Patent application title: Cottonwood Growth eIF-5A
Inventors:
John E. Thompson (Waterloo, CA)
Tzann-Wei Wang (Waterloo, CA)
Tzann-Wei Wang (Waterloo, CA)
Linda Nowack (Kitchener, CA)
Wendy Wu (Waterloo, CA)
Assignees:
Senesco Technologies, Inc.
IPC8 Class: AA01H500FI
USPC Class:
800290
Class name: Multicellular living organisms and unmodified parts thereof and related processes method of introducing a polynucleotide molecule into or rearrangement of genetic material within a plant or plant part the polynucleotide alters plant part growth (e.g., stem or tuber length, etc.)
Publication date: 2010-12-30
Patent application number: 20100333233
Inventors list |
Agents list |
Assignees list |
List by place |
Classification tree browser |
Top 100 Inventors |
Top 100 Agents |
Top 100 Assignees |
Usenet FAQ Index |
Documents |
Other FAQs |
Patent application title: Cottonwood Growth eIF-5A
Inventors:
John E. Thompson
Tzann-Wei Wang
Linda Nowack
Wendy Wu
Agents:
MORGAN LEWIS & BOCKIUS LLP
Assignees:
Origin: WASHINGTON, DC US
IPC8 Class: AA01H500FI
USPC Class:
Publication date: 12/30/2010
Patent application number: 20100333233
Abstract:
The present invention relates to unique isoforms of eukaryotic initiation
Factor 5A ("eIF-5A"): senescence-induced eIF-5A; wounding/pathogen
induced eIF-5A; stress eIF-5A and growth eIF-5A, as well as
polynucleotides that encode these three factors. The present invention
also relates to methods involving modulating the expression of these
factors. The present invention also relates to deoxyhypusine synthase
("DHS"), polynucleotides that encode DHS, and methods involving
modulating the expression of DHS.Claims:
1-2. (canceled)
3. An isolated cottonwood growth eIF-5A polynucleotide encoding a cottonwood growth eIF-5A protein wherein the protein comprises SEQ ID NO:204.
4. An isolated cottonwood growth eIF-5A polynucleotide encoding a cottonwood growth eIF-5A protein wherein the polynucleotide comprises SEQ ID NO:203.
5. A vector comprising the cottonwood growth eIF5A polynucleotide of claim 3 and regulatory sequences operatively linked to the polynucleotide to provide transcription of the polynucleotide in the plant.
6. A vector comprising the cottonwood growth eIF5A polynucleotide of claim 4 and regulatory sequences operatively linked to the polynucleotide to provide transcription of the polynucleotide in the plant.
7. A method of increasing biomass in a cottonwood plant, the method comprising incorporating into the genome of at least one cottonwood plant cell, the vector of claim 5 whereby transcription of the polynucleotide results in a plant having an increased biomass as compared to a wild type plant.
8. A method of increasing biomass in a cottonwood plant, the method comprising incorporating into the genome of at least one plant cell, the vector of claim 6 whereby transcription of the polynucleotide results in a plant having an increased biomass as compared to a wild type plant.
9. A cottonwood plant produced by the method of claim 7.
10. A cottonwood plant produced by the method of claim 8.
11. Progeny, plant parts or seeds of the cottonwood plant of claim 9 wherein the progeny, plant parts or seeds of the plant comprise the vector of claim 5.
12. Progeny, plant parts or seeds of the cottonwood plant of claim 10 wherein the progeny, plant parts or seeds of the plant comprise the vector of claim 6.
13. A vector comprising a growth eIF5A polynucleotide having at least 80% homology with SEQ ID NO: 203, and regulatory sequences operatively linked to the polynucleotide to provide transcription of the polynucleotide in the plant.
14. A method of increasing biomass in a plant, the method comprising incorporating into the genome of at least one plant cell, the vector of claim 13 whereby transcription of the polynucleotide results in a plant having an increased biomass as compared to a wild type plant.
15. The method of claim 14, wherein the plant is a cottonwood plant.
16. A plant produced by the method of claim 14.
17. Progeny, plant parts or seeds of the plant of claim 15 wherein the progeny, plant parts or seeds of the plant comprise the vector of claim 13.
18. A cotton wood plant produced by the method of claim 15.
19. Progeny, plant parts or seeds of the plant of claim 18 wherein the progeny, plant parts or seeds of the plant comprise the vector of claim 13.
Description:
GOVERNMENT INTEREST
[0001]This application is a divisional application of U.S. application Ser. No. 11/293,392, filed on Dec. 5, 2005 (now U.S. Pat. No. 7,663,028), which claims priority to U.S. provisional application 60/632,567, filed on Dec. 3, 2004, U.S. provisional 60/690,160 filed on Jun. 14, 2005, U.S. provisional 60/710,999 filed on Aug. 25, 2005 and U.S. provisional 60/728,737 filed on Oct. 21, 2005. U.S. application Ser. No. 11/293,392 is a continuation-in-part of U.S. application Ser. No. 10/862,440, filed Jun. 8, 2004 (now U.S. Pat. No. 7,358,418), which is a continuation-in-part application of Ser. No. 09/725,019, filed Nov. 29, 2000 (now U.S. Pat. No. 6,878,860), which is a continuation-in-part of 09/597,771, filed Jun. 19, 2000 (now U.S. Pat. No. 6,538,182), which is a continuation in part of 09/348,675, filed Jul. 6, 1999, now abandoned, all of which are herein incorporated by reference in their entireties.
FIELD OF THE INVENTION
[0002]The present invention relates to unique isoforms of eukaryotic initiation Factor 5A ("eIF-5A") and polynucleotides that encode eIF-5A and deoxyhypusine synthase ("DHS"), and polynucleotides that encode DHS, and methods involving modulating the expression of the isoforms eIF-5A and DHS.
DESCRIPTION OF THE PRIOR ART
[0003]Senescence is the terminal phase of biological development in the life of a plant. It presages death and occurs at various levels of biological organization including the whole plant, organs, flowers and fruit, tissues and individual cells.
[0004]The onset of senescence can be induced by different factors both internal and external. Senescence is a complex, highly regulated developmental stage in the life of a plant or plant tissue, such as fruit, flowers and leaves. Senescence results in the coordinated breakdown of cell membranes and macromolecules and the subsequent mobilization of metabolites to other parts of the plant.
[0005]In addition to the programmed senescence which takes place during normal plant development, death of cells and tissues and ensuing remobilization of metabolites occurs as a coordinated response to external, environmental factors. External factors that induce premature initiation of senescence, which is also referred to as necrosis or apoptosis, include environmental stresses such as temperature, drought, poor light or nutrient supply, as well as pathogen attack. Plant tissues exposed to environmental stress also produce ethylene, commonly known as stress ethylene (Buchanan-Wollaston, V., 1997, J. Exp. Botany, 48:181-199; Wright, M., 1974, Plant, 120:63-69). Ethylene is known to cause senescence in some plants.
[0006]Senescence is not a passive process, but, rather, is an actively regulated process that involves coordinated expression of specific genes. During senescence, the levels of total RNA decrease and the expression of many genes is switched off (Bate et al., 1991, J. Exper. Botany, 42, 801-11; Hensel et al., 1993, The Plant Cell, 5, 553-64). However, there is increasing evidence that the senescence process depends on de novo transcription of nuclear genes. For example, senescence is blocked by inhibitors of mRNA and protein synthesis and enucleation. Molecular studies using mRNA from senescing leaves and green leaves for in vitro translation experiments show a changed pattern of leaf protein products in senescing leaves (Thomas et al., 1992, J. Plant Physiol., 139, 403-12). With the use of differential screening and subtractive hybridization techniques, many cDNA clones representing senescence-induced genes have been identified from a range of different plants, including both monocots and dicots, such as Arabidopsis, maize, cucumber, asparagus, tomato, rice and potato. Identification of genes that are expressed specifically during senescence is hard evidence of the requirement for de novo transcription for senescence to proceed.
[0007]The events that take place during senescence appear to be highly coordinated to allow maximum use of the cellular components before necrosis and death occur. Complex interactions involving the perception of specific signals and the induction of cascades of gene expression must occur to regulate this process. Expression of genes encoding senescence related proteins is probably regulated via common activator proteins that are, in turn, activated directly or indirectly by hormonal signals. Little is known about the mechanisms involved in the initial signaling or subsequent co-ordination of the process.
[0008]Coordinated gene expression requires factors involved in transcription and translation, including initiation factors. Translation initiation factor genes have been isolated and characterized in a variety of organisms, including plants. Translation initiation factors can control the rate at which mRNA populations are moved out of the nucleus, the rate at which they are associated with a ribosome and to some extent can affect the stability of specific mRNAs. Zuk, et al., EMBO J. 17:2914-2925 (1998). Indeed, one such translation initiation factor, which is not required for global translation activity, is believed to shuttle specific subsets of mRNAs from the nucleus to the cytoplasm for translation. Jao, et al., J. Cell. Biochem. 86: 590-600, (2002); Wang et al., J Biol Chem 276:17541-17549 (2001); Rosorius et al., J. Cell Sci., 112, 2369-2380 (1999). This translation factor is known as the eukaryotic initiation factor 5A (eIF-5A), and is the only protein known to contain the amino acid hypusine. Park, et al., J Biol Chem 263:15264-15269 (1988).
[0009]Eukaryotic translation initiation factor 5A (eIF-5A) is an essential protein factor approximately 17 KDa in size, which is involved in the initiation of eukaryotic cellular protein synthesis. It is characterized by the presence of hypusine [N-(4-amino-2-hydroxybutyl)lysine], a unique modified amino acid, known to be present only in eIF-5A. Hypusine is formed post-translationally via the transfer and hydroxylation of the butylamino group from the polyamine, spermidine, to the side chain amino group of a specific lysine residue in eIF-5A. Activation of eIF-5A involves transfer of the butylamine residue of spermidine to the lysine of eIF-5A, forming hypusine and activating eIF-5A. In eukaryotes, deoxyhypusine synthase (DHS) mediates the post-translational synthesis of hypusine in eIF-5A. The hypusine modification has been shown to be essential for eIF-5A activity in vitro using a methionyl-puromycin assay.
[0010]Hypusine is formed on eIF-5A post-translationally through the conversion of a conserved lysine residue by the action of deoxyhypusine synthase (DHS; EC 1.1.1.249) and deoxyhypusine hydroxylase (DHH; EC 1.14.99.29). DHS has been isolated from several plant species, including tomato (GenBank Accession Number AF296077), Arabidopsis thaliana (AT-DHS; GenBank Accession Number AF296078), tobacco (Ober and Hartmann, 1999), carnation (GenBank Accession Number AF296079) and banana (GenBank Accession Number AF296080), whereas the gene for DHH has not been recognized.
[0011]DHS converts a conserved lysine residue of eIF-5A to deoxyhypusine through the addition of a butylamine group derived from spermidine. This intermediate form of eIF-5A is then hydroxylated by DHH to become hypusine. Park et al., Biol. Signals 6, 115-123 (1997). Both the deoxyhypusine and the hypusine form of eIF-5A are able to bind mRNA in vitro. Liu et al., Biol Signals 6:166-174 (1997). Although the function of eIF-5A is not fully understood, there is some evidence that it may regulate cell division (Park et al., J Biol Chem 263:15264-15269 (1998); Tome et al., Biol Signals 6:150-156, (1997)) and senescence. (Wang et al., J. Biol. Chem. 276(20): 17541-17549 (2001)). See also U.S. Pat. No. 6,538,182 and U.S. application Ser. No. 09/725,019, which are herein incorporated by reference in their entirety. It appears that several organisms are known to have more than one isoform of eIF-5A, which would suit the premise that each isoform is a specific shuttle to specific suites of mRNAs that are involved in such processes as cell division and senescence.
[0012]Wang et al. demonstrated that an increased level of DHS mRNA correlates with fruit softening and natural and stress-induced leaf senescence of tomato. Wang et al., J. Biol. Chem. 276(20): 17541-17549 (2001). Furthermore when the expression of DHS was suppressed in transgenic tomato plants by introducing a DHS antisense cDNA fragment under the regulation of a constitutive promoter, the tomato fruit from these transgenic plants exhibited dramatically delayed senescence as evidenced by delayed fruit softening and spoilage. See U.S. Pat. No. 6,538,182 and U.S. application Ser. No. 09/725,019, filed Nov. 29, 2003, incorporated herein by reference in their entirety. Since DHS is known to activate eIF-5A, these data suggest that the hypusine-modified eIF-5A (active eIF-5A) may regulate senescence through selective translation of mRNA species required for senescence. This is further demonstrated through the down-regulation of DHS in Arabidopsis thaliana ("AT") by antisense polynucleotides of the full length or 3'UTR cDNA under the control of a constitutive promoter. By down regulating Arabidopsis thaliana DHS ("AT-DHS") expression and making it less available for eIF-5A activation, senescence was delayed by approximately 2 weeks (See U.S. Pat. No. 6,538,182). Not only was senescence delayed, but also an increase in seed yield, an increase in stress tolerance and an increase in biomass were observed in the transgenic plants, where the extent of each phenotype was determined by the extent of the down-regulation of DHS. Since tomato and Arabidopsis thaliana only have one copy of DHS in their genome, as shown by Southern blot (Wang et al., 2001) and BLAST analysis, in order to target the specific eIF-5A isoform responsible for shuttling of senescence transcripts out of the nucleus, the senescence specific isoform of eIF-5A must be identified and specifically down-regulated through the antisense constructs of senescence-induced eIF-5A (of the 3' UTR) or by taking advantage of the plant's natural ability for down-regulation of an over expressed gene (i.e. creating over-expression through the use of sense polynucleotides).
[0013]Plants lack immune systems and thus, have a unique way of dealing with viruses--called co-suppression, which results in sequence-specific degradation of the viral RNA. When a transgene is under a strong constitutive promoter, like the cauliflower mosaic virus double 35S promoter, it appears as a viral transcript to the plant and sequence-specific degradation occurs, but not just of the transgene, but also the endogenous gene. (reviewed in Fagard and Vaucheret, Annual Review. Plant Physiol. Plant Mol. Biol., June; 51:167-194 (2000). There is some evidence that co-suppression may be as effective, if not more effective, than antisense suppression of expression for the down-regulation of an endogenous gene.
SUMMARY OF THE INVENTION
[0014]The present invention provides isolated isoforms of eukaryotic initiation Factor 5A ("eIF-5A"): senescence-induced eIF-5A; wounding/pathogen-induced eIF-5A; growth eIF-5A; and stress eIF-5A as well as polynucleotides that encode these isoforms. The present invention provides antisense and sense polynucleotides of the eIF-5A isoforms. The invention also provide expression vectors comprising sense and antisense polynucleotides of the eIF-5A isoforms. The present invention also relates to methods involving modulating (increasing/up-regulating or inhibiting/down-regulating) the expression of these factors, in addition to plants and their plant parts and progeny having modified expression of eIF-5A. Further, the present invention contemplates knock down mutants of these isoforms of eIF-5A.
[0015]The present invention also relates to isolated deoxyhypusine synthase ("DHS") and polynucleotides that encode DHS. The present invention also provides antisense and sense polynucleotides of DHS. The invention also provide expression vectors comprising sense and antisense polynucleotides of DHS. The present invention also relates to methods involving modulating (increasing/up-regulating or inhibiting/down-regulating) the expression of DHS in addition to plants and their plant parts and progeny having modified expression of DHS. Further, the present invention contemplates knock down mutants of DHS.
BRIEF DESCRIPTION OF THE DRAWINGS
[0016]FIG. 1 shows the alignment of three isoforms of eIF-5A isolated Arabidopsis thaliana senescence-induced eIF-5A (SEQ ID NO: 58; line 1) (previously described in U.S. Pat. No. 6,538,182 and pending application Ser. No. 09/725,019); wounding/pathogen induced eIF-5A (SEQ ID NO: 59; line 2); and growth eIF-5A (SEQ ID NO: 60; line 3). Identical amino acids are highlighted by dashed lines and the regions that were used for peptide design are indicated by the solid lines. Each peptide contains eleven amino acids from the eIF-5A sequences as well as additional cysteine residue at the N-terminus, in order for conjugation to occur with KLH.
[0017]FIG. 2 shows the alignment of the coding regions of these three Arabidopsis thaliana isoforms. Line 1 is senescence-induced eIF-5A (SEQ ID NO: 61). Line 2 is wounding/pathogen induced eIF-5A (SEQ ID NO: 62). Line 3 is growth eIF-5A (SEQ ID NO: 63). Base pairs that are identical in all three isoforms are indicated in boxes. The sequences only include the coding region from the methionine (ATG) to the stop codon.
[0018]FIG. 3 provides the genomic sequence of the senescence-induced eIF-5A of Arabidopsis thaliana (SEQ ID NO: 78). The dashed underscore indicates the areas in which the primers were designed against. The 5' end primer also contained a HindIII restriction site and the 3' end primer contained a Sad restriction site to ensure proper orientation when ligated into the binary vector. The boxed area indicates the 3' end used as a probe for Northern blots.
[0019]FIG. 4 provides the genomic sequence of the wounding/pathogen induced eIF5A of Arabidopsis thaliana (SEQ ID NO: 79). The dashed underscore indicates the areas in which the primers were designed against. The 5' end primer also contained a XhoI restriction site and the 3' end primer contained a Sad restriction site to ensure proper orientation when ligated into the binary vector. The boxed area indicates the 3' end used as probe for Northern blots.
[0020]FIG. 5 provides the genomic sequence of the growth eIF5A of Arabidopsis thaliana (SEQ ID NO: 52). The dashed underscore indicates the areas in which the primers were designed against. The 5' end primer also contained a XhoI restriction site and the 3' end primer contained a Sad restriction site to ensure proper orientation when ligated into the binary vector. The boxed area indicates the 3' end used as probe for Northern blots.
[0021]FIG. 6 is a map of binary vector pKYLX71-35S2 (SEQ ID NO: 80). The binary vector pKYLX71-35S2 contains tetracycline resistance for transformant selection in E. coli, and Kanamycin resistance for seed transformant selection on MS plates containing Kanamycin. The promoter is a duplicate 35S promoter, which serves to give higher levels of expression than a single 35S. RbcS 3' is the UTR of ribulose-1,5-bisphosphate carboxylase.
[0022]FIG. 7 is a map of binary vector pGEM®-T Easy Vector. The T overhangs in the middle of the multiple cloning sites provide the insertion site of PCR products. The Ampr gene is useful for screening transformants based on growth in the presence of ampicillin.
[0023]FIG. 8 shows Western blots of all three isoforms of eIF-5A in different tissues of Arabidopsis thaliana wild type of the Columbia ecotype. The lane descriptions are as follows: lanes labeled 2, 3, 4, 5, 6, 7 are the total resette leaves collected at 2, 3, 4, 5, 6, 7 weeks of age; Tr are leaves from plants treated with 5% PEG; U are leaves from the PEG control plants watered with water; B are closed unopened flower buds; F1 are flowers of all ages ranging from closed buds to senescent flowers; Si are siliques that were collected at 6 weeks; Se are seeds that were imbibed for 1 day; and St are stems collected at 6 weeks.
[0024]FIG. 9 are Western blots for the senescence-induced eIF-5A and the wounding/pathogen induced eIF-5A of infected leaves after 72 hours of Arabidopsis thaliana wild type of the Columbia ecotype. The expression level of senescence-induced AteIF-5A remains constant as these plants are all 4 weeks old. The expression of wounding-induced AteIF-5A increases in the virulent treated plants. The expression of growth AteIF-5A was not detectable and thus not included in the figure.
[0025]FIG. 10 are Northern blots for the three isoforms of eIF-5A in wounded leaves after 72 hours of Arabidopsis thaliana wild type of the Columbia ecotype. Leaves were wounded with a hemostat and collected at 0 hours, immediately after treatment, 1 hour after wounding, and 9 hours after wounding. The expression of wounding-induced AteIF-5A transcript is the only one that increases at 9 hours post-wounding. The expression of growth AteIF-5A, though low to begin with, decreases in the event of wounding.
[0026]FIG. 11 depicts PCR products from genomic DNA of Arabidopsis senescence-induced eIF-5A (ATeIF-5A), wounding/pathogen-induced AteIF-5A, and growth AteIF-5A in lanes 1, 2 and 3 respectively. The single top band was excised and purified for ligation into pGEM.
[0027]FIG. 12 shows an agarose gel with senescence-induced AteIF-5A, wounding/pathogen induced AteIF-5A, and growth AteIF-5A genomic sequences in pGEM. The pGEM: senescence-induced AteIF-5A, pGEM: wounding-induced AteIF-5A, and pGEM: growth AteIF-5A were digested with EcoRI to identify positive transformant colonies that contain inserts of the proper size. These clones were then sent for sequencing to confirm sequence suitability for over expression in planta.
[0028]FIG. 13 shows an agarose gel with wounding/pathogen induced AteIF-5A, growth AteIF-5A, genomic sequences in pKYLX71. The colonies that were able to grow on tetracycline containing plates were screened for either the wounding-induced AteIF-5A insert or the growth AteIF-5A insert through both double digestion (D) with appropriate enzymes and PCR(P) with the corresponding primers.
[0029]FIG. 14 is a picture of a T1 plate for plants transformed with a construct having sense wounding/pathogen induced AteIF-5A. Two transformants on this plate are circled in black and correspond to lines 13 and 14. The wild type controls are circled in white.
[0030]FIG. 15 is a picture of T1 plants transformed with sense wounding/pathogen-induced AteIF-5A at 4 weeks of age. The transgenic lines are indicated by the P tags, the wildtype plants are indicated by the W tags and the binary vector control plants are indicated by the Y tags. Lines 6, 8, 10, 13, and 14 did not produce seeds.
[0031]FIG. 16 is a picture of T1 plants transformed with sense wounding/pathogen induced AteIF-5A at 5.5 weeks of age. Only the lines that were very small are included in the figure. Lines 1, 4, and 12 all produced seed and the rest died eventually without producing seed.
[0032]FIG. 17 is a picture of T2 plants transformed with sense wounding/pathogen induced AteIF-5A at 10 days post seeding. All the T2 lines remain heterozygous as indicated by the mix of kanamycin resistant (dark plants) and non-transormants lacking kanamycin resistance (light plants). Wild type control plants are indicated in the white circles. Line 12 is not included in the figure as it only had one transformant grow and has yet to be transplanted.
[0033]FIG. 18 is a picture of T1 plants transformed with sense growth AteIF-5A at 10 days post seeding. Three transformants are indicated in black circles for this plate and correspond to lines 6, 7, and 8. Wild type control plants are indicated in the white circle.
[0034]FIG. 19 is a picture of T1 plants transformed with sense growth AteIF-5A at 4 weeks of age. The transformant lines are indicated by the B tags and wild type control by the W tags or the lack of tags. The empty binary control (Y tags) is included at the bottom of the figure showing that it looks no different than wild type.
[0035]FIG. 20 is a Western blot of T2 plants transformed with sense growth AteIF-5A lines. A representative of each mother line was used to determine the general level of expression in each line.
[0036]FIG. 21 are T2 plants transformed with sense growth AteIF-5A (Lines 1A-1D) at 4 weeks of age (top), 5 weeks of age (bottom left) and 6 weeks of age (bottom right). The transformant lines are indicated by the B tags (grey circles) and wild type control by the W tags (white ellipse). The empty binary control is indicated by the Y tags (black circles). Line 1A (indicated in the black box) will be carried through to T3.
[0037]FIG. 22 are T2 plants transformed with sense growth AteIF-5A (Lines 2A-1D) at 4 weeks of age (top), 5 weeks of age (bottom left) and 6 weeks of age (bottom right). The transformant lines are indicated by the B tags (grey circles) and wild type control by the W tags (white ellipse). The empty binary control is indicated by Y tags (black circles). Line 2D (indicated in the black box) will be carried through to T3.
[0038]FIG. 23 are T2 plants transformed with sense growth AteIF-5A (Lines 4A-D) at 4 weeks of age (top), 5 weeks of age (bottom left) and 6 weeks of age (bottom right). The transformant lines are indicated by the B tags (grey circles) and wild type controls by the W tags (white ellipse). The empty binary control is indicated by the Y tags (black circle). Line 4D (indicated in the black box) will be carried through to T3.
[0039]FIG. 24 are T2 plants transformed with sense growth AteIF-5A (Lines 15A-D) at 4 weeks of age (top), 5 weeks of age (bottom left) and 6 weeks of age (bottom right). The transformant lines are indicated by the B tags (grey circles) and wild type control by the W tags (white ellipse). The empty binary control is indicated by the Y tags (black circles). Line 15A (indicated in the black box) will be carried through to T3.
[0040]FIG. 25 are T2 plants transformed with sense growth AteIF-5A (Lines 8A-D) at 4 weeks of age (top), 5 weeks of age (bottom left) and 6 weeks of age (bottom right). The transformant lines are indicated by the B tags (grey circles) and wild type control by the W tags (white ellipse). The empty binary control is indicated by the Y tags (black circles). Line 8D (indicated in the black box) will be carried through to T3.
[0041]FIG. 26 are T2 plants transformed with sense growth AteIF-5A (Lines 9E-H) at 4 weeks of age (top), 5 weeks of age (bottom left) and 6 weeks of age (bottom right). The transformant lines are indicated by the B tags (grey circles) and wild type control by the W tags (white ellipse). The empty binary control is indicated by the Y tags (black circles). Line 9H (indicated in the black box) will be carried through to T3.
[0042]FIG. 27 are T2 plants transformed with sense growth AteIF-5A (Lines 11A-D) at 4 weeks of age (top), 5 weeks of age (bottom left) and 6 weeks of age (bottom right). The transformant lines are indicated by the blue tags and wild type control by the white tags. The empty binary control is indicated by yellow tags. Line 11C (indicated in the blue box) will be carried through to T3.
[0043]FIG. 28 are T2 plants transformed with sense growth AteIF-5A (Lines 16A-D) at 4 weeks of age (top), 5 weeks of age (bottom left) and 6 weeks of age (bottom right). The transformant lines are indicated by the B tags (grey circles) and wild type control by the W tags (white ellipse). The empty binary control is indicated by the Y tags (black circles). Line 16C (indicated in the black box) will be carried through to T3.
[0044]FIG. 29 are photographs of Arabidopsis thaliana seeds from various plant lines (including wild type control and plant lines having been transformed with sense growth AteIF-5A). Lines 11C and 16C are only 88 and 87% of the average wild type seed size, whereas lines 2D and 2H are 273 and 299% larger than wild type, respectively.
[0045]FIG. 30 is a bar graph of average seed size for each plant subline having been transformed with sense growth AteIF-5A. Each line has sublines A-H not labeled separately in the figure. The binary control and the wild type controls correspond to the last two bars. The standard errors as represented by the error bars were calculated with n=10.
[0046]FIG. 31 is a bar graph of individual seed weight for each plant subline having been transformed with sense growth AteIF-5A. Each line has sublines A-H. The binary control and the wild type controls correspond to the last two bars.
[0047]FIG. 32 is a graph showing the proportional relationship between the weight of the individual seeds versus the volume of individual seeds.
[0048]FIG. 33 is a bar graph showing seed yield per plant for each plant subline having been transformed with sense growth AteIF-5A. Each line has sublines A-H. The binary control and the wild type controls correspond to the last two bars.
[0049]FIG. 34 is a summary of phenotypes displayed in T2 sense growth AteIF-5A plants. The phenotypes are categorized based on the level of expression as determined by Western blotting. The lines that demonstrate high level of expression are blocked in crosshatching, the lines that demonstrate medium levels of expression are blocked with single shading lines, and the lines that demonstrated low or no expression are in white. The lines showing no expression may be due to cosupression.
[0050]FIG. 35 shows a comparison of transgenic arabidopsis plant (transformed with antisense full length senescence-induced eIF-5A) with a wild type plant. The transgenic plant exhibits delayed senescence. The transgenic plant is dwarfed, has an increased number of small rosette leaves, and exhibits delayed senescence.
[0051]FIGS. 36-38 show photographs of a plant (transformed with antisense growth eIF-5A).
[0052]FIG. 39 shows the primers used to construct the vector for generating antisense arabidopsis thaliana 3' DHS. Forward and reverse primers are SEQ ID NO: 81 and SEQ ID NO: 82, respectively. The amplified Arabidopsis DHS sequences are shown in SEQ ID NO: 83 and a portion thereof is shown in SEQ ID NO: 84.
[0053]FIG. 40 shows the vector construct.
[0054]FIG. 41 shows the sequence for wounding/pathogen factor eIF-5A isolated from arabidopsis (DNA sequence disclosed as SEQ ID NO: 54; protein disclosed as SEQ ID NO: 55) and the location of the antisense construct. Primers disclosed as SEQ ID NOS: 85-86.
[0055]FIG. 42 shows the vector construct (SEQ ID NOS: 87-89, respectively, in order of appearance).
[0056]FIG. 43 shows plate counts of leaf discs inoculated with pseudomonas.
[0057]FIG. 44 shows a graph of CFUs in antisense transgenic plants versus wild-type.
[0058]FIG. 45 depict the nucleotide sequence of the tomato leaf DHS cDNA sequence (SEQ ID NO:1) and the derived amino acid sequence (SEQ ID NO. 2) obtained from a tomato leaf cDNA library.
[0059]FIG. 46 depicts the nucleotide sequence of an Arabidopsis DHS gene obtained by aligning the tomato DHS sequence with unidentified genomic sequences in the Arabidopsis gene bank (SEQ ID NO:5; protein disclosed as SEQ ID NO: 6). The gaps between amino acid sequences are predicted introns. FIG. 46B depicts the derived Arabidopsis DHS amino acid sequence (SEQ ID NO:6). FIG. 46C depicts the nucleotide sequence of a 600 base pair Arabidopsis DHS cDNA obtained by PCR (SEQ ID NO: 26). FIG. 46D depicts the derived amino acid sequence of the Arabidopsis DHS cDNA fragment (SEQ ID NO: 92).
[0060]FIG. 47 is an alignment of the derived full length tomato leaf DHS amino acid sequence (Residues 1-369 of SEQ ID NO: 2) and the derived full length Arabidopsis senescence-induced DHS amino acid sequence (SEQ ID NO: 6) with sequence of DHS proteins of human (SEQ ID NO: 3), yeast (SEQ ID NO: 45), fungi (SEQ ID NO: 44), and Archaeobacteria (SEQ ID NO: 46). Identical amino acids among three or four of the sequences are boxed.
[0061]FIG. 48 is a restriction map of the tomato DHS cDNA.
[0062]FIG. 49 is a Southern blot of genomic DNA isolated from tomato leaves and probed with 32P-dCTP-labeled full length tomato DHS cDNA.
[0063]FIG. 50 is a Northern blot of RNA isolated from tomato flowers at different stages of development. The top panel is the ethidium bromide stained gel of total RNA. Each lane contains 10 μg RNA. The bottom panel is an autoradiograph of the Northern blot probed with 32P-dCTP-labeled full length tomato DHS cDNA.
[0064]FIG. 51 is a Northern blot of RNA isolated from tomato fruit at various stages of ripening that was probed with 32P-dCTP-labeled full length tomato DHS cDNA. Each lane contains 10 μg RNA.
[0065]FIG. 52 is a Northern blot of RNA isolated from tomato leaves that had been drought-stressed by treatment with 2 M sorbitol for six hours. Each lane contains 10 μg RNA. The blot was probed with 32P-dCTP-labeled full length tomato DHS cDNA.
[0066]FIG. 53 is a Northern blot of RNA isolated from tomato leaves that had been exposed to chilling temperature. FIG. 53A is the ethidium bromide stained gel of total RNA. Each lane contained 10 μg RNA. FIG. 53B is an autoradiograph of the Northern blot probed with 32P-dCTP-labeled full length tomato DHS cDNA. FIG. 53C shows corresponding leakage data measured as conductivity of leaf diffusates.
[0067]FIG. 54 is the carnation DHS full-length (1384 base pairs) cDNA clone nucleotide sequence (SEQ ID NO: 9) not including the PolyA tail and 5' end non-coding region. The derived amino acid sequence is shown below the nucleotide sequence (373 amino acids). (SEQ ID NO:10)
[0068]FIG. 55 is a Northern blot of total RNA from senescing Arabidopsis leaves probed with 32P-dCTP-labeled full-length Arabidopsis DHS cDNA. The autoradiograph is at the top, the ethidium stained gel below.
[0069]FIG. 56 is a Northern blot of total RNA isolated from petals of carnation flowers at various stages. The blot was probed with 32P-dCTP-labeled full-length carnation DHS cDNA. The autoradiograph is at the top, the ethidium stained gel below.
[0070]FIG. 57 is the nucleotide (top) (SEQ ID NO:11) and derived amino acid (bottom) (SEQ ID NO:12) sequence of the tomato fruit senescence-induced eIF-5A gene.
[0071]FIG. 58 is the nucleotide (top) (SEQ ID NO:13) and derived amino acid (bottom) (SEQ ID NO:14) sequence of the carnation senescence-induced eIF-5A gene.
[0072]FIG. 59 is the nucleotide (top) (SEQ ID NO:15) and derived amino acid (bottom) (SEQ ID NO:16) sequence of the Arabidopsis senescence-induced eIF-5A gene.
[0073]FIG. 60 is a Northern blot of total RNA isolated from leaves of Arabidopsis plants at various developmental stages. The blot was probed with 32P-dCTP-labeled full-length Arabidopsis DHS cDNA and full-length senescence-induced eIF-5A. The autoradiograph is at the top, the ethidium stained gel below.
[0074]FIG. 61 is a Northern blot of total RNA isolated from tomato fruit at breaker (BK), red-firm (RF) and red-soft (RS) stages of development. The blot was probed with 32P-dCTP-labeled full-length DHS cDNA and full-length senescence-induced eIF-5A. DHS and eIF-5A are up-regulated in parallel in red-soft fruit coincident with fruit ripening. The autoradiograph is at the top, the ethidium stained gel below.
[0075]FIG. 62 is a Northern blot of total RNA isolated from leaves of tomato that were treated with sorbitol to induce drought stress. C is control; S is sorbitol treated. The blot was probed with 32P-dCTP-labeled full-length DHS cDNA and full-length senescence-induced eIF-5A. Both eIF-5A and DHS are up-regulated in response to drought stress. The autoradiograph is at the top, the ethidium stained gel below.
[0076]FIG. 63 is a Northern blot of total RNA isolated from flower buds and open senescing flowers of tomato plants. The blot was probed with 32P-dCTP-labeled full-length senescence-induced DHS cDNA and full-length senescence-induced eIF-5A. Both eIF-5A and DHS are up-regulated in open/senescing flowers. The autoradiograph is at the top, the ethidium stained gel below.
[0077]FIG. 64 is a Northern blot of total RNA isolated from chill-injured tomato leaves. The blot was probed with 32P-dCTP-labeled full-length DHS cDNA and full-length senescence-induced eIF-5A. Both eIF-5A and DHS are up-regulated with the development of chilling injury during rewarming The autoradiograph is at the top, the ethidium stained gel below.
[0078]FIG. 65 is a photograph of 3.1 week old Arabidopsis wild-type (left) and transgenic plants expressing the 3'-end of the DHS gene (sequence shown in FIG. 80) in antisense orientation showing increased leaf size in the transgenic plants.
[0079]FIG. 66 is a photograph of 4.6 week old Arabidopsis wild-type (left) and transgenic plants expressing the 3'-end of the DHS gene (sequence shown in FIG. 80) in antisense orientation showing increased leaf size in the transgenic plants.
[0080]FIG. 67 is a photograph of 5.6 week old Arabidopsis wild-type (left) and transgenic plants expressing the 3'-end of the DHS gene (sequence shown in FIG. 80) in antisense orientation showing increased leaf size in the transgenic plants.
[0081]FIG. 68 is a photograph of 6.1 week old Arabidopsis wild-type (left) and transgenic plants expressing the 3'-end of the DHS gene (sequence shown in FIG. 80) in antisense orientation showing increased size of transgenic plants.
[0082]FIG. 69 is a graph showing the increase in seed yield from three T1 transgenic Arabidopsis plant lines expressing the DHS gene in antisense orientation. Seed yield is expressed as volume of seed. SE for n=30 is shown for wild-type plants.
[0083]FIG. 70 is a photograph of transgenic tomato plants expressing the 3'-end of the DHS gene (sequence shown in FIG. 80) in antisense orientation (left) and wild-type plants (right) showing increased leaf size and increased plant size in the transgenic plants. The photograph was taken 18 days after transfer of the plantlets to soil.
[0084]FIG. 71 is a photograph of transgenic tomato plants expressing the 3'-end of the DHS gene (sequence shown in FIG. 36) in antisense orientation (left) and wild-type plants (right) showing increased leaf size and increased plant size in the transgenic plants. The photograph was taken 32 days after transfer of the plantlets to soil.
[0085]FIGS. 72 through 79 are photographs of tomato fruit from wild-type (top panels) and transgenic plants expressing the full-length DHS gene in antisense orientation (bottom panels). Fruit were harvested at the breaker stage of development and ripened in a growth chamber. Days after harvest are indicated in the upper left corner of each panel.
[0086]FIG. 80 is the nucleotide (top) (SEQ ID NO:30) and derived amino acid (bottom) (SEQ ID NO: 90) sequence of the 3'-end of the Arabidopsis senescence-induced DHS gene used in antisense orientation to transform plants.
[0087]FIG. 81 is the nucleotide (top) (SEQ ID NO:31) and derived amino acid (bottom) (SEQ ID NO: 91) sequence of the 3'-end of the tomato DHS gene used in antisense orientation to transform plants.
[0088]FIG. 82 is the nucleotide (top) (SEQ ID NO:26) and derived amino acid (bottom) (SEQ ID NO: 92) sequence of a 600 base pair Arabidopsis DHS probe used to isolate the full-length Arabidopsis gene.
[0089]FIG. 83 is the nucleotide (top) (SEQ ID NO:27) and derived amino acid (bottom) (SEQ ID NO: 93) sequence of the 483 base pair carnation DHS probe used to isolate the full-length carnation gene.
[0090]FIGS. 84 (a) and (b) are photographs of tomato fruits from transgenic tomato plants expressing the 3'-end of the DHS gene (sequence shown in FIG. 81) in antisense orientation (right) and tomato fruits from wild-type plants (left). While the wild-type fruit exhibits blossom end rot, the transgenic fruit does not.
[0091]FIG. 85 shows the alignment of various isoforms of eIF-5A from several plant species. It also provides alignment of the hypusine conserved region. SEQ ID NOS: 4, 94-106 and 108-125 are shown, respectively, in order of appearance.
[0092]FIG. 86 provides tomato senescence-induced eIF-5A polynucleotide (SEQ ID NO: 126) and amino acid (SEQ ID NO: 127) sequences.
[0093]FIG. 87 provides Arabidopsis senescence-induced eIF-5A and the construction of pKYLX71-sense senescence-induced eIF-5A. The primer sequences are disclosed as SEQ ID NOS: 128-129, respectively, in order of appearance. The vector sequences are disclosed as SEQ ID NOS: 130-132, respectively, in order of appearance.
[0094]FIG. 88 provides tomato senescence-induced eIF-5A and the construction of pKYLX71-sense senescence-induced eIF-5A. The primer sequences are disclosed as SEQ ID NOS: 133-134, respectively, in order of appearance. The vector sequences are disclosed as SEQ ID NOS: 135-137, respectively, in order of appearance.
[0095]FIG. 89 provides photographs of a comparison of Arabidopsis thaliana control and transgenic plants comprising a sense polynucleotide senescence-induced eIF-5A. The transgenic plant has thicker inflorescence stems over that of the control plant.
[0096]FIGS. 90 and 91 shows that transgenic plants comprising an sense polynucleotide senescence-induced eIF-5A (FIG. 90--arabidopsis and FIG. 91--tomato) have increased xylogenesis as indicated by the increased xylem in the transgenic plant. The xylem zones were stained grey with phlorogucinol-HCl, bar=100 μm.
[0097]FIG. 92 provides photographs of a comparison of Arabidopsis thaliana control and Arabidopsis thaliana transgenic plants comprising a sense polynucleotide senescence-induced eIF-5A. A tomato sense polynucleotide senescence-induced eIF-5A was used in Arabidopsis thaliana. The transgenic plant has thicker inflorescence stems over that of the control plant.
[0098]FIGS. 93 and 94 are bar graphs that show increased xylogenesis in transgenic plants comprising a sense polynucleotide senescence-induced eIF-5A. FIG. 94 concerned tomato sense polynucleotide senescence-induced eIF-5A.
[0099]FIG. 95 provides canola growth eIF-5A amino acid (SEQ ID NO: 67) and polynucleotide (SEQ ID NO: 66) sequences.
[0100]FIG. 96 provides canola growth eIF-5A and the construction of pKYLX71-sense growth eIF-5A. The primer is disclosed as SEQ ID NO: 138. The vector sequences are disclosed as SEQ ID NOS 139-141, respectively, in order of appearance.
[0101]FIG. 97 provides canola DHS amino acid (SEQ ID NO: 71) and polynucleotide (SEQ ID NO: 70) sequences.
[0102]FIG. 98 provides canola DHS and the construction of pKYLX71-sense DHS. The 3'-UTR sequence is disclosed as SEQ ID NO: 142. The vector sequences are disclosed as SEQ ID NOS 143-145, respectively.
[0103]FIG. 99 shows in bar graph form that inhibition of DHS expression increases seed yield in canola.
[0104]FIG. 100 shows in bar graph form that up regulation of growth isoforms of eIF-5A from left to right arabidopsis, canola, tomato, and up regulation of tomato DHS.
[0105]FIG. 101 provides tomato growth eIF-5A amino acid (SEQ ID NO: 65) and polynucleotide (SEQ ID NO: 64) sequences.
[0106]FIG. 102 provides tomato growth eIF-5A and the construction of pKYLX71-sense tomato growth eIF-5A. The primer sequences are disclosed as SEQ ID NOS 146-147, respectively, in order of appearance. The vector sequences are disclosed as SEQ ID NOS 148-150, respectively, in order of appearance.
[0107]FIG. 103 provides tomato wounding/pathogen induced eIF-5A amino acid (SEQ ID NO: 57) and polynucleotide (SEQ ID NO: 56) sequences.
[0108]FIG. 104a and b provides tomato wounding/pathogen induced eIF-5A and the construction of pKYLX71-sense tomato wounding/pathogen induced eIF-5A. The primer sequences are disclosed as SEQ ID NOS 151-152, respectively, in order of appearance. The vector sequences are disclosed as SEQ ID NOS 153-155, respectively, in order of appearance.
[0109]FIG. 105 provides portions of lettuce DHS polynucleotide sequences. The primer sequences are disclosed as SEQ ID NOS 156-157, respectively, in order of appearance. The lettuce sequences are disclosed as SEQ ID NOS 158-159, respectively, in order of appearance.
[0110]FIG. 106 provides the construct of pTA7001-3'UTR antisense lettuce DHS.
[0111]FIGS. 107A and B provides alfalfa DHS amino acid and polynucleotide sequences.
[0112]FIG. 107A discloses nucleotides 1-1861 of SEQ ID NO: 72 coding SEQ ID NO: 73.
[0113]FIG. 107B discloses SEQ ID NOS: 72 (nucleotide) and 73 (protein).
[0114]FIGS. 108 A and B provides banana DHS amino acid (SEQ ID NO: 75) and polynucleotide (SEQ ID NO: 74) sequences.
[0115]FIGS. 109 A and B provides cottonwood DHS amino acid (SEQ ID NO: 77) and polynucleotide (SEQ ID NO: 76) sequences.
[0116]FIG. 110 provides partial mycosphaerella fijiensis DHS amino acid and polynucleotide sequences. Figure discloses SEQ ID NOS: 68, 160, 69, 161-163, 165, 163 coding 164/53, 47 (primer), 107 and 53, respectively, in order of appearance.
[0117]FIG. 111 shows a schematic of the activation of eIF-5A.
[0118]FIG. 112 is a Northern analysis of carnation aging and expression of eIF-5A.
[0119]FIG. 113 shows that transgenic carnations (antisense senescence-induced eIF-5A) show longer shelf life of cut flowers.
[0120]FIG. 114 shows delayed spoilage of bananas (transgenic plants with antisense DHS).
[0121]FIG. 115 shows delayed spoilage of tomatoes (transgenic plants with antisense DHS).
[0122]FIG. 116 shows expression of arabidopsis senescence-induced eIF-5A (AteIF-5A1) in cotyledons, leaves and flowers.
[0123]FIG. 117 shows delayed senescence of Arabidopsis leaves in transgenic arabidopsis (arabidopsis antisense senescence-induced eIF-5A) (AteIF-5A1).
[0124]FIG. 118 shows expression of arabidopsis senescence-induced eIF-5A::GUS in first true leaf pair.
[0125]FIG. 119 shows transgenic plants (antisense senescence-induced eIF-5A) have less xylem than wild type plants.
[0126]FIG. 120 shows enhanced xylem formation in transgenic plants (sense constructs of senescence-induced eIF-5A).
[0127]FIG. 121 shows resistance to drought in transgenic plants (reduced expression of stress-induced eIF-5A through antisense constructs).
[0128]FIG. 122 is a bar graph showing survival in wild type and transgenic Arabidopsis under drought stress. Transgenic plants (reduced expression of stress-induced eIF-5A through antisense constructs) have increased survival.
[0129]FIG. 123 shows a comparison of wild type canola and transgenic canola (reduced expression of stress-induced eIF-5A through antisense constructs) and shows that transgenic canola has an increased biomass.
[0130]FIG. 124 shows transgenic canola lines (reduced expression of stress-induced eIF-5A through antisense constructs) have an increase in seed yield over wild type plants.
[0131]FIG. 125 indicates that wounding/pathogen induced eIF-5A is up-regulated after infection with P. syringae.
[0132]FIG. 126 shows that antisense suppression of wounding/pathogen-induced isoform of eIF-5A inhibits infection by virulent pathogens. Transgenic plants showed a greater than 99% increase in an inhibition of infection over control plants.
[0133]FIGS. 127-130 show that transgenic canola (antisense wounding/pathogen-induced isoform of eIF-5A) shows a resistance to infection by Sclerotinia sclerotiorum. "WT" is a leaf from a wild type plant. "TG-1" and "TG-5" are leaves from transgenic plants.
[0134]FIG. 131 shows a photograph of transgenic bananas (antisense DHS) (left side) and wild type bananas (right side) and shows that transgenic banana plants have a larger biomass than the wild type control.
[0135]FIG. 132 shows transgenic tomatoes (growth eIF-5A in sense orientation) (left side) have an increased biomass over wild type control plants (right side).
[0136]FIG. 133 shows transgenic canola (growth eIF-5A in sense orientation) have an increased biomass over wild type control plants.
[0137]FIG. 134 shows transgenic canola (growth eIF-5A in sense orientation) have an increased seed yield over wild type control plants.
[0138]FIG. 135 provides lettuce DHS sequences. The nucleic acid is SEQ ID NO: 166 and the amino acid sequence is SEQ ID NO: 167.
[0139]FIG. 136 provides a partial petunia DHS nucleotide (SEQ ID NO: 170) and amino acid sequence (SEQ ID NO: 171) as well as two primers used to isolate the DHS sequences (SEQ ID NOS: 168-169)
[0140]FIG. 137 provides alfalfa nucleic acid sequence for wounding/pathogen eIF-5A (SEQ ID NO: 172) and the amino acid sequence (SEQ ID NO: 173).
[0141]FIG. 138 provides alfalfa nucleic acid sequence for senescence eIF-5A (SEQ ID NO: 174) and the amino acid sequence (SEQ ID NO: 175).
[0142]FIG. 139 provides alfalfa nucleic acid sequence for stress eIF-5A (SEQ ID NO: 176) and the amino acid sequence (SEQ ID NO: 177).
[0143]FIG. 140 provides alfalfa nucleic acid sequence for growth eIF-5A (SEQ ID NO: 178) and the amino acid sequence (SEQ ID NO: 179).
[0144]FIGS. 141-144 provide partial sequences for different isoforms of eIF-5A in lettuce (LF5A-1, LF5A-3, LFA-2, and LFA-1). FIG. 141 discloses SEQ ID NOS: 180 (nucleotide) and 181 (protein). FIG. 142 discloses SEQ ID NOS: 182 (nucleotide) and 183 (protein). FIG. 143 discloses SEQ ID NOS: 184 (nucleotide) and 185 (protein). FIG. 144 discloses SEQ ID NOS: 186-187 and 210-211, respectively, in order of appearance.
