method comprises amplifying, in a multiplex PCR or multiplex RT-PCR amplification procedure, nucleotide sequences of interest using a template derived from an isolated single cell or a population of isogenic cells and effecting linkage of the amplified nucleotide sequences of interest. Further, the method comprises an optional step of performing an additional amplification of the linked products.
[0081]A further embodiment of the present invention encompasses a method of producing a library of cognate pairs comprising linked variable region encoding sequences. The method comprises providing a lymphocyte-containing cell fraction from a donor, which is optionally enriched for a particular lymphocyte population from said cell fraction. Further, a population of isolated single cells is obtained by distributing cells from the lymphocyte-containing cell fraction, or the enriched cell fraction, individually among a plurality of vessels. Multiplex molecular amplification (multiplex RT-PCR amplification) of the variable region encoding sequences contained in the population of isolated single cells is performed and linkage of pairs of variable region encoding sequences, wherein an individual pair is derived from a single cell, within the population of isolated single cell, is effected. Further, the technique comprises two optional steps: in the first step the individual isolated single cell in the population of single cells is expanded to a population of isogenic cells prior to performing multiplex RT-PCR amplification. Thereby obtaining a plurality of vessels with a diverse population of isogenic cells (one population of isogenic cells in one vessel). The second optional step encompasses performing an additional amplification of the linked variable region encoding sequences.
[0082]In preferred embodiments of the present invention, an individual member of said library of cognate pairs comprised of an immunoglobulin light chain variable region encoding sequence is associated with an immunoglobulin heavy chain variable region encoding sequence, originating from the same cell or of sequences encoding a T cell receptor binding domain, constituted of an alpha chain variable region associated with a beta chain variable region or a gamma chain variable region associated with a delta chain variable region, where the associated variable regions originate from the same cell.
[0083]The multiplex RT-PCR amplification of the present invention can be performed either as a two-step process, where reverse transcription (RT) is preformed separate from the multiplex PCR amplification (or alternative multiplex molecular amplification), or as a single-step process, where the RT and multiplex PCR amplification steps are performed with the same primers in one tube.
[0084]The reverse transcription (RT) is performed with an enzyme containing reverse transcriptase activity resulting in the generation of cDNA from total RNA, mRNA or target specific RNA from an isolated single cell. Primers which can be utilized for the reverse transcription are for example oligo-dT primers, random hexamers, random decamers, other random primers, or primers that are specific for the nucleotide sequences of interest.
[0085]The two-step multiplex RT-PCR amplification procedure, allows for the cDNA generated in the RT step, to be distributed to more than one vessel permitting for the storage of a template fraction before proceeding with the amplification. Additionally, the distribution of cDNA to more than one tube, allows for the performance of more than one multiplex PCR amplification of nucleic acid derived from the same template. Although, this results in an increased number of separate reactions, it opens for the possibility to decrease the complexity of the multiplex primer mix if this should be desired. This two-step approach can for example be applied to amplify and link heavy chain variable region and kappa light chain variable region encoding sequences in one tube and heavy chain variable region and lambda light chain variable region encoding sequences in a different tube utilizing the same template. A single cell usually only expresses one of the light chains. However, it will often be easier to perform the reactions simultaneously instead of awaiting the result of one of the reactions before performing the other. Further, the amplification of both kappa and lambda serves as an internal negative control, since it would be expected that only kappa or lambda amplify from a single cell.
[0086]In the single-step multiplex RT-PCR procedure, reverse transcription and multiplex PCR amplification is carried out in the same vessel. All the components necessary to perform both the reverse transcription and the multiplex PCR in a single step are initially added into the vessels and the reaction is performed. Generally, there is no need to add additional components once the reaction has been started. The advantage of single-step multiplex RT-PCR amplification is that it reduces the number of steps necessary to generate the linked nucleotide sequences of the present invention even further. This is particular useful when performing multiplex RT-PCR on an array of single cells, where the same reaction needs to be carried out in a plurality of vessels. Single-step multiplex RT-PCR is performed by utilizing the reverse primers present in the multiplex primer mix needed for the multiplex PCR amplification as primers for the reverse transcription as well. Generally, the composition needed for the single-step multiplex RT-PCR comprises a nucleic acid template, an enzyme with reverse transcriptase activity, an enzyme with DNA polymerase activity, deoxynucleoside triphosphate mix (dNTP mix comprising dATP, dCTP, dGTP and dTTP) and a multiplex primer mix. The nucleic acid template is preferably total RNA or mRNA derived from an isolated single cell either in a purified form, as a lysate of the cell or still within the intact cell. Generally, the exact composition of the reaction mixture requires some optimization for each multiplex primer mixture to be used with the present invention. This applies both for the two-step and the single-step multiplex RT-PCR procedures.
[0087]In alternative embodiments of the present invention it may be appropriate to use genomic DNA instead of RNA as template. In such cases the reverse transcription step is omitted, and the remaining steps of the invention are performed as described throughout the application.
[0088]For some single-step multiplex RT-PCR reactions it may be an advantage to add additional components during the reaction. For example, addition of the polymerase following the RT step. Other components could for example be a dNTP mixture or a multiplex primer mix possibly with a different primer composition. This can then be considered as a one-tube multiplex RT-PCR, which generally has the same advantages as the single-step multiplex RT-PCR, since it also limits the number of tubes necessary to obtain the desired linked products.
[0089]The nucleotide sequences of interest, amplified by the multiplex RT-PCR, can be linked to one another by several methods, such as multiplex overlap-extension RT-PCR, ligation or recombination, using different multiplex primer mixes. Preferably the multiplex RT-PCR amplification and linkage process is a single step or a two step process. However, the linkage process may also be performed as a multi step process, using for example a stuffer fragment to link the nucleic acid sequences of interest, either with PCR, ligation or recombination. Such a stuffer fragment may contain cis-elements, promoter elements or a relevant coding sequence or recognition sequence. In a preferred embodiment the linkage process is performed in the same vessel as the multiplex RT-PCR amplification.
[0090]In one embodiment of the present invention the linkage of a plurality of non-contiguous nucleotide sequences of interest is performed in association with the multiplex PCR amplification, utilizing a multiplex overlap-extension primer mix. This results in the combined amplification and linkage of the target sequences. Generally, the composition needed for the multiplex overlap-extension PCR comprises, a nucleic acid template, an enzyme with DNA polymerase activity, deoxynucleoside triphospate mix (dNTP mix comprising dATP, dCTP, dGTP and dTTP) and a multiplex overlap-extension primer mix.
[0091]In a particular embodiment of the present invention, the linkage of a plurality of non-contiguous nucleotide sequences of interest is performed by multiplex overlap-extension RT-PCR using a template derived from an isolated single cell or a population of isogenic cells. Further, the method comprises an optional step of performing an additional molecular amplification of linked products. Preferably, the multiplex overlap-extension RT-PCR is performed as a single-step/one-tube reaction.
[0092]A multiplex overlap-extension primer mix of the present invention comprises at least two primer sets capable of priming the amplification and linkage of at least two variable region encoding sequences, for example, amplification and linkage of sequences from immunoglobulin heavy chain variable region families with kappa or lambda light chain variable region families, or amplification and linkage of sequences from T cell receptor families α, β, γ, or δ.
[0093]In another embodiment of the present invention the plurality of nucleotide sequences of interest, amplified by multiplex RT-PCR, are linked by ligation. To achieve this, the multiplex primer mix used for the multiplex RT-PCR, is designed such that the amplified target sequences can be cleaved with appropriate restriction enzymes, and covalent linkage by DNA ligation can be performed (the primer design is described in the section "Primer Mixtures and Design"). Following multiplex RT-PCR amplification with such a multiplex primer mix, the restriction enzymes needed to form compatible ends of the target sequences, are added to the mixture together with the ligase. No purification of the PCR products is needed prior to this step, although purification may be performed. The reaction temperature for the combined restriction cleavage and ligation is approximately between 0 and 40° C. However, if the polymerase from the multiplex PCR reaction is still present in the mixture, an incubation temperature below room temperature is preferred, most preferred are temperatures between 4 and 16° C.
[0094]In yet another embodiment of the present invention, the plurality of nucleotide sequences of interest, amplified by multiplex RT-PCR, are linked by recombination. In this approach, the target sequences amplified can be joined using identical recombination sites. Linkage is then performed by adding the recombinases facilitating recombination. Some suitable recombinase systems are Flp recombinase with a variety of FRT sites, Cre recombinase with a variety of lox sites, integrase ΦC31 which carries out recombination between the attP site and the attB site, the β-recombinase-six system as well as the Gin-gix system. Linkage by recombination has been exemplified for two nucleotide sequences (VH linked with VL) (Chapal, N. et al. 1997 BioTechniques 23, 518-524), hereby incorporated by reference.
[0095]In a preferred embodiment of the present invention, the nucleotide sequences of interest comprise variable region encoding sequences and the linkage generates a cognate pair of variable region encoding sequences. Such a cognate pair may comprise one or more constant region encoding sequences in addition to the variable regions.
[0096]In an even more preferred embodiment of the present invention, the nucleotide sequences of interest comprise immunoglobulin variable region encoding sequences and the linkage generates a cognate pair of light chain variable region and heavy chain variable region encoding sequences. Such a cognate pair may comprise one or more constant region encoding sequences in addition to the variable regions. Further, such a cognate pair may be isolated from template derived from cells of the B-lymphocyte lineage enriched from a lymphocyte-containing cell fraction, such as whole blood, mononuclear cells or white blood cells.
[0097]In a just as preferred embodiment of the present invention, the nucleotide sequences of interest comprise TcR variable region encoding sequences and the linkage generates a cognate pair of a chain variable region and β chain variable region encoding sequences or γ chain variable region and δ chain variable region encoding sequences. Such a cognate pair may comprise one or more constant region encoding sequences in addition to the variable regions. Further, such a cognate pair may be isolated from template derived from cells of the T-lymphocyte lineage enriched from a lymphocyte-containing cell fraction, such as whole blood, mononuclear cells or white blood cells.
[0098]Another aspect of the present invention, is to utilize the multiplex RT-PCR with a population of genetically diverse cells as template source. The majority of heteromeric protein encoding sequences do not vary from cell to cell as is the case with variable region encoding sequences from binding proteins. Thus, when utilizing the present invention for the cloning of such non-variable heteromeric protein encoding sequences there is no need to perform an initial isolation of single cells.
[0099]In this embodiment of the present invention, a plurality of non-contiguous nucleotide sequences of interest are linked randomly by a method comprising, performing multiplex RT-PCR amplification of nucleotide sequences of interest using a template derived from a population of genetically diverse cells and effecting linkage of the amplified nucleotide sequences of interest. Further, the method comprises an optional step of performing an additional amplification of the linked products. As with the single cell approach the linkage can either be performed utilizing a multiplex overlap-extension primer mix for the amplification or alternatively by ligation or recombination. Preferably the template derived from the population of cells is not strictly contained within the cells. The population of cells may for example be lysed.
[0100]Application of the process of random linkage on a population of cells expressing variant binding proteins, allows for a simplified generation of combinatorial libraries of variable region encoding sequences. Preferably, the population of cells constitutes cells that express variable region binding proteins, such as B lymphocytes, T lymphocytes, hybridoma cells, plasma cells, or a mixture of these cells.
[0101]The population of cells in the above mentioned embodiment can for example be permeabilized or lysed, without additional purification, or the template nucleic acids can be isolated from the cells by standard procedures. The single-step multiplex RT-PCR procedure is preferred. However, the two-step procedure may also be used in the embodiment.
[0102]The invention also provides a combinatorial library comprised of linked pairs of immunoglobulin light chain variable region and heavy chain variable region encoding sequences.
[0103]An efficient way to increase the specificity, sensitivity, and yield of the multiplex RT-PCR-linkage process, is by performing an additional molecular amplification of the linked nucleotide sequences obtained from the multiplex RT-PCR followed by linkage by ligation or recombination or linkage using the multiplex overlap-extension RT-PCR. This additional amplification is preferably performed with PCR amplification, utilizing a primer mix adapted for amplifying the linked nucleic acid sequences of interest. The primer mix utilized may be the outer primers of the multiplex primer mix or multiplex overlap-extension primer mix, meaning the primers which anneal to the outermost 5' end and 3' end of the sense strand of the linked variable region encoding sequences, thereby enabling the amplification of the entire linked product. The outer primers can also be described as the primers of the multiplex overlap-extension primer mixture that do not contain overlap extension tails. Alternatively, a nested or semi-nested primer set can be used for the additional amplification of the linked nucleotide sequences. Such a nested PCR especially serves to increase the specificity of the method as well as to increase the amount of linked product. For the present invention, semi-nested PCR (as described in the section entitled Primer Mixtures and Design) is considered to function as well as the nested PCR. Thus, it is desired although not necessary for the present invention to perform an additional PCR amplification of the linked products from the multiplex overlap-extension RT-PCR or of the products linked by ligation or recombination, preferably using nested PCR or semi-nested PCR.
[0104]The additional amplification can either be performed directly using a fraction or the entire multiplex overlap-extension RT-PCR reaction product or ligation product or recombination product, or a fraction of any one of these products or using partially purified linked products from any one of these reactions, e.g. by performing an agarose gel electrophoresis of the linked products, and excising the fragment corresponding to the expected size of the linked variable region encoding sequences. For products linked by multiplex overlap-extension RT-PCR, the additional amplification is preferably performed directly on a fraction from the multiplex overlap-extension RT-PCR reaction, since this would assist linkage of the individual target sequences that were not linked in the first reaction.
Sequences of Interest
[0105]The nucleotide sequences of interest of the present invention can be selected from sequences that encode different subunits or domains, which when expressed, forms a protein or part of a protein. Such proteins that are composed of at least two non-identical subunits are known as heteromeric proteins. Heteromeric proteins are common in all kinds of species. Some of the classes to which such proteins belong are for example enzymes, inhibitors, structural proteins, toxins, channel proteins, G-proteins, receptor proteins, immunoglobulin superfamily proteins, transportation proteins etc. The nucleotide sequences encoding such heteromeric proteins are non-contiguous, meaning for example that they originate from different genes, or different mRNA molecules. However, non-contiguous as used in the present invention may also mean nucleotide sequences encoding domains of the same protein, where the domains are separated by nucleotide sequences which are not of interest.
[0106]In one embodiment of the present invention the nucleotide sequences of interest contain variable region encoding sequences from the immunoglobulin superfamily, such as immunoglobulins (antibodies), B cell receptors and T cell receptors (TcR's). Especially variable region encoding sequences from immunoglobulins are of interest. Such variable region encoding sequences comprise full-length antibodies as well as Fab's, Fv's, scFv's and combinations of fragments of the variable region encoding sequences, e.g. complementarity determining regions (CDR's), joining genes or V-genes or combinations of these. Generally the present invention can be applied with any combinations of variable region encoding sequences and fragments thereof. The present application exemplifies the linkage of the entire light chain with the variable domain of the heavy chain. However, the present invention also allow for the linkage of only the variable domains of the heavy and light chains generating Fv or scFv encoding sequences, or the linkage of the entire light chain with the heavy chain variable region+constant region domain CHi+parts of the hinge region, generating Fab, Fab' or F(ab)2. Further, it is possible to add any region of the heavy chain constant region domains to the variable heavy chain, thereby generating truncated antibody encoding sequences or full-length antibody encoding sequences.
[0107]In a further embodiment of the present invention variable region encoding sequences comprise one type of immunoglobulin light chain (kappa or lambda) encoding sequence and one immunoglobulin heavy chain variable region encoding sequence.
[0108]Variable region encoding sequences derived from T cell receptors (TcR's) are also of interest. Such TcR encoding sequences comprise encoding sequences for full-length alpha and beta chains or gamma and delta chains as well as soluble TcR's or only the variable domains of these chains or single chain fusion proteins thereof (e.g. single chain a43 or single chain γδ).
Template Sources
[0109]One feature of the present invention is the ability to link nucleotide sequences derived from an isolated single cell, a population of isogenic cells, or a genetically diverse population of cells which have not been separated into single vessels. The cells utilized in the present invention can for example be bacteria, yeast, fungi, insect cells, plant cells or mammalian cells or fractions of such cells. Blood cells derived from mammals are one example of a fraction of cells that can be utilized in the present invention.
[0110]A preferred feature of the present invention is the use of isolated single cells or a population of isogenic cells as template source, since scrambling of the nucleic acid sequences of interest, in particular variable region encoding sequences is avoided. This is of importance if one wishes to obtain an original pair of for example variable region encoding sequences.
[0111]Another preferred feature of the present invention, is obtaining a single cell or population of single cells from a cell fraction comprising lymphocytes, such as B lymphocytes, T lymphocytes, plasma cells and/or various developmental stages of these cell lineages. Other populations of cells that express binding proteins from the immunoglobulin superfamily might also be used to obtain single cells. Cell lines such as hybridoma cells, cell lines of B lymphocyte or T lymphocyte lineage or virus immortalized cell lines or donor derived cells participating in the immune response are also applicable in the present is invention. Donor derived lymphocyte-containing cell fractions may be obtained from natural tissue or fluid which is rich in such cells, e.g. blood, bone marrow, lymph nodes, spleen tissue, tonsil tissue or from infiltrations in and around tumors or inflammatory tissue infiltrations. Suitable cell donors for the present invention can be selected from vertebrates that all contain an acquired immune system. Donors can either be naive or hyperimmune with respect to a desired target. For the isolation of antigen binding proteins with binding specificities toward a desired target, hyperimmune donors are preferred. Such hyperimmune donors can either be donors immunized with the target, or fragments of the target, or it can be convalescent patients, or non-healthy individuals which are running a natural immune response towards the target e.g. autoimmune patients, cancer patients, patients with infectious diseases e.g. HIV patients, Hepetitis A, B or C patients, SARS patients etc., or patients with chronic diseases.
[0112]When utilizing recombinant proteins for treatment, it is preferable that they are derived from sequences that have species identity with the individual to be treated (e.g. human sequences for treatment of humans). Firstly, because recombinant proteins derived from a foreign sequence (i.e. non-human) will be recognized by the immune system leading to an immune response implicating polyclonal anti-protein antibodies. These anti-protein antibodies can block the drug action by occupying the active site, they will accelerate drug clearance and they could potentially induce adverse reactions such as hypersensitivity reactions upon repeated exposure.
[0113]Immunogenicity may however, also be seen in cases where the recombinant protein is derived from a sequence that have species identity. Such immunogenicity can for example be induced by post-translational modifications that might differ from those seen in vivo. Combinatorial libraries of variable region encoding sequences might also give rise to immunogenicity, since they consist of random pairs of variable region encoding sequences created in vitro. The rules that govern formation of antibody heavy and light chain encoding sequence pairs (or T cell receptors) in vivo are not completely understood. Hence, it follows that some of the in vitro formed pairs could be recognized as foreign by the immune system, even though both sequences constituting the pair are perfectly human. Binding proteins obtained from cognate libraries do on the other hand not create said abnormal combinations and they are consequently of lesser potential immunogenicity than binding proteins from combinatorial libraries. This does not mean that products from combinatorial libraries are unsuitable for treatment, they just require a larger degree of monitoring with respect to the above mentioned side effects.
[0114]For use in the present invention, cell donors should preferably be of the same species as the species to be treated with the products obtainable from the linked nucleotide sequences of the present invention. Preferably, a cell donor is a domestic animal, a pet, a human or a transgenic animal. Transgenic animals carrying human immunoglobulin loci are described in U.S. Pat. No. 6,111,166 and Kuroiwa, Y. et al. Nature Biotechnology; 2002; 20: 889-893. Such transgenic animals are capable of producing human immunoglobulins. Thus, fully human antibodies against a specific target can be raised by usual immunization techniques of such transgenic animals. This allows for generation of libraries encoding for binding proteins with specificities towards more difficult targets such as human antigens to which no or limited natural human antibody response exist. Such transgenic animals can likewise be developed to produce human T cell receptors.
[0115]In a further embodiment of the present invention, the lymphocyte-containing cell fraction is constituted of whole blood, bone marrow, mononuclear cells, or white blood cells obtained from a donor. Mononuclear cells can be isolated from blood, bone marrow, lymph nodes, spleen, infiltrations around cancer cells and inflammatory infiltrations. Mononuclear cells can be isolated by density centrifugation techniques, e.g. Ficoll gradients. If the mononuclear cells are isolated from samples composed of tissue, the tissue is disintegrated before the gradient centrifugation is performed. Disintegration can be performed, for example, by mechanical methods such as grinding, electroporation and/or by chemical methods such as enzymatic treatments. The isolation of white blood cells can be performed directly from donors using leukopheresis. Raw preparations of for example bone marrow or tissue, which contain lymphocytes, can also be used in the present invention. Such preparations will need to be disintegrated, for example as described above, in order to facilitate single cell distribution.
[0116]A further feature of the present invention is enrichment of the lymphocyte-containing cell fraction e.g. whole blood, mononuclear cells, white blood cells or bone marrow, with respect to a particular lymphocyte population, such as cells from the B lymphocyte or T lymphocyte lineage. Enrichment of B lymphocytes can for example be performed, using magnetic bead cell sorting or fluorescence activated cell sorting (FACS) taking advantage of lineage-specific cell surface marker proteins such as CD19 or other B cell lineage-specific markers. Enrichment of T lymphocytes can for example be performed, utilizing a cell surface marker such as CD3 or other T cell lineage-specific markers.
[0117]A preferred feature of the present invention is to sort the enriched B lymphocytes further in order to acquire plasma cells, before distributing the cells individually among a plurality of vessels. Isolation of plasma cells is generally performed by FACS sorting, utilizing surface markers such as CD38 possibly in combination with CD45. Other plasma cell-specific surface markers or combinations thereof can be utilized as well, for example CD138, CD20, CD21, CD40, CD9, HLA-DR or CD62L, the exact choice of marker depends on the plasma cell source, e.g. tonsils, blood or bone marrow. Plasma cells can also be obtained from a non-enriched lymphocyte-containing cell population obtained from any of these sources. The plasma cells isolated from blood are sometimes called early plasma cells or plasmablasts. In the present invention these cells are also termed plasma cells although they are CD19 positive in contrast to plasma cells residing in the bone marrow. Plasma cells are desired for the isolation of cognate pairs of immunoglobulin encoding sequences because a higher frequency of these cells produces antigen-specific antibodies that reflect the acquired immunity toward the desired antigen and most of the cells have undergone somatic hypermutation and therefore encode for high-affinity antibodies. Further, the mRNA levels in plasma cells are elevated compared to the remaining B lymphocyte population, thus the reverse transcription procedure is more efficient when using single plasma cells. As an alternative to plasma cell isolation, memory B cells may be isolated from a lymphocyte containing cell fraction utilizing a cell surface marker such as CD22.
[0118]An alternative feature of the present invention, is selecting the enriched B lymphocytes for antigen specificity before distributing the cells among a plurality of vessels. Isolation of antigen-specific B lymphocytes is performed by contacting the enriched B lymphocytes with the desired antigen or antigens enabling binding of antigen to surface exposed immunoglobulin, followed by isolation of binders. This can be done, for example, by coating magnetic beads with the desired antigen or antigens followed by magnetic bead cell sorting, by FACS, by coating a column with the antigens followed by affinity chromatography, by filter screening assays or other methods known in the art. Plasma cells as well as B lymphocytes, non-enriched mononuclear cells, white blood cells, whole blood, bone marrow or tissue preparations can be subjected to isolation with respect to antigen specificity if this is desired.
[0119]Another feature of the present invention, is to sort enriched T lymphocytes (e.g. CD3 positive cells) using surface markers CD45R0 and/or CD27 to obtain a fraction of memory T cells. T lymphocytes can also be selected for MHC-antigen specificity using MHC-peptide complexes (e.g. Callan, M. F. et al. 1998. J. Exp. Med. 187, 1395-1402; Novak, E. J. et al. 1999. J. Clin. Invest 104, R63-R67).
[0120]A further feature of the present invention is immortalization of any of the isolated cell fractions described in the above (e.g. B lymphocytes, plasma cells, memory cells or T lymphocytes). Immortalization may for example be performed with Epstein-Barr virus (Traggiai, E., et al., 2004. Nat Med 10, 871-875) prior to cell distribution. Alternatively, isolated single cells may be immortalized and expanded prior to reverse transcription. Traggiai et al., Nat. Med. 2004 August; 10(8):871-5.
[0121]A further feature of the present invention, is the distribution of a population of desired cells (e.g. hybridoma cells, cell lines of B lymphocyte or T lymphocyte lineage, whole blood cells, bone marrow cells, mononuclear cells, white blood cells, B lymphocytes, plasma cells, antigen-specific B lymphocytes, memory B cells, T lymphocytes, antigen/MHC-specific T lymphocytes, or memory T cells) individually, into a plurality of vessels, in order to obtain a population of isolated single cells. This isolation of single cells refers to the physical separation of cells from a population of cells in such a way that a single vessel contains a single cell, or a micro array, chip or gelmatrix is loaded in a manner that produce single cells. The cells may be distributed directly into multitudes of vessels such as arrays of single vessels by limiting dilution. The single vessels utilized in the present invention are preferably those applicable in PCR (e.g. PCR tubes and 96 well or 384 well PCR plates or larger arrays of vessels). However other vessels may also be used. When distributing single cells into a large number of single vessels (e.g. 384 well plates), a population of single cells is obtained. Such a distribution may be performed, for example, by dispensing a volume into a single vessel that on average encompasses a cell concentration of one, 0.5 or 0.3 cell, thereby obtaining vessels that on average contain a single cell or less. Since distribution of cells by limiting dilution is a statistical event, a fraction of the vessels will be empty, a major fraction will contain a single cell, and a minor fraction will contain two or more cells. Where two or more cells are present in a vessel some scrambling of the variable region encoding sequences may occur among the cells present in the vessel. However, since it is a minor event it will not affect the overall utility of the present invention. Additionally, combinations of variable region encoding sequences which do not posses the desired binding affinity and specificity will most likely not be selected and hence eliminated during a screening process. Therefore, minor events of scrambling will not significantly affect the final library of the present invention.
[0122]There are alternatives to cell distribution by limiting dilution using, for example, cell sorters such as FACS machines or robots that can be programmed to accurately dispense single cells into single vessels. These alternatives are preferable, since they are less laborious and are more efficient in uniformly obtaining a distribution of single cells into single vessels.
[0123]The enrichment, sorting and isolation procedures described in the above, are performed such that the majority of the cells are kept intact. Rupture of cells during enrichment and sorting might result in scrambling of the variable region encoding sequences. However, this is not expected to be a problem since the frequency of rupture is expected to be low. Washing and possible RNAse treatment of the cells prior to distribution into single vessels will remove any RNA that has leaked during the process.
[0124]Further, when considering the above descriptions of how to distribute cells in order to obtain a population of single cells in a population of single vessels, it is not to be interpreted as an absolutely required feature that every vessel must contain a single cell. Rather, it indicates that a majority of the vessels contain single cells, e.g. the number of vessels with two or more cells is below 25% of the total amount of cells distributed, or even better it is below 10%.
[0125]A further feature of the present invention is the performance of a reverse transcription using template derived from cells distributed individually among a plurality of vessels.
[0126]For the purpose of reverse transcription (RT), in accordance with the present invention, the nucleic acids within a single cell that is to serve as template source for the RT, are considered to be derived from a single cell although they have not necessarily been separated from the remaining contents of that single cell.
[0127]When the final distribution of the single cells to their single vessels has been performed, the single cells may be expanded in order to obtain a population of isogenic cells prior to reverse transcription. This process yields more mRNA to be used as template, which might be important if a rare target is to be amplified and linked. However, the cells should remain genetically identical with respect to the target gene during the expansion. The isolated cells or the population of isogenic cells can either be kept intact or lysed, as long as the template for the reverse transcription is not degraded. Preferentially, the cells are lysed in order to ease the following reverse transcription and PCR amplification.
[0128]In a different embodiment of the present invention, the disclosed multiplex overlap-extension RT-PCR method or multiplex RT-PCR followed by linkage by ligation or recombination may also be utilized on template derived from a genetically diverse population of cells which have not been separated into single vessels, but all remain together as a pool of cells. This method may be used for the generation of combinatorial libraries. Such an approach will not require the distribution of single cells. However, the cells which may be used in this approach are the same as those described for the single cell approach, for example a population (pool) of sorted B lymphocytes or T lymphocytes. When performing the single-step multiplex overlap-extension RT-PCR or single-step multiplex RT-PCR followed by linkage by ligation or recombination on such a population of cells, it is preferable to lyse the cells prior to the reaction and if desired total RNA or mRNA may be isolated from the lysate.
[0129]The sensitivity of the single-step multiplex overlap-extension RT-PCR of the present invention enables the use of a very low amount of template. As shown in Example 2 and 3, single-step multiplex overlap-extension RT-PCR may be carried out on an amount of template corresponding to the lysate of a single cell.
Primer Mixtures and Design
[0130]The primer mixtures of the present invention comprise at least four primers that form primer sets two by two, which are capable of amplifying at least two different target sequences of interest. Mixtures of two or more of such primer sets constitute a multiplex primer mix. Preferably, a multiplex mix comprise at least 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 or 20, 30, 40, 50, 60, 70, 80, 90 100, 110, 120, 130, 140 or 150 primer sets (primer pairs). In particular for the amplification of variable region encoding sequences, may an individual primer set within the multiplex primer mix constitute several more than two primers. Preferably, an individual primer set comprise at least 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 20, 25, 30, 35, 45, 50, 60, 70, 80, 90, 100, 110, 120, 130, 140, 150, 160, 170, 180, 190, 200, 220, 240, 260, 280 or 300 primers. Preferably the total number of primers in a multiplex primer mix is at least 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 20, 25, 30, 35, 45, 50, 60, 70, 80, 90, 100, 125, 150 or 200 and at the most 225, 250, 275, 300, 325, 350, 375 or 400 primers.
[0131]All the primers of the present invention comprise a gene-specific region, and preferably all primers are additionally equipped with a primer tail at the 5' end of the primer, i.e. 5' non-coding sequences which are fused to the 3' end of the gene-specific primer part. Such a primer tail is approximately from 6 to 50 nucleotides long, but it may also be longer if desired. Upon amplification the primer tails are added to the target sequences.
[0132]Primer tails of the present invention are for example, cloning tails and linkage tails such as, tails adapted for linkage by ligation, tails adapted for linkage by recombination or overlap-extension tails.
[0133]Cloning tails may be from 6 to 20 nucleotides long or longer and comprise restriction sites and/or recombination sites, which are useful for the insertion of the linked product into an appropriate vector.
[0134]To enable linkage by ligation, the primer sets of the multiplex primer mix are designed such that one part (forward or reverse primer(s)) of the first primer set is equipped with a linkage-tail containing a restriction site that upon cleavage will be compatible with a restriction site located in the linkage tail of one part of the second primer set. For linkage of more than two target sequences, the second part of the second primer set is equipped with a restriction site that upon cleavage will be compatible with a restriction site located in one part of the third primer set. This second restriction site located in the second primer set should be non-compatible with that of the first primer set. A considerable number of target sequences can be linked by designing primer sets this way. Restriction sites with a low frequency or no occurrence, in the target sequences should be chosen. Further, it is preferable that compatible restriction sites are not identical, such that the site of ligation becomes cleavage-resistant for the particular restriction enzymes used. This will drive the reaction towards linkage of target sequence one with target sequence two, since linkage between identical target sequences will be cleavable by the restriction enzymes. Suitable pairs of restriction sites are for example, SpeI with XbaI (alternatively NheI or AvrII can substitute one or both of these), NcoI with BspHI, EcoRI with MfeI or PstI with NsiI. For linkage, SpeI can for example be located in target sequence one, XbaI can be located in target sequence two, NcoI can be located at the other end of target sequence two and BspHI in target sequence three and so forth. To simplify the process further, it is an advantage if the restriction enzymes function in the same buffer.
[0135]To enable linkage by recombination the primer sets of the multiplex primer mix can for example be designed as exemplified in the article by Chapal (1997 BioTechniques 23, 518-524), which is hereby incorporated by reference.
[0136]To enable the linkage of the nucleotide sequences of interest in the same step as the multiplex PCR amplification, tails adapted for overlap-extension PCR are added to at least one primer of each primer set of the multiplex primer mix, thereby generating a multiplex overlap-extension primer mixture.
[0137]The overlap-extension tails are typically longer, ranging from 8 to 75 nucleotides in length and may contain restriction sites or recombination sites which allow for subsequent insertion of regulatory elements such as promoters, ribosomal binding sites, termination sequences, or linker sequences such as in a scFv. The overlap-extension tail may also contain a stop codon if that is desired. Generally there are three types of overlap-extension tails, as illustrated in FIG. 1. In type I the overlap-extension tails of two primer sets solely overlap with each other. Not necessarily all of the nucleotides of two overlap-extension tails are complementary to each other. In one aspect of the present invention the complementary nucleotides represent between 60 to 85% of the overlap-extension tail. In type II overlap-extension tails, 4 to 6 of the 5' nucleotides are complementary to the gene-specific region of the adjacent target sequence. In type III overlap-extension tails, the entire overlap is complementary to the adjacent target sequence. The type I and II overlap-extension tails are preferred when regulatory elements and the like are later to be inserted between the linked target sequences. Type II overlap-extension tails are preferred if the target sequences are to be linked by a defined linker as seen with scFv. Type III overlap-extension tails are preferred if the target sequences are to be linked in-frame to each other.
[0138]Design of overlap-extension tails is dependent on sequence features such as length, relative GC content (GC %), presence of restriction sites, palindromes, melting temperature, the gene-specific part to which they are coupled etc. The length of the overlap-extension tails should be between 8 and 75 nucleotides long, preferably they are from 15 to 40 nucleotides long. Even more preferred they are from 22 to 28 nucleotides long. The use of very long overlap-extension tails (50 to 75 nucleotides) could favor the linkage of the products produced by each primer set. However, the proportion between the length of the overlap-extension tail and the gene-specific region probably will need to be adjusted when using very long overlap-extension tails. The GC % preference is dependent on the length of the overlap-extension tail. Since shorter tails have a shorter area where they are complementary they need a higher GC % to strengthen the interaction than longer tails. Other principles of primer design should likewise be observed, e.g. primer dimerization and hairpin formation should be minimized. Neither shall they engage in false priming. Further, it is known that Taq DNA polymerase often adds an adenosine (A) at the 3' end of the newly synthesized DNA strand, and this can be accommodated for in overlap-extension tail design by enabling overlap-extension tails to accommodate 3' non-template A addition.
[0139]The choice of primers that carry the linkage tail, e.g. the overlap-extension tail, tail adapted for linkage by ligation or tail adapted for linkage by recombination, defines the order and direction of linkage of the target sequences. It is not essential to the invention whether it is the forward primer(s) or reverse primer(s) of a primer set or possibly both forward and reverse primers that are equipped with the linkage tail. However, some consideration should be given to this anyway since the order and direction of the target sequences in the final product might be of relevance e.g. for the insertion of regulatory elements such as promoters and termination sequences or for the in-frame linkage of the individual target sequences.
[0140]For the linkage of two nucleotide sequences of interest the linkage tail may be added either to the reverse primer(s) or forward primer(s) of each primer set used for the PCR amplification of each target sequence. The present invention exemplifies addition of overlap-extension tails and tails adapted for linkage by ligation, to the VH and VL forward primers of each set (e.g. FIG. 2 and example 9, respectively). This results in a linking direction of the products that is 5' to 5' (head-to-head and bi-directional). However, linkage tails might as well be added to the reverse primer(s) of each set (e.g. C.sub.κ and/or C.sub.λ in the first set and CH or JH in the second set). This results in a linking direction of the product that is 3' to 3' (tail-to-tail and bi-directional). A third option is adding the linkage tails to the reverse primer(s) of the first primer set (e.g. C.sub.κ and/or C.sub.λ primers) and the forward primer(s) of the second primer set (e.g. VH primer(s)) or visa versa. This results in a 3' to 5' orientation (head-to-tail and uni-directional). FIG. 3 illustrates the possible directions that can be generated depending on which primer of each primer set that is equipped with the linkage tail.
[0141]When linking more than two nucleotide sequences of interest some of the primer sets need to have linkage tails on both the forward and reverse primers, such that one tail is complementary to a tail of the preceding primer set and the other tail is complementary to one of the primers of the subsequent primer set. This principle holds for all the primer sets that amplify target sequences that are to be linked between two other target sequences.
[0142]The design of the gene-specific primer part generally should observe known primer design rules such as minimizing primer dimerization, hairpin formation and non-specific annealing. Further, multiple G or C nucleotides as the 3' bases are to be avoided when possible. The melting temperature (Tm) of the gene-specific regions in a primer set should preferably be equal to each other plus/minus 5° C. In the present invention Tm values between 45° C. and 75° C. are desirable and Tm values of about 60° C. are optimal for most applications. Advantageously, the initial primer design can be aided by computer programs developed for this task. However, primer designs generally need laboratory testing and routine optimization. This may be done, for example, by analyzing size, restriction fragment length polymorphism (RFLP) and sequencing of the amplification products obtained using the primer sets. The use of degenerate positions within primers is a useful approach when amplifying sequences with variable regions or when searching for new family members belonging to a specified class of proteins. The numbers of degenerate positions may also require optimization.
[0143]The present invention encompasses improved primer sets that can be used together in a highly multiplexed format. The primer set described by de Haard (de Haard, H. J. et al. 1999. J. Biol. Chem. 274, 18218-18230) were used as starting point, and was modified by trimming the 3' ends of the primers to reduce non-specific interactions and adding overlap-extension tails or tails adapted for linkage by ligation.
[0144]One feature of the present invention, are primer mixes composed of at least two primer sets that are able to prime amplification and promote linkage of at least two nucleotide sequences of interest. The primer mixes of the present invention are capable of priming the amplification of at least two subunits or domains from heteromeric proteins, e.g. belonging to the class of enzymes, inhibitors, structural proteins, toxins, channel proteins, G-proteins, receptor proteins, immunoglobulin superfamily proteins, transportation proteins etc.
[0145]A further feature of the present invention is a multiplex overlap-extension primer mix comprising primer sets wherein at least one primer set member of each primer set comprises an overlap-extension tail capable of hybridizing to the overlap-extension tail of a primer set member of a second primer set.
[0146]The overlap-extension tails enables the immediate linkage of the nucleotides of interest during the multiplex overlap-extension PCR amplification by equipping each individual product arising from the primer sets with a tail that is complementary to an adjoining product. This however does not mean that the linkage necessarily occur during this first PCR amplification. Depending on the reaction setup, the majority of the actual linkage may be performed during an additional amplification with the outer primers of the first PCR amplification (multiplex PCR amplification).
[0147]A further feature of the present invention, is a primer set designed to amplify a family of nucleotide sequences containing variable region encoding sequences. Examples of such families are kappa light chains (e.g. VKI-VI in humans), lambda light chains (e.g. VL1-10 in humans) and variable heavy chains (e.g. VH1-7 in humans and VH1-15 in mice) from immunoglobulins, and α, β, γ or δ TcR variable regions. A primer set for the amplification of a family of nucleotide sequences containing variable region encoding sequences often comprise a plurality of primers where several primers can be degenerate primers. Amplification of families of immunoglobulin light chain variable region encoding sequences is for example performed using a primer set comprised of a plurality of primers complementary to the 5' end of the variable region of the kappa chain (VD, primer(s)) or the kappa leader sequence (VLκL primer(s)) and/or the lambda chain (VD, primer(s)) or the lambda leader sequence (VLλL primer(s)) (forward primers) together with constant region kappa (C.sub.κ primer(s)) and/or lambda primers (C2, primer(s)) (reverse primers) or a plurality of such primers. Alternatively, light chain joining region primers (JLκ and/or JLλ primer(s)) may be used as reverse primers instead of the constant region primers. Alternatively, forward primers anneal in the UTR region preceding the leader sequence of the variable light chain. Equally, families of immunoglobulin heavy chain variable region encoding sequences can be amplified with one primer set utilizing various primer combinations. For example, a plurality of primers complementary to the 5' end of the heavy chain variable region (VET primer(s)) or the leader sequence of this region (VHL primer(s)) (forward primers) together with a plurality of heavy chain joining region primers (JH primer(s)) or heavy chain constant region primer(s) (reverse primers). The CH primer may be isotype-specific and in principle any CH primer can be utilized (e.g. CH1, CH2, CH3, or CH4), also one that would result in a full-length heavy chain. Alternatively, forward primers anneal in the UTR region preceding the leader sequence of the variable heavy chain.
[0148]The use of forward primers annealing in the leader sequence instead of the 5' end of the variable region is particular useful if cross-hybridization is observed for the variable region primers. Since mutations due to cross-hybridization will be eliminated from the final protein because leader sequences are cleaved off during protein processing within the cell. to Example 9 describes the design of antibody variable heavy chain and kappa light chain leader primers. The aspect that the site of priming is located in the 3' end of the leader encoding sequence (C-terminal) is an advantage over previous antibody leader primers, since this allows for shuttling of the amplified sequences between bacterial and eukaryotic expression vectors in a way that allow functional leader sequences in both system. The design system described in Example 9 can easily be applied to antibody lambda light chains, TcR α, β, γ or δ chains as well.
[0149]One feature of the present invention are primers which anneal in the 3' end of the leader encoding sequence preceding a variable region encoding sequence, and their use for amplification of variable region encoding sequences.
[0150]A preferred feature of the present invention is the application of primers with at least 90% sequence identity (preferably at least 95% identity) with the gene-specific region of SEQ ID NO 86 to 92, which correspond to primers annealing in the C-terminal of heavy chain leader encoding sequences (VHL primers). The gene-specific sequence of these SEQ ID NO's correspond to base number 18 to the 3' end of the sequences (see also table 11).
[0151]Another preferred feature of the present invention is the application of primers with at least 90% sequence identity (preferably at least 95% identity) with the gene-specific region of SEQ ID NO 93 to 98, which correspond to primers annealing in the C-terminal of kappa light chain leader encoding sequences (VLκL primers). The gene-specific sequence of these SEQ ID NO's correspond to base number 25 to the 3' end of the sequences (see also table 11).
[0152]In one embodiment of the present invention, the multiplex overlap-extension primer mix utilized for the multiplex overlap-extension PCR and possibly for the reverse transcription step as well comprises a) at least one CL or JL primer complementary to the sense strand of an immunoglobulin light chain region encoding sequence; b) at least one VL 5' primer or VL leader primer complementary to the antisense strand of an immunoglobulin light chain variable region encoding sequence and capable of forming a primer set with the primer(s) in a); c) at least one CH or JH primer complementary to the sense strand of an immunoglobulin constant heavy chain domain encoding sequence or the heavy chain joining region and d) at least one VH 5' primer or VH leader primer complementary to the antisense strand of an immunoglobulin heavy chain variable region encoding sequence, and capable of forming a primer set with the primer(s) in c).
[0153]Primer sets of the present invention can for example be VLκ+C.sub.κ, VLλ, VLκ+JLκ, VLλ+JLλ, VLκL, VLλL+C.sub.λ, VLκL+JLκ, VLλL+JLλ, VH+JH, VH+CH, VHL+JH or VHL+CH or combinations of these, capable of amplifying a variable region encoding target sequence.
[0154]In a further embodiment are the CL (JL) primer(s) and VL (VLL) primer(s) adapted for amplifying sequences comprising kappa light chain variable regions or lambda light chain variable regions.
[0155]In a preferred embodiment of the present invention the are the CL (JL) primer(s) and VL (VLL) primer(s) adapted for amplifying both kappa and lambda light chain variable region encoding sequences.
[0156]In an even more preferred embodiment of the present invention, are the forward primers for light chain amplification VLL primers with at least 90% sequence identity (preferably at least 95% identity) with the gene-specific region of SEQ ID 93 to 98, and the forward primers for heavy chain amplification are VHL primers with at least 90% identity (preferably at least 95%) with the gene-specific region of SEQ ID 86 to 92.
[0157]In a further embodiment of the present invention, carries the immunoglobulin VL/VLL and VH/VHL primers linkage tails, preferably in the form of complementary overlap-extension tails. This generates variable region encoding sequences that are linked in a head-to-head fashion. For the linkage of variable region encoding sequences in a head-to-tail fashion, either the CL/JL and VH/VHL primers contain linkage tails or the VL/VLL and CH/JH primers contain linkage tails, preferably in the faun of complementary overlap-extension tails. For the linkage of variable region encoding sequences in a tail-to-tail fashion, the CL/JL and CH/JH primers contain linkage tails, preferably in the form of complementary overlap-extension tails (FIG. 3).
[0158]Preferentially, the multiplex primer mixes, including multiplex overlap-extension primer mixes, of the present invention comprise two primer sets. Thus, a multiplex primer mix comprises at least four different primers. In a further aspect of the present invention a multiplex primer mix comprises more than four different primers. A multiplex primer mix of the present invention is used for the amplification of target sequences in a single vessel. For example are kappa, lambda and heavy chain variable regions all amplified in the same vessel. One multiplex primer mix of the present invention was comprised of 16 different degenerate primers distributed as follows: eight VH primers, one CH1 primer, six VLκ primers, and one C.sub.κ primer (FIG. 2). Another set was comprised of 19 degenerate primers distributed as to follows: eight VH primers, four JH primers, six VLκ primers, and one C.sub.κ primer. A third set was comprised of 22 degenerate primers distributed as follows: eight VH primers, one CH1 primer, eleven VLλ primers, and two C.sub.λ primers. A fourth set was comprised of 27 degenerate primers distributed as follows: eight VH primers, one CH1 primer, six VLκ primers, one C.sub.κ primer, eleven VLλ primers, and two C.sub.λ primers.