[0145]FIG. 145 provides the sequences for growth eIF-5A sequences for tomato (nucleic acid--SEQ ID NO: 188) and amino acid (SEQ ID NO: 189).
[0146]FIG. 146 provides the sequences for stress eIF-5A sequences in tomato (nucleic acid--SEQ ID NO: 190) and amino acid (SEQ ID NO: 191).
[0147]FIGS. 147A-C provides nucleic acid and amino acid eIF-5A sequences isolated from four eIF-5A from cottonwood trees ("F1," (SEQ ID NO: 192 coding SEQ ID NO: 193) "F3," (SEQ ID NO: 194 coding SEQ ID NO: 195) "F3" (SEQ ID NO: 196 coding SEQ ID NO: 197) and "F4" (SEQ ID NO: 198 coding SEQ ID NO: 199)).
[0148]FIGS. 148 (a) and (b) provides an amino acid alignment of the various DHS showing the highly conserved nature of DHS across species. Figure discloses SEQ ID NOS: 45, 2, 200, 73, 77, 201, 10, 167, 75, 202 and 3, respectively, in order of appearance.
[0149]FIG. 149 shows the hydroponic conditions used for growing lettuce having a down-regulation of DHS.
[0150]FIG. 150 shows delayed browning in cut lettuce having expression of lettuce DHS down-regulated.
[0151]FIG. 151 shows a Western blot indicating that transgenic lettuce having antisense 3' end of lettuce DHS exhibit a decreased expression of DHS.
[0152]FIG. 152 shows a vector constructed used to transform Arabidopsis with antisense 3' end of DHS.
[0153]FIG. 153 shows arabidopsis plants having DHS down regulated have an increased growth rate/biomass over wild type plants (transgenic arabidopsis generated with the vector shown in FIG. 152).
[0154]FIG. 154 shows arabidopsis plants having DHS down regulated have an increased growth rate/biomass over wild type plants (transgenic arabidopsis generated with the vector shown in FIG. 152).
[0155]FIG. 155 shows arabidopsis plants having DHS down regulated have an increased seed yield over wild type plants (transgenic arabidopsis generated with the vector shown in FIG. 152).
[0156]FIG. 156 shows arabidopsis plants having DHS down regulated have an increased root development over wild type plants (transgenic arabidopsis generated with the vector shown in FIG. 152).
[0157]FIG. 157 shows canola plants having DHS down regulated have an increased growth rate/biomass over wild type plants.
[0158]FIG. 158 shows a Western blot indicating that antisense DHS has caused a decreased expression of DHS.
[0159]FIG. 159 shows canola plants having DHS down regulated have an increase in silique size over wild type plants.
[0160]FIG. 160 shows canola plants having DHS down regulated have an increase in seed yield over wild type plants.
[0161]FIGS. 161(a) and (b) shows that in canola plants having DHS down regulated, there is no change in the quantity of oil per seed and no change is the fatty acid composition of the oil.
[0162]FIG. 162 shows the effect of introducing cottonwood growth eIF-5A into arabidopsis in sense orientation to achieve up-regulation of growth eIF-5A. The resulting plants have an increased growth rate/biomass.
[0163]FIG. 163 shows the effect of introducing cottonwood growth eIF-5A into arabidopsis in sense orientation to achieve up-regulation of growth eIF-5A. The resulting plants have an increased growth rate/biomass.
[0164]FIG. 164 shows the effect of introducing cottonwood growth eIF-5A into arabidopsis in sense orientation to achieve up-regulation of growth eIF-5A. The resulting plants have an increased growth rate/biomass.
[0165]FIG. 165 show that there is an increase in seed yield in canola plants when either a canola or arabidopsis growth eIF-5A is used in sense orientation in canola plants.
[0166]FIG. 166 shows that a transgenic arabidopsis having been transformed with a mutant senescence eIF-5A (not able to be hypusinated) has increased biomass over wild type arabidopsis.
[0167]FIG. 167 shows that a transgenic arabidopsis having been transformed with a mutant senescence eIF-5A (not able to be hypusinated) has increased biomass over wild type arabidopsis.
[0168]FIG. 168 shows that growth eIF5A isolated from cottonwood (populas deltoides) expressed in sense orientation in arabidopsis, results in an increase in seed yield over the control plants.
[0169]FIG. 169 compares the seed yield from three different transgenic arabidopsis lines having cottonwood growth factor eIF5A expressed in sense orientation, showing that the three lines all had an increase of seed yield.
[0170]FIG. 170 provides the full length cDNA (SEQ ID NO: 203) of populus deltoides (cottonwood) growth eIF-5A and the full length amino acid sequence (SEQ ID NO: 204) of the protein. Figure legend discloses nucleotides 40-57 and 530-550 of SEQ ID NO: 203.
[0171]FIG. 171 provides the vector construct used to transform arabidopsis with cottonwood growth eIF5A in the sense orientation. The primer sequences are disclosed as SEQ ID NOS 205-206, respectively, in order of appearance. The vector sequences are disclosed as SEQ ID NOS 207-209, respectively, in order of appearance.
[0172]FIGS. 172 and 173 show that transgenic arabidopsis plants (having been transformed with the vector shown in FIG. 171) have increased biomass over control plants when treated with different levels of fertilizer, including optimal, suboptimal, and supraoptimal concentrations.
[0173]FIGS. 174 and 175 show that transgenic arabidopsis plants (having been transformed with a mutant senescence-induced eIF-5a) have increased biomass over control plants when treated with different levels of fertilizer, including optimal, suboptimal, and supraoptimal concentrations.
DETAILED DESCRIPTION
[0174]As used herein, the term "plant" refers to either a whole plant, a plant part, a plant cell or a group of plant cells. The type of plant which can be used in the methods of the invention is not limited and includes, for example, ethylene-sensitive and ethylene-insensitive plants; fruit bearing plants such as apricots, apples, oranges, bananas, grapefruit, pears, tomatoes, strawberries, avocados, etc.; vegetables such as carrots, peas, lettuce, cabbage, turnips, potatoes, broccoli, asparagus, etc.; flowers such as carnations, roses, mums, etc.; agronomic crop plants such as corn, rice, soybean, alfalfa and the like, and forest species such as deciduous trees, conifers, evergreens, etc., and in general, any plant that can take up and express DNA molecules of the present invention. It may include plants of a variety of ploidy levels, including haploid, diploid, tetraploid and polyploid. The plant may be either a monocotyledon or dicotyledon.
[0175]A transgenic plant is defined herein as a plant which is genetically modified in some way, including but not limited to a plant which has incorporated heterologous or homologous senescence-induced eIF-5A, wounding/pathogen induced eIF-5A, growth eIF-5A, stress eIF-5A or DHS into its genome. The altered genetic material may encode a protein, comprise a regulatory or control sequence, or may be or include an antisense sequence or sense sequence or encode an antisense RNA or sense RNA which is antisense or sense to senescence-induced eIF-5A, wounding/pathogen induced eIF-5A, growth eIF-5A, stress eIf-5A or DHS DNA or mRNA sequence or portion thereof of the plant.
[0176]The term "hybridization" as used herein is generally used to mean hybridization of nucleic acids at appropriate conditions of stringency as would be readily evident to those skilled in the art depending upon the nature of the probe sequence and target sequences. Conditions of hybridization and washing are well known in the art, and the adjustment of conditions depending upon the desired stringency by varying incubation time, temperature and/or ionic strength of the solution are readily accomplished. See, for example, Sambrook, J. et al., Molecular Cloning: A Laboratory Manual, 2nd edition, Cold Spring Harbor Press, Cold Spring Harbor, N.Y., 1989. The choice of conditions is dictated by the length of the sequences being hybridized, in particular, the length of the probe sequence, the relative G-C content of the nucleic acids and the amount of mismatches to be permitted. Low stringency conditions are preferred when partial hybridization between strands that have lesser degrees of complementarity is desired. When perfect or near perfect complementarity is desired, high stringency conditions are preferred. What is meant herein as high stringency conditions is as follows: the hybridization solution contains 6×S.S.C., 0.01 M EDTA, 1×Denhardt's solution and 0.5% SDS. Hybridization is carried out at about 68° C. for about 3 to 4 hours for fragments of cloned DNA and for about 12 to about 16 hours for total eukaryotic DNA. For lower stringencies the temperature of hybridization is reduced to about 42° C. below the melting temperature (TM) of the duplex. The TM is known to be a function of the G-C content and duplex length as well as the ionic strength of the solution.
[0177]As used herein, the term "substantial sequence identity" or "substantial homology" is used to indicate that a nucleotide sequence or an amino acid sequence exhibits substantial structural or functional equivalence with another nucleotide or amino acid sequence. Any structural or functional differences between sequences having substantial sequence identity or substantial homology will be de minimis; that is, they will not affect the ability of the sequence to function as indicated in the desired application. Differences may be due to inherent variations in codon usage among different species, for example. Structural differences are considered de minimis if there is a significant amount of sequence overlap or similarity between two or more different sequences or if the different sequences exhibit similar physical characteristics even if the sequences differ in length or structure. Such characteristics include, for example, ability to hybridize under defined conditions, or in the case of proteins, immunological crossreactivity, similar enzymatic activity, etc. Each of these characteristics can readily be determined by the skilled practitioner by art known methods.
[0178]Additionally, two nucleotide sequences are "substantially complementary" if the sequences have at least about 70 percent, more preferably, 80 percent and most preferably about 90 percent sequence similarity between them. Two amino acid sequences are substantially homologous if they have at least 70% similarity between the active portions of the polypeptides.
[0179]As used herein, the phrase "hybridizes to a corresponding portion" of a DNA or RNA molecule means that the molecule that hybridizes, e.g., oligonucleotide, polynucleotide, or any nucleotide sequence (in sense or antisense orientation) recognizes and hybridizes to a sequence in another nucleic acid molecule that is of approximately the same size and has enough sequence similarity thereto to effect hybridization under appropriate conditions. For example, a 100 nucleotide long antisense molecule from the 3' coding or non-coding region of tomato wounding/pathogen induced eIF-5A will recognize and hybridize to an approximately 100 nucleotide portion of a nucleotide sequence within the 3' coding or non-coding region, respectively of AT wounding/pathogen induced eIF-5A gene or any other plant wounding/pathogen induced eIF-5A gene so long as there is about 70% or more sequence similarity between the two sequences. It is to be understood that the size of the "corresponding portion" will allow for some mismatches in hybridization such that the "corresponding portion" may be smaller or larger than the molecule which hybridizes to it, for example 20-30% larger or smaller, preferably no more than about 12-15% larger or smaller.
[0180]The term "functional derivative" of a nucleic acid (or polynucleotide) as used herein means a fragment, variant, homolog, or analog of the gene or nucleotide sequence encoding senescence-induced eIF-5A, wounding/pathogen induced eIF-5A, growth eIF-5A or DHS. A functional derivative retains at least a portion of the function of the senescence-induced eIF-5A, wounding/pathogen induced eIF-5A, growth eIF-5A, stress eIF-5A or DHS encoding DNA, which permits its utility in accordance with the invention. Such function may include the ability to hybridize under high stringency conditions with native isolated senescence-induced eIF-5A, wounding/pathogen induced eIF-5A, growth eIF-5A, stress eIF-5A or DHS or substantially homologous DNA from another plant or an mRNA transcript thereof, and which senescence-induced eIF-5A, wounding/pathogen induced eIF-5A, growth eIF-5A, stress eIF-5A or DHS in antisense orientation inhibits expression of senescence-induced eIF-5A, wounding/pathogen induced eIF-5A, growth eIF-5A, stress eIF-5A or DHS.
[0181]A "fragment" of the gene or DNA sequence refers to any subset of the molecule, e.g., a shorter polynucleotide or oligonucleotide. A "variant" refers to a molecule substantially similar to either the entire gene or a fragment thereof, such as a nucleotide substitution variant having one or more substituted nucleotides, but which maintains the ability to hybridize with the particular gene or to encode mRNA transcript which hybridizes with the native DNA. A "homolog" refers to a fragment or variant sequence from a different plant genus or species. An "analog" refers to a non-natural molecule substantially similar to or functioning in relation to either the entire molecule, a variant or a fragment thereof.
[0182]By "modulating expression" it is meant that either the expression is inhibited or up-regulated. "Inhibition of expression" refers to the absence or detectable decrease in the level of protein and/or mRNA product from a target gene, such as senescence-induced eIF-5A, wounding/pathogen induced eIF-5A, growth eIF-5A, stress eIF-5A or DHS. "Up-regulation" or "over expression" refers to a detectable increase in the level of protein and/or mRNA product from a target gene, such as senescence-induced eIF-5A, wounding/pathogen induced eIF-5A, growth eIF-5A, stress eIF-5A or DHS. Included within the meaning of modulating expression, are decreasing the activity of DHS or any one of the isoforms of eIF-5A. For example, one may create a knock down mutant of DHS in a plant and the resulting plant will have a DHS that activates eIF-5A at a lower level or efficiency of a wild type DHS. Modulating expression also includes the use of chemicals or drugs that inhibit or decrease the activity of DHS or eIF-5A.
[0183]Isolated polynucleotides and peptides of the present invention include those isolated from natural sources, recombinantly produced or synthesized. Isolated proteins of the present invention include senescence-induced eIF-5A, wounding/pathogen induced eIF-5A, growth eIF-5A or DHS expressed as a fusion protein, preferably comprising eIF-5A or DHS fused with maltose binding protein.
[0184]"Functional derivatives" of senescence-induced eIF-5A, wounding/pathogen induced eIF-5A, growth eIF-5A, stress eIF-5A or DHS peptides of the present invention include fragments, variants, analogs, or chemical derivatives, which retain at least a portion of the activity or immunological cross reactivity with an antibody specific for the eIF-5A isoform or DHS. A fragment of eIF-5A or DHS peptide refers to any subset of the molecule. Variant peptides may be made by direct chemical synthesis, for example, using methods well known in the art. An analog of eIF-5A or DHS peptide refers to a non-natural protein substantially similar to either the entire protein or a fragment thereof. Chemical derivatives of eIF-5A or DHS contain additional chemical moieties not normally a part of the peptide or peptide fragment. Modifications may be introduced into peptides or fragments thereof by reacting targeted amino acid residues of the peptide with an organic derivatizing agent that is capable of reacting with selected side chains or terminal residues.
[0185]A eIF-5A or DHS peptide according to the invention may be produced by culturing a cell transformed with a nucleotide sequence of this invention (in the sense orientation), allowing the cell to synthesize the protein and then isolating the protein, either as a free protein or as a fusion protein, depending on the cloning protocol used, from either the culture medium or from cell extracts. Alternatively, the protein can be produced in a cell-free system. Ranu, et al., Meth. Enzymol., 60:459-484, (1979).
[0186]Preparation of plasmid DNA, restriction enzyme digestion, agarose gel electrophoresis of DNA, polyacrylamide gel electrophoresis of protein, PCR, RT-PCR, Southern blots, Northern blots, DNA ligation and bacterial transformation were carried out using conventional methods well-known in the art. See, for example, Sambrook, J. et al., Molecular Cloning: A Laboratory Manual, 2nd ed., Cold Spring Harbor Press, Cold Spring Harbor, N.Y., 1989. Techniques of nucleic acid hybridization are disclosed by Sambrook.
[0187]Procedures for constructing recombinant nucleotide molecules in accordance with the present invention are disclosed in Sambrook, et al., In: Molecular Cloning: A Laboratory Manual, 2nd ed., Cold Spring Harbor Press, Cold Spring Harbor, N.Y. (1989), and Maniatis, T. et al., Molecular mechanisms in the Control of Gene expression, eds., Nierlich, et al., eds., Acad. Press, N.Y. (1976), which are both incorporated herein in its entirety.
[0188]Transgenic plants made in accordance with the present invention may be prepared by DNA transformation using any method of plant transformation known in the art. Plant transformation methods include, but are not limited to, direct co-cultivation of plants, tissues or cells with Agrobacterium tumefaciens or direct infection (Miki, et al., Meth. in Plant Mol. Biol. and Biotechnology, (1993), p. 67-88); direct gene transfer into protoplasts or protoplast uptake (Paszkowski, et al., EMBO J., 12:2717 (1984); electroporation (Fromm, et al., Nature, 319:719 (1986); particle bombardment (Klein et al., BioTechnology, 6:559-563 (1988); injection into meristematic tissues of seedlings and plants (De LaPena, et al., Nature, 325:274-276 (1987); injection into protoplasts of cultured cells and tissues (Reich, et al., BioTechnology, 4:1001-1004 (1986)).
[0189]Generally a complete plant is obtained from the transformation process. Plants may be regenerated from protoplasts, callus, tissue parts or explants, etc. Plant parts obtained from regenerated transgenic plants in which the expression of an eIF-5A isoform or DHS is altered, such as leaves, flowers, fruit, seeds, pollen and the like are included in the definition of "plant" as used herein. Progeny, variants and mutants of the regenerated plants are also included in the definition of "plant."
eIF-5A Generally
[0190]The present invention relates to eIF-5A (eukaryotic translation initiation factor 5A). eIF-5A regulates programmed cell death in natural senescence of leaves, flowers and fruit; stress-induced premature senescence, cell death associated with xylogenesis and cell death associated with pathogen ingression. eIF-5A is post translationally activated by two enzymes deoxyhypusine synthase (DHS) and deoxyhypusine hydrolase (DHH). See FIG. 111. eIF-5A is not a conventional translation initiation factor but rather functions as a shuttle protein. Three or four distinct isoforms of eIF-5A have been identified. The three different isoforms selectively translocate different subpopulations of mRNAs from the nucleus to ribosomes for translation. One isoform controls cell division and growth, and two or three isoforms control cell death. For example, FIG. 1 shows three different isoforms of eIF-5A isolated in Arabidopsis. FIGS. 8-10 show Western blots probed with isoform specific antibodies against eIF-5A ("At" means arabidopsis). These figures show expression levels of the three different isoforms and different life cycles and events in Arabidopsis. In plants with larger genomes, e.g. crop plants, there are four isoforms of eIF-5A. One isoform is involved in cell division and cell growth and three isoforms are involved in cell death.
[0191]The present invention relates to different isoforms of eIF-5A: senescence-induced eIF-5A; wounding induced eIF-5A; stress eIF-5A, and growth eIF-5A. Sensecence-induced eIF-5A induces senescence at the end of the lifespan of a plant or plant tissue. Stress eIF-5A is also a death isoform that induces senescence prematurely in response to environmental stress e.g. drought stress. Disease/wounding elf-5A is a death isoform that induces cell death in response to pathogen ingression or a wounding event. Growth eIF-5A promotes plant growth. eIF-5A appears to function as a biological switch, regulating growth in one position and death in another. In the death position, one or more of the death isoforms are strongly expressed, and the growth isoform is only weakly expressed or not expressed at all. In the growth position, the growth isoform is strongly expressed, and the death isoforms are only weakly expressed or not expressed at all.
[0192]Although there are 3 to 4 isoforms of eIF-5A, it is believed that there is only one isoform of DHS, and this single isoform activates (hypusinates) all of the isoforms of eIF-5A.
[0193]The present invention provides various isoforms of eIF-5A isolated from various plant species and methods of isolating the isoforms of eIF-5A. The present invention also provides polynucleotides that encode the various isoforms of eIF-5A of the present invention. The invention also provides antisense polynucleotides of the isoforms of eIF-5A and expression vectors containing such polynucleotides or antisense polynucleotides. In some embodiments, there are provided methods of inhibiting expression of endogenous eIF-5A through the use of expression vectors containing antisense polynucleotides of the isoforms of eIF-5A to transform plants. In some embodiments, there are provided methods of up-regulating endogenous eIF-5A isoforms by providing expression vectors containing polynucleotides of the isoforms of eIF-5A in the sense orientation.
[0194]The different isoforms are naturally up or down-regulated depending upon the life stage of the plant or the plant's condition. For example in senescing tissues, the senescence-induced eIF-5A isoform is up-regulated. The senescence-induced eIF-5A is thought to participate in further senescence of the plant or plant tissues by shuttling specific subsets of mRNAs (those involved in the senescence pathway) from the nucleus to the cytoplasm for translation. By down regulating or inhibiting the expression of senescence-induced eIF-5A, senescence can be delayed in the plant and/or plant tissues. Delayed senescence is manifested in the transformed/transgenic plants by having a larger bio-mass, increased shelf life for fruit, increased shelf life of flowers, increased seed size and increased seed yield as compared to non-transformed or wild type plants.
[0195]When a plant and/or plant tissues are exposed to a wounding event, such as chilling, dehydration, or mechanical forces, or exposed to a pathogen, wounding/pathogen induced eIF-5A isoform is up-regulated. By down regulating the expression of wounding/pathogen induced eIF-5A, an increased resistance to virulent damage arising from pathogen ingression is conferred on the plants as compared to resistance to virulent damage in non-transformed or wild type plants.
[0196]When a plant is in the growth phase, growth eIF-5A isoform is up-regulated. By up-regulating growth eIF-5A, the resulting transgenic plants have an increased seed size, increased biomass and increased seed yield.
[0197]When certain plants undergo stress, stress eIF-5A isoform is up-regulated. By down regulating stress eIF-5A, the resulting transgenic plants show an increased tolerance to the stress, such as an increased tolerance to drought. Further, these plants have an increased seed yield over wild type plants.
[0198]FIG. 1 shows the alignment of three isoforms of eIF-5A isolated from Arabidopsis thaliana ("At"). FIG. 2 shows the alignment of the coding regions of these three isoforms. FIGS. 3-5 provide the genomic sequence of the three isoforms.
[0199]Western blots (see FIG. 8) show the expression of three isoforms of eIF-5A at different plant life stages. FIG. 8 reveals that the amount of the senescence-induced factor eIF-5A isoform increases as the ages of the leaves increases. It is not seen in the unopened flower buds, siliques or stems but it is seen in the imbibed seeds. In the imbibed seeds there is cotyledon tissue as well as growing embryo. Thus, senescence-induced eIF-5A is present in the imbibed seeds because the cotyledon tissue is senescing as the embryo is growing. Growth eIF-5A is seen in the imbibed seeds because there the embryo is actively growing. The wounding/pathogen induced eIF-5A is seen in the siliques, seeds and stems as the harvesting of these tissues induces some wounding.
[0200]Although there is a high degree of homology (about 85%) between the different isoforms and between the isoforms in different plant species, the different isoforms vary from each other in the 3'UTR. One region that is highly conserved between the isoforms and between species as well, is the area that is believed to be the hypusine site. The hypusine site is believed to be the following amino acids: 5'-CKVVEVSTSKTGKHGHAKCHFV-3' (SEQ ID NO: 32). See FIG. 85 for alignment of various eIF-5A isoforms and of several plant species.
Senescence-Induced eIF-5A
[0201]Senescence-induced eIF-5A is expressed in senescing tissues. See FIG. 116. The present invention relates to the discovery of senescence-induced eIF-5A in various plants including Arabidopsis thaliana, tomato, carnation, lettuce and alfalfa plants. Senescence-induced eIF-5A is up-regulated in senescing tissues and is involved in the induction of senescence related morphological changes in plants and plant tissues. When the senescence isoform is down regulated, natural senescence is delayed. Down regulation of senescence-induced eIF-5A (either through the use of antisense constructs of senescence-induced eIF-5A, use of sense constructs to achieve co-suppression, knock down mutants of senescence-induced eIF-5A, decreased expression of DHS (which is required to activate eIF-5A), knock down mutants of DHS, or use of inhibitors of DHS) results in various morphological changes in the transgenic plants, including increased plant bio-mass, delayed fruit softening or spoilage, delayed browning of cut flowers or plant tissues, such as lettuce leaves, increased seed yield and increased seed size. See FIGS. 35, 36 and 117 (delayed leaf senescence in Arabidopsis), FIGS. 112 and 113 (longer shelf life of cut carnation flowers), FIG. 114 (delayed browning in bananas), FIG. 115 (delayed spoilage of tomato), and FIG. 119 (decreased xylem formation).
[0202]When senescence-induced eIF-5A is up-regulated with sense constructs of senescence-induced eIF-5A, a gain-of-function phenotype results. For example, an increase in the formation of xylem is noticed. See FIG. 120.
[0203]To further elucidate the roles of DHS and senescence-induced eIF-5A, the promoter activities of both DHS and senescence-induced eIF-5A were characterized during growth and development of transgenic Arabidopsis plants using GUS as a reporter gene. FIG. 118 shows that senescence-induced eIF-5A is expressed in vascular tissues. The levels of DHS::GUS and senescence-induced eIF-5A::GUS expression were strongly upregulated at the onset of leaf senescence, and this high expression was maintained until leaf senescence terminated. Furthermore, and unexpectedly, DHS and senescence-induced eIF-5A were transiently and co-ordinately expressed in the vascular tissues of developing cotyledons, rosette leaves and stems. The timing and localization of the GUS expression imply a role for senescence-induced eIF-5A in xylogenesis. In addition, DHS and senescence-induced eIF-5A were co-ordinately expressed in another tissues during the degenerative stages of pollen production. Taken together, these results suggest that DHS and senescence-induced eIF-5A regulate programmed cell death associated with xylogenesis, leaf senescence and flower senescence in Arabidopsis.
[0204]Senescence-induced eIF-5A is also expressed in developing xylem. In order to determine whether senescence-induced eIF-5A plays a role in xylem formation, its full-length cDNA was constitutively over-expressed in transgenic Arabidopsis plants. These plants had larger rosette leaves as well as taller and thicker inflorescence stems in comparison to control plants, reflecting more rapid growth. In addition, their main inflorescence stems exhibited a significant enhancement in xylem development. In both vascular and interfascicular regions, cell layers and tissue thicknesses were increased by up to 80% and 60%, respectively, depending on the line. Increased xylem growth was more pronounced in secondary xylem than in primary xylem. Moreover, constitutive antisense suppression of senescence-induced eIF-5A resulted in stunted statures and delayed leaf senescence as well as ˜30% reduction in xylem tissue in comparison with control plants.
[0205]eIF5A is post-translationally modified by conversion of a conserved lysine to hypusine. To confirm that it is the hypusinated form of senescence-induced eIF-5A that regulates xylogenesis, we also generated transgenic Arabidopsis plants with constitutive overexpression of a mutant form of senescence-induced eIF-5A that cannot be hypusinated. The mutation was made by changing the codon, AAG, for the conserved lysine to GCG (for alanine), thereby disabling hypusination of senescence-induced eIF-5A. In the mutant population, there was no change in xylem development relative to control plants. Thus, senescence-induced eIF-5A appears to positively regulate xylogenesis, possibly by facilitating the programmed death of treachery elements, and such a function requires activation of the protein through hypusination.
[0206]Thus, one embodiment of the present invention is isolated senescence-induced eIF-5A from Arabidopsis thaliana. The amino acid sequence is provided in FIG. 59 and is SEQ ID NO: 16. The polynucleotide encoding the amino acid is provided in FIG. 59 and is SEQ ID NO: 15.
[0207]Another embodiment of the present invention is isolated senescence-induced eIF-5A from tomato. Amino acid sequences (SEQ ID NOS: 12, 127) and polynucleotides encoding the same (SEQ ID NOS: 11, 126, respectively) are shown in FIG. 57 (SEQ ID NOS: 11-12) and 86 (SEQ ID NOS: 126-127).
[0208]Another embodiment of the present invention is isolated senescence-induced eIF-5A from carnation. The amino acid sequence is provided in FIG. 58 and is SEQ ID NO: 14. The polynucleotide encoding the amino acid is provided in FIG. 58 and is SEQ ID NO: 13.
[0209]Another embodiment of the present invention is isolated senescence-induced eIF-5A from alfalfa. The amino acid sequence is provided in FIG. 138 and is SEQ ID NO: 175. The polynucleotide encoding the amino acid is provided in FIG. 138 and is SEQ ID NO: 174.
[0210]The present invention also provides isolated polynucleotides of senescence-induced eIF-5A that have 65-100% sequence homology to the above enumerated SEQ ID NOs, and hybridize under high stringency conditions to the complement of the enumerated SEQ ID NOs and which encode senescence-induced eIF-5A.
[0211]The present invention also provides senescence-induced eIF-5A isolated polypeptides of senescence-induced eIF-5A that have 65-100% amino acid sequence homology to the above enumerated SEQ ID NOs.
[0212]The present invention also provides antisense polynucleotides of the senescence-induced eIF-5As. The antisense polynucleotides may be of any length as long as they are able to inhibit expression. In some embodiments the antisense polynucleotides comprise the full length coding sequence and in other particularly preferred embodiments the antisense polynucleotides are directed at the 3'UTR since the different isoforms of eIF-5A have a higher degree of variation at the 3'UTR. In some embodiments the antisense polynucleotides are directed at the 5'-non-coding sequence. Antisense polynucleotides primarily complementary to 5'-non-coding sequences are known to be effective inhibitors of expression of genes encoding transcription factors. Branch, M. A., Molec. Cell Biol., 13:4284-4290 (1993).
[0213]The term "antisense polynucleotide of senescence-induced eIF5A" as used herein and in the claims encompasses not only those antisense polynucleotides that share 100% homology of the complement of an enumerated SEQ ID NO but also includes those antisense polynucleotides that are a functional variants. Functional variants are those variants, either natural or man made, that have at least 80% sequence homology to and hybridizes under high stringency conditions with the corresponding portion of the senescence-induced eIF-5A. Further the variant must have the function as intended by the present invention, that is it is capable of modulating expression of endogenous senescence-induced eIF-5A when introduced into an expression vector and wherein such vector is incorporated into the genome of at least one plant cell. One skilled in the art can appreciate that insubstantial changes can be made in the sequence that would not detrimentally effect the ability of the antisense polynucleotide to bind to the transcript and reduce or inhibition expression of the gene. Thus, the term "antisense polynucleotide" encompasses those polynucleotides that are substantially complementary to the transcript and that still maintain the ability to specifically bind to the transcript and inhibit or reduce gene expression. For a general discussion of antisense see Alberts, et al., Molecular Biology of the Cell, 2nd ed., Garland Publishing, Inc. New York, N.Y., 1989 (in particular pages 195-196, incorporated herein by reference).
[0214]One embodiment of the present invention provides expression vectors comprising either the senescence-induced eIF-5A polynucleotides (of the present invention as described above) or antisense polynucleotides of senescence-induced eIF-5A (of the present invention as described above). Vectors can be plasmids, preferably, or may be viral or other vectors known in the art to replicate and express genes encoded thereon in plant cells or bacterial cells. The vector becomes chromosomally integrated such that it can be transcribed to produce the desired antisense polynucleotide of senescence-induced eIF-5A RNA. Such plasmid or viral vectors can be constructed by recombinant DNA technology methods that are standard in the art. For example, the vector may be a plasmid vector containing a replication system functional in a prokaryotic host and an antisense polynucleotide according to the invention. Alternatively, the vector may be a plasmid containing a replication system functional in Agrobacterium and an antisense polynucleotide according to the invention. Plasmids that are capable of replicating in Agrobacterium are well known in the art. See, Miki, et al., Procedures for Introducing Foreign DNA Into Plants, Methods in Plant Molecular Biology and Biotechnology, Eds. B. R. Glick and J. E. Thompson. CRC Press (1993), PP. 67-83.
[0215]The vector further comprises regulatory sequences operatively linked to the polynucleotides to allow expression of such polynucleotides. The regulatory sequences may include a promoter functional in the transformed plant cell. The promoter may be inducible, constitutive, or tissue specific. Such promoters are known by those skilled in the art.
[0216]Promoter regulatory elements that are useful in combination with the various isoforms of eIF-5A and DHS of the present invention to generate sense or antisense transcripts of the gene include any plant promoter in general, and more particularly, a constitutive promoter such as the fig wart mosaic virus 35S promoter, the cauliflower mosaic virus promoter, CaMV35S promoter, or the MAS promoter, or a tissue-specific or senescence-induced promoter, such as the carnation petal GST1 promoter or the Arabidopsis SAG12 promoter (See, for example, J. C. Palaqui et al., Plant Physiol., 112:1447-1456 (1996); Morton et al., Molecular Breeding, 1:123-132 (1995); Fobert et al., Plant Journal, 6:567-577 (1994); and Gan et al., Plant Physiol., 113:313 (1997), incorporated herein by reference). Preferably, the promoter used in the present invention is a constitutive promoter. The SAG12 promoter is preferably preferred when using antisense polynucleotides of senescence-induced eIF-5A. See example 23.
[0217]Expression levels from a promoter which is useful for the present invention can be tested using conventional expression systems, for example by measuring levels of a reporter gene product, e.g., protein or mRNA in extracts of the leaves, flowers, fruit or other tissues of a transgenic plant into which the promoter/reporter gene have been introduced. An exemplary reporter gene is GUS.
[0218]Optionally, the regulatory sequences include a 5' non-translated leader sequence or a polyadenylation signal or enhancers. The present invention further contemplates other regulatory sequences as known by those skilled in the art.
[0219]The invention also provides a transgenic plant cell transformed with a vector or combination of vectors of the present invention comprising polynucleotides of senescence-induced eIF-5A in sense or antisense orientation, a transgenic plantlet or mature transgenic plant generated from such a cell, or a plant part, such as a flower, fruit, leaves, seeds, etc. of the transgenic plant.
[0220]The present invention also provides methods of inhibiting expression of endogenous senescence-induced eIF-5A. These methods comprise integrating into the genome of at least one cell of a plant, expression vectors of the present invention comprising antisense polynucleotides of senescence-induced eIF-5A. The antisense polynucleotides of senescence-induced eIF-5A are transcribed and inhibit expression of endogenous senescence-induced eIF-5A.
[0221]In another method of inhibiting expression of endogenous senescence-induced eIF-5A, an expression vector containing a senescence-induced eIF-5A polynucleotide of the present invention in a sense orientation is integrated into the genome of at least one cell of a plant. The polynucleotide of senescence-induced eIF-5A is transcribed and the resulting co-expression of exogenous senescence-induced eIF-5A causes a down-regulation or inhibition of expression of endogenous senescence-induced eIF-5A.
Stress-Induced eIF-5A
[0222]The present invention relates to the discovery of stress-induced eIf-5A in various plants including alfalfa, tomato and lettuce. Down regulating the stress isoform of eIF-5A ameliorates stress-induced senescence. Down regulation of stress-isoform eIF-5A provides an increase tolerance to stress and results in enhanced growth. FIGS. 121-123 shows that transgenic plants (having a reduced expression of stress-induced eIF-5A) show an increased resistance to drought. FIG. 124 shows that transgenic plants (having a reduced expression of stress-induced eIF-5A) have an increased seed yield over control (wild-type) plants.
[0223]Thus, one embodiment of the present invention is isolated stress eIF-5A from alfalfa. The amino acid sequence is provided in FIG. 139 and is SEQ ID NO: 177. The polynucleotide encoding the amino acid is provided in FIG. 139 and is SEQ ID NO: 176.
[0224]Thus, one embodiment of the present invention is isolated stress eIF-5A from tomato. The amino acid sequence is provided in FIG. 146 and is SEQ ID NO: 191. The polynucleotide encoding the amino acid is provided in FIG. 146 and is SEQ ID NO: 190.
[0225]The present invention also provides isolated polynucleotides of stress eIF-5A that have 65-100% sequence homology to the above enumerated SEQ ID NOs, and hybridize under high stringency conditions to the complement of the enumerated SEQ ID NOs and which encode stress eIF-5A.
[0226]The present invention also provides stress eIF-5A isolated polypeptides of stress eIF-5A that have 65-100% amino acid sequence homology to the above enumerated SEQ ID NOs.
[0227]The present invention also provides antisense polynucleotides of the stress eIF-5As. The antisense polynucleotides may be of any length as long as they are able to inhibit expression. In some embodiments the antisense polynucleotides comprise the full length coding sequence and in other particularly preferred embodiments the antisense polynucleotides are directed at the 3'UTR since the different isoforms of eIF-5A have a higher degree of variation at the 3'UTR. In some embodiments the antisense polynucleotides are directed at the 5'-non-coding sequence. Antisense polynucleotides primarily complementary to 5'-non-coding sequences are known to be effective inhibitors of expression of genes encoding transcription factors. Branch, M. A., Molec. Cell Biol., 13:4284-4290 (1993).
[0228]The term "antisense polynucleotide of stress eIF5A" as used herein and in the claims encompasses not only those antisense polynucleotides that share 100% homology of the complement of an enumerated SEQ ID NO but also includes those antisense polynucleotides that are a functional variants. Functional variants are those variants, either natural or man made, that have at least 80% sequence homology to and hybridizes under high stringency conditions with the corresponding portion of the stress eIF-5A. Further the variant must have the function as intended by the present invention, that is it is capable of modulating expression of endogenous stress eIF-5A when introduced into an expression vector and wherein such vector is incorporated into the genome of at least one plant cell. One skilled in the art can appreciate that insubstantial changes can be made in the sequence that would not detrimentally effect the ability of the antisense polynucleotide to bind to the transcript and reduce or inhibition expression of the gene. Thus, the term "antisense polynucleotide" encompasses those polynucleotides that are substantially complementary to the transcript and that still maintain the ability to specifically bind to the transcript and inhibit or reduce gene expression. For a general discussion of antisense see Alberts, et al., Molecular Biology of the Cell, 2nd ed., Garland Publishing, Inc. New York, N.Y., 1989 (in particular pages 195-196, incorporated herein by reference).
[0229]One embodiment of the present invention provides expression vectors comprising either the stress eIF-5A polynucleotides (of the present invention as described above) or antisense polynucleotides of stress eIF-5A (of the present invention as described above). Vectors can be plasmids, preferably, or may be viral or other vectors known in the art to replicate and express genes encoded thereon in plant cells or bacterial cells. The vector becomes chromosomally integrated such that it can be transcribed to produce the desired antisense polynucleotide of stress eIF-5A RNA. Such plasmid or viral vectors can be constructed by recombinant DNA technology methods that are standard in the art. For example, the vector may be a plasmid vector containing a replication system functional in a prokaryotic host and an antisense polynucleotide according to the invention. Alternatively, the vector may be a plasmid containing a replication system functional in Agrobacterium and an antisense polynucleotide according to the invention. Plasmids that are capable of replicating in Agrobacterium are well known in the art. See, Mild, et al., Procedures for Introducing Foreign DNA Into Plants, Methods in Plant Molecular Biology and Biotechnology, Eds. B. R. Glick and J. E. Thompson. CRC Press (1993), PP. 67-83.
[0230]The vector further comprises regulatory sequences and promoter regulatory elements as discussed above.
[0231]The invention also provides a transgenic plant cell transformed with a vector or combination of vectors of the present invention comprising polynucleotides of stress eIF-5A in sense or antisense orientation, a transgenic plantlet or mature transgenic plant generated from such a cell, or a plant part, such as a flower, fruit, leaves, seeds, etc. of the transgenic plant.
[0232]The present invention also provides methods of inhibiting expression of endogenous stress eIF-5A. These methods comprise integrating into the genome of at least one cell of a plant, expression vectors of the present invention comprising antisense polynucleotides of stress eIF-5A. The antisense polynucleotides of stress eIF-5A are transcribed and inhibit expression of endogenous stress eIF-5A.
[0233]In another method of inhibiting expression of endogenous stress eIF-5A, an expression vector containing a stress eIF-5A polynucleotide of the present invention in a sense orientation is integrated into the genome of at least one cell of a plant. The polynucleotide of stress eIF-5A is transcribed and the resulting co-expression of exogenous stress eIF-5A causes a down-regulation or inhibition of expression of endogenous stress eIF-5A.
Wounding/Pathogen-Induced eIF-5A
[0234]Wounding/pathogen induced eIF-5A (also referred to as eIF-5A3) is expressed in wounded tissues and infected tissues. The present invention relates to the discovery of wounding/pathogen induced eIF-5A in various plants including Arabidopsis thaliana, tomato, alfalfa and lettuce. The present inventors have discovered that this isoform is upregulated during a wounding event or pathogen ingression of the plant. FIG. 125 indicates that wounding/pathogen induced eIF-5A is up-regulated after infection with P. syringae. The up-regulation occurs at the transcriptional level. Further, it is up-regulated exclusively at the protein level following virulent infection, which then gives rise to cell death, leading to the inference that wounding/pathogen induced eIF-5A is driving cell death in the event of ingression by pathogens.
[0235]Down regulation of the wounding/pathogen-induced isoform of eIF-5A inhibits pathogen-induced cell death. FIG. 126 shows that antisense suppression of wounding/pathogen-induced isoform of eIF-5A inhibits infection by virulent pathogens. Transgenic plants showed a greater than 99% increase in an inhibition of infection over control plants. FIGS. 127-130 show that transgenic canola (antisense/pathogen-induced isoform of eIF-5A) shows a resistance to infection by Sclerotinia sclerotiorum.
[0236]Wounding/pathogen induced eIF-5A regulates programmed cell death accompanying infection by necrotrophic bacterial and fungal pathogens. Within 72 h of infection with virulent Pseudomonas syringae pv Tomato DC 3000, wounding/pathogen induced eIF-5A protein is strongly up-regulated in the leaves of Arabidopsis wild-type plants. The up-regulation coincides with visible symptoms of disease progression. Moreover, wounding/pathogen induced eIF-5A is the only eIF5A isoform to show increased expression following infection. In addition, the up-regulation appears to be post-transcriptionally regulated. To further test the role of wounding/pathogen induced eIF-5A in disease development, transgenic Arabidopsis plants with constitutively suppressed wounding/pathogen induced eIF-5A expression were developed using antisense T-DNA insertions. The suppressed plants showed marked resistance to virulent Pseudomonas syringae pv Tomato DC 3000, with some lines exhibiting a 99% decrease in bacterial load, relative to wild-type plants. This suppression of pathogen growth correlated with the absence of disease symptoms. The same lines also showed strong resistance to infection by the necrotrophic fungal pathogen, Sclerotinia sclerotiorum. The results have been interpreted as indicating that wounding/pathogen induced eIF-5A regulates programmed cell death induced by necrotrophic pathogens, and that suppression of wounding/pathogen induced eIF-5A inhibits host cell death upon infection and the ensuing release of nutrients that normally support growth of the pathogen.
[0237]FIG. 9 shows that senescence-induced eIF-5A remains constant in the control plant, the mock treated plant, the Avr treated plant and the Vir treated plant (it is detected as the plants were 4 weeks old). But wounding/pathogen induced eIF-5A is up-regulated in the Vir treated plant.
[0238]FIG. 10 shows the results of an experiment where leaves of a plant were wounded with a hemostat. Levels of senescence-induced eIF-5A, wounding/pathogen induced eIF-5A and growth eIF-5A in arabidopsis thaliana ("At") were measured immediately after the wounding, 1 hour, and 9 hours after the wounding. The Northern Blots show that senescence-induced eIF-5A remained constant, but there was a noticeable increase in the levels expression of the wounding/pathogen induced eIF-5A. The levels of expression of the growth eIF-5A began to decrease in the event of wounding.
[0239]The present inventors have demonstrated that when wounding/pathogen induced eIF-5A is up-regulated and a wounding event is imposed upon the plants (such as occurs when the seedlings are transplanted), this wounding results in a very strong suppression of growth eIF-5A. See FIGS. 14-17. The resulting plants have very stunted growth. But when the seeds are soaked in kanomycin and are planted directly into the soil (no need to transplant and thus no transplant wounding), the seeds develop into normal sized plants.
[0240]The differences seen between the various test plants all having a sense wounding/pathogen induced eIF-5A construct (FIG. 15) incorporated is due to varying degrees of expression of the wounding/pathogen induced eIF-5A. One skilled in the art will appreciate that when a gene is introduced (either sense or antisense) one gets varying degrees of either gene up-regulation or down-regulation. The degree of differences depends on where the gene gets incorporated and how many copies get incorporated. By having varying degrees of expression, one can correlate the various phenotypes to the gene expression. Once the desired phenotype is produced, that plant can be picked and used to create the desired progeny. Thus in FIG. 15, the plants that were strongly up-regulated for wounding/pathogen induced eIF-5A barely grew after the wounding event (plant tag 10), but the plants that grew a little better (but not as good as wild type) (plant tag 4) were not as strongly up-regulated.
[0241]One embodiment of the present invention is isolated wounding/pathogen induced eIF-5A from Arabidopsis thaliana. The amino acid sequence is provided in FIG. 41 and is SEQ ID NO: 55. The polynucleotide encoding the amino acid is provided in FIG. 41 and is SEQ ID NO: 54.
[0242]Another embodiment of the present invention is isolated wounding/pathogen induced eIF-5A from tomato. The amino acid sequence is provided in FIG. 103 and is SEQ ID NO: 57. The polynucleotide encoding the amino acid is provided in FIG. 103 and is SEQ ID NO: 56.
[0243]Another embodiment of the present invention is isolated wounding/pathogen induced eIF-5A from alfalfa. The amino acid sequence is provided in FIG. 137 and is SEQ ID NO: 173. The polynucleotide encoding the amino acid is provided in FIG. 137 and is SEQ ID NO: 172.
[0244]The present invention also provides isolated polynucleotides of wounding/pathogen induced eIF-5A that have 65-100% sequence homology to the above enumerated SEQ ID NOs, and hybridize under high stringency conditions to the complement of the enumerated SEQ ID NOs and which encode wounding/pathogen induced eIF-5A.
[0245]The present invention also provides wounding/pathogen induced eIF-5A isolated polypeptides of wounding/pathogen induced eIF-5A that have 65-100% amino acid sequence homology to the above enumerated SEQ ID NOs.
[0246]The present invention also provides antisense polynucleotides of the wounding/pathogen induced eIF-5As. The antisense polynucleotides may be of any length as long as they are able to inhibit expression. In some embodiments the antisense polynucleotides comprise the full length coding sequence and in other particularly preferred embodiments the antisense polynucleotides are directed at the 3'UTR since the different isoforms of eIF-5A have a higher degree of variation at the 3'UTR. In some embodiments the antisense polynucleotides are directed at the 5'-non-coding sequence. Antisense polynucleotides primarily complementary to 5'-non-coding sequences are known to be effective inhibitors of expression of genes encoding transcription factors. Branch, M. A., Molec. Cell Biol., 13:4284-4290 (1993).
[0247]The term "antisense polynucleotide of wounding/pathogen induced eIF5A" as used herein and in the claims encompasses not only those antisense polynucleotides that share 100% homology of the complement of an enumerated SEQ ID NO but also includes those antisense polynucleotides that are a functional variants. Functional variants are those variants, either natural or man made, that have at least 80% sequence homology to and hybridizes under high stringency conditions with the corresponding portion of the wounding/pathogen induced eIF-5A. Further the variant must have the function as intended by the present invention, that is it is capable of modulating expression of endogenous wounding/pathogen induced eIF-5A when introduced into an expression vector and wherein such vector is incorporated into the genome of at least one plant cell. One skilled in the art can appreciate that insubstantial changes can be made in the sequence that would not detrimentally effect the ability of the antisense polynucleotide to bind to the transcript and reduce or inhibition expression of the gene. Thus, the term "antisense polynucleotide" encompasses those polynucleotides that are substantially complementary to the transcript and that still maintain the ability to specifically bind to the transcript and inhibit or reduce gene expression.
[0248]One embodiment of the present invention provides expression vectors comprising either the wounding/pathogen induced eIF-5A polynucleotides (of the present invention as described above) or antisense polynucleotides of wounding/pathogen induced eIF-5A (of the present invention as described above). Vectors, regulatory sequences and promoter regulatory elements are as discussed above.
[0249]The invention also provides a transgenic plant cell transformed with a vector or combination of vectors of the present invention comprising polynucleotides of wounding/pathogen induced eIF-5A in sense or antisense orientation, a transgenic plantlet or mature transgenic plant generated from such a cell, or a plant part, such as a flower, fruit, leaves, seeds, etc. of the transgenic plant.
[0250]The present invention also provides methods of inhibiting expression of endogenous wounding/pathogen induced eIF-5A. These methods comprise integrating into the genome of at least one cell of a plant, expression vectors of the present invention comprising antisense polynucleotides of wounding/pathogen induced eIF-5A. The antisense polynucleotides of wounding/pathogen induced eIF-5A are transcribed and inhibit expression of endogenous wounding/pathogen induced eIF-5A.
[0251]In another method of inhibiting expression of endogenous wounding/pathogen induced eIF-5A, an expression vector containing a wounding/pathogen induced eIF-5A polynucleotide of the present invention in a sense orientation is integrated into the genome of at least one cell of a plant. The polynucleotide of wounding/pathogen induced eIF-5A is transcribed and the resulting co-expression of exogenous wounding/pathogen induced eIF-5A causes a down-regulation or inhibition of expression of endogenous wounding/pathogen induced eIF-5A.
[0252]By inhibiting expression of endogenous wounding/pathogen induced eIF-5A, resulting transgenic plants have an increased resistance to virulent damage arising from pathogen ingression. Pathogens include, but are not limited to Black Sigatoka in bananas, Sclerotinia sclerotiorum (a major fungal disease of lettuce and turf grass) Pythium (a major fungal disease in green house vegetable crops) and Pseudomonas syringae. See example 16 and FIGS. 43 and 44.
Growth eIF-5A
[0253]The present invention also relates to growth eIF-5A. Growth eIF-5A is expressed in growing tissues. When eIF-5A is up-regulated with polynucleotides of growth eIF-5A in sense orientation, three phenotypic changes are noticed: increased seed size, increased biomass, and increased seed yield. See FIGS. 132-134.
[0254]One embodiment of the present invention is isolated growth eIF-5A from Arabidopsis thaliana. The amino acid sequence is provided in FIG. 1 and is SEQ ID NO: 60. The polynucleotide encoding the amino acid is provided in FIG. 2 and is SEQ ID NO: 63.
[0255]Another embodiment of the present invention is isolated growth eIF-5A from tomato. The amino acid sequence is provided in FIG. 101 and is SEQ ID NO: 65. The polynucleotide encoding the amino acid is provided in FIG. 101 and is SEQ ID NO: 64.
[0256]Another embodiment of the present invention is isolated growth eIF-5A from canola. The amino acid sequence is provided in FIG. 95 and is SEQ ID NO: 67. The polynucleotide encoding the amino acid is provided in FIG. 95 and is SEQ ID NO: 66.
[0257]Another embodiment of the present invention is isolated growth eIF-5A from alfalfa. The amino acid sequence is provided in FIG. 140 and is SEQ ID NO: 179. The polynucleotide encoding the amino acid is provided in FIG. 140 and is SEQ ID NO: 178.
[0258]The present invention also provides isolated polynucleotides of growth eIF-5A that have 65-100% sequence homology to the above enumerated SEQ ID NOs, and hybridize under high stringency conditions to the complement of the enumerated SEQ ID NOs and which encode growth eIF-5A.
[0259]The present invention also provides isolated polypeptides of growth eIF-5A that have 65-100% amino acid sequence homology to the above enumerated SEQ ID NOs.
[0260]The present invention also provides antisense polynucleotides of the growth eIF-5As. The antisense polynucleotides may be of any length as long as they are able to inhibit expression. In some embodiments the antisense polynucleotides comprise the full length coding sequence and in other particularly preferred embodiments the antisense polynucleotides are directed at the 3'UTR since the different isoforms of eIF-5A have a higher degree of variation at the 3'UTR. In some embodiments the antisense polynucleotides are directed at the 5'-non-coding sequence. Antisense polynucleotides primarily complementary to 5'-non-coding sequences are known to be effective inhibitors of expression of genes encoding transcription factors. Branch, M. A., Molec. Cell Biol., 13:4284-4290 (1993).
[0261]The term "antisense polynucleotide of growth eIF5A" as used herein and in the claims encompasses not only those antisense polynucleotides that share 100% homology of the complement of an enumerated SEQ ID NO but also includes those antisense polynucleotides that are a functional variants. Functional variants are those variants, either natural or man made, that have at least 80% sequence homology to and hybridizes under high stringency conditions with the corresponding portion of the growth eIF-5A. Further the variant must have the function as intended by the present invention, that is it is capable of modulating expression of endogenous growth eIF-5A when introduced into an expression vector and wherein such vector is incorporated into the genome of at least one plant cell. One skilled in the art can appreciate that insubstantial changes can be made in the sequence that would not detrimentally effect the ability of the antisense polynucleotide to bind to the transcript and reduce or inhibition expression of the gene. Thus, the term "antisense polynucleotide" encompasses those polynucleotides that are substantially complementary to the transcript and that still maintain the ability to specifically bind to the transcript and inhibit or reduce gene expression.
[0262]One embodiment of the present invention provides expression vectors comprising either the growth eIF-5A polynucleotides (of the present invention as described above) or antisense polynucleotides of growth eIF-5A (of the present invention as described above). Vectors, regulatory sequences and promoter regulatory elements are as discussed above.
[0263]The invention also provides a transgenic plant cell transformed with a vector or combination of vectors of the present invention comprising polynucleotides of growth eIF-5A in sense or antisense orientation, a transgenic plantlet or mature transgenic plant generated from such a cell, or a plant part, such as a flower, fruit, leaves, seeds, etc. of the transgenic plant.
[0264]The present invention also provides methods of inhibiting expression of endogenous growth eIF-5A. These methods comprise integrating into the genome of at least one cell of a plant, expression vectors of the present invention comprising antisense polynucleotides of growth eIF-5A. The antisense polynucleotides of growth eIF-5A are transcribed and inhibit expression of endogenous growth eIF-5A.
[0265]In another method of inhibiting expression of endogenous growth eIF-5A, an expression vector containing a growth eIF-5A polynucleotide of the present invention in a sense orientation is integrated into the genome of at least one cell of a plant. The polynucleotide of growth eIF-5A is transcribed and the resulting co-expression of exogenous growth eIF-5A causes a down-regulation or inhibition of expression of endogenous growth eIF-5A.
[0266]In another embodiment of the present invention there is provided a method of up-regulating expression of growth eIF-5A. An expression vector containing a growth eIF-5A polynucleotide of the present invention in a sense orientation is integrated into the genome of at least one cell of a plant. The polynucleotide of growth eIF-5A is transcribed and the resulting co-expression of exogenous growth eIF-5A causes the cells to express more growth eIF-5A than non-transgenic cells.
[0267]FIG. 19 shows that plants that were up-regulated for growth eIF-5A had an increased biomass over that of the control plants. Growth eIF-5A was inserted into Arabidopsis thaliana plants in a sense orientation to up-regulate the expression of growth eIF-5A. Sixteen mother lines (1-16) were assayed to determine the general level of growth eIF-5A expression. From each mother line, 8 sister lines were produced (A-H). The level of expression of growth eIF-5A in each mother line was tested and the results shown in FIG. 20. Various degrees of expression are noticed throughout the mother lines. For example, lines 2 and 10 have very high levels of expression whereas lines 11 and 16 have very low or no expression.
[0268]FIGS. 21 and 22 show the plants from lines 1 and 2. These plants are bigger than the control plants. Because the growth eIF-5A is a cell-division isoform and because it is constitutively expressed, there is increased cell division. A reduction in senescence occurs because the plant is locked into a growth mode and can not make the switch to the senescence pathway.
[0269]FIGS. 23 and 24 are from lines that had medium level of expression of growth eIF-5A. They appear to have bigger leaves and delayed senescence.
[0270]FIGS. 25 and 26 are from lines that had low levels of up-regulation. They have large leaves and large rosettes.
[0271]FIGS. 27 and 28 are from lines that have no up-regulation (which may be due to co-suppression of the gene). Since the plant is kanomycin resistant, the gene must be present in order for the plants to grow on the media. It appears that the senescence-induced eIF-5A is also co-suppressed as well thus giving rise to an increase in size.
[0272]In addition to increased biomass, there is also increased seed size in plants having growth eIF-5A up-regulated. The seed size of all of the lines was measured. In the lines having the highest levels of growth eIF-5A expression, a greater than 3× increase in seed size is seen. This occurs because up-regulation of growth eIF-5A, increases cell division and thus increases seed size.
[0273]The growth eIF-5A (from Arabidopsis thaliana) in the above examples was being constitutively expressed, i.e. is being expressed everywhere in the plant through the use of a universal promoter. In contrast, by using a tissue specific promoter, one may direct the up-regulation in particular tissues. For example, by using a seed specific promoter, the growth eIF-5A would only be up-regulated in the seed, allowing the leaves to grow normally, but produce an increase in the amount of seeds. Thus, using a specific promoter, the growth eIF-5A can be up-regulated in the desired plant part to get a desired phenotype.
[0274]By up-regulating growth eIF-5A, at least three phenotypes result--increased biomass, increased seed yield, or increased seed size, but not all three phenotypes are present at the same time (or in the same plant). For example, if a plant exhibits an increase in seed size, a smaller plant will be present. In the plant lines that had the highest up-regulation of growth eIF-5A, the biggest seeds were produced, but the plants were smaller because there was massive cell division going on throughout the whole plant, which was at the expense of cell enlargement (needed for bigger leaves). At lower levels of up-regulation of expression of growth AteIF-5A, one sees an impact on the leaves (bigger) without impacting the seed. Thus, one may use tissue specific expression and pick the phenotype desired. For example, one may place growth eIF-5A under a xylem specific promoter to achieve an increase in the amount of xylem produced. Thus, any desired promoter may be used to achieve the desired tissue-specific up-regulation.
[0275]Growth eIF-5A as well as the other isoforms of eIF-5A are highly conserved across plant species. FIG. 85 shows an amino acid alignment of various eIF-5A isoforms from several plant species indicating that the hypusine region is very highly conserved. In fact, eIF-5A is so highly conserved that an isolated polynucleotide encoding one plant growth eIF-5A can be inserted into another plant species in sense orientation and achieve an up-regulation of growth eIF-5A. FIG. 162 shows the effect of introducing cottonwood growth eIF-5A sense orientation into arabidopsis. FIG. 162 shows T1 plants at 7 weeks old. The larger plants are from two different transgenic lines. The smaller plants are the controls. FIG. 163 shows T2 plants from three separate lines (labeled lines 2, 3 and 4) and the plants labeled "BIN" is the control empty vector. The plants are 28 days old. The transgenic arabidopsis have an increased growth rate as compared to the control plants. FIG. 164 show T2 plants (Line 3) at 32 days old compared to control plants.
[0276]Further evidence of the interchangeability of growth eIF-5A, is shown in FIG. 165. Growth factor eIF-5A was up-regulated in canola by with either canola growth factor eIF-5A or arabidopsis growth eIF-5A (full length) in the sense orientation under the control of a 35S promoter. The resulting transgenic plants had an increase in seed yield over the wild-type control plants. The increase in seed yield in the transgenic canola plant having an arabidopsis growth eIF-5A in the sense orientation and the transgenic canola plants having an exogenous canola growth eIF-5A in sense orientation is very similar.
DHS
[0277]DHS is necessary for the activation of eIF-5A and is expressed in senescing tissues. The present invention provides isolated DHS from Arabidopsis thaliana, tomato, carnation, canola, lettuce, alfalfa, banana, cottonwood, yeast, petunia and mycosphaerella.
[0278]Thus one embodiment of the present invention is isolated DHS from Arabidopsis thaliana. The amino acid sequence is provided in FIG. 46 and is SEQ ID NO: 6. The polynucleotide encoding the amino acid is provided in FIG. 46 and is SEQ ID NO: 5.
[0279]Another embodiment of the present invention is isolated DHS from tomato. The amino acid sequence is provided in FIGS. 45 A and B and is SEQ ID NO: 2. The polynucleotide encoding the amino acid is provided in FIGS. 45 A and B and is SEQ ID NO: 1.
[0280]Another embodiment of the present invention is isolated DHS from carnation. The amino acid sequence is provided in FIG. 54 and is SEQ ID NO: 10. The polynucleotide encoding the amino acid is provided in FIG. 54 and is SEQ ID NO: 9.
[0281]Another embodiment of the present invention is isolated DHS from canola. The amino acid sequence is provided in FIG. 97 and is SEQ ID NO: 71. The polynucleotide encoding the amino acid is provided in FIG. 97 and is SEQ ID NO: 70.
[0282]Another embodiment of the present invention is isolated DHS from lettuce. FIG. 105 provides a portion of lettuce DHS polynucleotide sequence.
[0283]Another embodiment of the present invention is isolated DHS from alfalfa. The amino acid sequence is provided in FIGS. 107 A and B and is SEQ ID NO: 73. The polynucleotide encoding the amino acid is provided in FIGS. 107 A and B and is SEQ ID NO: 72.
[0284]Another embodiment of the present invention is isolated DHS from banana. The amino acid sequence is provided in FIGS. 108 A and B and is SEQ ID NO: 75. The polynucleotide encoding the amino acid is provided in FIGS. 108 A and B and is SEQ ID NO: 74.
[0285]Another embodiment of the present invention is isolated DHS from cottonwood. The amino acid sequence is provided in FIGS. 109 A and B and is SEQ ID NO: 77. The polynucleotide encoding the amino acid is provided in FIGS. 109 A and B and is SEQ ID NO: 76.
[0286]Another embodiment of the present invention is isolated DHS from mycosphaerella. The sequence is provided at FIG. 110.
[0287]The present invention also provides isolated polynucleotides of DHS that have 65-100% sequence homology to the above enumerated SEQ ID NOs, and hybridize under high stringency conditions to the complement of the enumerated SEQ ID NOs and which encode DHS.
[0288]The present invention also provides isolated polypeptides of DHS that have 65-100% amino acid sequence homology to the above enumerated SEQ ID NOs. FIG. 47 and FIGS. 148 (a) and (b) provide an amino acid alignment of the various DHS showing the highly conserved nature of DHS across species.
[0289]The present invention also provides antisense polynucleotides of DHS. The antisense polynucleotides may comprise the full length coding sequence, or may be directed at the 3'UTR, or may be directed at the 5'-non-coding sequence. Antisense polynucleotides primarily complementary to 5'-non-coding sequences are known to be effective inhibitors of expression of genes encoding transcription factors. Branch, M. A., Molec. Cell Biol., 13:4284-4290 (1993). The antisense polynucleotides may be of any length as long as they are able to inhibit expression.
[0290]The term "antisense polynucleotide of DHS" as used herein and in the claims encompasses not only those antisense polynucleotides that share 100% homology of the complement of an enumerated SEQ ID NO but also includes those antisense polynucleotides that are a functional variants. Functional variants are as described above. The variant functions as intended by the present invention, are capable of modulating expression of endogenous DHS when introduced into an expression vector and wherein such vector is incorporated into the genome of at least one plant cell.
[0291]The present invention also provides sense polynucleotides of DHS. The sense polynucleotides may comprise the full length coding sequence, or may be directed at the 3'UTR, or may be directed at the 5'-non-coding sequence. The sense polynucleotides may be of any length as long as they are able to either down-regulate expression through co-suppression, or alternatively, up-regulate expression.
[0292]One embodiment of the present invention provides expression vectors comprising either DHS polynucleotides (of the present invention as described above) or antisense polynucleotides of DHS (of the present invention as described above) or sense polynucleotides of DHS (of the present invention as described above). Vectors are as described above.
[0293]The invention also provides a transgenic plant cell transformed with a vector or combination of vectors of the present invention comprising a polynucleotide of DHS either in the sense or antisense orientation, a transgenic plantlet or mature transgenic plant generated from such a cell, or a plant part, such as a flower, fruit, leaves, seeds, etc. of the transgenic plant.
[0294]The present invention also provides methods of inhibiting/decreasing expression of endogenous DHS. These methods comprise integrating into the genome of at least one cell of a plant, expression vectors of the present invention comprising antisense polynucleotides of DHS. The antisense polynucleotides of DHS are transcribed and inhibit/decrease expression of endogenous DHS. The resulting plants have an increased biomass, increased seed yield, and longer shelf life.
[0295]For example, FIGS. 65-68 show pictures of wild type arabidopsis plants compared to the arabidopsis plants having a down-regulation of DHS brought about by the use of antisense constructs. The arabidopsis plants having DHS down-regulated with antisense arabidopsis DHS constructs have an increased biomass as compared to wild type arabidopsis (not having a down-regulation of DHS). See FIGS. 153 and 154. Further, FIGS. 69 and 155 illustrates that the down-regulated DHS arabidopsis plants have an increased seed yield over the wild type plants. FIG. 156 shows that down regulation of DHS causes an increase in root development. See Example 13 of a description of various constructs used to transform arabidopsis with either full length or the 3' region of arabidopsis DHS (SEQ ID NO:30) in the antisense orientation.
[0296]Example 14 and 15 describe the transformation of tomato plants with full-length or 3' region of tomato DHS in the antisense orientation. FIGS. 70-79 show that the resulting tomato plants have a delayed softening and spoilage of tomato as compared to wild type tomato plants. FIGS. 84(a) and 84(b) are photographs of tomato fruits showing that tomatoes having been transformed with tomato DHS in the antisense orientation have a reduced incidence of blossom end rot, thus indicating that reducing the expression of DHS prevents the onset of tissue and cell death arising from physiological disease.
[0297]Suppression of DHS expression in canola increases plant biomass and increases seed yield. See example 28 and FIGS. 157-161. FIG. 157 shows the transgenic canola has increase biomass as demonstrated by increased leaf mass. FIG. 158 shows the results of a Western blot showing partial down-regulation of DHS in one of the transgenic lines of canola. FIG. 159 shows that down-regulation of DHS in canola results in larger siliques. FIG. 160 shows that down-regulation of DHS in canola results in an increased seed yield. FIG. 161 (a) illustrates that total yield of triacylglyercol expressed on a "per weight of seed" basis. This demonstrates that there is no change in the quantity of oil per seed. Thus, the ˜65% increase in seed yield translates into and ˜65% increase in seed oil. FIG. 161 (b) illustrates that there is no change in the fatty acid composition of oil in the DHS down regulated canola plants.
[0298]FIG. 99 shows the result of an experiment where canola plants were grown in small pots, which puts a small amount of stress on the plants (sub-lethal). When DHS is down-regulated, an increase in seed yield can be observed. The 3'UTR of canola DHS was used as an antisense nucleotide. The antisense oligonucleotide was expressed constitutively using a 35S promoter. Using a constitutive promoter, the DHS antisense polynucleotide is expressed throughout the growth and development of the entire plant. When the plant is exposed to episodes of sub-lethal stress (such as being grown in too small of a pot, or periods of suboptimal growing conditions, etc.), the plant may enter the senescence pathway. However, when the plant is a transgenic plant having a reduced expression of DHS, the plants do not enter into the senescence mode when subjected to sub-lethal stress. FIG. 99 shows an increase in canola yield in plants having DHS down-regulated. FIG. 99 shows T1 through T4 generations, and two particular T4 generation of a particular plant line (designated as the "P" and "T" line).
[0299]Down regulating DHS in carnations causes the flowers to have an extended vase life. Example 29 shows that down regulation brought about the use of antisense constructs, as well as the use of inhibitors of the DHS reaction extend the shelf life of flowers by up to 83%.
[0300]Transgenic bananas having antisense DHS exhibit increased biomass as evidenced by larger plants as compared to wild type. See FIG. 131. The fruit from the transgenic plant also show delayed spoilage as compared to the wild type. See FIG. 114. A nucleotide encoding banana DHS was inserted into a vector in an antisense orientation either under the control of 35S promoter or a fruit specific promoter (pPDK). Either the full length banana DHS or the 3' UTR was used.
[0301]Commercially-available pre-packaged salad is commonly stored under conditions of controlled atmosphere, whereby the level of oxygen is greatly reduced below its atmospheric concentration in order to extend the shelf life of the product. The most common symptom of spoiled pre-packaged salad is browning on the cut surfaces of lettuce. Although controlled atmosphere packaging does achieve a delay in browning, it can also result in off-odor and off-flavor. Down-regulation of DHS is shown to have potential as an alternative strategy for delaying browning on the cut surfaces of lettuce. DHS catalyzes the activation of eukaryotic translation initiation factor 5A (eIF5A), which acts as a nucleocytoplasmic shuttle protein for select populations of mRNAs. DHS appears to play a role in browning of cut lettuce inasmuch as suppression of DHS expression (by antisense technology) resulted in a significant delay in the onset of browning under atmospheric conditions. Specifically, 80% of the cut segments of wild-type lettuce plants showed browning at 6 days after cutting, whereas only 27%, on average, of the cut segments of transgenic plants from 5 segregating lines turned brown over the same period, with some individual plants showing 0% browning. Suppression of DHS expression in lettuce through the use of antisense constructs results in a delay in the onset of browning of cut lettuce as compared to wild type lettuce. See Example 27 and FIGS. 149-151.
[0302]Jatropha curcas L. is a plant known for its high oil content and may possible be a source of bio-fuel. The clear oil expressed from the seed has been used for illumination and lubricating, and more recently has been suggested for energetic purposes, one ton of nuts can yield 70 kg refined petroleum, 40 kg "gasoil leger" (light fuel oil), 40 kg regular fuel oil, 34 kg dry tar/pitch/rosin, 270 kg coke-like char, and 200 kg ammoniacal water, natural gas, creosote, etc. Thus, the ability to enhance the seed yield (and thus oil content) may prove valuable. Increasing the seed yield can be achieved through three strategies. The first strategy involves constitutively up-regulating growth eIF-5A. Preferably this is done with a heterologous isoform of growth eIF-5A (i.e. using a canola or arabidopsis growth eIF-5A) to minimize the prospect of co-suppression. This has a dual effect--there is up-regulation of hypusinated growth eIF-5A, and there is also an up-regulation of unhypusinated senescence eIF-5A because there is more eIF-5A being produced than there is DHS available to activate (hypusinate) it. Any plant growth eIF-5A may be used, for example, a tomato or cottonwood growth eIF-5A could be used in Jatropha. The second strategy involves constitutively up-regulating mutated senescence eIF-5A in which the conserved lysine that normally gets hypusinated is converted to alanine. This mutated eIF-5A can not be hypusinated (activated), and in its unhypusinated form promotes growth. See FIGS. 166 and 167 showing transgenic arabidopsis plants having a mutated senescence eIF-5A are larger than wild type arabidopsis plants. Larger plants have mor bolts and more seeds. Also, unhypusinated senescence eIF-5A confer upon the plants enhanced nitrogen utilization efficiency, thus enabling them to out perform control plants under conditions of low nitrogen in the soil. The third strategy involves constitutively down-regulating DHS. This must be a partial down-regulation of DHS because the plant needs some DHS to function properly. But as seen above, partial down-regulation of DHS substantially promotes growth and increases seed yield, presumably because of an accumulation of unhypusinated senescence eIF-5A.
[0303]FIGS. 47 and 148 (a) and (b) show an amino acid alignment of various DHS, revealing that DHS is highly conserved. FIG. 148(a) and (b) indicate the very highly conserved lysine of DHS as well as other regions. In fact, DHS is so highly conserved that an isolated polynucleotide encoding one plant DHS can be inserted into another plant species in antisense orientation and achieve down-regulation of DHS.
[0304]FIG. 100 shows a bar graph measuring canola yield (made by enhancing the capacity for plant growth. This figure shows that can increase the yield both by increasing growth eIF-5A expression (use of sense constructs) or by increasing expression of DHS (use of sense constructs). The second bar labeled "canola" shows the increase achieved when upregulated canola DHS by using a canola DHS in the sense orientation. The fourth bar shows an increase in canola yield by using a tomato DHS in sense orientation in a canola plant.
[0305]In another method of inhibiting expression of endogenous DHS, an expression vector containing a DHS polynucleotide of the present invention in a sense orientation is integrated into the genome of at least one cell of a plant. The polynucleotide of DHS is transcribed and the resulting co-expression of exogenous DHS causes a down-regulation or inhibition of expression of endogenous DHS.
[0306]By inhibiting expression of endogenous DHS, resulting transgenic plants have no or substantially less DHS protein to activate eIF-5A. As discussed earlier, eIF-5A must be activated to render it biologically useful. Thus, by inhibiting or reducing the expression of DHS either by antisense polynucleotides or by co-suppression with sense polynucleotides, the resulting transgenic plants will either have no active eIF-5A or reduced active eIF-5A. These transgenic plants will exhibit an increase in biomass of the plant, increased seed yield and/or increased seed size. Transgenic plants having antisense polynucleotides of DHS show an increase in photosynthesis and also have an increased starch content. See Examples 24 and 25.
[0307]The present invention also provides for up-regulation of DHS. By providing a sense DHS nucleotide into a plant to cause up-regulation, the resulting plant has an increased amount of DHS. The resulting transgenic plants have an increased biomass and seed yield.
[0308]Further evidence to support the contention that DHS and eIF-5A play regulatory roles in senescence was provided by treating carnation flowers with inhibitors that are specific for DHS. Spermidine and eIF-5A are the substrates of DHS reaction (Park et al., 1993; Park et al., 1997). Several mono-, di-, and polyamines that have structural features similar to spermidine inhibit DHS activity in vitro (Jakus et al., 1993). Some polyamines, such as spermidine, putrescine, and spermine, have been generally used to extend carnation vase life (Wang and Baker, 1980). Through treatment with different polyamines at different concentrations Wang et al (unpublished b) were able to extend the vase life of carnation flowers by 2 fold. Further studies employing a transient infection system to down-regulate DHS is in progress. Preliminary data indicates that the percent survival rate is almost 4 fold higher at day 8 in cut carnations that were vacuum infiltrated with a transient infection system expressing antisense DHS than untreated flowers (Wang et al., unpublished b).
[0309]A further major loss in agriculture besides the loss of growth due to stress is post harvest stress-induced senescence (McCabe et al., 2001). This is especially true for plants that are partially processed such as cut lettuce. A symptom of cutting lettuce is browning which is a result of phenolics production (Matile et al., 1999). A field trial of lettuce with antisense polynucleotides of lettuce eIF-5A (LeIF-5A) or antisense full length DHS demonstrated that the transgenic lettuce was significantly more resistant to browning after cutting than the control lettuce. See FIGS. 149-151. It appears that even though stress induced senescence due to harvesting has distinct circuitry (Page et al., 2001), the translational control upstream of browning and likely other senescence symptoms is regulated at least in part by DHS and eIF-5A. Downstream of the regulation of senescence are the execution genes. These are the effectors of senescence and cause the metabolic changes that bring on the senescence syndrome. It appears that eIF-5A and DHS when down-regulated are capable of dampening down a whole range of symptoms caused by senescence.
[0310]Example 1 describes the isolation and mRNA encoding DHS in various plants. Example 2 describes the induction of senescence-induced DHS in tomatoes by treating tomato leaves with sorbitol. FIGS. 52 and 62 show that DHS expression is increased when leaves are treated with sorbitol. Example 3 and FIGS. 50 and 63 show that DHS is up-regulated in senescing flowers. Example 4 and FIGS. 51 and 61 show that DHS is up-regulated in ripening fruit.
[0311]The present invention also relates to antibodies that recognize the three isoforms of eIF-5A (senescence-induced factor eIF-5A); (wounding factor eiF-5A) and (growth factor eIF-5A).
[0312]The present invention also provides a method of identifying senescence-induced eIF-5A, wounding/pathogen induced eIF-5A, growth eIF-5A and DHS in other plants and fungi. By using the methods described herein and the sequences provided, probes are designed to isolate/identify the desired isoforms or DHS. Since the isoforms of eIF-5A (senescence-induced eIF-5A, wounding/pathogen induced eIF-5A, and growth eIF-5A) are often highly homologous in the coding region (see FIG. 2), to ensure identification and even alter amplification of the desired isoform, probes or primers are preferably designed from the beginning of the 5'UTR and at the end of the 3'' UTR. (See FIGS. 3, 4 and 5). A preferred set of primers for amplification of wounding/pathogen induced eIF-5A or probes for identification of wounding/pathogen induced eIF-5A are as follows. The downstream primer is 5' GAG CTC AAG AAT AAC ATC TCA TAA GAAAC3' (SEQ ID NO: 33) The upstream primer is 5' CTC GAG TGC TCA CTT CTC TCT CTT AGG 3' (SEQ ID NO: 34).
[0313]Before isolating wounding/pathogen induced eIF5A from a plant or plant part, it is best to introduce a wounding event to allow the plant to begin expressing wounding/pathogen induced eIF-5A. Any wounding event is acceptable and one such exemplary wound events included crushing the leaves at the central vein. Similarly, before isolating senescence-induced eIF-5A, it best to stress the plant tissue to induce senescence.
[0314]Having now generally described the invention, the same will be more readily understood through reference to the following examples, which are provided by way of illustration, and are not intended to be limiting to the present invention.
EXAMPLES
Example 1
Messenger RNA (mRNA) Isolation
[0315]Total RNA was isolated from tomato flowers and tomato fruit at various developmental stages and from leaves (untreated or after chilling or sorbitol treatment). The tissue (5 g) was briefly ground in liquid nitrogen. The ground powder was mixed with 30 ml guanidinium buffer (4 M guanidinium isothiocyanate, 2.5 mM NaOAc pH 8.5, 0.8%-mercaptoethanol). The mixture was filtered through four layers of cheesecloth and centrifuged at 10,000×g at 4° C. for 30 minutes. The supernatant was then subjected to cesium chloride density gradient centrifugation at 26,000×g for 20 hours. The pelleted RNA was rinsed with 75% ethanol, resuspended in 600 μl DEPC-treated water and the RNA precipitated at -70° C. with 0.75 ml 95% ethanol and 30 μl of 3M NaOAc. Ten μg of RNA were fractionated on a 1.2% denaturing formaldehyde agarose gel and transferred to a nylon membrane. Randomly primed 32P-dCTP-labeled full length DHS cDNA (SEQ ID NO:1) was used to probe the membrane at 42° C. overnight. The membrane was then washed once in 1×SSC containing 0.1% SDS at room temperature for 15 minutes and three times in 0.2×SSC containing 0.1% SDS at 65° C. for 15 minutes each. The membrane was exposed to x-ray film overnight at -70° C.
[0316]PolyA.sup.+ mRNA was isolated from total RNA using the PolyA.sup.+ tract mRNA Isolation System available from Promega. PolyA.sup.+ mRNA was used as a template for cDNA synthesis using the ZAP Express® cDNA synthesis system available from Stratagene (La Jolla, Calif.)
[0317]Tomato Leaf cDNA Library Screening
[0318]A cDNA library made using mRNA isolated from Match F1 hybrid tomato leaves that had been exposed to 2 M sorbitol for six hours was diluted to approximately 5×106 PFU/ml. The cDNA library was screened using a 32P-labeled 600 by RT-PCR fragment. Three positive cDNA clones were excised and recircularized into a pBK-CMV® (Stratagene) phagemid using the method in the manufacturer's instructions. The full length cDNA was inserted into the pBK-CMV vector.
[0319]Plasmid DNA Isolation, DNA Sequencing
[0320]The alkaline lysis method described by Sambrook et al., (Supra) was used to isolate plasmid DNA. The full length positive cDNA clone was sequenced using the dideoxy sequencing method. Sanger, et al., Proc. Natl. Acad. Sci. USA, 74:5463-5467. The open reading frame was compiled and analyzed using BLAST search (GenBank, Bethesda, Md.) and alignment of the five most homologous proteins with the derived amino acid sequence of the encoded gene was achieved using a BCM Search Launcher: Multiple Sequence Alignments Pattern-Induced Multiple Alignment Method (See F. Corpet, Nuc. Acids Res., 16:10881-10890, (1987)). Functional motifs present in the derived amino acid sequence were identified by MultiFinder.
[0321]Northern Blot Hybridizations of Tomato RNA
[0322]Ten μg of total RNA isolated from tomato flowers at various stages (bud and blossom and senescing petals that are open widely or drying), tomato leaves, and tomato fruit at various stages of ripening (breaker, i.e., green fruit with less than 10% red color, pink, i.e., the entire fruit is orange or pink, and red, either soft or firm) were separated on 1% denatured formaldehyde agarose gels and immobilized on nylon membranes. The full length tomato cDNA labeled with 32P-dCTP using a random primer kit (Boehringer Mannheim) was used to probe the filters (7×107 cpm). The filters were washed once with 1×SSC, 0.1% SDS at room temperature and three times with 0.2×SSC, 0.1% SDS at 65° C. The filters were dried and exposed to X-ray film overnight at -70° C. The results are shown in FIGS. 50-52.
[0323]Northern Blot Hybridization of Arabidopsis RNA
[0324]Total RNA from leaves of Arabidopsis plants at five weeks of age (lane 1), six weeks (lane 2) and seven weeks (lane 3) was isolated as above, separated on 1% denatured formaldehyde agarose gels and immobilized on nylon membranes. The full-length Arabidopsis senescence-induced DHS cDNA labeled with 32P-dCTP using a random primer kit (Boehringer Mannheim) was used to probe the filters (7×107 cpm). The filters were washed once with 1×SSC, 0.1% SDS at room temperature and three times with 0.2×SSC, 0.1% SDS at 65° C. The filters were dried and exposed to X-ray film overnight at -70° C. The results are shown in FIG. 55.
[0325]Northern Blot Hybridization of Carnation RNA
[0326]Total RNA from petals of carnation plants at various stages of flower development, i.e., tight-bud flowers (lane 1), beginning to open (lane 2), fully open flowers (lane 3), flowers with inrolling petals (lane 4), was isolated as above, separated on 1% denatured formaldehyde agarose gels and immobilized on nylon membranes. The full-length carnation senescence-induced DHS cDNA labeled with 32P-dCTP using a random primer kit (Boehringer Mannheim) was used to probe the filters (7×107 cpm). The filters were washed once with 1×SSC, 0.1% SDS at room temperature and three times with 0.2×SSC, 0.1% SDS at 65° C. The filters were dried and exposed to X-ray film overnight at -70° C. The results are shown in FIG. 56.
Example 2
Sorbitol Induction of Tomato Senescence-Induced DHS Gene
[0327]Tomato leaves were treated with 2 M sorbitol in a sealed chamber for six hours. RNA was extracted from the sorbitol treated leaves as follows.
[0328]Leaves (5 g) were ground in liquid nitrogen. The ground powder was mixed with 30 ml guanidinium buffer (4 M guanidinium isothiocyanate, 2.5 mM NaOAc pH 8.5, 0.8%-mercaptoethanol). The mixture was filtered through four layers of cheesecloth and centrifuged at 10,000×g at 4° C. for 30 minutes. The supernatant was then subjected to cesium chloride density gradient centrifugation at 26,000×g for 20 hours. The pelleted RNA was rinsed with 75% ethanol, resuspended in 600 μl DEPC-treated water and the RNA precipitated at -70° C. with 0.75 ml 95% ethanol and 30 μl of 3M NaOAc. Ten μg of RNA were fractionated on a 1.2% denaturing formaldehyde agarose gel and transferred to a nylon membrane. Randomly primed 32P-dCTP-labeled full length DHS cDNA (SEQ ID NO:1) was used to probe the membrane at 42° C. overnight. The membrane was then washed once in 1×SSC containing 0.1% SDS at room temperature for 15 minutes and three times in 0.2×SSC containing 0.1% SDS at 65° C. for 15 minutes each. The membrane was exposed to x-ray film overnight at -70° C.
[0329]The results are shown in FIG. 52. As can be seen, transcription of DHS is induced in leaves by sorbitol.
Example 3
Induction of the Tomato DHS Gene in Senescing Flowers
[0330]Tight flower buds and open, senescing flowers of tomato plants were harvested, and RNA was isolated as in Example 2. Ten μg RNA were fractionated on a 1.2% denaturing formaldehyde agarose gel and transferred to a nylon membrane. Randomly primed 32P-dCTP-labeled full length DHS cDNA (SEQ ID NO. 1) was used to probe the membrane at 42° C. overnight. The membrane then was washed once in 1×SSC containing 0.1% SDS at room temperature for 15 minutes and then washed three times in 0.2×SSC containing 0.1% SDS at 65° C. for fifteen minutes each. The membrane was exposed to x-ray film overnight at -70° C.
[0331]The results are shown in FIG. 50. As can be seen, transcription of DHS is induced in senescing flowers.
Example 4
Induction of the Tomato DHS Gene in Ripening Fruit
[0332]RNA was isolated from breaker, pink and ripe fruit as in Example 2. Ten μg RNA were fractionated on a 1.2% denaturing formaldehyde agarose gel and transferred to a nylon membrane. Randomly primed 32P-dCTP-labeled full length DHS cDNA (SEQ ID NO. 1) (FIG. 45) was used to probe the membrane at 42° C. overnight. The membrane then was washed once in 1×SSC containing 0.1% SDS at room temperature for 15 minutes and then washed three times in 0.2×SSC containing 0.1% SDS at 65° C. for fifteen minutes each. The membrane was exposed to x-ray film overnight at -70° C.
[0333]The results are shown in FIG. 51. As can be seen, transcription of DHS is strongest in ripe, red fruit just prior to the onset of senescence leading to spoilage.
Example 5
Induction of Tomato Senescence-Induced DHS Gene by Chilling
[0334]Tomato plants in pots (7-8 weeks old) were exposed to 6° C. for two days, three days or six days in a growth chamber. The light cycle was set for eight hours of dark and sixteen hours of light. Plants were rewarmed by moving them back into a greenhouse. Plants that were not rewarmed were harvested immediately after removal from the growth chamber. RNA was extracted from the leaves as follows.
[0335]Leaves (5 g) were ground in liquid nitrogen. The ground powder was mixed with 30 ml guanidinium buffer (4 M guanidinium isothiocyanate, 2.5 mM NaOAc pH 8.5, 0.8%-mercaptoethanol). The mixture was filtered through four layers of cheesecloth and centrifuged at 10,000 g at 4° C. for 30 minutes. The supernatant was then subjected to cesium chloride density gradient centrifugation at 26,000 g for 20 hours. The pelleted RNA was rinsed with 75% ethanol, resuspended in 600 μl DEPC-treated water and the RNA precipitated at -70° C. with 0.75 ml 95% ethanol and 30 μl of 3M NaOAc. Ten μg of RNA were fractionated on a 1.2% denaturing formaldehyde agarose gel and transferred to a nylon membrane. Randomly primed 32P-dCTP-labeled full length DHS cDNA (SEQ ID NO:1) was used to probe the membrane at 42° C. overnight. The membrane was then washed once in 1×SSC containing 0.1% SDS at room temperature for 15 minutes and three times in 0.2×SSC containing 0.1% SDS at 65° C. for 15 minutes each. The membrane was exposed to x-ray film overnight at -70° C.
[0336]The results are shown in FIG. 53. As can be seen, transcription of DHS is induced in leaves by exposure to chilling temperature and subsequent rewarming, and the enhanced transcription correlates with chilling damage measured as membrane leakiness.
Example 6
Generation of an Arabidopsis PCR Product Using Primers Based on Unidentified Arabidopsis Genomic Sequence
[0337]A partial length senescence-induced DHS sequence from an Arabidopsis cDNA template was generated by PCR using a pair of oligonucleotide primers designed from Arabidopsis genomic sequence. The 5' primer is a 19-mer having the sequence, 5'-GGTGGTGT5TGAGGAAGATC (SEQ ID NO:7); the 3' primer is a 20 mer having the sequence, GGTGCACGCCCTGATGAAGC-3' (SEQ ID NO:8). A polymerase chain reaction using the Expand High Fidelity PCR System (Boehringer Mannheim) and an Arabidopsis senescing leaf cDNA library as template was carried out as follows.
Reaction Components:
TABLE-US-00001 [0338] cDNA 1 μl (5 × 107 pfu) dNTP (10 mM each) 1 μl MgCl2 (5 mM) + 10x buffer 5 μl Primers 1 and 2 (100 μM each) 2 μl Expand High Fidelity DNA polymerase 1.75 U Reaction volume 50 μl
[0339]Reaction Parameters:
[0340]94° C. for 3 min
[0341]94° C./1 min, 58° C./1 min, 72° C./2 min, for 45 cycles 72° C. for 15 min.
Example 7
Isolation of Genomic DNA and Southern Analysis
[0342]Genomic DNA was extracted from tomato leaves by grinding 10 grams of tomato leaf tissue to a fine powder in liquid nitrogen. 37.5 ml of a mixture containing 25 ml homogenization buffer [100 mM Tris-HCl, pH 8.0, 100 mm EDTA, 250 mM NaCl, 1% sarkosyl, 1% 2-mercaptoethanol, 10 μg/ml RNase and 12.5 ml phenol] prewarmed to 60° C. was added to the ground tissue. The mixture was shaken for fifteen minutes. An additional 12.5 ml of chloroform/isoamyl alcohol (24:1) was added to the mixture and shaken for another 15 minutes. The mixture was centrifuged and the aqueous phase reextracted with 25 ml phenol/chloroform/isoamylalcohol (25:24:1) and chloroform/isoamylalcohol (24:1). The nucleic acids were recovered by precipitation with 15 ml isopropanol at room temperature. The precipitate was resuspended in 1 ml of water.
[0343]Genomic DNA was subjected to restriction enzyme digestion as follows: 10 μg genomic DNA, 40 μl 10× reaction buffer and 100 U restriction enzyme (XbaI, EcoRI, EcoRV or HinDIII) were reacted for five to six hours in a total reaction volume of 400 μl. The mixture was then phenol-extracted and ethanol-precipitated. The digested DNA was subjected to agarose gel electrophoresis on a 0.8% agarose gel at 15 volts for approximately 15 hours. The gel was submerged in denaturation buffer [87.66 g NaCl and 20 g NaOH/Liter] for 30 minutes with gentle agitation, rinsed in distilled water and submerged in neutralization buffer [87.66 g NaCl and 60.55 g tris-HCl, pH 7.5/Liter] for 30 minutes with gentle agitation. The DNA was transferred to a Hybond-N.sup.+ nylon membrane by capillary blotting.
[0344]Hybridization was performed overnight at 42° C. using 1×106 cpm/ml of 32P-dCTP-labeled full length DHS cDNA or 3'-non-coding region of the DHS cDNA clone. Prehybridization and hybridization were carried out in buffer containing 50% formamide, 6×SSC, 5×Denhardt's solution, 0.1% SDS and 100 mg/ml denatured salmon sperm DNA. The membrane was prehybridized for two to four hours; hybridization was carried out overnight.
[0345]After hybridization was complete, membranes were rinsed at room temperature in 2×SSC and 0.1% SDS and then washed in 2×SSC and 0.1% SDS for 15 minutes and 0.2×SSC and 0.1% SDS for 15 minutes. The membrane was then exposed to x-ray film at -80° C. overnight. The results are shown in FIG. 49.
Example 8
Isolation of a Senescence-Induced eIF-5A Gene from Arabidopsis
[0346]A full-length cDNA clone of the senescence-induced eIF-5A gene expressed in Arabidopsis leaves was obtained by PCR using an Arabidopsis senescing leaf cDNA library as template. Initially, PCR products corresponding to the 5'- and 3'-ends of the gene were made using a degenerate upstream primer <AAARRYCGMCCYTGCAAGGT> (SEQ ID NO:17) paired with vector T7 primer <AATACGACTCACTATAG> (SEQ ID NO:18), and a degenerate downstream primer <TCYTTNCCYTCMKCTAAHCC> (SEQ ID NO:19) paired with vector T3 primer <ATTAACCCTCACTAAAG> (SEQ ID NO: 20). The PCR products were subcloned into pBluescript for sequencing. The full-length cDNA was then obtained using a 5'-specific primer <CTGTTACCAAAAAATCTGTACC> (SEQ ID NO: 21) paired with a 3'-specific primer <AGAAGAAGTATAAAAACCATC> (SEQ ID NO: 22), and subcloned into pBluescript for sequencing.
Example 9
Isolation of a Senescence-Induced eIF-5A Gene from Tomato Fruit
[0347]A full-length cDNA clone of the senescence-induced eIF-5A gene expressed in tomato fruit was obtained by PCR using a tomato fruit cDNA library as template. Initially, PCR products corresponding to the 5'- and 3'-ends of the gene were made using a degenerate upstream primer (SEQ ID NO:17) paired with vector T7 primer (SEQ ID NO:18), and a degenerate downstream primer (SEQ ID NO:19) paired with vector T3 primer (SEQ ID NO: 20). The PCR products were subcloned into pBluescript for sequencing. The full-length cDNA was then obtained using a 5'-specific primer <AAAGAATCCTAGAGAGAGAAAGG> (SEQ ID NO: 23) paired with vector T7 primer (SEQ ID NO: 18), and subcloned into pBluescript for sequencing.
Example 10
Isolation of a Senescence-Induced eIF-5A Gene from Carnation
[0348]A full-length cDNA clone of the senescence-induced eIF-5A gene expressed in carnation flowers was obtained by PCR using a carnation senescing flower cDNA library as template. Initially, PCR products corresponding to the 5'- and 3'-ends of the gene were made using a degenerate upstream primer (SEQ ID NO:17) paired with vector T7 primer (SEQ ID NO:18), and a degenerate downstream primer (SEQ ID NO:19) paired with vector T3 primer (SEQ ID NO: 20). The PCR products were subcloned into pBluescript for sequencing. The full-length cDNA was then obtained using a 5'-specific primer <TTTTACATCAATCGAAAA> (SEQ ID NO: 24) paired with a 3'-specific primer <ACCAAAACCTGTGTTATAACTCC> (SEQ ID NO: 25), and subcloned into pBluescript for sequencing.
Example 11
Isolation of a Senescence-Induced DHS Gene from Arabidopsis
[0349]A full-length cDNA clone of the senescence-induced DHS gene expressed in Arabidopsis leaves was obtained by screening an Arabidopsis senescing leaf cDNA library. The sequence of the probe (SEQ ID NO: 26) that was used for screening is shown in FIG. 82. The probe was obtained by PCR using the senescence leaf cDNA library as a template and primers designed from the unidentified genomic sequence (AB017060) in GenBank. The PCR product was subcloned into pBluescript for sequencing.
Example 12
Isolation of a Senescence-Induced DHS Gene from Carnation
[0350]A full-length cDNA clone of the senescence-induced DHS gene expressed in carnation petals was obtained by screening a carnation senescing petal cDNA library. The sequence of the probe (SEQ ID NO: 27) that was used for screening is shown in FIG. 83. The probe was obtained by PCR using the senescence petal cDNA library as a template and degenerate primers (upstream: 5' TTG ARG AAG ATY CAT MAA RTG CCT 3') (SEQ ID NO: 28); downstream: 5' CCA TCA AAY TCY TGK GCR GTG TT 3') (SEQ ID NO: 29). The PCR product was subcloned into pBluescript for sequencing.
Example 13
Transformation of Arabidopsis with Full-Length or 3' Region of Arabidopsis DHS in Antisense Orientation
[0351]Agrobacteria were transformed with the binary vector, pKYLX71, containing the full-length senescence-induced Arabidopsis DHS cDNA sequence or the 3' end of the DHS gene (SEQ ID NO:30) (FIG. 80), both expressed in the antisense configuration, under the regulation of double 35S promoter. Arabidopsis plants were transformed with the transformed Agrobacteria by vacuum infiltration, and transformed seeds from resultant T0 plants were selected on ampicillin.
[0352]FIGS. 65-68 are photographs of the transformed Arabidopsis plants, showing that expression of the DHS gene or 3' end thereof in antisense orientation in the transformed plants results in increased biomass, e.g., larger leaves and increased plant size. FIG. 69 illustrates that the transgenic Arabidopsis plants have increased seed yield.
[0353]In another experiment, a vector was constructed using the 3' end of the arabidopsis DHS gene (SEQ ID NO:30), in the antisense configuration under the regulation of an arabidopsis Rubisco promoter (At-rbcS promoter). The vector construct used plasmid pBI101. The GUS gene was removed but the NOS-ter remained. Rusbiso access number is X14564. The promoter sequence that was used is from nucleotide 2821 to 4780. See FIG. 152 for the construct. This vector was used to transform Arabidopsis. FIGS. 153 and 154 show that resulting transgenic plants have increased biomass as compared to the wild type control and the empty binary vector control. FIG. 155 shows that the transgenic lines had about a ˜100% increase in seed yield. FIG. 156 shows that the transgenic lines have increased root development.
Example 14
Transformation of Tomato Plants with Full-Length or 3' Region of Tomato DHS in Antisense Orientation
[0354]Agrobacteria were transformed with the binary vector, pKYLX71, containing the full-length senescence-induced tomato DHS cDNA sequence or the 3' end of the DHS gene (SEQ ID NO:31) (FIG. 81), both expressed in the antisense configuration, under the regulation of double 35S promoter. Tomato leaf explants were formed with these Agrobacteria, and transformed callus and plantlets were generated and selected by standard tissue culture methods. Transformed plantlets were grown to mature fruit-producing T1 plants under greenhouse conditions.