[0159]The present invention also encompasses primers for an additional PCR amplification of the linked products obtained by multiplex RT-PCR followed by linkage by ligation or recombination or by multiplex overlap-extension RT-PCR. This additional PCR amplification can be performed using a primer mix adapted for amplifying the linked target sequences. Such a primer mix may comprise the outer primers of the multiplex primer mix or multiplex overlap-extension primer mix, meaning the primers that anneals to the outermost 5' end and 3' end of the sense strand of the linked nucleotide sequences, thereby enabling the amplification of the entire linked product. One example of primers that can be utilized as outer in the present invention are the C.sub.κ/J.sub.κ and/or C.sub.λ/J.sub.λ primers forming a primer set with the JH or CH primers. This process generally serves to increase the amount of linked product obtained from the multiplex RT-PCR followed by linkage by ligation or recombination or from the multiplex overlap-extension RT-PCR.
[0160]Alternatively, a primer set which is nested compared to the outer primers used in the primary multiplex RT-PCR or multiplex overlap-extension RT-PCR reaction can be used for the additional amplification of the linked nucleotide sequences. In the present invention such a primer set is termed a nested primer set. The design of nested primers generally observes the same design rules as for the gene-specific primers previously described, except that they prime 3' to the annealing position of the outer primers used in the multiplex RT-PCR or multiplex overlap-extension RT-PCR. The product resulting from a nested PCR is therefore shorter than the linked product obtained by the multiplex RT-PCR followed by linkage by ligation or recombination or by multiplex overlap-extension RT-PCR. In addition to increase the amount of linked product, the nested PCR further serves to increase the overall specificity, especially of the multiplex overlap-extension RT-PCR technology. However, it should be noted that not all multiplex primer mixes/multiplex overlap-extension primer mixes that have been described previously are suitable for combination with a nested primer set when performing the additional amplification. In such cases the outer primers of the multiplex primer mix/multiplex overlap-extension primer mix can be used for the additional amplification or a semi-nested PCR can be applied as described later.
[0161]In one embodiment of the present invention, a mixture of JL and JH primers is used as nested primers for the additional amplification of the linked immunoglobulin variable region encoding sequences.
[0162]Nested primer sets of the present invention can also be comprised of a reverse (or forward) outer primer(s) from the first multiplex primer mix/multiplex overlap-extension primer mix and a second nested primer that prime 3' to the annealing position of the forward (or reverse) outer primer(s) of the first multiplex primer mix/multiplex overlap-extension primer mix. The use of such a primer set for an additional PCR amplification is generally known as a semi-nested PCR. Semi-nested PCR can for example be applied if it is difficult to design a nested primer in one specific region e.g. for the variable region sequences (V and J primers), because such a primer would have to anneal in the complementarity determining regions (CDRs). Further, semi-nested PCR can be used when it is desirable to keep one end of the linked sequences intact e.g. for cloning purposes.
[0163]One feature of the present invention keeps the constant light chain encoding sequence kept intact during the additional amplification reaction. The CL (reverse) primer(s) used for the additional amplification is only slightly modified compared to the outer primer(s) used for the primary reaction (the multiplex RT-PCR or multiplex overlap-extension RT-PCR reaction). The modification comprises the addition of a few bases to the 3' end of the CL primer(s) used for the primary amplification. Further, a different cloning tail might be added to the nested CL primer(s). The forward primer(s) used in the additional amplification is completely nested compared to the constant heavy chain-specific outer primer(s) used in the primary reaction. The combined use of the outer primer(s) in the primary reaction and the slightly modified nested CL primer(s) together with the completely nested forward primer(s) in the additional amplification results in an increase in specificity that is comparable to what is achievable with a nested PCR using a completely nested primer set.
[0164]In a preferred embodiment of the present invention, the nested PCR is performed with JH primer(s) and modified CL primer(s) where between 2 and 10 gene-specific base pairs have been added at the 3' end compared to the first CL primer(s) of the multiplex primer mix/multiplex overlap-extension primer mix.
Optimization of Multiplex Overlap-Extension PCR
[0165]The parameters of the multiplex overlap-extension PCR step of both the two-step and the single-step procedure can be optimized on several parameters (see, for example, Henegariu, O. et al. 1997. BioTechniques 23, 504-511; Markoulatos, P. et al. 2002. J. Clin. Lab. Anal. 16, 47-51). Generally the same optimization parameters apply for multiplex RT-PCR, although the ratio between outer and inner primers is less important for such a reaction.
[0166]a. Primer Concentration
[0167]The concentration of the primers carrying the overlap-extension tail (for example the VH and VL primers) is preferably lower than the concentration of the outer primers without overlap-extension tail (for example JH and kappa primers).
[0168]If one of the target sequences amplifies with a lower efficiency than the others, for example, as a result of a higher GC %, it may be possible to equalize the amplification efficacy. This may be done by using a higher concentration of the primer set which mediates amplification with low efficiency, or lowering the concentration of the other primer set. For example, sequences encoding for heavy chain variable regions tend to have a higher GC % and hence lower amplification efficiency than light chain variable regions. This points towards using VL primers at a lower concentration than the VH primers.
[0169]Further, when using a large number of primers the total primer concentration might be an issue. The upper limit is determined experimentally by titration experiments. For the "AmpliTaq Gold" PCR system from Applied Biosystems the upper limit was found to be 1.1 μM total oligonucleotide concentration, for other systems it may however be as high as 2.4 μM. Such an upper limit of total oligonucleotide concentration influences the maximal concentration of individual primers. If the individual primer concentration is too low it is likely to cause a poor PCR sensitivity.
[0170]The quality of the oligonucleotide primers have also been found to be important for the multiplex overlap-extension PCR. HPLC-purified oligonucleotides, have produced the best results.
[0171]b. PCR Cycling Conditions:
[0172]Preferentially the cycling conditions are as follows:
TABLE-US-00001 Denaturation: 10-30 s 94° C. Annealing: 30-60 s 50-70° C. Approximately 5° C. below Tm of primers. Extension: 1 min × EPL 65-72° C. EPL is Expected Product Length in kb. Cycle number: 30-80 Final extension: 10 min 65-72° C.
[0173]For the single-step multiplex overlap-extension RT-PCR the following steps were built into the cycling program prior to the amplification cycling outlined above:
TABLE-US-00002 Reverse 30 min 42-60° C. These conditions are also used transcription: where separate reverse transcription is performed. Polymerase 10-15 min 95° C. Hot-start polymerases are activation: favorable in single-step RT- PCR. Activation according to manufacturer.
[0174]It is possible to optimize on all these parameters. Especially the annealing temperature is important. Thus, initially all the individual primer sets that are to constitute the final primer mix should be tested separately in order to identify optimal annealing temperature and time, as well as elongation and denaturing times. This will give a good idea about the window within which these parameters can be optimized for the multiplex overlap-extension primer mix.
[0175]Problems with poor PCR sensitivity, for example due to low primer concentration or template concentration can be overcome by using a high number of thermal cycles. A high number of thermal circles constitute between 35 and 80 cycles, preferably around 40 cycles.
[0176]Further, longer extension times can improve the multiplex overlap-extension PCR process. Long extension times constitute 1.5-4 min×EPL compared to the normal 1 min extension.
[0177]c. Use of Adjuvants
[0178]Multiplex PCR reactions can be significantly improved by using a PCR additive, such as DMSO, glycerol, formamide, or betaine, which relax DNA, thus making template denaturation easier.
[0179]d. dNTP and MgCl2
[0180]Deoxynucleoside triphosphate (dNTP) quality and concentration is important for the multiplex overlap-extension PCR. The best dNTP concentration is between 200 and 400 μM of each dNTP (dATP, dCTP, dGTP and dTTP), above which the amplification is rapidly inhibited. Lower dNTP concentrations (100 μM of each dNTP) suffice to achieve PCR amplification. dNTP stocks are sensitive to thawing/freezing cycles. After three to five such cycles, multiplex PCR often do not work well. To avoid such problems, small aliquots of dNTP can be made and kept frozen at -20° C.
[0181]Optimization of Mg2+ concentration is critical since most DNA polymerases are magnesium-dependent enzymes. In addition to the DNA polymerase, the template DNA primers and dNTP's bind Mg2+. Therefore, the optimal Mg2+ concentration will depend on the dNTP concentration, template DNA, and sample buffer composition. If primers and/or template DNA buffers contain chelators such as EDTA or EGTA, the apparent Mg2+ optimum may be altered. Excessive Mg2+ concentration stabilizes the DNA double strand and prevents complete denaturation of DNA, which reduces yield. Excessive Mg2+ can also stabilize spurious annealing of primer to incorrect template sites, thereby decreasing specificity. On the other hand, an inadequate Mg2+ concentration reduces the amount of product.
[0182]A good balance between dNTP and MgCl2 is approximately 200 to 400 μM dNTP (of each) to 1.5 to 3 mM MgCl2.
[0183]e. PCR Buffer Concentration
[0184]Generally KCl based buffers suffice for multiplex overlap-extension PCR; however, buffers based on other components such as (NH4)2SO4, MgSO4, Tris-HCl, or combinations thereof may also be optimized to function with the multiplex overlap-extension PCR. Primer pairs involved in the amplification of longer products work better at lower salt concentrations (e.g. 20 to 50 mM KCl), whereas primer pairs involved in the amplification of short products work better at higher salt concentrations (e.g. 80 to 100 mM KCl). Raising the buffer concentration to 2× instead of 1× may improve the efficiency of the multiplex reaction.
[0185]f. DNA Polymerase
[0186]The present invention is exemplified with Taq polymerase. Alternatively, other types of heat-resistant DNA polymerases including, for example, Pfu, Phusion, Pwo, Tgo, Tth, Vent, Deep-vent may be used. Polymerases without or with 3' to 5' exonuclease activity may either be used alone or in combination with each other.
Vectors and Libraries
[0187]The linkage of nucleotide sequences of interest according to the present invention produces a nucleotide segment comprising the linked nucleotide sequences of interest. Further, libraries of such linked nucleic acid sequences of interest are produced by the methods of the present invention, in particular libraries of variable region encoding sequences.
[0188]One feature of the present invention is the insertion of a segment containing linked nucleotide sequences of interest or a library of linked nucleotide sequences of interest, generated by a method of the present invention, into suitable vectors. The libraries may be combinatorial libraries or libraries of cognate pairs of variable region encoding sequences. The restriction sites generated by the outer primers, nested primers or semi-nested primers are preferably designed to match appropriate restriction sites of the vector of choice. The linked nucleic acid sequences of interest can also be inserted into vectors by recombination, if one of the semi-nested, nested primers or outer primers were equipped with a suitable recombination site and the vector of choice contains one as well.
[0189]Basically there are no limitations to the vectors that can be used as carriers of the products generated by one of the multiplex RT-PCR-linkage methods of the present invention. Vectors of choice may be those suitable for amplification and expression in cells including, for example, bacteria, yeast, other fungi, insect cells, plant cells, or mammalian cells. Such vectors may be used to facilitate further cloning steps, shuttling between vector systems, display of the product inserted into the vector, expression of the inserted product and/or integrate into the genome of a host cell.
[0190]Cloning and shuttle vectors are preferably bacterial vectors. However, the other types of vectors may also be applied in cloning and shuttle procedures.
[0191]Display vectors can for example be phage vectors or phagemid vectors originating from the class of fd, M13, or f1 filamentous bacteriophages. Such vectors are capable of facilitating the display of a protein including, for example, a binding protein or a fragment thereof, on the surface of a filamentous bacteriophage. Display vectors suitable for display on ribosomes, DNA, yeast cells or mammalian cells are also known in the art. These comprise for example viral vectors or vectors encoding for chimeric proteins.
[0192]Expression vectors exist for all the mentioned species and the one to be chosen completely depend on the protein to be expressed. Some expression vectors are additionally capable of integrating into the genome of a host cell either by random integration, or by site-specific integration, utilizing appropriate recombination sites. Expression vectors may be designed to provide additional encoding sequences that, when the linked product is inserted in-frame to these sequences, enable the expression of a larger protein, e.g. a full-length monoclonal antibody, when introduced into an appropriate host cell. This in-frame insertion may also facilitate the expression of chimeric proteins that facilitate the display on the surface of a filamentous bacteriophage or cell. In a bacteriophage display system, the linked nucleotide sequences of interest may be inserted in-frame to a sequence encoding a coat protein such as pIII or pVIII (Barbas, C. F. et al. 1991. Proc. Natl. Acad. Sci. USA 88, 7978-7982; Kang, A. S. et al. 1991. Proc. Natl. Acad. Sci. USA 88, 4363-4366).
[0193]In one embodiment of the present invention, the individual segments of linked nucleotide sequences of interest is comprised of an immunoglobulin heavy chain variable region encoding sequence associated with a light chain variable region encoding sequence. Preferably, these linked sequences are inserted into a vector that contains sequences encoding one or more immunoglobulin constant domains. The insertion is engineered such that the linked heavy chain variable region and/or light chain variable region encoding sequences are inserted in-frame to the constant region encoding sequences. Such an insertion can for example generate a Fab expression vector, a full-length antibody expression vector or an expression vector encoding a fragment of a full-length antibody. Preferentially such a vector is an expression vector suitable for screening (e.g. E. coli, phagemid, or mammalian vectors) and the constant region heavy chain encoding sequences are chosen from the human immunoglobulin classes IgG1, IgG2, IgG3, IgG4, IgM, IgA1, IgA2, IgD, or IgE, thereby enabling the expression of a Fab or full-length recombinant antibody. In addition to the constant heavy chain encoding sequences the vector may also contain a constant light chain encoding sequence chosen from human lambda or kappa chains. This is appropriate when the linked nucleotide sequences only encode the immunoglobulin variable region encoding sequences (Fv's).
[0194]In another embodiment of the present invention, the individual segments of the linked nucleotide sequences is comprised of a TcR α chain variable region encoding sequence associated with a β chain variable region encoding sequence or a γ chain variable region encoding sequence associated with a δ chain variable region encoding sequence. Preferably, these linked sequences are inserted into a vector that contains sequences encoding one or more TcR constant domains. The insertion is engineered such that the inserted linked variable region encoding sequences are in-frame to the corresponding TcR constant region encoding sequences. In a further embodiment, such a vector is a chimeric expression vector comprising sequences that encode a leucine zipper in-frame to the TcR constant regions. It has been shown that such constructs increase stability of soluble TcR's (Willcox, B. E. et al. 1999. Protein Sci 8, 2418-2423).
[0195]Libraries of cognate pairs of the present invention may be introduced into vectors by two different approaches. In the first approach, the single cognate pairs are inserted individually into a suitable vector. This library of vectors may then either be kept separate or be pooled. In the second approach, all the cognate pairs are pooled prior to vector insertion, followed by in-mass insertion into suitable vectors generating a pooled library of vectors (illustrated in FIG. 12). Such a library of vectors comprises a large diversity of pairs of variable region encoding sequences.
[0196]One aspect of the present invention is a library of cognate pairs of linked variable region encoding sequences. Preferably the individual cognate pairs of the library comprise an immunoglobulin light chain variable region encoding sequence associated with a heavy chain variable region encoding sequence.
[0197]Another preferred library of cognate pairs comprise linked TcR region encoding sequences, where each individual TcR region encoding sequences comprise an alpha chain variable region encoding sequence associated with a beta chain variable region encoding sequence and/or a TcR gamma chain variable region encoding sequence associated with a delta chain variable region encoding sequence.
[0198]An embodiment of the present invention is a sub-library of cognate pairs of linked variable region encoding sequences which encode for desired binding specificities directed against a particular target. Preferably these cognate pairs comprise linked immunoglobulin light chain variable region and heavy chain variable region encoding sequences, TcR alpha chain variable region and beta chain variable region encoding sequences and/or TcR gamma chain variable region and delta chain variable region encoding sequences.
[0199]A further embodiment is a sub-library selected from a parent library of cognate pairs of variable region encoding sequences as described throughout the invention.
[0200]A preferred embodiment of the present invention is a library or sub-library encoding for cognate pairs of full-length immunoglobulins selected from human immunoglobulin classes IgA1, IgA2, IgD, IgE, IgG1, IgG2, IgG3, IgG4, or IgM.
[0201]Another preferred feature of the present invention is a library or sub-library encoding for soluble and stable cognate pairs of TcRs.
[0202]A feature of the present invention is the diversity of said libraries, which are comprised of at least 5, 10, 20, 50, 100, 1000, 104, 105 or 106 different cognate pairs.
[0203]In a further embodiment of the present invention, said libraries of cognate pairs of linked variable region encoding sequences are obtainable by a method comprising the steps described herein. This library is also termed the parent library.
Screening and Selection
[0204]The parent library of pairs of linked variable region encoding sequences isolated from a donor, utilizing one of the methods of the present invention, is expected to represent a diversity of binding proteins of which some will be irrelevant, i.e. not binding to a desired target, in particular for combinatorial libraries. Therefore, the present invention encompasses enrichment and screening, for a sub-library encoding a subset of diversities of binding specificities directed against a particular target.
[0205]For libraries of cognate pairs the diversity of the library is expected to represent the diversity present in the donor material, with only a minor number of randomly linked variable regions. Thus, an enrichment step may not be necessary prior to the screening for target-specific binding affinities in a library composed of cognate pairs.
[0206]In a further embodiment of the present invention, the method of generating a library of pairs of linked variable region encoding sequences, further comprises creating a sub-library by selecting a subset of pairs of linked variable region sequences that encode binding proteins with a desired target specificity. Such a selection of linked variable region encoding sequences, is also termed a library of target-specific cognate pairs.
[0207]In a preferred embodiment of the present invention, is the library of target-specific cognate pairs of variable region encoding sequences transferred to a mammalian expression vector.
[0208]Immunological assays are generally suitable for the selection of target-specific immunoglobulin variable region encoding sequences. Such assays are well know in the art and constitute for example ELISPOTS, ELISA, membrane assays (e.g. Western blots), arrays on filters, or FACS. The assays can either be performed in a direct manner, utilizing the polypeptides produced from the immunoglobulin variable region encoding sequences. Alternatively, the immunoassays can be performed in combination with or following enrichment methods such as phage display, ribosome display, bacterial surface display, yeast display, eukaryotic virus display, RNA display or covalent display (reviewed in FitzGerald, K., 2000. Drug Discov. Today 5, 253-258). As illustrated in FIG. 10, both cognate Fab expression libraries and cognate full-length antibody expression libraries can be subjected to screening, thereby generating a sub-library of positive clones. Such screening assays and enrichment procedures are also suitable for Fv or scFv fragments or combinatorial libraries of linked variable regions.
[0209]In a preferred embodiment of the present invention, the selection of a sub-library of target-specific cognate pairs or combinatorial pairs of variable region encoding sequences is performed by using a high-throughput screening assay. High-throughput screening assays could be, but are not restricted to, ELISA assays performed with semi-automated or fully automated equipment. It could also be a membrane assays in which bacteria are robotically picked and gridded onto an appropriate membrane on top of agar plates generating arrays of colonies expressing antigen-binding molecules. The molecules are secreted through the membrane onto a second underlying antigen-coated membrane which can be developed separately and used to identify clones that secrete antigen binding molecules towards the desired target (de Wildt, R. M., et al. 2000. Nat. Biotechnol. 18, 989-994).
[0210]When a sub-library of cognate pairs or combinatorial pairs of antigen-binding clones has been selected by an appropriate technology it is possible to perform an additional analysis by DNA sequencing of the linked immunoglobulin light chain variable region and heavy chain variable region encoding sequences. First of all such a DNA sequencing will provide information about the library diversity such as germline origin, family distribution and maturation within the CDR regions. Such an analysis will enable the selection of clones which represent a broad diversity, and leaving out repeated clones. Secondly, DNA sequencing will reveal mutations introduced during the isolation process.
[0211]When analyzing variable region encoding sequences there are three types of mutations to consider when assessing whether a mutation is acceptable: i) The most frequent type of mutations result from intra-family cross-priming, where V gene primers prime to the wrong subset within one particular V gene family. The changes introduced are mainly substitutions of naturally occurring codons at one particular position. Due to the high degree of sequence homology within a V gene family these changes usually can be regarded as conservative and acceptable changes; ii) Less frequent mutations are induced by inter-family cross-priming (e.g., a VH3 family primer primes a VH1 family encoding sequence) and induces more significant structural changes, sometimes with no natural counterpart. Such changes could potentially affect the immunogenicity of the variable region by creating new epitopes. Such changes can easily be identified and subsequently repaired using standard molecular biological techniques or the clones can be excluded from the library; iii) Errors created by the Taq DNA polymerase are most easily identified in the constant region encoding sequences and can easily be eliminated. However, Taq induced mutations will of course also be present in the variable region encoding sequences where they are indistinguishable from the naturally occurring somatic mutations, which are also the result of random mutations in the variable region encoding sequences. Considering that the mutations are non-systematic and only affect particular pairs in distinct ways, it appears reasonable to disregard such changes.
[0212]Further the sequence analysis can be used to identify the degree of scrambling in a cognate pair library, as illustrated in table 20 for VH group H4.
[0213]As described in Example 9 the presence of the mutations described in i) and ii) can be circumvented in the expression library, when utilizing primers that anneal in the leader sequence of the variable region encoding sequences instead of primers that anneal in the 5' region of the variable regions.
[0214]In a further embodiment of the present invention, the sub-library of target-specific and possibly sequence analysed pairs of linked immunoglobulin light chain variable region and heavy chain variable region encoding sequences are transferred to a mammalian expression vector. Such a transfer can be performed into any of the vectors described in the previous section, enabling the expression of a full-length recombinant antibody. If the screening is performed with a mammalian cognate full-length antibody expression library such a transfer may not be needed.
[0215]In another embodiment of the present invention, the parent library is generated from a lymphocyte-containing cell fraction which is enriched for T lymphocytes. The pairs of linked variable region encoding sequences constituting the parent library, may be selected for encoding a subset of pairs of linked variable region sequences, composed of alpha and beta and/or gamma and delta chains that encode binding proteins with a desired target specificity, generating a sub-library of cognate pairs or combinatorial pairs. Antigen-specific T cell receptors can subsequently be identified from a pool of transfected cells using standard methodology such as staining with tetrameric MHC-peptide complexes (e.g., Callan, M. F. et al. 1998. J. Exp. Med. 187, 1395-1402; Novak, E. J. et al. 1999. J. Clin. Invest 104, R63-R67), by measuring cellular responses in the form of IL-2 release or by more sophisticated means such as yeast or retroviral display techniques.
Host Cells and Expression
[0216]The libraries of the present invention can be transferred to vectors suitable for expression and production of proteins encoded from the linked nucleic acid sequences of interest, in particular variable region containing binding proteins or fragments thereof. Such vectors are described in the Vectors and Libraries section, and provide for the expression of for example full-length antibodies, Fab fragments, Fv fragments, scFv, membrane bound or soluble TcRs or TcR fragments of a species of choice.
[0217]One feature of the present invention is the introduction into a host cell of a library or a sub-library of vectors of cognate pairs of linked variable region encoding sequences or a single clone encoding a cognate pair of linked variable region encoding sequences, for amplification and/or expression. Host cells can be chosen from bacteria, yeast, other fungi, insect cells, plant cells, or mammalian cells. For expression purposes mammalian cells, such as Chinese hamster ovary (CHO) cells, COS cells, BHK cells, myeloma cells (e.g., Sp2/0 cells, NS0), NIH 3T3, fibroblast or immortalized human cells such as HeLa cells, HEK 293 cells, or PER.C6 are preferred.
[0218]The introduction of vectors into host cells may be accomplished by a number of transformation or transfection methods known to those skilled in the art, including calcium phosphate precipitation, electroporation, microinjection, liposome fusion, RBC ghost fusion, protoplast fusion, viral infection and the like. The production of monoclonal full-length antibodies, Fab fragments, Fv fragments and scFv fragments is well known.
[0219]The production of recombinant polyclonal antibodies to be used for treatment is a quite new area. A recombinant polyclonal manufacturing technology has been described in PCT application WO 2004/061104. In brief, this technology involves the generation of a collection of cells, suitable as a manufacturing cell line. The following description of the technique is made for a library of cognate pairs, it is however just as applicable for a combinatorial library. The individual cells in the collection of cells are capable of expressing a distinct member of the recombinant polyclonal binding protein for example from a library of cognate pairs. In order to ensure that the individual cells express a single cognate pair and not several cognate pairs of the polyclonal binding protein, the nucleic acid sequences encoding the cognate pairs are introduced into a single site-specific site in the genome of each individual cell. This is an important feature of the collection of cells, since this prevent scrambling of the heavy and light chains expressed from each cell, but also because it generates cells that are virtually identical to one another, except for the small differences in the variable regions of the individual cognate pairs. This trait will enable an unbiased growth of the collection of cells over the period of time necessary for the production. To ensure single site-specific integration, a host cell line with only one integration site should be used, these are commercially available e.g. Invitrogen's CHO Flp-In cells containing a single FRT site. Appropriate vectors for this cell line contain a corresponding FRT site and are introduced into the genome using the Flp recombinase. There are several other known recombinases e.g. Cre, beta-recombinase, Gin, Pin, PinB, PinD, R/RS, lambda integrase, or phage ΦC31 integrase that can be used in combination with their corresponding recombination sites. Further, appropriate vectors contain a selection marker that enables the selection of site-specific integrants.
[0220]The generation of a polyclonal manufacturing cell line and the production of a recombinant polyclonal protein from such a cell line can be obtained by several different transfection and manufacturing strategies.
[0221]One way, is to use a library of vectors mixed together into a single composition, for the transfection of a host cell line with a single integration site per cell. This method is termed bulk transfection or transfection in bulk. Generally, the vector and host cell design previously described will ensure that a polyclonal cell line capable of unbiased growth will be obtained upon appropriate selection. A frozen stock of the polyclonal cell line will be generated before initiation of the recombinant polyclonal protein manufacturing.
[0222]Another way, is to use a library of vectors split into fractions, containing approximately 5 to 50 individual vectors of the library in a composition, for transfection.
[0223]Preferably, a fraction of the library constitutes 10 to 20 individual vectors. Each composition is then transfected into an aliquot of host cells. This method is termed semi-bulk transfection. The number of aliquots transfected will depend on the size of the library and the number of individual vectors in each fraction. If the library for example constitutes 100 distinct cognate pairs, which are split into fractions containing 20 distinct members in a composition, 5 aliquots of host cells would need to be transfected with a library composition constituting a distinct fraction of the original library. The aliquots of host cells are selected for site-specific integration. Preferably, the distinct aliquots are selected separately. However, they can also be pooled before selection. The aliquots can be analyzed for their clonal diversity and only those with sufficient diversity will be used to generate a polyclonal cognate pair library stock. To obtain the desired polyclonal cell line for manufacturing, the aliquots can be mixed before generating the freezing stock, immediately after they have been retrieved from the stock or after a short proliferation and adaptation time. Optionally, the aliquots of cells are kept separate throughout production, and the polyclonal protein composition is assembled by combining the products of each aliquot rather than the aliquots of cells before production.
[0224]A third way, is a high throughput method in which host cells are transfected separately using the individual vectors constituting the library of cognate pairs. This method is termed individual transfection. The individually transfected host cells are preferably selected for site specific integration separately. The individual cell clones generated upon selection may be analyzed with respect to proliferation time and preferably, those with similar growth rates are used to generate a polyclonal cognate pair library stock. The individual cell clones can be mixed to obtain the desired polyclonal cell line before generating the stock, immediately after they have been retrieved from the stock, or after a short proliferation and adaptation time. This approach may eliminate any possible residual sequence bias during transfection, integration and selection. Alternatively the individually transfected host cells are mixed before selection is performed, this will enable control of sequence bias due to transfection.
[0225]A shared feature in the manufacturing strategies outlined in the above is that all the individual cognate pairs constituting the recombinant polyclonal protein can be produced in one, or a limited number of bioreactors. The only difference is the stage at which one chooses to generate the collection of cells that constitutes the polyclonal manufacturing cell line.
[0226]One embodiment of the present invention, is a population of host cells comprising a cognate library or sub-library of linked pairs of variable region encoding sequences.
[0227]In a further embodiment, a population of host cells comprises a library obtained from a population of isolated single cells constituting lymphocytes, utilizing the multiplex RT-PCR amplification followed by linkage by ligation or recombination or the multiplex overlap-extension RT-PCR technology of the present invention, to link the cognate pairs.
[0228]Another embodiment of the present invention, is a population of host cells comprising a combinatorial library or sub-library of linked pairs of variable region encoding sequences.
[0229]A population of host cells according to the present invention, will encompass a diverse population of cells corresponding to the diversity of the library the cells have been transformed/transfected with. Preferably, each cell of the population of cells only constitutes one cognate pair of the entire library of cognate pairs, and no individual member of the library of cognate pairs exceeds more than 50%, more preferred 25%, or most preferred 10%, of the total number of individual members expressed from the population of host cells.
[0230]In a preferred embodiment of the present invention, the population of host cells is mammalian cells.
[0231]A population of host cells as described in the above can be utilized for the expression of a recombinant polyclonal binding protein, since individual cells of the population constitute variable region encoding sequences of different diversity.
[0232]One embodiment of the present invention, is a recombinant polyclonal protein expressed from a population of host cells comprising a library of vectors encoding diverse cognate pairs of linked variable region encoding sequences, where such a library is obtainable by the method of the present invention. Typically, a recombinant polyclonal protein of the present invention is comprised of at least 2, 5, 10, 20, 50, 100, 1000, 104, 105 or 106 proteins composed of different cognate pairs.
[0233]A preferred embodiment of the present invention, is a recombinant polyclonal immunoglobulin expressed from a population of host cells comprising a library of vectors encoding diverse cognate pairs of heavy chain variable region and light chain variable region encoding sequences.
[0234]Another preferred embodiment of the present invention, is a recombinant polyclonal TcR expressed from a population of host cells comprising a library of vectors encoding diverse cognate pairs of TcR alpha chain variable region linked with beta chain variable region encoding sequences and/or TcR gamma chain variable region linked with delta chain variable region encoding sequences.
[0235]Another embodiment of the present invention is a host cell suitable for production of a monoclonal protein. In particular a monoclonal antibody comprised of a cognate pair of a light chain variable region with a heavy chain variable region or a monoclonal TcR comprised of a cognate pair of an alpha variable region with a beta variable region or a delta variable region with a gamma variable region. Preferably such a monoclonal production cell line is not a hybridoma cell line.
[0236]Such a monoclonal antibody or TcR can be generated by adding the following steps to the method of linking a plurality of non-contiguous nucleotide sequences of interest a) inserting said linked nucleic acid sequences into a vector; b) introducing said vector into a host cell; c) cultivating said host cells under conditions suitable for expression; and d) obtaining the protein product expressed from the vector inserted into said host cell. Preferably, the vector introduced into the host cell encodes an individual cognate pair of variable region encoding sequences.
Applications of the Invention
[0237]One of the major applications of the present invention is the linkage of cognate pairs of variable region encoding sequences, especially immunoglobulin heavy and light chain variable region encoding sequences or TcR alpha and beta chain or gamma and delta chain variable region encoding sequences, by a high-throughput method for the generation of libraries of cognate pairs. In addition to the generation of cognate pair libraries, the multiplex RT-PCR followed by linkage by ligation or recombination or the multiplex overlap-extension RT-PCR techniques of the present invention may be utilized in the generation of combinatorial libraries by performing the technique on a population of genetically diverse cells, cell lysates from such a population of cells, or on RNA purified from such a population of cells. The libraries, sub-libraries, or single clones from one of these libraries facilitate the expression of polyclonal or monoclonal proteins. Especially monoclonal or polyclonal antibodies may be obtained from the libraries of the present invention.
[0238]The use of recombinant monoclonal antibodies in diagnostics, treatment, and prophylaxis is well known. Recombinant monoclonal and polyclonal antibodies generated by the present invention will have the same applications as antibody products generated by existing technologies. In particular, a pharmaceutical composition comprising a polyclonal recombinant immunoglobulin as active ingredient, combined with at least one pharmaceutically acceptable excipient, can be produced by means of the present invention. More preferred are pharmaceutical compositions where the polyclonal recombinant immunoglobulin is comprised of cognate pairs of variable region encoding sequences. Such pharmaceutical compositions of polyclonal recombinant immunoglobulins can be used as medicaments. The polyclonal recombinant immunoglobulin of the composition can be specific for or reactive against a predetermined disease target, and the composition can thus be used for the treatment, amelioration or prevention of diseases such as cancer, infections, inflammatory diseases, allergy, asthma and other respiratory diseases, autoimmune diseases, immunological malfunctions, cardiovascular diseases, diseases in the central nervous system, metabolic and endocrine diseases, transplant rejection, or undesired pregnancy, in a mammal such as a human, a domestic animal, or a pet.
[0239]The present invention has a further application which cannot be obtained with conventional monoclonal combinatorial antibody techniques. In situations where protective antigens are either poorly characterized or completely unknown, such as in emerging infectious diseases, it is impossible to screen for a monoclonal antibody that will provide protection against the disease.
[0240]However, with the present invention it will be possible to obtain antibody-expressing cells directly from donors with an established protective antibody response, e.g. convalescent patients, and use the starting material from these individuals to generate a library of cognate pairs of immunoglobulin heavy and light chain variable region encoding sequences.
[0241]In situations where for example the virus is known, but the protective antigens are unknown, it will be possible to generate a sub-library of cognate antibody gene pairs with broad reactivity towards antigenic structures on the virus. If a recombinant polyclonal antibody is produced from such a sub-library, it will likely contain protective antibodies.
[0242]In situations where antigens are completely unknown, a recombinant polyclonal antibody generated from a cognate pair library from e.g. convalescent patients can be used in the same way as hyperimmune immunoglobulins are being used today. The reason for this is the cognate pairing which ensures that the recombinant polyclonal antibody produced closely resemble the antibody immune response of the convalescent patient.
[0243]Another application of the techniques for linking cognate pairs of variable regions, described in the present invention, is for diagnostic and analytical purposes. When administering a medicament to a patient, the possibility of an immune response directed towards the medicament always exists. This immunogenicity can be assessed by conventional techniques, such as medicament-specific binding assays with serum or plasma derived from the individual treated with the medicament. Alternatively, the methods of the present invention can be used to mirror the patients' immune response shortly after administration of the medicament, by isolating cognate pairs of variable heavy chain and light chain encoding sequences. Antibodies expressed from such a library can then be screened for reactivity towards the medicament or components of the medicament. This method is particular useful if the presence of the medicament in plasma or serum would interfere with the conventional method. Such medicaments are for example antibodies. With conventional methods, identification of an anti-medicament immune response (e.g. an anti-idiotypic, anti-framework or anti-Fc immune response) in relation to treatment with an antibody cannot be performed until the antibody used for treatment has been cleared completely from the blood. With the present invention sequences encoding such anti-medicament antibodies can be isolated and analyzed from the individual treated with the medicament within a couple of weeks. Thus, it would be an alternative method for assessing whether drugs in general and in particular antibody-based drugs are immunogenic.
[0244]A further application of the present invention is in the validation and comparison of vaccines and immunization programmes. This is particular useful during vaccine development, since it will be possible to assess and compare the sequence diversity of antibody responses generated in response to novel candidate vaccines, in addition to the current comparisons of antibody binding affinities and serum titer. Further, the present invention can be used to analyze, monitor and compare antibody responses in surveillance of vaccine efficacy in populations.
[0245]The present invention also finds applications outside the field of variable region containing binding proteins. It is merely a matter of optimization for one skilled in the art to adapt the technique disclosed in the present invention to link two or more transcribed nucleotide sequences encoding a heteromeric protein other that a binding protein. Such a linkage of sequences encoding for domains or subunits from a heteromeric protein may be an advantage in the isolation of these protein encoding sequences since it would reduce the number of steps considerably. Further, it is possible to isolate for example splice variants, mutations or new family members of such proteins by using only one multiplex primer mix/multiplex overlap-extension primer mix.
[0246]The linkage of sequences which encode distant domains of a single protein is also a possibility. Such a technique would ease the research in relation to the importance of certain domains in a multidomain protein, since it would ease the deletion of intermediate domains.
[0247]The generation of chimeric or coupled proteins is also an area where the present invention may be applied. Even if the proteins to be linked are of different origin, e.g. a chimera between a human and a mouse protein, the present invention may be utilized by mixing the cells prior to reverse transcription. Such a cell mixture may constitute either a population of cells from each species or a single cell from each species.
EXAMPLES
Example 1
Two-Step Multiplex Overlap-Extension RT-PCR
[0248]In this example, reverse transcription (RT) was performed using a template derived from an isolated single cell and the cDNA produced was used as template for more than one multiplex overlap-extension PCR.
[0249]a. Cells
[0250]An IgG1-kappa expressing Chinese hamster ovary (CHO) cell line was generated using the Flp-In technology (Invitrogen, Carlsbad, Calif., USA).
[0251]An IgG-kappa mammalian expressing plasmid vector pLL113 (FIG. 4) was constructed based on the Flp-In expression vector, pcDNA5/FRT. CHO-Flp-In cells were co-transfected, using Lipofectamin2000 (Invitrogen, Carlsbad, Calif., USA) according to the manufacturer's instructions, with pLL113 containing the antibody encoding genes and pOG44 conferring transient expression of the Flp recombinase. Transformants were selected and verified for production of IgG-kappa by immunoassays. The selected cell line was named CHO Flp-In pLL113, and maintained in Ham's F-12 medium, supplemented with 2 mM L-glutamine, 10% FCS and 900 Hygromycin B (maintenance media). CHO Flp-In pLL113 cells were harvested using trypsin and washed 3× in maintenance media. After the final wash the cells were re-suspended in the original volume of maintenance media. The cell concentration was determined using a Casy-1 System (Scharfe System GmbH, Reutlingen, Germany) and diluted in maintenance media to a concentration of 1 cell/5 μl.
[0252]b. Reverse Transcription
[0253]CHO Flp-In pLL113 cell suspension, on average containing one cell, was dispensed into single wells of a 96-well PCR plate (Thermo-Fast 96, skirted AB-0800, ABgene, Epsom, Surrey, UK).
[0254]The cDNA was synthesized from the distributed cells by utilizing the reverse transcriptase (RT) step in the Qiagen One-Step RT-PCR protocol (Qiagen OneStep RT-PCR Kit, Cat# 210210, Hilden, Germany).
[0255]Each well contained the following reagents in a total volume of 20 [0256]1× One Step RT-PCR buffer, [0257]dNTP's in a final concentration of 1 mM of each, [0258]5 pmol Oligo poly-dT (18), [0259]26 U RNase inhibitor (RNasin, Promega, Madison USA, cat. No N2111), [0260]2 μl One Step RT-PCR enzyme mix, and [0261]5 μl CHO Flp-In pLL113 cell suspension.
[0262]The RT reaction was performed by incubating the reaction mixes at 55° C. for 30 min. Subsequently the reverse transcriptase was inactivated by incubating the reaction mixture at to 94° C. for 10 min.
[0263]c. Multiplex Overlap-Extension PCR
[0264]A fraction of the cDNA products generated in step b) was used as template for multiplex overlap-extension PCR. The reactions were performed in 96-well plates.
[0265]Each well contained, in a total volume of 40 μA, the following reagents: [0266]1× One Step RT-PCR buffer, [0267]dNTP's in a final concentration of 500 μM of each, [0268]Multiplex overlap-extension primer mix in the concentrations as indicated in Table 1, [0269]26 U RNase inhibitor (RNasin, Promega, Madison USA, cat. No N2111), [0270]2 μl One Step RT-PCR enzyme mix, and [0271]1 μl cDNA template (derived from a single cell (step b)).
[0272]The multiplex overlap-extension primer mix used comprised the primers shown in table 1.
TABLE-US-00003 TABLE 1 Primer SEQ ID name Conc. NO primer sequence in 5' to 3' direction VH 20 nM of 1 agcctatactattgattaggcgcgccCAGRTGCAGCTGGTGCART each 2 agcctatactattgattaggcgcgccSAGGTCCAGCTGGTRCAGT 3 agcctatactattgattaggcgcgccCAGRTCACCTTGAAGGAGT 4 agcctatactattgattaggcgcgccSAGGTGCAGCTGGTGGAG 5 agcctatactattgattaggcgcgccCAGGTGCAGCTACAGCAGT 6 agcctatactattgattaggcgcgccCAGSTGCAGCTGCAGGAGT 7 agcctatactattgattaggcgcgccGARGTGCAGCTGGTGCAGT 8 agcctatactattgattaggcgcgccCAGGTACAGCTGCAGCAGTC CH 100 nM 9 GACSGATGGGCCCTTGGTGG VL 20 nM of 10 cgcctaatcaatagtataggctagccGACATCCAGWTGACCCAGTCT each 11 cgcctaatcaatagtataggctagccGATGTTGTGATGACTCAGTCT 12 cgcctaatcaatagtataggctagccGAAATTGTGWTGACRCAGTCT 13 cgcctaatcaatagtataggctagccGATATTGTGATGACCCACACT 14 cgcctaatcaatagtataggctagccGAAACGACACTCACGCAGT 15 cgcctaatcaatagtataggctagccGAAATTGTGCTGACTCAGTCT CLκ 100 nM 16 atatatatgcggccgcttaTTAACACTCTCCCCTGTTG W = A/T, S = G/C, R = A/G. Capitalized sequences correspond to the gene-specific region.
[0273]The reactions were performed with a 96-well MWG Primus HT thermocycler (MWG Biotech AG, Ebersberg, Germany) with the following cycling conditions:
TABLE-US-00004 Denature 30 sec 95° C. Anneal 30 sec 50° C. {close oversize brace} 50 cycles Extend 5 min 72° C. Final extension 10 min 72° C.
[0274]d. Nested PCR
[0275]Nested PCR was performed using the multiplex overlap-extension products as template. The reactions were performed in 96-well plates.
[0276]Each well contained, in a total volume of 50 μl, the following reagents: [0277]1× BioTaq buffer, [0278]dNTP's in a final concentration of 400 μM of each, [0279]2 mM MgCl2, [0280]Nested PCR primer mix, [0281]1.25 U BIOTAQ DNA Polymerase (Cat. No BIO-21040, Bioline, UK.), and [0282]1 μl Multiplex overlap-extension PCR product (step c).
[0283]The primers used are shown in table 2.
TABLE-US-00005 TABLE 2 Primer SEQ primer sequence in name Conc. ID NO 5' to 3' direction JH 100 nM 17 atattctcGAGACGGTGACCAGGGTG of each 18 atattctcGAGACGGTGACCATTGT 19 atattctcGAGACGGTGACCAGGGTTC 20 atattctcGAGACGGTGACCGTGGT CLκ 100 nM 21 accgcctccaccggcggccgctta TTAACACTCTCCCCTGTTGAAGCTCTT Capitalized sequences correspond to the gene-specific region.
[0284]The reactions were performed with a 96-well MWG Primus HT thermocycler (MWG Biotech AG, Ebersberg, Germany) with the following cycling conditions:
TABLE-US-00006 Denature 30 sec 95° C. Anneal 30 sec 50° C. {close oversize brace} 25 cycles Extend 90 sec 72° C. Final extension 10 min 72° C.
[0285]The nested PCR products were analyzed by 1% agarose gel electrophoresis using to ethidium bromide for detection (FIG. 5). The expected size of the overlap-extension product was 1076 bp. Such a product can be observed in lanes 1, 5, 6, 7, 8 and 12 indicated by the arrows (FIG. 5A). In the experiment shown, the negative control is contaminated and a similar contamination is also present in the samples. FIG. 5B illustrates the fragments of FIG. 5A, which are relevant to the present experiment.
[0286]The present experiment illustrates one way to perforin two-step multiplex overlap-extension PCR, utilizing template derived from a single cell. Further, it was shown that it is possible to perform approximately twenty multiplex overlap-extension PCR's using cDNA generated from an isolated single cell.
Example 2
Single-Step Multiplex Overlap-Extension RT-PCR
[0287]In this example reverse transcription and multiplex overlap-extension PCR was performed in a single step using template from lysed cells, in concentrations corresponding to 100, 10, or 1 cells.
[0288]a. Cells
[0289]The human hybridoma cell line HB-8501 producing an anti-tetanus IgG1-kappa antibody was acquired from American Type Culture Collection and cultured in Iscove's Modified Dulbecco's Medium (Vitacell, Kiev, Ukraine, cat. No 30-2005) containing 10% fetal bovine serum. Before multiplex overlap-extension RT-PCR was performed, the cells were harvested, counted and frozen at -80° C. in culture medium in a concentration of 200 cells/μl.
[0290]b. Single-Step Multiplex Overlap-Extension RT-PCR
[0291]The Qiagen One-Step RT-PCR kit (Qiagen cat. No 210212, Hilden, Germany) was used for the multiplex overlap-extension RT-PCR essentially according to the manufacturer's recommendation. Before addition to the PCR tubes cell lysates were thawed and diluted in H2O to yield a lysate concentration corresponding to 100, 10, and 1 cells per 5 μl.
[0292]Each PCR tube contained the following reagents in a total volume of 50 μl: [0293]1× One-Step RT-PCR buffer, [0294]dNTP's in a final concentration of 400 μM of each, [0295]Multiplex overlap-extension primer mix in the concentrations as indicated in table 3, [0296]2 μl One-step RT-PCR enzyme mix, [0297]50 U RNase inhibitor (RNasin, Promega, Madison USA, cat. No N2515), and [0298]5 μl diluted cell lysate
[0299]The multiplex overlap-extension primer mix used comprised the primers shown in table 3.