[0355]FIGS. 70-79 are photographs showing that reduced expression of the senescence-induced tomato DHS gene in the transformed plants results in increased biomass, e.g., larger leaf size and larger plants as seen in the transformed Arabidopsis plants, as well as delayed softening and spoilage of tomato fruit.
Example 15
Transformation of Tomato Plants with the 3' Region of Tomato DHS in Antisense Orientation
[0356]Agrobacteria were transformed with the binary vector, pKYLX71, containing the 3' end of the DHS gene (FIG. 81) expressed in the antisense configuration, under the regulation of double 35S promoter. Tomato leaf explants were formed with these Agrobacteria, and transformed callus and plantlets were generated and selected by standard tissue culture methods. Transformed plantlets were grown to mature fruit producing T1 plants under green house conditions.
[0357]Fruit from these transgenic plants with reduced DHS expression were completely free of blossom end rot under conditions in which about 33% of fruit from control plants developed this disease. Blossom end rot is a physiological disease attributable to nutrient stress that causes the bottom (blossom) end of the fruit to senesce and rot. FIGS. 84A and 84B are photographs showing a control fruit exhibiting blossom end rot and a transgenic fruit that is free of blossom end rot.
[0358]The results indicate that reducing the expression of DHS prevents the onset of tissue and cell death arising from physiological disease.
Example 16
Expression of Arabidopsis thaliana Translation Initiation Factor 5A (AteIF-5A) Isoforms in Wild Type Columbia
Plant Material
[0359]Seeds of Arabidopsis thaliana, ecotype Columbia, were grown in Promix BX soil (Premier Brands, Brampton, ON, Canada) in 6-inch pots. Freshly seeded pots were maintained at 4° C. for 2 days and then transferred to a growth chamber operating at 22° C. with 16-h light/8-h dark cycles. Lighting at 150 μmol radiation m-2.s-1 was provided by cool-white fluorescent bulbs. Whole rosettes were collected one week intervals at 2 weeks to 7 weeks of age, cauline leaves were collected at 5 weeks, stem, siliques, buds, and flowers were collected at 6 weeks and imbibed seeds (24 hours in water) were also collected, flash frozen in liquid nitrogen and stored at -80° C.
[0360]Infection of Arabidopsis thaliana Plants with Pseudomonas syringae
[0361]Seeds of Arabidopsis thaliana ecotype Columbia were sown onto Promix BX soil (Premier Brands, Brampton, ON, Canada) in flats containing 64 growth cells. The seeded flats were maintained at 4° C. for 2 days and transferred to a growth chamber with photoperiod of 9-h light/15-h dark. All plants were treated at 4 weeks of age, though physiologically due to the shortened photoperiod these appear to be slower in development.
[0362]Rosette leaves of 4-week-old plants were infected with avirulent (avr) and virulent (vir) strains Pseudomonas syringae pv. Tomato DC 3000 obtained from Dr. Robin Cameron (university of Toronto, Toronto, Canada). The abaxial surface of the rosette leaves of each plant was inoculated using 1 ml syringe without a needle. Plants were treated using one of four treatments: no inoculation, mock-inoculation with 10 mM MgCl2, inoculation with avr P. syringae strain (106 cfu/ml 10 mM MgCl2) or inoculation with vir P. syringae strain (106 cfu/ml 10 mM MgCl2). Two bacterial counts were made, one immediately after inoculation and the second 3 days later, to ensure that a sufficient amount of bacteria was infiltrated to induce systemic acquired resistance in the avr treatment. The inoculated leaves were harvested at predetermined time points for subsequent analysis.
[0363]Plants with reduced DHS or wounding/pathogen induced eIF-5A expression were developed using antisense T-DNA insertions for either gene. These plant lines have shown marked resistance to Pseudomonas syringae pv Tomato DC 300, with transgenic lines exhibiting up to a 99% decrease in bacterial load, relative to the wild type plants. See FIGS. 43 and 44. Data using crop plants have also indicated enhanced pathogen resistance.
[0364]Wounding of Arabidopsis thaliana Plants with Hemostat
[0365]4-week-old plants grown under normal lighting conditions were wounded by crushing with hemostat along the midvein (approximately 10% of the leaf surface) according to Stotz et al (2000). Tissue was harvested at 0 minutes, 1 hour and 9 hours and immediately frozen in liquid nitrogen and stored at -80° C. for further analysis.
[0366]RNA Isolation and Northern Blotting
[0367]Total RNA for Northern blot analysis was isolated from Arabidopsis thaliana rosette leaves according to Davis et al. (1986). The RNA was fractionated on a 1% agarose gel and transferred to nylon membranes. (Davis et. al., 1986) Immobilized RNA was hybridized overnight at 42° C. with radiolabeled 3'UTR portions of senescence-induced AteIF-5A, wounding/pathogen induced AteIF-5A or growth AteIF-5A. The 3'UTRs were labeled with [α-32P]-dCTP using a random primer kit (Boehringer Mannheim). The hybridized membranes were washed twice in 2×SSC containing 0.1% SDS at 42° C. for 15 minutes and twice in 1×SSC containing 0.1% SDS at 42° C. for 30 minutes. Hybridization was visualized by autoradiography after an overnight exposure at -80° C.
[0368]Antibody Production and Purification
[0369]Eukaryotic translation initiation factor 5A (eIF-5A) isoforms of Arabidopsis thaliana (At) are highly homologous at the amino acid level, especially at the N-terminal region and the central region of the proteins (FIG. 1). In order to obtain antibodies that will be isoform specific, peptides were designed against regions in the isoforms of AteIF-5A that appeared to be unique to each other. An additional cysteine residue was added to each peptide at the N-terminus for conjugation with KLH. The sequences used were: CNDDTLLQQIKS (SEQ ID NO: 35) for senescence-induced AteIF-5A, CTDDGLTAQMRL (SEQ ID NO: 36) for wounding/pathogen induced AteIF5A, and CTDEALLTQLKN (SEQ ID NO: 37) for growth AteIF-5A. When these sequences were submitted to protein BLAST (short nearly exact sequences; limited by Arabidopsis thaliana; expected number 20000; word size 2; Matrix PAM90; Gap cost 91) the significant sequences that found in the database were only the matched AteIF-5A and no other. The peptides were synthesized at the University of Western Ontario Peptide Synthesis facility. The carrier protein, Keyhole Limpet Hemocyanin (Sigma), was conjugated to the N-terminal cysteine of the peptide using m-maleimidobenzoyl-N-hydroxysuccinimide ester according to Drenckhahn et al. (1993) and Collawn and Patterson (1999). The rabbits were injected four times at two-week intervals with the linked peptide. Two weeks after the final injection blood is collected by exsanguination of the rabbits and clotting of the collected blood in order to amass the antisera.
[0370]Protein Fractionation and Western Blotting
[0371]Tissues list above were homogenized (˜0.5 g/ml) in buffer (50 mM EPPS, pH 7.4, 0.25M sorbitol, 10 mM EDTA, 2 mM EGTA, 1 mM DTT, 10 mM amino-n-caproic acid, Protease Inhibitor Cocktail for Plant tissues (Sigma)) in an eppendorf tube with a small pestle, or in a large mortar and pestle. The homogenates were centrifuged briefly in the microcentrifuge at maximum speed and the pellet was discarded. The total protein was quantified according to Ghosh et al. (1988). SDS-PAGE was performed on Mini protein Dual Slab cells (BioRad, Mississauga, Ontario), and the gels (12% polyacrlyamide) were stained with Coomassie brilliant blue R250 (Fairbanks et. al. 1971) or transferred to polyvinyldiene difluoride (PVDF) membranes using the semi-dry transfer method (semi-dry transfer cell, Bio-Rad, Hercules, Calif.). The blots were blocked for 30 s in 1 mg/ml polyvinyl alcohol (Miranda et. al., 1993) and for 1 hour in phosphate-buffered saline (PBS) containing 0.1% (v/v) Tween 20 and 5% (w/v) powdered milk. Primary antibody (from bleeds after second injection) was diluted 1:50 in PBS containing 0.1% (v/v) Tween 20 and 1% (w/v) powdered milk. Antigen was visualized using secondary antibody made in goat against rabbit antibody coupled to alkaline phosphatase (Bioshop, Burlington, Ontario) and the phosphatase substrates, NBT and BCIP (BioRad, Mississauga, ON).
Example 17
Production of Transformed Arabidopsis thaliana Plants Over Expressing Three eIF-5A Isoforms
[0372]Primer Design
[0373]Eukaryotic translation initiation factor 5A (eIF-5A) isoforms of Arabidopsis thaliana (At) are highly homologous in the coding region (FIG. 2). To avoid problems with amplification of the correct genes, primers for senescence-induced AteIF-5A, wounding/pathogen induced eIF-5A and growth eIF-5A were designed from the approximate beginning of the 5'UTR and at the end of the 3'UTR as shown in FIG. 3, 4 and 5 respectively. The 5'UTR and 3'UTR were estimated based on EST information and other sequence information in the GenBank database. The appropriate restriction sites were added to the ends of the primers for ligation in the sense orientation in the pKYLX71 binary vector (FIG. 6). For senescence-induced AteIF-5A the upstream primer is 5' AAGCTT GATCGTGGTCAACTTCCTCTGTTACC 3' (SEQ ID NO: 38) and the downstream primer is 5' GAGCT CAGAAGAAGTATAAAAACCATC 3' (SEQ ID NO: 39). For wounding/pathogen induced AteIF-5A the upstream primer is 5' CTC GAGTGCTCACTTCTCTCTCTTAGG 3' (SEQ ID NO: 40) and the downstream primer is 5' GAGCTCA AGAATAACATCTCATAAGAAAC 3' (SEQ ID NO: 41). The upstream primer for growth AteIF-5A is 5' CTC GAGCTAAACTCCATTCGCTGACTTCGC 3' (SEQ ID NO: 42) and the downstream primer is 5' GAGC TCTAGTAAATATAAGAGTGTCTTGC 3' (SEQ ID NO: 43). The restriction sites that were added into the primers were HindIII and SacI for senescence-induced AteIF-5A, XhoI and SacI for wounding/pathogen induced AteIF-5A, and XhoI and SacI for growth AteIF-5A as indicated by underlining in the primers listed above.
[0374]Isolation of Genomic DNA from Arabidopsis thaliana
[0375]Genomic DNA was isolated from 3-week-old rosette leaf. The tissue was homogenized in extraction buffer (200 mM Tris pH 7.5, 250 mM NaCl, 25 mM EDTA, 0.5% SDS) and the resulting homogenate was vortexed for 15 seconds. The remaining debris was removed by centrifugation in a microcentrifuge at maximum speed for 1 minute. The supernatant was collected and mixed in a 1:1 ratio with isopropanol, vortexed and left at room temperature for 2 minutes. A pellet was collected by centrifugation in a microcentrifuge at maximum speed for 5 minutes, washed with 70% ethanol and vacuum dried for 2 minutes. The dried pellet was resuspended in water and treated with 1:1 volume of chloroform and vortexed. After centrifugation in a microcentrifuge at maximum speed for 2 minutes the top layer was collected and treated with 20 μl salt (3M sodium acetate) and 2 volumes of ethanol for precipitation at -20° C. for 30 minutes. The purified genomic DNA was then centrifuged at maximum speed for 30 minutes in a microcentrifuge, dried and resuspended in water for PCR.
[0376]PCR from Genomic DNA
[0377]PCR was performed with the primers described above. The PCR reaction mixture contained 1×Tsg or Taq polymerase reaction buffer, 1 U of Tsg or Taq polymerase, 0.2 mM dNTP, 2 mM MgCl2, and 15 pmols of each specific primer accordingly. The reaction began with a hot start at 95° C. for 10 minutes and first cycle consisted of 1 minute denaturing temperature of 95° C., 2 minutes annealing temperature of 55° C., and a 2 minute extension temperature of 72° C. The following 29 cycles proceeded a touchdown program where the annealing temperature was decreased by 0.5° C. per cycle, and the final cycle had an annealing temperature of 40° C. The final extension of 72° C. was held for 10 minutes. The PCR products were separated by 1% agarose gel electrophoresis, cut out and retrieved by Millipore Ultrafree-DA for DNA Extraction from Agarose spin columns (Millipore Corporation, Bedford, Mass.) according to directions.
[0378]Ligation into pGEM®-T Easy
[0379]Purified PCR products were ligated into pGEM®-T Easy Vector (FIG. 7) according to directions provided by Promega. Briefly, PCR products were mixed in a 3:1 ratio with pGEM T-Easy Vector, 3 Weiss Units T4 DNA ligase in Rapid Ligation Buffer (30 mM Tris-HCl, 10 mM MgCl2, 10 mM DTT, 1 mM ATP, and 5% polyethylene glycol (MW8000, ACS Grade) pH 7.8) provided in the Promega pGEM®-T Easy Vector System (Promega Corporation, Madison Wis.). The ligation reaction was incubated overnight at 15° C. and transformed into competent E. coli DH5-α cell suspension (made competent using RbCl/CaCl; Kushner, 1978). The transformation mixture was first incubated on ice for 30 minutes, heat shocked for 90 seconds at 42° C., and allowed to recover at 37° C. for 1 hour after the addition of 1 ml 2xYT broth. The transformed cells were pelleted, resuspended in a small volume of 2xYT broth and plated on agar plates containing 50 μg/ml ampicillin for selection. Only transformants are able to grow on the ampicillin-containing plates as the pGEM®-T Easy Vector provides ampicillin resistance to the cells. Transformants were selected and screened for the PCR product insert ligated into the pGEM®-T Easy Vector.
[0380]Screening for PCR Product Inserts in pGEM®-T Easy Vector Through Restriction Enzyme Digestions
[0381]Colonies that grew on selection media were grown in 5 ml 2xYT broth containing 50 μg/ml ampicillin overnight at 37° C. The recombinant plasmids from the selected colonies were purified using Wizard Prep DNA Purification Kit (Promega). The plasmid DNA was digested with EcoRI for 1 hour at 37° C. and visualized on a 1% agarose gel for verification that the AteIF-5As insert sizes were present. The positive plasmids were then sequenced by the Core Molecular Biology Facility (University of Waterloo, Waterloo, ON) for confirmation that the sequence is suitable for over expression in planta.
[0382]Ligation into pKYLX71
[0383]The constructs of pGEM:wounding/pathogen induced AteIF-5A, and pGEM:growth AteIF-5A were double digested with XhoI and SacI and sub-cloned into the binary vector, pKYLX71 that had also been digested with XhoI and SacI. These enzyme digestions ensured that wounding/pathogen induced AteIF-5A and growth AteIF-5A would be inserted in the sense orientation in the binary vector pKYLX71 under the control of the cauliflower mosaic virus double 35S promoter. The ligation reactions used 1 μg of binary vector and 3 μg of either wounding/pathogen induced AteIF-5A or growth AteIF-5A. Ligation took place in ligation buffer (30 mM Tris-HCl, 10 mM MgCl2, 10 mM DTT, 1 mM ATP, and 5% polyethylene glycol (MW8000, ACS Grade) pH 7.8) with 3 Weiss units of T4 DNA Ligase (Fermentas). The ligation reaction was incubated overnight at 15° C. and transformed into competent E. coli DH5-α cell suspension (made competent using RbCl/CaCl; Kushner, 1978). The transformation mixture was first incubated on ice for 30 minutes; heat shocked for 90 seconds at 42° C. and allowed to recover at 37° C. for 1 hour after the addition of 1 ml 2xYT broth. The transformed cells were pelleted, resuspended in a small volume of 2xYT broth and plated on agar plates containing 50 μg/ml tetracycline for selection. Only transformants are able to grow on the tetracycline-containing plates as the binary vector pKYLX71 provides tetracycline resistance to bacterial cells. Transformants were selected and screened for wounding/pathogen induced AteIF-5A or growth AteIF5A insert by PCR and double digestion with XhoI and SacI. Following PCR amplification (same as was done with genomic DNA explained above) and digestion, the products were separated using 1% agarose electrophoresis for conformation of the correct sized insert.
[0384]Agrobacterium Electroporation and Selection
[0385]The constructs pKYLX71:wounding/pathogen induced AteIF-5A and pKYLX71:growth AteIF-5A was electroporated into competent Agrobacterium tumefaciens GV3010. The preparation of competent Agrobacterium cells a single colony was inoculated in 5 ml of 2xYT broth containing 50 μg/ml of rifampicin, and 50 μg/ml gentamycin. This grew overnight at 28° C. in a Form a Scientific Orbital Shaker (Fisher Scientific) at 280 rpm and was used to inoculate 30 ml cultures of 2xYT also with 50 μg/ml of rifampicin, and 50 μg/ml gentamycin at various dilutions (1:500, 1:1000, 1:2000). The newly inoculated cultures grew until OD600 was between 0.5 and 0.8 before being cooled and centrifuged down in an SS-34 rotor (Sorvall) at 2000 g for 15 minutes. The pellets were resuspended in 50 ml of ice-cold water and centrifuged at 2000 g for 15 minutes. This washing procedure was repeated for a total of four times to remove the salts and the dead cells from the culture. The final pellet was resuspended in 40 ml ice cold 10% (v/v) glycerol and centrifuged at 2000 g for 15 minutes and repeated once. The pellet was then resuspended in 100 μl ice-cold 10% glycerol and mixed well. Cells were split up into aliquots of 100 μl and stored on ice.
[0386]For electroporation of the DNA constructs into the competent Agrobacterium cells the 100 μl aliquots were each mixed well with 500 ng of DNA construct. The bacteria:vector mixture was then transferred to a pre-cooled electroporation cuvette and placed in the Gene Pulser (Biorad) adjusted to the following settings: 2.5 kV, 25 μF, and 200Ω. After electroporation 1 ml 2xYT broth was added and the whole suspension was transferred to a culture tube. The electroporated cultures were incubated at 28° C., 280 rpm, for 3 hours to allow them to recover and then 2 ml 2×YT both was added as well as 50 μg/ml of rifampicin, and 50 μg/ml gentamycin. After 2 days of growing in culture the electroporated cells were plated on tetracycline, gentamycin and rifampicin (all at 50 μg/ml) and colonies grew after an addition 2 days. The resulting colonies were screened for pKYLX71:wounding/pathogen induced AteIF-5A or pKYLX71:growth AteIF-5A by PCR and double digestion with SacI and XhoI, and visualized by separation on a 1% agarose gel.
[0387]Plant Transformation
[0388]A positive colony of Agrobacterium tumefaciens GV3010 containing either pKYLX71:wounding/pathogen induced AteIF-5A or pKYLX71:growth AteIF-5A were used for the transformation of wild type Arabidopsis thaliana ecotype Columbia. In preparation of the bacterial slurry used for plant transformation a single colony positive for pKYLX71:wounding/pathogen induced AteIF-5A or pKYLX71:growth AteIF-5A construct was inoculated in 5 ml of 2xYT broth containing 50 μg/ml of tetracycline, 50 μg/ml of rifampicin, and 50 μg/ml gentamycin. This grew for 2 days at 28° C. in a Form a Scientific Orbital Shaker (Fisher Scientific) at 280 rpm and was used to inoculate 35 ml (total) 2xYT also with 50 μg/ml of rifampicin, and 50 μg/ml gentamycin. The 35 ml culture was grown overnight at 28° C., 280 rpm, and used to inoculate 535 ml (total) 2xYT with 50 μg/ml of rifampicin, and 50 μg/ml gentamycin. Again the culture was grown overnight at 28° C., 280 rpm, to an OD600 of about 2.0.
[0389]The cultures were transferred to two 250 ml tubes before centrifugation for 15 minutes at 1945 g at 4° C. in a GSA rotor (Sorvall). The pellets were resuspended in 500 ml of infiltration media (1.1 g MS salts, 25 g sucrose, 0.25 g MES, pH5.7 with KOH, 100 ng/ml benzylaminopurine and 50 μl Vac-In-Stuff (Silwet L-77; Lehle Seeds)) and placed in a large plastic dish in a vacuum desiccator with 4 large rubber stoppers. Five pots containing 8 plants each at the right stage of development were used sequentially for infiltration. Each pot was first inverted over a trash can to remove any loose soil, then was placed (still inverted) into plastic container in the glass desiccator so that the 4 large rubber stoppers acted as stand for the inverted pot thus allowing the bolts to be dipped into the Agrobacterium slurry, but not the rosettes. The plants were then subjected to a vacuum (400 mm Hg) in this inverted state for 10 minutes. The vacuum infiltrated plants were then allowed to recover and grown as usual in the growth chamber conditions explained in the plant material section. After several weeks when the siliques were dry and seed matured, the seeds were collected with each pot pooled together.
[0390]Selecting Plant Transformants and Segregation Analysis
[0391]To identify primary transformants, seeds from the vacuum-infiltrated plants were surface sterilized in a solution of 1% (v/v) sodium hypochlorite and 0.1% (v/v) Tween 80 for 20 minutes on a rotator (Barnstead/Thermolyne), rinsed four times with sterile water, and resuspended in a sterile 0.8% agar. The resuspended seeds were then planted onto sterile, half-strength Murashige and Skoog (MS) medium (2.2 g/L) supplemented with 1% (w/v) sucrose, 0.5 g/L 2-[N-Morpholino]ethanesulfonic acid (MES), 0.7% (w/v) bacteriological agar and 40 to 50 μg/ml kanamycin (Murashige and Shoog, 1962). Only transformants are able to grow on the kanamycin-containing plates since the binary vector provides the kanamycin resistance gene to the transformant seedlings (FIG. 6). Seedlings that do not harbour the binary vector become yellow and die, as there is no kanamycin resistance gene. Wild-type seedlings were used as controls and plated onto MS medium without kanamycin added to the medium, as well seeds from a homozygous line containing empty pKYLX71 vectors were seeded as controls on kanamycin containing plates. The empty vector control is useful in demonstrating the effect kanamycin has on growth of the seedlings as well as the effect of random integration of the binary vector into the genome of Arabidopsis thaliana. A small amount of wild type seed was plated onto a small area of each plate containing MS medium and 40 to 50 μg/ml kanamycin. This was done in order to make sure the medium was selective enough for the transformants and to test the strength of the kanamycin.
[0392]The seeded plates were kept at 4° C. for 3 days to synchronize the germination. After 3 days the plates were transferred to growth chambers where they grew for an additional 7 days under 16-h light/8-h dark cycles at 20±2° C. Lighting was maintained at 150 μmol radiation m-2.s-1 and was provided by cool-white fluorescent bulbs. The efficiency for transformation of Arabidopsis thaliana plants with the pKYLX71:wounding/pathogen induced AteIF-5A and pKYLX71:growth AteIF-5A vectors was determined.
[0393]After a total of 10 days since seeding, the 14 transformants or the 16 transformants for Sense wounding/pathogen induced AteIF-5A and Sense growth AteIF-5A respectively were transplanted to Promix BX soil (Premier Brands, Brampton, ON, Canada) in flats containing 32 cells. These transplanted T1 generation plants were then transferred into another growth chamber operating at 22° C. with 16-h light/8-h dark cycles. Lighting at 150 mmol radiation m-2.s-1 was provided by cool-white fluorescent bulbs. The T1 generation plants grew to maturity and produced T2 generation seeds. These were harvested and stored at -20° C. until further screening was done. The T1 generation was named 1, 2, 3, etc. All 16 lines of Sense growth AteIF-5A plants survived and produced seeds, but only 9 out of 14 transformants of the Sense wounding/pathogen induced AteIF-5A plants survived and produced seeds.
[0394]The selection of T2 generation transformants was conducted in the same way as the T1 generation transformants. Line 12 of the Sense growth AteIF-5A plants produced no transformants on the selectable media and was not included in any further work. Lines 1 through to 16 (minus line 12) of the Sense growth AteIF-5A plants each had 8 sublines carried through. These were named A through H so that for example in the T1 line 1, the T2 generation plants were named 1A, 1B, 1C, etc. Lines 1, 2, 3, 4, 5, 7, 9, and 11 of the Sense wounding/pathogen induced AteIF-5A plants each had 8 sublines (A-H) carried through. Line 12 T1 plants had only produced about 30 T2 seeds and only 1 subline in the T2 generation will be carried through. T2 plants of Sense wounding/pathogen induced AteIF-5A are still growing and being characterized. The T2 plants for the Sense growth AteIF-5A have matured and produced seeds, which were harvested and stored at -20° C. until further analysis.
[0395]The selection of the T3 generation transformants of Sense growth AteIF-5A was conducted in the same manner as the T2. Eight lines were chosen based on phenotype analysis as well as the degree of over expression of Sense growth AteIF-5A. The levels of expression were broken down into four categories: high-level expression, medium-level expression, low-level expression, and no expression (due to co-suppression). Two lines were chosen for each of the levels of expression and 12 plants from each line were transplanted. The corresponding lines for these four levels of expression are: 1A, 2D, 4D, 15A, 8D, 9H, 11C and 16C. The T3 generation for Sense growth AteIF-5A plants are still growing and being characterized.
Example 18
Phenotype Analysis of Sense Wounding/Pathogen Induced AteIF5A and Sense Growth AteIF5A
[0396]Photographic Record
[0397]Morphological phenotypes of the Sense wounding/pathogen induced AteIF-5A and Sense growth AteIF-5A lines were recorded photographically during segregation, as were the phenotypes of the corresponding control wild type plants (Arabidopsis thaliana ecotype Columbia) and plants transformed with an empty binary vector pKYLX71.
[0398]Seed Measurements
[0399]T3 seeds collected from T2 plants of Sense growth AteIF-5A were measured for total seed yield (both weight and volume), seed size (length and width), and calculated individual weight and volume of produced seed. Total seed yield by weight was measured on a Sartorius analytical digitized scale, and the volume was determined by pouring and packing down the total seed yielded by each plant into a glass 1 ml syringe that was graduated every 100 μl. To determine the seed size by length, width and calculated volume, the seeds were placed on a slide containing a micrometer and viewed on an Olympus BX51 Microscope. Photographs of the seeds on the micrometer were taken with a Spot Insight Color Camera (Diagnostic Instruments Inc.) attached to a Compaq Evo D500 (Compaq Company Corporation; Intel® Pentium 4 CPU 1.7 GHz, 262 MG RAM, running Windows 2000). Using Image-Pro Express Version 4.0 for Windows. Measurements of 10 seeds in each subline were made using the micrometer in the image for size calibration. The measurements were imported into Microsoft Excel, and calculations such as standard error and volume were performed.
Example 19
Biochemical Analysis of Sense Wounding/Pathogen Induced AteIF5A and Sense Growth AteIF5A
Protein Fractionation and Western Blotting
[0400]The first cauline leaf from each subline of Sense growth AteIF-5A T2 plants were collected and proteins extracted as described above. Total protein from lines 1A, 2A, up to 16A were fractionated by 12% SDS-PAGE and transferred to a PVDF membrane. The blot was probed with growth αAteIF-5A at a 1:50 dilution. Control total protein was extracted from the first cauline leaf from wild type and empty binary vector control plants.
Example 20
Expression of Arabidopsis thaliana Translation Initiation Factor 5A (AteIF-5A) Isoforms in Wild Type Columbia
[0401]Several tissues were collected at different developmental stages and the extracted proteins from these tissues were used for Western blotting. The Western blot in FIG. 8 demonstrates that senescence-induced AteIF-5A is not present in the 2 week old rosette leaves, but is upregulated in the 3 week old rosette leaves and increases in abundance until 5 weeks and declines in abundance, but is still present at 7 weeks. No senescence AteIF-5A was detected in the PEG treated plants or control, but was present in the flower lane (which included senescent flowers) and in the imbibed seed lane reflecting senescence of cotyledonary tissues. When the blot was probed with the wounding/pathogen induced αATeIF-5A antibody, faint bands appeared in the siliques, imbibed seed and stem lanes. The band seen in the siliques and stem lanes may be due to the wounding that occurred with collection of the tissue. Since it is difficult to collect the siliques and stem, they were not flash frozen immediately allowing for some up-regulation of the wounding/pathogen induced isoform of AteIF-5A. The only band that appeared when the blot was probed with growth αAT-eIF5A was imbibed seeds, keeping with the notion that this is the isoform involved in cell division.
[0402]Plants that were treated with either no treatment, mock inoculation with MgCl2, avr P. syringae or with vir P. syringae were collected at several time points to analyze the expression of the AteIF-5As during pathogen ingress. The avr strain is recognizable by the plant and induces the hypersensitive response that leads to cell death or necrosis in the region of infection, thus disallowing the pathogen to cause disease. Furthermore the localized response eventually becomes a systemic response in order to protect the plant from further ingress. This is known as Systemic Acquired Resistance (SAR), which involves the expression of a suite of genes known as the Pathogenesis Response (PR) genes. On the other hand the vir strain will not be recognized by the plant, and will not induce a hypersensitive response and will lead to disease. The diseased state of Arabidopsis thaliana includes yellowing leaves and cell death after a few days post infection. After 72 hours post treatment control plants, mock treated plants, avr treated plants and vir treated plants were collected for western blotting with the three αAteIF-5A antibodies (FIG. 9). At this point both SAR and disease were visible in the avr treated and the vir treated plants respectively. When probed with the senescence-induced αAteIF-5A antibody, a band that was relatively the same in all the samples was observed. Since all of the plants were 4 weeks old this came with no surprise, since the senescence isoform was seen starting at 3 weeks in FIG. 8. When the blot was then probed with the wounding/pathogen induced αAteIF-5A antibody, a faint band was detectable in the untreated, mock treated and avr treated plants where there was a strong band detected in the vir treated plants. This upregulation of the wounding isoform may be due to cell death caused by disease (also a type of cellular wounding). The blot probed with growth αAteIF-5A did not show any bands and thus was not included in the figure. As the senescence-induced AteIF-5A did not change in expression during these treatments demonstrates its specificity for natural senescence. The increase in wounding/pathogen induced AteIF-5A expression also demonstrates its specificity for death due to wounding. To further investigate this possibility, an experiment was performed with wounding leaves of Arabidopsis thaliana.
[0403]The wounding experiment showed similar results as the pathogenesis experiment (FIG. 10). Northern blots were used to show the transcriptional change in of senescence-induced AteIF-5A, wounding/pathogen induced AteIF-5A and growth AteIF-5A. The probes were specific to each of the AteIF-5As and consisted of the 3' UTR of each. It was observed that like the pathogenesis experiment senescence-induced AteIF-5A expression did not change, as these were 4-week-old plants and samples were only taken over a 9-hour interval. This again is consistent with the fact that senescence-induced AteIF-5A is natural senescence specific isoform. The expression of wounding/pathogen induced AteIF-5A however did increase after 9 hours. There is probably some translational control occurring, as the transcript appears fairly constitutive (FIG. 10), but the protein does not appear as highly expressed when not induced (FIG. 9). The transcript for growth AteIF-5A was barely detectable in all the samples, and shows a decline in expression post wounding.
Example 21
Production of Transformed Arabidopsis thaliana Plants Over Expressing Three eIF-5A Isoforms
[0404]The AteIF-5As were isolated from genomic DNA by PCR (FIG. 11). The products were ligated in pGEM (FIG. 12) and the sequence was verified for suitability for over-expression in planta. Wounding/pathogen induced AteIF-5A and growth AteIF-5A were double digested out of pGEM with XhoI and SacI and ligated in the sense orientation behind the cauliflower mosaic virus 35S2 promoter in pKYLX71. Positive ligation was confirmed by digestion and PCR (FIG. 13). The pKYLX71:senescence-induced AteIF-5A and the pKYLX71:growth AteIF-5A were then electroporated into Agrobacterium tumefaciens GV3010 for transformation via vacuum infiltration of Arabidopsis thaliana wild type of the ecotype Columbia. After plant transformation the seeds were collected and transformants selected for on Kanamycin containing MS plates.
[0405]Arabidopsis thaliana plants over expressing wounding/pathogen induced AteIF-5A (Sense wounding/pathogen induced AteIF-5A)
[0406]T1 generation plants were seeded on MS plates containing 50 μg/ml Kanamycin and were stored at 4° C. for 3 days and in the growth chamber for 7 days (FIG. 14). There were 14 transformants that were transplanted to soil. A common phenotype in these 14 T1 generation plants was stunted growth. Lines 1, 4, 6, 8, 10, 11, 12, 13, and 14 were severely stunted in their growth and 6, 8, 10, 13 and 14 did not produce any seed. Lines 2 and 3 were moderately stunted whereas lines 5, 7 and 9 grew similarly to wild type plants (FIG. 15 and FIG. 16). Some other phenotypes observed in the T1 generation of Sense wounding/pathogen induced AteIF-5A plants included yellow leaves, purple cotyledons, curled up leaves and differences in flower shape. It is interesting to note that the appearance in the stunted growth was not observed until the plants were transplanted to soil. A possible explanation of this would be that during transplant the roots are damaged slightly (a consequence of transplanting that is unavoidable) and were unable to recover. In fact a preliminary experiment where seeds were soaked in a Kanamycin solution and seeded to soil directly no stunted plants were observed (whereas previously 70% of the plants had some degree of stunting), as no root damage would be invoked without transplantation.
[0407]Lines 1, 2, 3, 4, 5, 7, 8, 11 and 12 produced T2 seeds and were carried through (FIG. 17). Each T2 line has sublines A-H, except for 12, which only grew one transformant, and are currently being analyzed.
[0408]Arabidopsis thaliana plants over expressing growth AteIF-5A (Sense growth AteIF-5A)
[0409]The T1 generation seeds of Sense growth AteIF-5A were grown on selective media and 16 transformants grew (FIG. 18). The transformants were photographed over their lifetime. The phenotypes varied from similar to wild type (Lines 1, 2, 5, 6, 7, 8, 10, 11, 12, 13, 14, 15, and 16) to moderately stunted and yellow (Lines 2, 4 and 9; FIG. 19). All the lines were carried through to T2 and each line had 8 sublines labeled A-H. Line 12 did not produce any transformants in T2 and was deemed to be wild type. The T2 generation plants had much more exaggerated phenotypes than that of T1 generation plants. The lines that were carried to T3 will be discussed in detail.
[0410]The Sense growth AteIF-5A T2 generation lines were characterized in groups according to the level of expression of the growth AteIF-5A transgene. A Western blot was performed on protein extracted from cauline leaves from each line (FIG. 20). Since most of the sublines A-H demonstrated similar phenotypes within a line, the Western blot was only done with subline A of each line to get a general overview of level of expression of growth AteIF-5A. Protein from the cauline leaves of wild type plants and plants containing the empty binary vector were used as controls on the gels. The level of expression observed in these sublines can be categorized as high (Lines 1, 2, 3, 10, 13), medium (Lines 4, 5, 6, 15), low (Lines 7, 8, 9, 14) or none (Lines 11, 16, wild type and binary control). The blots were also probed with antibodies against senescence-induced AteIF-5A and wounding/pathogen induced AteIF-5A. These westerns indicated that the increase in expression in the Sense growth AteIF-5A lines is due to growth AteIF-5A and not a general upregulation of other AteIF-5A isoforms, as no significant amount of either isoform was detected. This also demonstrated that the specificity of the isoform specific antibodies is acceptable.
[0411]The Sense growth AteIF-5A lines be carried through to the T3 generation were chosen based on phenotype as well as the level of expression of growth AteIF-5A (See Table 1 for a summary of phenotypes within each line). Two lines from each category of level of expression were chosen. The lines that will be carried through are 1A, 2D, 4D, 15A, 8D, 9H, 11C, and 16C.
[0412]Line 1 according to the western blot in FIG. 20, has a high level of growth AteIF-5A expression. These plants had large, dark green rosettes with leaves that were quite round in comparison to wild type plants (FIG. 21). The rosettes of line 1 also had a whorled phenotype, where the leaves all curl in the same direction. These Sense growth AteIF-5A plants bolted slightly later than wild type. Line 2 also demonstrated high level of growth AteIF-5A expression, but differed from line 1 in that these plants were small and yellowed (FIG. 22). Line 2 plants also bolted later than the wild type and binary control plants, as well produced smaller bolts (about half the size) and fewer siliques.
[0413]Of the medium level of expression lines, line 4 appeared similar to wild type in leaf/rosette size and in bolt size, though appeared to bolt just a few days before the wild type and binary control plants. The second line with a medium level of expression of growth AteIF-5A is line 15. These plants are, like line 4, very similar to wild type, but the area that the rosette occupied was larger than the controls (FIGS. 23 and 24). The leaves of the rosette also appeared to be rounder at the tips than the controls. The bolts however did not appear to have any distinctive phenotype.
[0414]The low expressing Sense growth AteIF-5A lines that will be carried through to T3 are from lines 8 and 9. Line 8 had very large leaves and large rosettes compared to the control plants (FIG. 25). The leaves also appeared to be wider and rounder than the control plants. The time of bolting, bolt size and number seemed to be consistent with the controls. The Sense growth AteIF-5A line 9 had similar leaf shape as in line 8, but was far more yellow and smaller (FIG. 26). As in line 2 (one of the high expressing lines), these plants show stunted growth, shorter bolts, but unlike line 2, line 9 bolted about the same time as the control plants.
[0415]The two lines 11 and 16 of the Sense growth AteIF-5A plants according to the western blot (FIG. 20) have no upregulated expression of growth AteIF-5A. This may be due to cosuppression of the transgene as well as the endogenous gene. Though these plants do look similar to the controls (FIG. 27 and FIG. 28), it is believed that the transgene is incorporated into the genome of lines 11 and 16 for several reasons. Firstly, they do have Kanamycin resistance as demonstrated by the selectivity on the Kanamycin containing MS plates. Secondly, the rosette size, leaf size, and bolt size of line 16 (FIG. 28) are at least 50% larger than the controls. But the strongest evidence is in the size and composition of the T3 seeds that they produced.
[0416]The T3 seeds were measured from all lines of T2 Sense growth AteIf-5A plants. Photographs were taken of each line (the largest and the smallest highlighted in FIG. 29), and measurements were made in silico with a micrometer in the photographs used for calibration. For each line and for the controls, ten of the largest seeds in the field of view were measured and used for calculations. It was found that the high expression line 2 had seeds that were up to 3 times as large as the wild type and binary controls. Whereas the lines that demonstrated the lowest expression (Lines 11 and 16) had some of the smallest seeds that were only about 88% the size of wild type or binary control seeds. The average seed size for each line was expressed as nm3 (FIG. 30) and was calculated using an equation for the volume of an ellipsoid as seeds from Arabidopsis thaliana are approximately ellipsoid. The measured size of the control seeds fell into published guidelines as determined by Boyes et al (2001). From the measured size of individual seeds and the total seed yield (both weight and volume), the average individual seed weight was calculated and plotted (FIG. 31). It appeared that most of the lines that demonstrated a different size than that of the control seeds also had the same trend in individual seed weight. In fact when the seed weight was plotted against the seed size (volume) the relationship was mostly linear with an R2=0.7412. There were 5 lines that were outliers that had either an increased density (3 of them) or a decreased density (2 of them). One of the lines with the increased density is 8D and will be carried through T3 generation. The total seed yield from all the T2 generation plants were quite variable, with few trends. One notable line however is the medium expressing Sense growth AteIF-5A line 4D, which produced the most seeds (both weight and volume). In fact 4D produced 2.5 fold more than the control plants and will be carried through T3.
[0417]T3 seeds were plated on selection media as described previously. Lines 1A, 2D, 4D, 15A, 8D, 9H, 11C and 16C were transplanted to soil. Several other sublines of Sense growth AteIF-5A line 1 did not germinate, as well as line 2H, which had the largest seeds of all the sublines did not germinate. Plants from line 11 (one of the cosuppression lines) were not as healthy as typically found at this age. These seeds were also one of the smallest measured. It appears that these lines are still segregating, as there were still non-Kanamycin resistant plants as well as seeds that did not germinate from all the lines. This is probably a side effect of the transgene and not technique as the control seeds that were treated in the same manner, all germinated.
Example 22
Characterization of Arabidopsis Senescence-Induced eIF-5A
[0418]Methodology for Obtaining Full-Length Arabidopsis Senescence-Induced eIF-5A
[0419]Degenerate primers based on several plant eIF-5A genes, in combination with vector primers T3 & T7 were used in order to PCR an eIF-5A gene from an Arabidopsis cDNA library. Specifically, the 5' region of the eIF-5A gene was obtained from a PCR reaction utilizing both the T3 primer (located upstream of the F5A gene in the library vector) and one of the downstream (reverse-orientation) degenerate primers. Likewise, the 3' region of the gene was obtained from a PCR reaction utilizing both the T7 primer (located downstream of the eIF-5A gene in the library vector) and one of the upstream (forward-orientation) degenerate primers. The full-length eIF-5A gene was derived from alignment analysis of the 5' region and 3' region of the gene.
[0420]There are 2-3 major products for each PCR reaction. These fragments were cloned to pBluescript plasmid and sequenced. The eIF-5A positive PCR fragments were identified based on the mapping analysis against the gene bank. There is only one upstream and downstream positive eIF-5A PCR fragments for Arabidopsis.
[0421]The specific 5'- and 3'-end primers for the Arabidopsis eIF-5A gene were designed according to the 5' and 3' PCR fragment sequencing results. The full-length Arabidopsis eIF-5A gene was obtained from a PCR reaction utilizing their specific 5'- and 3'-end primers and the corresponding cDNA library as a template. The full-length gene was further confirmed by sequencing. In the end, we cloned one Arabidopsis eIF-5A isoform gene, which was termed senescence-induced eIF-5A.
TABLE-US-00002 T3 and T7 Primers: T3: 5'-ATT AAC CCT CAC TAA AG-3' (SEQ ID NO: 20) T7: 5'-AAT ACG ACT CAC TAT AG-3' (SEQ ID NO: 18) Degenerate Primers for Arabidopsis eIFSA: Forward (upstream) primer: 5'-AAA RRY CGM CCY TGC AAG GT-3' (SEQ ID NO: 17) Reverse (downstream) primer: 5'-TCY TTN CCY TCM KCT AAH CC-3' (SEQ ID NO: 19) Subcloning Arabidopsis antisense full-length senescence-induced eIF-5A into pKYLX71 vector (containing the SAG12 promoter)
[0422]Specific (Homologous) Primers for Arabidopsis senescence-induced eIF-5A, antisense full-length construct: Forward Full-length senescence-induced eIF-5A primer (30-mer):
5'-CCGAGCTCCTGTTACCAAAAAATCTGTACC-3' (SEQ ID NO: 48) (note: underlined portion is the SacI recognition sequence, used for ligating the 5'-end of the PCR fragment into the SacI site in the Multiple Cloning Site (MCS) of pBluescript). Reverse full-length senescence-induced eIF-5A primer (36-mer): 5'-ACCTCGAGCGGCCGCAGAAGAAGTATAAAAACCATC-3' (SEQ ID NO: 49) (note: underlined portion is the NotI recognition sequence, used for ligation into the MCS of pBluescript).
[0423]The orientation of the SacI and NotI sites within the MCS of the pBluescript vector was such that the gene was subcloned in its antisense orientation (i.e. the NotI site is upstream of the SacI site).
Example 23
SAG 12 Promoter was Used to Express the Antisense Senescence-Induced Arabidopsis Full-Length eIF-5A
[0424]Experimental evidence shows that transcription of a set of "senescence-associated genes" or SAGs increases during the onset of senescence (Lohman et al., 1994; Weaver et al., 1998). In fact, senescence appears to begin with the synthesis of new mRNAs and probably down-regulation of other mRNAs, indicating that selective synthesis of proteins is necessary for senescence (Nooden, 1988). That the leaf senescence program is accompanied by changes in gene expression was first demonstrated by Watanabe and Imaseki (1982) using in vitro translation followed by gel electrophoresis to detect changes occurring in translatable mRNA populations. This initial work and subsequent analysis of the in vitro translated proteins revealed the abundance of most mRNAs diminished significantly during the progression of senescence while other translatable mRNAs increased (Watanabe and Imaseki, 1982; Davies and Grierson, 1989; Becker and Apel, 1993; Buchanan-Wollaston, 1994; Smart et al., 1995). Differential screening of cDNA libraries made from mRNAs of senescent leaf tissues also demonstrated that the expression of many genes is down-regulated, whereas the expression of other genes is up-regulated during senescence. SAGs have been identified from a variety of plant species, including Arabidopsis (Hensel et al., 1993; Taylor et al., 1993; Lohman et al., 1994; Oh et al., 1996), asparagus (King et al., 1995), barley (Becker and Apel, 1993), Brassica napus (Buchanan-Wollaston, 1994), maize (Smart et al., 1995), radish (Azumi and Watanabe, 1991) and tomato (Davies and Grierson, 1989; Drake et al., 1996).
[0425]Senescence can be morphologically identified as a characteristically patterned leaf yellowing that begins at the edges of a leaf and reaches the veins last (Weaver et al., 1998). Visible senescence in Arabidopsis thaliana rosette leaves appears approximately 21 days after germination with dramatic upregulation of SAG 12 at the time (Noh an Amasino, 1999). SAG 12 is a gene with the closest specificity for natural senescence and is thus termed a senescence marker. With no detectable expression in young leaves, SAG 12 is induced in older leaves after they are ˜20% yellow but cannot be induced by treatment that does not induce yellowing of leaves (Weaver et al., 1998). Its high degree of specificity for natural senescence can be explained by the fact that the gene product of SAG 12 shows similarity to cysteine proteases and may be involved in protein turnover during senescence (Lohman et al., 1994; Weaver et al., 1998).
[0426]Description of Transgenic Plants
[0427]Transgenic Arabidopsis plants were generated expressing the full-length antisense senescence-induced eIF-5A transgene under the control of the SAG 12 (leaf senescence-specific) promoter, which is activated at the onset of natural leaf senescence, approximately 21 days after germination (Noh and Amasino, 1994), but not in the event of stress-induced senescence. At this point, the transgenic plants express phenotypes characteristic of suppressed full-length senescence-induced eIF-5A expression. Rosette leaves were harvested from 3 to 8-week-old transgenic Arabidopsis antisense full-length senescence-induced eIF-5A plants.
[0428]Methodology for the production of homozygous transgenic antisense senescence-induced eIF-5A Arabidopsis thaliana plants under control of the SAG 12 promoter
[0429]Inserting the SAG 12-antisense-full-length senescence-induced eIF-5A construct in pKYLX71
[0430]First, the plasmid pKYLX71 was cut with EcoRI and HindIII to remove its double 35S promoter, and resultant sticky ends were filled in with Klenow enzyme to create blunt ends. pKYLX71 without the promoter was then ligated to re-circularize the plasmid.
[0431]Secondly, the Arabidopsis SAG 12 promoter was amplified from genomic DNA by PCR using primers containing SalI and XbaI, as described below. This promoter sequence was then inserted into the Multiple Cloning Site (MCS) of pBlueScript using the restriction enzymes SalI and XbaI followed by ligation with T4 DNA ligase.
[0432]The forward SAG 12 Primer was 5'-GGC CGTCGACGATATCTCTTTTTATATTCAAAC-3' (SEQ ID NO: 50) (underlined portion is SalI recognition site, used for ligating the 5'-end of the PCR fragment into the SalI site in the Multiple Cloning Site (MCS) of pBluescript). The Reverse SAG 12 Primer was 5'-CGTCTAGACATTGTTTTAGGAAAGTTAAATGA-3' (SEQ ID NO: 51) (underlined portion is the XbaI recognition site, used for ligating the 5'-end of the PCR fragment into the SacI site in the Multiple Cloning Site (MCS) of pBluescript).
[0433]Thirdly, to create the pBlueScript-SAG 12:antisense-full length-senescence-induced eIF-5A construct, full length senescence-induced eIF-5A was amplified by PCR from the Arabidopsis cDNA library using primers with SacI and NotI restriction sites, as outlined below, and subcloned into the pBluescript-SAG 12 described in the previous paragraph. Note that the orientation of the SacI and NotI sites within the MCS of the pBluescript-SAG 12 vector was such that the gene was subcloned in its antisense orientation (i.e. the NotI site is upstream of the SacI site).