TABLE-US-00007 TABLE 3 Primer SEQ ID name Conc. NO primer sequence in 5' to 3' direction VH 40 nM 22 gctagcattattattaccatggccCAGRTGCAGCTGGTGCART of each 23 gctagcattattattaccatggccSAGGTCCAGCTGGTRCAGT 24 gctagcattattattaccatggccCAGRTCACCTTGAAGGAGT 25 gctagcattattattaccatggccSAGGTGCAGCTGGTGGAG 26 tattggcgcgccatggccsaggtgCAGCTGGTGGAG 27 gctagcattattattaccatggccCAGGTGCAGCTACAGCAGT 28 gctagcattattattaccatggccCAGSTGCAGCTGCAGGAGT 29 gctagcattattattaccatggccGARGTGCAGCTGGTGCAGT 30 gctagcattattattaccatggccCAGGTACAGCTGCAGCAGTC JH 200 nM 31 atattctcGAGACGGTGACCAGGGTG of each 32 atattctcGAGACGGTGACCATTGTCC 33 atattctcGAGACGGTGACCAGGGTTC 34 atattctcGAGACGGTGACCGTGGTCC VL 40 nM 35 ccatggtaataataatgctagccGACATCCAGWTGACCCAGTCT of each 36 ccatggtaataataatgctagccGATGTTGTGATGACTCAGTCT 37 ccatggtaataataatgctagccGAAATTGTGWTGACRCAGTCT 38 ccatggtaataataatgctagccGATATTGTGATGACCCACACT 39 ccatggtaataataatgctagccGAAACGACACTCACGCAGT 40 ccatggtaataataatgctagccGAAATTGTGCTGACTCAGTCT CLκ 200 nM 16 atatatatgcggccgcttaTTAACACTCTCCCCTGTTG W = A/T, S = G/C, R = A/G. Capitalized sequences correspond to the gene-specific region
[0300]The reactions were performed using the following cycling conditions:
TABLE-US-00008 Reverse transcription: 30 min 55° C. Polymerase activation: 15 min 95° C. inactivating reverse transcriptase and activating Taq polymerase.
[0301]PCR reaction:
TABLE-US-00009 Denature 30 sec 94° C. Anneal 30 sec 44° C. {close oversize brace} 50 cycles Extend 3 min 72° C. Final extension 10 min 72° C.
[0302]Ten microliters of the reaction products were analyzed by 1.5% agarose gel electrophoresis using ethidium bromide for detection (FIG. 6A).
[0303]Expected size of the fragments (the exact size depends upon the lengths of the variable regions):
[0304]VH: 410 bp
[0305]LC: 680 bp
[0306]Overlap-extension fragment: 1070 bp
[0307]Discrete DNA fragments with mobilities corresponding to the heavy chain variable region (VH) and the light chain variable and constant region (LC) lengths are seen for all dilutions of cell lysate. A less intense fragment with mobility corresponding to the putative overlap-extension fragment is seen in lysates from 100 and 10 cells. The overlap-extension fragment of the one cell lysate sample, is difficult to recognize, although it can be seen on the original gel (see arrow in photo shown in FIG. 6A and sketch shown in FIG. 6B).
[0308]c. Identification of the Overlap-Extension Fragment
[0309]From an experiment reproduced as the one described above the presence of the overlap-extension band was verified. Regions in the agarose gel around 1070 bp from a lane corresponding to 1 cell and from a lane corresponding to 100 cells were excised and the DNA purified using Qiaex II (Qiagen cat. No 20051, Hilden, Germany) and eluted in 20 μl of water.
[0310]One microliter of the eluate was subjected to PCR (Biotaq kit, Bioline, UK cat. No BIO-21040) according to the manufacturers instructions with primers flanking the putative overlap-extension fragment (JH primers corresponding to SEQ ID NO 32 to 35 and CLκ primer corresponding to SEQ ID NO: 17, each primer at a concentration of 0.5 μM). Cycling parameters were:
TABLE-US-00010 Denature 30 sec 95° C. Anneal 30 sec 55° C. {close oversize brace} 30 cycles Extend 1 min 72° C.
[0311]Ten microliters of each reaction product was analyzed by 1% agarose gel electrophoresis using ethidium bromide for detection. Several fragments can be seen including, both from 1 and 100 cells, a fragment with mobility of the expected overlap-extension fragment (arrow in FIG. 6C). The 1 kb fragment from the gel lane corresponding to one cell was cut out of the gel and purified using Qiaex II as described above. The purified fragment was digested with the restriction enzymes NheI and NcoI (separately) and the reaction products were analyzed by 1% agarose gel electrophoresis using ethidium bromide for detection (FIG. 6D). These restriction sites are present in the overlap between VH and LC and the expected sizes after digestion are approximately 410 and 680 bp long, respectively (FIG. 2). The NheI digestion was partial since a large fraction is still present at the original size.
Example 3
Combined Single-Step Multiplex Overlap-Extension RT-PCR and Nested PCR
[0312]In this example, reverse transcription and multiplex overlap-extension PCR reactions were performed in a single step followed by a semi-nested PCR amplification, using template from lysed cells, in concentrations corresponding to 100, 10 or 1 cells.
[0313]a. Single-Step Multiplex Overlap-Extension RT-PCR
[0314]Multiplex overlap-extension RT-PCR using HB-8501 cell lysate was performed as described in Example 2 utilizing a multiplex overlap-extension primer mix comprising the primers shown in table 4.
TABLE-US-00011 TABLE 4 Primer SEQ name Conc. ID NO primer sequence in 5' to 3' direction VH 40 nM 41 tattggcgcgccatggccCAGRTGCAGCTGGTGCART of each 42 tattggcgcgccatggccSAGGTCCAGCTGGTRCAGT 43 tattggcgcgccatggccCAGRTCACCTTGAAGGAGT 44 tattggcgcgccatggccSAGGTGCAGCTGGTGGAG 45 tattggcgcgccatggccCAGGTGCAGCTACAGCAG 46 tattggcgcgccatggccCAGSTGCAGCTGCAGGAGT 47 tattggcgcgccatggccGARGTGCAGCTGGTGCAGT 48 tattggcgcgccatggccCAGGTACAGCTGCAGCAGTC CH1-5 200 nM 49 AAGTAGTCCTTGACCAGGCAGCC of one 50 TGAGTTCCACGACACCGTCA of these 51 AGAGGTGCTCTTGGAGGAGG primers 52 AGTTTTGTCACAAGATTTGGG 53 GTTGCAGATGTAGGTCTGGGTGC VL 20 nM 54 gccatggcgcgccaatagctagccGACATCCAGWTGACCCAGTCT of each 55 gccatggcgcgccaatagctagccGATGTTGTGATGACTCAGTCT 56 gccatggcgcgccaatagctagccGAAATTGTGWTGACRCAGTCT 57 gccatggcgcgccaatagctagccGATATTGTGATGACCCACACT 58 gccatggcgcgccaatagctagccGAAACGACACTCACGCAGT 59 gccatggcgcgccaatagctagccGAAATTGTGCTGACTCAGTCT CLκ 200 nM 60 atatatatgcggccgcttaTTAACACTCTCCCCTGTTGAA W = A/T, S = G/C, R = A/G. Capitalized sequences correspond to the gene-specific region.
[0315]It should however be noted that for each multiplex overlap-extension primer mix only one of the CH1-5 primers were used, resulting in five different multiplex overlap-extension primer mixes.
[0316]Remaining parameters were as described for the multiplex overlap-extension RT-PCR reaction in Example 2, only with a change in annealing temperature to 50° C. A reaction was performed for each heavy chain constant region reverse primer (CH1, CH2, CH3, CH4, CH5 corresponding to SEQ ID NOs: 49 to 53, respectively) utilizing lysates corresponding to 100, 10, 1 and 0 cells.
[0317]b. Semi-Nested PCR
[0318]One microliter of the multiplex overlap-extension RT-PCR reaction product was subjected to semi-nested PCR (Biotaq kit, Bioline, UK cat. No BIO-21040), essentially as proposed by the manufacturer. The total volume of the reactions were 50 μl. The primers used are shown in table 5.
TABLE-US-00012 TABLE 5 SEQ Primer ID primer sequence in name Conc. NO 5' to 3' direction JH 200 nM 61 ggaggcgctcGAGACGGTGACCAGGGTGCC of each 62 ggaggcgctcGAGACGGTGACCATTGTCCC 63 ggaggcgctcGAGACGGTGACCAGGGTTCC 64 ggaggcgctcGAGACGGTGACCGTGGTCCC CLκ 200 nM 21 accgcctccaccggcggccgcttaTTAACA CTCTCCCCTGTTGAAGCTCTT Capitalized sequences correspond to the gene-specific region.
[0319]Cycling conditions were as follows:
TABLE-US-00013 Denature 30 sec 95° C. Anneal 30 sec 50° C. {close oversize brace} 25 cycles Extend 1.5 min 72° C. Final extension 5 min 72° C.
[0320]Ten microliters of each reaction was analyzed by 1.5% agarose gel electrophoresis using ethidium bromide for detection (FIG. 7).
[0321]The putative overlap-extension fragment from the semi-nested PCR corresponding to lysate from one cell and where the CH4 primer was used in the first reaction (see arrow in FIG. 7), was excised from the agarose gel, purified using a Qiaex II kit (Qiagen cat. No. 20051, Hilden, Germany) and inserted into pCR2.1-TOPO using an Invitrogen TOPO TA cloning kit (Invitrogen cat. No 45-0641, Carlsbad, Calif., USA). Inserts of eight clones were sequenced and seven of these appeared to consist of a heavy chain variable region linked to a light chain variable and constant region, by the expected overlap region.
[0322]In summary, the sensitivity of the combined multiplex overlap-extension RT-PCR and the semi-nested PCR reaction was very satisfactory, with 4 out of 5 constant region primers capable of amplifying significant amounts of overlap-extension products from lysate corresponding to a single cell.
Example 4
Combined Single-Step Multiplex Overlap-Extension RT-PCR and Nested PCR Using Enriched Human B Lymphocytes as Template Source
[0323]In this example, reverse transcription and multiplex overlap-extension PCR reactions were performed in a single step followed by a semi-nested amplification PCR, using isolated single human B lymphocytes as template source.
[0324]a. B-Cell Isolation
[0325]A human male donor was immunized with Tetanus toxoid. A blood sample of 120 ml was collected from the donor 6 days post immunization and peripheral blood mononuclear cells (PBMC's) were isolated using Lymphoprep (Axis-Shield, Oslo Norway, prod. No 1001967) according to the manufacturer's instructions. The CD 19-positive cell population was enriched utilizing magnetic bead cell sorting. The PBMC's were stained with FITC-conjugated anti-CD19 antibody (Becton Dickinson, N.J., USA, cat. No 345776). Magnetic bead cell sorting using anti-FITC-conjugated magnetic microbeads and column purification was performed according to manufactures instructions (Miltenyi Biotec, Gladbach, Germany, cat. No 130-042-401). The cells were diluted to a concentration of 200 cells per ml in PBS containing 2 nM EDTA and 0.5% BSA. Five microliters of the diluted cells was distributed to PCR tubes obtaining approximately a single cell pr. tube. The tubes were stored at -80° C. until use.
[0326]b. Multiplex Overlap-Extension RT-PCR and Semi-Nested PCR
[0327]Conditions for multiplex overlap-extension RT-PCR and semi-nested PCR were as described in Example 3. However, the reactions were only performed with the multiplex overlap-extension primer mix constituting primer CH3 corresponding to SEQ ID NO: 51. Sixteen samples from the combined multiplex overlap-extension RT-PCR and semi-nested PCR reactions were analyzed by subjecting 10 μl from each semi-nested PCR reaction to 1% agarose gel electrophoresis using ethidium bromide for detection (FIG. 8).
[0328]As can be seen in FIG. 8, 2 out of 16 lanes (lanes 5 and 6) contained fragments of the expected mobility (around 1 kb). Further, lane 2 contained a less intense fragment at the expected mobility. The 1 kb fragments of lane 5 and 6 were excised from the agarose gel, purified using a Qiaex II kit (Qiagen cat. No 20051) and inserted into pCR2.1-TOPO using the Invitrogen TOPO TA cloning kit (Invitrogen cat. No 45-0641, Carlsbad, Calif., USA). Two clones from each isolated fragment had inserts of the correct size. Restriction enzyme digestion (NcoI and NheI, separately) showed fragments of the expected sizes (410 and 680 bp) indicating a correct linkage between the heavy chain variable region and the light chain variable and constant region encoding sequences.
[0329]The two clones originating from the fragment in lane 5 were sequenced and shown to be identical, indicating that the linked VH and LC were cognate pairs.
Example 5
Combined Single-Step Multiplex Overlap-Extension RT-PCR and Nested PCR Using V.sub.λ-Specific Primers
[0330]In this example, reverse transcription and multiplex overlap-extension PCR reactions were performed in a single step using V.sub.λ-specific primers, followed by a semi nested PCR reaction. Total RNA purified from two different cell lines expressing lambda gene families 1b and 1e in combination with the same heavy chain variable region were used as templates.
[0331]a. Cells
[0332]Two IgG1-lambda expressing Chinese hamster ovary (CHO) cell lines were generated using the Flp-In technology (Invitrogen, Carlsbad, Calif., USA).
[0333]IgG1-lamda mammalian expressing vectors pEm465/01P581 and pEm465/01P582 (FIG. 9A) representing two lambda gene families were constructed based on the Flp-In expression vector, pcDNA5/FRT. CHO-Flp-In cells were co-transfected, using Fugene 6 (Roche, Mannheim, Germany) according to the manufacturer's instructions, with the above mentioned plasmids containing the antibody encoding genes and pOG44 conferring transient expression of the Flp recombinase. Transformants were selected for insertions. The selected cell lines were named CHO Flp-In/Em464/01P581 & CHO Flp-In/Em464/01P582, and maintained in Ham's F-12 medium, supplemented with 2 mM L-glutamine, 10% FCS and 900 μg/ml Hygromycin B (maintenance media).
[0334]The cell lines were harvested using trypsin and washed 3× in maintenance media. Approximately 107 cells were used for Total RNA purification using Nucleo Spin L kit (Macherey-Nagel, Duren, Germany) according to the manufacturer's description. The final RNA concentrations were determined by OD260 measurements.
[0335]b. Single-Step Multiplex Overlap-Extension RT-PCR
[0336]Multiplex overlap-extension RT-PCR using 50 pg, 5 pg, or 0.5 pg total RNA as template was essentially performed as described in Example 3 utilizing a multiplex overlap-extension primer mix comprising the primers shown in Table 6 and the cycling conditions specified below.
TABLE-US-00014 Reverse transcription: 30 min 55° C. Polymerase activation: 15 min 95° C. inactivating reverse transcriptase and activating Taq polymerase.
[0337]PCR reaction:
TABLE-US-00015 Denature 30 sec 95° C. Anneal 30 sec 50° C. {close oversize brace} 35 cycles Extend 5 min 72° C. Final extension 10 min 72° C.
TABLE-US-00016 TABLE 6 Primer SEQ ID name Conc. NO primer sequence in 5' to 3' direction VH 40 nM 1 agcctatactattgattaggcgcgccCAGRTGCAGCTGGTGCART of each 2 agcctatactattgattaggcgcgccSAGGTCCAGCTGGTRCAGT 3 agcctatactattgattaggcgcgccCAGRTCACCTTGAAGGAGT 4 agcctatactattgattaggcgcgccSAGGTGCAGCTGGTGGAG 5 agcctatactattgattaggcgcgccCAGGTGCAGCTACAGCAGT 6 agcctatactattgattaggcgcgccCAGSTGCAGCTGCAGGAGT 7 agcctatactattgattaggcgcgccGARGTGCAGCTGGTGCAGT 8 agcctatactattgattaggcgcgccCAGGTACAGCTGCAGCAGTC CH 100 nM 9 GACSGATGGGCCCTTGGTGG V.sub.λ 100 nM 65 cgcctaatcaatagtataggctagccCAGTCTGTGCTGACTCAGCCA of each 66 cgcctaatcaatagtataggctagccCAGTCTGTGYTGACGCAGC 67 cgcctaatcaatagtataggctagccCAGTCTGTCGTGACGCAGC 68 cgcctaatcaatagtataggctagccCARTCTGCCCTGACTCAGCCT 69 cgcctaatcaatagtataggctagccTCCTATGWGCTGACTCAGCCA 70 cgcctaatcaatagtataggctagccTCTTCTGAGCTGACTCAGGAC 71 cgcctaatcaatagtataggctagccCACGTTATACTGACTCAACCG 72 cgcctaatcaatagtataggctagccCAGGCTGTGCTGACTCAGC 73 cgcctaatcaatagtataggctagccAATTTTATGCTGACTCAGCCCCA 74 cgcctaatcaatagtataggctagccCAGRCTGTGGTGACYCAGGA 75 cgcctaatcaatagtataggctagccCWGCCTGTGCTGACTCAGC C.sub.λ2/Cλ7 100 nM 76 gccgcttattaTGAACATTCTGTAGGGGCCA 77 gccgcttattaAGAGCATTCTGCAGGGGCCA Y = C/T, W = A/T, S = G/C, R = A/G. Capitalized sequences correspond to the gene-specific region.
[0338]c. Semi-Nested PCR
[0339]One microliter of the multiplex overlap-extension RT-PCR reaction product was subjected to semi-nested PCR (Biotaq kit, Bioline, UK cat. No BIO-21040), essentially as proposed by the manufacturer. The total volumes of the reactions were 204 The primers used are shown in table 7.
TABLE-US-00017 TABLE 7 SEQ Primer ID primer sequence in name Conc. NO 5' to 3' direction JH 200 nM 61 ggaggcgctcGAGACGGTGACCAGGGTGCC of each 62 ggaggcgctcGAGACGGTGACCATTGTCCC 63 ggaggcgctcGAGACGGTGACCAGGGTTCC 64 ggaggcgctcGAGACGGTGACCGTGGTCCC CLλ 200 nM 78 atatatatgcggccgcttattaTGAACATT CTGTAGGGGCCACTG 79 atatatatgcggccgcttattaAGAGCATT CTGCAGGGGCCACTG Capitalized sequences correspond to the gene-specific region.
[0340]Cycling conditions were as follows:
TABLE-US-00018 Denature 30 sec 95° C. Anneal 30 sec 50° C. {close oversize brace} 25 cycles Extend 1.5 min 72° C. Final extension 5 min 72° C.
[0341]Ten microliters of the nested products were analyzed by 1.5% agarose gel electrophoresis using ethidium bromide for detection (FIGS. 9B and 9C). The arrows indicate overlap-extension products with the expected migration properties of approximately 1 kb.
[0342]The present experiment illustrates that the lambda multiplex overlap-extension primer mix shown in Table 6 is applicable in single-step multiplex overlap-extension RT-PCR independent on the template used. Further, specific overlap-extension PCR products were produced at 0.5 pg total RNA. This sensitivity suggest that cognate linked heavy chain variable region and light chain variable region encoding sequences can be amplified from a single cell.
Example 6
Generation of a Sub-Library of Tetanus-Specific Cognate Pairs of Antibody Encoding Sequences
[0343]In the present example the steps outlined in the flow chart illustrated in FIG. 10, are exemplified using Tetanus Toxoid (TT) as target antigen.
[0344]a. Donors
[0345]Donors which previously have been immunized with the Tetanus vaccine are boosted with Tetanus vaccine (Statens Serum Institut, Denmark). Six days post the Tetanus vaccine boost a blood sample of approximately 200 ml is drawn from the donors into a tube containing anticoagulant.
[0346]It is required that the donors generally are healthy with no silent or chronic infections. They should not suffer from autoimmune diseases or receive any immunosuppressive medication, and they should not have had any vaccinations within the last 3 months. Also, at the time of the TT-vaccine boost, the donors must not have had any serious infections within the last month.
[0347]b. Preparation of Peripheral Blood Mononuclear Cells (PBMC)
[0348]PBMC are isolated from the blood samples using Lymphoprep (Axis-Shield PoC AS, Norway, prod. No 1001967) according to the manufactures recommendations. Briefly, the blood is diluted 1:1 in PBS and this suspension is layered over Lymphoprep in a 2:1 ratio. Vials are centrifuged for 20 min, 25° C. at 800 g and the white inter phase band is collected. Cells are washed in PBS containing 2 mM EDTA.
[0349]c. Enrichment of B Cells
[0350]The B-cell lineage (CD19+ cells) of the PBMC's is enriched by magnetic bead cell sorting using the following procedure.
[0351]The isolated PBMC are stained with anti-CD19-FITC (Becton Dickenson, N.J., USA, cat. No 345776). All steps are performed at 4° C. in the dark. Staining is performed with 10 μl anti-CD19-FITC pr. 1×106 cells in a volume of 100 μl per 1×106 cells using M-buffer (PBS, pH 7.2, 0.5% BSA, 2 mM EDTA). This will stain the B-cell lineage of the PBMCs. Cells are incubated for 20 min followed by two washing steps with M-buffer. The anti-CD 19-FITC stained cells are magnetically labeled with anti-FITC conjugated microbeads, using 10 μl of anti-FITC magnetic beads (Miltenyi Biotec, Gladbach, Germany, cat. No 130-042-401) per 1×106 cells in a volume of 100 μM-buffer per 1×106 cells. Incubation is performed for 15 min followed by a single washing step with M-buffer. The cells are resuspended in degassed M-buffer.
[0352]A MACS LS column (Miltenyi Biotec, Gladbach, Germany, cat. No 130-042-401) is pretreated with degassed M-buffer according to the manufactures prescriptions. The suspension of cells stained with anti-CD19-FITC and labeled with anti-FITC-magnetic beads are applied to the column and allowed to run through. Stained and labeled cells (CD19+) will be retained in the magnetic field surrounding the column while unstained cells (CD19-) will pass through the column. The column is washed with degassed M-buffer. The magnetic field is removed and the CD19+ cells are collected.
[0353]d. Sorting of Plasma Cells
[0354]The eluate from the MACS column is centrifuged and resuspended in FACS buffer (PBS, pH 7.2, 2% BSA) in a concentration of 1×106 cells/60 μl FACS buffer. Anti-CD19-FITC (Becton Dickenson, N.J., USA, cat. No 345776) (10 μl/106 cells), anti-CD38-PE (Becton Dickenson, N.J., USA, cat. No 555460) (10 μl/106 cells) and anti-CD45-PerCP (Becton Dickenson, N.J., USA, cat. No 345809) (20 μl/106 cells) are added. Incubation is performed at 4° C. for 20 min in the dark, followed by two times washing and resuspension in FACS buffer.
[0355]The cells are sorted by fluorescence activated cell sorting (FACS) using the following gating parameters: [0356]1. Forward scatter and side scatter in order to retain lymphocytes and monocytes including plasma blasters and plasma cells and to avoid dead cells and cells with very high side scatter which may be aggregates or granulocytes. [0357]2. Cells that are CD19 positive and express increased levels of CD38 (CD38hi). This is basically only a gate on CD38 since the PBMCs have been enriched on a MACS column for CD19 expression, but this will disclose any contaminants. [0358]3. CD45 positive cells. All lymphocytes express CD45. However, plasma cells down regulate their CD45 expression compared to earlier lymphocyte differentiation stages. Therefore a discrete population of cells corresponding to plasma cells can be obtained when gating on CD45.
[0359]FACS sorted cells are collected as single cells directly in single wells of 96 well plates containing 5 μl PBS buffer supplemented with 5 U RNase inhibitor (RNasin, Promega, Madison USA, cat. No N2515) per well. At this point the cells may be frozen for later RT-PCR or the cells may proceed immediately for RT-PCR.
[0360]e. Linkage of Cognate Pairs of Immunoglobulin Variable Region Encoding Sequences
[0361]The multiplex overlap-extension RT-PCR technology is applied to the single cells, thereby achieving cognate linkage of transcribed anti-Tetanus Toxoid heavy chain variable region and light chain variable region encoding sequences.
[0362]e-1. Single-Step Multiplex Overlap-Extension RT-PCR
[0363]The Qiagen One-Step RT-PCR kit (Qiagen cat. No 210212, Hilden, Germany) is used for the multiplex overlap-extension RT-PCR essentially according to the manufacturer's recommendation. Frozen 96-well plates containing a single cell per well are removed from the freezer and when the wells are free of ice crystals 15 μl RT-PCR reaction mixture is added immediately to each sample (single cell).
[0364]The RT-PCR reaction mixture contains, in a total volume of 20 μl, the following reagents: [0365]1× One-Step RT-PCR buffer, [0366]dNTP's in a final concentration of 400 μM of each, [0367]Multiplex overlap-extension primer mix in the concentrations as indicated in table 8, [0368]0.8 μl One-step RT-PCR enzyme mix, and [0369]20 U RNase inhibitor (RNasin, Promega, Madison USA, cat. No N2515).
[0370]The composition of the multiplex overlap-extension primer mix is shown in table 8.
TABLE-US-00019 TABLE 8 Primer SEQ ID name Conc. NO primer sequence in 5' to 3' direction VH 40 nM 41 tattggcgcgccatggccCAGRTGCAGCTGGTGCART of each 42 tattggcgcgccatggccSAGGTCCAGCTGGTRCAGT 43 tattggcgcgccatggccCAGRTCACCTTGAAGGAGT 44 tattggcgcgccatggccSAGGTGCAGCTGGTGGAG 45 tattggcgcgccatggccCAGGTGCAGCTACAGCAG 46 tattggcgcgccatggccCAGSTGCAGCTGCAGGAGT 47 tattggcgcgccatggccGARGTGCAGCTGGTGCAGT 48 tattggcgcgccatggccCAGGTACAGCTGCAGCAGTC CH(IgG) 200 nM 9 GACSGATGGGCCCTTGGTGG VL 40 nM 54 gccatggcgcgccaatagctagccGACATCCAGWTGACCCAGTCT of each 55 gccatggcgcgccaatagctagccGATGTTGTGATGACTCAGTCT 56 gccatggcgcgccaatagctagccGAAATTGTGWTGACRCAGTCT 57 gccatggcgcgccaatagctagccGATATTGTGATGACCCACACT 58 gccatggcgcgccaatagctagccGAAACGACACTCACGCAGT 59 gccatggcgcgccaatagctagccGAAATTGTGCTGACTCAGTCT CLκ 200 nM 60 atatatatgcggccgcttaTTAACACTCTCCCCTGTTGAA W = A/T, S = G/C, R = A/G. Capitalized sequences correspond to the gene specific region.
[0371]Cycling conditions are as follows:
TABLE-US-00020 Reverse transcription: 30 min 55° C. Polymerase activation: 15 min 95° C. inactivates reverse transcriptase and activates Taq polymerase.
[0372]PCR reaction:
TABLE-US-00021 Denature 30 sec 94° C. Anneal 30 sec 52° C. {close oversize brace} 35 cycles Extend 5 min 72° C. Final extension 10 min 72° C.
[0373]e-2. Additional Amplification
[0374]One microliter of the multiplex overlap-extension RT-PCR reaction product, from each sample, is subjected to semi-nested PCR (Biotaq kit, Bioline, UK cat. No BIO-21040), essentially as proposed by the manufacturer utilizing 96 well plates. The total volume of each reaction is 50 μl, containing a final concentration of 1× Biotaq buffer, 200 μM dNTP's (of each), 2 mM MgCl2, 1.25 U Bio Taq polymerase and the primers shown in Table 9.
[0375]Cycling conditions are as follows:
TABLE-US-00022 Denature 30 sec 95° C. Anneal 30 sec 55° C. {close oversize brace} 30 cycles Extend 1.5 min 72° C. Final extension 5 min 72° C.
TABLE-US-00023 TABLE 9 SEQ Primer ID primer sequence in name Conc. NO 5' to 3' direction JH 200 nM 61 ggaggcgctcGAGACGGTGACCAGGGTGCC of each 62 ggaggcgctcGAGACGGTGACCATTGTCCC 63 ggaggcgctcGAGACGGTGACCAGGGTTCC 64 ggaggcgctcGAGACGGTGACCGTGGTCCC CLκ 200 nM 21 accgcctccaccggcggccgcttaTTAACA CTCTCCCCTGTTGAAGCTCTT Capitalized sequences correspond to the gene specific region.
[0376]Ten microliters of a limited set of samples are analyzed by 1.5% agarose gel electrophoresis using ethidium bromide for visualization, in order to verify that the multiplex overlap-extension RT-PCR has been successful.
[0377]Expected size of the fragments (the exact size depends upon the lengths of the variable regions):
[0378]VH: ˜410 bp
[0379]LC: ˜680 bp
[0380]Overlap-extension fragment: ˜1070 bp
[0381]Two microliters from all the samples, run in the 96 well plates, originating from the same donor, are combined in a single tube. The pooled samples are digested with XhoI and NotI. The digested overlap-extension fragments are purified by preparative 1% agarose gel electrophoresis; the overlap-extension fragments are excised from the agarose gel and purified using a Qiaex II kit (Qiagen cat. No 20051, Hilden, Germany). It is not necessary to pool the cognate pairs at this stage if this is not desired. In that case each individual reaction will be subjected to restriction cleavage and the products are cloned individually into the vector described below.
[0382]f. Cognate Fab Expression Library
[0383]A library of Fab vectors is generated by inserting the pool of XhoI/NotI digested overlap-extension fragments into E. coli vector JSK301 (FIG. 11) by ligation. E. coli (TOP10) is transformed by electroporation with the library of Fab vectors and the transformants are selected on 2×YT agar containing 100 μg/ml carbenicillin. Plasmid DNA is prepared from colonies directly off the agar plates. The plasmid preparation is derived from a minimum number of colonies per donor, corresponding to 3× the total number of single plasma cells originally sorted, in order to maintain diversity. An Fab expression library is generated by inserting a prokaryotic promoter and leader cassette derived from phh3 (Den, W. et al. 1999. J. Immunol. Methods 222, 45-57) into the resulting plasmid preparation by digesting the plasmid preparation with AscI and NheI and subsequent ligation. The library generation process is outlined in FIG. 12. E. coli (TG1) is transformed by electroporation with the resulting Fab expression library and the transformants are selected on 2×YT agar containing 100 μg/ml carbenicillin. The individual donors are kept separate during the described procedure in Den et al, J Immunol Methods. 1999 Jan. 1; 222(1-2):45-57. They can, however, be combined at any stage where this might be desired.
[0384]g. Screening of Clones
[0385]Fab expressing clones are screened for TT antigen-binding by antigen-specific ELISA assays.
[0386]g-1. Master Plate Generation and Fab Expression
[0387]Individual selected colonies of the TG1 cells, each harbouring a cognate Fab expression vector from the library generated in step (f) are picked into single wells of 96 well plates containing 2×YT/100 μg/mL Amp/1% glucose. The colonies are grown overnight at 37° C. with gentle shaking. These plates are referred to as master plates, which are stored at -80° C. upon addition of glycerol to a final concentration of 15%.
[0388]Before the master plates are stored, they are used for inoculation of one or more 384 well plates containing 2×YT/100 μg/mL Amp 0.1% glucose using a 96 pin replicator. The plates are sealed and shaken for 2-3 h at 37° C.
[0389]Fab expression is induced by adding an equal volume of 2×YT/100 μg/mL Amp/0.2 mM IPTG, obtaining a final IPTG concentration of 0.1 mM. The plates are sealed and shaken overnight at 30° C. The following day, the Fab containing supernatants are analyzed for TT antigen-binding specificity by ELISA.
[0390]g-2. ELISA Analysis
[0391]Three hundred and eighty four (384) well ELISA plates (Nunc, Roskilde, Denmark, cat. No 265196) are coated overnight at 4° C. with tetanus toxoid (TT) antigen, diluted to a final concentration of 1 μg/mL in PBS in a volume of 25 μl per well. Excess binding sites of the wells are blocked for 1 h at RT by adding 2% M-PBS-T (2% skim milk powder in PBS, 0.05% Tween 20). The wells are washed 2 times with PBS-T (PBS, 0.05% Tween 20).
[0392]The Fab containing bacterial supernatants from g-1. are diluted 1:2 in 2% M-PBS-T, and transferred to the ELISA wells in duplicate. Incubation is performed for 1 h at RT. The wells are washed 4 times with PBS-T. Goat-anti-human Fab/HRP (Sigma, St. Louis, Mo., USA, cat. No A0293) 1:10.000 dilution in 2% M-PBS-T is added to the wells. Incubation is performed for 1 h at RT. The wells are washed 4 times with PBS-T. TMB Plus (KemEnTec, Copenhagen, Denmark, cat. No 4390L) substrate is added and incubation is performed for 5-15 minutes. The reactions are stopped by adding an equal volume of 1 M H2SO4. The extents of the reactions are read at 450 nm in an ELISA reader (Multiscan Ascent, Labsystems, Franklin, USA).
[0393]The original bacterial clones corresponding to TT antigen-binding clones are subsequently retrieved from the original master plates. Plasmid DNA is prepared from the isolated antigen positive Fab clones, generating a cognate Fab expression sub-library of TT antigen-binding clones.
[0394]The clones may also be analyzed by an anti-light chain ELISA assay, in order to get a correlation between the number of antigen-binding clones and the number of Fab expressing clones. Further, such an analysis provides information of kappa and lambda light chain representation in the clones.
[0395]h. Cognate Antibody Expression Library
[0396]In order to facilitate the expression of full-length human antibodies the cognate variable regions from the bacterial Fab expression vector must be transferred to a mammalian expression vector containing the constant domains from one of the human immunoglobulin isotypes. Such a transfer can either be performed clone by clone or in mass. Below it is described how to perform the mass transfer.
[0397]The mass transfer is performed in two steps. First, the plasmid preparations of the individually isolated antigen-binding Fab clones are pooled. The prokaryotic promoter and leader containing cassette is exchanged with a mammalian promoter and leader cassette consisting of human alkaline phosphatase leader sequence (AP Leader), human elongation factor 1-alpha promoter (EFP), adenovirus major late promoter (AdMLP), IgK leader sequence by digesting the pooled Fab expression sub-library with AscI and NheI followed by insertion of a mammalian promoter and leader cassette by a subsequent ligation. E. coli (TOP10) is transformed by electroporation with the resulting promoter exchanged Fab expression library and the transformants are selected on 2×YT agar containing 100 μg/ml carbenicillin. Plasmid DNA is prepared from colonies directly off the agar plates. The plasmid preparation is derived from a minimum number of colonies per donor, corresponding to approximately 3× the total number of clones in the original library, in order to maintain diversity.
[0398]Second, the cognate variable regions from the promoter exchanged Fab expression vector are isolated by digesting the plasmid preparation with XhoI and NotI. The isolated fragment is inserted by ligation into a mammalian expression vector resulting in a cognate antibody expression library. The vector used is essentially the same as in FIG. 9A, except that IgL in this case corresponds to the kappa light chain encoding sequence. The mammalian expression vector used above is based on the site-specific Flp-In system (Invitrogen Corporation, Carlsbad, Calif., USA, Cat No K6010-01). E. coli (TOP10) is transformed by electroporation with the resulting cognate antibody expression library and the transformants are selected on 2×YT agar containing 100 μg/ml carbenicillin. Plasmid DNA is prepared from colonies directly off the agar plates. The plasmid preparation is again derived from a minimum number of colonies per donor, corresponding to approximately 3× the total number of clones in the original library, in order to maintain diversity. The resulting plasmid preparation of the cognate antibody expression library can be used to transfect a mammalian host cell, e.g. the CHO cells from Invitrogen's Flp-In system (Invitrogen Corporation, Carlsbad, Calif., USA, Cat No K6010-01) in order to generate a stable mammalian expression cell lines by Flp recombinase-mediated integration. Such a cell line can be used in the production of recombinant polyclonal antibodies.
Example 7
Screening by High Density Membrane Assay
[0399]The Fab expressing clones from Example 6f. are screened by a high density membrane assay.
[0400]a. Master Plate Generation
[0401]Master plates are generated as described in step (g-1) of Example 6.
[0402]b. PVDF Membrane Preparation
[0403]PVDF membranes (Amersham, Uppsala, Sweden, cat. NoRPN2020F) are coated according to the manufacturer's instructions overnight at 4° C. with TT antigen diluted to a final concentration of 1 μg/ml in PBS. Excess binding sites on the membranes are blocked for 1 h in 2% M-PBS (2% skim milk powder in PBS). The membranes are rinsed 3 times in PBS. The membranes are soaked in 2×YT for a few minutes.
[0404]c. Clone Consolidation
[0405]Consolidation of clones into 384 well plates is performed by transferring clones from the master plates into one or more 384 well plates containing 20 μA 2×YT/well, using a 96 pin replicator.
[0406]Using a 384 pin replicator the bacteria are transferred to one or more nylon membranes in duplicate (Amersham, Uppsala, Sweden, cat. No RPN2020B) placed on 2×YT/Carb/1% glucose agar plates. The plates are incubated 4-6 h at 37° C.
[0407]d. Fab Induction
[0408]The PVDF membranes prepared in step b, are placed on a 2×YT/Carb/0.1 mM IPTG agar plates. The nylon membranes with the growing bacterial colonies are placed on top of the PVDF membranes (one nylon membrane per PVDF membrane) on the IPTG-containing agar plate. The IPTG-containing agar plate is incubated at 30° C. overnight to facilitate IPTG induced Fab expression from the colonies. The Fab molecules diffuse through the nylon membrane onto the PVDF membrane, where TT-antigen binding Fab' s will be retained on the membrane.
[0409]e. Detection
[0410]The nylon membranes are removed and the PVDF membranes are washed 2×5 minutes with PBS-T (PBS, 0.05% Tween 20). The membranes are incubated for 30 minutes with 2% M-PBS. The PVDF membranes are washed 2×5 minutes with PBS-T, followed by incubation for 1 h with goat-anti-human IgG/HRP (Sigma, St. Louis, Mo., USA, cat. No A0293), 1:10.000 diluted in 2% M-PBS. The PVDF membranes are washed 3×5 minutes with PBS-T. Finally, the PVDF membranes are incubated for 5 minutes with SuperSignal West Femto chemiluminiscent substrate (Pierce, Rockford, Il, USA, cat. No 34095) according to the manufactures instructions. Excess substrate is removed and the PVDF membranes are placed under a CCD camera for detection of the chemiluminescent signal generated at positions on the PVDF membrane where a Fab fragment binds the TT-antigen.
[0411]Positive TT-antigen binding clones will appear as dots on the image. The original bacterial clone is subsequently retrieved from the original master plates. Plasmid DNA is prepared from the isolated antigen positive Fab clones, generating a cognate Fab expression sub-library of TT-antigen binding clones.
[0412]f. Correlation Between Antigen Binding Clones and Fab Expressing Clones
[0413]The nylon membranes removed in step e. are placed on a second series of PVDF membranes coated with anti-light chain antibody or anti-Fab antibody according to the procedure in step b. Step d. and e. are then repeated.
[0414]Fab positive clones will then appear as dots on the image and a correlation between the number of TT-antigen binding clones and the number of Fab expressing clones can be made.
Example 8
Illustrative Example Describing the Isolation of Sequences Encoding a Heteromeric Protein
[0415]The present example illustrates how a trimeric guanine nucleotide binding protein (G-protein) can be isolated in a single step utilizing the multiplex overlap-extension RT-PCR technique of the present invention. The family of G-protein subunits is large, alone in humans there are approximately 16 different alpha subunits, 5 beta subunits and 12 gamma subunits. For the present example subunit GαS (GenBank accession No X04408), subunit Gβ1 (GenBank accession No AF501822) and subunit Gγ1 (GenBank accession No BC029367) were chosen. The linkage procedure is illustrated in FIG. 13, where step 1 is the multiplex overlap-extension RT-PCR step and step 2 is the additional amplification of the linked product.
[0416]a. Cells
[0417]A population of human liver cells is obtained from surgical discharge samples. The liver cells are disintegrated and lysed, total RNA is isolated from the lysate using RNA L kit (Macherey--Nagel, Duren, Germany, cat. No 740 962.20) according to the manufactures instructions.
[0418]b. Multiplex Overlap-Extension RT-PCR
[0419]The Qiagen One-Step RT-PCR kit (Qiagen cat. No 210212, Hilden, Germany) is used for the multiplex overlap-extension RT-PCR essentially according to the manufacturer's recommendation.
[0420]Each PCR tube contains, in a total volume of 50 the following reagents: [0421]1× One Step RT-PCR buffer, [0422]dNTP's in a final concentration of 400 μM of each, [0423]Multiplex overlap-extension primer mix in the concentrations as indicated in table 10, [0424]2 μl One step RT-PCR enzyme mix, [0425]1 U/μl RNase inhibitor (RNasin, Promega, Madison USA, cat. No N2515), and [0426]50 ng total RNA.
[0427]The multiplex overlap-extension primer mix used comprises the primers shown in table 10.
TABLE-US-00024 TABLE 10 Primer SEQ Name Conc. ID NO primer sequence in 5' to 3' direction α1 400 nM 80 atatatatgcggccgcttaATGGGCTGCCTCGGGAA α2 80 nM 81 gccatggcgcgccaatagctagccTTAGAGCAGCTCGTACTGACGAA β1 80 nM 82 tattggcgcgccatggccATGAGTGAGCTTGACCAGTTACG β2 80 nM 83 ggtacctaataataatcgatcgcTTAGTTCCAGATCTTGAGGAAGCT γ1 80 nM 84 cgatcgattattattaggtaccccATGCCAGTAATCAATATTGAGGAC γ2 400 nM 85 ggaggcgctcgagTTATGAAATCACACAGCCTCCTTTG Capitalized sequences correspond to the gene-specific region.
[0428]The cycling conditions used are:
TABLE-US-00025 Reverse transcription: 30 min 42° C. Polymerase activation: 15 min 95° C. inactivating reverse transcriptase and activating Taq polymerase.
[0429]PCR reaction:
TABLE-US-00026 Denature 30 sec 94° C. Anneal 30 sec 49° C. {close oversize brace} 45 cycles Extend 3 min 72° C. Final extension 10 min 72° C.
[0430]c. Amplification PCR
[0431]One microliter of the multiplex overlap-extension RT-PCR reaction product is subjected to an additional PCR amplification step (Biotaq kit, Bioline, UK cat. No BIO-21040) essentially as proposed by the manufacturer. The primers used are α1 and γ2 as shown in table 10 corresponding to SEQ ID NO 80 and 85. The total volume of the reaction is 50 μl.
[0432]Cycling conditions are as follows:
TABLE-US-00027 Denature 30 sec 95° C. Anneal 30 sec 53° C. {close oversize brace} 25 cycles 81 Extend 3 min 72° C. Final extension 10 min 72° C.
[0433]Five microliters of the reaction product is analyzed by 1% agarose gel electrophoresis using ethidium bromide for detection. The size of the product is expected to be 2215 bp. However, additional unlinked fragments of the subunits might be present. The expected sizes of these fragments are Gα: 1228 bp, Gβ: 1064 bp and Gγ: 262 bp.
[0434]d. Cloning
[0435]The remaining reaction product is subjected to 1% agarose gel electrophoresis using ethidium bromide for detection and the overlap-extension fragment of 2215 bp is excised from the agarose gel, purified using a Qiaex II kit (Qiagen, Hilden, Germany, cat. No 20051) and inserted into pCR2.1-TOPO using Invitrogen's TOPO TA cloning kit (Invitrogen cat. No 45-0641, Carlsbad, Calif., USA).
[0436]Furthermore, promoter sequences or ribosome binding sites might be inserted upstream of each subunit encoding sequence in order to facilitate their expression from the vector. The restriction sites present in the overlap-extension sequences are applicable for such insertion.
[0437]e. General Considerations
[0438]The linkage of sequences encoding subunits constituting a heteromeric protein, as described above, can be performed for most heteromeric proteins where the sequence of the individual subunits is known. Moreover, when utilizing a multiplex overlap-extension primer mixture capable of amplifying several members of a particular family, e.g. all the G-protein α-, β-, and γ-subunits, it is possible to identify and link any combination of subunits from a cell type without first having to analyze which family members are expressed by what cell type. For example, the above example can be performed utilizing template derived from isolated single liver cells employing a multiplex overlap-extension primer mix capable of amplifying and linking all 16 Gα subunits with the 5 Gβ subunits and 12 Gγ subunits, thereby identifying which combination of subunits are expressed in each cell and simultaneously isolating the encoding sequences responsible for that combination of subunits. In addition, the use of degenerate primers might even serve to identify new members of a particular family.
[0439]When linking subunits of a heteromeric protein using the multiplex overlap-extension RT-PCR technique of the present invention, the overall size of the linked product should be considered, since there are limits to the number of base pairs a DNA polymerase can amplify. The Deep Vent polymerase from New England Biolabs, MA, USA is one DNA polymerase capable of generating very long primer extensions, of up to 14,000 bp in length. Theoretically, 14,000 bp can encode a protein with an average weight of 510 kDa. Thus, it should be possible to isolate encoding sequences for very large heteromeric proteins utilizing the method of the present invention. The extension capability could possibly be increased even further by utilizing mixtures of DNA polymerases.
Example 9
Design of Leader Primers
[0440]In this example, a multiplex set of primers in which the site of priming is kept in the sequence encoding the leader region of the human antibody heavy chain and kappa light chain families was designed.
[0441]When using primers annealing to the N-terminal encoding sequences of the variable region families, some degree of cross-hybridization may be observed. Primers of one particular gene family sequence can hybridize to the cDNA of another gene family, which in many cases cause creation of novel sequences. Proteins produced from such sequences are potentially immunogenic when used in treatment. Generally, these novel sequences can be corrected by PCR, or eliminated from the library.
[0442]An alternative solution, is to position the site of priming in the sequence encoding the leader peptide region of the antibody variable region. Hybrid sequences, generated as a result of cross-priming, will be eliminated during intra-cellular processing of the antibody, in which the peptide leader containing the potential immunogenic sequences will be cleaved off, and hence will not be present in the secreted antibody product. Previous sets of leader primers for antibody cloning, have been located in the 5' end of the leader encoding sequence, allowing for direct eukaryotic expression (Campbell M. J. et al. 1992 Mol Immunol. 29, 193-203). Unfortunately, eukaryotic leader peptides are poorly suited for prokaryotic processing.
[0443]The ease by which nucleic acid sequences are manipulated in bacteria makes it attractive to develop cloning systems in which sequence shuttling between bacterial vectors and vectors for other host organisms is possible. Therefore, a multiplex primer set, in which the site of priming is kept at a position in the leader region encoding sequence, that allow for functional expression of antibodies or antibody fragments in both eukaryotic and prokaryotic expression systems, was designed in the present example.
[0444]The sequence requirement for signal peptidase cleavage at the C-terminal region of any leader peptide seems very similar for prokaryote and eukaryote systems (Nielsen H. et al. 1997 Protein Eng. 10, 1-6.). Further, it follows that mutations in the C-terminal region of the leader peptide is likely to have little effect on the conformation of the N-terminal region and vice versa. Thus, it is expected that C-terminal sequences could be transferable among species without loss of function.
[0445]The basic concept of the primer design in the present example, was to place the site of priming at the last six codons of the C-terminal region of the leader peptide. The remaining part of the leader encoding sequence, up-stream of the site of priming (approximately 50 nucleotides) should be supplied by the appropriate expression vector matching the host species. For example, vectors suitable for expression in gram negative bacteria can be designed with a partial or full-length PelB leader sequence. For expression of antibodies in eukaryotes, a vector with a partial native antibody heavy chain leader encoding sequence and a partial variable kappa chain leader encoding sequence will be suitable. Regardless of expression system, the last six amino acids of the leader, will originate from the endogenous antibody leader encoding sequence contained in the nucleic acid segment inserted into the vector.
[0446]The following primer sequences were designed for use in the multiplex overlap-extension RT-PCR (table 11). However, the overlapping tails (lowercase letters) can easily be redesigned to function in the linkage by ligation or recombination procedures.
TABLE-US-00028 TABLE 11 SEQ Primer ID name NO primer sequence in 5' to 3' direction VHL 86 tattcccagcggccgccGCCACAGGTGCCCACTC 87 tattcccagcggccgccCCTTCMTGGGTCTTGTC 88 tattcccagcggccgccTTARAAGGTGTCCAGTGT 89 tattcccagcggccgccCCCAGATGGGTCCTG 90 tattcccagcggccgccCTCCAAGGAGTCTGTTC 91 tattcccagcggccgccCCATGGGGTGTCCTGTCA 92 tattcccagcggccgccGCAACAGGTGCCCACT CH(IgG) 9 GACSGATGGGCCCTTGGTGG VLκL 93 ggcggccgctgggaatagctagcgYTCCCAGGTGCCAGA TG 94 ggcggccgctgggaatagctagcgGTCCCTGGATCCAGT G 95 ggcggccgctgggaatagctagcgCTCCCAGATACCACC GGA 96 ggcggccgctgggaatagctagcgATCTCTGGTGCCTAC 97 ggcggccgctgggaatagctagcgATCTCTGATACCAGG GCA 98 ggcggccgctgggaatagctagcgGTTCCAGCCTCCAGG GGT CLκ 99 atatatatggcgcgccTTAACACTCTCCCCTGTTGAA M = A/T, R = A/G, Y = C/T, S = G/C. Capitalized sequences correspond to the gene-specific region.
[0447]It should be noted that the restriction sites in this primer design were slightly modified compared to the primers described in the previous examples. Thus, the overlap sequence between the heavy chain and light chain variable region encoding sequences comprised a NotI and a NheI restriction site, and in the constant kappa primer (CL,), the previous NotI site has been exchanged with an AscI site. The later modification should of course be observed when designing the primers for the additional PCR amplification. Further, the NotI site in the cloning vector, shown in FIG. 11, should be changed to a AscI site, and the AscI site in the promoter unit should be changed to a NotI site.
[0448]The functionality with respect to leader cleavage has been tested in silico using SignalP provided by CBS at the Danish Technical University.
[0449]The chimeric leader peptides tested for the variable heavy chain, were encoded by nucleic acid sequences composed of a truncated PelB sequence, a C-terminal NotI site (SEQ ID NO 100: ATG AAA TAT CTT CTA CCA ACA GCG GCA GCT GGA TTA TTG GCG GCC GCC), and in association with the NotI site the gene specific part of one of SEQ ID NO 86 to 92, encoding the six C-terminal amino acids from native VHL family members. All seven sequences showed signal peptide cleavage in gram negative bacteria, using the SignalP program.
[0450]The chimeric leader peptides tested for the variable light chain, were encoded by nucleic acid sequences composed of a truncated PelB sequence, a C-terminal NheI site (SEQ ID NO 101: ATG AAA TAT TTG CTA CCA ACA GCG GCA GCT GGA TTA TTG TTA CTA GCG), and in association with the NheI site the gene specific part of one of SEQ ID NO 93 to 98, encoding the six C-terminal amino acids from native VLκL family members. All six sequences showed signal peptide cleavage in gram negative bacteria, using the SignalP program.
[0451]The sequences of SEQ ID NO 100 and 101 are suitable for the construction of a bacterial expression vector similar to the one illustrated in FIG. 11, in that the AscI site illustrated in FIG. 11 is substituted with SEQ ID NO 100 and the NheI site is substituted with SEQ ID NO 101. Further, the NotI site illustrated in FIG. 11 needs to be substituted for a AscI site.
[0452]The mammalian expression vector (FIG. 9) can likewise be re-designed to enable expression in mammalian cells following transfer of the variable region encoding segments from the bacterial vector just described.
Example 10
One Tube Multiplex RT-PCR and Linkage by Ligation
[0453]The present example illustrates how a cognate pair comprising linked heavy chain variable region and light chain variable region encoding sequences can be generated in a single tube reaction using ligation instead of overlap-extension PCR, to obtain linkage.
[0454]a. Multiplex RT-PCR
[0455]An in-house produced cell line expressing Fab molecules with specificity towards Tetanus Toxoid are distributed by limiting dilution to obtain a single cell in a single PCR tube.
[0456]In addition to the single cell, each PCR tube contains the following reagents in a total volume of 20 μl:
[0457]1× Phusion HF buffer [0458]dNTP's in a final concentration of 200 μM each [0459]Multiplex primer mix in the concentrations as indicated in table 12 [0460]0.8 U Phusion polymerase (FinnZymes Cat. No F-530-L) [0461]1 μl Sensiscript reverse transcriptase (Qiagen Cat. No 205213 [0462]20 U Rnase inhibitor
TABLE-US-00029 [0462]TABLE 12 Pimer SEQ ID name Conc. NO primer sequence in 5' to 3' direction VH 200 nM 102 tgttgttctagatgaaggcgcgccCAGRTGCAGCTGGTGCART of each 103 tgttgttctagatgaaggcgcgccSAGGTCCAGCTGGTRCAGT 104 tgttgttctagatgaaggcgcgccCAGRTCACCTTGAAGGAGT 105 tgttgttctagatgaaggcgcgccSAGGTGCAGCTGGTGGAG 106 tgttgttctagatgaaggcgcgccCAGGTGCAGCTACAGCAGT 107 tgttgttctagatgaaggcgcgccCAGSTGCAGCTGCAGGAGT 108 tgttgttctagatgaaggcgcgccGARGTGCAGCTGGTGCAGT 109 tgttgttctagatgaaggcgcgccCAGGTACAGCTGCAGCAGTC JH 200 nM 61 ggaggcgctcGAGACGGTGACCAGGGTGCC of each 62 ggaggcgctcGAGACGGTGACCATTGTCCC 63 ggaggcgctcGAGACGGTGACCAGGGTTCC 64 ggaggcgctcGAGACGGTGACCGTGGTCCC VL 200 nM 110 aacaacactagtgataggctagccGACATCCAGWTGACCCAGTCT of each 111 aacaacactagtgataggctagccGATGTTGTGATGACTCAGTCT 112 aacaacactagtgataggctagccGAAATTGTGWTGACRCAGTCT 113 aacaacactagtgataggctagccGATATTGTGATGACCCACACT 114 aacaacactagtgataggctagccGAAACGACACTCACGCAGT 115 aacaacactagtgataggctagccGAAATTGTGCTGACTCAGTCT CLκ 200 nM 16 atatatatgcggccgcttaTTAACACTCTCCCCTGTTG W = A/T, R = A/G, S = G/C. Capitalized sequences correspond to the gene-specific region
[0463]The cycling conditions are:
TABLE-US-00030 Reverse 30 min 37° C. transcription Denature 30 sec 98° C. Denature 10 sec 98° C. Anneal 30 sec 55° C. {close oversize brace} 40 cycles Extend 30 sec 72° C. Final extension 5 min 72° C.
[0464]b. Linkage by Ligation
[0465]To perform the linkage by ligation, restriction enzymes and ligase are added directly to the multiplex RT-PCR reaction products. Alternatively, the multiplex RT-PCR can be purified prior to this addition in order to free the mixture from the polymerase.
[0466]The following reagents are added to each tube in a total volume of 40 [0467]1×NEBII buffer [0468]1 mM ATP [0469]50 U XbaI (New England Biolabs Cat. No R0145L) [0470]25 U SpeI (New England Biolabs Cat. No R0133S) [0471]100 mg/ml BSA [0472]400 U T4 DNA ligase (New England Biolabs Cat. No M0202S)
[0473]The reaction is incubated at 16° C. over night.
[0474]The linked heavy chain and light chain variable region encoding sequences are purified from the reaction by gel electrophoresis and excision of the band of approximately 1000 bp.
[0475]In an alternatively version of this method, the multiplex RT-PCR reaction is performed with a CH primer instead of the JH primers in the multiplex primer set. This primer may either be equipped with a cloning tail that allows in-frame insertion of the heavy chain variable region into an expression vector, or a semi nested PCR may be performed with the primers of table 5.
Example 11
Generation of a Recombinant Polyclonal Immunoglobulin with Specificity Towards Tetanus Toxoid
[0476]In the present example results from a single donor (TT03) immunized with Tetanus Toxoid (TT) is used to illustrate the steps outlined in the flow chart in FIG. 10.
[0477]a. Donors
[0478]Eight Donors which previously have been immunized with the Tetanus vaccine were boosted with Tetanus vaccine (Statens Serum Institut, Denmark). The donors were assigned with numbers TT01 to TT08. Six days after the Tetanus vaccine boost a blood sample of approximately 200 ml was drawn from the donors into a tube containing anticoagulant.
[0479]The donors were healthy without chronic infections, autoimmune disease, or immunosuppressive medication, and they had not had any vaccinations within the last 3 months.
[0480]a-1. Monitoring Donor Quality
[0481]Pre-bleedings of the donors were taken at the time of immunization. Fourteen days later additional bleedings were taken to determine the serum titer. All donors responded with an increase in the TT-titer. An ELISPOT assay was also set up to measure the frequency of TT-specific plasma cells. Different cell fractions can be used for the ELISPOT, e.g. the major bleed from day 6, the PBMC fraction, the magnetic sorted fraction, or the FACS sorted fraction. ELISPOT can be used to evaluate the donor material and to identify the best responders, which then may proceed through the sorting step and multiplex overlap-extension RT-PCR. For donor TT03 an ELISPOT was performed using the CD19+ cell fraction (see section c). The ELISPOT assay was performed as described by Lakew (Lakew, M. et al. 1997. J. Immunol. Methods 203, 193-198) with minor modifications. The frequency of TT specific plasma cells in the PBMC fraction was calculated to 0.021%
[0482]b. Preparation of Peripheral Blood Mononuclear Cells (PBMC)
[0483]PBMC's were isolated from the blood sample using Lymphoprep (Axis-Shield PoC AS, Norway, prod. No 1001967) according to the manufactures recommendations. Briefly, the blood was diluted 1:1 in PBS and this suspension was layered over Lymphoprep in a 2:1 ratio. Vials were centrifuged for 20 min, 25° C. at 800 g and the white inter phase band was collected. Cells were washed in PBS containing 2 mM EDTA.
[0484]From the 200 ml full blood drawn from donor TT03 approximately 2×108 PBMC's were obtained.
[0485]c. Enrichment of B Cells
[0486]The B-cell lineage (CD19+ cells) of the PBMC's was enriched by magnetic bead cell sorting using the following procedure.
[0487]The isolated PBMC's were stained with anti-CD19-FITC (Becton Dickenson, N.J., USA, cat. No 345776). All steps were performed at 4° C. in the dark. Staining was performed with 10 μl anti-CD19-FITC pr. 1×106 cells in a volume of 100 μl per 1×106 cells using M-buffer (PBS, pH 7.2, 0.5% BSA, 2 mM EDTA). This will stain the B-cell lineage of the PBMC's. Cells were incubated for 20 min followed by two washing steps with M-buffer. The anti-CD19-FITC stained cells were magnetically labeled with anti-FITC-conjugated microbeads, using 10 μl of anti-FITC magnetic beads (Miltenyi Biotec, Gladbach, Germany, cat. No 130-042-401) per 1×106 cells in a volume of 100 μl M-buffer per 1×106 cells. Incubation was performed for 15 min followed by a single washing step with M-buffer. The cells were resuspended in degassed M-buffer.
[0488]A MACS LS column (Miltenyi Biotec, Gladbach, Germany, cat. No 130-042-401) was pretreated with degassed M-buffer according to the manufacturers descriptions. The suspension of cells stained with anti-CD19-FITC and labeled with anti-FITC-magnetic beads were applied to the column and allowed to run through. Stained and labeled cells (CD19+) were retained in the magnetic field surrounding the column while unstained cells (CD19/) passed through the column. The column was washed with degassed M-buffer. The magnetic field was removed and the CD19+ cells were collected.
[0489]An analytic staining of the starting material (PBMC), the unlabelled fraction and the CD19 labelled fraction from TT03, using anti-CD19-FITC, anti-CD38-PE, and anti-CD45-PerCP was performed (FIG. 14). This shows that the Magnetic cell sorting results in two distinct fractions compared to the PBMC fraction. The CD19 negative cells shown in panel B and the CD19 positive cells shown in panel C. Of the PBMC fraction 11% of the cells were CD19+, the extent of CD19 positive cell purification was 99.5% of R1 (the scatter gate in FIG. 14c) for TT03.
[0490]As seen in FIG. 14c, the anti-CD38/anti-CD45 plot showed a distinct population of CD38hi, CD45in cells (R2) corresponding to 1.1% of R1. This population contained the plasma cells and was collected during the later sorting step. This indicated that in the PBMC fraction 0.12% of the cells corresponded to plasma cells.
[0491]The CD19 positive cell fraction was frozen in FCS (Invitrogen, Cat. No. 16000-044)+10% DMSO (Sigma, Cat. No. D2650) for later sorting. An analysis like the one seen in FIG. 14 was made on the MACS purified cells that had been frozen to make sure that the cells were intact (FIG. 15). The staining patterns seen in FIG. 15 were the same as seen in FIG. 14c although a slightly broader staining intensity were observed. The cells that were collected by sorting is the subset of R1 and R2. This corresponds to approx. 1.1% of the MACS purified cells.
[0492]d. Sorting of Plasma Cells
[0493]The frozen eluate from the MACS column was thawed, centrifuged and resuspended in FACS buffer (PBS, pH 7.2, 2% BSA) in a concentration of 1×106 cells/60 μl FACS buffer. Anti-CD19-FITC (Becton Dickenson, N.J., USA, cat. No 345776) (10 μl/106 cells), anti-CD38-PE (Becton Dickenson, N.J., USA, cat. No 555460) (10 μl/106 cells) and anti-CD45-PerCP (Becton Dickenson, N.J., USA, cat. No 345809) (20 μl/106 cells) were added, and the mixture was incubated at 4° C. for 20 min in the dark, followed by two times washing and resuspension in FACS buffer.
[0494]The cells were sorted by fluorescence activated cell sorting (FACS) using the following gating parameters: [0495]1. Forward scatter and side scatter in order to retain lymphocytes and monocytes including plasma blasters and plasma cells and to avoid dead cells and cells with very high side scatter which may be aggregates or granulocytes. [0496]2. Cells that are CD19 positive and express increased levels of CD38 (CD38hi) This is basically only a gate on CD38 since the PBMC's have been enriched on a MACS column for CD19 expression, but this will disclose any contaminants. [0497]3. CD45 intermediate positive cells. All lymphocytes express CD45. However, plasma cells down regulate their CD45 expression compared to earlier lymphocyte differentiation stages. Therefore, a discrete population of cells corresponding to plasma cells can be obtained when gating on CD45.
[0498]FACS sorted cells from donor TT03 were collected as the subset of the two gates P2 and P1 (FIGS. 16A and 16B, respectively). The cells were CD38hi (FL2-A), CD45 in (FL3-A) and CD19+, the latter due to the MACS purification. Briefly, the cells were collected in bulk, counted and diluted in RPMI (Invitrogen, Cat No. 21875-034) containing 10% FCS (Invitrogen Cat. No. 16000-044), 100 units/ml Penicillin-streptomycin (Invitrogen, cat. No. 15140-122), 2 mM L-glutamine (cat No. 25030-024). Cells were then dispensed into fifty 96 well PCR plates (ABgene, cat. No. AB-0800) with one cell per well in 5 μl medium. The plates were sealed and frozen immediately, and stored for later RT-PCR analysis.
[0499]e. Linkage of Cognate Pairs of Immunoglobulin Variable Region Encoding Sequences
[0500]The multiplex overlap-extension RT-PCR technology was applied to the single cells obtained from donor TT03, thereby achieving cognate linkage of transcribed anti-Tetanus Toxoid heavy chain variable region and light chain variable region encoding sequences.
[0501]e-1. Single-Step Multiplex Overlap-Extension RT-PCR
[0502]The Qiagen One-Step RT-PCR kit (Qiagen cat. No 210212, Hilden, Germany) was used for the multiplex overlap-extension RT-PCR essentially according to the manufacturer's recommendation. Fifty frozen 96-well PCR plates containing approximately a single cell per well were removed from the freezer and when the wells were free of ice crystals, 15 μl RT-PCR reaction mixture was added immediately to each well.
[0503]The RT-PCR reaction mixture contained, in a total volume of 20 μl, the following reagents: [0504]1× One Step RT-PCR buffer [0505]dNTP's in a final concentration of 400 μM of each [0506]Multiplex overlap-extension primer mix in the concentrations as indicated in table 13 [0507]0.8 μl One-step RT-PCR enzyme mix [0508]20 U RNase inhibitor (RNasin, Promega, Madison USA, cat. No N2515)
[0509]The composition of the multiplex overlap-extension primer mix is shown in table 13.
TABLE-US-00031 TABLE 13 Primer SEQ ID name Conc. NO primer sequence in 5' to 3' direction VH 40 nM 116 tattcccatggcgcgccCAGRTGCAGCTGGTGCART of each 117 tattcccatggcgcgccSAGGTCCAGCTGGTRCAGT 118 tattcccatggcgcgccCAGRTCACCTTGAAGGAGT 119 tattcccatggcgcgccSAGGTGCAGCTGGTGGAG 120 tattcccatggcgcgccCAGGTGCAGCTACAGCAGT 121 tattcccatggcgcgccCAGSTGCAGCTGCAGGAGT 122 tattcccatggcgcgccGARGTGCAGCTGGTGCAGT 123 tattcccatggcgcgccCAGGTACAGCTGCAGCAGTC CH(IgG) 200 nM 9 GACSGATGGGCCCTTGGTGG VL 40 nM 124 ggcgcgccatgggaatagctagccGACATCCAGWTGACCCAGTCT of each 125 ggcgcgccatgggaatagctagccGATGTTGTGATGACTCAGTCT 126 ggcgcgccatgggaatagctagccGAAATTGTGWTGACRCAGTCT 127 ggcgcgccatgggaatagctagccGATATTGTGATGACCCACACT 128 ggcgcgccatgggaatagctagccGAAACGACACTCACGCAGT 129 ggcgcgccatgggaatagctagccGAAATTGTGCTGACTCAGTCT CLκ 200 nM 130 atatatatgcggccgcTTAACACTCTCCCCTGTTGAA W = A/T, S = G/C, R = A/G. Capitalized sequences correspond to the gene-specific region
[0510]Cycling conditions were as follows:
TABLE-US-00032 Reverse transcription: 30 min 55° C. Polymerase activation: 15 min 95° C. inactivates reverse transcriptase and activates Taq polymerase.
[0511]PCR reaction:
TABLE-US-00033 Denature 30 sec 94° C. Anneal 30 sec 52° C. {close oversize brace} 35 cycles Extend 5 min 72° C. Final extension 10 min 72° C.
[0512]e-2. Additional Amplification
[0513]One microliter of the multiplex overlap-extension RT-PCR reaction product, from each sample, was subjected to semi-nested PCR (Biotaq kit, Bioline, UK cat. No BIO-21040), essentially as proposed by the manufacturer utilizing 96 well PCR plates (ABgene, cat. No. AB-0800). The total volume of each reaction was 50 μl, containing a final concentration of 1× Biotaq buffer, 200 μM dNTP's (of each), 2 mM MgCl2, 1.25 U Bio Taq polymerase and the primers shown in table 14.
TABLE-US-00034 TABLE 14 SEQ Primer ID primer sequence in name Conc. NO 5' to 3' direction JH 200 nM 61 ggaggcgctcGAGACGGTGACCAGGGTGCC of each 62 ggaggcgctcGAGACGGTGACCATTGTCCC 63 ggaggcgctcGAGACGGTGACCAGGGTTCC 64 ggaggcgctcGAGACGGTGACCGTGGTCCC CLκ 200 nM 21 accgcctccaccggcggccgcttaTTAACA CTCTCCCCTGTTGAAGCTCTT Capitalized sequences correspond to the gene-specific region.
[0514]Cycling conditions were as follows:
TABLE-US-00035 Denature 30 sec 95° C. Anneal 30 sec 55° C. {close oversize brace} 30 cycles Extend 1.5 min 72° C. Final extension 5 min 72° C.
[0515]Ten microliters of samples from row A, well 1-12 from each plate were analyzed by 1.5% agarose gel electrophoresis using ethidium bromide for visualization, in order to verify that the multiplex overlap-extension RT-PCR was successful. The expected size of the overlap-extension fragment was approximately 1070 bp (the exact size depends upon the lengths of the variable regions). FIG. 17 shows samples from eight 96 well plates. The average number of successful overlap-extension RT-PCR fragments from fifty 96 well plates, was estimated to eight per plate. Thus, the total number of successful overlap-extension RT-PCR fragments were estimated to approximately 400.
[0516]Ten microliters from all the performed reactions, run in the 96 well plates, originating from the same donor, were consolidated in a single tube. An aliquot consisting of 200 μl of the pooled PCR products were subsequently purified using QIAquick PCR purification kit according to the manufactures procedure (Qiagen cat. No. 28106, Hilden, Germany) using 60 microliters buffer EB for elution. The purified pool of overlap PCR products was digested with XhoI and NotI and subsequently purified by preparative 1% agarose gel electrophoresis; the overlap-extension fragments are excised from the agarose gel and purified using a Qiaex II kit (Qiagen cat. No 20051, Hilden, Germany).
[0517]f. Cognate Fab Expression Library
[0518]A Fab expression library was generated by a two-step ligation procedure. Initially the pool of digested overlap-extension fragments described above was ligated into the XhoI/NotI digested E. coli vector JSK301 (FIG. 11). The ligation reaction was subsequently transformed into electrocompetent E. coli cells (XL1-Blue electroporation competent, Stratagen, cat. No. 200228, La Jolla, USA), according to manufactures instructions. The to transformed E. coli cells were plated onto 2×YT agar containing 100 μg/ml carbenicillin.
[0519]Biomass corresponding to approximately 1010-1011 cells originating from a number of independent colonies that exceeded at least 5 times the total number of overlap PCR products was used as starting material for plasmid preparation using Qiagen Plasmid preparation Maxi kit (Qiagen cat. No. 12163, Hilden, Germany). To enable Fab expression of the cloned cognate linked VH and VL encoding sequences, a prokaryotic promoter and leader cassette was inserted in a second ligation step. The used bi-directional promoter fragment (SEQ ID NO 321) was extracted from the parental JSK301 by AscI/NheI digestion. The purified pool of plasmids was likewise digested with AscI/NheI restriction endonucleases and gel purified as previously described. The purified fragments were subsequently ligated and transformed into electrocompetent E. coli cells (TG1, Stratagene, cat. No. 200123, La Jolla, USA) and plated on 2×YT agar containing 100 μg/ml carbenicillin, and 1% glycose. The Fab expression library generation process is outlined in FIG. 12.
[0520]g. Screening of Clones
[0521]Fab expressing clones were screened for TT antigen-binding by antigen-specific ELISA assays.
[0522]g-1. Master Plate Generation and Fab Expression
[0523]Individual selected colonies of the TG1 cells, each harbouring a cognate Fab expression vector from the library generated as described in section (f) were picked into single wells of 96 well plates containing 2×YT/100 μg/ml Carb/1% glucose. The plates were incubated overnight at 37° C. with gentle shaking. Four 96 well plates were consolidated into the wells of one 384 well plate, containing 2×YT/100 μg/ml Carb/1% glucose, using 96 pin replicators. The 384 well plates were incubated overday at 37° C. These plates are referred to as master plates, and they were stored at -80° C. after addition of glycerol to a final concentration of 15%.
[0524]The masterplates were used for inoculation of 384 DeepWell plates containing 2×YT/100 μg/ml-Carb 0.1% glucose using a 96 pin replicator. The plates were sealed and incubated for 2-3 h at 37° C. with shaking.
[0525]Fab expression was induced by adding an equal volume of 2×YT/100 μg/ml Carb/0.2 mM IPTG, obtaining a final IPTG concentration of 0.1 mM. The plates were sealed and incubated overnight at 30° C. with shaking. The following day, the Fab-containing supernatants were analyzed for TT antigen-binding specificity by ELISA.
[0526]g-2. ELISA Analysis
[0527]Three hundred and eighty four (384) well ELISA plates (Corning Inc., Corning, N.Y., USA, cat. No 3700) were coated overnight at 4° C. with Tetanus Toxoid (TT) antigen, diluted to a final concentration of 1 μg/ml in PBS in a volume of 25 μl per well. Excess binding sites of the wells were blocked for 1 h at room temperature (RT) by adding 2% M-PBS-T (2% skim milk powder in PBS, 0.05% Tween 20). The wells were washed 2 times with PBS-T (PBS, 0.05% Tween 20).
[0528]The Fab-containing bacterial supernatants from section (g-1) were diluted 1:2 in 2% M-PBS-T, and transferred to the ELISA wells in duplicate. Incubation was performed for 1 h at RT. The wells were washed 4 times with PBS-T. Goat-anti-human Fab/HRP (Sigma, St. Louis, Mo., USA, cat. No A0293) 1:10.000 dilution in 2% M-PBS-T was added to the wells. Incubation was performed for 1 h at RT. The wells were washed 4 times with PBS-T. TMB Plus (KemEnTec, Copenhagen, Denmark, cat. No 4390L) substrate was added and incubation was performed for 5-15 minutes. The reactions were stopped by adding an equal volume of 1 M H2SO4, and analyzed using a spectrophotometer at 450 nm (Multiscan Ascent, Labsystems, Franklin, USA).
[0529]The original bacterial clones corresponding to TT antigen-binding clones may subsequently be retrieved from the original master plates. Plasmid DNA can be prepared from the isolated antigen positive Fab clones, generating a cognate Fab expression sub-library of TT antigen-binding clones.
[0530]A subset of the clones were further analyzed by an anti-kappa ELISA assay, in order to get a correlation between the number of antigen-binding clones and the number of Fab expressing clones. The anti-kappa assay was generally performed as the TT-assay, except that the wells were coated with a 1:1000 dilution of goat anti-human kappa antibody (Caltag, Calif., USA, Cat. No H16000), using a carbonate buffer, pH 9.6.
[0531]g-3. Screening Results
[0532]Clones from four 384 well plates were screened for reactivity with anti-kappa Ab and TT using ELISA assays according to the procedure described in g-2. The results obtained results are summarized in the tables 15 through 19. The anti-kappa ELISA results inform about the expression of Fab fragments in a given clone, the TT ELISA results inform about the functionality of the Fab fragments.
TABLE-US-00036 TABLE 15 ELISA screening, anti-kappa coat (1440 clones in total) Plate # 2 × background 3 × background 4 × background G050 105 79 73 G051 93 79 68 G052 120 96 73 G053 164 141 125 Sum 482 395 339 % Fab pos. clones 34% 27% 24%
[0533]Of 1440 single clones analyzed, 482 clones or 34% exhibited anti-Kappa reactivity at a level exceeding 2× background reactivity. 395 clones or 27% of the clones showed reactivity above 3× background, etc.
[0534]The same clones were analyzed for TT-antigen reactivity by ELISA, and results are given in table 16 below:
TABLE-US-00037 TABLE 16 ELISA screening, TT coat (1440 clones in total) Plate # 2 × background 3 × background 4 × background G050 26 19 17 G051 24 18 15 G052 34 31 24 G053 46 36 30 Sum 130 104 86 % TT pos. clones 9.0% 7.2% 6.1%
[0535]From this table it is seen that 9.0% clones showed reactivity with TT at 2× background (defined as the signal obtained by the reactivity of a Fab fragment with a irrelevant specificity). 104 clones reacted at 3× background (7.2%) etc.
[0536]Of the Fab-positive clones (482 clones in total), approximately 27% of the clones (130/482) exhibited reactivity with TT at the 2× background level. This level does not change significantly at the other background levels.
[0537]Six 384 well plates were screened for clones with TT reactivity. The number of clones giving rise to reactivity with the antigen are shown in the table 17. Anti-kappa ELISA was not carried out with these plates.
TABLE-US-00038 TABLE 17 ELISA screening, TT coat (2160 clones in total) Plate # 2 × background 3 × background 4 × background G054 82 65 52 G055 30 24 21 G056 25 23 21 G057 30 28 26 G058 24 21 19 G059 18 14 13 Sum 209 175 151 % TT pos. clones 9.7% 8.1% 7.0%
[0538]The percentage of TT positive clones in plates G054-G059 was comparable to what was found in plates G050-G053 (table 16).
[0539]The results from all the clones screened against TT (table 16 and 17) are summarized in table 18.
TABLE-US-00039 TABLE 18 All clones screened, TT reactive clones Plates 2 × background 3 × background 4 × background G050-G053 130 104 86 G054-G059 209 175 151 Sum 339 279 237 % of total (3600) 9.4% 7.8% 6.6%
[0540]In summary, a total of 339 clones showed reactivity with TT at least at 2× background levels (table 18). This corresponds to 9.4% of all clones screened.
[0541]All positive clones exhibiting reactivity with TT at 2× background was inoculated into 96 well plates as previously described, from the master plate. The next day, the bacteria was collected by centrifugation at 4000 rpm for 15 minutes, and the pellet was resuspended in 0.8 mM EDTA, 0.4×PBS, 0.8 M NaCl, and incubated on ice for 15 minutes. The periplasmic extract was collected by centrifugation, and the reactivity of the clones was further analyzed. Here we show results from one such plate (G060) for reactivity with anti-Kappa (FIG. 18), Ovalbumin (unrelated antigen) (FIG. 19), TT (FIG. 20), and a one-step competitive assay using 10-7 M concentration of TT-antigen in solution (FIG. 21). These ELISA assays were performed on the same periplasmic extract, at the same dilution.
[0542]Most of the clones expressed Fab fragments (90/96) (FIG. 13), and no clones reacted with Ovalbumin (FIG. 19). The reactivity with immobilized TT was reduced or completely inhibited by TT in solution (FIG. 21), indicating that the clones react specifically with TT. The reactivity of the clones in plate G060 is summarized in table 19.
TABLE-US-00040 TABLE 19 Summary of G060 plate (96 clones in total) Background level Anti-Kappa positive TT positive 2× 94% (90/96) 74% (71/96) 3× 91% (87/96) 72% (69/96) 4× 84% (81/96) 69% (66/96)
[0543]h. Diversity Analysis and Clone Approval
[0544]Plasmid isolated from 47 clones (from plate G060) from the cognate TT antigen-binding Fab expression sub-library were subjected to sequence analysis. The variable heavy chain encoding sequences were sequenced using primer LSN-HCP: AGGAAACAGGAGATATACAT (SEQ ID NO131), annealing to the P tac promoter, and the light chain encoding sequences were sequenced using primer LSN-LCP: TCGCCAAGGAGACAGTCATA (SEQ ID NO132), annealing to the P lac promoter. The sequence data was analysed using the Vector NTI software (Informax, Frederick, Md., USA). The resulting sequence data of the heavy chain was trimmed to one base pair 5' of the upstream AscI restriction site and immediately 3' of the downstream XhoI restriction site. The light chain sequence data was trimmed to the second 5' base pair of the upstream NheI restriction site and 3' of the last codon "AAA" encoding the variable chain C-terminal lysine.
[0545]The trimming was performed in order to facilitate further analysis such as translation of each DNA sequence.
[0546]The variable heavy and light chain encoding sequences were analyzed for germline gene usage by comparing the sequences to the V-base germline variable region sequence database (MRC Centre for Protein Engineering, Cambridge, UK). The closest related germline allele was thus determined for each sequence showing a V-gene repertoire originating from 12 different variable heavy chain germline alleles belonging to VH-family VH1 through VH5 and 8 different variable kappa light chain germline alleles belonging to VKI, VKIII and VKIV (Table 20).
[0547]Furthermore, the variable gene sequences were translated into protein sequences which were aligned using the AlignX software (Informax, Frederick, Md., USA) as depicted in FIG. 22 and FIG. 23. Based on the protein sequence alignments, the variable chain sequences could be categorized into groups according to V(D)J rearrangement events, designated with H for the variable heavy chain sequence and L for the variable kappa chain sequence followed by a unique number. In each rearrangement group, the sequences were categorized according to the maturation genotype (M-type in table 20) within the CDR1, 2 and 3 regions, represented by a lower case letter in alphabetical order. Seven of the clones had premature stop codons, Whereof 6 clones were an amber mutation (TAG) (clone IDs g060:b12, d08, f06, c12, f03, c04) and 1 clone was an opal mutation (TGA) (clone ID g060: h12). These codons were most likely suppressed by E. coli resulting in functional Fab fragments. Four of these clones were members of groups containing similar clones making them redundant. Additional clones would have to be analyzed in order to replace the remaining 3 clones with premature stop codons as they were single members of their groups. Alternatively, the sequences could be corrected by standard molecular biology techniques such as PCR, replacing the stop codon with an appropriate codon.
[0548]The 47 analyzed V-region sequences could be divided into 20 unique V(D)J rearrangement groups (designated "groups" in table 20) both for the variable heavy and variable kappa gene. Four of the rearrangement groups could further be divided into 1 to 4 maturation types (a through d), resulting in 27 unique antibody encoding sequences (Table 12). Generally, specific heavy chain rearrangement groups combine with specific light chain rearrangement groups (e.g. H1 with L1, H12 with L24 and so forth). Further, the maturation type match among pairs of variable heavy chain and the variable light encoding sequences (e.g. H4c match with L13c). Such stringency in paring among rearrangement groups and maturation types indicating cognate paring of the variable region encoding sequences.
[0549]However, heavy chain rearrangement group H4 is an exception. H4 pairs with two different light chains L28 and L13. L13 was unique to H4, comprising maturation types a, b and c that matches with L13 maturations types. L28, on the other hand, was also found paired with the single member in heavy chain group H2. Two out of seven H4a heavy chain sequences pair with heavy chain L13a and five out of seven H4a heavy chain sequences pair with L28a. In summary, these observations suggested a multiplex overlap-extension RT-PCR event where two TT-specific antibody producing plasma cells, with the genotype combinations H2-L28 and H4a-L13a, were present in a single well. Rare scrambling events of the light chain and heavy chain gene pairing, such as for H2 and H4a with L28, was expected as the experiment was based on limiting dilution of plasma cells.
[0550]The sequence identity among individual cognate pairs of the variable heavy chain and variable light chain encoding sequences from the same group and maturation type is at least 90% and preferably at least 95%. Take for example g060g03 and g060a01 from group H1 maturation type a. Clone g060g03 correspond to nucl. SEQ ID pair 168:215, where the variable heavy chain encoding sequence correspond to SEQ ID NO 168 and the variable light chain encoding sequences correspond SEQ ID NO 215. Clone g060a01 correspond to nucl. SEQ ID pair 133:180. When SEQ ID NO 168 is aligned with SEQ ID NO 133 (the variable heavy chains) 4/369 bases are not identical and for the variable lights chains (SEQ ID NOs 215 and 180) 8/327 bases are not identical, this correspond to a sequence identity of 98.3% between these two cognate pairs (g060g03 and g060a01). However, when looking at sequence identity among different groups it is not expected that the sequence identity will be high, since it is a polyclonal sub-library and diversity is desired. In this particular example the lowest identity among the cognate pairs is approximately 40% (e.g. g060b11 and g060h11 have a sequence identity of 39.5%).
[0551]One embodiment of the present invention is a sub-library of cognate pairs of immunoglobulin heavy chain variable region and light chain variable region encoding sequences, where the immunoglobulins obtainable from said library are capable of reacting with or binding to Tetanus Toxin.
[0552]A further embodiment of the present invention is such a sub-library of cognate pairs of immunoglobulin heavy chain variable region and light chain variable region encoding sequences, comprising individual cognate pairs with at least 90% sequence identity with one individual SEQ ID pair, selected from the group consisting of SEQ ID pairs 135:182, 168:215, 146:193, 151:198, 173:220, 152:199, 164:211, 148:195, 137:184, 169:216, 138:185, 143:190, 161:208, 166:213, 157:204, 139:186, 134:181, 150:197, 156:203, 158:205, 170:217, 178:225, 141:188 or 144:191.
TABLE-US-00041 TABLE 20 VH Vkappa Clone ID Group M-type Family Germ-line Seq nucl. Seq prot. Group M-type Family Germ-line Seq nucl. Seq prot. g060a03 H1 b VH1 DP-14 135 229 L1 b VKI DPK9 182 276 g060g03 H1 a VH1 DP-14 168 262 L1 a VKI DPK9 215 309 g060a01 H1 a VH1 DP-14 133 227 L1 a VKI DPK9 180 274 g060b12 H11 a VH3 DP-77 145 239 L10 a VKIII DPK22 192 286 g060c03 H11 a VH3 DP-77 146 240 L10 a VKIII DPK22 193 287 g060d08 H11 a VH3 DP-77 154 248 L10 a VKIII DPK22 201 295 g060f06 H11 a VH3 DP-77 162 256 L10 a VKIII DPK22 209 303 g060d04 H12 a VH1 DP-10 151 245 L24 a VKI DPK4 198 292 g060h02 H12 a VH1 DP-10 175 269 L24 a VKI DPK4 222 316 g060h10 H12 a VH1 DP-10 177 271 L24 a VKI DPK4 224 318 g060g11 H12 b VH1 DP-10 173 267 L24 b VKI DPK4 220 314 g060d05 H12 c VH1 DP-10 152 246 L24 c VKI DPK4 199 293 g060f09 H12 d VH1 DP-10 164 258 L24 d VKI DPK4 211 305 g060c07 H15 a VH5 DP-73 148 242 L16 a VKIII DPK22 195 289 g060d11 H15 a VH5 DP-73 155 249 L16 a VKIII DPK22 202 296 g060g02 H15 a VH5 DP-73 167 261 L16 a VKIII DPK22 214 308 g060a09 H16 -- VH1 DP-73 137 231 L4 -- VKI DPK9 184 278 g060g04 H17 a VH4 DP-66 169 263 L3 a VKI DPK9 216 310 g060h04 H17 a VH4 DP-66 176 270 L3 a VKI DPK9 223 317 g060a10 H18 a VH2 VII-5 138 232 L21 a VKI L12a 185 279 g060h12 H18 a VH2 VII-5 179 273 L21 a VKI L12a 226 320 g060b09 H19 a VH2 VII-5 143 237 L23 a VKI DPK8 190 284 g060f10 H19 a VH2 VII-5 165 259 L23 a VKI DPK8 212 306 g060f05 H2* -- VH1 DP-88 161 255 L28* a VKIV DPK24 208 302 g060f11 H21 a VH3 DP-32 166 260 L31 a VKI DPK9 213 307 g060g12 H21 a VH3 DP-32 174 268 L31 a VKI DPK9 221 315 g060f01 H10 -- VH3 COS-8 157 251 L7 -- VKI DPK8 204 298 g060a11 H23 a VH3 DP-77 139 233 L5 a VKI DPK9 186 280 g060c12 H23 b VH3 DP-77 149 243 L5 b VKI DPK9 196 290 g060f03 H24 -- VH3 DP-38 159 253 L20 -- VKI DPK8 206 300 g060a02 H3 -- VH3 DP-77 134 228 L9 -- VKI O16 181 275 g060d03 H4 a VH1 DP-88 150 244 L13 a VKIII DPK22 197 291 g060f08 H4 a VH1 DP-88 163 257 L13 a VKIII DPK22 210 304 g060b05 H4* a VH1 DP-88 140 234 L28* a VKIV DPK24 187 281 g060b07 H4* a VH1 DP-88 142 236 L28* a VKIV DPK24 189 283 g060f04 H4* a VH1 DP-88 160 254 L28* a VKIV DPK24 207 301 g060g10 H4* a VH1 DP-88 172 266 L28* a VKIV DPK24 219 313 g060d07 H4* a VH1 DP-88 153 247 L28* a VKIV DPK24 200 294 g060e05 H4 c VH1 DP-88 156 250 L13 c VKIII DPK22 203 297 g060f02 H4 b VH1 DP-88 158 252 L13 b VKIII DPK22 205 299 g060g09 H4 c VH1 DP-88 171 265 L13 c VKIII DPK22 218 312 g060c04 H6 -- VH1 DP-88 147 241 L12 -- VKIII DPK22 194 288 g060g07 H20 -- VH4 DP-71 170 264 L8 a VKI DPK9 217 311 g060h11 H7 -- VH2 VII-5 178 272 L18 -- VKI L12a 225 319 g060b06 H8 a VH3 DP-46 141 235 L17 a VKI DPK1 188 282 g060a04 H8 a VH3 DP-46 136 230 L17 a VKI DPK1 183 277 g060b10 H9 -- VH1 DP-14 144 238 L30 -- VKI DPK1 191 285 Asterisk indicates V encoding sequences implicated in scrambling. Seq prot. indicates SEQ ID numbers from the sequence list corresponding to the protein sequences and Seq nucl. indicate SEQ ID numbers corresponding to the nucleic acid sequence in the sequence list
[0553]i. Apparent Affinities
[0554]A competition assay was set up in order to determine the apparent affinity or IC50 of selected clones from plate G060.
[0555]Briefly, Fab fragments were expressed in 50 ml cultures as follows: 50 ml 2×YT/100 μg/ml Carb/0.1% glucose was added 0.5 ml overnight culture, and shaken for approximately 2 h at 37° C. IPTG was added to a final concentration of 0.1 mM, and shaking was continued overnight at 30° C. The next day, the bacteria were collected by centrifugation at 4000 rpm for 15 minutes, and the pellet was resuspended in 1 ml 0.8 mM EDTA, 0.4×PBS, 0.8 M NaCl, and incubated on ice for 15 minutes. The periplasmic extract was collected by centrifugation, and stored at -20° C.
[0556]The competition assay was performed as follows: the periplasmic extracts in appropriate dilutions estimated by titration were added to a series of tubes. As a positive control the phage display derived Fab fragment mp584, derived from the human hybridoma cell line HB8501 expressing an anti-TT antibody was used. Soluble TT was added to the first tube at 100 nM concentration, and subsequently diluted at four-fold steps in the following tubes, making a total of seven dilutions of TT (from 100 nM to 25 μM). The reactions were incubated for approximately 45 minutes at room temperature. The samples were transferred to ELISA plates coated with TT at 1 μg/ml, and blocked as previously described. The plates were incubated for 1 h at room temperature, followed by 4× wash with PBS-T, goat-anti-human Fab/HRP was added at a 1:10.000 dilution, and incubated for 1 h. TMB Plus substrate (KemEnTech, Denmark, cat. No. 4390A) was added, incubation was performed for approximately 10 minutes, and the reactions were stopped with 1 M H2SO4. The plates were read at 450 nm. The data are plotted in FIG. 24.
[0557]The apparent affinities of the analyzed clones are given in the table 21:
TABLE-US-00042 TABLE 21 Clone Apparent affinity g060a10 0.50 nM g060b06 0.60 nM g060b12 1.2 nM g060c04 0.55 nM g060d04 1.1 nM g060f01 1.2 nM g060g04 0.9 nM g060g07 0.35 nM g060h11 3.0 nM mp584 (pos. cont.) 6.0 nM
[0558]As seen in table 21, the Fab fragments have apparent affinities in the lower nano-molar or upper pico-molar range. Furthermore, all cognate paired Fab fragments exhibit a higher apparent affinity than that of the phage display derived Fab fragment.
[0559]j. Summary
[0560]In donor TT03 the frequency of TT specific plasma cells in the peripheral blood monocyte cell fraction was calculated to 0.022%. Approximately 400 cognate pairs were generated from TT03. 3600 clones from the cells transformed with the cognate pair library were screened using ELISA, of these 339 clones showed TT reactivity in the ELISA screening. 47 of these clones have been analysed with respect to their clonal diversity. Of these 47 clones, 27 proved to resemble unique non-scrambled variable region encoding sequences. Three of these clones contained a premature stop codon that will need correction before transfer to a mammalian expression vector. The transfer to mammalian expression vectors was described in example 1 section h. The apparent affinities were measured on selected clones, ranging from the lower nano-molar to upper pico-molar range.
[0561]k. Prospects
[0562]The Tetanus Toxin is one of the most toxic substances known with a lethal dose of a few nanogram. The toxin is produced by Clostridium tetani, a soil bacteria also present in the digestive tract of up to 25% of humans. The tetanus immunization program has effectively abolished the disease in the western world, although 100-200 cases are still observed in the major western countries annually, with a case-fatality ratio of 50%. In the developing world the number of cases is significantly greater. Bacterial growth in contaminated penetrating wounds may lead to toxin release, ultimately leading to rigidity, spasms, respiratory arrest and death.