[0434]The forward full-length senescence-induced eIF-5A Primer was 5'-CCGAGCTCCTGTTACCAAAAAATCTGTACC-3' (SEQ ID NO: 48) (note: underlined portion is the SacI recognition sequence, used for ligating the 5'-end of the PCR fragment into the SacI site in the Multiple Cloning Site (MCS) of pBluescript-SAG 12 vector). The reverse Full-length senescence-induced eIF-5A Primer was 5'-ACCTCGAGCGGCCGCAGAAGAAGTATAAAAACCATC-3' (SEQ ID NO: 49) (note: underlined portion is the NotI recognition sequence, used for ligation into the Multiple Cloning Site (MCS) of pBluescript-SAG 12 vector).
[0435]Finally, the desired construct was created in the binary vector, pKYLX71, by digesting pKYLX71 was digested with SacI and XhoI, and also cutting out the SAG 12:full-length senescence-induced eIF-5A cassette from pBluescript with SalI and SacI.
[0436]The XhoI and SalI sticky ends are partially complementary. Hence, these two sets of digested overhangs (specifically, SacI with SacI, and XhoI with SalI) were able to be ligated together with T4 DNA ligase, creating the final construct (SAG 12:antisense-senescence-induced eIF-5A in pKYLX71).
[0437]Transformation and T1 Seed Harvest
[0438]The pKYLX71-SAG 12:antisense-eIF-5A construct was proliferated in E. coli DHα cells, isolated and electroporated into a competent Agrobacterium strain. The bacteria were then used to infiltrate 4.5 week old wildtype Arabidopsis plants and the resulting infiltrated plants were designated as "T0" plants, which were then grown to the end of their life-cycle. Seeds were harvested, collected and designated as T1 seeds. 10 plates of T1 seeds were plated and screened for kanamycin resistance (1/2 MS salt and 50 μg kanamycin/mL) with wildtype as a control; only those seeds containing pKYLX71-SAG 12-antisense-eIF-5A construct survive and grow on kanamycin (K50) media. 24 T1 seedlings were chosen from these plates and placed in soil. The seeds harvested from T1 transgenic plants were labeled as T2 seeds. Each seedling yielded one plant line (#1=1 line containing 1 plant, #2=1 line containing 1 plant, etc.).
[0439]Screening and Identification of Phenotypes
[0440]Once kanamycin resistant T1 seeds were identified, successive generations of T2, T3 and T4 plants were grown. By screening seeds on K50 media, it was possible to distinguish between those plants which inherited the genetic construct and were homozygous for the construct. A phenotypic expression of stunted growth was observed in one T3 plant line when grown in a pot. However, when the same set of seeds was re-grown in identical conditions, the phenotype was not observed.
[0441]From the 24 T1 plants, 4 lines were chosen on the basis of high seed yield (lines T2.14, T2.18, T2.19 and T2.23) and plated on K50 media with wildtype seeds as a control. Approximately 75% of the seeds from each line survived on K50 media and fell into size categories of Small, Medium and Large. From each line, small, medium and large seedlings were removed from plates and planted in soil. Under greenhouse conditions, the Small seedlings did not recover as quickly as their Medium and Large counterparts. At week 6, the Small plants were just beginning to show signs of bolting while the other plants had bolted and flowered. In total, six transgenic T2 plants (from a total of 3 lines×8 plants=96 transgenic plants) demonstrated dramatic delay in bolting and were deemed "Late Bolt" plants. The seed yields of these plants were also dramatically lower than other transgenics.
[0442]From the 96 T2 plants, 3 lines were selected to produce T3 plants (T3.19.S8 and T3.14.L7 which were Late Bolts; and, T3.23.S3 which was not a Late Bolt). When planted on K50 media plates, these lines showed homozygous survival. 13 seedlings were transplanted into pots (10 seedlings per pot). From this set of plants, a dramatic dwarf phenotype was observed in T3.14.L7 plant line. T4 seeds were collected, and lower seed yield was observed in that line. A dense growth (dense silique growth, more branches) phenotype was observed in line T3.19.S8, while a phenotype similar to wildtype was observed in line T3.23.S3. Seed sizes from the 3 transgenic lines were compared but no statistically significant differences were determined. Chlorophyll levels were also analyzed but no statistically significant differences from wildtype control were determined.
[0443]T4 seeds of lines T3.19.S8, T3.14.L7 and T3.23.S3 were screened on K50 to obtain the next generation of plants and showed evidence of inherited gene construct (uniform green growth on plates) compared with wild-type seed that died. However, when planted in individual flats, the dwarf phenotype was not expressed suggesting that the eIF-F5A antisense transgene had been lost. Finally, seeds collected from all T5 plants were screened on K50 plates and showed evidence of kanamycin resistance. Work is now underway to confirm that the antisense transgene has been lost, and these T4 plants are azygous.
[0444]Eight daughter lines were chosen from mother lines T2.14, T2.19 and T2.23 and screened on K50 media with wild-type seeds as a control. Three lines were chosen based on low seed yield: T3.14.L8, T3.14.S8, and T3.23.S1. The other five lines chosen are: T3.18.S7, T3.18.S2, T3.19.S1, T3.19.S5, and T3.23.S6. All the lines screened on K50 media showed homozygous survival, while T3.14.L8, T3.14.S8 and T3.23.S6 showed heterozygous survival. Seedlings from lines T3.14.L8 and T3.14.S8 that survived were white in color with green vascular tissue, while seedlings from T3.23.S6 that survived were entirely dark green in color. These seedlings were selected for transplantation. In total, 28 seedlings from each line were transplanted into cells and grown in greenhouse conditions.
[0445]At week 3, all lines started bolting except for lines T3.14.L8 and T3.23.S1 and several plants within lines T3.18.S7, T3.18.S2, T3.19.S1, T3.19.S5, T3.23.S1 and T3.23.S6. An irregular rosette leaf morphology (elongation of 2nd pair leaves phenotype) was observed in T3.14.L8 and T3.14.S8 lines. At week 5, additional irregular leaf morphologies of increased number of rosette leaves and crinkle-edged rosette leaves phenotypes were also observed in lines T3.18.S7 and T3.23.S6. Rosettes smaller than wild-type were observed in lines T3.23.S1, T3.19.S1, and T3.19.S5. At week 7, spindly stem and no stem elongation phenotypes were observed in lines T3.18.S7, T3.18.S2, T3.19.S1, T3.19.S5, T3.23.S1 and T3.23.S6. The first and second cauline leaf of each plant was collected at week 5 and 6, respectively, for investigation of senescence eIF-5A protein expression.
Example 24
Determination of Oxygen Output
[0446]The leaves were harvested and the areas were measured before they were weighed. The leaves were ground to a fine powder using 1 mL of cold degassed grinding buffer with a mortar and pestle. Then the homogenate was transferred into an eppendorf tube and placed immediately on ice. For tomato leaves, the homogenate isolated required to be filtered through a piece of Miracloth.
[0447]50 μl of homogenate from all samples were added into 10 ml test tubes containing 5 ml grinding buffer and 25 μl DCPIP (2,6-dichlorophenol indophenol). The samples were shaken well and then one set of samples were placed for 15 mins under illumination by a pair of lamps and the second set of samples were placed in the dark for 15 mins. After the 15 minute incubation, 50 μL of DCMU (3-(3,4-dichlorophenyl)-1,1 dimethylurea) was added to both set of samples in order to stop the reaction and then centrifuged in a microcentrifuge for 2 mins at 14,000 g. The absorbencies of the supernatant collected were read at 590 nm using grinding buffer as a blank.
[0448]The molar extinction coefficient for this assay is 16×103, that is, a change in concentration of 1 mole per liter changes the absorbance of the solution by 16×103 μmole of DCPIP reduced/h/ml=(difference in absorbance)×[1/16×103 (moles/l)]×[reaction volume(ml)/103 (ml/l)]×[106 (μmole/mole)]×[60 (min/hr)/reaction time (min)]×[1/sample volume(ml)].
[0449]For every 2 moles of DCPIP that are reduced, 1 mole of O2 is generated. [0450]Reference: Allen J. F. and Holmes N. G., 1986 Electron Transport and Redox Titration s in Photosynthesis: Energy Transduction. Edited by M. F. Hipkins & N. R. Baker., IRL Press, Oxford Pp 107-108.
Example 25
Quantitative Determination of Starch
[0451]Starch content in tomato stems was determined using a method adapted from Lustinec et al. Quantitative determination of starch, amylose, and amylopectin in plant tissues using glass fiber paper. Anal. Biochem. 132:265-271 (1983). Tomato stem tissue was homogenized in three volumes of water using an Omnimixer (12 reps of 5 sec each), followed by a Polytron homogenizer (30 sec). Homogenate was stored in 10 ml aliquots at -20° C. prior to analysis. For analysis, 10 ml homogenate was thawed and mixed with an equal volume of concentrated perchloric acid (HClO4, 70% w/w) and incubated for 20 min at room temperature to dissolve the starch. Simultaneously, several solutions of potato starch (in the range of 0.1-1.0 mg/ml) were processed alongside the tomato stem sample to generate a standard curve. The homogenate (or potato starch standard solution) was stirred and filtered through Whatman GF/A glass microfiber paper (9.0 cm diameter) using a vacuum flask attached to an aspirator. One ml of filtrate was mixed with 3 ml of iodine solution A (8 mM I2, 17 mM KI, 514 mM NaCl) and incubated for 30 min at 4° C. to form a starch-iodine precipitate. The precipitate was collected on Whatman GF/A glass microfiber paper (9.0 cm diameter) using a vacuum flask attached to an aspirator, and then wash the filtrate with the following solutions: once with 10 mL iodine solution B (83 mM I2, 180 mM KI, 8% perchloric [HClO4] acid); once with 5 mL ethanol-NaCl solution (67% ethanol, 342 mM NaCl); twice with 3 ml ethanol-NaOH solution (67% ethanol, 250 mM NaOH). Once ethanol had evaporated, the microfiber paper was removed from aspirator and inserted into screw-capped glass tube. Sulfuric [H2SO4] acid (9 mL of 0.75 M solution) was added to the tube and the tube was incubated in a boiling water bath for 30 min. Three 1 mL-aliquots of eluate were pipetted into glass test tubes and mixed with 1 mL of 5% phenol, quickly followed by 5 mL of concentrated H2SO4. The tubes were vortexed and incubated at room temperature for 30 min to allow the color to develop. Simultaneously, a blank for the spectrophotometer measurements was prepared by mixing 1 mL of 0.75 M H2SO4 with 1 mL of 5% phenol, and quickly adding 5 mL concentrated H2SO4; the blank was also incubated at room temperature for 30 min. A spectrophotometer was calibrated at 480 nm using the blank, and the O.D. of all samples and potato starch standards were measured and recorded. A standard curve was prepared using the potato starch solutions, and used to interpolate the quantity of starch in each sample.
Example 26
[0452]Arabidopsis thaliana (Columbia ecotype) was transformed by the Arabidopsis thaliana sense Senescence-induced eIF-5A (At-eIF) and Tomato sense senescence-induced eIF-5A genes independently. These genes were constitutively expressed in the whole life cycle of the transgenic plants. The inflorescence stems of these plants exhibited a significant increase of xylem development. See FIGS. 89-94.
[0453]The seeds of transgenic and control plants were sown on 1/2 MS medium agar plates, and kept in a growth chamber at 22° C., 80% rh, and 16 h light/day, for 9 days. Then, the seedlings were transferred to 32-well-flats with a commercial soil, and were maintained under the same conditions as above, for 48 days. The main inflorescence stems were selected for microscopic observation. Cross sections were hand-cut from the base of the stems within 2 mm above the rosette. The sections were stained with the phloroglucinol-HCl method. We found that the stem xylem at this age has achieved its maximum development. A comparison was made between transgenic and control plants in the sizes (sectional areas) of xylem. In addition, measurements were done for phloem and pith in both transgenic and control plants.
[0454]Measurement of tissue areas was as follows. Cross sections were photographed with a Zeiss microscope, and the micrographs were digitalized using Photoshop®. These images were printed out on paper and different tissues were cut out, and their areas were measured by an area-measuring meter. To calculate the actual area of each tissue, the following formula was used: The actual area=(The area of an individual tissue on paper)/(Magnification)2
[0455]It thus appears that senescence-induced eIF-5A is also involved in programmed cell death associated with xylogenesis. Constitutive antisense suppression of senescence-induced AteIF-5A in Arabidopsis reduced the thickness of the inflorescence stem as well as the number of xylem cell layers. By contrast, the inflorescence stems of plants in which Arabidopsis or tomato senescence-induced eIF-5A was constitutively over-expressed were, on average, 1.7-fold thicker than those of corresponding wild-type plants, and the total xylem area per cross-section of inflorescence stem was 2 fold higher. The over-expressing transgenic plants also had greatly increased rosette leaf biomass and grew faster than wild-type plants, which may reflect enhanced nutrient uptake. The same phenotype was observed when the senescence-induced isoform of eIF-5A from tomato was over-expressed in Arabidopsis plants. These results collectively indicate that the senescence-induced isoform of eIF-5A not only regulates leaf and flower senescence, but is also involved in xylogenesis.
Example 27
Suppression of Deoxyhypusine Synthase Delays Browning of Pre-Packaged Cut Lettuce in Ambient Atmosphere
[0456]The lettuce was grown hydroponically, as this was an ideal for obtaining a controlled and continuous induction of the transgene (antisense DHS) by the glucocorticoid-inducible promoter. The inducer, dexamethasone, was applied directly to the hydroponic solution.
[0457]FIG. 149(a) shows 6 week old plants immediately prior to harvest. FIG. 149 (b) shows a view of air stone inside the pot. FIG. 149(c) shows the lid with three openings to support plants, and an airline in the center. FIG. 150 shows browning of cut lettuce in ambient atmosphere.
[0458]FIG. 150(a) shows the onset of browning plotted as a factor of time for three transgenic lines: Line 13 (n=4), Line 21 (n=5) and Line 34 (n=7), expressing the 3' UTR of lettuce DHS in the antisense orientation, as well as for wildtype romaine lettuce (Green Towers variety; n=12). The transgene was expressed under the control of a glucocorticoid-inducible promoter. The lettuce heads were grown hydroponically, and were harvested and chopped into small segments. The chopped lettuce was maintained in perforated zipped bags at 7.5° C. for 6 days. Browning of cut lettuce was evaluated on a scale of 0 to 4, where a rating of 1 was defined as the onset of browning. Standard error bars are shown. FIG. 150(b) shows the results of two individual plants of Line 21 that showed 0% browning through the 6-day study. FIG. 150(c) shows the result of one individual plant of line 34 that also did not turn brown over the same period, where as the wild type lettuce turned brown.
[0459]FIG. 151 shows a Western blot of transgenic lettuce showing decreased expression of DHS protein. Total protein from four individual lettuce plants expressing the antisense 3'UTR of lettuce DHS, as well as one plant that had lost the transgene as the gene segregated in the T2 generation ("null"), and one wild type plant, was isolated and 20 μg of each sample was loaded on a 14% acrylamide gel stained with Coomassie blue. The gel was blotted onto a PVDF membrane and probed with a polyclonal antibody raised against tomato DHS. The blot was developed colormetrically, an shows that the expression of DHS in the transgenic plants was reduced relative to the null and wild type plants.
Example 28
Suppression of Deoxyhypusine Synthase Expression in Canola Increases Seed Yield
[0460]Deoxyhypusine synthase (DHS) mediates the first of two enzymatic reactions that convert inactive eukaryotic translation initiation factor-5A (eIF-5A) to an activated form able to facilitate translation. A full-length cDNA clone encoding canola (Brassica napus cv Westar) DHS was isolated from a cDNA expression library prepared from senescing leaves. DHS was suppressed in transgenic canola plants by expressing the antisense 3'-UTR of canola DHS cDNA under the regulation of the constitutive cauliflower mosaic virus (CaMV-35S) promoter. See FIG. 158 showing decreased expression of DHS protein. The transgenic canola plants were obtained by vacuum infiltration of canola inflorescences by modifying the protocol developed for Arabidopsis. The efficiency of transformation was enhanced by removal of new flowers that formed after the plants had been vacuum-infiltrated, and transformation rates of 50 to 60% were routinely obtained. Transgenic plants had reduced levels of leaf DHS protein and exhibited delayed natural leaf senescence. Suppression of DHS also increased leaf size by 1.5- to 2-fold (see FIG. 157) and resulted in increases in seed yield of up to 65% (see FIG. 160. This was attributable in part to an increase in the size of the siliques, which were on average 18% to 26% longer than wild-type siliques depending on the line (see FIG. 159). When wild-type and transgenic plants were grown in 6-inch pots, the increase in seed yield accruing from suppression of DHS was ˜4.5-fold greater than when the plants were grown in 12 inch pots. Thus suppression of DHS appears to ameliorate the effects of sub-lethal stress engendered by growth in small containers. The increase in seed yield for transgenic plants translates into a corresponding increase in seed oil content based on measurements of triacylglycerol, and there was no change in the fatty acid composition of the oil in transgenic seeds. See FIG. 161(a) and (b).
Example 29
Extending the Vase Life of Carnation Flowers by Administering Inhibitors of Deoxyhypusine Synthase and by Antisense Suppression of Deoxyhypusine Synthase
[0461]A full-length cDNA clone (AF296079) encoding deoxyhypusine synthase (DHS) was isolated from carnation petals. DHS mediates the first of two enzymatic reactions that convert inactive eukaryotic translation initiation factor-5A (eIF-5A) to an activated form able to facilitate translation. Northern analysis revealed that DHS expression is correlated with senescence of carnation flower petals. Treatment of cut carnation flowers with inhibitors of the DHS reaction, including diaminobutane (putrescine), diaminopropane, diaminohexane, diaminooctane and spermidine, extended the vase life of the flowers by up to 83%. In order to evaluate the role of DHS in carnation flower senescence more definitively, expression of the protein was suppressed in transgenic plants by introducing the antisense 3'-UTR of carnation DHS cDNA under regulation of the constitutive cauliflower mosaic virus promoter through Agrobacterium transformation. Three lines of transgenic flowers with reduced DHS expression were analyzed and found to have longer vase-life relative to wild-type flowers. Indeed, one of the lines exhibited an increase in vase life of >100%. These findings indicate that DHS plays a central role in flower senescence.
Example 30
The Delayed Bolting Phenotype Induced by Suppression of Deoxyhypusine Synthase in Arabidopsis can be Rescued by Treatment with Ga3
[0462]Deoxyhypusine synthase (DHS) is a ubiquitous enzyme required for post-translational activation of eukaryotic translation initiation factor 5A (eIF-5A) and appears to be essential for normal plant growth and development. DHS was suppressed in Arabidopsis by expressing full-length antisense Arabidopsis DHS cDNA in transgenic plants under the regulation of the senescence-specific SAG12 promoter. Plants expressing the transgene had reduced levels of leaf DHS protein, and exhibited delayed bolting and a pronounced delay (2 to 5 weeks) in the onset of leaf senescence. The bolts were also shorter, although this did not result in a reduction in biomass or seed yield. Treatment of the transgenic plants with GA3 reversed the delayed bolting phenotype. A similar phenotype was obtained by antisense suppression of DHS under the regulation of GCI, a glucacorticoid-inducible promoter that can be activated by administering dexamethasone (DEX). Again, administering GA3 rescued this phenotype; that is, the GA3-treated transgenic plants bolted normally, the bolts were of normal size and there was no delay in the onset of leaf senescence. These results collectively indicate that DHS, through activation of one or more of the three isoforms of eIF-5A in Arabidopsis, influences GA metabolism.
Example 31
Transformation of Arabidopsis Plants with Sense Growth eIF-5A from Cottonwood to Increase Biomass
[0463]4 week-old-Arabidopsis plants were infiltrated with Agrobacterium tumefaciens strain GV3101 containing pKYLX71-sense-poplar Growth-eIF-5A3 (see FIG. 171).
[0464]Seeds of the first generation (T1) of transgenic as well as control plants (transformed with the empty pKYLX71 vector only, conferring kanamycin resistance) were sown on 1/2 MS medium agar plates supplemented with kanamycin. The plates were kept in a growth chamber at 22° C., 80% rh, and 16 h light/day, for 9 days. Then, the seedlings were transferred to 32-well-flats with a commercial soil mix, and were maintained under the same conditions as above, until T2 seeds were harvested. Different lines of T2 seeds as well as control plants were screened on 1/2 MS medium agar plates again. Seedlings were transferred into soil and kept under the same condition as for T1 plants.
[0465]Transgenic Line 3, which had the best phenotype, was chosen for this experiment. eIF5A protein was confirmed to be over-expressed by Western analysis. Transgenic Line 3 plants displayed a 1.5-fold increase in inflorescence stem height relative to vector-only control plants at the 6th week, and produced 4-fold more seed yield than the control plants did.
[0466]Seeds from transgenic line 3 were sown on plates with 1/2 MS medium (supplemented with kanamycin) as for screening transgenic seedlings harboring over-expressed poplar eIF5A3.
[0467]1-week old seedlings were transferred to soil, Sunshine LC1/LG3, 1:1 (basic soil, no added fertilizer). See Wang et al. (2003) Plant Mol Biol 53: 1223-1235 for a discussion on the soil analysis for this brand of soil, as compared with a richer soil brand that is routinely used (Promix BX)
[0468]When watering is needed (that is about 3 days later), a series of fertilizer (20:20:20) at 0 g/L, 0.5 g/L, 1 g/L, 3 g/L and 5 g/L was applied (to separate flats of plants) until the soil was saturated. The recommended amount of fertilizer for optimal growth is 1 g/L. Fertilizer was reapplied once per week.
[0469]Morphology and phenotype were observed for leaf size and color, bolting time and inflorescence stem height, silique size and number, etc. The resulting transgenic plants have increased biomass on optimal, suboptimal and supraoptimal concentrations of fertilizer. See FIGS. 172 and 173.
Example 32
Transformation of Arabidopsis Plants with Mutant Senescence eIF-5A in Sense Orientation to Increase Biomass
[0470]Arabidopsis plants were transformed with a mutant senescence eIF-5A as described above (not able to be hypusinated). The resulting transgenic plants have increased biomass on optimal, suboptimal and supraoptimal concentrations of fertilizer. See FIGS. 174 and 175.
REFERENCES
[0471]Azumi, Y and Watanabe, A (1991) Evidence for a senescence-associated gene induced by darkness. Plant Physiol 95: 577-583. [0472]Becker W, Apel K (1993) Differences in gene expression between natural and artificially induced leaf senescence. Planta 189: 74-79 [0473]Bevec, D., Jaksche, H., Oft, M., Wohl, T., Himmelspach, M., Pacher, A., Schebesta, M., Koettnitz, K., Dobrovnik, M., Csonga, R., Lottspeich, F., and Hauber, J (1996) Inhibition of HIV-1 replication in lymphocytes by mutants of the Rev cofactor of eIF-5A. Science 271:1858-1860. [0474]Bevec, D, and Hauber, J (1997) Eukaryotic initiation factor 5A activity and HIV-1 Rev function. Biol Signals 6:124-133. [0475]Bleecker, A. B., and Patterson, S E (1997) Last exit: Senescence, abscission, and meristem arrest in Arabidopsis. Plant Cell. 9:1169-1179. [0476]Buchanan-Wollaston, V (1997) The molecular biology of leaf senescence. J Exp Bot 48:181-191. [0477]Buchanan-Wollaston V (1994) Isolation of cDNA clones for genes that are expressed during leaf senescence in Brassica napus. Plant Physiol 105: 839-846 [0478]Chen, K Y, and Liu, A Y C. (1997) Biochemistry and function of hypusine formation on eukaryotic initiation factor 5A. Biol Signals 6:105-109. [0479]Davies K M, and Grierson D (1989) Identification of cDNA clones for tomato (Lycopersicon esculentum Mill.) mRNAs that accumulate during fruit ripening and leaf senescence in response to ethylene. Plant Cell 179: 73-80. [0480]Drake R, John I, Farrell A, Cooper W, Schuch W, and Grierson D (1996) Isolation and analysis of cDNAs encoding tomato cysteine proteases expressed during leaf senescence. Plant Mol Biol 30: 755-767 [0481]Gan, S and Amasino, R M (1997). Molecular genetic regulation and manipulation of leaf senescence. Plant Physiol 113: 313-319. [0482]Gan, S and Amasino, R M (1995) Inhibition of leaf senescence by autoregulated production of cytokinin. Science 270: 1986-1988. [0483]Hanauske-Abel, H. M., Park, M. H., Hanauske, A.-R., Popowicz, A. M., Lalande, M., and Folk, J. E. Inhibition of G1-S transition by inhibitors of deoxyhypusine hydroxylation. Biochem Biophys Acta 1221: 115-124, 1994. [0484]Henderson, B R, and Percipalle, P. (1997) Interactions between HIV Rev and nuclear import and export factors: the Rev nuclear localization signal mediate specific binding to human importin-beta. J Mol Biol 274: 693-707. [0485]Hensel, L. L., V. Grbic, D. B. Baumgarten, and A. B. Bleecker. (1993). Developmental and age-related processes that influence the longevity and senescence of photosynthetic tissues in Arabidopsis. Plant Cell 5:553-564. [0486]Jakus, J., Wolff, E. C., Park, M. H., and Folk, J. E. Features of the spermidine-binding site of deoxyhypusine synthase as derived from inhibition studies: effective inhibition by bis- and mono-guanylated diamines and polyamines. J Biol Chem 268:13151-13159, 1993. [0487]Kang, H A, and Hershey, J W B (1994) Effect of initiation factor eIF-5A depletion on protein synthesis and proliferation of Saccharomyces cerevisiae. J Biol Chem 269: 3934-3940. [0488]Katahira, J, Ishizaki, T, Sakai, H, Adachi, A, Yamamoto, K, and Shida, H. (1995) Effects of translation initiation factor eIF-5A on the functioning of human T-cell leukemia virus type I Rex and human immunodeficiency virus Rev inhibited trans dominantly by a Rex mutant deficient in RNA binding. J Virol 69: 3125-3133 [0489]Kemper, W M, Berry, K W, and Merrick, W C (1976) Purification and properties of rabbit reticulocyte protein synthesis initiation factors M2Bα and M2Bβ. J Biol Chem 251: 5551-5557. [0490]Lohman, K N, Gan, S, John, M C and Amasino, R (1994) Molecular analysis of natural leaf senescence in Arabidopsis thaliana. Phys Plant 92: 322:328. [0491]Lipowsky, G., Bischoff, F. R., Schwarzmaier, P, Kraft, R., Kostka, S., Hartmann, E., Kutay, U., and Gorlich, D. (2000) Exportin 4: a mediator of a novel nuclear export pathway in higher eukaryotes. EMBO J. 19:4362-4371. [0492]Liu, Y P, Nemeroff, M, Yan, Y P, and Chen, K Y. (1997) Interaction of eukaryotic initiation facto 5A with the human immunodeficiency virus type 1 Rev response element RNA and U6 snRNA requires deoxyhypusine or hypusine modification. Biol Signals 6:166-174. [0493]Martinez-Zapater, J M and Salinas, J (1994) Arabidopsis Protocols. Humana Press: p. 197. [0494]Mattaj, I. W., and Englmeier, L. (1998) Nucleocytoplasmic transport: The soluble phase. Annu Rev Biochem 67: 265-306 [0495]Mehta, A M, Saftner, R A, Mehta, R A, and Davies, P J (1994) Identification of posttranslationally modified 18-kilodalton protein from rice as eukaryotic translation initiation factor 5A. Plant Physiol 106:1413-1419. [0496]Noh, Y-S and Amasino, R (1999) Identification of a promoter region responsible for the senescence-specific expression of SAG12. Plant Mol Biol 41:181-194. [0497]Nooden, L D, Guaimet, J J and John, I (1997) Senescence mechanisms. Physiol Plant 101: 746-753. [0498]Nooden, L D and Leopold, A C (eds) (1988) The phenomena of senescence and aging. Senescence and Aging in Plants. Academic Press: pp. 1-50. [0499]Ober, D and Hartmann, T (1999) Deoxyhypusine synthase from tobacco. J Biol Chem 274: 32040-32047. [0500]Oh S A, Lee S Y, Chung I K, Lee C H, and Nam H G (1996) A senescence-associated gene of Arabidopsis thaliana is distinctively regulated during natural and artificially induced leaf senescence. Plant Mol Biol 30: 739-754 [0501]Page, D R and Grossniklaus, U (2002) The art and design of genetic screens: Arabidopsis Thaliana. Nature Reviews Genetics 3:124-136 [0502]Park, M H, Joe, Y A, and Kang K R (1998) Deoxyhypusine synthase activity is essential for cell viability in the yeast Saccharomyces cerevisiae. J Biol Chem 16:1677-1683. [0503]Park, M H, Wolff, E C, Lee, Y B and Folk, J E (1994) Antiproliferative effects of inhibitors of deoxyhypusine synthase: inhibition of growth of Chinese hamster ovary cells by guanyl diamines. J Biol Chem 269:27827-27832. [0504]Park, M. H., Wolff E. C., and Folk J. E. (1993) Hypusine: its post-translational formation in eukaryotic initiation factor 5A and its potential role in cellular regulation. BioFactors 4:95-104. [0505]Park, M H, Wolff, E C and Folk, J E (1993) Is hypusine essential for eukaryotic cell proliferation? Trends Biochem Sci 18:475-479. [0506]Park, M H, Wolff, E C, Smit-McBride, Z, Hershey, J W B and Folk, J E (1991) Comparison of the activities of variant forms of eIF-4D: the requirement for hypusine or deoxyhypusine. J Biol Chem 266:7988-7994, 1991. [0507]Park, M H and Wolff, E C (1988) Cell-free synthesis of deoxyhypusine. J Biol Chem 263:15264-15269. [0508]Quirino, BF, Noh, Y-S, Himelblau, E and Amasino, R (2000) Molecular aspects of leaf senescence. Trends Plant Sci 5:278-282. [0509]Rosorius, O., Reichart, B., Kratzer, F., Heger, P., Dabauvalle, M. C. & Hauber, J. (1999) Nuclear pore localization & nucleocytoplasmic transport of eIF-5A: evidence for direct interaction with the export receptor CRM1. J. Cell Sci. 112, 2369-2380. [0510]Ruhl, M., Himmelspach, M., Bahr, G. M., Hammerschmid, F., Jaschke, H., Wolff, B., Aschauer, H., Farrington, G. K., Probst, H., Bevec, D., and Hauber, J. (1993) Eukaryotic initiation factor 5A is a cellular target of the human immunodeficiency virus type 1 Rev activation domain mediating transactivation. J Cell Biol 123:1309-1320. [0511]Schardl, C L, Byrd, A D, Bension, G, Altschuler, M S, Hildebrand, D F and Hunt, A G (1987) Design and construction of a versatile system for the expression of foreign genes in plants. Genes 61: 1-11. [0512]Schnier J, Schwelberger H G, Smit-McBride Z, Kang H A and Hershey J W B (1991) Translation initiation factor 5A and its hypusine modification are essential for cell viability in the yeast Saccharomyces cerevisiae. Mol Cell Biol 11:3105-3114 [0513]Smart C M, Hosken S E, Thomas H, Greaves J A, Blair B G, Schuch W (1995) The timing of maize leaf senescence and characterization of senescence-related cDNAs. Physiol Plant 93: 673-682 [0514]Smart, C M (1994) Gene expression during leaf senescence. New Phytologist 126:419-448. [0515]Taylor C B, Bariola P A, Delcardayre S B, Raines R T, Green R T (1993) RNS2: A senescence-associated RNase of Arabidopsis that diverged from the S-RNases before speciation. Proc Natl Acad Sci USA 90: 5118-5122 [0516]Tome, M., Fiser, S. M., Payne, C. M., and Gerner, E W. (1997) Excess putrescine accumulation inhibits the formation of modified eukaryotic initiation factor 5A (eIF-5A) and induces apoptosis. Biochem. J. 328: 847-854. [0517]Tome, M., and Gerner, E W. (1997) Cellular eukaryotic initation factor 5A content as a mediator of polyamine effects on growth and apoptosis. Biol Signals 6:150-156. [0518]Walbot V (2000) A green chapter in the book of life. Nature 408: 794-795. [0519]Wang, T-W, Lu, L, Wang, D, and Thompson, J E (2001) Isolation and characterization of senescence-induced cDNAs encoding deoxyhypusine synthase and eucaryotic translation initiation factor 5A from tomato. J Biol Chem 276:17541-17549. [0520]Watanabe, A and Imaseki, H (1982) Changes in translatable mRNA in senescing wheat leaves. Plant Cell Physiol 23:489-497. [0521]Weaver, L. M., Gan, S, Quirino, B and Amasino, R (1998) A comparison of the expression patterns of several senescence-associated genes in response to stress and hormone treatment. Plant Mol Biol 37:455-469. [0522]Xu, A. and Chen, K. Y. (2001) Hypusine is required for a sequence-specific interaction of eukaryotic initiation factor 5A with post-SELEX RNA. J. Biol. Chem. 276:2555-2561.
[0523]Zuk, D., and Jacobson, A. (1998) A single amino acid substitution in yeast eIF-5A results in mRNA stabilization. EMBO J. 17:2914-2925. [0524]Boyes, D. C., A. M. Zayed, R. Ascenzi, A. J. McCaskill, N. E. Hoffman, K. R. Davis, and J. Goerlach. 2001. Growth stage-based phenotypic analysis of Arabidopsis: A model for high throughput functional genomics in plants. Plant Cell, 13: 1499-1510. [0525]Collawn, J. F., and Y. Patterson. 1999. Production of antipeptide antibodies. In F M Ausubel, R Brent, R E Kingston, D D Moore, J A Smith, J G Seidman, K Struhl, eds, Current Protocols in Molecular Biology on CD. John Wiley & Sons, New York. [0526]Chamot, D. and C. Kuhlemeier. 1992. Differential expression of genes encoding the hypusine-containing translation initiation factor, eIF-5A, in tobacco. Nucleic Acids Res 20: 665-669. [0527]Clemens, M. J., and U.-A. Bommer 1999. Translational control: The cancer connection. Int. J. of Biochem. Cell Biol. 31: 1-23. [0528]Davis, L. G., M. D. Dibner, and J. B. Battey. 1986. Basic methods in molecular biology. Elsevier Science Publishing Co., Inc, New York, pp. 130-5. [0529]Drenckhahn, D., T. Jons, and F. Schmitz. 1993. Production of polyclonal antibodies against proteins and peptides. In D J Asai, ed, Methods in Cell Biology, Vol 37. Academic Press, New York, pp 7-56. [0530]Dresselhaus, T., C. Simone, and H. Lorz. 1999. A transcript encoding translation factor eIF-5A is stored in unfertilized egg cells of maize. Plant Mol. Biol. 39: 1063-1071. Fagard, M. and H. Vaucheret. 2000. (Trans)Gene silencing in plants: How many mechanisms? Annu. Rev. Plant Physiol. Mol. Biol. 51: 167-94. [0531]Fairbanks, G., T. L. Steck, and D. F. H. Wallach. 1971. Coomassie blue R250 used in isopropanol-acetic acid. Biochem 10: 2606-2618. [0532]Ghosh, S., S. Gepstein, J. J. Heikkila, and E. B. Dumbroff. 1988. Use of a scanning densitometer or an ELISA plate reader for measurement of nanogram amounts of protein in crude extracts from biological tissues. Anal. Biochem. 169: 227-233. [0533]Jao, D. L.-E., and K. Y. Chen. 2002. Subcellular localization of the hypusin-containing eukaryotic initiation factor 5A by immunofluorescent staining and green fluorescent protein tagging. J. Cell. Biochem. 86: 590-600. [0534]Jakus, J., E. C. Wolff, M. H. Park, and J. E. Folk. 1993. Features of the spermidine-binding site of deoxyhypusine synthase as derived from inhibition studies: effective inhibition by bis- and mono-guanylated diamines and polyamines. J. Biol. Chem. 268, 13151-13159. [0535]Kushner, S. R. 1978. An improved method for transformation of Escherichia coli with ColE1 derived plasmids. In: Genetic Engineering: Proceedings of the International Symposium on Genetic Engineering (H. W. Boyer and S, Nicosia, eds.), Elsevier/North-Holland Press, New York. Pp. 17-23. [0536]Liu, Y. P., M. Nemeroff, Y. P. Yan, and K. Y. Chen. 1997. Interaction of eukaryotic initiation factor 5A with the human immunodeficiency virus type 1 Rev response element RNA and U6 snRNA requires deoxyhypusine or hypusine modification. Biol. Signals 6 (3), 166-74. [0537]Matile, P., S. Hoertensteiner, and H. Thomas. 1999. Chlorophyll degradation. Annu. Rev. Plant Physiol. Plant Mol. Biol. 50: 67-95. [0538]McCabe, M. S, L. C. Garratt, F. Schepers, W. J. R. M. Jordi, G. M. Stoopen, E. Davelaar, J. H. A. van Rhijn, J. B. Power, and M. R. Davey. 2001. Effects of P.sub.SAG12-IPT gene expression on development and senescence in transgenic lettuce. Plant Physiol. 127: 505-516. [0539]Miranda, P. V., A. Brandelli, and J. G. Tezon. 1993. Instantaneous blocking for immunoblots. Anal. Biochem. 209: 376-377. [0540]Murashige, T. and F. Skoog. 1962. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 15:473-497. [0541]Nowack, L. N. 2002 personal communications. [0542]Ober, D., and T. Hartmann. 1999. Deoxyhypusine synthase from tobacco. J. Biol. Chem. 274(45): 32040-32047. [0543]Page, T., G. Griffiths, and V. Buchanan-Wollaston. 2001. Molecular and biochemical characterization of postharvest senescence in broccoli. Plant Physiol. 125: 718-727. [0544]Park, J.-H., S. A. Oh, Y. H. Kim, H. R. Woo, and H. G. Nam. 1998. Differential expression of senescence-associated mRNAs during leaf senescence induced by different senescence-inducing factors in Arabidopsis. Plant Mol. Biol. 37: 445-454. [0545]Park, M. H., Y. B. Lee, and Y. A. Joe. 1997. Hypusine is essential for eukaryotic cell proliferation. Biol. Signals 6, 115-123. [0546]Park, M. H., E. C. Wolff, and J. E. Folk. 1993. Hypusine: its post-translational formation in eukaryotic initiation factor 5A and its potential role in cellular regulation. BioFactors 4 (2), 95-104. [0547]Park, M. H., and E. C. Wolff 1988. Cell-free synthesis of deoxyhypusine. J. Biol. Chem. 263(30): 15264-15269. [0548]Rosorius, O., B. Reicher, F. Kraetzer, P. Heger, M.-C. Dabauvalle, and J. Hauber. 1999. Nuclear pore localization and nucleocytoplasmic transport of eIF-5A: evidence for direct interaction with the export receptor CRM1. J. Cell Sci. 112: 2369-2380. Stotz 2000. [0549]Tome, M. E., and E. W. Gerner. 1997. Cellular eukaryotic initiation factor 5A content as a mediator of polyamine effects of growth and apoptosis. Biol. Signals 6, 150-156. [0550]Wang, C. Y. and J. E. Baker. 1980. Extending vase life of carnations with minooxyacetic acid, polyamines, EDU, and CCCP. HortSci. 15, 805-806. [0551]Wang, T.-W., L. Lu, C.-G. Zhang, C. A. Taylor, and J. E. Thompson. Unpublished a. Suppression of deoxyhypusine synthase expression in Arabidopsis thaliana delays leaf senescence. [0552]Wang, T.-W., W. Wu, C. Taylor, and J. E. Thompson. Unpublished b. Characterization of eukaryotic translation initiation factor-5A cDNA isoforms from tomato. [0553]Wang, T.-W., C.-G. Zhang, L. Lu, L. N. Nowack, and J. E. Thompson. Unpublished c. Extending vase life of carnation by deoxyhypusine synthase inhibitors and transient transfection with antisense deoxyhypusine synthase. [0554]Wang, T.-W., L. Lu, D. Wang, and J. E. Thompson. 2001. Isolation and characterization of senescence-induced cDNAs ecoding deoxyhypusine synthase and eucaryotic translation initiation factor 5A from tomato. J. Biol. Chem. 276(20): 17541-17549. [0555]Zuk, D., and A. Jacobson. 1998. A single amino acid substitution in yeast eIF-5A results in mRNA stabilization. EMBO J. 17(10): 2914-2925.
Sequence CWU
1
21111584DNALycopersicon esculentumCDS(54)..(1196) 1cgcagaaact cgcggcggca
gtcttgttcc gtacataatc ttggtctgca ata atg 56
Met
1gga gaa gct ctg aag tac agt atc atg gac tca gta
aga tcg gta gtt 104Gly Glu Ala Leu Lys Tyr Ser Ile Met Asp Ser Val
Arg Ser Val Val 5 10
15ttc aaa gaa tcc gaa aat cta gaa ggt tct tgc act aaa atc gag ggc
152Phe Lys Glu Ser Glu Asn Leu Glu Gly Ser Cys Thr Lys Ile Glu Gly
20 25 30tac gac ttc aat aaa ggc gtt
aac tat gct gag ctg atc aag tcc atg 200Tyr Asp Phe Asn Lys Gly Val
Asn Tyr Ala Glu Leu Ile Lys Ser Met 35 40
45gtt tcc act ggt ttc caa gca tct aat ctt ggt gac gcc att gca
att 248Val Ser Thr Gly Phe Gln Ala Ser Asn Leu Gly Asp Ala Ile Ala
Ile 50 55 60 gtt
aat caa atg cta gat tgg agg ctt tca cat gag ctg ccc acg gag 296Val
Asn Gln Met Leu Asp Trp Arg Leu Ser His Glu Leu Pro Thr Glu65
70 75 80gat tgc agt gaa gaa gaa
aga gat gtt gca tac aga gag tcg gta acc 344Asp Cys Ser Glu Glu Glu
Arg Asp Val Ala Tyr Arg Glu Ser Val Thr 85
90 95tgc aaa atc ttc ttg ggg ttc act tca aac ctt gtt
tct tct ggt gtt 392Cys Lys Ile Phe Leu Gly Phe Thr Ser Asn Leu Val
Ser Ser Gly Val 100 105 110aga
gac act gtc cgc tac ctt gtt cag cac cgg atg gtt gat gtt gtg 440Arg
Asp Thr Val Arg Tyr Leu Val Gln His Arg Met Val Asp Val Val 115
120 125gtt act aca gct ggt ggt att gaa gag
gat ctc ata aag tgc ctc gca 488Val Thr Thr Ala Gly Gly Ile Glu Glu
Asp Leu Ile Lys Cys Leu Ala 130 135
140 cca acc tac aag ggg gac ttc tct tta cct gga gct tct
cta cga tcg 536Pro Thr Tyr Lys Gly Asp Phe Ser Leu Pro Gly Ala Ser
Leu Arg Ser145 150 155
160aaa gga ttg aac cgt att ggt aac tta ttg gtt cct aat gac aac tac
584Lys Gly Leu Asn Arg Ile Gly Asn Leu Leu Val Pro Asn Asp Asn Tyr
165 170 175tgc aaa ttt gag aat
tgg atc atc cca gtt ttt gac caa atg tat gag 632Cys Lys Phe Glu Asn
Trp Ile Ile Pro Val Phe Asp Gln Met Tyr Glu 180
185 190gag cag att aat gag aag gtt cta tgg aca cca tct
aaa gtc att gct 680Glu Gln Ile Asn Glu Lys Val Leu Trp Thr Pro Ser
Lys Val Ile Ala 195 200 205cgt ctg
ggt aaa gaa att aat gat gaa acc tca tac ttg tat tgg gct 728Arg Leu
Gly Lys Glu Ile Asn Asp Glu Thr Ser Tyr Leu Tyr Trp Ala 210
215 220 tac aag aac cgg att cct gtc
ttc tgt cct ggc ttg acg gat gga tca 776Tyr Lys Asn Arg Ile Pro Val
Phe Cys Pro Gly Leu Thr Asp Gly Ser225 230
235 240ctt ggt gac atg cta tac ttc cat tct ttc aaa aag
ggt gat cca gat 824Leu Gly Asp Met Leu Tyr Phe His Ser Phe Lys Lys
Gly Asp Pro Asp 245 250
255aat cca gat ctt aat cct ggt cta gtc ata gac att gta gga gat att
872Asn Pro Asp Leu Asn Pro Gly Leu Val Ile Asp Ile Val Gly Asp Ile
260 265 270agg gcc atg aat ggt gaa
gct gtc cat gct ggt ttg agg aag aca gga 920Arg Ala Met Asn Gly Glu
Ala Val His Ala Gly Leu Arg Lys Thr Gly 275 280
285atg att ata ctg ggt gga ggg ctg cct aag cac cat gtt tgc
aat gcc 968Met Ile Ile Leu Gly Gly Gly Leu Pro Lys His His Val Cys
Asn Ala 290 295 300
aat atg atg cgc aat ggt gca gat ttt gcc gtc ttc att aac acc gca
1016Asn Met Met Arg Asn Gly Ala Asp Phe Ala Val Phe Ile Asn Thr Ala305
310 315 320caa gag ttt gat
ggt agt gac tct ggt gcc cgt cct gat gaa gct gta 1064Gln Glu Phe Asp
Gly Ser Asp Ser Gly Ala Arg Pro Asp Glu Ala Val 325
330 335tca tgg gga aag ata cgt ggt ggt gcc aag
act gtg aag gtg cat tgt 1112Ser Trp Gly Lys Ile Arg Gly Gly Ala Lys
Thr Val Lys Val His Cys 340 345
350gat gca acc att gca ttt ccc ata tta gta gct gag aca ttt gca gct
1160Asp Ala Thr Ile Ala Phe Pro Ile Leu Val Ala Glu Thr Phe Ala Ala
355 360 365aag agt aag gaa ttc tcc cag
ata agg tgc caa gtt tgaacattga 1206Lys Ser Lys Glu Phe Ser Gln
Ile Arg Cys Gln Val 370 375
380ggaagctgtc cttccgacca cacatatgaa ttgctagctt ttgaagccaa cttgctagtg
1266tgcagcacca tttattctgc aaaactgact agagagcagg gtatattcct ctaccccgag
1326ttagacgaca tcctgtatgg ttcaaattaa ttatttttct ccccttcaca ccatgttatt
1386tagttctctt cctcttcgaa agtgaagagc ttagatgttc ataggttttg aattatgttg
1446gaggttggtg ataactgact agtcctctta ccatatagat aatgtatcct tgtactatga
1506gattttgggt gtgtttgata ccaaggaaaa atgtttattt ggaaaacaat tggattttta
1566atttattttt tcttgttt
15842381PRTLycopersicon esculentum 2Met Gly Glu Ala Leu Lys Tyr Ser Ile
Met Asp Ser Val Arg Ser Val1 5 10
15Val Phe Lys Glu Ser Glu Asn Leu Glu Gly Ser Cys Thr Lys Ile
Glu 20 25 30Gly Tyr Asp Phe
Asn Lys Gly Val Asn Tyr Ala Glu Leu Ile Lys Ser 35
40 45Met Val Ser Thr Gly Phe Gln Ala Ser Asn Leu Gly
Asp Ala Ile Ala 50 55 60Ile Val Asn
Gln Met Leu Asp Trp Arg Leu Ser His Glu Leu Pro Thr65 70
75 80Glu Asp Cys Ser Glu Glu Glu Arg
Asp Val Ala Tyr Arg Glu Ser Val 85 90
95Thr Cys Lys Ile Phe Leu Gly Phe Thr Ser Asn Leu Val Ser
Ser Gly 100 105 110Val Arg Asp
Thr Val Arg Tyr Leu Val Gln His Arg Met Val Asp Val 115
120 125Val Val Thr Thr Ala Gly Gly Ile Glu Glu Asp
Leu Ile Lys Cys Leu 130 135 140Ala Pro
Thr Tyr Lys Gly Asp Phe Ser Leu Pro Gly Ala Ser Leu Arg145
150 155 160Ser Lys Gly Leu Asn Arg Ile
Gly Asn Leu Leu Val Pro Asn Asp Asn 165
170 175Tyr Cys Lys Phe Glu Asn Trp Ile Ile Pro Val Phe
Asp Gln Met Tyr 180 185 190Glu
Glu Gln Ile Asn Glu Lys Val Leu Trp Thr Pro Ser Lys Val Ile 195
200 205Ala Arg Leu Gly Lys Glu Ile Asn Asp
Glu Thr Ser Tyr Leu Tyr Trp 210 215
220Ala Tyr Lys Asn Arg Ile Pro Val Phe Cys Pro Gly Leu Thr Asp Gly225
230 235 240Ser Leu Gly Asp
Met Leu Tyr Phe His Ser Phe Lys Lys Gly Asp Pro 245
250 255Asp Asn Pro Asp Leu Asn Pro Gly Leu Val
Ile Asp Ile Val Gly Asp 260 265
270Ile Arg Ala Met Asn Gly Glu Ala Val His Ala Gly Leu Arg Lys Thr
275 280 285Gly Met Ile Ile Leu Gly Gly
Gly Leu Pro Lys His His Val Cys Asn 290 295
300Ala Asn Met Met Arg Asn Gly Ala Asp Phe Ala Val Phe Ile Asn
Thr305 310 315 320Ala Gln
Glu Phe Asp Gly Ser Asp Ser Gly Ala Arg Pro Asp Glu Ala
325 330 335Val Ser Trp Gly Lys Ile Arg
Gly Gly Ala Lys Thr Val Lys Val His 340 345
350Cys Asp Ala Thr Ile Ala Phe Pro Ile Leu Val Ala Glu Thr
Phe Ala 355 360 365Ala Lys Ser Lys
Glu Phe Ser Gln Ile Arg Cys Gln Val 370 375
3803369PRTHomo sapiens 3Met Glu Gly Ser Leu Glu Arg Glu Ala Pro Ala
Gly Ala Leu Ala Ala1 5 10
15Val Leu Lys His Ser Ser Thr Leu Pro Pro Glu Ser Thr Gln Val Arg
20 25 30Gly Tyr Asp Phe Asn Arg Gly
Val Asn Tyr Arg Ala Leu Leu Glu Ala 35 40
45Phe Gly Thr Thr Gly Phe Gln Ala Thr Asn Phe Gly Arg Ala Val
Gln 50 55 60Gln Val Asn Ala Met Ile
Glu Lys Lys Leu Glu Pro Leu Ser Gln Asp65 70
75 80Glu Asp Gln His Ala Asp Leu Thr Gln Ser Arg
Arg Pro Leu Thr Ser 85 90
95Cys Thr Ile Phe Leu Gly Tyr Thr Ser Asn Leu Ile Ser Ser Gly Ile
100 105 110Arg Glu Thr Ile Arg Tyr
Leu Val Gln His Asn Met Val Asp Val Leu 115 120
125Val Thr Thr Ala Gly Gly Val Glu Glu Asp Leu Ile Lys Cys
Leu Ala 130 135 140Pro Thr Tyr Leu Gly
Glu Phe Ser Leu Arg Gly Lys Glu Leu Arg Glu145 150
155 160Asn Gly Ile Asn Arg Ile Gly Asn Leu Leu
Val Pro Asn Glu Asn Tyr 165 170
175Cys Lys Phe Glu Asp Trp Leu Met Pro Ile Leu Asp Gln Met Val Met
180 185 190Glu Gln Asn Thr Glu
Gly Val Lys Trp Thr Pro Ser Lys Met Ile Ala 195
200 205Arg Leu Gly Lys Glu Ile Asn Asn Pro Glu Ser Val
Tyr Tyr Trp Ala 210 215 220Gln Lys Asn
His Ile Pro Val Phe Ser Pro Ala Leu Thr Asp Gly Ser225
230 235 240Leu Gly Asp Met Ile Phe Phe
His Ser Tyr Lys Asn Pro Gly Leu Val 245
250 255Leu Asp Ile Val Glu Asp Leu Arg Leu Ile Asn Thr
Gln Ala Ile Phe 260 265 270Ala
Lys Cys Thr Gly Met Ile Ile Leu Gly Gly Gly Val Val Lys His 275
280 285His Ile Ala Asn Ala Asn Leu Met Arg
Asn Gly Ala Asp Tyr Ala Val 290 295
300Tyr Ile Asn Thr Ala Gln Glu Phe Asp Gly Ser Asp Ser Gly Ala Arg305
310 315 320Pro Asp Glu Ala
Val Ser Trp Gly Lys Ile Arg Val Asp Ala Gln Pro 325
330 335Val Lys Val Tyr Ala Asp Ala Ser Leu Val
Phe Pro Leu Leu Val Ala 340 345
350Glu Thr Phe Ala Gln Lys Met Asp Ala Phe Met His Glu Lys Asn Glu
355 360 365Asp 46PRTArtificial
SequenceDescription of Artificial Sequence Synthetic peptide
fragment 4Thr Gly Lys His Gly His1 552272DNAArabidopsis
thalianaCDS(68)..(265)CDS(348)..(536)CDS(624)..(842)CDS(979)..(1065)CDS(1-
154)..(1258)CDS(1575)..(1862) 5gaactcccaa aaccctctac tactacactt tcagatccaa
ggaaatcaat tttgtcattc 60gagcaac atg gag gat gat cgt gtt ttc tct tcg
gtt cac tca aca gtt 109 Met Glu Asp Asp Arg Val Phe Ser Ser
Val His Ser Thr Val 1 5 10ttc aaa
gaa tcc gaa tca ttg gaa gga aag tgt gat aaa atc gaa gga 157Phe Lys
Glu Ser Glu Ser Leu Glu Gly Lys Cys Asp Lys Ile Glu Gly15
20 25 30tac gat ttc aat caa gga gta
gat tac cca aag ctt atg cga tcc atg 205Tyr Asp Phe Asn Gln Gly Val
Asp Tyr Pro Lys Leu Met Arg Ser Met 35 40
45ctc acc acc gga ttt caa gcc tcg aat ctc ggc gaa gct
att gat gtc 253Leu Thr Thr Gly Phe Gln Ala Ser Asn Leu Gly Glu Ala
Ile Asp Val 50 55 60gtc aat
caa atg gttcgtttct cgaattcatc aaaaataaaa attccttctt 305Val Asn
Gln Met 65tttgttttcc tttgttttgg gtgaattagt aatgacaaag ag ttt gaa
ttt gta 359 Phe Glu
Phe Val 70ttg
aag cta gat tgg aga ctg gct gat gaa act aca gta gct gaa gac 407Leu
Lys Leu Asp Trp Arg Leu Ala Asp Glu Thr Thr Val Ala Glu Asp
75 80 85tgt agt gaa gag gag aag aat
cca tcg ttt aga gag tct gtc aag tgt 455Cys Ser Glu Glu Glu Lys Asn
Pro Ser Phe Arg Glu Ser Val Lys Cys 90 95
100aaa atc ttt cta ggt ttc act tca aat ctt gtt tca tct ggt
gtt aga 503Lys Ile Phe Leu Gly Phe Thr Ser Asn Leu Val Ser Ser Gly
Val Arg 105 110 115gat act att cgt
tat ctt gtt cag cat cat atg gtttgtgatt tttgctttat 556Asp Thr Ile Arg
Tyr Leu Val Gln His His Met 120 125caccctgctt
ttttatagat gttaaaattt tcgagcttta gttttgattt caatggtttt 616tctgcag gtt
gat gtt ata gtc acg aca act ggt ggt gtt gag gaa gat 665 Val
Asp Val Ile Val Thr Thr Thr Gly Gly Val Glu Glu Asp 130
135 140ctc ata aaa tgc ctt gca cct aca ttt aaa ggt
gat ttc tct cta cct 713Leu Ile Lys Cys Leu Ala Pro Thr Phe Lys Gly
Asp Phe Ser Leu Pro 145 150 155gga gct
tat tta agg tca aag gga ttg aac cga att ggg aat ttg ctg 761Gly Ala
Tyr Leu Arg Ser Lys Gly Leu Asn Arg Ile Gly Asn Leu Leu160
165 170 175gtt cct aat gat aac tac tgc
aag ttt gag gat tgg atc att ccc atc 809Val Pro Asn Asp Asn Tyr Cys
Lys Phe Glu Asp Trp Ile Ile Pro Ile 180
185 190ttt gac gag atg ttg aag gaa cag aaa gaa gag
gtattgcttt atctttcctt 862Phe Asp Glu Met Leu Lys Glu Gln Lys Glu Glu
195 200tttatatgat ttgagatgat tctgtttgtg
cgtcactagt ggagatagat tttgattcct 922ctcttgcatc attgacttcg ttggtgaatc
cttctttctc tggtttttcc ttgtag aat 981
Asngtg ttg tgg act cct tct aaa ctg tta gca cgg
ctg gga aaa gaa atc 1029Val Leu Trp Thr Pro Ser Lys Leu Leu Ala Arg
Leu Gly Lys Glu Ile 205 210 215aac aat
gag agt tca tac ctt tat tgg gca tac aag gtatccaaaa 1075Asn Asn
Glu Ser Ser Tyr Leu Tyr Trp Ala Tyr Lys220 225
230ttttaacctt tttagttttt taatcatcct gtgaggaact cggggattta aattttccgc
1135ttcttgtggt gtttgtag atg aat att cca gta ttc tgc cca ggg tta aca
1186 Met Asn Ile Pro Val Phe Cys Pro Gly Leu Thr
235 240gat ggc tct ctt ggg gat
atg ctg tat ttt cac tct ttt cgt acc tct 1234Asp Gly Ser Leu Gly Asp
Met Leu Tyr Phe His Ser Phe Arg Thr Ser 245 250
255ggc ctc atc atc gat gta gta caa ggtacttctt ttactcaata
agtcagtgtg 1288Gly Leu Ile Ile Asp Val Val Gln 260
265ataaatattc ctgctacatc tagtgcagga atattgtaac tagtagtgca ttgtagcttt
1348tccaattcag caacggactt tactgtaagt tgatatctaa aggttcaaac gggagctagg
1408agaatagcat aggggcattc tgatttaggt ttggggcact gggttaagag ttagagaata
1468ataatcttgt tagttgttta tcaaactctt tgatggttag tctcttggta atttgaattt
1528tatcacagtg tttatggtct ttgaaccagt taatgtttta tgaaca gat atc aga
1583 Asp Ile Arggct atg
aac ggc gaa gct gtc cat gca aat cct aaa aag aca ggg atg 1631Ala Met
Asn Gly Glu Ala Val His Ala Asn Pro Lys Lys Thr Gly Met270
275 280 285ata atc ctt gga ggg ggc ttg
cca aag cac cac ata tgt aat gcc aat 1679Ile Ile Leu Gly Gly Gly Leu
Pro Lys His His Ile Cys Asn Ala Asn 290
295 300atg atg cgc aat ggt gca gat tac gct gta ttt ata
aac acc ggg caa 1727Met Met Arg Asn Gly Ala Asp Tyr Ala Val Phe Ile
Asn Thr Gly Gln 305 310 315gaa
ttt gat ggg agc gac tcg ggt gca cgc cct gat gaa gcc gtg tct 1775Glu
Phe Asp Gly Ser Asp Ser Gly Ala Arg Pro Asp Glu Ala Val Ser 320
325 330tgg ggt aaa att agg ggt tct gct aaa
acc gtt aag gtc tgc ttt tta 1823Trp Gly Lys Ile Arg Gly Ser Ala Lys
Thr Val Lys Val Cys Phe Leu 335 340
345att tct tca cat cct aat tta tat ctc act cag tgg ttt tgagtacata
1872Ile Ser Ser His Pro Asn Leu Tyr Leu Thr Gln Trp Phe350
355 360tttaatattg gatcattctt gcaggtatac tgtgatgcta
ccatagcctt cccattgttg 1932gttgcagaaa catttgccac aaagagagac caaacctgtg
agtctaagac ttaagaactg 1992actggtcgtt ttggccatgg attcttaaag atcgttgctt
tttgatttta cactggagtg 2052accatataac actccacatt gatgtggctg tgacgcgaat
tgtcttcttg cgaattgtac 2112tttagtttct ctcaacctaa aatgatttgc agattgtgtt
ttcgtttaaa acacaagagt 2172cttgtagtca ataatccttt gccttataaa attattcagt
tccaacaaca cattgtgatt 2232ctgtgacaag tctcccgttg cctatgttca cttctctgcg
22726362PRTArabidopsis thaliana 6Met Glu Asp Asp
Arg Val Phe Ser Ser Val His Ser Thr Val Phe Lys1 5
10 15Glu Ser Glu Ser Leu Glu Gly Lys Cys Asp
Lys Ile Glu Gly Tyr Asp 20 25
30Phe Asn Gln Gly Val Asp Tyr Pro Lys Leu Met Arg Ser Met Leu Thr
35 40 45Thr Gly Phe Gln Ala Ser Asn Leu
Gly Glu Ala Ile Asp Val Val Asn 50 55
60Gln Met Phe Glu Phe Val Leu Lys Leu Asp Trp Arg Leu Ala Asp Glu65
70 75 80Thr Thr Val Ala Glu
Asp Cys Ser Glu Glu Glu Lys Asn Pro Ser Phe 85
90 95Arg Glu Ser Val Lys Cys Lys Ile Phe Leu Gly
Phe Thr Ser Asn Leu 100 105
110Val Ser Ser Gly Val Arg Asp Thr Ile Arg Tyr Leu Val Gln His His
115 120 125Met Val Asp Val Ile Val Thr
Thr Thr Gly Gly Val Glu Glu Asp Leu 130 135
140Ile Lys Cys Leu Ala Pro Thr Phe Lys Gly Asp Phe Ser Leu Pro
Gly145 150 155 160Ala Tyr
Leu Arg Ser Lys Gly Leu Asn Arg Ile Gly Asn Leu Leu Val
165 170 175Pro Asn Asp Asn Tyr Cys Lys
Phe Glu Asp Trp Ile Ile Pro Ile Phe 180 185
190Asp Glu Met Leu Lys Glu Gln Lys Glu Glu Asn Val Leu Trp
Thr Pro 195 200 205Ser Lys Leu Leu
Ala Arg Leu Gly Lys Glu Ile Asn Asn Glu Ser Ser 210
215 220Tyr Leu Tyr Trp Ala Tyr Lys Met Asn Ile Pro Val
Phe Cys Pro Gly225 230 235
240Leu Thr Asp Gly Ser Leu Gly Asp Met Leu Tyr Phe His Ser Phe Arg
245 250 255Thr Ser Gly Leu Ile
Ile Asp Val Val Gln Asp Ile Arg Ala Met Asn 260
265 270Gly Glu Ala Val His Ala Asn Pro Lys Lys Thr Gly
Met Ile Ile Leu 275 280 285Gly Gly
Gly Leu Pro Lys His His Ile Cys Asn Ala Asn Met Met Arg 290
295 300Asn Gly Ala Asp Tyr Ala Val Phe Ile Asn Thr
Gly Gln Glu Phe Asp305 310 315
320Gly Ser Asp Ser Gly Ala Arg Pro Asp Glu Ala Val Ser Trp Gly Lys
325 330 335Ile Arg Gly Ser
Ala Lys Thr Val Lys Val Cys Phe Leu Ile Ser Ser 340
345 350His Pro Asn Leu Tyr Leu Thr Gln Trp Phe
355 360719DNAArtificial SequenceDescription of
Artificial Sequence Synthetic Primer 7ggtggtgttg aggaagatc
19820DNAArtificial
SequenceDescription of Artificial Sequence Synthetic Primer
8ggtgcacgcc ctgatgaagc
2091660DNADianthus caryophyllusCDS(256)..(1374) 9gtcattacaa tgcataggat
cattgcacat gctaccttcc tcattgcact tgagcttgcc 60atacttttgt ttttgacgtt
tgataataat actatgaaaa tattatgttt tttcttttgt 120gtgttggtgt ttttgaagtt
gtttttgata agcagaaccc agttgtttta cacttttacc 180attgaactac tgcaattcta
aaactttgtt tacattttaa ttccatcaaa gattgagttc 240agcataggaa aaagg atg
gag gat gct aat cat gat agt gtg gca tct gcg 291 Met
Glu Asp Ala Asn His Asp Ser Val Ala Ser Ala 1
5 10cac tct gca gca ttc aaa aag tcg gag aat tta gag
ggg aaa agc gtt 339His Ser Ala Ala Phe Lys Lys Ser Glu Asn Leu Glu
Gly Lys Ser Val 15 20 25aag att
gag ggt tat gat ttt aat caa ggt gta aac tat tcc aaa ctc 387Lys Ile
Glu Gly Tyr Asp Phe Asn Gln Gly Val Asn Tyr Ser Lys Leu 30
35 40ttg caa tct ttc gct tct aat ggg ttt caa gcc
tcg aat ctt gga gat 435Leu Gln Ser Phe Ala Ser Asn Gly Phe Gln Ala
Ser Asn Leu Gly Asp45 50 55
60gcc att gaa gta gtt aat cat atg cta gat tgg agt ctg gca gat gag
483Ala Ile Glu Val Val Asn His Met Leu Asp Trp Ser Leu Ala Asp Glu
65 70 75gca cct gtg gac gat
tgt agc gag gaa gag agg gat cct aaa ttc aga 531Ala Pro Val Asp Asp
Cys Ser Glu Glu Glu Arg Asp Pro Lys Phe Arg 80
85 90gaa tct gtg aag tgc aaa gtg ttc ttg ggc ttt act
tca aat ctt att 579Glu Ser Val Lys Cys Lys Val Phe Leu Gly Phe Thr
Ser Asn Leu Ile 95 100 105tcc tct
ggt gtt cgt gac aca att cgg tat ctc gtg caa cat cat atg 627Ser Ser
Gly Val Arg Asp Thr Ile Arg Tyr Leu Val Gln His His Met 110
115 120gtt gac gtg ata gta acg aca acc gga ggt ata
gaa gaa gat cta ata 675Val Asp Val Ile Val Thr Thr Thr Gly Gly Ile
Glu Glu Asp Leu Ile125 130 135
140aaa gga aga tcc atc aag tgc ctt gca ccc act ttc aaa ggc gat ttt
723Lys Gly Arg Ser Ile Lys Cys Leu Ala Pro Thr Phe Lys Gly Asp Phe
145 150 155gcc tta cca gga gct
caa tta cgc tcc aaa ggg ttg aat cga att ggt 771Ala Leu Pro Gly Ala
Gln Leu Arg Ser Lys Gly Leu Asn Arg Ile Gly 160
165 170aat ctg ttg gtt ccg aat gat aac tac tgt aaa ttt
gag gat tgg atc 819Asn Leu Leu Val Pro Asn Asp Asn Tyr Cys Lys Phe
Glu Asp Trp Ile 175 180 185att cca
att tta gat aag atg ttg gaa gag caa att tca gag aaa atc 867Ile Pro
Ile Leu Asp Lys Met Leu Glu Glu Gln Ile Ser Glu Lys Ile 190
195 200tta tgg aca cca tcg aag ttg att ggt cga tta
gga aga gaa ata aac 915Leu Trp Thr Pro Ser Lys Leu Ile Gly Arg Leu
Gly Arg Glu Ile Asn205 210 215
220gat gag agt tca tac ctt tac tgg gcc ttc aag aac aat att cca gta
963Asp Glu Ser Ser Tyr Leu Tyr Trp Ala Phe Lys Asn Asn Ile Pro Val
225 230 235ttt tgc cca ggt tta
aca gac ggc tca ctc gga gac atg cta tat ttt 1011Phe Cys Pro Gly Leu
Thr Asp Gly Ser Leu Gly Asp Met Leu Tyr Phe 240
245 250cat tct ttt cgc aat ccg ggt tta atc gtc gat gtt
gtg caa gat ata 1059His Ser Phe Arg Asn Pro Gly Leu Ile Val Asp Val
Val Gln Asp Ile 255 260 265aga gca
gta aat ggc gag gct gtg cac gca gcg cct agg aaa aca ggc 1107Arg Ala
Val Asn Gly Glu Ala Val His Ala Ala Pro Arg Lys Thr Gly 270
275 280atg att ata ctc ggt gga ggg ttg cct aag cac
cac atc tgc aac gca 1155Met Ile Ile Leu Gly Gly Gly Leu Pro Lys His
His Ile Cys Asn Ala285 290 295
300aac atg atg aga aat ggc gcc gat tat gct gtt ttc atc aac act gcc
1203Asn Met Met Arg Asn Gly Ala Asp Tyr Ala Val Phe Ile Asn Thr Ala
305 310 315gaa gag ttt gac ggc
agt gat tct ggt gct cgc ccc gat gag gct att 1251Glu Glu Phe Asp Gly
Ser Asp Ser Gly Ala Arg Pro Asp Glu Ala Ile 320
325 330tca tgg ggc aaa att agc gga tct gct aag act gtg
aag gtg cat tgt 1299Ser Trp Gly Lys Ile Ser Gly Ser Ala Lys Thr Val
Lys Val His Cys 335 340 345gat gcc
acg ata gct ttc cct cta cta gtc gct gag aca ttt gca gca 1347Asp Ala
Thr Ile Ala Phe Pro Leu Leu Val Ala Glu Thr Phe Ala Ala 350
355 360aaa aga gaa aaa gag agg aag agc tgt
taaaactttt ttgattgttg 1394Lys Arg Glu Lys Glu Arg Lys Ser
Cys365 370aaaaatctgt gttatacaag tctcgaaatg cattttagta
attgacttga tcttatcatt 1454tcaatgtgtt atctttgaaa atgttggtaa tgaaacatct
cacctcttct atacaacatt 1514gttgatccat tgtactccgt atcttgtaat tttggaaaaa
aaaaaccgtc tattgttacg 1574agagagtaca tttttgaggt aaaaatatag gatttttgtg
cgatgcaaat gctggttatt 1634cccttgaaaa aaaaaaaaaa aaaaaa
166010373PRTDianthus caryophyllus 10Met Glu Asp Ala
Asn His Asp Ser Val Ala Ser Ala His Ser Ala Ala1 5
10 15Phe Lys Lys Ser Glu Asn Leu Glu Gly Lys
Ser Val Lys Ile Glu Gly 20 25
30Tyr Asp Phe Asn Gln Gly Val Asn Tyr Ser Lys Leu Leu Gln Ser Phe
35 40 45Ala Ser Asn Gly Phe Gln Ala Ser
Asn Leu Gly Asp Ala Ile Glu Val 50 55
60Val Asn His Met Leu Asp Trp Ser Leu Ala Asp Glu Ala Pro Val Asp65
70 75 80Asp Cys Ser Glu Glu
Glu Arg Asp Pro Lys Phe Arg Glu Ser Val Lys 85
90 95Cys Lys Val Phe Leu Gly Phe Thr Ser Asn Leu
Ile Ser Ser Gly Val 100 105
110Arg Asp Thr Ile Arg Tyr Leu Val Gln His His Met Val Asp Val Ile
115 120 125Val Thr Thr Thr Gly Gly Ile
Glu Glu Asp Leu Ile Lys Gly Arg Ser 130 135
140Ile Lys Cys Leu Ala Pro Thr Phe Lys Gly Asp Phe Ala Leu Pro
Gly145 150 155 160Ala Gln
Leu Arg Ser Lys Gly Leu Asn Arg Ile Gly Asn Leu Leu Val
165 170 175Pro Asn Asp Asn Tyr Cys Lys
Phe Glu Asp Trp Ile Ile Pro Ile Leu 180 185
190Asp Lys Met Leu Glu Glu Gln Ile Ser Glu Lys Ile Leu Trp
Thr Pro 195 200 205Ser Lys Leu Ile
Gly Arg Leu Gly Arg Glu Ile Asn Asp Glu Ser Ser 210
215 220Tyr Leu Tyr Trp Ala Phe Lys Asn Asn Ile Pro Val
Phe Cys Pro Gly225 230 235
240Leu Thr Asp Gly Ser Leu Gly Asp Met Leu Tyr Phe His Ser Phe Arg
245 250 255Asn Pro Gly Leu Ile
Val Asp Val Val Gln Asp Ile Arg Ala Val Asn 260
265 270Gly Glu Ala Val His Ala Ala Pro Arg Lys Thr Gly
Met Ile Ile Leu 275 280 285Gly Gly
Gly Leu Pro Lys His His Ile Cys Asn Ala Asn Met Met Arg 290
295 300Asn Gly Ala Asp Tyr Ala Val Phe Ile Asn Thr
Ala Glu Glu Phe Asp305 310 315
320Gly Ser Asp Ser Gly Ala Arg Pro Asp Glu Ala Ile Ser Trp Gly Lys
325 330 335Ile Ser Gly Ser
Ala Lys Thr Val Lys Val His Cys Asp Ala Thr Ile 340
345 350Ala Phe Pro Leu Leu Val Ala Glu Thr Phe Ala
Ala Lys Arg Glu Lys 355 360 365Glu
Arg Lys Ser Cys 37011780DNALycopersicon esculentumCDS(43)..(522)
11aaagaatcct agagagagaa agggaatcct agagagagaa gc atg tcg gac gaa
54 Met Ser Asp Glu
1gaa cac cat ttt gag tca aag gca gat
gct ggt gcc tca aaa act ttc 102Glu His His Phe Glu Ser Lys Ala Asp
Ala Gly Ala Ser Lys Thr Phe5 10 15
20cca cag caa gct gga acc atc cgt aag aat ggt tac atc gtt
atc aaa 150Pro Gln Gln Ala Gly Thr Ile Arg Lys Asn Gly Tyr Ile Val
Ile Lys 25 30 35ggc cgt
ccc tgc aag gtt gtt gag gtc tcc act tca aaa act gga aaa 198Gly Arg
Pro Cys Lys Val Val Glu Val Ser Thr Ser Lys Thr Gly Lys 40
45 50cac gga cat gct aaa tgt cac ttt gtg
gca att gac att ttc aat gga 246His Gly His Ala Lys Cys His Phe Val
Ala Ile Asp Ile Phe Asn Gly 55 60
65aag aaa ctg gaa gat atc gtt ccg tcc tcc cac aat tgt gat gtg cca
294Lys Lys Leu Glu Asp Ile Val Pro Ser Ser His Asn Cys Asp Val Pro 70
75 80cat gtt aac cgt acc gac tat cag ctg
att gat atc tct gaa gat ggt 342His Val Asn Arg Thr Asp Tyr Gln Leu
Ile Asp Ile Ser Glu Asp Gly85 90 95
100ttt gtc tca ctt ctt act gaa agt gga aac acc aag gat gac
ctc agg 390Phe Val Ser Leu Leu Thr Glu Ser Gly Asn Thr Lys Asp Asp
Leu Arg 105 110 115ctt ccc
acc gat gaa aat ctg ctg aag cag gtt aaa gat ggg ttc cag 438Leu Pro
Thr Asp Glu Asn Leu Leu Lys Gln Val Lys Asp Gly Phe Gln 120
125 130gaa gga aag gat ctt gtg gtg tct gtt
atg tct gcg atg ggc gaa gag 486Glu Gly Lys Asp Leu Val Val Ser Val
Met Ser Ala Met Gly Glu Glu 135 140
145cag att aac gcc gtt aag gat gtt ggt acc aag aat tagttatgtc
532Gln Ile Asn Ala Val Lys Asp Val Gly Thr Lys Asn 150
155 160atggcagcat aatcactgcc aaagctttaa gacattatca
tatcctaatg tggtactttg 592atatcactag attataaact gtgttatttg cactgttcaa
aacaaaagaa agaaaactgc 652tgttatggct agagaaagta ttggctttga gcttttgaca
gcacagttga actatgtgaa 712aattctactt tttttttttt gggtaaaata ctgctcgttt
aatgttttgc aaaaaaaaaa 772aaaaaaaa
78012160PRTLycopersicon esculentum 12Met Ser Asp
Glu Glu His His Phe Glu Ser Lys Ala Asp Ala Gly Ala1 5
10 15Ser Lys Thr Phe Pro Gln Gln Ala Gly
Thr Ile Arg Lys Asn Gly Tyr 20 25
30Ile Val Ile Lys Gly Arg Pro Cys Lys Val Val Glu Val Ser Thr Ser
35 40 45Lys Thr Gly Lys His Gly His
Ala Lys Cys His Phe Val Ala Ile Asp 50 55
60Ile Phe Asn Gly Lys Lys Leu Glu Asp Ile Val Pro Ser Ser His Asn65
70 75 80Cys Asp Val Pro
His Val Asn Arg Thr Asp Tyr Gln Leu Ile Asp Ile 85
90 95Ser Glu Asp Gly Phe Val Ser Leu Leu Thr
Glu Ser Gly Asn Thr Lys 100 105
110Asp Asp Leu Arg Leu Pro Thr Asp Glu Asn Leu Leu Lys Gln Val Lys
115 120 125Asp Gly Phe Gln Glu Gly Lys
Asp Leu Val Val Ser Val Met Ser Ala 130 135
140Met Gly Glu Glu Gln Ile Asn Ala Val Lys Asp Val Gly Thr Lys
Asn145 150 155
16013812DNADianthus caryophyllusCDS(67)..(546) 13ctcttttaca tcaatcgaaa
aaaaattagg gttcttattt tagagtgaga ggcgaaaaat 60cgaacg atg tcg gac gac
gat cac cat ttc gag tca tcg gcc gac gcc 108 Met Ser Asp Asp
Asp His His Phe Glu Ser Ser Ala Asp Ala 1 5
10gga gca tcc aag act tac cct caa caa gct ggt aca atc cgc aag agc
156Gly Ala Ser Lys Thr Tyr Pro Gln Gln Ala Gly Thr Ile Arg Lys Ser15
20 25 30ggt cac atc
gtc atc aaa aat cgc cct tgc aag gtg gtt gag gtt tct 204Gly His Ile
Val Ile Lys Asn Arg Pro Cys Lys Val Val Glu Val Ser 35
40 45acc tcc aag act ggc aag cac ggt cat
gcc aaa tgt cac ttt gtt gcc 252Thr Ser Lys Thr Gly Lys His Gly His
Ala Lys Cys His Phe Val Ala 50 55
60att gac att ttc aac ggc aag aag ctg gaa gat att gtc ccc tca tcc
300Ile Asp Ile Phe Asn Gly Lys Lys Leu Glu Asp Ile Val Pro Ser Ser
65 70 75cac aat tgt gat gtt cca cat
gtc aac cgt gtc gac tac cag ctg ctt 348His Asn Cys Asp Val Pro His
Val Asn Arg Val Asp Tyr Gln Leu Leu 80 85
90gat atc act gaa gat ggc ttt gtt agt ctg ctg act gac agt ggt gac
396Asp Ile Thr Glu Asp Gly Phe Val Ser Leu Leu Thr Asp Ser Gly Asp95
100 105 110acc aag gat gat
ctg aag ctt cct gct gat gag gcc ctt gtg aag cag 444Thr Lys Asp Asp
Leu Lys Leu Pro Ala Asp Glu Ala Leu Val Lys Gln 115
120 125atg aag gag gga ttt gag gcg ggg aaa gac
ttg att ctg tca gtc atg 492Met Lys Glu Gly Phe Glu Ala Gly Lys Asp
Leu Ile Leu Ser Val Met 130 135
140tgt gca atg gga gaa gag cag atc tgc gcc gtc aag gac gtt agt ggt
540Cys Ala Met Gly Glu Glu Gln Ile Cys Ala Val Lys Asp Val Ser Gly
145 150 155ggc aag tagaagcttt tgatgaatcc
aatactacgc ggtgcagttg aagcaatagt 596Gly Lys 160aatctcgaga
acattctgaa ccttatatgt tgaattgatg gtgcttagtt tgttttggaa 656atctctttgc
aattaagttg taccaaatca atggatgtaa tgtcttgaat ttgttttatt 716tttgttttga
tgtttgctgt gattgcatta tgcattgtta tgagttatga cctgttataa 776cacaaggttt
tggtaaaaaa aaaaaaaaaa aaaaaa
81214160PRTDianthus caryophyllus 14Met Ser Asp Asp Asp His His Phe Glu
Ser Ser Ala Asp Ala Gly Ala1 5 10
15Ser Lys Thr Tyr Pro Gln Gln Ala Gly Thr Ile Arg Lys Ser Gly
His 20 25 30Ile Val Ile Lys
Asn Arg Pro Cys Lys Val Val Glu Val Ser Thr Ser 35
40 45Lys Thr Gly Lys His Gly His Ala Lys Cys His Phe
Val Ala Ile Asp 50 55 60Ile Phe Asn
Gly Lys Lys Leu Glu Asp Ile Val Pro Ser Ser His Asn65 70
75 80Cys Asp Val Pro His Val Asn Arg
Val Asp Tyr Gln Leu Leu Asp Ile 85 90
95Thr Glu Asp Gly Phe Val Ser Leu Leu Thr Asp Ser Gly Asp
Thr Lys 100 105 110Asp Asp Leu
Lys Leu Pro Ala Asp Glu Ala Leu Val Lys Gln Met Lys 115
120 125Glu Gly Phe Glu Ala Gly Lys Asp Leu Ile Leu
Ser Val Met Cys Ala 130 135 140Met Gly
Glu Glu Gln Ile Cys Ala Val Lys Asp Val Ser Gly Gly Lys145
150 155 16015702DNAArabidopsis
thalianaCDS(56)..(529) 15ctgttaccaa aaaatctgta ccgcaaaatc ctcgtcgaag
ctcgctgctg caacc atg 58
Met
1tcc gac gag gag cat cac ttt gag tcc agt gac gcc gga gcg tcc aaa
106Ser Asp Glu Glu His His Phe Glu Ser Ser Asp Ala Gly Ala Ser Lys
5 10 15acc tac cct caa caa gct
gga acc atc cgt aag aat ggt tac atc gtc 154Thr Tyr Pro Gln Gln Ala
Gly Thr Ile Arg Lys Asn Gly Tyr Ile Val 20 25
30atc aaa aat cgt ccc tgc aag gtt gtt gag gtt tca acc
tcg aag act 202Ile Lys Asn Arg Pro Cys Lys Val Val Glu Val Ser Thr
Ser Lys Thr 35 40 45ggc aag cat
ggt cat gct aaa tgt cat ttt gta gct att gat atc ttc 250Gly Lys His
Gly His Ala Lys Cys His Phe Val Ala Ile Asp Ile Phe 50
55 60 acc agc aag aaa ctc gaa gat att
gtt cct tct tcc cac aat tgt gat 298Thr Ser Lys Lys Leu Glu Asp Ile
Val Pro Ser Ser His Asn Cys Asp65 70 75
80gtt cct cat gtc aac cgt act gat tat cag ctg att gac
att tct gaa 346Val Pro His Val Asn Arg Thr Asp Tyr Gln Leu Ile Asp
Ile Ser Glu 85 90 95gat
gga tat gtc agt ttg ttg act gat aac ggt agt acc aag gat gac 394Asp
Gly Tyr Val Ser Leu Leu Thr Asp Asn Gly Ser Thr Lys Asp Asp
100 105 110ctt aag ctc cct aat gat gac
act ctg ctc caa cag atc aag agt ggg 442Leu Lys Leu Pro Asn Asp Asp
Thr Leu Leu Gln Gln Ile Lys Ser Gly 115 120
125ttt gat gat gga aaa gat cta gtg gtg agt gta atg tca gct atg
gga 490Phe Asp Asp Gly Lys Asp Leu Val Val Ser Val Met Ser Ala Met
Gly 130 135 140 gag
gaa cag atc aat gct ctt aag gac atc ggt ccc aag tgagactaac 539Glu
Glu Gln Ile Asn Ala Leu Lys Asp Ile Gly Pro Lys145 150
155aaagcctccc ctttgttatg agattcttct tcttctgtag gcttccatta
ctcgtcggag 599attatcttgt ttttgggtta ctcctatttt ggatatttaa acttttgtta
ataatgccat 659cttcttcaac cttttccttc tagatggttt ttatacttct tct
70216158PRTArabidopsis thaliana 16Met Ser Asp Glu Glu His His
Phe Glu Ser Ser Asp Ala Gly Ala Ser1 5 10
15Lys Thr Tyr Pro Gln Gln Ala Gly Thr Ile Arg Lys Asn
Gly Tyr Ile 20 25 30Val Ile
Lys Asn Arg Pro Cys Lys Val Val Glu Val Ser Thr Ser Lys 35
40 45Thr Gly Lys His Gly His Ala Lys Cys His
Phe Val Ala Ile Asp Ile 50 55 60Phe
Thr Ser Lys Lys Leu Glu Asp Ile Val Pro Ser Ser His Asn Cys65
70 75 80Asp Val Pro His Val Asn
Arg Thr Asp Tyr Gln Leu Ile Asp Ile Ser 85
90 95Glu Asp Gly Tyr Val Ser Leu Leu Thr Asp Asn Gly
Ser Thr Lys Asp 100 105 110Asp
Leu Lys Leu Pro Asn Asp Asp Thr Leu Leu Gln Gln Ile Lys Ser 115
120 125Gly Phe Asp Asp Gly Lys Asp Leu Val
Val Ser Val Met Ser Ala Met 130 135
140Gly Glu Glu Gln Ile Asn Ala Leu Lys Asp Ile Gly Pro Lys145
150 1551720DNAArtificial SequenceDescription of
Artificial Sequence Synthetic Primer 17aaarrycgmc cytgcaaggt
201817DNAArtificial
SequenceDescription of Artificial Sequence Synthetic Primer
18aatacgactc actatag
171920DNAArtificial SequenceDescription of Artificial Sequence Synthetic
Primer 19tcyttnccyt cmkctaahcc
202017DNAArtificial SequenceDescription of Artificial Sequence
Synthetic Primer 20attaaccctc actaaag
172122DNAArtificial SequenceDescription of Artificial
Sequence Synthetic Primer 21ctgttaccaa aaaatctgta cc
222221DNAArtificial SequenceDescription of
Artificial Sequence Synthetic Primer 22agaagaagta taaaaaccat c
212323DNAArtificial
SequenceDescription of Artificial Sequence Synthetic Primer
23aaagaatcct agagagagaa agg
232418DNAArtificial SequenceDescription of Artificial Sequence Synthetic
Primer 24ttttacatca atcgaaaa
182523DNAArtificial SequenceDescription of Artificial Sequence
Synthetic Primer 25accaaaacct gtgttataac tcc
2326581DNAArabidopsis thalianaCDS(1)..(579) 26ggt ggt
gtt gag gaa gat ctc ata aaa tgc ctt gca cct aca ttt aaa 48Gly Gly
Val Glu Glu Asp Leu Ile Lys Cys Leu Ala Pro Thr Phe Lys1 5
10 15ggt gat ttc tct cta cct gga gct
tat tta agg tca aag gga ttg aac 96Gly Asp Phe Ser Leu Pro Gly Ala
Tyr Leu Arg Ser Lys Gly Leu Asn 20 25
30cga att ggg aat ttg ctg gtt cct aat gat aac tac tgc aag ttt
gag 144Arg Ile Gly Asn Leu Leu Val Pro Asn Asp Asn Tyr Cys Lys Phe
Glu 35 40 45gat tgg atc att ccc
atc ttt gac gag atg ttg aag gaa cag aaa gaa 192Asp Trp Ile Ile Pro
Ile Phe Asp Glu Met Leu Lys Glu Gln Lys Glu 50 55
60gag aat gtg ttg tgg act cct tct aaa ctg tta gca cgg ctg
gga aaa 240Glu Asn Val Leu Trp Thr Pro Ser Lys Leu Leu Ala Arg Leu
Gly Lys65 70 75 80gaa
atc aac aat gag agt tca tac ctt tat tgg gca tac aag atg aat 288Glu
Ile Asn Asn Glu Ser Ser Tyr Leu Tyr Trp Ala Tyr Lys Met Asn
85 90 95att cca gta ttc tgc cca ggg
tta aca gat ggc tct ctt agg gat atg 336Ile Pro Val Phe Cys Pro Gly
Leu Thr Asp Gly Ser Leu Arg Asp Met 100 105
110ctg tat ttt cac tct ttt cgt acc tct ggc ctc atc atc gat
gta gta 384Leu Tyr Phe His Ser Phe Arg Thr Ser Gly Leu Ile Ile Asp
Val Val 115 120 125caa gat atc aga
gct atg aac ggc gaa gct gtc cat gca aat cct aaa 432Gln Asp Ile Arg
Ala Met Asn Gly Glu Ala Val His Ala Asn Pro Lys 130
135 140aag aca ggg atg ata atc ctt gga ggg ggc ttg cca
aag cac cac ata 480Lys Thr Gly Met Ile Ile Leu Gly Gly Gly Leu Pro
Lys His His Ile145 150 155
160tgt aat gcc aat atg atg cgc aat ggt gca gat tac gct gta ttt ata
528Cys Asn Ala Asn Met Met Arg Asn Gly Ala Asp Tyr Ala Val Phe Ile
165 170 175aac acc ggg caa gaa
ttt gat ggg agc gac tcg ggt gca cgc cct gat 576Asn Thr Gly Gln Glu
Phe Asp Gly Ser Asp Ser Gly Ala Arg Pro Asp 180
185 190gaa gc
581Glu27523DNADianthus caryophyllusCDS(1)..(522) 27mga
aga tcc atc aag tgc ctt gca ccc act ttc aaa ggc gat ttt gcc 48Arg
Arg Ser Ile Lys Cys Leu Ala Pro Thr Phe Lys Gly Asp Phe Ala1
5 10 15tta cca gga gct caa tta cgc
tcc aaa ggg ttg aat cga att ggt aat 96Leu Pro Gly Ala Gln Leu Arg
Ser Lys Gly Leu Asn Arg Ile Gly Asn 20 25
30ctg ttg gtt ccg aat gat aac tac tgt aaa ttt gag gat tgg
atc att 144Leu Leu Val Pro Asn Asp Asn Tyr Cys Lys Phe Glu Asp Trp
Ile Ile 35 40 45cca att tta gat
aag atg ttg gaa gag caa att tca gag aaa atc tta 192Pro Ile Leu Asp
Lys Met Leu Glu Glu Gln Ile Ser Glu Lys Ile Leu 50 55
60tgg aca cca tcg aag ttg att ggt cga tta gga aga gaa
ata aac gat 240Trp Thr Pro Ser Lys Leu Ile Gly Arg Leu Gly Arg Glu
Ile Asn Asp65 70 75
80gag agt tca tac ctt tac tgg gcc ttc aag aac aat att cca gta ttt
288Glu Ser Ser Tyr Leu Tyr Trp Ala Phe Lys Asn Asn Ile Pro Val Phe
85 90 95tgc cca ggt tta aca gac
ggc tca ctc gga gac atg cta tat ttt cat 336Cys Pro Gly Leu Thr Asp
Gly Ser Leu Gly Asp Met Leu Tyr Phe His 100
105 110tct ttt cgc aat ccg ggt tta atc atc gat gtt gtg
caa gat ata aga 384Ser Phe Arg Asn Pro Gly Leu Ile Ile Asp Val Val
Gln Asp Ile Arg 115 120 125gca gta
aat ggc gag gct gtg cac gca gcg cct agg aaa aca ggc atg 432Ala Val
Asn Gly Glu Ala Val His Ala Ala Pro Arg Lys Thr Gly Met 130
135 140att ata ctc ggt gga ggg ttg cct aag cac cac
atc tgc aac gca aac 480Ile Ile Leu Gly Gly Gly Leu Pro Lys His His
Ile Cys Asn Ala Asn145 150 155
160atg atg aga aat ggc gcc gat tat gct gtt ttc atc aac acc g