[0563]Hyperimmune immunoglobulin products isolated from human blood donors with a high titer of antibody response against Tetanus Toxoid may be used to prevent tetanus or if instituted early to treat established tetanus, also in conjunction with active immunization. However, due to shortage of human product, equine hyperimmune anti-Tetanus Toxoid is used in the developing world. Recombinant monoclonal or polyclonal antibodies against Tetanus Toxoid have potential to substitute hyperimmune globulin products for therapeutic and/or prophylactic use. Recombinant monoclonal antibodies originating from the conventional hybridoma technology has been described to be effective against TT (Chin, J. et al. 2003. Biologicals 31, 45-53). Interestingly, a synergistic effect was observed when mixing two monoclonal antibodies. Thus, a recombinant polyclonal anti-Tetanus Toxoid antibody capable of reacting with or binding to Tetanus Toxin could potentially be very effective in treatment or prophylaxis protection of patients at risk of developing tetanus.
[0564]One embodiment of the present invention is a recombinant polyclonal immunoglobulin or fragments thereof capable of reacting with or binding to Tetanus Toxin.
[0565]A preferred embodiment of the present invention is a recombinant polyclonal immunoglobulin capable of reacting with or binding to Tetanus Toxin comprised of cognate pairs of immunoglobulin heavy chain variable region and light chain variable region.
[0566]A further embodiment of the present invention is a recombinant polyclonal immunoglobulin capable of reacting with or binding to Tetanus Toxin obtained by the method according to the present invention.
[0567]A further embodiment of the present invention is a recombinant polyclonal immunoglobulin capable of reacting with or binding to Tetanus Toxin comprising individual cognate pairs of immunoglobulin heavy chain variable region and light chain variable region with at least 90% sequence identity with one individual SEQ ID pair, selected among SEQ ID pairs 229:276, 262:309, 240:287, 245:292, 267:314, 246:293, 258:305, 242:289, 231:278, 263:310, 232:279, 237:284, 255:302, 260:307, 251:298, 233:280, 228:275, 244:291, 250:297, 252:299, 264:311, 272:319, 235:282 or 238:285.
[0568]Another embodiment of the invention, is a pharmaceutical composition comprising a recombinant polyclonal antibody capable of reacting with or binding to Tetanus Toxin as active ingredient intended for the treatment or prevention of a tetanus. Preferably, the recombinant polyclonal antibody is combined with a pharmaceutically acceptable excipient.
[0569]A further embodiment of the present invention is the use of a recombinant polyclonal immunoglobulin capable of reacting with or binding to Tetanus Toxin as a medicament for the treatment or prophylactic protection of a patient at risk of developing tetanus.
[0570]An additional embodiment is a method of preventing or treating a patient at risk of developing tetanus by administering to a patient in need thereof a composition comprising a recombinant polyclonal antibody capable of reacting with or binding to Tetanus Toxin.
Example 12
Comparing Results Obtained from Two Donors
[0571]In the following example results obtained from donor TT08 are compared with the results obtained from donor TT03 in Example 11. The results are summarized in table 22.
TABLE-US-00043 TABLE 22 TT03 TT08 TT specific plasma cells in PBMC fraction (ELISPOT) 0.021% 0.011% CD19+ cells in PBMC fraction (FACS) 11% 5% Plasma cells in PBMC fraction (FACS CD38hi, CD45in) 0.12% 0.08% Estimated number of linked VH and VL sequences 400 400 Number of clones screened for Fab expression 1400 2000 Number of Fab positive clones (2 × background) 482 558 Number of clones screened for TT-specificity 3600 3400 Number of TT-specific clones (2 × background) 339 188 Number of sequences analyzed from TT-specific clones 47 102 VH germline alleles 12 (VH1-5) NA VL germline alleles 8 (VKI, III, IV) NA Number of unique cognate pairs 27 40 Apparent affinity range 0.35-3.0 nM NA NA = not analyzed
[0572]This clearly illustrated that libraries of similar quality could be isolated from two different donors immunized with TT.
[0573]The reason for the higher number of unique cognate pairs in the library obtained from donor TT08 is most likely due to the larger number of sequences analyzed.
Example 13
Comparing the Library of Cognate Pairs from Example 11 with a Combinatorial Phage Display Library Generated from the Same Donor
[0574]In the present example a combinatorial phage display library was prepared from the same donor previously used to prepare the library of cognate pairs, in order to compare library diversity, affinity and specificity between libraries of cognate pairs and combinatorial libraries.
[0575]a. Combinatorial Phage Library Construction
[0576]A phage display library was generated from the CD19.sup.+ fraction of cells from donor TT03 (identical to the cell fraction obtained in Example 11c).
[0577]Total RNA was prepared from app. 5×106 CD19.sup.+ cells using NucleoSpin RNA L kit (Machery-Nagel cat. no. 740 962.20). cDNA was subsequently synthesized in an oligo(dT)-primed reaction using ThermoScript reverse transcriptase (Invitrogen cat. no. 11146-016).
[0578]The VH and Kappa chains were PCR amplified using HotMasterTaq DNA polymerase (Eppendorf cat. no. 0032 002.692) and primers essentially as described by de Haard et al. (J. Biol. Chem. 274, 18218-18230; 1999), only modified with respect to restriction enzyme recognition sequences in the 5'-end.
[0579]The combinatorial phage display library was generated by successive insertion of Kappa and VH PCR products into Em351 phage display vector (modified from phh3 described in Den, W. et al. 1999 J. Immunol. Methods 222, 45-57).
[0580]The final library was electroporated into TG1 E. coli strain (Stratagene). The size of the combinatorial library contained 3×106 independent clones with a high insert frequency.
[0581]b. Panning of the Combinatorial Library
[0582]Fab displaying phage particles were prepared according to standard procedures (e.g. Antibody Engineering, A Practical Approach 1996, ed. McCafferty, Hoogenboom and Chiswell).
[0583]Panning was performed on tetanus toxoid (TT; SSI batch no. 89-2) diluted to 1 μg/mL in PBS, and immobilized in MaxiSorp immunotubes (Nunc cat. no. 444202). Following a one hour incubation period and several washing steps, bound phage particles were eluted using 100 mM TEA (Triethylamine).
[0584]The phage particle eluate was neutralized, and used to infect exponentially growing TG1 cells, from which Fab displaying phage particles enriched for TT specificity were obtained. A second round of panning using the eluted phages was performed following the general procedure outlined above.
[0585]In parallel, three panning rounds were performed on the C-fragment of the tetanus toxin molecule (Sigma cat. no. T3694), following the procedure outlined above.
[0586]Single colonies were screened for binding to tetanus toxoid and the C-fragment from the unselected library, and after each panning round, from both sets of panning described above.
[0587]c. Comparing Specificity and Affinity of Combinatorial Phage Particle Clones and Cognate Pair Clones
[0588]Single colonies were picked from the unselected library and after each round of panning (on intact tetanus toxoid (TT) and the tetanus toxoid C-fragment, respectively). Fab displaying phage particles were analyzed for reactivity by ELISA assays. The number of Fab-positive clones exhibiting TT and/or C-fragment reactivity of at least 2× background reactivity and at least 4× background reactivity are given in table 23 to 25 below. All the results are shown as number of specific clones/number of Fab positive clones.
TABLE-US-00044 TABLE 23 Clones selected on TT and analyzed by TT-specific ELISA Enrichment 2 × background 4 × background Unselected clones -- 13/app. 5000 After 1 round of panning 61/99 59/93 After 2 rounds of panning 165/169 163/166
TABLE-US-00045 TABLE 24 Clones selected on TT and analyzed by C-fragment-specific ELISA Enrichment 2 × background 4 × background Unselected clones 0/13 0/13 After 1 round of panning 0/85 0/85 After 2 rounds of panning 0/85 0/85
TABLE-US-00046 TABLE 25 Clones selected on C-fragment, analyzed by C-fragment-specific ELISA Enrichment 2 × background 4 × background After 1 round of panning 1/35 0/35 After 2 rounds of panning 12/47 9/47 After 3 rounds of panning 64/126 26/126
[0589]Panning of the phage display library against TT revealed an increasing number of TT specific clones when increasing the number of panning rounds, and only a few clones were identified in the unselected library. No clones were found to be reactive with the teatanus toxoid C-fragment from the unselected library or after two rounds of panning of the library against TT. To obtain Fab fragments with C-fragment specificity from the combinatorial phage display library, this library had to be panned against the C-fragment specifically.
[0590]For comparison, thirteen TT-specific clones from Example 11 were subjected to C-fragment-specific ELISA, of these seven showed reactivity above 2× background. Thus, Fab fragments with specificity towards the tetanus toxin C-fragment could be expressed from a library of cognate pairs obtained from a TT-immunized individual.
[0591]This clearly illustrates the disadvantage of using panning to identify clones with specificity towards a particular antigen. If important antigen fragments or epitopes are unknown, they may be discharged during panning, thereby resulting in a less efficient product in the end.
[0592]Further, apparent affinities of the combinatorial clones were measured as described in the assay of Example 11i. Ten of the combinatorial clones obtained after two rounds of panning on TT were analyzed, revealing apparent binding affinities between 1 and 15 nM.
[0593]For comparison four of the nine clones analyzed from the library of cognate pairs (table 21), showed affinities in the pico-molar range.
[0594]This indicates that the pairing of variable regions as originally selected for by the donors immune system, in combination with the somatic hyper-mutations the pairs have been subjected to as a pair, potentially results in higher apparent binding affinities than random combinations of such variable regions.
[0595]d. Comparing Sequences of TT Specific Combinatorial Phage Particle Clones and Cognate Pair Clones
[0596]The large amount of sequence data generated from the two libraries precludes direct comparisons of raw sequences. In order to visualize the difference between VH and VL sequence pairs in the phage display library an in the library of cognate pairs, phylogenetic threes were generated for the VH and VL sequences, and pairs were illustrated in a dot matrix. The pairing of phylogenetic information in a dot matrix (FIG. 25) revealed very different distribution profiles of VH and VL sequence pairs in the two libraries. The scattered appearance of the VH and VL sequence pairs in the phage display library (FIG. 25A) indicated little phylogenetic relation between VH and VL genes in agreement with the random paring of the V genes in this library. In contrast, VH and VL sequence pairs from the library of cognate pairs (FIG. 25B) show a clustered appearance indicating co-evolution of V genes as expected for cognate pairs. Also, the genetic diversity is much greater for the V-genes in the library of cognate pairs compared to the V-genes from the combinatorial library, indicating that the method of isolating cognate paired V-genes is less biased.
Example 14
Combined Single-Step Multiplex Overlap-Extension RT-PCR and Nested PCR Using T-Cells as Template Source
[0597]In this example, it is described how a single-step multiplex overlap-extension RT-PCR can be performed on T lymphocytes derived from a human donor.
[0598]a. Obtaining a Lymphocyte Containing Cell Fraction
[0599]A blood sample is obtained from human donors who have been subjected to the desired antigen, for example by immunization, natural infection, malignancy, through an autoimmune reaction, or other diseases. The peripheral blood mononuclear cells (PBMC's) are isolated using Lymphoprep (Axis-Shield, Oslo Norway, prod. No 1001967) according to the manufacturer's instructions.
[0600]In the present example antigen-specific T cells are generated by further stimulation of the PBMC fraction. However, the multiplex overlap-extension RT-PCR can also be performed directly on single cells from the PBMC fraction or on a cell fraction enriched for T cells (e.g. by FACS-sorting for CD3-positive cells).
[0601]b. Generating Antigen-Specific T Cells
[0602]The PBMC cell fraction is re-suspended in an appropriate culture medium containing relevant cytokines such as IL2. Further, the desired antigen is added to the culture, where it will be presented to the T lymphocytes by antigen presenting cells (APC) present in the PBMC fraction. Alternatively, APC feeder cells treated in beforehand such that they are presenting the desired antigen may be added to the PBMC fraction. Such APC feeder cells can be APC exposed to the desired antigen in the form of for example peptide, protein or other molecular form, or microbially infected APC or cells transfected to express and present the antigen, or APC co-cultured with other antigenic cells, e.g. such as cancer cell in the than of primary tissue or cell lines. Many different types of APC feeder cell are known from the literature, including transformed cell lines, B cell lines, dendritic cells, etc.
[0603]The PBMC fraction is cultivated for approximately 3 to 5 weeks, during which fresh antigen presenting cells are added together with cytokines, e.g. on a weekly basis. This results in T lymphocyte proliferation, activation and maturation. At the end of the cultivation period the cell culture will be dominated by antigen-specific T cells. Whether these cells are CD4+ or CD8+ depends on the disease, the antigen, the APC cells, and the cytokine mixtures used during the stimulation period.
[0604]The antigen specificity can for example be tested with a CTL assay, proliferation assays and MHC tetramers loaded with the desired antigen (Altman, J. D., et al. 1996. Science 274, 94-96).
[0605]The antigen-specific T cells are distributed to PCR tubes, either by limiting dilution or using a FACS in order to obtain a single cell pr. vessel. The vessel can be stored at -80° C. until use.
[0606]b. Single-Step Multiplex Overlap-Extension RT-PCR
[0607]The Qiagen One-Step RT-PCR kit (Qiagen cat. No 210212, Hilden, Germany) is used for the multiplex overlap-extension RT-PCR essentially according to the manufacturer's recommendation. Before addition of PCR reaction mixture to the PCR tubes, the cells are thawed.
[0608]PCR reaction mixtures and cycling conditions are initially set up as described in Example 11 e-1. However, a certain amount of optimization may be expected for a new set of primers.
[0609]The multiplex overlap-extension primer mix used comprise the primers shown in table 26.
TABLE-US-00047 TABLE 26 Primer SEQ ID name Conc. NO primer sequence in 5' to 3' direction Vα 10 nM 322 tattggcgcgccatggccGCCCAGTCTGTGACCCAGC of each 323 tattggcgcgccatggccGCCCAGAAGATAACTCAA 324 tattggcgcgccatggccGATGCTAAGACCACCCAG 325 tattggcgcgccatggccGCCCAGACAGTCACTCAG 326 tattggcgcgccatggccAAACAGGAGGTGACACAGA 327 tattggcgcgccatggccGGAGACTCGGTTACCCAGA 328 tattggcgcgccatggccGGAGATTCAGTGACCCAGA 329 tattggcgcgccatggccAGCAATTCAGTCAAGCAGA 330 tattggcgcgccatggccGGACAAAACATTGACCAG 331 tattggcgcgccatggccAAAAATGAAGTGGAGCAGA 332 tattggcgcgccatggccGAAGACCAGGTGACGCAGA 333 tattggcgcgccatggccGGAATACAAGTGGAGCAGA 334 tattggcgcgccatggccCTACATACACTGGAGCAGA 335 tattggcgcgccatggccGAAGACAAGGTGGTACAAA 336 tattggcgcgccatggccGAAAACCAGGTGGAGCACA 337 tattggcgcgccatggccCAGGAGAATGTGGAGCAG 338 tattggcgcgccatggccGGAGAGAGTGTGGGGCTG 339 tattggcgcgccatggccGGAGAGGATGTGGAGCAGA 340 tattggcgcgccatggccAGCCAAAAGATAGAACAGA 341 tattggcgcgccatggccAGTCAACAGGGAGAAGAG 342 tattggcgcgccatggccCAACAACCAGTGCAGAGT 343 tattggcgcgccatggccAGCCAAGAACTGGAGCAGA 344 tattggcgcgccatggccGGTCAACAGCTGAATCAGA 345 tattggcgcgccatggccACCCAGCTGCTGGAGCAGA 346 tattggcgcgccatggccCAGCAGCAGGTGAAACAA 347 tattggcgcgccatggccATACTGAACGTGGAACAA 348 tattggcgcgccatggccGACCAGCAAGTTAAGCAA 349 tattggcgcgccatggccGGACAACAGGTAATGCAA 350 tattggcgcgccatggccAAGGACCAAGTGTTTCAG Cα 200 nM 351 TAGGCAGACAGACTTGTCACT Vβ 10 nM 352 gccatggcgcgccaatagctagccGGTGAAGAAGTCGCCCAGA of each 353 gccatggcgcgccaatagctagccGATGCCATGGTCATCCAGA 354 gccatggcgcgccaatagctagccGAAGCCCAAGTGACCCAGA 355 gccatggcgcgccaatagctagccCATGCCAAAGTCACACAGA 356 gccatggcgcgccaatagctagccGACACAGCCGTTTCCCAGA 357 gccatggcgcgccaatagctagccGAAACGGGAGTTACGCAGA 358 gccatggcgcgccaatagctagccGAACCTGAAGTCACCCAGA 359 gccatggcgcgccaatagctagccGAAGCTGACATCTACCAGA 360 gccatggcgcgccaatagctagccATATCTGGAGTCTCCCACA 361 gccatggcgcgccaatagctagccGATGCTGGAATCACCCAGA 362 gccatggcgcgccaatagctagccAATGCTGGTGTCACTCAGA 363 gccatggcgcgccaatagctagccAGTGCTGTCGTCTCTCAA 364 gccatggcgcgccaatagctagccGACGCTGGAGTCACACAA 365 gccatggcgcgccaatagctagccGATGGTGGAATCACTCAGT 366 gccatggcgcgccaatagctagccACTGCTGGGATCACCCAG 367 gccatggcgcgccaatagctagccGAAGCTGGAGTTGCCCAGT 368 gccatggcgcgccaatagctagccGAAGCTGGAGTGGTTCAGT 369 gccatggcgcgccaatagctagccGAAGCTGGAGTTACTCAGT 370 gccatggcgcgccaatagctagccGATGCTGTAGTTACACAA 371 gccatggcgcgccaatagctagccGATGCTGGAGTTATCCAGT 372 gccatggcgcgccaatagctagccGGTGCTGGAGTCTCCCAG 373 gccatggcgcgccaatagctagccGATGCTGATGTTACCCAGA 374 gccatggcgcgccaatagctagccTCTCAGACTATTCATCAAT Cβ 200 nM 375 CAGGCACACCAGTGTGGCCTT
[0610]c. Semi-Nested PCR
[0611]Semi-nested PCR is likewise proposed to be performed as described in Example 11 e-2, and some optimization may be expected.
[0612]The primers used are shown in table 27.
TABLE-US-00048 TABLE 27 SEQ Primer ID primer sequence in name Conc. NO 5' to 3' direction Cα 200 nM 376 caccttagagctcTTAGAGTCTCTCAGCTGGTA CAC Cβ 200 nM 377 gacattatgCAtGATCTCTGCTTCTGATGGCTC Capitalized sequences correspond to the gene-specific region.
[0613]In order to verify that the multiplex overlap-extension RT-PCR is successful a proportion of the samples from the semi-nested PCR reactions are analyzed by subjecting 10 μl from each semi-nested PCR reaction to 1% agarose gel electrophoresis using ethidium bromide for detection. The expected size of the overlap-extension fragment is approximately 850 bp (the exact size depends upon the lengths of the variable regions).
[0614]Approximately ten microliters from all the performed reactions originating from the same donor, are consolidated in a single tube. An aliquot of the pooled PCR products is subsequently purified using QIAquick PCR purification kit according to the manufactures procedure (Qiagen cat. No. 28106, Hilden, Germany. The purified pool of overlap PCR products can be digested with appropriate restriction enzymes and subsequently purified and inserted into a suitable vector.
[0615]In the present experiment constant region primers (SEQ ID NO 376 and 377) used in the semi-nested PCR are designed for sub-cloning of semi-nested PCR product into a suitable vector. The design of the Cβ primer relies on changing a SER to Met at pos. 21 in the β chain constant region peptide, whereby a NsiI site can be introduced into the nucleic acid sequence:
TABLE-US-00049 NsiI tcc cac -> atg cat SER HIS MET HIS
[0616]This transition should be relative safe since SER 21 is exposed on the β chain constant region in a loop structure at the membrane proximal end of the domain. This relative conservative change is therefore not likely to disturb the overall domain structure.
[0617]The design of the Cα primer relies on changing nucleotides corresponding to pos. 15-17 in the α chain constant region peptide, whereby a Sad site can be introduced into the nucleic acid sequence:
TABLE-US-00050 SacI aaa tcc agt -> aag agc tct LYS SER SER LYS SER SER
[0618]Suitable vectors will therefore contain the remaining parts of the constant regions of the TcR α and β chains and will contain appropriate modification in terms of restriction sites. These changes can be performed using standard PCR and sub-cloning techniques.
[0619]e. Additional Considerations
[0620]The large amount of variable region primers may potentially interact in an inhibitory manner during the multiplex overlap-extension PCR amplification. To avoid this, variable region primers may be divided into sub-sets and used separately in proper combinations.
Other Embodiments
[0621]All publications and patent applications cited in this specification are herein incorporated by reference as if each individual publication or patent application were specifically and individually indicated to be incorporated by reference. Although the foregoing invention has been described in some detail by way of illustration and example for purposes of clarity of understanding, it will be readily apparent to those of ordinary skill in the art in light of the teachings of this invention that certain changes and modifications may be made thereto without departing from the spirit or scope of the appended claims.
Sequence CWU
1
377145DNAArtificialPrimer sequence 1agcctatact attgattagg cgcgcccagr
tgcagctggt gcart 45245DNAArtificialPrimer sequence
2agcctatact attgattagg cgcgccsagg tccagctggt rcagt
45345DNAArtificialPrimer sequence 3agcctatact attgattagg cgcgcccagr
tcaccttgaa ggagt 45444DNAArtificialPrimer sequence
4agcctatact attgattagg cgcgccsagg tgcagctggt ggag
44545DNAArtificialPrimer sequence 5agcctatact attgattagg cgcgcccagg
tgcagctaca gcagt 45645DNAArtificialPrimer sequence
6agcctatact attgattagg cgcgcccags tgcagctgca ggagt
45745DNAArtificialPrimer sequence 7agcctatact attgattagg cgcgccgarg
tgcagctggt gcagt 45846DNAArtificialPrimer sequence
8agcctatact attgattagg cgcgcccagg tacagctgca gcagtc
46920DNAArtificialPrimer sequence 9gacsgatggg cccttggtgg
201047DNAArtificialPrimer sequence
10cgcctaatca atagtatagg ctagccgaca tccagwtgac ccagtct
471147DNAArtificialPrimer sequence 11cgcctaatca atagtatagg ctagccgatg
ttgtgatgac tcagtct 471247DNAArtificialPrimer sequence
12cgcctaatca atagtatagg ctagccgaaa ttgtgwtgac rcagtct
471347DNAArtificialPrimer sequence 13cgcctaatca atagtatagg ctagccgata
ttgtgatgac ccacact 471445DNAArtificialPrimer sequence
14cgcctaatca atagtatagg ctagccgaaa cgacactcac gcagt
451547DNAArtificialPrimer sequence 15cgcctaatca atagtatagg ctagccgaaa
ttgtgctgac tcagtct 471638DNAArtificialPrimer sequence
16atatatatgc ggccgcttat taacactctc ccctgttg
381726DNAArtificialPrimer sequence 17atattctcga gacggtgacc agggtg
261825DNAArtificialPrimer sequence
18atattctcga gacggtgacc attgt
251927DNAArtificialPrimer sequence 19atattctcga gacggtgacc agggttc
272025DNAArtificialPrimer sequence
20atattctcga gacggtgacc gtggt
252151DNAArtificialPrimer sequence 21accgcctcca ccggcggccg cttattaaca
ctctcccctg ttgaagctct t 512243DNAArtificialprimer sequence
22gctagcatta ttattaccat ggcccagrtg cagctggtgc art
432343DNAArtificialPrimer sequence 23gctagcatta ttattaccat ggccsaggtc
cagctggtrc agt 432443DNAArtificialPrimer sequence
24gctagcatta ttattaccat ggcccagrtc accttgaagg agt
432542DNAArtificialPrimer sequence 25gctagcatta ttattaccat ggccsaggtg
cagctggtgg ag 422636DNAArtificialPrimer sequence
26tattggcgcg ccatggccsa ggtgcagctg gtggag
362743DNAArtificialPrimer sequence 27gctagcatta ttattaccat ggcccaggtg
cagctacagc agt 432843DNAArtificialPrimer sequence
28gctagcatta ttattaccat ggcccagstg cagctgcagg agt
432943DNAArtificialPrimer sequence 29gctagcatta ttattaccat ggccgargtg
cagctggtgc agt 433044DNAArtificialPrimer sequence
30gctagcatta ttattaccat ggcccaggta cagctgcagc agtc
443126DNAArtificialPrimer sequence 31atattctcga gacggtgacc agggtg
263227DNAArtificialPrimer sequence
32atattctcga gacggtgacc attgtcc
273327DNAArtificialPrimer sequence 33atattctcga gacggtgacc agggttc
273427DNAArtificialPrimer sequence
34atattctcga gacggtgacc gtggtcc
273544DNAArtificialPrimer sequence 35ccatggtaat aataatgcta gccgacatcc
agwtgaccca gtct 443644DNAArtificialPrimer sequence
36ccatggtaat aataatgcta gccgatgttg tgatgactca gtct
443744DNAArtificialPrimer sequence 37ccatggtaat aataatgcta gccgaaattg
tgwtgacrca gtct 443844DNAArtificialPrimer sequence
38ccatggtaat aataatgcta gccgatattg tgatgaccca cact
443942DNAArtificialPrimer sequence 39ccatggtaat aataatgcta gccgaaacga
cactcacgca gt 424044DNAArtificialPrimer sequence
40ccatggtaat aataatgcta gccgaaattg tgctgactca gtct
444137DNAArtificialPrimer sequence 41tattggcgcg ccatggccca grtgcagctg
gtgcart 374237DNAArtificialPrimer sequence
42tattggcgcg ccatggccsa ggtccagctg gtrcagt
374337DNAArtificialPrimer sequence 43tattggcgcg ccatggccca grtcaccttg
aaggagt 374436DNAArtificialPrimer sequence
44tattggcgcg ccatggccsa ggtgcagctg gtggag
364536DNAArtificialPrimer sequence 45tattggcgcg ccatggccca ggtgcagcta
cagcag 364637DNAArtificialPrimer sequence
46tattggcgcg ccatggccca gstgcagctg caggagt
374737DNAArtificialPrimer sequence 47tattggcgcg ccatggccga rgtgcagctg
gtgcagt 374838DNAArtificialPrimer sequence
48tattggcgcg ccatggccca ggtacagctg cagcagtc
384923DNAArtificialPrimer sequence 49aagtagtcct tgaccaggca gcc
235020DNAArtificialPrimer sequence
50tgagttccac gacaccgtca
205120DNAArtificialPrimer sequence 51agaggtgctc ttggaggagg
205221DNAArtificialPrimer sequence
52agttttgtca caagatttgg g
215323DNAArtificialPrimer sequence 53gttgcagatg taggtctggg tgc
235445DNAArtificialPrimer sequence
54gccatggcgc gccaatagct agccgacatc cagwtgaccc agtct
455545DNAArtificialPrimer sequence 55gccatggcgc gccaatagct agccgatgtt
gtgatgactc agtct 455645DNAArtificialPrimer sequence
56gccatggcgc gccaatagct agccgaaatt gtgwtgacrc agtct
455745DNAArtificialPrimer sequence 57gccatggcgc gccaatagct agccgatatt
gtgatgaccc acact 455843DNAArtificialPrimer sequence
58gccatggcgc gccaatagct agccgaaacg acactcacgc agt
435945DNAArtificialPrimer sequence 59gccatggcgc gccaatagct agccgaaatt
gtgctgactc agtct 456040DNAArtificialPrimer sequence
60atatatatgc ggccgcttat taacactctc ccctgttgaa
406130DNAArtificialPrimer sequence 61ggaggcgctc gagacggtga ccagggtgcc
306230DNAArtificialPrimer sequence
62ggaggcgctc gagacggtga ccattgtccc
306330DNAArtificialPrimer sequence 63ggaggcgctc gagacggtga ccagggttcc
306430DNAArtificialPrimer sequence
64ggaggcgctc gagacggtga ccgtggtccc
306547DNAArtificialPrimer sequence 65cgcctaatca atagtatagg ctagcccagt
ctgtgctgac tcagcca 476645DNAArtificialPrimer sequence
66cgcctaatca atagtatagg ctagcccagt ctgtgytgac gcagc
456745DNAArtificialPrimer sequence 67cgcctaatca atagtatagg ctagcccagt
ctgtcgtgac gcagc 456847DNAArtificialPrimer sequence
68cgcctaatca atagtatagg ctagcccart ctgccctgac tcagcct
476947DNAArtificialPrimer sequence 69cgcctaatca atagtatagg ctagcctcct
atgwgctgac tcagcca 477047DNAArtificialPrimer sequence
70cgcctaatca atagtatagg ctagcctctt ctgagctgac tcaggac
477147DNAArtificialPrimer sequence 71cgcctaatca atagtatagg ctagcccacg
ttatactgac tcaaccg 477245DNAArtificialPrimer sequence
72cgcctaatca atagtatagg ctagcccagg ctgtgctgac tcagc
457349DNAArtificialPrimer sequence 73cgcctaatca atagtatagg ctagccaatt
ttatgctgac tcagcccca 497446DNAArtificialPrimer sequence
74cgcctaatca atagtatagg ctagcccagr ctgtggtgac ycagga
467545DNAArtificialPrimer sequence 75cgcctaatca atagtatagg ctagcccwgc
ctgtgctgac tcagc 457631DNAArtificialPrimer sequence
76gccgcttatt atgaacattc tgtaggggcc a
317731DNAArtificialPrimer sequence 77gccgcttatt aagagcattc tgcaggggcc a
317845DNAArtificialPrimer sequence
78atatatatgc ggccgcttat tatgaacatt ctgtaggggc cactg
457945DNAArtificialPrimer sequence 79atatatatgc ggccgcttat taagagcatt
ctgcaggggc cactg 458036DNAArtificialPrimer sequence
80atatatatgc ggccgcttaa tgggctgcct cgggaa
368147DNAArtificialPrimer sequence 81gccatggcgc gccaatagct agccttagag
cagctcgtac tgacgaa 478241DNAArtificialPrimer sequence
82tattggcgcg ccatggccat gagtgagctt gaccagttac g
418347DNAArtificialPrimer sequence 83ggtacctaat aataatcgat cgcttagttc
cagatcttga ggaagct 478448DNAArtificialPrimer sequence
84cgatcgatta ttattaggta ccccatgcca gtaatcaata ttgaggac
488538DNAArtificialPrimer sequence 85ggaggcgctc gagttatgaa atcacacagc
ctcctttg 388634DNAArtificialPrimer sequence
86tattcccagc ggccgccgcc acaggtgccc actc
348734DNAArtificialPrimer sequence 87tattcccagc ggccgcccct tcmtgggtct
tgtc 348835DNAArtificialPrimer sequence
88tattcccagc ggccgcctta raaggtgtcc agtgt
358932DNAArtificialPrimer sequence 89tattcccagc ggccgccccc agatgggtcc tg
329034DNAArtificialPrimer sequence
90tattcccagc ggccgccctc caaggagtct gttc
349135DNAArtificialPrimer sequence 91tattcccagc ggccgcccca tggggtgtcc
tgtca 359233DNAArtificialPrimer sequence
92tattcccagc ggccgccgca acaggtgccc act
339341DNAArtificialPrimer sequence 93ggcggccgct gggaatagct agcgytccca
ggtgccagat g 419440DNAArtificialPrimer sequence
94ggcggccgct gggaatagct agcggtccct ggatccagtg
409542DNAArtificialPrimer sequence 95ggcggccgct gggaatagct agcgctccca
gataccaccg ga 429639DNAArtificialPrimer sequence
96ggcggccgct gggaatagct agcgatctct ggtgcctac
399742DNAArtificialPrimer sequence 97ggcggccgct gggaatagct agcgatctct
gataccaggg ca 429842DNAArtificialPrimer sequence
98ggcggccgct gggaatagct agcggttcca gcctccaggg gt
429937DNAArtificialPrimer sequence 99atatatatgg cgcgccttaa cactctcccc
tgttgaa 3710048DNAArtificialVector sequence
100atgaaatatc ttctaccaac agcggcagct ggattattgg cggccgcc
4810148DNAArtificialVector sequence 101atgaaatatt tgctaccaac agcggcagct
ggattattgt tactagcg 4810243DNAArtificialPrimer sequence
102tgttgttcta gatgaaggcg cgcccagrtg cagctggtgc art
4310343DNAArtificialPrimer sequence 103tgttgttcta gatgaaggcg cgccsaggtc
cagctggtrc agt 4310443DNAArtificialPrimer sequence
104tgttgttcta gatgaaggcg cgcccagrtc accttgaagg agt
4310542DNAArtificialPrimer sequence 105tgttgttcta gatgaaggcg cgccsaggtg
cagctggtgg ag 4210643DNAArtificialPrimer sequence
106tgttgttcta gatgaaggcg cgcccaggtg cagctacagc agt
4310743DNAArtificialPrimer sequence 107tgttgttcta gatgaaggcg cgcccagstg
cagctgcagg agt 4310843DNAArtificialPrimer sequence
108tgttgttcta gatgaaggcg cgccgargtg cagctggtgc agt
4310944DNAArtificialPrimer sequence 109tgttgttcta gatgaaggcg cgcccaggta
cagctgcagc agtc 4411045DNAArtificialPrimer sequence
110aacaacacta gtgataggct agccgacatc cagwtgaccc agtct
4511145DNAArtificialPrimer sequence 111aacaacacta gtgataggct agccgatgtt
gtgatgactc agtct 4511245DNAArtificialPrimer sequence
112aacaacacta gtgataggct agccgaaatt gtgwtgacrc agtct
4511345DNAArtificialPrimer sequence 113aacaacacta gtgataggct agccgatatt
gtgatgaccc acact 4511443DNAArtificialPrimer sequence
114aacaacacta gtgataggct agccgaaacg acactcacgc agt
4311545DNAArtificialPrimer sequence 115aacaacacta gtgataggct agccgaaatt
gtgctgactc agtct 4511636DNAArtificialPrimer sequence
116tattcccatg gcgcgcccag rtgcagctgg tgcart
3611736DNAArtificialPrimer sequence 117tattcccatg gcgcgccsag gtccagctgg
trcagt 3611836DNAArtificialPrimer sequence
118tattcccatg gcgcgcccag rtcaccttga aggagt
3611935DNAArtificialPrimer sequence 119tattcccatg gcgcgccsag gtgcagctgg
tggag 3512036DNAArtificialPrimer sequence
120tattcccatg gcgcgcccag gtgcagctac agcagt
3612136DNAArtificialPrimer sequence 121tattcccatg gcgcgcccag stgcagctgc
aggagt 3612236DNAArtificialPrimer sequence
122tattcccatg gcgcgccgar gtgcagctgg tgcagt
3612337DNAArtificialPrimer sequence 123tattcccatg gcgcgcccag gtacagctgc
agcagtc 3712445DNAArtificialPrimer sequence
124ggcgcgccat gggaatagct agccgacatc cagwtgaccc agtct
4512545DNAArtificialPrimer sequence 125ggcgcgccat gggaatagct agccgatgtt
gtgatgactc agtct 4512645DNAArtificialPrimer sequence
126ggcgcgccat gggaatagct agccgaaatt gtgwtgacrc agtct
4512745DNAArtificialPrimer sequence 127ggcgcgccat gggaatagct agccgatatt
gtgatgaccc acact 4512843DNAArtificialPrimer sequence
128ggcgcgccat gggaatagct agccgaaacg acactcacgc agt
4312945DNAArtificialPrimer sequence 129ggcgcgccat gggaatagct agccgaaatt
gtgctgactc agtct 4513037DNAArtificialPrimer sequence
130atatatatgc ggccgcttaa cactctcccc tgttgaa
3713120DNAArtificialVector Sequence 131aggaaacagg agatatacat
2013220DNAArtificialVector sequence
132tcgccaagga gacagtcata
20133369DNAHomo Sapiens 133gggcgcgccc aggtgcagct ggtgcaatct ggacctgaag
tgaagaagcc tggggcctca 60gtgagggtct cctgcaaggc ctctggttac tcctttaaga
actatggtat ccactgggtg 120cgacaggccc ctggacaggg gcttgagtgg atggggtgga
tcagcgctga caatggtgac 180acaaccactg cactgaacct ccggggcaga gtctccatga
ccacagacac atccacaaac 240acagtctaca tggaggtgaa gagcctaaga tctgacgaca
cggccatata tttctgtgcg 300cgagattttt atagtgggag ttaccggtcc tttgactgct
ggggccaggg caccctggtc 360accgtctcg
369134378DNAHomo Sapiens 134gggcgcgccc aggtgcagct
acagcagtct gggggaggcc tggtcaagcc tggggggtcc 60ctgagactct cctgtgcagc
ctctggattc accttcagta gctatagcat gaactgggtc 120cgccaggctc cagggaaggg
gctggagtgg gtctcatcca ttagtagtag tagtagttac 180atatactacg cagactcagt
gaagggccga ttcaccatct ccagagacaa cgccaagaac 240tcactgtatc tgcaaatgaa
cagcctgaga gccgaggaca cggctgtgta ttactgtgcg 300agacgccctg gttgggctgc
tactcgggcg gcgggtgctt ttgatatctg gggccaaggg 360acaatggtca ccgtctcg
378135357DNAHomo Sapiens
135gggcgcgccc aggtgcagct ggtgcaatct ggacctgaac tgaagaagcc tggggcctca
60gtgaggatct cctgcaaggc ctctgagaaa tgtgatatcc actgggtgcg acaggcccct
120ggacaggggc ttgagtggat gggatggatc agcgctgacg atggtgggac aaccactgcg
180ctgaacctcc ggggcagagt ctccatgacc acagacagag ccacaaacac agtatacatg
240gaactgaaga gcctaagatc tgacgacacg gccatttatt tctgtgcgcg agatttttat
300agtgggactt accggtcctt tgactactgg ggccagggaa ccctggtcac cgtctcg
357136396DNAHomo Sapiens 136gggcgcgcca aggtgcagct ggtggagtct gggggaggcg
tggtccagcc tgggacgtcc 60ctcagactct cctgtgtagt ctctggattc accttcagaa