523Met Met Arg Asn Gly Ala Asp Tyr Ala Val Phe Ile Asn Thr
165 1702824DNAArtificial SequenceDescription of
Artificial Sequence Synthetic Primer 28ttgargaaga tycatmaart gcct
242923DNAArtificial
SequenceDescription of Artificial Sequence Synthetic Primer
29ccatcaaayt cytgkgcrgt gtt
2330484DNAArabidopsis thalianaCDS(2)..(112) 30t gca cgc cct gat gaa gct
gtg tct tgg ggt aaa att agg ggt tct gct 49 Ala Arg Pro Asp Glu Ala
Val Ser Trp Gly Lys Ile Arg Gly Ser Ala 1 5
10 15aaa acc gtt aag gtc tgc ttt tta att tct tca cat
cct aat tta tat 97Lys Thr Val Lys Val Cys Phe Leu Ile Ser Ser His
Pro Asn Leu Tyr 20 25 30ctc
act cag tgg ttt tgagtacata tttaatattg gatcattctt gcaggtatac 152Leu
Thr Gln Trp Phe 35tgtgatgcta ccatagcctt cccattgttg gttgcagaaa
catttgccac aaagagagac 212caaacctgtg agtctaagac ttaagaactg actggtcgtt
ttggccatgg attcttaaag 272atcgttgctt tttgatttta cactggagtg accatataac
actccacatt gatgtggctg 332tgacgcgaat tgtcttcttg cgaattgtac tttagtttct
ctcaacctaa aatgatttgc 392agattgtgtt ttcgtttaaa acacaagagt cttgtagtca
ataatccttt gccttataaa 452attattcagt tccaacaaaa aaaaaaaaaa aa
48431559DNALycopersicon
esculentumCDS(1)..(156)modified_base(542)a, t, c, g, unknown or other
31ggt gct cgt cct gat gaa gct gta tca tgg gga aag ata cgt ggt ggt
48Gly Ala Arg Pro Asp Glu Ala Val Ser Trp Gly Lys Ile Arg Gly Gly1
5 10 15gcc aag act gtg aag gtg
cat tgt gat gca acc att gca ttt ccc ata 96Ala Lys Thr Val Lys Val
His Cys Asp Ala Thr Ile Ala Phe Pro Ile 20 25
30tta gta gct gag aca ttt gca gct aag agt aag gaa ttc
tcc cag ata 144Leu Val Ala Glu Thr Phe Ala Ala Lys Ser Lys Glu Phe
Ser Gln Ile 35 40 45agg tgc caa
gtt tgaacattga ggaagctgtc cttccgacca cacatatgaa 196Arg Cys Gln
Val 50ttgctagctt ttgaagccaa cttgctagtg tgcagcacca tttattctgc
aaaactgact 256agagagcagg gtatattcct ctaccccgag ttagacgaca tcctgtatgg
ttcaaattaa 316ttatttttct ccccttcaca ccatgttatt tagttctctt cctcttcgaa
agtgaagagc 376ttagatgttc ataggttttg aattatgttg gaggttggtg ataactgact
agtcctctta 436ccatatagat aatgtatcct tgtactatga gattttgggt gtgtttgata
ccaaggaaaa 496atgtttattt ggaaaacaat tggattttta atttaaaaaa aattgnttaa
aaaaaaaaaa 556aaa
5593222PRTArtificial SequenceDescription of Artificial
Sequence Synthetic peptide fragment 32Cys Lys Val Val Glu Val Ser
Thr Ser Lys Thr Gly Lys His Gly His1 5 10
15Ala Lys Cys His Phe Val 203329DNAArtificial
SequenceDescription of Artificial Sequence Synthetic Primer
33gagctcaaga ataacatctc ataagaaac
293427DNAArtificial SequenceDescription of Artificial Sequence Synthetic
Primer 34ctcgagtgct cacttctctc tcttagg
273512PRTArtificial SequenceDescription of Artificial Sequence
Synthetic peptide 35Cys Asn Asp Asp Thr Leu Leu Gln Gln Ile Lys Ser1
5 103612PRTArtificial SequenceDescription
of Artificial Sequence Synthetic peptide 36Cys Thr Asp Asp Gly Leu
Thr Ala Gln Met Arg Leu1 5
103712PRTArtificial SequenceDescription of Artificial Sequence Synthetic
peptide 37Cys Thr Asp Glu Ala Leu Leu Thr Gln Leu Lys Asn1
5 103832DNAArtificial SequenceDescription of
Artificial Sequence Synthetic Primer 38aagcttgatc gtggtcaact
tcctctgtta cc 323927DNAArtificial
SequenceDescription of Artificial Sequence Synthetic Primer
39gagctcagaa gaagtataaa aaccatc
274027DNAArtificial SequenceDescription of Artificial Sequence Synthetic
Primer 40ctcgagtgct cacttctctc tcttagg
274129DNAArtificial SequenceDescription of Artificial Sequence
Synthetic Primer 41gagctcaaga ataacatctc ataagaaac
294230DNAArtificial SequenceDescription of Artificial
Sequence Synthetic Primer 42ctcgagctaa actccattcg ctgacttcgc
304329DNAArtificial SequenceDescription of
Artificial Sequence Synthetic Primer 43gagctctagt aaatataaga
gtgtcttgc 2944353PRTUnknown
OrganismDescription of Unknown Organism Fungi sequence 44Met Ala Asp Asn
Gln Ile Pro Ser Ser Val Ala Asp Ala Val Leu Val1 5
10 15Lys Ser Ile Glu Met Pro Glu Gly Ser Gln
Lys Val Glu Glu Leu Asp 20 25
30Phe Asn Lys Phe Lys Gly Arg Pro Ile Thr Val Asp Asp Leu Leu Gln
35 40 45Gly Met Lys His Met Gly Phe Gln
Ala Ser Ser Met Cys Glu Ala Val 50 55
60Arg Ile Ile Asn Glu Met Arg Ala Tyr Arg Asp Pro Thr Thr Ser Glu65
70 75 80Lys Thr Thr Ile Phe
Leu Gly Tyr Thr Ser Asn Leu Ile Ser Ser Gly 85
90 95Leu Arg Gly Thr Leu Arg Tyr Leu Val Gln His
Lys His Val Ser Ala 100 105
110Ile Val Thr Thr Ala Gly Gly Ile Glu Glu Asp Phe Ile Lys Cys Leu
115 120 125Gly Asp Thr Tyr Met Ser Ser
Phe Ser Ala Val Gly Ala Asp Leu Arg 130 135
140Ser Lys Gly Leu Asn Arg Ile Gly Asn Leu Val Val Pro Asn Ser
Asn145 150 155 160Tyr Cys
Ala Phe Glu Asp Trp Val Val Pro Ile Leu Asp Lys Met Leu
165 170 175Glu Glu Gln Glu Ala Ser Arg
Gly Thr Glu Asn Glu Ile Asn Trp Thr 180 185
190Pro Ser Lys Val Ile His Arg Leu Gly Lys Glu Ile Asn Asp
Glu Arg 195 200 205Ser Val Tyr Tyr
Trp Ala Trp Lys Asn Asp Ile Pro Val Phe Cys Pro 210
215 220Ala Leu Thr Asp Gly Ser Leu Gly Asp Met Leu Tyr
Phe His Thr Phe225 230 235
240Lys Ala Ser Pro Glu Gln Leu Arg Ile Asp Ile Val Glu Asp Ile Arg
245 250 255Lys Ile Asn Thr Ile
Ala Val Arg Ala Lys Arg Ala Gly Met Ile Ile 260
265 270Leu Gly Gly Gly Ile Val Lys His His Ile Ala Asn
Ala Cys Leu Met 275 280 285Arg Asn
Gly Ala Glu Ser Ala Val Tyr Ile Asn Thr Ala Gln Glu Phe 290
295 300Asp Gly Ser Asp Ala Gly Ala Arg Pro Asp Glu
Ala Val Ser Trp Gly305 310 315
320Lys Ile Lys Val Gly Ala Asp Ala Val Lys Val Tyr Met Glu Ala Thr
325 330 335Ala Ala Phe Pro
Phe Ile Val Ala Asn Thr Phe Ala Lys Glu Asp Gly 340
345 350Leu 45387PRTUnknown OrganismDescription of
Unknown Organism Yeast sequence 45Met Ser Asp Ile Asn Glu Lys Leu Pro Glu
Leu Leu Gln Asp Ala Val1 5 10
15Leu Lys Ala Ser Val Pro Ile Pro Asp Asp Phe Val Lys Val Gln Gly
20 25 30Ile Asp Tyr Ser Lys Pro
Glu Ala Thr Asn Met Arg Ala Thr Asp Leu 35 40
45Ile Glu Ala Met Lys Thr Met Gly Phe Gln Ala Ser Ser Val
Gly Thr 50 55 60Ala Cys Glu Ile Ile
Asp Ser Met Arg Ser Trp Arg Gly Lys His Ile65 70
75 80Asp Glu Leu Asp Asp His Glu Lys Lys Gly
Cys Phe Asp Glu Glu Gly 85 90
95Tyr Gln Lys Thr Thr Ile Phe Met Gly Tyr Thr Ser Asn Leu Ile Ser
100 105 110Ser Gly Val Arg Glu
Thr Leu Arg Tyr Leu Val Gln His Lys Met Val 115
120 125Asp Ala Val Val Thr Ser Ala Gly Gly Val Glu Glu
Asp Leu Ile Lys 130 135 140Cys Leu Ala
Pro Thr Tyr Leu Gly Glu Phe Ala Leu Lys Gly Lys Ser145
150 155 160Leu Arg Asp Gln Gly Met Asn
Arg Ile Gly Asn Leu Leu Val Pro Asn 165
170 175Asp Asn Tyr Cys Lys Phe Glu Glu Trp Ile Val Pro
Ile Leu Asp Lys 180 185 190Met
Leu Glu Glu Gln Asp Glu Tyr Val Lys Lys His Gly Ala Asp Cys 195
200 205Leu Glu Ala Asn Gln Asp Val Asp Ser
Pro Ile Trp Thr Pro Ser Lys 210 215
220Met Ile Asp Arg Phe Gly Lys Glu Ile Asn Asp Glu Ser Ser Val Leu225
230 235 240Tyr Trp Ala His
Lys Asn Lys Ile Pro Ile Phe Cys Pro Ser Leu Thr 245
250 255Asp Gly Ser Ile Gly Asp Met Leu Phe Phe
His Thr Phe Lys Ala Ser 260 265
270Pro Lys Gln Leu Arg Val Asp Ile Val Gly Asp Ile Arg Lys Ile Asn
275 280 285Ser Met Ser Met Ala Ala Tyr
Arg Ala Gly Met Ile Ile Leu Gly Gly 290 295
300Gly Leu Ile Lys His His Ile Ala Asn Ala Cys Leu Met Arg Asn
Gly305 310 315 320Ala Asp
Tyr Ala Val Tyr Ile Asn Thr Gly Gln Glu Tyr Asp Gly Ser
325 330 335Asp Ala Gly Ala Arg Pro Asp
Glu Ala Val Ser Trp Gly Lys Ile Lys 340 345
350Ala Glu Ala Lys Ser Val Lys Leu Phe Ala Asp Val Thr Thr
Val Leu 355 360 365Pro Leu Ile Val
Ala Ala Thr Phe Ala Ser Gly Lys Pro Ile Lys Lys 370
375 380Val Lys Asn38546370PRTUnknown OrganismDescription
of Unknown Organism Archaebacterial sequence 46Met Arg Asp Ile Lys
Asp Asn Pro Ile Arg Arg Gly Ile Ala Glu Gln1 5
10 15Ser Glu Ala Met His Pro Gly Tyr Thr Asn Arg
Ala Lys Pro Tyr Gly 20 25
30Cys Lys Arg Asp Pro Lys Asp Ile Val Leu Lys Glu Ser Glu Asp Ile
35 40 45Glu Gly Ile Ala Ile Glu Gly Pro
Trp Leu Glu Asp Asp Ile Ser Leu 50 55
60Glu Glu Ile Ile Lys Lys Tyr Tyr Leu Lys Ile Gly Phe Gln Ala Ser65
70 75 80His Ile Gly Lys Ala
Ile Lys Ile Trp Lys His Ile Glu Glu Lys Arg 85
90 95Lys Lys Gly Asp Glu Ile Thr Val Phe Phe Gly
Tyr Thr Ser Asn Ile 100 105
110Val Ser Ser Gly Leu Arg Glu Ile Ile Ala Tyr Leu Val Lys His Lys
115 120 125Lys Ile Asp Ile Ile Val Thr
Thr Ala Gly Gly Val Glu Glu Asp Phe 130 135
140Ile Lys Cys Leu Lys Pro Phe Ile Leu Gly Asp Trp Glu Val Asp
Gly145 150 155 160Lys Met
Leu Arg Glu Lys Gly Ile Asn Arg Ile Gly Asn Ile Phe Val
165 170 175Pro Asn Asp Arg Tyr Ile Ala
Phe Glu Glu Tyr Met Met Glu Phe Phe 180 185
190Glu Glu Ile Leu Asn Leu Gln Arg Glu Thr Gly Lys Ile Ile
Thr Ala 195 200 205Ser Glu Phe Cys
Tyr Lys Leu Gly Glu Phe Met Asp Lys Lys Leu Lys 210
215 220Ser Lys Glu Lys Glu Lys Ser Ile Leu Tyr Trp Ala
Tyr Lys Asn Asn225 230 235
240Ile Pro Ile Phe Cys Pro Ala Ile Thr Asp Gly Ser Ile Gly Asp Met
245 250 255Leu Tyr Phe Phe Lys
Lys Tyr Asn Lys Asp Glu Glu Leu Lys Ile Asp 260
265 270Val Ala Asn Asp Ile Val Lys Leu Asn Asp Ile Ala
Ile Asn Ser Lys 275 280 285Glu Thr
Ala Cys Ile Val Leu Gly Gly Ser Leu Pro Lys His Ser Ile 290
295 300Ile Asn Ala Asn Leu Phe Arg Glu Gly Thr Asp
Tyr Ala Ile Tyr Val305 310 315
320Thr Thr Ala Leu Pro Trp Asp Gly Ser Leu Ser Gly Ala Pro Pro Glu
325 330 335Glu Gly Val Ser
Trp Gly Lys Ile Gly Ala Lys Ala Asp Tyr Val Glu 340
345 350Ile Trp Gly Asp Ala Thr Ile Ile Phe Pro Leu
Leu Val Tyr Cys Val 355 360 365Met
Lys 3704723DNAArtificial SequenceDescription of Artificial Sequence
Synthetic Primer 47gcngarttyg ayggntccga yca
234830DNAArtificial SequenceDescription of Artificial
Sequence Synthetic Primer 48ccgagctcct gttaccaaaa aatctgtacc
304936DNAArtificial SequenceDescription of
Artificial Sequence Synthetic Primer 49acctcgagcg gccgcagaag
aagtataaaa accatc 365030DNAArtificial
SequenceDescription of Artificial Sequence Synthetic Primer
50cgtcgacgat atctcttttt atattcaaac
305132DNAArtificial SequenceDescription of Artificial Sequence Synthetic
Primer 51cgtctagaca ttgttttagg aaagttaaat ga
32521235DNAArabidopsis thaliana 52accctagatc gctttcttca
gtgttctata aaaactaaac tccattcgct gacttcgcaa 60agaagaacac tttctctctg
aaatctcaaa ttcatctctt ctcttccgat ttcgctgaat 120catgtcagac gacgagcatc
acttcgaatc cagcgacgcc ggagcttcta agacttatcc 180tcaacaagcc ggtaacattc
gtaaaggtgg tcacatcgtc atcaagggac gtccctgcaa 240ggttttgtct ctgatttgat
tattattgat tttagaggaa tcatcttcat ggattgtatt 300aaagcagtgt tccgttacct
gatcgttgtg aatttttgag gtttagtgat tctggattgt 360gatctggtgt ttagtgttga
gaaaaacctc tgtttttgaa gtttatggat ttatagggtt 420tttaaatcta taatagggtt
taattcaatt ggtgatatgt ggggtttatg atataggtgg 480ttgaggtatc gacttcgaag
actgggaagc atggtcacgc caagtgtcac tttgttgcca 540ttgatatctt tacttctaag
aagcttgaag atatcgttcc ttcttcccac aattgtgatg 600tgagtcttgt gaatggatta
gaaacgttat acaaagtcta taatttttga ctcacaacac 660aaaactgttt cctttttatt
ggcacaggtt ccacatgtga atcgtgttga ttatcagttg 720attgatatct ctgaagatgg
ctttgtatgt catcttcttt ttcactagtt cagctttgtg 780ttttgtcttt gcccatatgg
ttgaattaga gggttttgtt ctttgattac atttacaggt 840tagtcttctt actgataatg
gtagcactaa ggatgatctg aagctgccaa cagatgaagc 900tttactcaca cagctcaaga
atggatttga ggagggtaag gatattgttg tgtctgtcat 960gtctgcaatg ggagaggagc
agatgtgtgc tctcaaggaa gttggtccca agtaataata 1020ataagtaagc attctctctt
ttacagaggc tatgtattat caagtttgac agagtcaaat 1080gttataagaa caaagtttgg
tccttttttt tggtcttctt agtataattt aagcccacat 1140gtgtttccca tgcaagacac
tcttatattt actagtatat cttactattg gttttggttg 1200tggagaagtt actgttgaca
gttccaaacc tctac 123553161PRTMycosphaerella
fijiensis 53Gly Leu Asn Arg Ile Gly Asn Phe Leu Val Pro Asn Asp Asn Tyr
Cys1 5 10 15Arg Phe Glu
Asp Trp Val Met Pro Ile Leu Asp Thr Met Leu Glu Glu 20
25 30Gln Glu Ala Cys Lys Gly Ser Gly Glu Ala
Ile His Trp Thr Pro Ser 35 40
45Lys Ile Ile Asn Arg Leu Gly Lys Glu Val Asn Asp Glu Ser Ser Val 50
55 60Tyr Tyr Trp Ala Trp Lys Asn Asp Ile
Pro Val Phe Cys Pro Ala Leu65 70 75
80Thr Asp Gly Ser Leu Gly Asp Met Leu Tyr Phe His Thr Phe
Lys Ser 85 90 95Ser Pro
Gln Gln Leu Arg Val Asp Ile Val Glu Asp Ile Arg Lys Ile 100
105 110Asn Thr Leu Ala Val Arg Ala Lys Arg
Thr Gly Met Ile Ile Leu Gly 115 120
125Gly Gly Ile Val Lys His His Ile Ala Asn Ala Asn Leu Met Arg Asn
130 135 140Gly Ala Glu Ser Ala Val Tyr
Ile Asn Thr Ala Glu Phe Asp Gly Ser145 150
155 160Asp54861DNAArabidopsis sp. 54tttttccctt
ctcccaatct catcttctcc gaaaaccttt cttctctcaa atttctgtga 60aaacatgtct
gacgacgagc accactttga ggccagcgaa tccggagctt ccaagaccta 120tcctcaatca
gccggtaaca tccgtaaagg tggtcacatc gtcatcaaaa accgtccctg 180caaggttgtt
gaggtttcga cttccaaaac tggcaagcac ggtcacgcca aatgtcactt 240tgttgctatt
gatatcttca ctgctaagaa gcttgaagat attgttccat cttcccacaa 300ttgtgatgtt
ccacatgtga accgtgttga ttaccagttg attgatatca ctgaggatgg 360cttcgtgagc
cttctcactg acagtggtgg caccaaggat gatctcaagc ttcccaccga 420tgatggtctc
accgcccaga tgaggcttgg attcgatgag ggaaaggata ttgtggtgtc 480tgtcatgtct
tccatgggag aggagcagat ctgtgccgtc aaggaagttg gtggtggcaa 540gtaaacaagt
atcattcgat atattattac cagtttgaca acggacgtca atgttataag 600aaccaaaaga
tgtttttctt tttcctaatt tagacccttt gtgtgtgttt cttgttgcaa 660gacaaccata
tctattggtt ttggattgtt ggaaaagttt gtgttgaaac attcaaagtt 720tcttatgaga
tgttattctt aaaaccactt tttgtttgtt cactggatat gtttgttcat 780gaagcttgtt
ttaagcaact ctttacatga tattcattgc tatttgcacg attcaagagt 840gaaatataca
ttttatttaa c
86155159PRTArabidopsis thaliana 55Met Ser Asp Asp Glu His His Phe Glu Ala
Ser Glu Ser Gly Ala Ser1 5 10
15Lys Thr Tyr Pro Gln Ser Ala Gly Asn Ile Arg Lys Gly Gly His Ile
20 25 30Val Ile Lys Asn Arg Pro
Cys Lys Val Val Glu Val Ser Thr Ser Lys 35 40
45Thr Gly Lys His Gly His Ala Lys Cys His Phe Val Ala Ile
Asp Ile 50 55 60Phe Thr Ala Lys Lys
Leu Glu Asp Ile Val Pro Ser Ser His Asn Cys65 70
75 80Asp Val Pro His Val Asn Arg Val Asp Tyr
Gln Leu Ile Asp Ile Thr 85 90
95Glu Asp Gly Phe Val Ser Leu Leu Thr Asp Ser Gly Gly Thr Lys Asp
100 105 110Asp Leu Lys Leu Pro
Thr Asp Asp Gly Leu Thr Ala Gln Met Arg Leu 115
120 125Gly Phe Asp Glu Gly Lys Asp Ile Val Val Ser Val
Met Ser Ser Met 130 135 140Gly Glu Glu
Gln Ile Cys Ala Val Lys Glu Val Gly Gly Gly Lys145 150
15556698DNALycopersicon esculentumCDS(52)..(528)
56cttcctgaat ttttctcctt ctccttctcc gttcaatcga atttttcagc c atg tct
57 Met Ser
1gac gag gag cat caa ttt
gag tct aag gct gat gcc gga gca tca aaa 105Asp Glu Glu His Gln Phe
Glu Ser Lys Ala Asp Ala Gly Ala Ser Lys 5 10
15act tac cct caa caa gct ggt act att cgt aag aac ggt tat
atc gtc 153Thr Tyr Pro Gln Gln Ala Gly Thr Ile Arg Lys Asn Gly Tyr
Ile Val 20 25 30atc aaa ggc cgt
cca tgc aag gtt gtg gaa gtc tct aca tcc aaa act 201Ile Lys Gly Arg
Pro Cys Lys Val Val Glu Val Ser Thr Ser Lys Thr 35 40
45 ggc aag cac ggt cac gcc aaa tgt cat
ttc gtt gct att gac atc ttc 249Gly Lys His Gly His Ala Lys Cys His
Phe Val Ala Ile Asp Ile Phe50 55 60
65act ggg aag aag ctt gag gat att gtc ccc tct tca cac aat
tgt gat 297Thr Gly Lys Lys Leu Glu Asp Ile Val Pro Ser Ser His Asn
Cys Asp 70 75 80gtg ccc
cat gtt aat cgt aca gat tat cag ctt att gac atc tct gaa 345Val Pro
His Val Asn Arg Thr Asp Tyr Gln Leu Ile Asp Ile Ser Glu 85
90 95gat gga ttt gtg agt ctg ctt act gac
aat ggt aac acc aag gat gac 393Asp Gly Phe Val Ser Leu Leu Thr Asp
Asn Gly Asn Thr Lys Asp Asp 100 105
110ctc agg ctt cct act gat gaa aat ctg ctt tca ctg atc aag gac ggg
441Leu Arg Leu Pro Thr Asp Glu Asn Leu Leu Ser Leu Ile Lys Asp Gly 115
120 125 ttt gcc gag ggt
aag gac ctc gtt gtg tct gtt atg tca gct atg ggt 489Phe Ala Glu Gly
Lys Asp Leu Val Val Ser Val Met Ser Ala Met Gly130 135
140 145gag gaa cag att aat gct ttg aag gat
att ggc ccc aag tgatctcttg 538Glu Glu Gln Ile Asn Ala Leu Lys Asp
Ile Gly Pro Lys 150 155attggatgga
ttgcttgacg cgatggttct ttacgacctt gagtgagata gatatttata 598gtcatggaaa
aaaattgtga tcttatggaa tattcgtatc atgatttatg gaccattgtg 658agttagattt
ttatttatgt tgttttaaat tgtggtattc
69857159PRTLycopersicon esculentum 57Met Ser Asp Glu Glu His Gln Phe Glu
Ser Lys Ala Asp Ala Gly Ala1 5 10
15Ser Lys Thr Tyr Pro Gln Gln Ala Gly Thr Ile Arg Lys Asn Gly
Tyr 20 25 30Ile Val Ile Lys
Gly Arg Pro Cys Lys Val Val Glu Val Ser Thr Ser 35
40 45Lys Thr Gly Lys His Gly His Ala Lys Cys His Phe
Val Ala Ile Asp 50 55 60Ile Phe Thr
Gly Lys Lys Leu Glu Asp Ile Val Pro Ser Ser His Asn65 70
75 80Cys Asp Val Pro His Val Asn Arg
Thr Asp Tyr Gln Leu Ile Asp Ile 85 90
95Ser Glu Asp Gly Phe Val Ser Leu Leu Thr Asp Asn Gly Asn
Thr Lys 100 105 110Asp Asp Leu
Arg Leu Pro Thr Asp Glu Asn Leu Leu Ser Leu Ile Lys 115
120 125Asp Gly Phe Ala Glu Gly Lys Asp Leu Val Val
Ser Val Met Ser Ala 130 135 140Met Gly
Glu Glu Gln Ile Asn Ala Leu Lys Asp Ile Gly Pro Lys145
150 15558158PRTArabidopsis thaliana 58Met Ser Asp Glu Glu
His His Phe Glu Ser Ser Asp Ala Gly Ala Ser1 5
10 15Lys Thr Tyr Pro Gln Gln Ala Gly Thr Ile Arg
Lys Asn Gly Tyr Ile 20 25
30Val Ile Lys Asn Arg Pro Cys Lys Val Val Glu Val Ser Thr Ser Lys
35 40 45Thr Gly Lys His Gly His Ala Lys
Cys His Phe Val Ala Ile Asp Ile 50 55
60Phe Thr Ser Lys Lys Leu Glu Asp Ile Val Pro Ser Ser His Asn Cys65
70 75 80Asp Val Pro His Val
Asn Arg Thr Asp Tyr Gln Leu Ile Asp Ile Ser 85
90 95Glu Asp Gly Tyr Val Ser Leu Leu Thr Asp Asn
Gly Ser Thr Lys Asp 100 105
110Asp Leu Lys Leu Pro Asn Asp Asp Thr Leu Leu Gln Gln Ile Lys Ser
115 120 125Gly Phe Asp Asp Gly Lys Asp
Leu Val Val Ser Val Met Ser Ala Met 130 135
140Gly Glu Glu Gln Ile Asn Ala Leu Lys Asp Ile Gly Pro Lys145
150 15559159PRTArabidopsis thaliana 59Met Ser
Asp Asp Glu His His Phe Glu Ala Ser Glu Ser Gly Ala Ser1 5
10 15Lys Thr Tyr Pro Gln Ser Ala Gly
Asn Ile Arg Lys Gly Gly His Ile 20 25
30Val Ile Lys Asn Arg Pro Cys Lys Val Val Glu Val Ser Thr Ser
Lys 35 40 45Thr Gly Lys His Gly
His Ala Lys Cys His Phe Val Ala Ile Asp Ile 50 55
60Phe Thr Ala Lys Lys Leu Glu Asp Ile Val Pro Ser Ser His
Asn Cys65 70 75 80Asp
Val Pro His Val Asn Arg Val Asp Tyr Gln Leu Ile Asp Ile Thr
85 90 95Glu Asp Gly Phe Val Ser Leu
Leu Thr Asp Ser Gly Gly Thr Lys Asp 100 105
110Asp Leu Lys Leu Pro Thr Asp Asp Gly Leu Thr Ala Gln Met
Arg Leu 115 120 125Gly Phe Asp Glu
Gly Lys Asp Ile Val Val Ser Val Met Ser Ser Met 130
135 140Gly Glu Glu Gln Ile Cys Ala Val Lys Glu Val Gly
Gly Gly Lys145 150 15560158PRTArabidopsis
thaliana 60Met Ser Asp Asp Glu His His Phe Glu Ser Ser Asp Ala Gly Ala
Ser1 5 10 15Lys Thr Tyr
Pro Gln Gln Ala Gly Asn Ile Arg Lys Gly Gly His Ile 20
25 30Val Ile Lys Gly Arg Pro Cys Lys Val Val
Glu Val Ser Thr Ser Lys 35 40
45Thr Gly Lys His Gly His Ala Lys Cys His Phe Val Ala Ile Asp Ile 50
55 60Phe Thr Ser Lys Lys Leu Glu Asp Ile
Val Pro Ser Ser His Asn Cys65 70 75
80Asp Val Pro His Val Asn Arg Val Asp Tyr Gln Leu Ile Asp
Ile Ser 85 90 95Glu Asp
Gly Phe Val Ser Leu Leu Thr Asp Asn Gly Ser Thr Lys Asp 100
105 110Asp Leu Lys Leu Pro Thr Asp Glu Ala
Leu Leu Thr Gln Leu Lys Asn 115 120
125Gly Phe Glu Glu Gly Lys Asp Ile Val Val Ser Val Met Ser Ala Met
130 135 140Gly Glu Glu Gln Met Cys Ala
Leu Lys Glu Val Gly Pro Lys145 150
15561477DNAArabidopsis thaliana 61atgtccgacg aggagcatca ctttgagtcc
agtgacgccg gagcgtccaa aacctaccct 60caacaagctg gaaccatccg taagaatggt
tacatcgtca tcaaaaatcg tccctgcaag 120gttgttgagg tttcaacctc gaagactggc
aagcatggtc atgctaaatg tcattttgta 180gctattgata tcttcaccag caagaaactc
gaagatattg ttccttcttc ccacaattgt 240gatgttcctc atgtcaaccg tactgattat
cagctgattg acatttctga agatggatat 300gtcagtttgt tgactgataa cggtagtacc
aaggatgacc ttaagctccc taatgatgac 360actctgctcc aacagatcaa gagtgggttt
gatgatggaa aagatctagt ggtgagtgtg 420atgtcagcta tgggagagga acagatcaat
gctcttaagg acatcggtcc caagtga 47762480DNAArabidopsis thaliana
62atgtctgacg acgagcacca ctttgaggcc agcgaatccg gagcttccaa gacctatcct
60caatcagccg gtaacatccg taaaggtggt cacatcgtca tcaaaaaccg tccctgcaag
120gttgttgagg tttcgacttc caaaactggc aagcacggtc acgccaaatg tcactttgtt
180gctattgata tcttcactgc taagaagctt gaagatattg ttccatcttc ccacaattgt
240gatgttccac atgtgaaccg tgttgattac cagttgattg atatcactga ggatggcttc
300gtgagccttc tcactgacag tggtggcacc aaggatgatc tcaagcttcc caccgatgat
360ggtctcaccg cccagatgag gcttggattc gatgagggaa aggatattgt ggtgtctgtc
420atgtcttcca tgggagagga gcagatctgt gccgtcaagg aagttggtgg tggcaagtaa
48063477DNAArabidopsis thaliana 63atgtcagacg acgagcatca cttcgaatcc
agcgacgccg gagcttctaa gacttatcct 60caacaagccg gtaacattcg taaaggtggt
cacatcgtca tcaagggacg tccctgcaag 120gtggttgagg tatcgacttc gaagactggg
aagcatggtc acgccaagtg tcactttgtt 180gccattgata tctttacttc taagaagctt
gaagatatcg ttccttcttc ccacaattgt 240gatgttccac atgtgaatcg tgttgattat
cagttgattg atatctctga agatggcttt 300gttagtcttc ttactgataa tggtagcact
aaggatgatc tgaagctgcc aacagatgaa 360gctttactca cacagctcaa gaatggattt
gaggagggta aggatattgt tgtgtctgtc 420atgtctgcaa tgggagagga gcagatgtgt
gctctcaagg aagttggtcc caagtaa 47764700DNALycopersicon
esculentumCDS(56)..(532) 64aaatttctcc ttctccttaa tcctctccac cggcgaaccg
gcgaagatca aaacg atg 58
Met
1tcg gac gaa gag cac cac ttc gaa tcc aag gcc gat gcc gga gct tca
106Ser Asp Glu Glu His His Phe Glu Ser Lys Ala Asp Ala Gly Ala Ser
5 10 15aag acg tat cct caa caa
gct ggt act att cgt aaa ggt ggt cac atc 154Lys Thr Tyr Pro Gln Gln
Ala Gly Thr Ile Arg Lys Gly Gly His Ile 20 25
30gtc ata aaa aat cgt cct tgc aag gtg gtt gaa gtt tca act
tcc aag 202Val Ile Lys Asn Arg Pro Cys Lys Val Val Glu Val Ser Thr
Ser Lys 35 40 45aca ggc aag cac ggt
cat gct aaa tgt cac ttc gtg gca att gac att 250Thr Gly Lys His Gly
His Ala Lys Cys His Phe Val Ala Ile Asp Ile50 55
60 65ttc act gga aag aaa ctt gag gat att gtt
ccc tct tct cac aat tgt 298Phe Thr Gly Lys Lys Leu Glu Asp Ile Val
Pro Ser Ser His Asn Cys 70 75
80gat gtt cct cat gtg aat agg act gat tat caa ctt att gat atc tct
346Asp Val Pro His Val Asn Arg Thr Asp Tyr Gln Leu Ile Asp Ile Ser
85 90 95gag gat ggc ttt gtg agt
ctg ttg act gaa aat ggt aac acc aag gat 394Glu Asp Gly Phe Val Ser
Leu Leu Thr Glu Asn Gly Asn Thr Lys Asp 100 105
110gac ttg aga ctc cca act gat gat act ctt ctg gct cag gtc
aaa gat 442Asp Leu Arg Leu Pro Thr Asp Asp Thr Leu Leu Ala Gln Val
Lys Asp 115 120 125ggt ttt gct gag ggg
aaa gac ctg gtt cta tca gtg atg tct gcc atg 490Gly Phe Ala Glu Gly
Lys Asp Leu Val Leu Ser Val Met Ser Ala Met130 135
140 145gga gag gag cag att tgt ggt atc aag gac
att ggc cct aag 532Gly Glu Glu Gln Ile Cys Gly Ile Lys Asp
Ile Gly Pro Lys 150 155tagctgcaga
tggtattggt gtatgtttac agagtttcta taaaagatgt attaagaacc 592aaaacttctt
tactttctct tgcagttgct ctatataact gccatttaac tattattata 652tgtgttgtga
ttagattctt gtctcactac agtatttcct ttactctg
70065159PRTLycopersicon esculentum 65Met Ser Asp Glu Glu His His Phe Glu
Ser Lys Ala Asp Ala Gly Ala1 5 10
15Ser Lys Thr Tyr Pro Gln Gln Ala Gly Thr Ile Arg Lys Gly Gly
His 20 25 30Ile Val Ile Lys
Asn Arg Pro Cys Lys Val Val Glu Val Ser Thr Ser 35
40 45Lys Thr Gly Lys His Gly His Ala Lys Cys His Phe
Val Ala Ile Asp 50 55 60Ile Phe Thr
Gly Lys Lys Leu Glu Asp Ile Val Pro Ser Ser His Asn65 70
75 80Cys Asp Val Pro His Val Asn Arg
Thr Asp Tyr Gln Leu Ile Asp Ile 85 90
95Ser Glu Asp Gly Phe Val Ser Leu Leu Thr Glu Asn Gly Asn
Thr Lys 100 105 110Asp Asp Leu
Arg Leu Pro Thr Asp Asp Thr Leu Leu Ala Gln Val Lys 115
120 125Asp Gly Phe Ala Glu Gly Lys Asp Leu Val Leu
Ser Val Met Ser Ala 130 135 140Met Gly
Glu Glu Gln Ile Cys Gly Ile Lys Asp Ile Gly Pro Lys145
150 15566658DNABrassica napusCDS(1)..(477) 66atg tct gac
gag gag cac cac ttc gag tcc agc gac gcc gga gct tcc 48Met Ser Asp
Glu Glu His His Phe Glu Ser Ser Asp Ala Gly Ala Ser1 5
10 15aaa acc tac cct cag cag gct ggt aac
atc cgc aag ggt ggt cac atc 96Lys Thr Tyr Pro Gln Gln Ala Gly Asn
Ile Arg Lys Gly Gly His Ile 20 25
30gtc atc aag ggc cgt ccc tgc aag gtt gtt gag gtt tcg act tcg aag
144Val Ile Lys Gly Arg Pro Cys Lys Val Val Glu Val Ser Thr Ser Lys
35 40 45act ggg aag cac ggt cac gca
aag tgt cac ttt gtt gct atc gac atc 192Thr Gly Lys His Gly His Ala
Lys Cys His Phe Val Ala Ile Asp Ile 50 55
60ttc act gct aag aag ctc gag gat att gtt ccc tct tcc cac aat tgt
240Phe Thr Ala Lys Lys Leu Glu Asp Ile Val Pro Ser Ser His Asn Cys65
70 75 80gat gtt ccc cat
gtg aac cgt att gac tac cag ttg att gat atc tct 288Asp Val Pro His
Val Asn Arg Ile Asp Tyr Gln Leu Ile Asp Ile Ser 85
90 95gag aat ggc ttt gtt agc ctt ttg acc gac
agt ggt ggc acc aag gac 336Glu Asn Gly Phe Val Ser Leu Leu Thr Asp
Ser Gly Gly Thr Lys Asp 100 105
110gac ctc aag ctt ccc acc gat gat aat ctc agc gct ctg atg aag agt
384Asp Leu Lys Leu Pro Thr Asp Asp Asn Leu Ser Ala Leu Met Lys Ser
115 120 125gga ttc gag gag gga aag gat
gtg gtg gtg tct gtc atg tct tcc atg 432Gly Phe Glu Glu Gly Lys Asp
Val Val Val Ser Val Met Ser Ser Met 130 135
140gga gag gag cag atc tgt gcc gtc aag gaa gtt ggt ggt ggc aag
477Gly Glu Glu Gln Ile Cys Ala Val Lys Glu Val Gly Gly Gly Lys145
150 155taaaacccat tctctgagag aggataatct
tattaccagt ggtcaatgtt ataagaacaa 537gaacttgttt tttttccttt ttctaattta
gatcatttgt gttgtgtttc tttgttgcaa 597gacaaccatt atctattatt ggttttggat
tgtttaaaaa aaaaaaaaaa aaaaaaaaaa 657a
65867159PRTBrassica napus 67Met Ser Asp
Glu Glu His His Phe Glu Ser Ser Asp Ala Gly Ala Ser1 5
10 15Lys Thr Tyr Pro Gln Gln Ala Gly Asn
Ile Arg Lys Gly Gly His Ile 20 25
30Val Ile Lys Gly Arg Pro Cys Lys Val Val Glu Val Ser Thr Ser Lys
35 40 45Thr Gly Lys His Gly His Ala
Lys Cys His Phe Val Ala Ile Asp Ile 50 55
60Phe Thr Ala Lys Lys Leu Glu Asp Ile Val Pro Ser Ser His Asn Cys65
70 75 80Asp Val Pro His
Val Asn Arg Ile Asp Tyr Gln Leu Ile Asp Ile Ser 85
90 95Glu Asn Gly Phe Val Ser Leu Leu Thr Asp
Ser Gly Gly Thr Lys Asp 100 105
110Asp Leu Lys Leu Pro Thr Asp Asp Asn Leu Ser Ala Leu Met Lys Ser
115 120 125Gly Phe Glu Glu Gly Lys Asp
Val Val Val Ser Val Met Ser Ser Met 130 135
140Gly Glu Glu Gln Ile Cys Ala Val Lys Glu Val Gly Gly Gly Lys145
150 155688PRTMycosphaerella fijiensis 68Gly
Leu Asn Arg Ile Gly Asn Leu1 5698PRTMycosphaerella
fijiensis 69Ala Glu Phe Asp Gly Ser Asp Gln1
5701335DNABrassica napusCDS(57)..(1160) 70cttgctagaa ccctaaaact
ccctcccaaa actctccaca tcttccgaga aagaag atg 59
Met
1gag gag gat cgt gtt ctc tcg tct gtc cac tca
acg gtc ttc aag gaa 107Glu Glu Asp Arg Val Leu Ser Ser Val His Ser
Thr Val Phe Lys Glu 5 10
15tcc gaa tcg ttg gaa gga aag tgc gac aaa atc gaa gga tac gat ttc
155Ser Glu Ser Leu Glu Gly Lys Cys Asp Lys Ile Glu Gly Tyr Asp Phe
20 25 30aac caa gga gta aac tac ccg aag
ctc ctc cga tcc atg ctc aca acc 203Asn Gln Gly Val Asn Tyr Pro Lys
Leu Leu Arg Ser Met Leu Thr Thr 35 40
45ggc ttc caa gcc tca aac ctc ggc gac gta att gat gtc gtt aat caa
251Gly Phe Gln Ala Ser Asn Leu Gly Asp Val Ile Asp Val Val Asn Gln50
55 60 65atg cta gag tgg aga
ctc tct gat gaa act ata gca cct gaa gac tgt 299Met Leu Glu Trp Arg
Leu Ser Asp Glu Thr Ile Ala Pro Glu Asp Cys 70
75 80agt gaa gag gag aag gat cca gcg tat aga gag
tcc gtg aag tgt aaa 347Ser Glu Glu Glu Lys Asp Pro Ala Tyr Arg Glu
Ser Val Lys Cys Lys 85 90
95atc ttt cta ggc ttc act tcg aat ctt gtt tcc tct ggt gtt aga gag
395Ile Phe Leu Gly Phe Thr Ser Asn Leu Val Ser Ser Gly Val Arg Glu
100 105 110act att cga tac ctt gtt cag
cat cat atg gtt gat gtt ata gtt act 443Thr Ile Arg Tyr Leu Val Gln
His His Met Val Asp Val Ile Val Thr 115 120
125aca act ggt ggc gta gag gaa gat ctc atc aaa tgc ctt gct cct act
491Thr Thr Gly Gly Val Glu Glu Asp Leu Ile Lys Cys Leu Ala Pro Thr130
135 140 145ttc aaa ggt gat
ttc tct cta ccg ggt gcg tat ctt cgg tca aag gga 539Phe Lys Gly Asp
Phe Ser Leu Pro Gly Ala Tyr Leu Arg Ser Lys Gly 150
155 160ttg aac cgg atc ggg aac ttg ctt gtt cct
aat gat aac tac tgc aag 587Leu Asn Arg Ile Gly Asn Leu Leu Val Pro
Asn Asp Asn Tyr Cys Lys 165 170
175ttt gag gat tgg atc att ccc atc ttt gac cag atg ttg aag gaa cag
635Phe Glu Asp Trp Ile Ile Pro Ile Phe Asp Gln Met Leu Lys Glu Gln
180 185 190aaa gaa gag agt gtg ttg tgg
acg cct tct aaa ttg tta gcg cgg ctg 683Lys Glu Glu Ser Val Leu Trp
Thr Pro Ser Lys Leu Leu Ala Arg Leu 195 200
205ggg aaa gaa ata aat aat gag agt tca tat ctt tat tgg gca tac aag
731Gly Lys Glu Ile Asn Asn Glu Ser Ser Tyr Leu Tyr Trp Ala Tyr Lys210
215 220 225atg aat att cca
gtg ttc tgc cgg ggg tta acc gat ggc tct ctc ggt 779Met Asn Ile Pro
Val Phe Cys Arg Gly Leu Thr Asp Gly Ser Leu Gly 230
235 240gat atg ttg tat ttt cac tca ttt cgt acc
tct ggc ctt gtc atc gat 827Asp Met Leu Tyr Phe His Ser Phe Arg Thr
Ser Gly Leu Val Ile Asp 245 250
255gtt gtg caa gat att aga gct atg aac ggt gaa gca gtc cat gcg act
875Val Val Gln Asp Ile Arg Ala Met Asn Gly Glu Ala Val His Ala Thr
260 265 270cca aga aag aca ggg atg ata
atc ctt gga ggg ggc ttg ccg aag cac 923Pro Arg Lys Thr Gly Met Ile
Ile Leu Gly Gly Gly Leu Pro Lys His 275 280
285cac ata tgt aat gcc aac atg atg cgt aac ggt gcg gat tac gct gtg
971His Ile Cys Asn Ala Asn Met Met Arg Asn Gly Ala Asp Tyr Ala Val290
295 300 305ttt atc aac acc
ggg caa gag ttt gat gga agt gac tcg ggt gca cgc 1019Phe Ile Asn Thr
Gly Gln Glu Phe Asp Gly Ser Asp Ser Gly Ala Arg 310
315 320cct gat gaa gca gtg tct tgg ggt aaa ata
agg gga tct gct aaa act 1067Pro Asp Glu Ala Val Ser Trp Gly Lys Ile
Arg Gly Ser Ala Lys Thr 325 330
335gtc aag gtg tac tgt gat gct acc ata gcc ttc cct ttg ttg gtt gct
1115Val Lys Val Tyr Cys Asp Ala Thr Ile Ala Phe Pro Leu Leu Val Ala
340 345 350gaa aca ttt gcc tcc aag aga
gaa caa agc tgt gag cac aag acc 1160Glu Thr Phe Ala Ser Lys Arg
Glu Gln Ser Cys Glu His Lys Thr 355 360
365taagcccaag aaagcttacg tctcttttat cggtttgttc ttccatcttg ttgttgtacc
1220ctttgtcctg ctttacataa cattcatctc taaaacaata ctacctcctt ttgacaaaaa
1280ataaaaaaaa ttggaaaaat ggtttcacaa gaataaaaaa aaaaaaaaaa aaaaa
133571368PRTBrassica napus 71Met Glu Glu Asp Arg Val Leu Ser Ser Val His
Ser Thr Val Phe Lys1 5 10
15Glu Ser Glu Ser Leu Glu Gly Lys Cys Asp Lys Ile Glu Gly Tyr Asp
20 25 30Phe Asn Gln Gly Val Asn Tyr
Pro Lys Leu Leu Arg Ser Met Leu Thr 35 40
45Thr Gly Phe Gln Ala Ser Asn Leu Gly Asp Val Ile Asp Val Val
Asn 50 55 60Gln Met Leu Glu Trp Arg
Leu Ser Asp Glu Thr Ile Ala Pro Glu Asp65 70
75 80Cys Ser Glu Glu Glu Lys Asp Pro Ala Tyr Arg
Glu Ser Val Lys Cys 85 90
95Lys Ile Phe Leu Gly Phe Thr Ser Asn Leu Val Ser Ser Gly Val Arg
100 105 110Glu Thr Ile Arg Tyr Leu
Val Gln His His Met Val Asp Val Ile Val 115 120
125Thr Thr Thr Gly Gly Val Glu Glu Asp Leu Ile Lys Cys Leu
Ala Pro 130 135 140Thr Phe Lys Gly Asp
Phe Ser Leu Pro Gly Ala Tyr Leu Arg Ser Lys145 150
155 160Gly Leu Asn Arg Ile Gly Asn Leu Leu Val
Pro Asn Asp Asn Tyr Cys 165 170
175Lys Phe Glu Asp Trp Ile Ile Pro Ile Phe Asp Gln Met Leu Lys Glu
180 185 190Gln Lys Glu Glu Ser
Val Leu Trp Thr Pro Ser Lys Leu Leu Ala Arg 195
200 205Leu Gly Lys Glu Ile Asn Asn Glu Ser Ser Tyr Leu
Tyr Trp Ala Tyr 210 215 220Lys Met Asn
Ile Pro Val Phe Cys Arg Gly Leu Thr Asp Gly Ser Leu225
230 235 240Gly Asp Met Leu Tyr Phe His
Ser Phe Arg Thr Ser Gly Leu Val Ile 245
250 255Asp Val Val Gln Asp Ile Arg Ala Met Asn Gly Glu
Ala Val His Ala 260 265 270Thr
Pro Arg Lys Thr Gly Met Ile Ile Leu Gly Gly Gly Leu Pro Lys 275
280 285His His Ile Cys Asn Ala Asn Met Met
Arg Asn Gly Ala Asp Tyr Ala 290 295
300Val Phe Ile Asn Thr Gly Gln Glu Phe Asp Gly Ser Asp Ser Gly Ala305
310 315 320Arg Pro Asp Glu
Ala Val Ser Trp Gly Lys Ile Arg Gly Ser Ala Lys 325
330 335Thr Val Lys Val Tyr Cys Asp Ala Thr Ile
Ala Phe Pro Leu Leu Val 340 345
350Ala Glu Thr Phe Ala Ser Lys Arg Glu Gln Ser Cys Glu His Lys Thr
355 360 365721862DNAMedicago
sativaCDS(293)..(1396) 72gaaaccttct tcttctggag caaagtcgcc attccctacc
tccttcttca ttcttattct 60ctataacaaa cggtccgacc ggatccaagt tgcaccggtt
cgaaccgctt tagttactac 120taacggttcg aaccgttatt tttcaacccg tgacaaacgt
ggaaggcttc gttgtttctt 180cttcttcttc ttaattacca tgcgtttttg tttttctttt
gagtcattga agtcttgttt 240tttgtcgtgt ttctgtcttg agaccgtgaa agagaaaaca
aagagtacga ga atg agt 298
Met Ser
1gaa aca aag caa gaa gat gat aca att atg tcc tca gtt cac tcc act
346Glu Thr Lys Gln Glu Asp Asp Thr Ile Met Ser Ser Val His Ser Thr
5 10 15gtc ttc aaa gaa tcc gaa aat ctc
gca gga aag tgt gtc caa atc gag 394Val Phe Lys Glu Ser Glu Asn Leu
Ala Gly Lys Cys Val Gln Ile Glu 20 25
30ggt tat gat ttc aac cgc ggc gtc gat tat caa cag ctt ctc aaa tca
442Gly Tyr Asp Phe Asn Arg Gly Val Asp Tyr Gln Gln Leu Leu Lys Ser35
40 45 50atg ctc aca act ggt
ttt caa gct tcc aac ttt ggt gat gcc gtt aaa 490Met Leu Thr Thr Gly
Phe Gln Ala Ser Asn Phe Gly Asp Ala Val Lys 55
60 65gtc gtt aat caa atg cta gat tgg agg ttg gtt
gat gaa cca att gat 538Val Val Asn Gln Met Leu Asp Trp Arg Leu Val
Asp Glu Pro Ile Asp 70 75
80gag gat tgt gat gaa gat aag aag gat ttg gag tat agg aaa tct gtt
586Glu Asp Cys Asp Glu Asp Lys Lys Asp Leu Glu Tyr Arg Lys Ser Val
85 90 95aca tgc aaa gtg ttt ttg ggt ttc
act tct aat ctt atc tct tct ggt 634Thr Cys Lys Val Phe Leu Gly Phe
Thr Ser Asn Leu Ile Ser Ser Gly 100 105
110gtt aga gat gtt gtt cgt tac ctt tgt cag cat cac atg gtt cat gta
682Val Arg Asp Val Val Arg Tyr Leu Cys Gln His His Met Val His Val115