cctatggtat gaactgggtc 120cgccaggctc caggcaaggg gctggagtgg ctggcatttc
tatcatctga tgggagcgat 180gaattctacg cggactccgt gaagggccga ttcaccgtct
ccagggacaa ttccaagagc 240actctctttc tgaaaatgaa cagtctgaga cctgatgaca
cggctgtcta ttactgtgcg 300agggatcgtg gggcccaaat tacacttttt ggagcccctc
ttataaggcc ttcgtccttt 360gactcctggg gccagggcac cctggtcacc gtctcg
396137362DNAHomo Sapiens 137gggcgcgccc aggtgcagct
gcaggagtct ggacctgagg tgaagaagcc tggggcctca 60gtgaaggtgt cctgcaaggc
ttctggctac acgtttaaca gttatggaat cgcctgggtg 120cgacaggtcc ctggacaagg
gcttgagtgg atgggatgga tcagccctta cagtggtcac 180acaaactatg cagagaaggt
ccagggcaga gtcaccatga ccacagacac cgccacgagc 240acagcctgca tggagctgac
gagcctgaga tctgacgaca cggccgttta tttctgtgcg 300agagactaca gtagtccgta
ccactttgac tactggggcc agggcacctg gtcaccgtct 360cg
362138366DNAHomo Sapiens
138gggcgcgccc agatcacctt gaaggagtct ggtcctacgc tggtgaaacc cacacagacc
60ctcacgctga cctgcacctt gtctgggttc tcactctaca ctactggagt gggtgtgggc
120tggatccgtc agcccccagg aaaggccctg gagtggctgg cacgcattta ttgggatgat
180gatgagcgct acaacccgtc tctgaagagc aggctcacca tcaccaagga cgcctccaaa
240aaccaggtgg tccttaaaat gaccaacatg gaccctgtgg acacagccac atattactgt
300gcccggacca tgggcgtcgt tcttccattt gactactggg gccagggaac cctggtcacc
360gtctcg
366139351DNAHomo Sapiens 139gggcgcgcca aggtgcagct ggtggagtct gggggaggcc
tggtcaagcc gggggggtcc 60ctgagactct cctgtgtagt ttctgggttc cccctcaata
gatacaccat gaactgggtc 120cgccaggctc cagggaaggg gctggagtgg ctctcgtcca
ttagtagtac tagttcttac 180atatactacg cagactcagt gaagggccga ttcaccatct
ccagagacaa cgccaagaat 240tctctgtttc tgcagatgaa cagcctgaga gccgacgaca
cggctctcta tttctgtgcg 300agtggcaata ctcatgacta ctggggccag ggaaccctgg
tcaccgtctc g 351140378DNAHomo Sapiens 140gggcgcgccc
aggtgcagct ggtgcagtct ggggctgagg tgaagaagct tgggacgtca 60gtgagggtct
cctgcaagac tcctggaggc tatgttttca gctgggtgcg acaggcccct 120ggacaagggc
ctgagtggat gggagggatc atcaccaact ttgggacaac aaactacgca 180cggaagttcc
agggcagaat cacggttacc gcggacaaat ccacgaacac agtgtacatg 240gatttgagca
acctggcatc tgaggacacg gccgtgtatt actgtgcgag agccccccga 300ggcacgtcga
ctatagcagc tcgttttaat cgatatttct ttgactcctg gggccagggc 360accctggtca
ccgtctcg
378141396DNAHomo Sapiens 141gggcgcgccc aggtgcagct tcaggagtct gggggaggcg
tggtccagcc tgggacgtcc 60ctcagactct cctgtgtagt ctctggattc accttcagaa
cctatggtat gaactgggtc 120cgccaggctc caggcaaggt gctggagtgg ctggcatttg
tatcatctga tgggagcgat 180gaattctacg cggactccgt gaagggccga ttcaccgtct
ccagggacaa ttccaagagc 240actctctttc tgaaaatgaa cagtctgaga gctgatgaca
cggctgtcta ttactgtgcg 300agagatcgtg gggcccaaat tacacttttt ggagcccctc
ttataaggcc ttcgtccttt 360gactcctggg gccagggaac cctggtcacc gtctcg
396142378DNAHomo Sapiens 142gggcgcgccg aggtgcagct
ggtggagtct ggggctgagg tgaagaagcc tgggacctca 60gtgagggtct cctgcaagac
ttctggaggc tatgttttca gctgggtgcg acaggcccct 120ggacaagggc ctgagtggat
gggagggatc atcaccagct ttgggacaac aagctacgca 180cagaagttcc agggcagagt
cacgattacc gcggacaaag ccacgaacac agtgtacatg 240gatttgagcg acctgacatc
tgaggacacg gccatatatt actgtgcgaa agccccccga 300ggcacgtcga ctatagcagc
tcgttttaat cgctatttct ttgactcctg gggccagggc 360accctggtca ccgtctcg
378143366DNAHomo Sapiens
143gggcgcgccc aggtgcagct gcaggagtct ggtcctacgc tggtgaaacc cacacagacc
60ctcacgctga cctgctcgtt ctctggtttc tcactcggca ctactggagt caatgtgggc
120tggatccgtc agcccccggg aaaggccctg gagtggcttg cactcatttc ttgggatggt
180ggtaagcact acagcccatc tctgaactcc aggatcaccc tcactaagga cgcctccaga
240gagcaggtgg tggtccctac aatgaccaac atggaccctg cggacacagg cagatattat
300tgtgcacgta tagtggggac tcacggcttt gactactggg gccagggaac cctggtcacc
360gtctcg
366144393DNAHomo Sapiens 144gggcgcgcca aggtgcagct ggtggagtct ggagctgagg
tgaagaagcc tggggccaca 60gtcagggtct cctgtaaggc ttctggttac aggtttaacg
actattgtat cagctgggtg 120cgacaggccc ctggacaagg gcttgagtgg atggggtgga
tcaacggtaa caatgctgac 180acattctatg caccgaagct ccagggcaga gtcaccatga
gcacagacac atccacgagc 240acagcctaca tggagctgag gaacttgaga tcggacgaca
cggccgttta tttctgtgcg 300cgagatcgag gacgtattac tctttttggc gaagttattt
taagggcggg atggttcgac 360tcctggggcc agggcaccct ggtcaccgtc tcg
393145372DNAHomo Sapiens 145gggcgcgccc aggtgcagct
ggtgcagtct gggggaggcc tggtcaagcc gggggggtcc 60ctgagactct cctgtgcagc
ctctggattc tcctttagta ataataacat gaattgggtc 120cgccagactc caggaaaggg
actggagtgg gtcgcatcta ttagttttgg aagtcattac 180atatcctacg cagactcagt
gaagggccga ttcaccatct ccagagacaa cgccaggaat 240gcagtttatc tgcagatgaa
cagcctgaga gtcgaggaca cggctgtcta ttactgcacg 300agatgcaggg gcggaactcg
tacctattat tacatggacg tctggggcaa aggcaccctg 360gtcaccgtct cg
372146372DNAHomo Sapiens
146gggcgcgcca aggtgcagct ggtggagtct gggggaggcc tggtcaagcc aggggggtcc
60ctgagactct cctgtgcagc ctctggatcc tcctttagta ataataacat gaattgggtc
120cgccagactc caggaaaggg actggagtgg gtcgcatcca ttagttttgg aagtcattac
180atatcctacg cagactcagt gaagggccga ttcaccatct ccagagacaa cgccaggaat
240gcagtttatc tgcagatgaa cagcctgaga gtcgaggaca cggctgtcta ttactgcacg
300agatgcaggg gcggaactcg tacctattat tacatggacg tctggggcaa aggcaccctg
360gtcaccgtct cg
372147390DNAHomo Sapiens 147gggcgcgccc agatgcagct ggtgcaatct ggggctgagg
tgaagaagcc tgggtcctcg 60gtgaaggtct cctgccagtc ttctggaggc ccccccaaaa
gttatactct cagctgggtg 120cggcaggccc ctggacaagg ccctgagtgg atgggcggaa
tcattctaat ctttggccca 180ccaaactacg cccagaagtt ccaggacaga ctcacgatca
ccgcggacaa gtccaccaac 240acagtctaca tggagttaag tagcctgaga tctgatgaca
cggccatgta ctactgtgtg 300acagcccccg acgacactgg cactatatta gctcgtcaca
accgttacta ctttgactcc 360tggggccagg gcaccctggt caccgtctcg
390148390DNAHomo Sapiens 148gggcgcgcca aggtgcagct
ggtgcagtct ggagcagagg tgaagaagcc cggggagtct 60ctgaaaatct cctgtcaggc
ttctggatac ggctttaccg tctactggat cggctgggtg 120cgccagccgc ccgggaaagg
cctggagtgg ctgggtatca tctatcctgg tgactctgat 180accagataca atccgtcctt
ccaaggccag gtcaccatct cagccgacaa gtccgtcagc 240accacctacc tgcagtggag
cagcctgaag gcctcggaca ccgccattta ctactgtgcg 300agacatctgg actcatacga
tgttttcact ggttataatt tggggggcta catggacgtc 360tggggcaagg gaaccctggt
caccgtctcg 390149351DNAHomo Sapiens
149gggcgcgcca aagtgtagct ggtgcagtct gggggaggcc tggtcaagcc tggggggtcc
60ctgagactct cctgtgtagt ctctggattc cccctcaata gatacatcat gaactgggtc
120cgccagactc cagggaaggg gctggagtgg ctctcgtcca ttagtagtac cagttcttac
180atatactacg cagactcagc gaagggccga ttcaccatct ccagagacaa cgccaagaat
240tctctgtttc tgcagatgaa cagcctgaga gccgaggaca caggtctcta ttactgtgcg
300agtggcaata ctcatgacta ttggggccag ggcaccctgg tcaccgtctc g
351150378DNAHomo Sapiens 150gggcgcgccc agatgcagct ggtgcagtct ggggctgagg
tgaagaagcc tgggacctca 60gtgagggtct cctgcaagac ttctggaggc tatgttttca
gctgggtgcg acaggcccct 120ggacaagggc ctgagtggat gggagggatc atcaccaact
ttgggacaac aaactacgca 180cagaagttcc agggcagagt cacgattacc gcggacaaat
ccacgaacac agtgtatatg 240gatttgagca acctgacatc tgaggacacg gccgtgtatt
actgtgcgag agccccccga 300ggcacgtcga ctatagcagc tcgttttaat cggtatttct
ttgactcctg gggccagggc 360accctggtca ccgtctcg
378151372DNAHomo Sapiens 151gggcgcgcca aggtgcagct
ggtggagtct ggggctgagg tgaagaagcc tgggtcctcg 60gtgaaggtct cctgcaaggc
ctctggaggc agcttcagca cctatgctat cacctgggtg 120cgccaggccc ctggacaggg
gcttgaatgg atgggaggga tcatccctat ctttgcttca 180agagactacg cacagaagtt
tcagggcaga gtcacagtca ccgcggacga atccacgagg 240acagtgtaca tggagctgag
cagcctgaga tctgacgaca cggccgtgta ttactgtgca 300agagtgctgg gaggtacaag
gctctactac gctctgaacg tctggggcca agggacaatg 360gtcaccgtct cg
372152372DNAHomo Sapiens
152gggcgcgcca aggtgcagct ggtggagtct ggggctgagg tgaagaagcc tgggtcctcg
60gtgaaggtct cctgcaagac atctggaggc agtttcagca catactctat cacctgggtg
120cgccaggccc ctggacaggg gcttgagtgg atgggaggga tcaaccctat cttcgctaca
180agagactacg cacagaagtt ccagggcaga gtcacgatca ccgcggacga atccacgagg
240acagtctaca tggagttgag gaacttgaga tctgaggaca cggccgtgta ttattgtgca
300agagtgttcg gaggaacaag actctactac gccctgaacg tctggggcca aggcaccctg
360gtcaccgtct cg
372153378DNAHomo Sapiens 153gggcgcgccg aagtgcagct ggtgcagtct ggggctgagg
tgaagaagcc tgggacgtca 60gtgagggtct cctgcaagac tcctggaggc tatgttttca
gctgggtgcg acaggcccct 120ggacaagggc ctgagtggat gggagggatc atcaccaact
ttgggacaac aaactacgca 180cggaagttcc agggcagaat cacggttacc gcggacaaat
ccacgaacac agtgtacatg 240gatttgagca acctggcatc tgaggacacg gccgtgtatt
actgtgcgag agccccccga 300ggcacgtcga ctatagcagc tcgttttaat cgatatttct
ttgactcctg gggccagggc 360accctggtca ccgtctcg
378154372DNAHomo Sapiens 154gggcgcgcca aggtgcagct
ggtggagtct gggggaggcc tggtcaagcc gggggggtcc 60ctgagactct cctgtgcagc
ctctggattc tcctttagta ataataacat gaattgggtc 120cgccagaccc caggaaaggg
actggagtgg gtcgcatcca ttagttttgg aagtcattac 180atatcctacg cagactcagt
gaagggccga ttcaccatct ccagagacaa cgccaggaat 240gcagtttatc tgcagatgaa
cagcctgaga gtcgaggaca cggctgtcta ttactgcacg 300agatgcaggg gcggaactcg
tacctattat tacatggacg tctggggcaa agggacaatg 360gtcaccgtct cg
372155390DNAHomo Sapiens
155gggcgcgcca aggtgcagct ggtgcagtct ggagcagagg tgaagaagcc cggggagtct
60ctgaaaatct cctgtcaggc ttctggatac ggctttaccg tctactggat cggctgggtg
120cgccagccgc ccgggaaagg cctggagtgg ctgggtatca tctatcctgg tgactctgat
180accagataca atccgtcctt ccaaggccag gtcaccatct cagccgacaa gtccgtcagc
240accacctacc tgcagtggag cagcctgaag gcctcggaca ccgccattta ctactgtgcg
300agacatctgg actcatacga tgttttcact ggttataatt tggggggcta catggacgtc
360tggggcaagg gaaccctggt caccgtctcg
390156378DNAHomo Sapiens 156gggcgcgccc aggtgcagct ggtggagtct ggggctgaag
tgaagaagcc tgggtcctca 60gtgaaggtct cctgcaagtc ttctggaggc tatgttttca
gctgggtgcg acaggcccct 120gggcaaggac tagagtggat gggagggatc atctccaact
ttcgcacggc agagtacgca 180cggaagttcc agggtagagt caccatgacc gcggacacat
ccacgaacac aatctacatg 240gagctgacca gcctgacatc tgaagacacg gccgtatatt
tctgtgtgag cgccccccga 300gacacgtcga ctatagcagc tcgttttaat cgatacttct
ttgacacctg gggccagggc 360accctggtca ccgtctcg
378157378DNAHomo Sapiens 157gggcgcgccc aggtgcagct
acagcagtcg gggggaggcg tggtccagcc tgggaggtcc 60ctgagactct cctgtgctgc
ctctggattc gccttcagag actatgccat gcactgggtc 120cgccaggccc caggcaaggg
gctggagtgg atgggagtta tctcatttaa tggagatcag 180atattttacg cagactccat
gaagggccgc ttcaccatct ccagagagaa ctccaagaac 240acgctgcatc tgcgcatgaa
cagcctgaga cctgaggaca cggctgtcta ttactgtgcg 300agagcccgac ttcttttttg
tagcggtggt aggtgcgaca tggactcttg gggccaggga 360accctggtca ccgtctcg
378158378DNAHomo Sapiens
158gggcgcgcca aagtgcagct ggtgcagtct ggggctgagg tgaagaagcc tgggacctca
60gtgagggtct cctgcaagac ttctggaggc tatgttttca gttgggtgcg acaggcccct
120ggacacgggc ctgaatggat gggagggatc atcaccaact ttgggacagc aacctacgca
180cagaagttcc agggcagagt ctcgattacc gcggacacat ccacgaacac attttacatg
240gatttgaaca atctgaaatc tgacgacacg gccgtttatt actgtgcgag cgccccccga
300gacacgtcga ctatagcggc tcggtttaat cggtacttct ttgacttctg gggcccgggc
360accctggtca ccgtctcg
378159348DNAHomo Sapiens 159gggcgcgcct aggtgcagct ggtgcagtct gggggaggct
tggtaaaacc tggggggtcc 60cttagactct cctgtgcagc ctctggattc actttcagta
acgcctggat gagctgggtc 120cgccaggctc cagggaaggg gctggagtgg gttggccatg
ttaaaagcat gactgatggt 180gggacaacag actacgctgc acccgtgaaa ggcagattca
ccatctcaag agatgattca 240gaaaacacgc tgtatctgca aatgaacagc ctgaagaccg
acgacacagc cgtgtattac 300tgtaccactc atgactactg gggccagggc accctggtca
ccgtctcg 348160378DNAHomo Sapiens 160gggcgcgcca
aagtgcagct ggtgcagtct ggggctgagg tgaagaagcc tgggacgtca 60gtgagggtct
cctgcaagac tcctggaggc tatgttttca gctgggtgcg acaggcccct 120ggacaagggc
ctgagtggat gggagggatc atcaccaact ttgggacaac aaactacgca 180cggaagttcc
agggcagaat cacggttacc gcggacaaat ccacgaacac agtgtacatg 240gatttgagca
acctggcatc tgaggacacg gccgtgtatt actgtgcgag agccccccga 300ggcacgtcga
ctatagcagc tcgttttaat cgatatttct ttgactcctg gggccagggc 360accctggtca
ccgtctcg
378161393DNAHomo Sapiens 161gggcgcgccc aggtgcagct ggtggagtct ggggctgaga
tgaagaagcc tgggtcctcg 60gtgaaggtct cctgccaggc ctctggaggc accttcagca
actatggcat caattgggtg 120cgacacgccc ctggacaagg gcttgagtgg atgggaggaa
tcgtccctat ctatggtcca 180ccgaagtacg cacagaagtt ccagggcaga gtcacgatta
ctgcggacac gtccacgagc 240acagcctaca tggagctgag cagcctgagc tctgatgaca
cggccgtgta ttactgtgcg 300cgactttccc ggactgtgtt tggctcgggg acttatcgtc
cgggctacta ctacatggac 360gtctggggca tagggaccac ggtcaccgtc tcg
393162372DNAHomo Sapiens 162gggcgcgcct aggtgcagct
acagcagtct gggggaggcc tggtcaagcc gggggggtcc 60ctgagactct cctgtgcagc
ctctggattc tcctttagta ataataacat gaattgggtc 120cgccagactc caggaaaggg
actggagtgg gtcgcatcca ttagttttgg aagtcattac 180atatcctacg cagactcagt
gaagggccga ttcaccatct ccagagacaa cgccaggaat 240gcagtttatc tgcagatgaa
cagcctgaga gtcgaggaca cggctgtcta ttactgcacg 300agatgcaggg gcggaactcg
tacctattat tacatggacg tctggggcaa aggcaccctg 360gtcaccgtct cg
372163378DNAHomo Sapiens
163gggcgcgcca aggtgcagct ggtgcagtct ggggctgagg tgaagaagcc tgggacctca
60gtgagggtct cctgcaagac ttctggaggc tatgttttca gctgggtgcg acaggcccct
120ggacaagggc ctgagtggat gggagggatc atcaccaact ttgggacaac aaactacgca
180cagaagttcc agggcagagt cacgattacc gcggacaaat ccacgaacac agtgtacatg
240gatttgagca acctgacatc tgaggacacg gccgtgtatt actgtgcgag agccccccga
300ggcacgtcga ctatagcagc tcgttttaat cggtatttct ttgactcctg gggccagggc
360accctggtca ccgtctcg
378164372DNAHomo Sapiens 164gggcgcgccg aagtgcagct ggtgcagtct ggggctgagg
tgaagaagcc tgggtcctcg 60gtgaaggtct cctgcaagac tgaaggaggc accttcagca
cctatgttat cagttggatg 120cgacaggccc ctggacaagg gcttgagtgg atgggaggga
tcgtccccat ctttaacaca 180ccaaactacg ctcagaaatt ccagggcaga gtcacaatta
ccgcggacag atccacgagc 240acagcctaca tggagctgag gagcctggga tctgaggaca
cggccgtcta ttactgtgcg 300agggtagtgg gaggtacaag gtcctactac gctttgggct
tctggggcca ggggaccacg 360gtcaccgtct cg
372165366DNAHomo Sapiens 165gggcgcgccc aggtcacctt
gaaggagtct ggtcctacgc tggtgaaacc cacacagacc 60ctcacgctga cctgcacgtt
ctctggtttc tcactcggca ctactggagt caatgtgggc 120tggatccgtc agcccccagg
aaaggccctg gagtggcttg cactcatttc ttggggtggt 180ggtaagcact acagcccatc
tctgaactcc aggatcaccc tcactaagga cgcctccaga 240gagcaggtgg tggtccttac
aatggccaac atggaccctg tggacacagg cagatattat 300tgtgcacgta tagtggggac
tcacggcttt gactactggg gccagggcac cctggtcacc 360gtctcg
366166372DNAHomo Sapiens
166gggcgcgccg aggtccagct ggtgcagtct gggggaggtg tggtgcggcc tggggggtcc
60ctgagactct cgtgtgcagg ctctggattc acctttgatg aatacgctat gagctgggtc
120cgccaagctc cagggaaggg gctggagtgg gtcgctttta ttaattggaa tggtgatagc
180acatattatg cagactctgt gaagggccga ttcaccgtct ccagaaccaa cgccaagaac
240tccctgtatc tgcaaatgaa cagtctgaga gccgaggaca cggccttcta ttactgtgcg
300agagaccccc gcactaaact ggggatgtcc tattttgact attggggcca gggcaccctg
360gtcaccgtct cg
372167390DNAHomo Sapiens 167gggcgcgccg aagtgcagct ggtgcagtct ggagcagagg
tgaagaagcc cggggagtct 60ctgaaaatct cctgtcaggc ttctggatac ggctttaccg
tctactggat cggctgggtg 120cgccagctgc ccgggaaagg cctggagtgg ctgggtatca
tctatcctgg tgactctgat 180accagataca atccgtcctt ccaaggccag gtcaccatct
cagccgacaa gtccgtcagc 240accacctacc tgcagtggag cagcctgaag gcctcggaca
ccgccattta ctactgtgcg 300agacatctgg actcatacga tgttttcact ggttataatt
tggggggcta catggacgtc 360tggggcaagg ggacaatggt caccgtctcg
390168369DNAHomo Sapiens 168gggcgcgccg aggtgcagct
ggtgcagtct ggacctgaag tgaagaagcc tggggcctca 60gtgagggtct cctgcaaggc
ctctggttac tcccttaaga actatggtat ccactgggtg 120cgacaggccc ctggacaggg
gcttgagtgg atggggtgga tcagcgctga caatggtgac 180acaaccactg cactgaacct
ccggggcaga gtctccatga ccacagacac atccacaaac 240acagtctaca tggaggtgaa
gagcctaaga tctgacgaca cggccatata tttctgtgcg 300cgagattttt atagtgggag
ttaccggtcc tttgactact ggggccaggg caccctggtc 360accgtctcg
369169378DNAHomo Sapiens
169gggcgcgccg aagtgcagct ggtgcagtcg ggcccaggac tggtgaagcc ttcggagacc
60ctgtccctca catgcgctgt ctctggtgcc tccgtcagcg gtggtgatta ctactggagc
120tggatccggc agcccccagg gaaggcactg gagtggattg ggtatatcta ttacataggg
180agcaccaact acaatccctc tctcaagagt cgactctccc tatcagtaga cacggccaag
240agccagttct ccctgaagtt gagctctgtg accgctgcgg acacggccat ttatttctgt
300gcgagagcac gtcggacgta tagtggctac gactccgcct ttgactactg gggccaggga
360accctggtca ccgtctcg
378170372DNAHomo Sapiens 170gggcgcgccc aggtgcagct gcagcagtcg ggcccaggac
tggtgaagcc ctcggagacc 60ctgcccctca cctgcactgt ctctggtggc tccttcagta
cctactactg gagctggatc 120cggcagtccc cagagaaggg actggagtgg attggatata
tccaaaacag tgtgaacacc 180aactacaacc cctccctcaa gagtcgtgtc atcatttcag
tggacacgtc caacaaccag 240ttctccctga agctgaggtc tgtgaccgct gcggacacgg
ccgtatatta ctgtgcgaga 300gtaagcggct ggggcccaag gggaggcatc tactttgact
actggggcca gggaaccctg 360gtcaccgtct cg
372171378DNAHomo Sapiens 171gggcgcgcca aggtgcagct
ggtgcagtct ggggctgaag tgaagaagcc tgggtcctca 60gtgaaggtct cctgcaagtc
ttctggaggc tatgttttca gctgggtgcg acaggcccct 120gggcaaggac tagagtggat
gggagggatc atctccaact ttcacacggc agagtacgca 180cagaagttcc agggtagagt
caccatgacc gcggacacat ccacgaacac aatctacatg 240gagctgacca gcctgacatc
tgaagacacg gccgtatatt tctgcgtgag cgccccccga 300gacacgtcga ctatagcagc
tcgttttaat cgatacttct ttgacacctg gggccagggc 360accctggtca ccgtctcg
378172378DNAHomo Sapiens
172gggcgcgccg aggtgcagct ggtgcagtct ggggctgagg tgaagaagcc tgggacgtca
60gtgagggtct cccgcaagac tcctggaggc tatgttttca gctgggtgcg acaggcccca
120ggacaagggc ctgagtggat gggagggatc atcaccaact ttgggacaac aaactacgca
180cggaagttcc agggcagaat cacggttacc gcggacaaat ccacgaacac agtgtacatg
240gatttgagca acctggcatc tgaggacacg gccgtgtatt actgtgcgag agccccccga
300ggcacgtcga ctatagcagc tcgttttaat cgatatttct ttgactcctg gagccagggc
360accctggtca ccgtctcg
378173372DNAHomo Sapiens 173gggcgcgccg aggtgcagct ggtggagtct ggggctgagg
tgaagaagcc tgggtcctcg 60gtgaaggtct cctgcaaggc ctctggaggc agcttcagca
cctatatttt cacctgggtg 120cgccaggccc ctggacaggg gcttgagtgg atgggaggga
tcaatcctat ctttgctaca 180agagactacc caaagaagtt ccagggcaga gtcacgatca
ccgcggacga atccacgagg 240actgtctata tggagttgag cagcctcaca tctgaggaca
cggccgtcta ttactgtgca 300agagtgttgg gaggtacaag gctctactac gctctgaacg
tctggggcca agggaccacg 360gtcaccgtct cg
372174372DNAHomo Sapiens 174gggcgcgccg aagtgcagct
ggtgcagtct gggggaggtg tggtgcggcc tggggggtcc 60ctgagactct cgtgtgcagg
ctctggattc acctttgatg aatacgccat gagctgggtc 120cgccaagctc cagggaaggg
gctgaagtgg gtcgctttta ttaattggaa tggtgatagc 180acatattatg cagactctgt
gaagggccga ttcaccgtct ccagatccaa cgccaagaac 240tccctgtatc tgcaaatgaa
cagtctgaga gccgaggaca cggccttcta ttgctgtgcg 300agagaccccc gcactaaact
ggggatgtcc tattttgact attggggcca gggcaccctg 360gtcaccgtct cg
372175372DNAHomo Sapiens
175gggcgcgccg aggtgcagct ggtgcagtct ggggctgagg tgaagaagcc tgggtcctcg
60gtgaaggtct cctgcaaggc ctctggaggc agcttcagca cctatgctat cacctgggtg
120cgccaggccc ctggacaggg gcttgaatgg atgggaggga tcatccctat ctttgcttca
180agagactacg cacagaagtt tcagggcaga gtcacaatca ccgcggacga atccacgagg
240acagtgtaca tggagccgag gagcctgaga tctgaagaca cggccgtgta ttactgtgca
300agagtgctgg gaggtacaag gctctactac gctctgaacg tctggggcca aggcaccctg
360gtcaccgtct cg
372176378DNAHomo Sapiens 176gggcgcgccg aggtgcagct ggtgcagtcg ggcccaggac
tggtgaagcc ttcggagacc 60ctgtccctca catgcgctgt ctctggtgcc tccgtcagcg
gtggtgatta ctactggagc 120tggatccggc agcccccagg gaaggcactg gagtggattg
ggtatatcta ttacataggg 180agcaccaact acaatccctc tctcaagagt cgactctccc
tatcagtaga cacggccaag 240agccagttct ccctgaagtt gagctctgtg accgctgcgg
acacggccac ttatttctgt 300gcgagagcac gtcggacgta tagtggctac gactccgcct
ttgactactg gggccaggga 360accctggtca ccgtctcg
378177372DNAHomo Sapiens 177gggcgcgccg aggtgcagct
ggtggagtct ggggctgagg tgaagaagcc tgggtcctcg 60gtgaaggtct cctgcaaggc
ctctggaggc agcttcagca cctatgctat cacctgggtg 120cgccaggccc ctggacaggg
gcttgaatgg atgggaggga tcatccctat ctttgcttca 180agagactacg cacagaagtt
tcagggcaga gtcacaatca ccgcggacga atccacgagg 240acagtgtaca tggagctgag
cagcctgaga tctgacgaca cggccgtgta ttactgtgca 300agagtgctgg gaggtacaag
gctctactac gctctgaacg tctggggcca agggaccacg 360gtcaccgtct cg
372178369DNAHomo Sapiens
178gggcgcgccc aggtcacctt gaaggagtct ggtcctacag tggtgaaacc cacacagacc
60ctcacgctga cctgtagcct ctctgggttc gcactcggca ctaccggagt ggctgtgggc
120tggatccgtc agcccccagg aaaggccctg gaatggcttg gactcatcga ttggaatgat
180gataggcgct acagaccctc tctgaagacc agactcacca tcacccagga catgtccagg
240aaccaggtgg tccttagact gaccaacttg gacccactgg acacaggcac atatttttgt
300gcacgttcag tagtgccggc gactagggcc tttgacttct ggggccaggg caccctggtc
360accgtctcg
369179366DNAHomo Sapiens 179gggcgcgccc aggtcacctt gaaggagtct ggtcctacgc
tggtgaaacc cacacagacc 60ctcacgctga cctgcacctt gtctgggttc tcactctaca
ctactggagt gggtgtgggc 120tggatccgtc agcccccagg aaaggccctg gagtgactgg
cacgcattta ttgggatgat 180gatgagcgct acaacccgtc tctgaagagc aggctcacca
tcaccaagga cgcctccaaa 240aaccaggtgg tccttaaaat gaccaacatg gaccctgtgg
acacagccac atattactgt 300gcccggacca tgggcgtcgt tcttccattt gactactggg
gccagggaac cctggtcacc 360gtctcg
366180327DNAHomo Sapiens 180ctagccgaaa ttgtgctgac
tcagtctcca tcctccctgt ctgcatctct aggagacaga 60gtcaccatca cttgccgggc
aagtcagcac attagcaatt atttaaattg gtatcagcag 120aaacctggga aagcccctaa
actcctgatc tgtgctgcat ccagtttgca aagtggggtc 180ccatcaaggt tcactggcag
tggatctggg gcagattaca ccctcaccat cagcagtctg 240caacctgaag attttgcaac
ttactactgt caacagagtt acagtacctc gtggacgttc 300ggccaaggga ccacggtgga
aatcaaa 327181327DNAHomo Sapiens
181ctagccgaaa ttgtgttgac acagtctcca tcctccctgt ctgcatctgt aggagacaga
60gtcaccatca cttgccgggc gagtcagggc attagcaatt atttagcctg gtatcagcag
120aaaccaggga aagttcctaa gctcctgatc tatgctgcat ccactttgca atcaggggtc
180ccatctcggt tcagtggcag tggatctggg acagatttca ctctcaccat cagcagcctg
240cagcctgaag atgttgcaac ttattactgt caaaagtata acagtgcccc gtacactttt
300ggccagggga ccaagctgga gatcaaa
327182327DNAHomo Sapiens 182ctagccgatg ttgtgatgac tcagtctcca tcctccctgt
ctgcatctct aggagacaga 60gtcagcatca cttgccgggc aagtcaacat attagcaatt
atataaattg gtatcagcag 120aaacctggga aagcccctaa actcctgatc tatgctgctt
ccactttgca aagtggggtc 180ccatcaaggt tcactggcag tggatctggg gcagattaca
ctctcaccat caccagtctg 240caacctgaag attttgcaac ttactactgt caacagagtt
acggtacctc gtggacgttc 300ggccaaggga ccacggtgga aatcaaa
327183327DNAHomo Sapiens 183ctagccgatg ttgtgatgac
tcagtctcca tcctccctgt ctgcatctgt gggagacaga 60gtcaccatca cttgccaggc
gagtcaagac attaccaact atttaaattg gtatcagcag 120aaaccaggga aagcccctaa
gctcctgatc tacgatgcat ccaatttgca accaggggtc 180ccatcaaggt tcagtggaag
tggatctgtg acagatttta ctttcaccat cagcagcctg 240cggcctgaag atattgcaac
atattactgt caacagtatg atggtccagt gcttactttc 300ggcggaggga ccaaggtaga
gatcaaa 327184327DNAHomo Sapiens
184ctagccgatg ttgtgatgac tcagtctcca tcctccctgt ctgcatctgt aggagacaga
60gtcaccatct cttgccgggc aagtcagccc attagcacct atttaaattg gtatcagcag
120aaaccaggga aagcccctaa gatcctgatc tttggtgcat ctcgtttgca aagtggtgtc
180ccatcaaggt tcagcggcag tggatctggg acagatttca gtctcaccat caccagtctc
240caacctgaag attttgcaac ttacatctgt caacaaagca agagtccccc gtacaatttt
300ggccggggga ccaagctgga gatcaaa
327185330DNAHomo Sapiens 185ctagccgaaa ttgtgttgac acagtctcct tccaccctgt
ctgcatctgt aggcgacaga 60gtcaccatca cttgccgggc cagtcagagt attggtagct
ggttggcctg gtatcaacag 120aaaccaggga aagcccctaa actcctgatc tataaggcgt
ctactttaca aagtgagacc 180ccttcaaggt tccgcggcag tggatctggg accgaattca
ctctcaccat cagcagcctg 240cagcctgatg attttgcaac ttactactgc caacagttta
atagtttttc tccgtggacg 300ttcggccaag ggaccaaggt ggaattcaaa
330186327DNAHomo Sapiens 186ctagccgatg ttgtgatgac
tcagtctcca tcgtccctgt ctgcatctgt tggtgacaga 60gtcaccatca cttgccgggc
aagtcagagc attaacatta atttaaattg gtttcagcag 120aaacccggga aagcccctaa
cctactgatc tattctgcat ccactttgca aactggggtc 180ccctcaaggt tcagtggaag
tggatctggg acagagttca ctctcaccat cagcagtcta 240caacctgaag attttgcaac
ttactactgt caacagattt acagtcatgt taggacgttc 300ggccaaggga ccaaggtgga
gatcaaa 327187345DNAHomo Sapiens
187ctagccgatg ttgtgatgac tcagtctcca gactccctgg ctgtgtctct gggcgagagg
60gccaccatca actgcaagtc cagccagagt cttttgtaca gctccaacaa taagaattac
120ttagcttggt accagcagaa accaggacag cctcctaagt tgctcattta ctgggcatct
180acccgggaat ccggggtccc tgaccgattc agtggcagcg ggtctgggac agatttcact
240ctcaccatca gcagcctgca ggctgaagat gtggcagttt attactgtca gcaatattat
300agtactcctc cgatgttcgg ccaagggacc aaggtggaaa tcaaa
345188327DNAHomo Sapiens 188ctagccgaaa ttgtgttgac acagtctcca tcctccctgt
ctgcatctgt gggagacaga 60atcaccatca cttgccaggc gagtcaagac attaccaact
atttaaattg gtatcagcag 120aaaccaggga aagcccctaa gctcctgatc tacgatgcat
ccaatttgca accaggggtc 180ccatcaaggt tcagtggaag tagatctggg acagatttta
ctttcaccat cagcagcctg 240cggcctgaag atattgcaac atattactgt caacagtatg
atggtccagt gcttactttc 300ggcggaggga ccaaggtaga gatcaaa
327189345DNAHomo Sapiens 189ctagccgaaa ttgtgttgac
acagtctcca gactccctgg ctgtgtctct gggcgagagg 60gccaccatca actgcaagtc
cagccagagt cttttataca gctccaacaa taagaactac 120ttagcttggt accagcagaa
accaggacag cctcctaagt tgctcattta ctgggcatct 180acccgggaat ccggggtccc
tgaccgattc agtggcagcg ggtctgggac agatttcacc 240ctcaccatca gcagcctgca
ggctgaagat gtggcagttt attactgtca gcaatattat 300agtactcctc cgatgttcgg
ccaagggacc aaggtggaaa tcaaa 345190330DNAHomo Sapiens
190ctagccgaaa ttgtgttgac acagtctcca tccttcctgt ctgcatctgt aggagacaga
60gtcaccatca cttgccgggc cagtcagggc attgggaatt ttttagcctg gtatcagcaa
120aaaccaggga aagcccctaa gctcctaatc tctggtgcat ccactttgca aagttgggtc
180ccatcaaggt tcagcggccg tggatctggg accgaattca ccctcacaat cagcagcctg
240cagcctgaag attttgcaac ttattactgt caacagctta atcgttaccc tccatacact
300tttggccagg ggaccaagct ggagatcaaa
330191327DNAHomo Sapiens 191ctagccgaaa ttgtgatgac gcagtctcca tcctccctgt
ctgcatctgt gggagacaga 60gtcaccatca cttgccaggc gagtcaggac attagcaaat
atttaaattg gtatcagcag 120aaaccaggga gagcccctaa actcctgatc tacgaagcat
ccaatttgga gacaggggtc 180ccaccaaggt tcagtggaag tggatctggg acacatttta
ctttcaccat caccggcctg 240cagcctgaag atattgcaac atattactgt caacagtgtg
atagtctgcc tccggtcttt 300ggccagggga ccaagctgga agtcaaa
327192330DNAHomo Sapiens 192ctagccgaaa ttgtgctgac
tcagtctcca ggcaccctgg ctttgtctcc aggtgataga 60gccaccctct cctgcggggc
cagtcagagc gtgttcggcg acttcttagc ctggtaccaa 120cacaagcctg gccaggctcc
caggctcctc atctatggtg cttccaccag ggccactggc 180atcccagaca ggttcagtgg
cagtggagct gggacagact tcactctcac catcagcaga 240ctggagcctg aggattttgc
agtttatttc tgtcagtagt atggtgactc agtgttcacg 300ttcggccaag ggaccaagtt
gggaatcaaa 330193330DNAHomo Sapiens
193ctagccgaaa ttgtgatgac acagtctcca ggcaccctgg ctttgtctcc aggtgataga
60gccaccctct cctgcggggc cagtcagagc gtgttcggcg acttcttagc ctggtaccaa
120cacaagcctg gccaggctcc caggctcctc atctatggtg cttccaccag ggccactggc
180atcccagaca ggttcagtgg cagtggagct gggacagact tcactctcac catcagcaga
240ctggagcctg aggattttgc agtttatttc tgtcagcagt atggtgactc agtgttcacg
300ttcggccaag ggaccaagtt ggaaatcaaa
330194330DNAHomo Sapiens 194ctagccgaaa ttgtgttgac acagtctcca ggcaccctgt
ctttgtctcc agggcaaaga 60gccaccctct cctgcagggc cagtcagagt gttaacagag
actacctagc ctggtaccag 120cagaaacctg gctaggctcc caggctcctc atctatggtg
catccagcag ggccactggc 180atcccagaca ggttcagtgg cagtgggtct ggaacagact
tcactctcac catcagcaga 240ctggagcgag acgattttgc agtgtatttc tgtcaccagt
atggtagctc acctaacact 300tttggccagg ggaccaagct ggagatcaaa
330195330DNAHomo Sapiens 195ctagccgaaa cgacactcac
gcagtctcca ggcaccctgt ctttgtctcc aggggaaaga 60gccaccctct cctgcagggc
cagtcagagc gttagcacca actacttagc ctggtaccga 120cagaaacctg gccaggctcc
caggctcctc atccatggtg catccagccg ggccactggc 180atcccagaca ggttcagtgg
cagtgggtct gggacagact tcactctcac catcagcaga 240ctggagcctg aagattttgc
agtgtatttc tgtcagcagt atggtagctc acctcagacg 300ttcggccaag ggaccaggtt
ggaaatcaaa 330196327DNAHomo Sapiens
196ctagccgaaa ttgtgctgac tcagtctcca tcctccctgt ctgcatctgt tggtgacaga
60gtcaccacca cttgccggac aagtcagagc attttcatta atttaaattg gtttcagcag
120aaacccggga aagcccctaa actcctgatc tattctgcat ccactttgca aactggggtc
180ccctcaaggt tcagtggcag tggatctggg acagagttca ctctcaccat cagcagtcta
240caacctgaag attttgcaac ttactactgt caacagattt acagtcaggt taggacgttc
300ggccaaggga ccaaggtgga gatcaaa
327197330DNAHomo Sapiens 197ctagccgaca tccagatgac ccagtctcca ggcaccctgt
ctttgtctcc aggggaaaga 60gccaccctct cctgcagggc cagtcagagt gttaacagcg
acaacttagc ctggtaccag 120cagaaacctg gccaggctcc caggctcctc atgtctggtg
caaccagtag ggccactgac 180gtcccagaca ggttcagtgg cagtgggtct gggacagact
tcactctcac catcagcaga 240ctggagcctg aagattttgc agtgtattac tgtcaccagt
atggtagctc agataacact 300tttggccagg ggaccaagct ggagatcaaa
330198330DNAHomo Sapiens 198ctagccgatg ttgtgatgac
tcagtttcct tcctctctgt ctgcatctgt gggagacaga 60gtcaccatca cttgccgggc
gagtcagggc attagcaatt ttttagcctg gtatcagcag 120aaaccaggga aagttcctga
gctccttatc tatggtgcat ccactttgca atcaggggtc 180ccatctcggt tcagtggcag
tggatctggg gcagagttca cactcaccat caacagcctg 240cagcctgaag atgttgcgac
ttattactgt caaaagtatg acagtggcct gagattcact 300ttcggccctg ggaccaaagt
ggatatcaaa 330199330DNAHomo Sapiens
199ctagccgatg ttgtgatgac tcagtctcct tcctctctgt ctgcatctgt aggagacaga
60gtcaccatca cttgccgggc gagtcagggc attagcaatt ttttagcctg gcatcagcag
120aaaccagggc aagttcctaa actcctgatc tatggtgcat ccactttgca atcaggggtc
180ccatctcgct tcagtggcag tggatctggc acagatttca ctctcaccat caacggcctg
240cagcctgaag atgttgcaac ttattactgt caaaagtatg acagtggcct gatattcact
300ttcggccctg ggaccagagt ggagatcaaa
330200345DNAHomo Sapiens 200ctagccgatg ttgtgatgac tcagtctcca gactccctgg
ctgtgtctct gggcgagagg 60gccaccatca actgcaagtc cagccagagt cttttataca
gctccaacaa taagaactac 120ttagcttggt accagcagaa accaggacag cctcctaagt
tgctcattta ctgggcatct 180acccgggaat ccggggtccc tgaccgattc agtggcagcg
ggtctgggac agatttcact 240ctcaccatca gcagcctgca ggctgaagat gtggcagttt
attactgtca gcaatattat 300agtactcctc cgatgctcgg ccaagggacc aaggtggaaa
tcaaa 345201330DNAHomo Sapiens 201ctagccgatg
ttgtgatgac ttagtctcca ggcaccctgg ctttgtctcc aggtgataga 60gccaccctct
cctgcggggc cagtcagagc gtgttcggcg acttcttagc ctggtaccaa 120cacaagcctg
gccaggctcc caggctcctc atctatgttg cttccaccag ggccactggc 180atcccagaca
ggttcagtgg cagtggagct gggacagact tcactctcac catcagcaga 240ctggagcctg
aggattttgc agtttatttc tgtcagcagt atggtgactc agtgttcacg 300ttcggccaag
ggaccaagtt ggaaatcaga
330202330DNAHomo Sapiens 202ctagccgaaa cgacactcac gcagtctcca ggcaccctgt
ctttgtctcc aggggaaaga 60gccaccctct cctgcagggc cagtcagagc gttagcacca
actacttagc ctggtaccga 120cagaaacctg gccaggctcc caggctcctc atccatggtg
catccagccg ggccactggc 180atcccagaca ggttcagtgg cagtgggtct gggacagact
tcactctcac catcagcaga 240ctggagcctg aagattttgc agtgtatttc tgtcagcagt
atggtagctc acctcagacg 300ttcggccaag ggaccaggtt ggaaatcaaa
330203330DNAHomo Sapiens 203ctagccgaaa ttgtgctgac
tcagtctcca ggcaccctgt ctttgtctcc aggggaaaga 60gccaccctct cctgcagggc
cagtcggagt gtaaacagca acaacttagc ctggtatcag 120cagaaacctg accaggctcc
caggctcctc atgtatggtg catccagtag ggccactggc 180atcccagaca ggttcactgg
cagtgggtct gggacagact tcactctcac catcagcaga 240ctggagcctg aagattttgc
agtgtattac tgtcatcagt atggtgcctc agataacact 300tttggccagg ggaccaagct
ggagatcaag 330204327DNAHomo Sapiens
204ctagccgaaa ttgtgatgac acagtctcca tccttcctgt ctgcatctgt aggagacaga
60gtcaccatca cttgccgggc cagtcaggac attatcactt atttagcctg gtatcaacaa
120aaaccaggga aagcccctga ggtcctgatc tttggtgcgt ccactttgca aagtggggtc
180ccatcaagat tcagcggcag tggatctggg accgaattca ctctcactat ctccagcctg
240cagcctgaag attttgcaac ttattactgt caacagatta ttcgttaccc tcgcactttc
300ggccaaggga ccagggtgga aatcaaa
327205330DNAHomo Sapiens 205ctagccgatg ttgtgatgac tcagtctcca ggcaccctgt
ctttgtctcc aggggaaaga 60gccaccctct cctgccgggc cagtcagagt gttaacagca
acaacttagc ctggtaccag 120cagaaacctg gccaggctcc caggctcctc gtctctggtg
caaccaatag ggccactgac 180atcccagaca ggttcagtgg cagtgggtct gggacagact
tcactctcac catcagcaga 240ctggagcctg aagattttgc agtgtattac tgtcaccagt
atggtagctc agagaacact 300tttggccagg ggaccaagct ggagatcaga
330206327DNAHomo Sapiens 206ctagccgaaa ttgtgatgac
gcagtctcca cccttcctgt ctgcatctgt aggagacaga 60gtcaccatca cttgccgggc
cagtcagggc ttaagcactt atttagcctg gtatcaggta 120aaaccaggga aagcccctaa
gctcctgatc tatgctgcat ccactttgca aagtggggtc 180ccatcaaggt tcagcggcag
tggatctggg acagaattca ctctcacaat caacagcctg 240cagcctgaag attttgcaac
ttattactgt caacaacttg atacttaccc tctcactctc 300ggcggaggga ccaaggtgga
gatcaaa 327207345DNAHomo Sapiens
207ctagccgaaa ttgtgttgac acagtctcca gactccctgg ctgtgtctct gggcgagagg
60gccaccatca actgcaagtc cagccagagt cttttataca gctccaacaa taagaactac
120ttagcttggt accagcagaa accaggacag cctcctaagt tgctcattta ctgggcatct
180acccgggaat ccggggtccc tgaccgattc agtggcagcg ggtctgggac agatttcact
240ctcaccatca gcagcctgca ggctgaagat gtggcagttt attactgtca gcaatattat
300agtactcctc cgatgttcgg ccaagggacc aaggtggaaa tcaaa
345208345DNAHomo Sapiens 208ctagccgaca tccagatgac ccagtctcca gactccctgg
ctgtgtctct gggcgagagg 60gccaccatca actgcaagtc cagccagagt cttttataca
gctccaacaa taagaactac 120ttagcttggt accagcagaa accaggacag cctcctaagt
tgctcattta ctgggcatct 180acccgggaat ccggggtccc tgaccgattc agtggcagcg
ggtctgggac agatttcact 240ctcaccatca gcagcctgca ggctgaagat gtggcagttt
attactgtca gcaatattat 300agtactcctc cgatgttcgg ccaagggacc agggtggaaa
tcaaa 345209330DNAHomo Sapiens 209ctagccgaaa
ttgtgatgac acagtctcca ggcaccctgg ctttgtctcc aggtgataga 60gccaccctct
cctgcggggc cagtcagagc gtgttcggcg acttcttagc ctggtaccaa 120cacaagcctg
gccaggctcc caggcccctc atctatggtg cttccaccag ggccactggc 180atcccagaca
ggttcagtgg cagtggagct gggacagact tcactctcac catcagcaga 240ctggagcctg
aggattttgc agtttatttc tgtcagcagt atggtgactc agtgttcacg 300ttcggccaag
ggaccaagtt ggaaatcaaa
330210330DNAHomo Sapiens 210ctagccgatg ttgtgatgac tcagtctcca ggcaccctgt
ctttgtctcc aggggaaaga 60gccaccctct cctgcagggc cagtcagagt gttaacagcg
acaacttagc ctggtaccag 120cagaaacctg gccaggctcc caggctcctc atgtctggtg
caaccagtag ggccactgac 180atcccagaca ggttcagtgg cagtgggtct gggacagact
tcactctcac catcagcaga 240ctggagcctg aagattttgc agtgtattac tgtcaccagt
atggtagctc agataacact 300tttggccagg ggaccaagct ggagatcaaa
330211330DNAHomo Sapiens 211ctagccgaca tccagttgac
ccagtctcca tcctccctgt ctgcatctgt tggagacagc 60gtcaccatca cttgccgggc
gagtcagggc attagcaatt ttttagcctg gtatcagaag 120aaaccgggaa aagttcctag
gctcctgatc tatggtgcat ccactttgca atcaggggtc 180ccatctcggt tcagtggcag
tggatctggg accgatttca ctctcaccat cagcagcctg 240cagcctgaag atgttgcgac
ttattactgt caggagtata gcaatgccct gatattcact 300ttcggccctg ggaccaaagt
tcatatcaaa 330212330DNAHomo Sapiens
212ctagccgaaa ttgtgatgac acagtctcca tccttcctgt ctgcgtctgt aggagacaga
60gtcaccatca cttgccgggc cagtcagggc attgggaatt ttttagcctg gtatcagcaa
120aaaccaggga aagcccctaa gctcctgatc tctggtgcat ccactttgca aagttgggtc
180ccatcaaggt tcagcggccg tggatctggg accgaattca ctctcacaat cagcagcctg
240cagcctgaag attttgcaac ttattactgt caacagctta atcgttaccc tccatacact
300tttggccagg ggaccaagct ggagatcaaa
330213327DNAHomo Sapiens 213ctagccgaaa ttgtgatgac acagtctcca tcctccctgt
ctgcatctgt aggagacaga 60gtcaccatca cttgccgggc aagtcagacc attagcaacc
atttaaattg gtatcagcag 120aaaccaggga gagcccctaa gctcctgatc tatgttgggt
ccagtctgca aagtggggtc 180ccatcaaggt tcagtggcag tggatctggg acagatttca
ctctcaccat cagtggtctg 240caacctgaag attttgcaac ttactactgt caacagagtt
acagtccctc gtacactttt 300ggccagggga ccaaggtgga gatcaaa
327214330DNAHomo Sapiens 214ctagccgaca tccagttgac
ccagtctcca ggcaccctgt ctttgtctcc aggggaaaga 60gccaccctct cctgcagggc
cagtcagagc gttagcacca actacttagc ctggtaccga 120cagaaacctg gccaggctcc
caggctcctc atccatggtg catccagccg ggccactggc 180atcccagaca ggttcagtgg
cagtgggtct gggacagact tcactctcac catcagcaga 240ctggagcctg aagattttgc
agtgtatttc tgtcagcagt atggtagctc acctcagacg 300ttcggccaag ggaccaggtt
ggaaatcaaa 330215327DNAHomo Sapiens
215ctagccgatg ttgtgatgac tcagtctcca tcctccctgt ctgcatctct aggagacaga
60gtcaccatca cttgccgggc aagtcagcat attagcaatt atttaaattg gtatcagcag
120aaacctggga aagcccctaa actcctgatc tatgctgcat ccagtttgca aagtggggtc
180ccatcaaggt tcactggcag tggatctggg gcagattaca ctctcaccat cagcagtctg
240caacctgaag attttgcaac ttactactgt caacagagtt acagcacctc gtggacgttc
300ggccaaggga ccacggtggg aatcaaa
327216327DNAHomo Sapiens 216ctagccgatg ttgtgatgac tcagtctcca tcctccctgt
ctgcatctgt aggagacaga 60gtcagcatca cttgccgggc aagtcggagc gttaagacct
atttaaattg gtatcagcag 120aaaccaggga aagcccctaa gctcctggtc tatggttcgt
ccagtttgga aagtggggtc 180ccatcaagat tcagtggcag tggatctggg acagatttca
ctctcaccat cagcagtctg 240caacctgagg attttgcaac ttactactgt caacagagtt
tcggtacccc ctacactttt 300ggccagggga ccaagctgga caccaaa
327217327DNAHomo Sapiens 217ctagccgaca tccagatgac
ccagtctcca tcctccctgt ctgcatctgt aagtgacaga 60gtcaccatta cttgccgggc
aagtcagagc attaacaagt tcttaaactg gtatcagcag 120aaaccaggga aagcccctca
gctcctcatc tatgctgcaa ccaatttgca gagtggggtc 180ccatcaaggt tcagtggcag
tggatctggg acagacttca ctctcaccat cagcagtctg 240caaactgaag attttgccac
ttactactgt caacagagtt acgatatgcc tcggacgttc 300ggccaaggga ccaaggtgga
gatcaaa 327218330DNAHomo Sapiens
218ctagccgaaa ttgtgctgac tcagtctcca ggcaccctgt ctttgtctcc aggggaaaga
60gccaccctct cctgcagggc cagtcggagt gtatacagca acaacttagc ctggtatcag
120cagaaacctg accaggctcc caggctcctc atgtatggtg catccagtag ggccactggc
180atcccagaca ggttcactgg cagtgggtct gggacagact tcactctcac catcagcaga
240ctggagcctg aagattttgc agtgtattac tgtcatcagt atggtgcctc agataacact
300tttggccagg ggaccaagct ggagatcaag
330219345DNAHomo Sapiens 219ctagccgaca tccagatgac ccagtctcca gactccctgg
ctgtgtctct gggcgagaag 60gccaccatca actgcaagtc cagccagagt cttttataca
gctccaacaa taagaactac 120ttagcttggt accagcagaa accaggacag cctcctaagt
tgctcattta ctgggcatct 180acccgggaat ccggggtccc tgaccgattc agtggcagcg
ggtctgggac agatttcact 240ctcaccatca gcagcctgca ggctgaagat gtggcagttt
attactgtca gcaatattat 300agtactcctc cgatgttcgg ccaagggacc aaggtggaaa
tcaaa 345220330DNAHomo Sapiens 220ctagccgaaa
ttgtgatgac gcagtctcct tcctctctgt ctgcatctgt aggagacaga 60gtcaccatct
cttgccgggc gagtcagggc attagcaatt ttttagcctg gtttcagcag 120aaaccaggtc
aagttcctaa gctcctgatc tatgttgcat ccactttgca atcaggggtc 180ccatctcggt
tcagtggcag tggatctggg acagatttca gtctcaccat caacggcctg 240cagcctgagg
atgttgcaac ttattactgt caaaggtatg acagtggcct gatattcact 300ttcggccctg
ggaccaaagt ggagatcaaa
330221327DNAHomo Sapiens 221ctagccgaca tccagatgac ccagtctcca tcctccctgt
ctgcatctgt aggagacaga 60gtcaccatca cttgccgggc aagtcagacc attagcaacc
atttaaattg gtatcagcag 120aaaccaggga gagcccctaa gctcctgatc tatgttgggt
ccagtctgcg aagtggggtc 180ccatcaaggt tcagtggcag tggatctggg acagatttca
ctctcaccat cagtggtctg 240caacctgaag attttgcaac ttactactgt caacagagtt
acagtccctc gtacactttt 300ggccagggga ccaaggtgga gatcaaa
327222330DNAHomo Sapiens 222ctagccgaaa ttgtgttgac
acagtctcct tcctctctgt ctgcatctgt gggagacaga 60gtcaccatca cttgccgggc
gagtcagggc attagcaatt ttttagcctg gtatcagcag 120aaaccaggga aagttcctga
gctccttatc tatggcgcat ccactttgca atcgggggtc 180ccatctcggt tcagtggcag
tggatctggg acagatttca ctctcaccat caacagcctg 240cagcctgaag atgttgcaac
ttattactgt caaaagtatg acagtggcct gagattcact 300ttcggccctg gggccaaagt
ggatatcaaa 330223327DNAHomo Sapiens
223ctagccgatg ttgtgatgac tcagtctcca tcctccctgt ctgcatctgt aggagacaga
60gtcagcatca cttgccgggc aagtcggagc gttaagacct atttaaattg gtatcagcag
120aaaccaggga aagcccctaa gctcctggtc tatggttcgt ccagtttgga aagtggggtc
180ccatcaagat tcagtggcag tggatctggg acagatttca ctctcaccat cagcagtctg
240caacctgagg attttgcaac ttactgctgt caacagagtt tcggtacccc ctacactttt
300ggccagggga ccaagctgga catcaaa
327224330DNAHomo Sapiens 224ctagccgaca tccagttgac ccagtctcct tcctctctgt
ctgcatctgt gggagacaga 60gtcaccatca cttgccgggc gagtcagggc attagcaatt
ttttagcctg gtatcagcag 120aaaccaggga aagttcctga gctccttatc tatggtgcat
ccaccttgca atcaggggtc 180ccatctcggt tcagtggcag tggatctggg acagagttca
ctctcaccat caacagcctg 240cagcctgaag atgttgcgac ttattactgt caaaagtatg
atagtggcct gagattcact 300ttcggccctg ggaccaaagt ggatatcaaa
330225327DNAHomo Sapiens 225ctagccgaca tccagatgac
ccagtcttct tccaccctgt ctgcatctgt aggagacaga 60gtcaccatca cttgccgggc
cagtcagaat attaatatct ggttggcctg gtatcagcag 120aaagcaggga aagcccccaa
actcctgatc tataaggcgt ctactttaga aaggggggtc 180ccctcaaggt tcagcggcag
tggatctggg acagagttca ctctcaccat caccggcctg 240cgccctgacg atttcggaag
ttattattgc caacactatg atggtaattc actaacgttc 300ggccaaggga ccagggtgga
tatccaa 327226330DNAHomo Sapiens
226ctagccgaca tccagttgac ccagtctcct tccaccctgt ctgcatctgt aggcgacaga
60gtcaccatca cttgccgggc cagtcagagt attggtagct ggttggcctg gtatcaacag
120aaaccaggga aagcccctaa actcctgatc tataaggcgt ctactttaca aagtgagacc
180ccttcaaggt tccgcggcag tggatctggg accgaattca ctctcaccat cagcagcctg
240cagcctgatg attttgcaac ttactactgc caacagttta atagtttttc tccgtggacg
300ttcggccaag ggaccaaggc ggaattcaaa
330227123PRThomo sapiens 227Gly Arg Ala Gln Val Gln Leu Val Gln Ser Gly
Pro Glu Val Lys Lys1 5 10
15Pro Gly Ala Ser Val Arg Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe
20 25 30Lys Asn Tyr Gly Ile His Trp
Val Arg Gln Ala Pro Gly Gln Gly Leu 35 40
45Glu Trp Met Gly Trp Ile Ser Ala Asp Asn Gly Asp Thr Thr Thr
Ala 50 55 60Leu Asn Leu Arg Gly Arg
Val Ser Met Thr Thr Asp Thr Ser Thr Asn65 70
75 80Thr Val Tyr Met Glu Val Lys Ser Leu Arg Ser
Asp Asp Thr Ala Ile 85 90
95Tyr Phe Cys Ala Arg Asp Phe Tyr Ser Gly Ser Tyr Arg Ser Phe Asp
100 105 110Cys Trp Gly Gln Gly Thr
Leu Val Thr Val Ser 115 120228126PRThomo sapiens
228Gly Arg Ala Gln Val Gln Leu Gln Gln Ser Gly Gly Gly Leu Val Lys1
5 10 15Pro Gly Gly Ser Leu Arg
Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe 20 25
30Ser Ser Tyr Ser Met Asn Trp Val Arg Gln Ala Pro Gly
Lys Gly Leu 35 40 45Glu Trp Val
Ser Ser Ile Ser Ser Ser Ser Ser Tyr Ile Tyr Tyr Ala 50
55 60Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp
Asn Ala Lys Asn65 70 75
80Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
85 90 95Tyr Tyr Cys Ala Arg Arg
Pro Gly Trp Ala Ala Thr Arg Ala Ala Gly 100
105 110Ala Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr
Val Ser 115 120 125229119PRThomo
sapiens 229Gly Arg Ala Gln Val Gln Leu Val Gln Ser Gly Pro Glu Leu Lys
Lys1 5 10 15Pro Gly Ala
Ser Val Arg Ile Ser Cys Lys Ala Ser Glu Lys Cys Asp 20
25 30Ile His Trp Val Arg Gln Ala Pro Gly Gln
Gly Leu Glu Trp Met Gly 35 40
45Trp Ile Ser Ala Asp Asp Gly Gly Thr Thr Thr Ala Leu Asn Leu Arg 50
55 60Gly Arg Val Ser Met Thr Thr Asp Arg
Ala Thr Asn Thr Val Tyr Met65 70 75
80Glu Leu Lys Ser Leu Arg Ser Asp Asp Thr Ala Ile Tyr Phe
Cys Ala 85 90 95Arg Asp
Phe Tyr Ser Gly Thr Tyr Arg Ser Phe Asp Tyr Trp Gly Gln 100
105 110Gly Thr Leu Val Thr Val Ser
115230132PRThomo sapiens 230Gly Arg Ala Lys Val Gln Leu Val Glu Ser Gly
Gly Gly Val Val Gln1 5 10
15Pro Gly Thr Ser Leu Arg Leu Ser Cys Val Val Ser Gly Phe Thr Phe
20 25 30Arg Thr Tyr Gly Met Asn Trp
Val Arg Gln Ala Pro Gly Lys Gly Leu 35 40
45Glu Trp Leu Ala Phe Leu Ser Ser Asp Gly Ser Asp Glu Phe Tyr
Ala 50 55 60Asp Ser Val Lys Gly Arg
Phe Thr Val Ser Arg Asp Asn Ser Lys Ser65 70
75 80Thr Leu Phe Leu Lys Met Asn Ser Leu Arg Pro
Asp Asp Thr Ala Val 85 90
95Tyr Tyr Cys Ala Arg Asp Arg Gly Ala Gln Ile Thr Leu Phe Gly Ala
100 105 110Pro Leu Ile Arg Pro Ser
Ser Phe Asp Ser Trp Gly Gln Gly Thr Leu 115 120
125Val Thr Val Ser 130231121PRThomo sapiens 231Gly Arg
Ala Gln Val Gln Leu Gln Glu Ser Gly Pro Glu Val Lys Lys1 5
10 15Pro Gly Ala Ser Val Lys Val Ser
Cys Lys Ala Ser Gly Tyr Thr Phe 20 25
30Asn Ser Tyr Gly Ile Ala Trp Val Arg Gln Val Pro Gly Gln Gly
Leu 35 40 45Glu Trp Met Gly Trp
Ile Ser Pro Tyr Ser Gly His Thr Asn Tyr Ala 50 55
60Glu Lys Val Gln Gly Arg Val Thr Met Thr Thr Asp Thr Ala
Thr Ser65 70 75 80Thr
Ala Cys Met Glu Leu Thr Ser Leu Arg Ser Asp Asp Thr Ala Val
85 90 95Tyr Phe Cys Ala Arg Asp Tyr
Ser Ser Pro Tyr His Phe Asp Tyr Trp 100 105
110Gly Gln Gly Thr Trp Ser Pro Ser Arg 115
120232122PRThomo sapiens 232Gly Arg Ala Gln Ile Thr Leu Lys Glu Ser
Gly Pro Thr Leu Val Lys1 5 10
15Pro Thr Gln Thr Leu Thr Leu Thr Cys Thr Leu Ser Gly Phe Ser Leu
20 25 30Tyr Thr Thr Gly Val Gly
Val Gly Trp Ile Arg Gln Pro Pro Gly Lys 35 40
45Ala Leu Glu Trp Leu Ala Arg Ile Tyr Trp Asp Asp Asp Glu
Arg Tyr 50 55 60Asn Pro Ser Leu Lys
Ser Arg Leu Thr Ile Thr Lys Asp Ala Ser Lys65 70
75 80Asn Gln Val Val Leu Lys Met Thr Asn Met
Asp Pro Val Asp Thr Ala 85 90
95Thr Tyr Tyr Cys Ala Arg Thr Met Gly Val Val Leu Pro Phe Asp Tyr
100 105 110Trp Gly Gln Gly Thr
Leu Val Thr Val Ser 115 120233117PRThomo sapiens
233Gly Arg Ala Lys Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys1
5 10 15Pro Gly Gly Ser Leu Arg
Leu Ser Cys Val Val Ser Gly Phe Pro Leu 20 25
30Asn Arg Tyr Thr Met Asn Trp Val Arg Gln Ala Pro Gly
Lys Gly Leu 35 40 45Glu Trp Leu
Ser Ser Ile Ser Ser Thr Ser Ser Tyr Ile Tyr Tyr Ala 50
55 60Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp
Asn Ala Lys Asn65 70 75
80Ser Leu Phe Leu Gln Met Asn Ser Leu Arg Ala Asp Asp Thr Ala Leu
85 90 95Tyr Phe Cys Ala Ser Gly
Asn Thr His Asp Tyr Trp Gly Gln Gly Thr 100
105 110Leu Val Thr Val Ser 115234126PRThomo
sapiens 234Gly Arg Ala Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys
Lys1 5 10 15Leu Gly Thr
Ser Val Arg Val Ser Cys Lys Thr Pro Gly Gly Tyr Val 20
25 30Phe Ser Trp Val Arg Gln Ala Pro Gly Gln
Gly Pro Glu Trp Met Gly 35 40
45Gly Ile Ile Thr Asn Phe Gly Thr Thr Asn Tyr Ala Arg Lys Phe Gln 50
55 60Gly Arg Ile Thr Val Thr Ala Asp Lys
Ser Thr Asn Thr Val Tyr Met65 70 75
80Asp Leu Ser Asn Leu Ala Ser Glu Asp Thr Ala Val Tyr Tyr
Cys Ala 85 90 95Arg Ala
Pro Arg Gly Thr Ser Thr Ile Ala Ala Arg Phe Asn Arg Tyr 100
105 110Phe Phe Asp Ser Trp Gly Gln Gly Thr
Leu Val Thr Val Ser 115 120
125235132PRThomo sapiens 235Gly Arg Ala Gln Val Gln Leu Gln Glu Ser Gly
Gly Gly Val Val Gln1 5 10
15Pro Gly Thr Ser Leu Arg Leu Ser Cys Val Val Ser Gly Phe Thr Phe
20 25 30Arg Thr Tyr Gly Met Asn Trp
Val Arg Gln Ala Pro Gly Lys Val Leu 35 40
45Glu Trp Leu Ala Phe Val Ser Ser Asp Gly Ser Asp Glu Phe Tyr
Ala 50 55 60Asp Ser Val Lys Gly Arg
Phe Thr Val Ser Arg Asp Asn Ser Lys Ser65 70
75 80Thr Leu Phe Leu Lys Met Asn Ser Leu Arg Ala
Asp Asp Thr Ala Val 85 90
95Tyr Tyr Cys Ala Arg Asp Arg Gly Ala Gln Ile Thr Leu Phe Gly Ala
100 105 110Pro Leu Ile Arg Pro Ser
Ser Phe Asp Ser Trp Gly Gln Gly Thr Leu 115 120
125Val Thr Val Ser 130236126PRThomo sapiens 236Gly Arg
Ala Glu Val Gln Leu Val Glu Ser Gly Ala Glu Val Lys Lys1 5
10 15Pro Gly Thr Ser Val Arg Val Ser
Cys Lys Thr Ser Gly Gly Tyr Val 20 25
30Phe Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Pro Glu Trp Met
Gly 35 40 45Gly Ile Ile Thr Ser
Phe Gly Thr Thr Ser Tyr Ala Gln Lys Phe Gln 50 55
60Gly Arg Val Thr Ile Thr Ala Asp Lys Ala Thr Asn Thr Val
Tyr Met65 70 75 80Asp
Leu Ser Asp Leu Thr Ser Glu Asp Thr Ala Ile Tyr Tyr Cys Ala
85 90 95Lys Ala Pro Arg Gly Thr Ser
Thr Ile Ala Ala Arg Phe Asn Arg Tyr 100 105
110Phe Phe Asp Ser Trp Gly Gln Gly Thr Leu Val Thr Val Ser
115 120 125237122PRThomo sapiens
237Gly Arg Ala Gln Val Gln Leu Gln Glu Ser Gly Pro Thr Leu Val Lys1
5 10 15Pro Thr Gln Thr Leu Thr
Leu Thr Cys Ser Phe Ser Gly Phe Ser Leu 20 25
30Gly Thr Thr Gly Val Asn Val Gly Trp Ile Arg Gln Pro
Pro Gly Lys 35 40 45Ala Leu Glu
Trp Leu Ala Leu Ile Ser Trp Asp Gly Gly Lys His Tyr 50
55 60Ser Pro Ser Leu Asn Ser Arg Ile Thr Leu Thr Lys
Asp Ala Ser Arg65 70 75
80Glu Gln Val Val Val Pro Thr Met Thr Asn Met Asp Pro Ala Asp Thr
85 90 95Gly Arg Tyr Tyr Cys Ala
Arg Ile Val Gly Thr His Gly Phe Asp Tyr 100
105 110Trp Gly Gln Gly Thr Leu Val Thr Val Ser 115
120238131PRThomo sapiens 238Gly Arg Ala Lys Val Gln Leu
Val Glu Ser Gly Ala Glu Val Lys Lys1 5 10
15Pro Gly Ala Thr Val Arg Val Ser Cys Lys Ala Ser Gly
Tyr Arg Phe 20 25 30Asn Asp
Tyr Cys Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu 35
40 45Glu Trp Met Gly Trp Ile Asn Gly Asn Asn
Ala Asp Thr Phe Tyr Ala 50 55 60Pro
Lys Leu Gln Gly Arg Val Thr Met Ser Thr Asp Thr Ser Thr Ser65
70 75 80Thr Ala Tyr Met Glu Leu
Arg Asn Leu Arg Ser Asp Asp Thr Ala Val 85
90 95Tyr Phe Cys Ala Arg Asp Arg Gly Arg Ile Thr Leu
Phe Gly Glu Val 100 105 110Ile
Leu Arg Ala Gly Trp Phe Asp Ser Trp Gly Gln Gly Thr Leu Val 115
120 125Thr Val Ser 130239124PRThomo
sapiens 239Gly Arg Ala Gln Val Gln Leu Val Gln Ser Gly Gly Gly Leu Val
Lys1 5 10 15Pro Gly Gly
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Phe 20
25 30Ser Asn Asn Asn Met Asn Trp Val Arg Gln
Thr Pro Gly Lys Gly Leu 35 40
45Glu Trp Val Ala Ser Ile Ser Phe Gly Ser His Tyr Ile Ser Tyr Ala 50
55 60Asp Ser Val Lys Gly Arg Phe Thr Ile
Ser Arg Asp Asn Ala Arg Asn65 70 75
80Ala Val Tyr Leu Gln Met Asn Ser Leu Arg Val Glu Asp Thr
Ala Val 85 90 95Tyr Tyr
Cys Thr Arg Cys Arg Gly Gly Thr Arg Thr Tyr Tyr Tyr Met 100
105 110Asp Val Trp Gly Lys Gly Thr Leu Val
Thr Val Ser 115 120240124PRThomo sapiens 240Gly
Arg Ala Lys Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys1
5 10 15Pro Gly Gly Ser Leu Arg Leu
Ser Cys Ala Ala Ser Gly Ser Ser Phe 20 25
30Ser Asn Asn Asn Met Asn Trp Val Arg Gln Thr Pro Gly Lys
Gly Leu 35 40 45Glu Trp Val Ala
Ser Ile Ser Phe Gly Ser His Tyr Ile Ser Tyr Ala 50 55
60Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn
Ala Arg Asn65 70 75
80Ala Val Tyr Leu Gln Met Asn Ser Leu Arg Val Glu Asp Thr Ala Val
85 90 95Tyr Tyr Cys Thr Arg Cys
Arg Gly Gly Thr Arg Thr Tyr Tyr Tyr Met 100
105 110Asp Val Trp Gly Lys Gly Thr Leu Val Thr Val Ser
115 120241130PRThomo sapiens 241Gly Arg Ala Gln Met
Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys1 5
10 15Pro Gly Ser Ser Val Lys Val Ser Cys Gln Ser
Ser Gly Gly Pro Pro 20 25
30Lys Ser Tyr Thr Leu Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Pro
35 40 45Glu Trp Met Gly Gly Ile Ile Leu
Ile Phe Gly Pro Pro Asn Tyr Ala 50 55
60Gln Lys Phe Gln Asp Arg Leu Thr Ile Thr Ala Asp Lys Ser Thr Asn65
70 75 80Thr Val Tyr Met Glu
Leu Ser Ser Leu Arg Ser Asp Asp Thr Ala Met 85
90 95Tyr Tyr Cys Val Thr Ala Pro Asp Asp Thr Gly
Thr Ile Leu Ala Arg 100 105
110His Asn Arg Tyr Tyr Phe Asp Ser Trp Gly Gln Gly Thr Leu Val Thr
115 120 125Val Ser 130242130PRThomo
sapiens 242Gly Arg Ala Lys Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys
Lys1 5 10 15Pro Gly Glu
Ser Leu Lys Ile Ser Cys Gln Ala Ser Gly Tyr Gly Phe 20
25 30Thr Val Tyr Trp Ile Gly Trp Val Arg Gln
Pro Pro Gly Lys Gly Leu 35 40
45Glu Trp Leu Gly Ile Ile Tyr Pro Gly Asp Ser Asp Thr Arg Tyr Asn 50
55 60Pro Ser Phe Gln Gly Gln Val Thr Ile
Ser Ala Asp Lys Ser Val Ser65 70 75
80Thr Thr Tyr Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr
Ala Ile 85 90 95Tyr Tyr
Cys Ala Arg His Leu Asp Ser Tyr Asp Val Phe Thr Gly Tyr 100
105 110Asn Leu Gly Gly Tyr Met Asp Val Trp
Gly Lys Gly Thr Leu Val Thr 115 120
125Val Ser 130243117PRThomo sapiens 243Gly Arg Ala Lys Val Gln Leu
Val Gln Ser Gly Gly Gly Leu Val Lys1 5 10
15Pro Gly Gly Ser Leu Arg Leu Ser Cys Val Val Ser Gly
Phe Pro Leu 20 25 30Asn Arg
Tyr Ile Met Asn Trp Val Arg Gln Thr Pro Gly Lys Gly Leu 35
40 45Glu Trp Leu Ser Ser Ile Ser Ser Thr Ser
Ser Tyr Ile Tyr Tyr Ala 50 55 60Asp
Ser Ala Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn65
70 75 80Ser Leu Phe Leu Gln Met
Asn Ser Leu Arg Ala Glu Asp Thr Gly Leu 85
90 95Tyr Tyr Cys Ala Ser Gly Asn Thr His Asp Tyr Trp
Gly Gln Gly Thr 100 105 110Leu
Val Thr Val Ser 115244126PRThomo sapiens 244Gly Arg Ala Gln Met
Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys1 5
10 15Pro Gly Thr Ser Val Arg Val Ser Cys Lys Thr
Ser Gly Gly Tyr Val 20 25
30Phe Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Pro Glu Trp Met Gly
35 40 45Gly Ile Ile Thr Asn Phe Gly Thr
Thr Asn Tyr Ala Gln Lys Phe Gln 50 55
60Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Asn Thr Val Tyr Met65
70 75 80Asp Leu Ser Asn Leu
Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala 85
90 95Arg Ala Pro Arg Gly Thr Ser Thr Ile Ala Ala
Arg Phe Asn Arg Tyr 100 105
110Phe Phe Asp Ser Trp Gly Gln Gly Thr Leu Val Thr Val Ser 115
120 125245124PRThomo sapiens 245Gly Arg Ala
Lys Val Gln Leu Val Glu Ser Gly Ala Glu Val Lys Lys1 5
10 15Pro Gly Ser Ser Val Lys Val Ser Cys
Lys Ala Ser Gly Gly Ser Phe 20 25
30Ser Thr Tyr Ala Ile Thr Trp Val Arg Gln Ala Pro Gly Gln Gly Leu
35 40 45Glu Trp Met Gly Gly Ile Ile
Pro Ile Phe Ala Ser Arg Asp Tyr Ala 50 55
60Gln Lys Phe Gln Gly Arg Val Thr Val Thr Ala Asp Glu Ser Thr Arg65
70 75 80Thr Val Tyr Met
Glu Leu Ser Ser Leu Arg Ser Asp Asp Thr Ala Val 85
90 95Tyr Tyr Cys Ala Arg Val Leu Gly Gly Thr
Arg Leu Tyr Tyr Ala Leu 100 105
110Asn Val Trp Gly Gln Gly Thr Met Val Thr Val Ser 115
120246124PRThomo sapiens 246Gly Arg Ala Lys Val Gln Leu Val Glu Ser
Gly Ala Glu Val Lys Lys1 5 10
15Pro Gly Ser Ser Val Lys Val Ser Cys Lys Thr Ser Gly Gly Ser Phe
20 25 30Ser Thr Tyr Ser Ile Thr
Trp Val Arg Gln Ala Pro Gly Gln Gly Leu 35 40
45Glu Trp Met Gly Gly Ile Asn Pro Ile Phe Ala Thr Arg Asp
Tyr Ala 50 55 60Gln Lys Phe Gln Gly
Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Arg65 70
75 80Thr Val Tyr Met Glu Leu Arg Asn Leu Arg
Ser Glu Asp Thr Ala Val 85 90
95Tyr Tyr Cys Ala Arg Val Phe Gly Gly Thr Arg Leu Tyr Tyr Ala Leu
100 105 110Asn Val Trp Gly Gln
Gly Thr Leu Val Thr Val Ser 115 120247126PRThomo
sapiens 247Gly Arg Ala Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys
Lys1 5 10 15Pro Gly Thr
Ser Val Arg Val Ser Cys Lys Thr Pro Gly Gly Tyr Val 20
25 30Phe Ser Trp Val Arg Gln Ala Pro Gly Gln
Gly Pro Glu Trp Met Gly 35 40
45Gly Ile Ile Thr Asn Phe Gly Thr Thr Asn Tyr Ala Arg Lys Phe Gln 50
55 60Gly Arg Ile Thr Val Thr Ala Asp Lys
Ser Thr Asn Thr Val Tyr Met65 70 75
80Asp Leu Ser Asn Leu Ala Ser Glu Asp Thr Ala Val Tyr Tyr
Cys Ala 85 90 95Arg Ala
Pro Arg Gly Thr Ser Thr Ile Ala Ala Arg Phe Asn Arg Tyr 100
105 110Phe Phe Asp Ser Trp Gly Gln Gly Thr
Leu Val Thr Val Ser 115 120
125248124PRThomo sapiens 248Gly Arg Ala Lys Val Gln Leu Val Glu Ser Gly
Gly Gly Leu Val Lys1 5 10
15Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Phe
20 25 30Ser Asn Asn Asn Met Asn Trp
Val Arg Gln Thr Pro Gly Lys Gly Leu 35 40
45Glu Trp Val Ala Ser Ile Ser Phe Gly Ser His Tyr Ile Ser Tyr
Ala 50 55 60Asp Ser Val Lys Gly Arg
Phe Thr Ile Ser Arg Asp Asn Ala Arg Asn65 70
75 80Ala Val Tyr Leu Gln Met Asn Ser Leu Arg Val
Glu Asp Thr Ala Val 85 90
95Tyr Tyr Cys Thr Arg Cys Arg Gly Gly Thr Arg Thr Tyr Tyr Tyr Met
100 105 110Asp Val Trp Gly Lys Gly
Thr Met Val Thr Val Ser 115 120249130PRThomo
sapiens 249Gly Arg Ala Lys Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys
Lys1 5 10 15Pro Gly Glu
Ser Leu Lys Ile Ser Cys Gln Ala Ser Gly Tyr Gly Phe 20
25 30Thr Val Tyr Trp Ile Gly Trp Val Arg Gln
Pro Pro Gly Lys Gly Leu 35 40
45Glu Trp Leu Gly Ile Ile Tyr Pro Gly Asp Ser Asp Thr Arg Tyr Asn 50
55 60Pro Ser Phe Gln Gly Gln Val Thr Ile
Ser Ala Asp Lys Ser Val Ser65 70 75
80Thr Thr Tyr Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr
Ala Ile 85 90 95Tyr Tyr
Cys Ala Arg His Leu Asp Ser Tyr Asp Val Phe Thr Gly Tyr 100
105 110Asn Leu Gly Gly Tyr Met Asp Val Trp
Gly Lys Gly Thr Leu Val Thr 115 120
125Val Ser 130250126PRThomo sapiens 250Gly Arg Ala Gln Val Gln Leu
Val Glu Ser Gly Ala Glu Val Lys Lys1 5 10
15Pro Gly Ser Ser Val Lys Val Ser Cys Lys Ser Ser Gly
Gly Tyr Val 20 25 30Phe Ser
Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met Gly 35
40 45Gly Ile Ile Ser Asn Phe Arg Thr Ala Glu
Tyr Ala Arg Lys Phe Gln 50 55 60Gly
Arg Val Thr Met Thr Ala Asp Thr Ser Thr Asn Thr Ile Tyr Met65
70 75 80Glu Leu Thr Ser Leu Thr
Ser Glu Asp Thr Ala Val Tyr Phe Cys Val 85
90 95Ser Ala Pro Arg Asp Thr Ser Thr Ile Ala Ala Arg
Phe Asn Arg Tyr 100 105 110Phe
Phe Asp Thr Trp Gly Gln Gly Thr Leu Val Thr Val Ser 115
120 125251126PRThomo sapiens 251Gly Arg Ala Gln Val
Gln Leu Gln Gln Ser Gly Gly Gly Val Val Gln1 5
10 15Pro Gly Arg Ser Leu Arg Leu Ser Cys Ala Ala
Ser Gly Phe Ala Phe 20 25
30Arg Asp Tyr Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
35 40 45Glu Trp Met Gly Val Ile Ser Phe
Asn Gly Asp Gln Ile Phe Tyr Ala 50 55
60Asp Ser Met Lys Gly Arg Phe Thr Ile Ser Arg Glu Asn Ser Lys Asn65
70 75 80Thr Leu His Leu Arg
Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Val 85
90 95Tyr Tyr Cys Ala Arg Ala Arg Leu Leu Phe Cys
Ser Gly Gly Arg Cys 100 105
110Asp Met Asp Ser Trp Gly Gln Gly Thr Leu Val Thr Val Ser 115
120 125252126PRThomo sapiens 252Gly Arg Ala
Lys Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys1 5
10 15Pro Gly Thr Ser Val Arg Val Ser Cys
Lys Thr Ser Gly Gly Tyr Val 20 25
30Phe Ser Trp Val Arg Gln Ala Pro Gly His Gly Pro Glu Trp Met Gly
35 40 45Gly Ile Ile Thr Asn Phe Gly
Thr Ala Thr Tyr Ala Gln Lys Phe Gln 50 55
60Gly Arg Val Ser Ile Thr Ala Asp Thr Ser Thr Asn Thr Phe Tyr Met65
70 75 80Asp Leu Asn Asn
Leu Lys Ser Asp Asp Thr Ala Val Tyr Tyr Cys Ala 85
90 95Ser Ala Pro Arg Asp Thr Ser Thr Ile Ala
Ala Arg Phe Asn Arg Tyr 100 105
110Phe Phe Asp Phe Trp Gly Pro Gly Thr Leu Val Thr Val Ser 115
120 125253116PRThomo sapiens 253Gly Arg Ala
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Leu Val Lys1 5
10 15Pro Gly Gly Ser Leu Arg Leu Ser Cys
Ala Ala Ser Gly Phe Thr Phe 20 25
30Ser Asn Ala Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
35 40 45Glu Trp Val Gly His Val Lys
Ser Met Thr Asp Gly Gly Thr Thr Asp 50 55
60Tyr Ala Ala Pro Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser65
70 75 80Glu Asn Thr Leu
Tyr Leu Gln Met Asn Ser Leu Lys Thr Asp Asp Thr 85
90 95Ala Val Tyr Tyr Cys Thr Thr His Asp Tyr
Trp Gly Gln Gly Thr Leu 100 105
110Val Thr Val Ser 115254126PRThomo sapiens 254Gly Arg Ala Lys
Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys1 5
10 15Pro Gly Thr Ser Val Arg Val Ser Cys Lys
Thr Pro Gly Gly Tyr Val 20 25
30Phe Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Pro Glu Trp Met Gly
35 40 45Gly Ile Ile Thr Asn Phe Gly Thr
Thr Asn Tyr Ala Arg Lys Phe Gln 50 55
60Gly Arg Ile Thr Val Thr Ala Asp Lys Ser Thr Asn Thr Val Tyr Met65
70 75 80Asp Leu Ser Asn Leu
Ala Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala 85
90 95Arg Ala Pro Arg Gly Thr Ser Thr Ile Ala Ala
Arg Phe Asn Arg Tyr 100 105
110Phe Phe Asp Ser Trp Gly Gln Gly Thr Leu Val Thr Val Ser 115
120 125255131PRThomo sapiens 255Gly Arg Ala
Gln Val Gln Leu Val Glu Ser Gly Ala Glu Met Lys Lys1 5
10 15Pro Gly Ser Ser Val Lys Val Ser Cys
Gln Ala Ser Gly Gly Thr Phe 20 25
30Ser Asn Tyr Gly Ile Asn Trp Val Arg His Ala Pro Gly Gln Gly Leu
35 40 45Glu Trp Met Gly Gly Ile Val
Pro Ile Tyr Gly Pro Pro Lys Tyr Ala 50 55
60Gln Lys Phe Gln Gly Arg Val Thr Ile Thr Ala Asp Thr Ser Thr Ser65
70 75 80Thr Ala Tyr Met
Glu Leu Ser Ser Leu Ser Ser Asp Asp Thr Ala Val 85
90 95Tyr Tyr Cys Ala Arg Leu Ser Arg Thr Val
Phe Gly Ser Gly Thr Tyr 100 105
110Arg Pro Gly Tyr Tyr Tyr Met Asp Val Trp Gly Ile Gly Thr Thr Val
115 120 125Thr Val Ser
130256124PRThomo sapiens 256Gly Arg Ala Gln Val Gln Leu Gln Gln Ser Gly
Gly Gly Leu Val Lys1 5 10
15Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Phe
20 25 30Ser Asn Asn Asn Met Asn Trp
Val Arg Gln Thr Pro Gly Lys Gly Leu 35 40
45Glu Trp Val Ala Ser Ile Ser Phe Gly Ser His Tyr Ile Ser Tyr
Ala 50 55 60Asp Ser Val Lys Gly Arg
Phe Thr Ile Ser Arg Asp Asn Ala Arg Asn65 70
75 80Ala Val Tyr Leu Gln Met Asn Ser Leu Arg Val
Glu Asp Thr Ala Val 85 90
95Tyr Tyr Cys Thr Arg Cys Arg Gly Gly Thr Arg Thr Tyr Tyr Tyr Met
100 105 110Asp Val Trp Gly Lys Gly
Thr Leu Val Thr Val Ser 115 120257126PRThomo
sapiens 257Gly Arg Ala Lys Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys
Lys1 5 10 15Pro Gly Thr
Ser Val Arg Val Ser Cys Lys Thr Ser Gly Gly Tyr Val 20
25 30Phe Ser Trp Val Arg Gln Ala Pro Gly Gln
Gly Pro Glu Trp Met Gly 35 40
45Gly Ile Ile Thr Asn Phe Gly Thr Thr Asn Tyr Ala Gln Lys Phe Gln 50
55 60Gly Arg Val Thr Ile Thr Ala Asp Lys
Ser Thr Asn Thr Val Tyr Met65 70 75
80Asp Leu Ser Asn Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr
Cys Ala 85 90 95Arg Ala
Pro Arg Gly Thr Ser Thr Ile Ala Ala Arg Phe Asn Arg Tyr 100
105 110Phe Phe Asp Ser Trp Gly Gln Gly Thr
Leu Val Thr Val Ser 115 120
125258124PRThomo sapiens 258Gly Arg Ala Glu Val Gln Leu Val Gln Ser Gly
Ala Glu Val Lys Lys1 5 10
15Pro Gly Ser Ser Val Lys Val Ser Cys Lys Thr Glu Gly Gly Thr Phe
20 25 30Ser Thr Tyr Val Ile Ser Trp
Met Arg Gln Ala Pro Gly Gln Gly Leu 35 40
45Glu Trp Met Gly Gly Ile Val Pro Ile Phe Asn Thr Pro Asn Tyr
Ala 50 55 60Gln Lys Phe Gln Gly Arg
Val Thr Ile Thr Ala Asp Arg Ser Thr Ser65 70
75 80Thr Ala Tyr Met Glu Leu Arg Ser Leu Gly Ser
Glu Asp Thr Ala Val 85 90
95Tyr Tyr Cys Ala Arg Val Val Gly Gly Thr Arg Ser Tyr Tyr Ala Leu
100 105 110Gly Phe Trp Gly Gln Gly
Thr Thr Val Thr Val Ser 115 120259122PRThomo
sapiens 259Gly Arg Ala Gln Val Thr Leu Lys Glu Ser Gly Pro Thr Leu Val
Lys1 5 10 15Pro Thr Gln
Thr Leu Thr Leu Thr Cys Thr Phe Ser Gly Phe Ser Leu 20
25 30Gly Thr Thr Gly Val Asn Val Gly Trp Ile
Arg Gln Pro Pro Gly Lys 35 40
45Ala Leu Glu Trp Leu Ala Leu Ile Ser Trp Gly Gly Gly Lys His Tyr 50
55 60Ser Pro Ser Leu Asn Ser Arg Ile Thr
Leu Thr Lys Asp Ala Ser Arg65 70 75
80Glu Gln Val Val Val Leu Thr Met Ala Asn Met Asp Pro Val
Asp Thr 85 90 95Gly Arg
Tyr Tyr Cys Ala Arg Ile Val Gly Thr His Gly Phe Asp Tyr 100
105 110Trp Gly Gln Gly Thr Leu Val Thr Val
Ser 115 120260124PRThomo sapiens 260Gly Arg Ala
Glu Val Gln Leu Val Gln Ser Gly Gly Gly Val Val Arg1 5
10 15Pro Gly Gly Ser Leu Arg Leu Ser Cys
Ala Gly Ser Gly Phe Thr Phe 20 25
30Asp Glu Tyr Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
35 40 45Glu Trp Val Ala Phe Ile Asn
Trp Asn Gly Asp Ser Thr Tyr Tyr Ala 50 55
60Asp Ser Val Lys Gly Arg Phe Thr Val Ser Arg Thr Asn Ala Lys Asn65
70 75 80Ser Leu Tyr Leu
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Phe 85
90 95Tyr Tyr Cys Ala Arg Asp Pro Arg Thr Lys
Leu Gly Met Ser Tyr Phe 100 105
110Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser 115
120261130PRThomo sapiens 261Gly Arg Ala Glu Val Gln Leu Val Gln Ser
Gly Ala Glu Val Lys Lys1 5 10
15Pro Gly Glu Ser Leu Lys Ile Ser Cys Gln Ala Ser Gly Tyr Gly Phe
20 25 30Thr Val Tyr Trp Ile Gly
Trp Val Arg Gln Leu Pro Gly Lys Gly Leu 35 40
45Glu Trp Leu Gly Ile Ile Tyr Pro Gly Asp Ser Asp Thr Arg
Tyr Asn 50 55 60Pro Ser Phe Gln Gly
Gln Val Thr Ile Ser Ala Asp Lys Ser Val Ser65 70
75 80Thr Thr Tyr Leu Gln Trp Ser Ser Leu Lys
Ala Ser Asp Thr Ala Ile 85 90
95Tyr Tyr Cys Ala Arg His Leu Asp Ser Tyr Asp Val Phe Thr Gly Tyr
100 105 110Asn Leu Gly Gly Tyr
Met Asp Val Trp Gly Lys Gly Thr Met Val Thr 115
120 125Val Ser130262123PRThomo sapiens 262Gly Arg Ala Glu
Val Gln Leu Val Gln Ser Gly Pro Glu Val Lys Lys1 5
10 15Pro Gly Ala Ser Val Arg Val Ser Cys Lys
Ala Ser Gly Tyr Ser Leu 20 25
30Lys Asn Tyr Gly Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu
35 40 45Glu Trp Met Gly Trp Ile Ser Ala
Asp Asn Gly Asp Thr Thr Thr Ala 50 55
60Leu Asn Leu Arg Gly Arg Val Ser Met Thr Thr Asp Thr Ser Thr Asn65
70 75 80Thr Val Tyr Met Glu
Val Lys Ser Leu Arg Ser Asp Asp Thr Ala Ile 85
90 95Tyr Phe Cys Ala Arg Asp Phe Tyr Ser Gly Ser
Tyr Arg Ser Phe Asp 100 105
110Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser 115
120263126PRThomo sapiens 263Gly Arg Ala Glu Val Gln Leu Val Gln Ser Gly
Pro Gly Leu Val Lys1 5 10
15Pro Ser Glu Thr Leu Ser Leu Thr Cys Ala Val Ser Gly Ala Ser Val
20 25 30Ser Gly Gly Asp Tyr Tyr Trp
Ser Trp Ile Arg Gln Pro Pro Gly Lys 35 40
45Ala Leu Glu Trp Ile Gly Tyr Ile Tyr Tyr Ile Gly Ser Thr Asn
Tyr 50 55 60Asn Pro Ser Leu Lys Ser
Arg Leu Ser Leu Ser Val Asp Thr Ala Lys65 70
75 80Ser Gln Phe Ser Leu Lys Leu Ser Ser Val Thr
Ala Ala Asp Thr Ala 85 90
95Ile Tyr Phe Cys Ala Arg Ala Arg Arg Thr Tyr Ser Gly Tyr Asp Ser
100 105 110Ala Phe Asp Tyr Trp Gly
Gln Gly Thr Leu Val Thr Val Ser 115 120
125264124PRThomo sapiens 264Gly Arg Ala Gln Val Gln Leu Gln Gln Ser
Gly Pro Gly Leu Val Lys1 5 10
15Pro Ser Glu Thr Leu Pro Leu Thr Cys Thr Val Ser Gly Gly Ser Phe
20 25 30Ser Thr Tyr Tyr Trp Ser
Trp Ile Arg Gln Ser Pro Glu Lys Gly Leu 35 40
45Glu Trp Ile Gly Tyr Ile Gln Asn Ser Val Asn Thr Asn Tyr
Asn Pro 50 55 60Ser Leu Lys Ser Arg
Val Ile Ile Ser Val Asp Thr Ser Asn Asn Gln65 70
75 80Phe Ser Leu Lys Leu Arg Ser Val Thr Ala
Ala Asp Thr Ala Val Tyr 85 90
95Tyr Cys Ala Arg Val Ser Gly Trp Gly Pro Arg Gly Gly Ile Tyr Phe
100 105 110Asp Tyr Trp Gly Gln
Gly Thr Leu Val Thr Val Ser 115 120265126PRThomo
sapiens 265Gly Arg Ala Lys Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys
Lys1 5 10 15Pro Gly Ser
Ser Val Lys Val Ser Cys Lys Ser Ser Gly Gly Tyr Val 20
25 30Phe Ser Trp Val Arg Gln Ala Pro Gly Gln
Gly Leu Glu Trp Met Gly 35 40
45Gly Ile Ile Ser Asn Phe His Thr Ala Glu Tyr Ala Gln Lys Phe Gln 50
55 60Gly Arg Val Thr Met Thr Ala Asp Thr
Ser Thr Asn Thr Ile Tyr Met65 70 75
80Glu Leu Thr Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Phe
Cys Val 85 90 95Ser Ala
Pro Arg Asp Thr Ser Thr Ile Ala Ala Arg Phe Asn Arg Tyr 100
105 110Phe Phe Asp Thr Trp Gly Gln Gly Thr
Leu Val Thr Val Ser 115 120
125266126PRThomo sapiens 266Gly Arg Ala Glu Val Gln Leu Val Gln Ser Gly
Ala Glu Val Lys Lys1 5 10
15Pro Gly Thr Ser Val Arg Val Ser Arg Lys Thr Pro Gly Gly Tyr Val
20 25 30Phe Ser Trp Val Arg Gln Ala
Pro Gly Gln Gly Pro Glu Trp Met Gly 35 40
45Gly Ile Ile Thr Asn Phe Gly Thr Thr Asn Tyr Ala Arg Lys Phe
Gln 50 55 60Gly Arg Ile Thr Val Thr
Ala Asp Lys Ser Thr Asn Thr Val Tyr Met65 70
75 80Asp Leu Ser Asn Leu Ala Ser Glu Asp Thr Ala
Val Tyr Tyr Cys Ala 85 90
95Arg Ala Pro Arg Gly Thr Ser Thr Ile Ala Ala Arg Phe Asn Arg Tyr
100 105 110Phe Phe Asp Ser Trp Ser
Gln Gly Thr Leu Val Thr Val Ser 115 120
125267124PRThomo sapiens 267Gly Arg Ala Glu Val Gln Leu Val Glu Ser
Gly Ala Glu Val Lys Lys1 5 10
15Pro Gly Ser Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Ser Phe
20 25 30Ser Thr Tyr Ile Phe Thr
Trp Val Arg Gln Ala Pro Gly Gln Gly Leu 35 40
45Glu Trp Met Gly Gly Ile Asn Pro Ile Phe Ala Thr Arg Asp
Tyr Pro 50 55 60Lys Lys Phe Gln Gly
Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Arg65 70
75 80Thr Val Tyr Met Glu Leu Ser Ser Leu Thr
Ser Glu Asp Thr Ala Val 85 90
95Tyr Tyr Cys Ala Arg Val Leu Gly Gly Thr Arg Leu Tyr Tyr Ala Leu
100 105 110Asn Val Trp Gly Gln
Gly Thr Thr Val Thr Val Ser 115 120268124PRThomo
sapiens 268Gly Arg Ala Glu Val Gln Leu Val Gln Ser Gly Gly Gly Val Val
Arg1 5 10 15Pro Gly Gly
Ser Leu Arg Leu Ser Cys Ala Gly Ser Gly Phe Thr Phe 20
25 30Asp Glu Tyr Ala Met Ser Trp Val Arg Gln
Ala Pro Gly Lys Gly Leu 35 40
45Lys Trp Val Ala Phe Ile Asn Trp Asn Gly Asp Ser Thr Tyr Tyr Ala 50
55 60Asp Ser Val Lys Gly Arg Phe Thr Val
Ser Arg Ser Asn Ala Lys Asn65 70 75
80Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr
Ala Phe 85 90 95Tyr Cys
Cys Ala Arg Asp Pro Arg Thr Lys Leu Gly Met Ser Tyr Phe 100
105 110Asp Tyr Trp Gly Gln Gly Thr Leu Val
Thr Val Ser 115 120269124PRThomo sapiens 269Gly
Arg Ala Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys1
5 10 15Pro Gly Ser Ser Val Lys Val
Ser Cys Lys Ala Ser Gly Gly Ser Phe 20 25
30Ser Thr Tyr Ala Ile Thr Trp Val Arg Gln Ala Pro Gly Gln
Gly Leu 35 40 45Glu Trp Met Gly
Gly Ile Ile Pro Ile Phe Ala Ser Arg Asp Tyr Ala 50 55
60Gln Lys Phe Gln Gly Arg Val Thr Ile Thr Ala Asp Glu
Ser Thr Arg65 70 75
80Thr Val Tyr Met Glu Pro Arg Ser Leu Arg Ser Glu Asp Thr Ala Val
85 90 95Tyr Tyr Cys Ala Arg Val
Leu Gly Gly Thr Arg Leu Tyr Tyr Ala Leu 100
105 110Asn Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser
115 120270126PRThomo sapiens 270Gly Arg Ala Glu Val
Gln Leu Val Gln Ser Gly Pro Gly Leu Val Lys1 5
10 15Pro Ser Glu Thr Leu Ser Leu Thr Cys Ala Val
Ser Gly Ala Ser Val 20 25
30Ser Gly Gly Asp Tyr Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys
35 40 45Ala Leu Glu Trp Ile Gly Tyr Ile
Tyr Tyr Ile Gly Ser Thr Asn Tyr 50 55
60Asn Pro Ser Leu Lys Ser Arg Leu Ser Leu Ser Val Asp Thr Ala Lys65
70 75 80Ser Gln Phe Ser Leu
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala 85
90 95Thr Tyr Phe Cys Ala Arg Ala Arg Arg Thr Tyr
Ser Gly Tyr Asp Ser 100 105
110Ala Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser 115
120 125271124PRThomo sapiens 271Gly Arg Ala
Glu Val Gln Leu Val Glu Ser Gly Ala Glu Val Lys Lys1 5
10 15Pro Gly Ser Ser Val Lys Val Ser Cys
Lys Ala Ser Gly Gly Ser Phe 20 25
30Ser Thr Tyr Ala Ile Thr Trp Val Arg Gln Ala Pro Gly Gln Gly Leu
35 40 45Glu Trp Met Gly Gly Ile Ile
Pro Ile Phe Ala Ser Arg Asp Tyr Ala 50 55
60Gln Lys Phe Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Arg65
70 75 80Thr Val Tyr Met
Glu Leu Ser Ser Leu Arg Ser Asp Asp Thr Ala Val 85
90 95Tyr Tyr Cys Ala Arg Val Leu Gly Gly Thr
Arg Leu Tyr Tyr Ala Leu 100 105
110Asn Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser 115
120272123PRThomo sapiens 272Gly Arg Ala Gln Val Thr Leu Lys Glu Ser
Gly Pro Thr Val Val Lys1 5 10
15Pro Thr Gln Thr Leu Thr Leu Thr Cys Ser Leu Ser Gly Phe Ala Leu
20 25 30Gly Thr Thr Gly Val Ala
Val Gly Trp Ile Arg Gln Pro Pro Gly Lys 35 40
45Ala Leu Glu Trp Leu Gly Leu Ile Asp Trp Asn Asp Asp Arg
Arg Tyr 50 55 60Arg Pro Ser Leu Lys
Thr Arg Leu Thr Ile Thr Gln Asp Met Ser Arg65 70
75 80Asn Gln Val Val Leu Arg Leu Thr Asn Leu
Asp Pro Leu Asp Thr Gly 85 90
95Thr Tyr Phe Cys Ala Arg Ser Val Val Pro Ala Thr Arg Ala Phe Asp
100 105 110Phe Trp Gly Gln Gly
Thr Leu Val Thr Val Ser 115 120273122PRThomo
sapiensMISC_FEATURE(52)..(52)Xaa unknown amino acid 273Gly Arg Ala Gln
Val Thr Leu Lys Glu Ser Gly Pro Thr Leu Val Lys1 5
10 15Pro Thr Gln Thr Leu Thr Leu Thr Cys Thr
Leu Ser Gly Phe Ser Leu 20 25
30Tyr Thr Thr Gly Val Gly Val Gly Trp Ile Arg Gln Pro Pro Gly Lys
35 40 45Ala Leu Glu Xaa Leu Ala Arg Ile
Tyr Trp Asp Asp Asp Glu Arg Tyr 50 55
60Asn Pro Ser Leu Lys Ser Arg Leu Thr Ile Thr Lys Asp Ala Ser Lys65
70 75 80Asn Gln Val Val Leu
Lys Met Thr Asn Met Asp Pro Val Asp Thr Ala 85
90 95Thr Tyr Tyr Cys Ala Arg Thr Met Gly Val Val
Leu Pro Phe Asp Tyr 100 105
110Trp Gly Gln Gly Thr Leu Val Thr Val Ser 115
120274109PRThomo sapiens 274Leu Ala Glu Ile Val Leu Thr Gln Ser Pro Ser
Ser Leu Ser Ala Ser1 5 10
15Leu Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln His Ile Ser
20 25 30Asn Tyr Leu Asn Trp Tyr Gln
Gln Lys Pro Gly Lys Ala Pro Lys Leu 35 40
45Leu Ile Cys Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg
Phe 50 55 60Thr Gly Ser Gly Ser Gly
Ala Asp Tyr Thr Leu Thr Ile Ser Ser Leu65 70
75 80Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln
Gln Ser Tyr Ser Thr 85 90
95Ser Trp Thr Phe Gly Gln Gly Thr Thr Val Glu Ile Lys 100
105275109PRThomo sapiens 275Leu Ala Glu Ile Val Leu Thr Gln
Ser Pro Ser Ser Leu Ser Ala Ser1 5 10
15Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly
Ile Ser 20 25 30Asn Tyr Leu
Ala Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu 35
40 45Leu Ile Tyr Ala Ala Ser Thr Leu Gln Ser Gly
Val Pro Ser Arg Phe 50 55 60Ser Gly
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu65
70 75 80Gln Pro Glu Asp Val Ala Thr
Tyr Tyr Cys Gln Lys Tyr Asn Ser Ala 85 90
95Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105276109PRThomo sapiens 276Leu Ala Asp Val
Val Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser1 5
10 15Leu Gly Asp Arg Val Ser Ile Thr Cys Arg
Ala Ser Gln His Ile Ser 20 25
30Asn Tyr Ile Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu
35 40 45Leu Ile Tyr Ala Ala Ser Thr Leu
Gln Ser Gly Val Pro Ser Arg Phe 50 55
60Thr Gly Ser Gly Ser Gly Ala Asp Tyr Thr Leu Thr Ile Thr Ser Leu65
70 75 80Gln Pro Glu Asp Phe
Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Gly Thr 85
90 95Ser Trp Thr Phe Gly Gln Gly Thr Thr Val Glu
Ile Lys 100 105277109PRThomo sapiens 277Leu
Ala Asp Val Val Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser1
5 10 15Val Gly Asp Arg Val Thr Ile
Thr Cys Gln Ala Ser Gln Asp Ile Thr 20 25
30Asn Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
Lys Leu 35 40 45Leu Ile Tyr Asp
Ala Ser Asn Leu Gln Pro Gly Val Pro Ser Arg Phe 50 55
60Ser Gly Ser Gly Ser Val Thr Asp Phe Thr Phe Thr Ile
Ser Ser Leu65 70 75
80Arg Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Tyr Asp Gly Pro
85 90 95Val Leu Thr Phe Gly Gly
Gly Thr Lys Val Glu Ile Lys 100
105278109PRThomo sapiens 278Leu Ala Asp Val Val Met Thr Gln Ser Pro Ser
Ser Leu Ser Ala Ser1 5 10
15Val Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln Pro Ile Ser
20 25 30Thr Tyr Leu Asn Trp Tyr Gln
Gln Lys Pro Gly Lys Ala Pro Lys Ile 35 40
45Leu Ile Phe Gly Ala Ser Arg Leu Gln Ser Gly Val Pro Ser Arg
Phe 50 55 60Ser Gly Ser Gly Ser Gly
Thr Asp Phe Ser Leu Thr Ile Thr Ser Leu65 70
75 80Gln Pro Glu Asp Phe Ala Thr Tyr Ile Cys Gln
Gln Ser Lys Ser Pro 85 90
95Pro Tyr Asn Phe Gly Arg Gly Thr Lys Leu Glu Ile Lys 100
105279110PRThomo sapiens 279Leu Ala Glu Ile Val Leu Thr Gln
Ser Pro Ser Thr Leu Ser Ala Ser1 5 10
15Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser
Ile Gly 20 25 30Ser Trp Leu
Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu 35
40 45Leu Ile Tyr Lys Ala Ser Thr Leu Gln Ser Glu
Thr Pro Ser Arg Phe 50 55 60Arg Gly
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu65
70 75 80Gln Pro Asp Asp Phe Ala Thr
Tyr Tyr Cys Gln Gln Phe Asn Ser Phe 85 90
95Ser Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Phe
Lys 100 105 110280109PRThomo
sapiens 280Leu Ala Asp Val Val Met Thr Gln Ser Pro Ser Ser Leu Ser Ala
Ser1 5 10 15Val Gly Asp
Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Asn 20
25 30Ile Asn Leu Asn Trp Phe Gln Gln Lys Pro
Gly Lys Ala Pro Asn Leu 35 40
45Leu Ile Tyr Ser Ala Ser Thr Leu Gln Thr Gly Val Pro Ser Arg Phe 50
55 60Ser Gly Ser Gly Ser Gly Thr Glu Phe
Thr Leu Thr Ile Ser Ser Leu65 70 75
80Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ile Tyr
Ser His 85 90 95Val Arg
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys 100
105281115PRThomo sapiens 281Leu Ala Asp Val Val Met Thr Gln Ser Pro Asp
Ser Leu Ala Val Ser1 5 10
15Leu Gly Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu
20 25 30Tyr Ser Ser Asn Asn Lys Asn
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro 35 40
45Gly Gln Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu
Ser 50 55 60Gly Val Pro Asp Arg Phe
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr65 70
75 80Leu Thr Ile Ser Ser Leu Gln Ala Glu Asp Val
Ala Val Tyr Tyr Cys 85 90
95Gln Gln Tyr Tyr Ser Thr Pro Pro Met Phe Gly Gln Gly Thr Lys Val
100 105 110Glu Ile Lys
115282109PRThomo sapiens 282Leu Ala Glu Ile Val Leu Thr Gln Ser Pro Ser
Ser Leu Ser Ala Ser1 5 10
15Val Gly Asp Arg Ile Thr Ile Thr Cys Gln Ala Ser Gln Asp Ile Thr
20 25 30Asn Tyr Leu Asn Trp Tyr Gln
Gln Lys Pro Gly Lys Ala Pro Lys Leu 35 40
45Leu Ile Tyr Asp Ala Ser Asn Leu Gln Pro Gly Val Pro Ser Arg
Phe 50 55 60Ser Gly Ser Arg Ser Gly
Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu65 70
75 80Arg Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln
Gln Tyr Asp Gly Pro 85 90
95Val Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys 100
105283115PRThomo sapiens 283Leu Ala Glu Ile Val Leu Thr Gln
Ser Pro Asp Ser Leu Ala Val Ser1 5 10
15Leu Gly Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser
Leu Leu 20 25 30Tyr Ser Ser
Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro 35
40 45Gly Gln Pro Pro Lys Leu Leu Ile Tyr Trp Ala
Ser Thr Arg Glu Ser 50 55 60Gly Val
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr65
70 75 80Leu Thr Ile Ser Ser Leu Gln
Ala Glu Asp Val Ala Val Tyr Tyr Cys 85 90
95Gln Gln Tyr Tyr Ser Thr Pro Pro Met Phe Gly Gln Gly
Thr Lys Val 100 105 110Glu Ile
Lys 115284110PRThomo sapiens 284Leu Ala Glu Ile Val Leu Thr Gln
Ser Pro Ser Phe Leu Ser Ala Ser1 5 10
15Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly
Ile Gly 20 25 30Asn Phe Leu
Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu 35
40 45Leu Ile Ser Gly Ala Ser Thr Leu Gln Ser Trp
Val Pro Ser Arg Phe 50 55 60Ser Gly
Arg Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu65
70 75 80Gln Pro Glu Asp Phe Ala Thr
Tyr Tyr Cys Gln Gln Leu Asn Arg Tyr 85 90
95Pro Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile
Lys 100 105 110285109PRThomo
sapiens 285Leu Ala Glu Ile Val Met Thr Gln Ser Pro Ser Ser Leu Ser Ala
Ser1 5 10 15Val Gly Asp
Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Asp Ile Ser 20
25 30Lys Tyr Leu Asn Trp Tyr Gln Gln Lys Pro
Gly Arg Ala Pro Lys Leu 35 40
45Leu Ile Tyr Glu Ala Ser Asn Leu Glu Thr Gly Val Pro Pro Arg Phe 50
55 60Ser Gly Ser Gly Ser Gly Thr His Phe
Thr Phe Thr Ile Thr Gly Leu65 70 75
80Gln Pro Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Cys Asp
Ser Leu 85 90 95Pro Pro
Val Phe Gly Gln Gly Thr Lys Leu Glu Val Lys 100
105286110PRThomo sapiens 286Leu Ala Glu Ile Val Leu Thr Gln Ser Pro Gly
Thr Leu Ala Leu Ser1 5 10
15Pro Gly Asp Arg Ala Thr Leu Ser Cys Gly Ala Ser Gln Ser Val Phe
20 25 30Gly Asp Phe Leu Ala Trp Tyr
Gln His Lys Pro Gly Gln Ala Pro Arg 35 40
45Leu Leu Ile Tyr Gly Ala Ser Thr Arg Ala Thr Gly Ile Pro Asp
Arg 50 55 60Phe Ser Gly Ser Gly Ala
Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg65 70
75 80Leu Glu Pro Glu Asp Phe Ala Val Tyr Phe Cys
Gln Gln Tyr Gly Asp 85 90
95Ser Val Phe Thr Phe Gly Gln Gly Thr Lys Leu Gly Ile Lys 100
105 110287110PRThomo sapiens 287Leu Ala
Glu Ile Val Met Thr Gln Ser Pro Gly Thr Leu Ala Leu Ser1 5
10 15Pro Gly Asp Arg Ala Thr Leu Ser
Cys Gly Ala Ser Gln Ser Val Phe 20 25
30Gly Asp Phe Leu Ala Trp Tyr Gln His Lys Pro Gly Gln Ala Pro
Arg 35 40 45Leu Leu Ile Tyr Gly
Ala Ser Thr Arg Ala Thr Gly Ile Pro Asp Arg 50 55
60Phe Ser Gly Ser Gly Ala Gly Thr Asp Phe Thr Leu Thr Ile
Ser Arg65 70 75 80Leu
Glu Pro Glu Asp Phe Ala Val Tyr Phe Cys Gln Gln Tyr Gly Asp
85 90 95Ser Val Phe Thr Phe Gly Gln
Gly Thr Lys Leu Glu Ile Lys 100 105
110288110PRThomo sapiens 288Leu Ala Glu Ile Val Leu Thr Gln Ser Pro
Gly Thr Leu Ser Leu Ser1 5 10
15Pro Gly Gln Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Asn
20 25 30Arg Asp Tyr Leu Ala Trp
Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg 35 40
45Leu Leu Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro
Asp Arg 50 55 60Phe Ser Gly Ser Gly
Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg65 70
75 80Leu Glu Arg Asp Asp Phe Ala Val Tyr Phe
Cys His Gln Tyr Gly Ser 85 90
95Ser Pro Asn Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110289110PRThomo sapiens 289Leu
Ala Glu Thr Thr Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser1
5 10 15Pro Gly Glu Arg Ala Thr Leu
Ser Cys Arg Ala Ser Gln Ser Val Ser 20 25
30Thr Asn Tyr Leu Ala Trp Tyr Arg Gln Lys Pro Gly Gln Ala
Pro Arg 35 40 45Leu Leu Ile His
Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg 50 55
60Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
Ile Ser Arg65 70 75
80Leu Glu Pro Glu Asp Phe Ala Val Tyr Phe Cys Gln Gln Tyr Gly Ser
85 90 95Ser Pro Gln Thr Phe Gly
Gln Gly Thr Arg Leu Glu Ile Lys 100 105
110290109PRThomo sapiens 290Leu Ala Glu Ile Val Leu Thr Gln Ser
Pro Ser Ser Leu Ser Ala Ser1 5 10
15Val Gly Asp Arg Val Thr Thr Thr Cys Arg Thr Ser Gln Ser Ile
Phe 20 25 30Ile Asn Leu Asn
Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu 35
40 45Leu Ile Tyr Ser Ala Ser Thr Leu Gln Thr Gly Val
Pro Ser Arg Phe 50 55 60Ser Gly Ser
Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu65 70
75 80Gln Pro Glu Asp Phe Ala Thr Tyr
Tyr Cys Gln Gln Ile Tyr Ser Gln 85 90
95Val Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105291110PRThomo sapiens 291Leu Ala Asp Ile Gln
Met Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser1 5
10 15Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala
Ser Gln Ser Val Asn 20 25
30Ser Asp Asn Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg
35 40 45Leu Leu Met Ser Gly Ala Thr Ser
Arg Ala Thr Asp Val Pro Asp Arg 50 55
60Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg65
70 75 80Leu Glu Pro Glu Asp
Phe Ala Val Tyr Tyr Cys His Gln Tyr Gly Ser 85
90 95Ser Asp Asn Thr Phe Gly Gln Gly Thr Lys Leu
Glu Ile Lys 100 105
110292110PRThomo sapiens 292Leu Ala Asp Val Val Met Thr Gln Phe Pro Ser
Ser Leu Ser Ala Ser1 5 10
15Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser
20 25 30Asn Phe Leu Ala Trp Tyr Gln
Gln Lys Pro Gly Lys Val Pro Glu Leu 35 40
45Leu Ile Tyr Gly Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg
Phe 50 55 60Ser Gly Ser Gly Ser Gly
Ala Glu Phe Thr Leu Thr Ile Asn Ser Leu65 70
75 80Gln Pro Glu Asp Val Ala Thr Tyr Tyr Cys Gln
Lys Tyr Asp Ser Gly 85 90
95Leu Arg Phe Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys 100
105 110293110PRThomo sapiens 293Leu Ala
Asp Val Val Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser1 5
10 15Val Gly Asp Arg Val Thr Ile Thr
Cys Arg Ala Ser Gln Gly Ile Ser 20 25
30Asn Phe Leu Ala Trp His Gln Gln Lys Pro Gly Gln Val Pro Lys
Leu 35 40 45Leu Ile Tyr Gly Ala
Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe 50 55
60Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn
Gly Leu65 70 75 80Gln
Pro Glu Asp Val Ala Thr Tyr Tyr Cys Gln Lys Tyr Asp Ser Gly
85 90 95Leu Ile Phe Thr Phe Gly Pro
Gly Thr Arg Val Glu Ile Lys 100 105
110294115PRThomo sapiens 294Leu Ala Asp Val Val Met Thr Gln Ser Pro
Asp Ser Leu Ala Val Ser1 5 10
15Leu Gly Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu
20 25 30Tyr Ser Ser Asn Asn Lys
Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro 35 40
45Gly Gln Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg
Glu Ser 50 55 60Gly Val Pro Asp Arg
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr65 70
75 80Leu Thr Ile Ser Ser Leu Gln Ala Glu Asp
Val Ala Val Tyr Tyr Cys 85 90
95Gln Gln Tyr Tyr Ser Thr Pro Pro Met Leu Gly Gln Gly Thr Lys Val
100 105 110Glu Ile Lys
115295110PRThomo sapiens 295Leu Ala Asp Val Val Met Thr Gln Ser Pro Gly
Thr Leu Ala Leu Ser1 5 10
15Pro Gly Asp Arg Ala Thr Leu Ser Cys Gly Ala Ser Gln Ser Val Phe
20 25 30Gly Asp Phe Leu Ala Trp Tyr
Gln His Lys Pro Gly Gln Ala Pro Arg 35 40
45Leu Leu Ile Tyr Val Ala Ser Thr Arg Ala Thr Gly Ile Pro Asp
Arg 50 55 60Phe Ser Gly Ser Gly Ala
Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg65 70
75 80Leu Glu Pro Glu Asp Phe Ala Val Tyr Phe Cys
Gln Gln Tyr Gly Asp 85 90
95Ser Val Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Arg 100
105 110296110PRThomo sapiens 296Leu Ala
Glu Thr Thr Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser1 5
10 15Pro Gly Glu Arg Ala Thr Leu Ser
Cys Arg Ala Ser Gln Ser Val Ser 20 25
30Thr Asn Tyr Leu Ala Trp Tyr Arg Gln Lys Pro Gly Gln Ala Pro
Arg 35 40 45Leu Leu Ile His Gly
Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg 50 55
60Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
Ser Arg65 70 75 80Leu
Glu Pro Glu Asp Phe Ala Val Tyr Phe Cys Gln Gln Tyr Gly Ser
85 90 95Ser Pro Gln Thr Phe Gly Gln
Gly Thr Arg Leu Glu Ile Lys 100 105
110297110PRThomo sapiens 297Leu Ala Glu Ile Val Leu Thr Gln Ser Pro
Gly Thr Leu Ser Leu Ser1 5 10
15Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Arg Ser Val Asn
20 25 30Ser Asn Asn Leu Ala Trp
Tyr Gln Gln Lys Pro Asp Gln Ala Pro Arg 35 40
45Leu Leu Met Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro
Asp Arg 50 55 60Phe Thr Gly Ser Gly
Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg65 70
75 80Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr
Cys His Gln Tyr Gly Ala 85 90
95Ser Asp Asn Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110298109PRThomo sapiens 298Leu
Ala Glu Ile Val Met Thr Gln Ser Pro Ser Phe Leu Ser Ala Ser1
5 10 15Val Gly Asp Arg Val Thr Ile
Thr Cys Arg Ala Ser Gln Asp Ile Ile 20 25
30Thr Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
Glu Val 35 40 45Leu Ile Phe Gly
Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe 50 55
60Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile
Ser Ser Leu65 70 75
80Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ile Ile Arg Tyr
85 90 95Pro Arg Thr Phe Gly Gln
Gly Thr Arg Val Glu Ile Lys 100
105299110PRThomo sapiens 299Leu Ala Asp Val Val Met Thr Gln Ser Pro Gly
Thr Leu Ser Leu Ser1 5 10
15Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Asn
20 25 30Ser Asn Asn Leu Ala Trp Tyr
Gln Gln Lys Pro Gly Gln Ala Pro Arg 35 40
45Leu Leu Val Ser Gly Ala Thr Asn Arg Ala Thr Asp Ile Pro Asp
Arg 50 55 60Phe Ser Gly Ser Gly Ser
Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg65 70
75 80Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys
His Gln Tyr Gly Ser 85 90
95Ser Glu Asn Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Arg 100
105 110300109PRThomo sapiens 300Leu Ala
Glu Ile Val Met Thr Gln Ser Pro Pro Phe Leu Ser Ala Ser1 5
10 15Val Gly Asp Arg Val Thr Ile Thr
Cys Arg Ala Ser Gln Gly Leu Ser 20 25
30Thr Tyr Leu Ala Trp Tyr Gln Val Lys Pro Gly Lys Ala Pro Lys
Leu 35 40 45Leu Ile Tyr Ala Ala
Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe 50 55
60Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Asn
Ser Leu65 70 75 80Gln
Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Leu Asp Thr Tyr
85 90 95Pro Leu Thr Leu Gly Gly Gly
Thr Lys Val Glu Ile Lys 100 105301115PRThomo
sapiens 301Leu Ala Glu Ile Val Leu Thr Gln Ser Pro Asp Ser Leu Ala Val
Ser1 5 10 15Leu Gly Glu
Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu 20
25 30Tyr Ser Ser Asn Asn Lys Asn Tyr Leu Ala
Trp Tyr Gln Gln Lys Pro 35 40
45Gly Gln Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser 50
55 60Gly Val Pro Asp Arg Phe Ser Gly Ser
Gly Ser Gly Thr Asp Phe Thr65 70 75
80Leu Thr Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr
Tyr Cys 85 90 95Gln Gln
Tyr Tyr Ser Thr Pro Pro Met Phe Gly Gln Gly Thr Lys Val 100
105 110Glu Ile Lys 115302115PRThomo
sapiens 302Leu Ala Asp Ile Gln Met Thr Gln Ser Pro Asp Ser Leu Ala Val
Ser1 5 10 15Leu Gly Glu
Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu Leu 20
25 30Tyr Ser Ser Asn Asn Lys Asn Tyr Leu Ala
Trp Tyr Gln Gln Lys Pro 35 40
45Gly Gln Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser 50
55 60Gly Val Pro Asp Arg Phe Ser Gly Ser
Gly Ser Gly Thr Asp Phe Thr65 70 75
80Leu Thr Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr
Tyr Cys 85 90 95Gln Gln
Tyr Tyr Ser Thr Pro Pro Met Phe Gly Gln Gly Thr Arg Val 100
105 110Glu Ile Lys 115303110PRThomo
sapiens 303Leu Ala Glu Ile Val Met Thr Gln Ser Pro Gly Thr Leu Ala Leu
Ser1 5 10 15Pro Gly Asp
Arg Ala Thr Leu Ser Cys Gly Ala Ser Gln Ser Val Phe 20
25 30Gly Asp Phe Leu Ala Trp Tyr Gln His Lys
Pro Gly Gln Ala Pro Arg 35 40
45Pro Leu Ile Tyr Gly Ala Ser Thr Arg Ala Thr Gly Ile Pro Asp Arg 50
55 60Phe Ser Gly Ser Gly Ala Gly Thr Asp
Phe Thr Leu Thr Ile Ser Arg65 70 75
80Leu Glu Pro Glu Asp Phe Ala Val Tyr Phe Cys Gln Gln Tyr
Gly Asp 85 90 95Ser Val
Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys 100
105 110304110PRThomo sapiens 304Leu Ala Asp Val Val
Met Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser1 5
10 15Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala
Ser Gln Ser Val Asn 20 25
30Ser Asp Asn Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg
35 40 45Leu Leu Met Ser Gly Ala Thr Ser
Arg Ala Thr Asp Ile Pro Asp Arg 50 55
60Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg65
70 75 80Leu Glu Pro Glu Asp
Phe Ala Val Tyr Tyr Cys His Gln Tyr Gly Ser 85
90 95Ser Asp Asn Thr Phe Gly Gln Gly Thr Lys Leu
Glu Ile Lys 100 105
110305110PRThomo sapiens 305Leu Ala Asp Ile Gln Leu Thr Gln Ser Pro Ser
Ser Leu Ser Ala Ser1 5 10
15Val Gly Asp Ser Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser
20 25 30Asn Phe Leu Ala Trp Tyr Gln
Lys Lys Pro Gly Lys Val Pro Arg Leu 35 40
45Leu Ile Tyr Gly Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg
Phe 50 55 60Ser Gly Ser Gly Ser Gly
Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu65 70
75 80Gln Pro Glu Asp Val Ala Thr Tyr Tyr Cys Gln
Glu Tyr Ser Asn Ala 85 90
95Leu Ile Phe Thr Phe Gly Pro Gly Thr Lys Val His Ile Lys 100
105 110306110PRThomo sapiens 306Leu Ala
Glu Ile Val Met Thr Gln Ser Pro Ser Phe Leu Ser Ala Ser1 5
10 15Val Gly Asp Arg Val Thr Ile Thr
Cys Arg Ala Ser Gln Gly Ile Gly 20 25
30Asn Phe Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys
Leu 35 40 45Leu Ile Ser Gly Ala
Ser Thr Leu Gln Ser Trp Val Pro Ser Arg Phe 50 55
60Ser Gly Arg Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser
Ser Leu65 70 75 80Gln
Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Leu Asn Arg Tyr
85 90 95Pro Pro Tyr Thr Phe Gly Gln
Gly Thr Lys Leu Glu Ile Lys 100 105
110307109PRThomo sapiens 307Leu Ala Glu Ile Val Met Thr Gln Ser Pro
Ser Ser Leu Ser Ala Ser1 5 10
15Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Thr Ile Ser
20 25 30Asn His Leu Asn Trp Tyr
Gln Gln Lys Pro Gly Arg Ala Pro Lys Leu 35 40
45Leu Ile Tyr Val Gly Ser Ser Leu Gln Ser Gly Val Pro Ser
Arg Phe 50 55 60Ser Gly Ser Gly Ser
Gly Thr Asp Phe Thr Leu Thr Ile Ser Gly Leu65 70
75 80Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
Gln Gln Ser Tyr Ser Pro 85 90
95Ser Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys 100
105308110PRThomo sapiens 308Leu Ala Asp Ile Gln Leu Thr
Gln Ser Pro Gly Thr Leu Ser Leu Ser1 5 10
15Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln
Ser Val Ser 20 25 30Thr Asn
Tyr Leu Ala Trp Tyr Arg Gln Lys Pro Gly Gln Ala Pro Arg 35
40 45Leu Leu Ile His Gly Ala Ser Ser Arg Ala
Thr Gly Ile Pro Asp Arg 50 55 60Phe
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg65
70 75 80Leu Glu Pro Glu Asp Phe
Ala Val Tyr Phe Cys Gln Gln Tyr Gly Ser 85
90 95Ser Pro Gln Thr Phe Gly Gln Gly Thr Arg Leu Glu
Ile Lys 100 105
110309109PRThomo sapiens 309Leu Ala Asp Val Val Met Thr Gln Ser Pro Ser
Ser Leu Ser Ala Ser1 5 10
15Leu Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln His Ile Ser
20 25 30Asn Tyr Leu Asn Trp Tyr Gln
Gln Lys Pro Gly Lys Ala Pro Lys Leu 35 40
45Leu Ile Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg
Phe 50 55 60Thr Gly Ser Gly Ser Gly
Ala Asp Tyr Thr Leu Thr Ile Ser Ser Leu65 70
75 80Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln
Gln Ser Tyr Ser Thr 85 90
95Ser Trp Thr Phe Gly Gln Gly Thr Thr Val Gly Ile Lys 100
105310109PRThomo sapiens 310Leu Ala Asp Val Val Met Thr Gln
Ser Pro Ser Ser Leu Ser Ala Ser1 5 10
15Val Gly Asp Arg Val Ser Ile Thr Cys Arg Ala Ser Arg Ser
Val Lys 20 25 30Thr Tyr Leu
Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu 35
40 45Leu Val Tyr Gly Ser Ser Ser Leu Glu Ser Gly
Val Pro Ser Arg Phe 50 55 60Ser Gly
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu65
70 75 80Gln Pro Glu Asp Phe Ala Thr
Tyr Tyr Cys Gln Gln Ser Phe Gly Thr 85 90
95Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Asp Thr Lys
100 105311109PRThomo sapiens 311Leu Ala Asp Ile
Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser1 5
10 15Val Ser Asp Arg Val Thr Ile Thr Cys Arg
Ala Ser Gln Ser Ile Asn 20 25
30Lys Phe Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Gln Leu
35 40 45Leu Ile Tyr Ala Ala Thr Asn Leu
Gln Ser Gly Val Pro Ser Arg Phe 50 55
60Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu65
70 75 80Gln Thr Glu Asp Phe
Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Asp Met 85
90 95Pro Arg Thr Phe Gly Gln Gly Thr Lys Val Glu
Ile Lys 100 105312110PRThomo sapiens 312Leu
Ala Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser1
5 10 15Pro Gly Glu Arg Ala Thr Leu
Ser Cys Arg Ala Ser Arg Ser Val Tyr 20 25
30Ser Asn Asn Leu Ala Trp Tyr Gln Gln Lys Pro Asp Gln Ala
Pro Arg 35 40 45Leu Leu Met Tyr
Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg 50 55
60Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
Ile Ser Arg65 70 75
80Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys His Gln Tyr Gly Ala
85 90 95Ser Asp Asn Thr Phe Gly
Gln Gly Thr Lys Leu Glu Ile Lys 100 105
110313115PRThomo sapiens 313Leu Ala Asp Ile Gln Met Thr Gln Ser
Pro Asp Ser Leu Ala Val Ser1 5 10
15Leu Gly Glu Lys Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Leu
Leu 20 25 30Tyr Ser Ser Asn
Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro 35
40 45Gly Gln Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser
Thr Arg Glu Ser 50 55 60Gly Val Pro
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr65 70
75 80Leu Thr Ile Ser Ser Leu Gln Ala
Glu Asp Val Ala Val Tyr Tyr Cys 85 90
95Gln Gln Tyr Tyr Ser Thr Pro Pro Met Phe Gly Gln Gly Thr
Lys Val 100 105 110Glu Ile Lys
115314110PRThomo sapiens 314Leu Ala Glu Ile Val Met Thr Gln Ser
Pro Ser Ser Leu Ser Ala Ser1 5 10
15Val Gly Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln Gly Ile
Ser 20 25 30Asn Phe Leu Ala
Trp Phe Gln Gln Lys Pro Gly Gln Val Pro Lys Leu 35
40 45Leu Ile Tyr Val Ala Ser Thr Leu Gln Ser Gly Val
Pro Ser Arg Phe 50 55 60Ser Gly Ser
Gly Ser Gly Thr Asp Phe Ser Leu Thr Ile Asn Gly Leu65 70
75 80Gln Pro Glu Asp Val Ala Thr Tyr
Tyr Cys Gln Arg Tyr Asp Ser Gly 85 90
95Leu Ile Phe Thr Phe Gly Pro Gly Thr Lys Val Glu Ile Lys
100 105 110315109PRThomo sapiens
315Leu Ala Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser1
5 10 15Val Gly Asp Arg Val Thr
Ile Thr Cys Arg Ala Ser Gln Thr Ile Ser 20 25
30Asn His Leu Asn Trp Tyr Gln Gln Lys Pro Gly Arg Ala
Pro Lys Leu 35 40 45Leu Ile Tyr
Val Gly Ser Ser Leu Arg Ser Gly Val Pro Ser Arg Phe 50
55 60Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
Ile Ser Gly Leu65 70 75
80Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Ser Pro
85 90 95Ser Tyr Thr Phe Gly Gln
Gly Thr Lys Val Glu Ile Lys 100
105316110PRThomo sapiens 316Leu Ala Glu Ile Val Leu Thr Gln Ser Pro Ser
Ser Leu Ser Ala Ser1 5 10
15Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser
20 25 30Asn Phe Leu Ala Trp Tyr Gln
Gln Lys Pro Gly Lys Val Pro Glu Leu 35 40
45Leu Ile Tyr Gly Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg
Phe 50 55 60Ser Gly Ser Gly Ser Gly
Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu65 70
75 80Gln Pro Glu Asp Val Ala Thr Tyr Tyr Cys Gln
Lys Tyr Asp Ser Gly 85 90
95Leu Arg Phe Thr Phe Gly Pro Gly Ala Lys Val Asp Ile Lys 100
105 110317109PRThomo sapiens 317Leu Ala
Asp Val Val Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser1 5
10 15Val Gly Asp Arg Val Ser Ile Thr
Cys Arg Ala Ser Arg Ser Val Lys 20 25
30Thr Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys
Leu 35 40 45Leu Val Tyr Gly Ser
Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe 50 55
60Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
Ser Leu65 70 75 80Gln
Pro Glu Asp Phe Ala Thr Tyr Cys Cys Gln Gln Ser Phe Gly Thr
85 90 95Pro Tyr Thr Phe Gly Gln Gly
Thr Lys Leu Asp Ile Lys 100 105318110PRThomo
sapiens 318Leu Ala Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala
Ser1 5 10 15Val Gly Asp
Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser 20
25 30Asn Phe Leu Ala Trp Tyr Gln Gln Lys Pro
Gly Lys Val Pro Glu Leu 35 40
45Leu Ile Tyr Gly Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe 50
55 60Ser Gly Ser Gly Ser Gly Thr Glu Phe
Thr Leu Thr Ile Asn Ser Leu65 70 75
80Gln Pro Glu Asp Val Ala Thr Tyr Tyr Cys Gln Lys Tyr Asp
Ser Gly 85 90 95Leu Arg
Phe Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys 100
105 110319109PRThomo sapiens 319Leu Ala Asp Ile Gln
Met Thr Gln Ser Ser Ser Thr Leu Ser Ala Ser1 5
10 15Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala
Ser Gln Asn Ile Asn 20 25
30Ile Trp Leu Ala Trp Tyr Gln Gln Lys Ala Gly Lys Ala Pro Lys Leu
35 40 45Leu Ile Tyr Lys Ala Ser Thr Leu
Glu Arg Gly Val Pro Ser Arg Phe 50 55
60Ser Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Thr Gly Leu65
70 75 80Arg Pro Asp Asp Phe
Gly Ser Tyr Tyr Cys Gln His Tyr Asp Gly Asn 85
90 95Ser Leu Thr Phe Gly Gln Gly Thr Arg Val Asp
Ile Gln 100 105320110PRThomo sapiens 320Leu
Ala Asp Ile Gln Leu Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser1
5 10 15Val Gly Asp Arg Val Thr Ile
Thr Cys Arg Ala Ser Gln Ser Ile Gly 20 25
30Ser Trp Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
Lys Leu 35 40 45Leu Ile Tyr Lys
Ala Ser Thr Leu Gln Ser Glu Thr Pro Ser Arg Phe 50 55
60Arg Gly Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile
Ser Ser Leu65 70 75
80Gln Pro Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Phe Asn Ser Phe
85 90 95Ser Pro Trp Thr Phe Gly
Gln Gly Thr Lys Ala Glu Phe Lys 100 105
110321402DNAArtificialVector sequence 321gctagcgctg gttgggcagc
gagtaataac aatccagcgg ctgccgtagg caataggtat 60ttcattatga ctgtctcctt
ggcgactagc tagtttagaa ttaattcgtg aaattgttat 120ccgctcacaa ttccacacaa
catacgagcc ggaagcataa agtgtaaagc ctggggtgcc 180taatgagtga gctaactcac
attaattgcg ttgcgctcac tgcccgcttt ccagatctta 240ctccccatcc ccctgttgac
aattaatcat cggctcgtat gatgtgtgga attgtgagcg 300gataacaatt tcacacagga
aacaggagat atacatatga aatacctgct gccgaccgct 360gctgctggtc tgctgctcct
cgctgcccag ccggggcgcg cc
40232237DNAArtificialPrimer sequence 322tattggcgcg ccatggccgc ccagtctgtg
acccagc 3732336DNAArtificialPrimer sequence
323tattggcgcg ccatggccgc ccagaagata actcaa
3632436DNAArtificialPrimer sequence 324tattggcgcg ccatggccga tgctaagacc
acccag 3632536DNAArtificialPrimer sequence
325tattggcgcg ccatggccgc ccagacagtc actcag
3632637DNAArtificialPrimer sequence 326tattggcgcg ccatggccaa acaggaggtg
acacaga 3732737DNAArtificialPrimer sequence
327tattggcgcg ccatggccgg agactcggtt acccaga
3732837DNAArtificialPrimer sequence 328tattggcgcg ccatggccgg agattcagtg
acccaga 3732937DNAArtificialPrimer sequence
329tattggcgcg ccatggccag caattcagtc aagcaga
3733036DNAArtificialPrimer sequence 330tattggcgcg ccatggccgg acaaaacatt
gaccag 3633137DNAArtificialPrimer sequence
331tattggcgcg ccatggccaa aaatgaagtg gagcaga
3733237DNAArtificialPrimer sequence 332tattggcgcg ccatggccga agaccaggtg
acgcaga 3733337DNAArtificialPrimer sequence
333tattggcgcg ccatggccgg aatacaagtg gagcaga
3733437DNAArtificialPrimer sequence 334tattggcgcg ccatggccct acatacactg
gagcaga 3733537DNAArtificialPrimer sequence
335tattggcgcg ccatggccga agacaaggtg gtacaaa
3733637DNAArtificialPrimer sequence 336tattggcgcg ccatggccga aaaccaggtg
gagcaca 3733736DNAArtificialPrimer sequence
337tattggcgcg ccatggccca ggagaatgtg gagcag
3633836DNAArtificialPrimer sequence 338tattggcgcg ccatggccgg agagagtgtg
gggctg 3633937DNAArtificialPrimer sequence
339tattggcgcg ccatggccgg agaggatgtg gagcaga
3734037DNAArtificialPrimer sequence 340tattggcgcg ccatggccag ccaaaagata
gaacaga 3734136DNAArtificialPrimer sequence
341tattggcgcg ccatggccag tcaacaggga gaagag
3634236DNAArtificialPrimer sequence 342tattggcgcg ccatggccca acaaccagtg
cagagt 3634337DNAArtificialPrimer sequence
343tattggcgcg ccatggccag ccaagaactg gagcaga
3734437DNAArtificialPrimer sequence 344tattggcgcg ccatggccgg tcaacagctg
aatcaga 3734537DNAArtificialPrimer sequence
345tattggcgcg ccatggccac ccagctgctg gagcaga
3734636DNAArtificialPrimer sequence 346tattggcgcg ccatggccca gcagcaggtg
aaacaa 3634736DNAArtificialPrimer sequence
347tattggcgcg ccatggccat actgaacgtg gaacaa
3634836DNAArtificialPrimer sequence 348tattggcgcg ccatggccga ccagcaagtt
aagcaa 3634936DNAArtificialPrimer sequence
349tattggcgcg ccatggccgg acaacaggta atgcaa
3635036DNAArtificialPrimer sequence 350tattggcgcg ccatggccaa ggaccaagtg
tttcag 3635121DNAArtificialPrimer sequence
351taggcagaca gacttgtcac t
2135243DNAArtificialPrimer sequence 352gccatggcgc gccaatagct agccggtgaa
gaagtcgccc aga 4335343DNAArtificialPrimer sequence
353gccatggcgc gccaatagct agccgatgcc atggtcatcc aga
4335443DNAArtificialPrimer sequence 354gccatggcgc gccaatagct agccgaagcc
caagtgaccc aga 4335543DNAArtificialPrimer sequence
355gccatggcgc gccaatagct agcccatgcc aaagtcacac aga
4335643DNAArtificialPrimer sequence 356gccatggcgc gccaatagct agccgacaca
gccgtttccc aga 4335743DNAArtificialPrimer sequence
357gccatggcgc gccaatagct agccgaaacg ggagttacgc aga
4335843DNAArtificialPrimer sequence 358gccatggcgc gccaatagct agccgaacct
gaagtcaccc aga 4335943DNAArtificialPrimer sequence
359gccatggcgc gccaatagct agccgaagct gacatctacc aga
4336043DNAArtificialPrimer sequence 360gccatggcgc gccaatagct agccatatct
ggagtctccc aca 4336143DNAArtificialPrimer sequence
361gccatggcgc gccaatagct agccgatgct ggaatcaccc aga
4336243DNAArtificialPrimer sequence 362gccatggcgc gccaatagct agccaatgct
ggtgtcactc aga 4336342DNAArtificialPrimer sequence
363gccatggcgc gccaatagct agccagtgct gtcgtctctc aa
4236442DNAArtificialPrimer sequence 364gccatggcgc gccaatagct agccgacgct
ggagtcacac aa 4236543DNAArtificialPrimer sequence
365gccatggcgc gccaatagct agccgatggt ggaatcactc agt
4336642DNAArtificialPrimer sequence 366gccatggcgc gccaatagct agccactgct
gggatcaccc ag 4236743DNAArtificialPrimer sequence
367gccatggcgc gccaatagct agccgaagct ggagttgccc agt
4336843DNAArtificialPrimer sequence 368gccatggcgc gccaatagct agccgaagct
ggagtggttc agt 4336943DNAArtificialPrimer sequence
369gccatggcgc gccaatagct agccgaagct ggagttactc agt
4337042DNAArtificialPrimer sequence 370gccatggcgc gccaatagct agccgatgct
gtagttacac aa 4237143DNAArtificialPrimer sequence
371gccatggcgc gccaatagct agccgatgct ggagttatcc agt
4337242DNAArtificialPrimer sequence 372gccatggcgc gccaatagct agccggtgct
ggagtctccc ag 4237343DNAArtificialPrimer sequence
373gccatggcgc gccaatagct agccgatgct gatgttaccc aga
4337443DNAArtificialPrimer sequence 374gccatggcgc gccaatagct agcctctcag
actattcatc aat 4337521DNAArtificialPrimer sequence
375caggcacacc agtgtggcct t
2137636DNAArtificialPrimer sequence 376caccttagag ctcttagagt ctctcagctg
gtacac 3637733DNAArtificialPrimer sequence
377gacattatgc atgatctctg cttctgatgg ctc
33
User Contributions:
Comment about this patent or add new information about this topic:
 |  |
 |  |
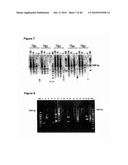 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
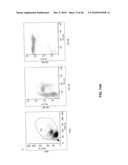 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
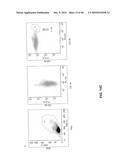 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
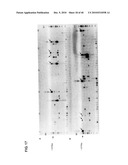 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
| Similar patent applications: | |
| Date | Title |
|---|---|
| 2014-05-08 | Nutraceutical composition for limiting the absorption of dietary lipids and for inducing weight loss, comprising, as active agent, at least one extract of carrot |
| 2014-04-03 | Styling agents with an interesting texture |
| 2014-04-10 | Botulinum toxin treatments of depression |
| 2011-10-20 | Method for increasing performance of offspring |
| New patent applications in this class: | |
| Date | Title |
|---|---|
| 2022-05-05 | Antibodies to oxidation-specific epitopes |
| 2022-05-05 | Chimeric antigen receptor comprising interleukin-15 intracellular domain and uses thereof |
| 2022-05-05 | Cd38 antibody |
| 2022-05-05 | Antibodies binding to human cd3 at acidic ph |
| 2022-05-05 | Anti-sirp alpha antibodies |
| New patent applications from these inventors: | |
| Date | Title |
|---|---|
| 2012-01-12 | Methods for recombinant manufacturing of anti-rsv antibodies |
| 2009-05-28 | Methods for recombinant manufacturing of anti-rsv antibodies |
| Top Inventors for class "Drug, bio-affecting and body treating compositions" | |
| Rank | Inventor's name |
|---|---|
| 1 | David M. Goldenberg |
| 2 | Hy Si Bui |
| 3 | Lowell L. Wood, Jr. |
| 4 | Roderick A. Hyde |
| 5 | Yat Sun Or |