120 125 130gtt gtt aca act
aca ggt ggt att gaa gaa gat ctt ata aag tgc ctt 730Val Val Thr Thr
Thr Gly Gly Ile Glu Glu Asp Leu Ile Lys Cys Leu 135
140 145gca cca aca tat aaa gga gag ttc tct ttg
ccc gga gct tat ctt cgc 778Ala Pro Thr Tyr Lys Gly Glu Phe Ser Leu
Pro Gly Ala Tyr Leu Arg 150 155
160tca aaa gga ttg aat cga atc ggt aat tta ttg gtc cct aat gaa aat
826Ser Lys Gly Leu Asn Arg Ile Gly Asn Leu Leu Val Pro Asn Glu Asn
165 170 175tat tgc aaa ttt gag gac tgg
att att cct att ttt gat caa atg ttg 874Tyr Cys Lys Phe Glu Asp Trp
Ile Ile Pro Ile Phe Asp Gln Met Leu 180 185
190aag gag caa aag gaa gag aaa gtg ctg tgg aca ccg tct aag tta ata
922Lys Glu Gln Lys Glu Glu Lys Val Leu Trp Thr Pro Ser Lys Leu Ile195
200 205 210gct cga ttg gga
aaa gag atc aac aat gaa aac tcc tac ctt tac tgg 970Ala Arg Leu Gly
Lys Glu Ile Asn Asn Glu Asn Ser Tyr Leu Tyr Trp 215
220 225gca tat aag aac aat att cca gtt tat tgt
cca gga tta acc gat ggc 1018Ala Tyr Lys Asn Asn Ile Pro Val Tyr Cys
Pro Gly Leu Thr Asp Gly 230 235
240tca ctg ggt gac atg ctg tac ttc cat tcc ttc cac aac cct ggt ctg
1066Ser Leu Gly Asp Met Leu Tyr Phe His Ser Phe His Asn Pro Gly Leu
245 250 255att gtg gac ata gtg caa gat
ata agg gcc atg aat ggt gaa gct gta 1114Ile Val Asp Ile Val Gln Asp
Ile Arg Ala Met Asn Gly Glu Ala Val 260 265
270cat gca aat cct agc aag acg ggc atg att att tta gga ggc ggc ctt
1162His Ala Asn Pro Ser Lys Thr Gly Met Ile Ile Leu Gly Gly Gly Leu275
280 285 290cca aaa cat cac
att tgc aat gcc aat atg atg cgc aat ggt gca gac 1210Pro Lys His His
Ile Cys Asn Ala Asn Met Met Arg Asn Gly Ala Asp 295
300 305tat gcg gtt ttt att aat act gca caa gaa
ttt gat gga agt gat tct 1258Tyr Ala Val Phe Ile Asn Thr Ala Gln Glu
Phe Asp Gly Ser Asp Ser 310 315
320gga gct cgt cca gat gag gct gtt tca tgg ggg aaa ata cga gga tct
1306Gly Ala Arg Pro Asp Glu Ala Val Ser Trp Gly Lys Ile Arg Gly Ser
325 330 335gct aaa act gtt aag gta cat
tgt gat gca acg ata gca ttc cct ctg 1354Ala Lys Thr Val Lys Val His
Cys Asp Ala Thr Ile Ala Phe Pro Leu 340 345
350ctg gtt gct gaa aca ttt gcc tca aga acg tca ccc ctt aat
1396Leu Val Ala Glu Thr Phe Ala Ser Arg Thr Ser Pro Leu Asn355
360 365tgataaaggt ccaccgtcaa aagtaaaagg tgtggctggg
aagtgtttta ccgcagctcc 1456acttgtgagt gccaaatgtt ttggtatgta acttataaga
ccaaggtcgg ctgtatgtca 1516tacttgagtt gaggtcaaag ttcatttgca atgcagtgtg
tttgaggatc ttgatggacc 1576agtttgccat tgacttttaa tttgactgtc ttgttattcg
caaggtccac ataacaagca 1636tttttaccat ttagaaacaa tttattagtc ctgaaggaat
tgagagtcat gaattcagat 1696gtaaattatg caatgctaac tatatttttt tggaactgtg
gtttctctta gatttgaggt 1756gttgaaaact gtaatatcta gagcaaataa gactagaaaa
gtttatcaac tattactgat 1816cagttatagt atcttcaata ttttccagaa aaaaaaaaaa
aaaaaa 186273368PRTMedicago sativa 73Met Ser Glu Thr Lys
Gln Glu Asp Asp Thr Ile Met Ser Ser Val His1 5
10 15Ser Thr Val Phe Lys Glu Ser Glu Asn Leu Ala
Gly Lys Cys Val Gln 20 25
30Ile Glu Gly Tyr Asp Phe Asn Arg Gly Val Asp Tyr Gln Gln Leu Leu
35 40 45Lys Ser Met Leu Thr Thr Gly Phe
Gln Ala Ser Asn Phe Gly Asp Ala 50 55
60Val Lys Val Val Asn Gln Met Leu Asp Trp Arg Leu Val Asp Glu Pro65
70 75 80Ile Asp Glu Asp Cys
Asp Glu Asp Lys Lys Asp Leu Glu Tyr Arg Lys 85
90 95Ser Val Thr Cys Lys Val Phe Leu Gly Phe Thr
Ser Asn Leu Ile Ser 100 105
110Ser Gly Val Arg Asp Val Val Arg Tyr Leu Cys Gln His His Met Val
115 120 125His Val Val Val Thr Thr Thr
Gly Gly Ile Glu Glu Asp Leu Ile Lys 130 135
140Cys Leu Ala Pro Thr Tyr Lys Gly Glu Phe Ser Leu Pro Gly Ala
Tyr145 150 155 160Leu Arg
Ser Lys Gly Leu Asn Arg Ile Gly Asn Leu Leu Val Pro Asn
165 170 175Glu Asn Tyr Cys Lys Phe Glu
Asp Trp Ile Ile Pro Ile Phe Asp Gln 180 185
190Met Leu Lys Glu Gln Lys Glu Glu Lys Val Leu Trp Thr Pro
Ser Lys 195 200 205Leu Ile Ala Arg
Leu Gly Lys Glu Ile Asn Asn Glu Asn Ser Tyr Leu 210
215 220Tyr Trp Ala Tyr Lys Asn Asn Ile Pro Val Tyr Cys
Pro Gly Leu Thr225 230 235
240Asp Gly Ser Leu Gly Asp Met Leu Tyr Phe His Ser Phe His Asn Pro
245 250 255Gly Leu Ile Val Asp
Ile Val Gln Asp Ile Arg Ala Met Asn Gly Glu 260
265 270Ala Val His Ala Asn Pro Ser Lys Thr Gly Met Ile
Ile Leu Gly Gly 275 280 285Gly Leu
Pro Lys His His Ile Cys Asn Ala Asn Met Met Arg Asn Gly 290
295 300Ala Asp Tyr Ala Val Phe Ile Asn Thr Ala Gln
Glu Phe Asp Gly Ser305 310 315
320Asp Ser Gly Ala Arg Pro Asp Glu Ala Val Ser Trp Gly Lys Ile Arg
325 330 335Gly Ser Ala Lys
Thr Val Lys Val His Cys Asp Ala Thr Ile Ala Phe 340
345 350Pro Leu Leu Val Ala Glu Thr Phe Ala Ser Arg
Thr Ser Pro Leu Asn 355 360
365741363DNAMusa acuminataCDS(93)..(1220) 74ggcacgagcg cgcggcgccc
gcaacgaata ttgcagagag taagaaggat cctcgccttt 60gtcaccaaac ccttggtttc
cagcgaggcg ac atg gaa ggc ggc gcc gcg gga 113
Met Glu Gly Gly Ala Ala Gly
1 5ggg cag cga gac cgg gaa acc ctg gac gcg gtg cgg tcg gtg
gtg ttt 161Gly Gln Arg Asp Arg Glu Thr Leu Asp Ala Val Arg Ser Val
Val Phe 10 15 20aag cct tcc gta
tcc ttg gag gag aag cgg ttc ccg agg gtc cag ggg 209Lys Pro Ser Val
Ser Leu Glu Glu Lys Arg Phe Pro Arg Val Gln Gly 25 30
35tac gac ttc aac cgg ggt tgt gac ctc atc ggc ctc ctc
gat tcc atc 257Tyr Asp Phe Asn Arg Gly Cys Asp Leu Ile Gly Leu Leu
Asp Ser Ile40 45 50
55tcc tct acc ggg ttc caa gct tcc aac ctc ggc gac gcc atc gat gtc
305Ser Ser Thr Gly Phe Gln Ala Ser Asn Leu Gly Asp Ala Ile Asp Val
60 65 70atc aat cag atg att gac
tgg agg ctc tcc cat gat gcg ccc acg gaa 353Ile Asn Gln Met Ile Asp
Trp Arg Leu Ser His Asp Ala Pro Thr Glu 75 80
85gat tgc agc gag gaa gag cgc aat ctg gct tac agg caa
tcg gtc acg 401Asp Cys Ser Glu Glu Glu Arg Asn Leu Ala Tyr Arg Gln
Ser Val Thr 90 95 100tgc aag atc
ttt ctg ggc ttc act tcg aac ctt gta tct tct ggc atc 449Cys Lys Ile
Phe Leu Gly Phe Thr Ser Asn Leu Val Ser Ser Gly Ile 105
110 115agg gag ata att cgg ttt ctt gtg cag cac cga atg
gtt gaa gtt tta 497Arg Glu Ile Ile Arg Phe Leu Val Gln His Arg Met
Val Glu Val Leu120 125 130
135gtc aca act gct ggc ggc att gaa gaa gat tta atc aaa tgc ctt gct
545Val Thr Thr Ala Gly Gly Ile Glu Glu Asp Leu Ile Lys Cys Leu Ala
140 145 150cca aca tat aag ggt
gac ttt tct ttg cct gga tcg tat ctg cgt tca 593Pro Thr Tyr Lys Gly
Asp Phe Ser Leu Pro Gly Ser Tyr Leu Arg Ser 155
160 165aaa gga ttg aat cgg ata gga aac ctt ctt gtc cct
aat gac aat tac 641Lys Gly Leu Asn Arg Ile Gly Asn Leu Leu Val Pro
Asn Asp Asn Tyr 170 175 180tgc aaa
ttc gag gac tgg atc atg cca att ctg gac cag atg tta ctt 689Cys Lys
Phe Glu Asp Trp Ile Met Pro Ile Leu Asp Gln Met Leu Leu 185
190 195gaa cag act aca gag aat gta gtt tgg aca cca
tct aaa gtg att gcg 737Glu Gln Thr Thr Glu Asn Val Val Trp Thr Pro
Ser Lys Val Ile Ala200 205 210
215cgc ctt gga aaa gaa ata aat gat gaa agt tca tac ctg tac tgg gca
785Arg Leu Gly Lys Glu Ile Asn Asp Glu Ser Ser Tyr Leu Tyr Trp Ala
220 225 230tac aag aac aat gtt
tct gtc tat tgc ccg gca ttg act gat gga tca 833Tyr Lys Asn Asn Val
Ser Val Tyr Cys Pro Ala Leu Thr Asp Gly Ser 235
240 245ttg ggg gat atg ttg tac tgc cat tca gtg cgg aat
cct ggt tta ctt 881Leu Gly Asp Met Leu Tyr Cys His Ser Val Arg Asn
Pro Gly Leu Leu 250 255 260att gat
att gtg caa gac ata cga gca atg aat gga gaa gct gta cat 929Ile Asp
Ile Val Gln Asp Ile Arg Ala Met Asn Gly Glu Ala Val His 265
270 275gtg ggt ctg aga aag act ggg gtc ata att ctt
ggt ggg ggc ctc cca 977Val Gly Leu Arg Lys Thr Gly Val Ile Ile Leu
Gly Gly Gly Leu Pro280 285 290
295aag cac cat ata tgt aat gcc aac atg ttt cgg aat ggt gca gat tat
1025Lys His His Ile Cys Asn Ala Asn Met Phe Arg Asn Gly Ala Asp Tyr
300 305 310gct gtt tat gtc aac
act gca cag gaa ttt gat gga agt gat tct gga 1073Ala Val Tyr Val Asn
Thr Ala Gln Glu Phe Asp Gly Ser Asp Ser Gly 315
320 325gca gag cct gat gag gcg att tca tgg gga aag ata
aaa ggt tct gcg 1121Ala Glu Pro Asp Glu Ala Ile Ser Trp Gly Lys Ile
Lys Gly Ser Ala 330 335 340aag act
att aaa gtt cat tgt gat gca act att gct ttt cct cta ttg 1169Lys Thr
Ile Lys Val His Cys Asp Ala Thr Ile Ala Phe Pro Leu Leu 345
350 355gta gct gca aca ttt gca aga aag ttt cag gaa
aga aac aac aaa tta 1217Val Ala Ala Thr Phe Ala Arg Lys Phe Gln Glu
Arg Asn Asn Lys Leu360 365 370
375gcc tgatgggggt gcaaaaggtg atcatcttat ttggattcaa ataccttaat
1270Alagtaatctgct aacatctgca gatgctgtat tcttgcaaac caaaaattta
atattagata 1330accgagagcc tacagagggt cctttcaaaa aaa
136375376PRTMusa acuminata 75Met Glu Gly Gly Ala Ala Gly Gly
Gln Arg Asp Arg Glu Thr Leu Asp1 5 10
15Ala Val Arg Ser Val Val Phe Lys Pro Ser Val Ser Leu Glu
Glu Lys 20 25 30Arg Phe Pro
Arg Val Gln Gly Tyr Asp Phe Asn Arg Gly Cys Asp Leu 35
40 45Ile Gly Leu Leu Asp Ser Ile Ser Ser Thr Gly
Phe Gln Ala Ser Asn 50 55 60Leu Gly
Asp Ala Ile Asp Val Ile Asn Gln Met Ile Asp Trp Arg Leu65
70 75 80Ser His Asp Ala Pro Thr Glu
Asp Cys Ser Glu Glu Glu Arg Asn Leu 85 90
95Ala Tyr Arg Gln Ser Val Thr Cys Lys Ile Phe Leu Gly
Phe Thr Ser 100 105 110Asn Leu
Val Ser Ser Gly Ile Arg Glu Ile Ile Arg Phe Leu Val Gln 115
120 125His Arg Met Val Glu Val Leu Val Thr Thr
Ala Gly Gly Ile Glu Glu 130 135 140Asp
Leu Ile Lys Cys Leu Ala Pro Thr Tyr Lys Gly Asp Phe Ser Leu145
150 155 160Pro Gly Ser Tyr Leu Arg
Ser Lys Gly Leu Asn Arg Ile Gly Asn Leu 165
170 175Leu Val Pro Asn Asp Asn Tyr Cys Lys Phe Glu Asp
Trp Ile Met Pro 180 185 190Ile
Leu Asp Gln Met Leu Leu Glu Gln Thr Thr Glu Asn Val Val Trp 195
200 205Thr Pro Ser Lys Val Ile Ala Arg Leu
Gly Lys Glu Ile Asn Asp Glu 210 215
220Ser Ser Tyr Leu Tyr Trp Ala Tyr Lys Asn Asn Val Ser Val Tyr Cys225
230 235 240Pro Ala Leu Thr
Asp Gly Ser Leu Gly Asp Met Leu Tyr Cys His Ser 245
250 255Val Arg Asn Pro Gly Leu Leu Ile Asp Ile
Val Gln Asp Ile Arg Ala 260 265
270Met Asn Gly Glu Ala Val His Val Gly Leu Arg Lys Thr Gly Val Ile
275 280 285Ile Leu Gly Gly Gly Leu Pro
Lys His His Ile Cys Asn Ala Asn Met 290 295
300Phe Arg Asn Gly Ala Asp Tyr Ala Val Tyr Val Asn Thr Ala Gln
Glu305 310 315 320Phe Asp
Gly Ser Asp Ser Gly Ala Glu Pro Asp Glu Ala Ile Ser Trp
325 330 335Gly Lys Ile Lys Gly Ser Ala
Lys Thr Ile Lys Val His Cys Asp Ala 340 345
350Thr Ile Ala Phe Pro Leu Leu Val Ala Ala Thr Phe Ala Arg
Lys Phe 355 360 365Gln Glu Arg Asn
Asn Lys Leu Ala 370 375761451DNAPopulus
deltoidesCDS(8)..(1126) 76gggattt atg aca ggc aaa aaa caa tgg gag gaa gat
ttg cta tca tca 49 Met Thr Gly Lys Lys Gln Trp Glu Glu Asp
Leu Leu Ser Ser 1 5 10gta cgg acc
aca gtg ttt aaa gaa tca gaa gct ctt gat ggg aaa tgc 97Val Arg Thr
Thr Val Phe Lys Glu Ser Glu Ala Leu Asp Gly Lys Cys15 20
25 30att aaa att gaa ggt tat gat ttt
aat caa gga gtg aac tac tct aag 145Ile Lys Ile Glu Gly Tyr Asp Phe
Asn Gln Gly Val Asn Tyr Ser Lys 35 40
45ctt ctc aaa tcc atg gtc tct acc ggg ttt caa gct tcc aac
ctt gga 193Leu Leu Lys Ser Met Val Ser Thr Gly Phe Gln Ala Ser Asn
Leu Gly 50 55 60gat gcc att
caa gtt gtt aat aac atg cta gac tgg agg ctt gct gat 241Asp Ala Ile
Gln Val Val Asn Asn Met Leu Asp Trp Arg Leu Ala Asp 65
70 75gaa gag ata aca gaa gat tgt agt gat gag gag
agg gag ttg gcc tat 289Glu Glu Ile Thr Glu Asp Cys Ser Asp Glu Glu
Arg Glu Leu Ala Tyr 80 85 90aga gag
tct gtg aga tgc aaa ctg ttc ttg ggt ttt aca tca aat ctt 337Arg Glu
Ser Val Arg Cys Lys Leu Phe Leu Gly Phe Thr Ser Asn Leu95
100 105 110gtt tct tca ggt gtc aga gat
aca att cga tat ctt gtt cag cat cat 385Val Ser Ser Gly Val Arg Asp
Thr Ile Arg Tyr Leu Val Gln His His 115
120 125atg gtt gat gta gtg gtt aca acg gca ggt ggc ata
gaa gaa gat ctt 433Met Val Asp Val Val Val Thr Thr Ala Gly Gly Ile
Glu Glu Asp Leu 130 135 140ata
aaa tgc ctg gca cca aca tac aaa ggt gac ttt tct cta ccc ggg 481Ile
Lys Cys Leu Ala Pro Thr Tyr Lys Gly Asp Phe Ser Leu Pro Gly 145
150 155gct caa tta cga tca aaa ggg ttg aat
cga att ggt aac ttg ttg gta 529Ala Gln Leu Arg Ser Lys Gly Leu Asn
Arg Ile Gly Asn Leu Leu Val 160 165
170cct aat gac aac tac tgc aaa ttt gag gat tgg atc att cca atc ttt
577Pro Asn Asp Asn Tyr Cys Lys Phe Glu Asp Trp Ile Ile Pro Ile Phe175
180 185 190gac caa atg ttg
aag gaa caa att gaa gag aat atc acc tgg aca cct 625Asp Gln Met Leu
Lys Glu Gln Ile Glu Glu Asn Ile Thr Trp Thr Pro 195
200 205tct aaa tta ata gct cgc atg ggg aaa gaa
ata aat aat gag agt tca 673Ser Lys Leu Ile Ala Arg Met Gly Lys Glu
Ile Asn Asn Glu Ser Ser 210 215
220tac ctt tat tgg gca tat aag aac gac att cca gta ttc tgt cca ggc
721Tyr Leu Tyr Trp Ala Tyr Lys Asn Asp Ile Pro Val Phe Cys Pro Gly
225 230 235tta aca gat ggt tct cta ggg
gac atg cta tac ttt cat tcc ttc cac 769Leu Thr Asp Gly Ser Leu Gly
Asp Met Leu Tyr Phe His Ser Phe His 240 245
250aac cct ggt cta att gtt gcc ata gtc caa gat att aga gcc atg aat
817Asn Pro Gly Leu Ile Val Ala Ile Val Gln Asp Ile Arg Ala Met Asn255
260 265 270ggt gaa gct gtc
cac gca agt cct aga aaa act ggt atc atc att ctt 865Gly Glu Ala Val
His Ala Ser Pro Arg Lys Thr Gly Ile Ile Ile Leu 275
280 285gga ggt ggg ctt cct aag cat cat ata tgc
aat gcc aat atg atg cgt 913Gly Gly Gly Leu Pro Lys His His Ile Cys
Asn Ala Asn Met Met Arg 290 295
300aac ggt gca gat tat gct gta ttc atc aat aca gca caa gaa ttt gat
961Asn Gly Ala Asp Tyr Ala Val Phe Ile Asn Thr Ala Gln Glu Phe Asp
305 310 315ggg agt gat tct gga gct cat
cct gat gag gct gta tcg tgg ggg aaa 1009Gly Ser Asp Ser Gly Ala His
Pro Asp Glu Ala Val Ser Trp Gly Lys 320 325
330ata cga ggt tct gct aaa act gtt aag gtc cac tgt gat gca act att
1057Ile Arg Gly Ser Ala Lys Thr Val Lys Val His Cys Asp Ala Thr Ile335
340 345 350gct ttt cct ctc
cta gtt gct gaa aca ttt gcc cct agg agg aac aga 1105Ala Phe Pro Leu
Leu Val Ala Glu Thr Phe Ala Pro Arg Arg Asn Arg 355
360 365ttc tgc agc agt act caa agc tagggctgtg
tgcagttctt ggccagaaaa 1156Phe Cys Ser Ser Thr Gln Ser
370ttgattcatt tttatttgta ttatgactga acgatccgca ggatgggtag tgggctccat
1216tgatgccata aacttctttt tttttcccct cagaattaag ggatccgcca gaacacactg
1276ctctcagccc caaaccattg ttgcctctac tgggagtagc ataaccaatt gaattgcgct
1336cctccaagca gcgcctctta gttgcgttat ttattgtaag tagcgcaacc aactaaatta
1396tgctagttcc cacatttatt gactgctatt ttcaaaagaa aaaaaaaaaa aaaaa
145177373PRTPopulus deltoides 77Met Thr Gly Lys Lys Gln Trp Glu Glu Asp
Leu Leu Ser Ser Val Arg1 5 10
15Thr Thr Val Phe Lys Glu Ser Glu Ala Leu Asp Gly Lys Cys Ile Lys
20 25 30Ile Glu Gly Tyr Asp Phe
Asn Gln Gly Val Asn Tyr Ser Lys Leu Leu 35 40
45Lys Ser Met Val Ser Thr Gly Phe Gln Ala Ser Asn Leu Gly
Asp Ala 50 55 60Ile Gln Val Val Asn
Asn Met Leu Asp Trp Arg Leu Ala Asp Glu Glu65 70
75 80Ile Thr Glu Asp Cys Ser Asp Glu Glu Arg
Glu Leu Ala Tyr Arg Glu 85 90
95Ser Val Arg Cys Lys Leu Phe Leu Gly Phe Thr Ser Asn Leu Val Ser
100 105 110Ser Gly Val Arg Asp
Thr Ile Arg Tyr Leu Val Gln His His Met Val 115
120 125Asp Val Val Val Thr Thr Ala Gly Gly Ile Glu Glu
Asp Leu Ile Lys 130 135 140Cys Leu Ala
Pro Thr Tyr Lys Gly Asp Phe Ser Leu Pro Gly Ala Gln145
150 155 160Leu Arg Ser Lys Gly Leu Asn
Arg Ile Gly Asn Leu Leu Val Pro Asn 165
170 175Asp Asn Tyr Cys Lys Phe Glu Asp Trp Ile Ile Pro
Ile Phe Asp Gln 180 185 190Met
Leu Lys Glu Gln Ile Glu Glu Asn Ile Thr Trp Thr Pro Ser Lys 195
200 205Leu Ile Ala Arg Met Gly Lys Glu Ile
Asn Asn Glu Ser Ser Tyr Leu 210 215
220Tyr Trp Ala Tyr Lys Asn Asp Ile Pro Val Phe Cys Pro Gly Leu Thr225
230 235 240Asp Gly Ser Leu
Gly Asp Met Leu Tyr Phe His Ser Phe His Asn Pro 245
250 255Gly Leu Ile Val Ala Ile Val Gln Asp Ile
Arg Ala Met Asn Gly Glu 260 265
270Ala Val His Ala Ser Pro Arg Lys Thr Gly Ile Ile Ile Leu Gly Gly
275 280 285Gly Leu Pro Lys His His Ile
Cys Asn Ala Asn Met Met Arg Asn Gly 290 295
300Ala Asp Tyr Ala Val Phe Ile Asn Thr Ala Gln Glu Phe Asp Gly
Ser305 310 315 320Asp Ser
Gly Ala His Pro Asp Glu Ala Val Ser Trp Gly Lys Ile Arg
325 330 335Gly Ser Ala Lys Thr Val Lys
Val His Cys Asp Ala Thr Ile Ala Phe 340 345
350Pro Leu Leu Val Ala Glu Thr Phe Ala Pro Arg Arg Asn Arg
Phe Cys 355 360 365Ser Ser Thr Gln
Ser 370781488DNAArabidopsis thaliana 78acaataaggc tttaaagccc
ataaaaccct taaatatatc aaagcccaaa agaaacgcct 60tttgcgcttt cccgatcgtg
gtcaacttcc tctgttacca aaaaatctgt accgcaaaat 120cctcgtcgaa gctcgctgct
gcaaccatgt ccgacgagga gcatcacttt gagtccagtg 180acgccggagc gtccaaaacc
taccctcaac aagctggaac catccgtaag aatggttaca 240tcgtcatcaa aaatcgtccc
tgcaaggttt cgttctcaaa catttctcca ctctcttcct 300ctgatcttat tagatctgtt
cattacttag attcctcaga ttcttttttt tgtcacctcc 360acgatgttcg actgatattt
gttcttgtca tcattgttaa attcacattt tattgcactt 420ttgttttagc gaaattatta
aattggtcat cttcagtttt gttcgattag ataagtttta 480ggattttttc ttacacaagt
tactggatca gctgctaaat gtcattttgt gtcgcaggtt 540gttgaggttt caacctcgaa
gactggcaag catggtcatg ctaaatgtca ttttgtagct 600attgatatct tcaccagcaa
gaaactcgaa gatattgttc cttcttccca caattgtgat 660gtatgtgaaa aaagctcctt
tgatcacttt catttcttgt ttgtttcttt caagtcccat 720ttgagatttt gtttttgttg
aattgggttt caggttcctc atgtcaaccg tactgattat 780cagctgattg acatttctga
agatggatat gtatgtgttc ttaaatagca cttgttcctt 840tatatggttt agttacttgt
tctgttttgt aatcattttg caggtcagtt tgttgactga 900taacggtagt accaaggatg
accttaagct ccctaatgat gacactctgc tccaacaggt 960taagttttgc atgttcatca
cattaaatgt tgctagttaa ttaaaatcaa ctctatgtcg 1020atttctgaaa atggaagaaa
aagtgcagag taatgagtga cctgattgtg ttaatgaaac 1080agatcaagag tgggtttgat
gatggaaaag atctagtggt gagtgtgatg tcagctatgg 1140gagaggaaca gatcaatgct
cttaaggaca tcggtcccaa gtgagactaa caaagcctcc 1200cctttgttat gagattcttc
ttcttcttct gtaggcttcc attactcatc ggagattatc 1260ttgtttttgg gtgactccta
ttttggatat ttaaactttt gttaataatg ccatcttctt 1320caaccttttc cttctagatg
gtttttatac ttcttctaat tgattgattc tttatggttg 1380tccaagtgtc aaagtgttcc
acccatatga ttctaacctt ttgatgagcg aagtctttac 1440tcgtgcgtta tgtagagacg
tagaagcaat accacaaaag agtataat 1488791566DNAArabidopsis
thaliana 79aggataataa tacagtaacc ctagaaaggt ttcctccacc ttcctcttcc
cctcctatat 60aaaaaaaatc gacatcgctt ttgctcactt ctctctctta ggtttttttt
cccttctccc 120aatctcatct tctccgaaaa cctttcttct ctcaaatttc tgtgaaaaca
tgtctgacga 180cgagcaccac tttgaggcca gcgaatccgg agcttccaag acctatcctc
aatcagccgg 240taacatccgt aaaggtggtc acatcgtcat caaaaaccgt ccctgcaagg
tctgatttct 300atttcatcat caaacatcgt tctcgatctc tttttcctga ttctagatct
cgtctctgta 360tagtagctcc ttgattttgt ttttatcctc ggatttgacc tggttctgtt
tagtttgaat 420ttttcttata gatcgctact tagatgaata tgatgaatct tatcctgtta
ttttgatggt 480ggtacctctc tagattcgtg gaattttggg aaatgaaaat gaaaaatgga
tagaaatcaa 540gcaatatcag acgacgcctt ttgtgatttt gaatctaagt agtctattga
ttgatttgat 600ttaaacgttt atggagaaca tagatttgat tttgatattt tggttttgat
taggttgttg 660aggtttcgac ttccaaaact ggcaagcacg gtcacgccaa atgtcacttt
gttgctattg 720atatcttcac tgctaagaag cttgaagata ttgttccatc ttcccacaat
tgtgatgtaa 780gttactacac aaactatgta gattcatttt cacagtattt gatatgattg
tgtgatctga 840ctcaaatatt gttcctttct ctttttttct caggttccac atgtgaaccg
tgttgattac 900cagttgattg atatcactga ggatggcttc gtatgttttt ctttatactc
actttcctca 960tcactccagc tttatttatc tattcttgcc ataacttttg tacttgttta
cattataggt 1020gagccttctc actgacagtg gtggcaccaa ggatgatctc aagcttccca
ccgatgatgg 1080tctcaccgcc caggttattt tcttgtcttt tcatactcgc acacaaatga
cttgactttg 1140tattcatctc tcgaattgtg atattgaaaa cagttgttgt gttttgttaa
tgcagatgag 1200gcttggattc gatgagggaa aggatattgt ggtgtctgtc atgtcttcca
tgggagagga 1260gcagatctgt gccgtcaagg aagttggtgg tggcaagtaa acaagtatca
ttcgatatat 1320tattaccagt ttgacaacgg acgtcaatgt tataagaacc aaaagatgtt
tttctttttc 1380ctaatttaga ccctttgtgt gtgtttcttg ttgcaagaca accatatcta
ttggttttgg 1440attgttggaa aagtttgtgt tgaaacattc aaagtttctt atgagatgtt
attcttaaaa 1500ccactttttg tttgttcact ggatatgttt gttcatgaag cttgttttaa
gcaactcttt 1560acatga
156680319DNAArtificial SequenceDescription of Artificial
Sequence Synthetic promoter sequence 80aagacgttcc aaccacgtct
tcaaagcaag tggattgatg tgatatctcc actgacgtaa 60gggatgacgc acaatcccac
tatccttcgc aagacccttc ctctatataa ggaagttcat 120ttcatttgga gaggacacgc
tgaaatcacc agtctctctc tcaagcttgg atcctcgagt 180actagttcag ggagctcgaa
ttgatcctct agagctttcg ttcgtatcat cggtttcgac 240aacgttcgtc aagttcaatg
catcagtttc attgcgcaca caccagaatc ctactgagtt 300tgagtattat ggcattggg
3198122DNAArtificial
SequenceDescription of Artificial Sequence Synthetic Primer
81gggagggact agtgtgcacg cc
228238DNAArtificial SequenceDescription of Artificial Sequence Synthetic
Primer 82gcgaagcggc catggctcga gttttttttt tttttttt
3883521DNAArabidopsis thaliana 83gggagggact agtgtgcacg
ccctgatgaa gctgtgtctt ggggtaaaat taggggttct 60gctaaaaccg ttaaggtata
ctgtgatgct accatagcct tcccattgtt ggttgcagaa 120acatttgcca caaagagaga
ccaaacctgt gagtctaaga cttaagaact gactggttcg 180tacctctggc ctcatcatcg
atgtagtaca agatatcaga gctatgaacg gcgaagctgt 240ccatgcaaat cctaaaaaga
caggcgtttt ggccatggat tcttaaagat cgttgctttt 300tgattttaca ctggagtgac
catataacac tccacattga tgtggctgtg acgcgaattg 360tcttcttgcg aattgtactt
tagtttctct caacctaaaa tgatttgcag attgtgtttt 420cgtttaaaac acaagagtct
tgtagtcaat aatcctttgc cttataaaat tattcagttc 480caacaaaaaa aaaaaaaaaa
ctcgagccat ggccgcttcg c 52184497DNAArabidopsis
thaliana 84ctagtgtgca cgccctgatg aagctgtgtc ttggggtaaa attaggggtt
ctgctaaaac 60cgttaaggta tactgtgatg ctaccatagc cttcccattg ttggttgcag
aaacatttgc 120cacaaagaga gaccaaacct gtgagtctaa gacttaagaa ctgactggtt
cgtacctctg 180gcctcatcat cgatgtagta caagatatca gagctatgaa cggcgaagct
gtccatgcaa 240atcctaaaaa gacaggcgtt ttggccatgg attcttaaag atcgttgctt
tttgatttta 300cactggagtg accatataac actccacatt gatgtggctg tgacgcgaat
tgtcttcttg 360cgaattgtac tttagtttct ctcaacctaa aatgatttgc agattgtgtt
ttcgtttaaa 420acacaagagt cttgtagtca ataatccttt gccttataaa attattcagt
tccaacaaaa 480aaaaaaaaaa aactcga
4978529DNAArtificial SequenceDescription of Artificial
Sequence Synthetic Primer 85ctcgagaaga ataacatctc ataagaaac
298629DNAArtificial SequenceDescription of
Artificial Sequence Synthetic Primer 86gagctcggca agtaaacaag
tatcattcg 2987906DNAArtificial
SequenceDescription of Artificial Sequence Synthetic construct
87cactgaatca aaggccatgg agtcaaagat tcaaatagag gacctaacag aactcgccgt
60aaagactggc gaacagttca tacagagtct cttacgactc aatgacaaga agaaaatctt
120cgtcaacatg gtggagcacg acacgcttgt ctacctccaa aaatatcaaa gatacagtct
180cagaagacca aagggaattg agacttttca acaaagggta atatccggaa acctcctcgg
240attccattgc ccagctatct gtcactttat tgtgaagata gtggaaaagg aaggtggctc
300ctacaaatgc catcattgcg ataaaggaaa ggccatcgtt gaagatgcct ctgccgacag
360tggtcccaaa gatggacccc cacccacgag gagcatcgtg gaaaaagaag acgttccaac
420cacgtcttca aagcaagtgg attgatgtga taacatggtg gagcacgaca cgcttgtcta
480cctccaaaaa tatcaaagat acagtctcag aagaccaaag ggaattgaga cttttcaaca
540aagggtaata tccggaaacc tcctcggatt ccattgccca gctatctgtc actttattgt
600gaagatagtg gaaaaggaag gtggctccta caaatgccat cattgcgata aaggaaaggc
660catcgttgaa gatgcctctg ccgacagtgg tcccaaagat ggacccccac ccacgaggag
720catcgtggaa aaagaagacg ttccaaccac gtcttcaaag caagtggatt gatgtgatat
780ctccactgac gtaagggatg acgcacaatc ccactatcct tcgcaagacc cttcctctat
840ataaggaagt tcatttcatt tggagaggac acgctgaaat caccagtctc tctctaagct
900tggatc
90688205DNAArtificial SequenceDescription of Artificial Sequence
Synthetic construct 88aagaataaca tctcataaga aactttgaat gtttcaacac
aaacttttcc aacaatccaa 60aaccaataga tatggttgtc ttgcaacaag aaacacacac
aaagggtcta aattaggaaa 120aagaaaaaca tcttttggtt cttataacat tgacgtccgt
tgtcaaactg gtaataatat 180atcgaatgat acttgtttac ttgcc
20589662DNAArtificial SequenceDescription of
Artificial Sequence Synthetic construct 89gaattgatcc tctagagctt
tcgttcgtat catcggtttc gacaacgttc gtcaagttca 60atgcatcagt ttcattgcgc
acacaccaga atcctactga gttcgagtat tatggcattg 120ggaaaactgt ttttcttgta
ccatttgttg tgcttgtaat ttactgtgtt ttttattcgg 180ttttcgctat cgaactgtga
aatggaaatg gatggagaag agttaatgaa tgatatggtc 240cttttgttca ttctcaaatt
aatattattt gttttttctc ttatttgttg tgtgttgaat 300ttgaaattat aagagatatg
caaacatttt gttttgagta aaaatgtgtc aaatcgtggc 360ctctaatgac cgaagttaat
atgaggagta aaacacttgt agttgtacca ttatgcttat 420tcactaggca acaaatatat
tttcagacct agaaaagctg caaatgttac tgaatacaag 480tatgtcctct tgtgttttag
acatttatga actttccttt atgtaatttt ccagaatcct 540tgtcagattc taatcattgc
tttataatta tagttatact catggatttg tagttgagta 600tgaaaatatt ttttaatgca
ttttatgact tgccaattga ttgacaacat gcatcaatcg 660at
6629037PRTArabidopsis
thaliana 90Ala Arg Pro Asp Glu Ala Val Ser Trp Gly Lys Ile Arg Gly Ser
Ala1 5 10 15Lys Thr Val
Lys Val Cys Phe Leu Ile Ser Ser His Pro Asn Leu Tyr 20
25 30Leu Thr Gln Trp Phe
359152PRTLycopersicon esculentum 91Gly Ala Arg Pro Asp Glu Ala Val Ser
Trp Gly Lys Ile Arg Gly Gly1 5 10
15Ala Lys Thr Val Lys Val His Cys Asp Ala Thr Ile Ala Phe Pro
Ile 20 25 30Leu Val Ala Glu
Thr Phe Ala Ala Lys Ser Lys Glu Phe Ser Gln Ile 35
40 45Arg Cys Gln Val 5092193PRTArabidopsis thaliana
92Gly Gly Val Glu Glu Asp Leu Ile Lys Cys Leu Ala Pro Thr Phe Lys1
5 10 15Gly Asp Phe Ser Leu Pro
Gly Ala Tyr Leu Arg Ser Lys Gly Leu Asn 20 25
30Arg Ile Gly Asn Leu Leu Val Pro Asn Asp Asn Tyr Cys
Lys Phe Glu 35 40 45Asp Trp Ile
Ile Pro Ile Phe Asp Glu Met Leu Lys Glu Gln Lys Glu 50
55 60Glu Asn Val Leu Trp Thr Pro Ser Lys Leu Leu Ala
Arg Leu Gly Lys65 70 75
80Glu Ile Asn Asn Glu Ser Ser Tyr Leu Tyr Trp Ala Tyr Lys Met Asn
85 90 95Ile Pro Val Phe Cys Pro
Gly Leu Thr Asp Gly Ser Leu Arg Asp Met 100
105 110Leu Tyr Phe His Ser Phe Arg Thr Ser Gly Leu Ile
Ile Asp Val Val 115 120 125Gln Asp
Ile Arg Ala Met Asn Gly Glu Ala Val His Ala Asn Pro Lys 130
135 140Lys Thr Gly Met Ile Ile Leu Gly Gly Gly Leu
Pro Lys His His Ile145 150 155
160Cys Asn Ala Asn Met Met Arg Asn Gly Ala Asp Tyr Ala Val Phe Ile
165 170 175Asn Thr Gly Gln
Glu Phe Asp Gly Ser Asp Ser Gly Ala Arg Pro Asp 180
185 190Glu 93174PRTDianthus caryophyllus 93Arg Arg
Ser Ile Lys Cys Leu Ala Pro Thr Phe Lys Gly Asp Phe Ala1 5
10 15Leu Pro Gly Ala Gln Leu Arg Ser
Lys Gly Leu Asn Arg Ile Gly Asn 20 25
30Leu Leu Val Pro Asn Asp Asn Tyr Cys Lys Phe Glu Asp Trp Ile
Ile 35 40 45Pro Ile Leu Asp Lys
Met Leu Glu Glu Gln Ile Ser Glu Lys Ile Leu 50 55
60Trp Thr Pro Ser Lys Leu Ile Gly Arg Leu Gly Arg Glu Ile
Asn Asp65 70 75 80Glu
Ser Ser Tyr Leu Tyr Trp Ala Phe Lys Asn Asn Ile Pro Val Phe
85 90 95Cys Pro Gly Leu Thr Asp Gly
Ser Leu Gly Asp Met Leu Tyr Phe His 100 105
110Ser Phe Arg Asn Pro Gly Leu Ile Ile Asp Val Val Gln Asp
Ile Arg 115 120 125Ala Val Asn Gly
Glu Ala Val His Ala Ala Pro Arg Lys Thr Gly Met 130
135 140Ile Ile Leu Gly Gly Gly Leu Pro Lys His His Ile
Cys Asn Ala Asn145 150 155
160Met Met Arg Asn Gly Ala Asp Tyr Ala Val Phe Ile Asn Thr
165 1709422PRTArabidopsis thaliana 94Cys Lys Val Val Glu
Val Ser Thr Ser Lys Thr Gly Lys His Gly His1 5
10 15Ala Lys Cys His Phe Val
209522PRTArabidopsis thaliana 95Cys Lys Val Val Glu Val Ser Thr Ser Lys
Thr Gly Lys His Gly His1 5 10
15Ala Lys Cys His Phe Val 209622PRTArabidopsis thaliana
96Cys Lys Val Val Glu Val Ser Thr Ser Lys Thr Gly Lys His Gly His1
5 10 15Ala Lys Cys His Phe Val
209722PRTBrassica napus 97Cys Lys Val Val Glu Val Ser Thr Ser
Lys Thr Gly Lys His Gly His1 5 10
15Ala Lys Cys His Phe Val 209822PRTDianthus
caryophyllus 98Cys Lys Val Val Glu Val Ser Thr Ser Lys Thr Gly Lys His
Gly His1 5 10 15Ala Lys
Cys His Phe Val 209922PRTLycopersicon esculentum 99Cys Lys Val
Val Glu Val Ser Thr Ser Lys Thr Gly Lys His Gly His1 5
10 15Ala Lys Cys His Phe Val
2010022PRTLycopersicon esculentum 100Cys Lys Val Val Glu Val Ser Thr Ser
Lys Thr Gly Lys His Gly His1 5 10
15Ala Lys Cys His Phe Val 2010122PRTLycopersicon
esculentum 101Cys Lys Val Val Glu Val Ser Thr Ser Lys Thr Gly Lys His Gly
His1 5 10 15Ala Lys Cys
His Phe Val 2010222PRTMedicago sativa 102Cys Lys Val Val Glu
Val Ser Thr Ser Lys Thr Gly Lys His Gly His1 5
10 15Ala Lys Cys His Phe Val
2010322PRTMedicago sativa 103Cys Lys Val Val Glu Val Ser Thr Ser Lys Thr
Gly Lys His Gly His1 5 10
15Ala Lys Cys His Phe Val 2010422PRTMedicago sativa 104Cys
Lys Val Val Glu Val Ser Thr Ser Lys Thr Gly Lys His Gly His1
5 10 15Ala Lys Cys His Phe Val
2010522PRTLactuca sativa 105Cys Lys Val Val Glu Val Ser Thr Ser Lys
Thr Gly Lys His Gly His1 5 10
15Ala Lys Cys His Phe Val 2010622PRTUnknown
OrganismDescription of Unknown Organism Tree sequence peptide 106Cys
Lys Val Val Glu Val Ser Thr Ser Lys Thr Gly Lys His Gly His1
5 10 15Ala Lys Cys His Phe Val
20107488DNAMycosphaerella fijiensis 107gggttaaatc gtattggaaa
cttcttagtg ccaaacgaca attactgccg ctttgaagac 60tgggtgatgc caatcctcga
cacaatgctc gaagaacagg aagcatgcaa gggttcgggc 120gaagcaatcc actggactcc
cagcaaaatc atcaaccggc ttggcaagga ggtcaacgac 180gaatcgtccg tgtactactg
ggcatggaag aacgacattc cagtgttctg tccggcgctt 240actgatggca gtctcggaga
catgctgtac ttccacacgt tcaaatcctc accgcagcag 300cttcgagtcg acattgtgga
agacatccga aagatcaaca ccctcgccgt ccgagccaag 360cgcactggca tgatcattct
cggaggcggc attgtcaagc accacatcgc aaatgccaac 420ctgatgcgca atggcgcgga
aagcgcagtg tacatcaata ccgcgccgga attcgacgga 480tccgacca
48810822PRTUnknown
OrganismDescription of Unknown Organism Tree sequence peptide 108Cys
Lys Val Val Glu Val Ser Thr Ser Lys Thr Gly Lys His Gly His1
5 10 15Ala Lys Cys His Phe Val
2010922PRTUnknown OrganismDescription of Unknown Organism Tree
sequence peptide 109Cys Lys Val Val Glu Val Ser Thr Ser Lys Thr Gly
Lys His Gly His1 5 10
15Ala Lys Cys His Phe Val 2011066DNAArabidopsis thaliana
110tgcaaggttg ttgaggtttc aacctcgaag actggcaagc atggtcatgc taaatgtcat
60tttgta
6611166DNAArabidopsis thaliana 111tgcaaggttg ttgaggtttc gacttccaaa
actggcaagc acggtcacgc caaatgtcat 60tttgtt
6611266DNAArabidopsis thaliana
112tgcaaggtgg ttgaggtatc gacttcgaag actgggaagc atggtcacgc caagtgtcac
60tttgtt
6611366DNABrassica napus 113tgcaaggttg ttgaggtttc gacttcgaag actgggaagc
acggtcacgc aaagtgtcac 60tttgtt
6611466DNADianthus caryophyllus 114tgcaaggtgg
ttgaggtttc tacctccaag actggcaagc acggtcatgc caaatgtcac 60tttgtt
6611566DNALycopersicon esculentum 115tgcaaggtgg ttgaggtttc aacttccaag
acaggcaagc acggtcatgc taaatgtcac 60tttgtt
6611666DNALycopersicon esculentum
116tgcaaggttg tggaagtctc tacatccaaa actggcaagc acggtcacgc caaatgtcat
60ttcgtt
6611766DNALycopersicon esculentum 117tgcaaggttg ttgaggtctc cacttccaaa
actggcaagc atggacatgc aaaatgtcac 60tttgtg
6611866DNAMedicago sativa
118tgcaaggtgg ttgaagtttc gacttcgaag accgggaagc atggacatgc caagtgtcat
60tttgtt
6611966DNAMedicago sativa 119tgcaaggttg ttgaggtttc tacttcaaaa acaggaaaac
atggacatgc aaagtgtcac 60tttgtt
6612066DNAMedicago sativa 120tgcaaggtag
ttgaagtttc aacttctaaa actggaaagc atggacatgc aaagtgtcac 60tttgtt
6612166DNALactuca sativa 121tgcaaggttg ttgaagtttc tacctccaag actgggaagc
atgggcatgc taagtgtcac 60tttgtc
6612266DNAUnknown OrganismDescription of Unknown
Organism Tree oligonucleotide sequence 122tgcaaggtcg tggaggtttc
aacctctaaa actggcaagc atggccatgc taaatgtcac 60tttgtt
6612366DNAUnknown
OrganismDescription of Unknown Organism Tree oligonucleotide
sequence 123tgcaaggttg ttgaggtttc cacctcaaag acaggcaagc acggacatgc
taagtgccac 60tttgtg
6612466DNAUnknown OrganismDescription of Unknown Organism
Tree oligonucleotide sequence 124tgcaaggttg tggaggtttc tacctctaaa
actggcaagc acggccatgc caaatgtcac 60tttgtt
6612566DNAUnknown OrganismDescription
of Unknown Organism Tree oligonucleotide sequence 125tgcaaggttg
ttgaggtttc cacctcaaag acaggcaagc atggacatgc taagtgccac 60tttgtg
66126745DNALycopersicon esculentumCDS(55)..(534) 126ttctccacag caaacacaga
gaagttcata gcagaagaag agagagattt agct atg 57
Met
1tct gat gaa gaa cac cat ttt gag tcc aaa gct gat
gct ggt gcc tca 105Ser Asp Glu Glu His His Phe Glu Ser Lys Ala Asp
Ala Gly Ala Ser 5 10 15aaa
act tac cct caa caa gct ggt act att cgc aag aat ggt tat ata 153Lys
Thr Tyr Pro Gln Gln Ala Gly Thr Ile Arg Lys Asn Gly Tyr Ile 20
25 30gtt atc aaa ggc aga cct tgc aag gtt
gtt gag gtc tcc act tcc aaa 201Val Ile Lys Gly Arg Pro Cys Lys Val
Val Glu Val Ser Thr Ser Lys 35 40
45act ggc aag cat gga cat gca aaa tgt cac ttt gtg gca atc gac att
249Thr Gly Lys His Gly His Ala Lys Cys His Phe Val Ala Ile Asp Ile50
55 60 65ttc aat gca aaa aag
ctt gaa gat att gtt cct tca tcc cac aat tgt 297Phe Asn Ala Lys Lys
Leu Glu Asp Ile Val Pro Ser Ser His Asn Cys 70
75 80gat gtg cca cat gtc aat cgt act gac tat cag
ctg att gac ata tct 345Asp Val Pro His Val Asn Arg Thr Asp Tyr Gln
Leu Ile Asp Ile Ser 85 90
95gaa gat ggt ttt gtg tct ctt ctt act gaa aat gga aac acc aaa gac
393Glu Asp Gly Phe Val Ser Leu Leu Thr Glu Asn Gly Asn Thr Lys Asp
100 105 110gac ctc aga ctt ccc acc gat
gac acc ctg ttg aac cag gtt aaa ggt 441Asp Leu Arg Leu Pro Thr Asp
Asp Thr Leu Leu Asn Gln Val Lys Gly 115 120
125gga ttt gag gaa gga aag gat ctc gtt ctg tct gtg atg tct gca atg
489Gly Phe Glu Glu Gly Lys Asp Leu Val Leu Ser Val Met Ser Ala Met130
135 140 145ggt gaa gag cag
atc tgt gct gtg aag gac att ggt acc aag acc 534Gly Glu Glu Gln
Ile Cys Ala Val Lys Asp Ile Gly Thr Lys Thr 150
155 160tagttgtgtg cattctgcag cataaataat tgctttttag
cgaagacgtt ttatatcttg 594ttatcgtggt acctttgcaa tctgttttat cgtgaaaaca
ccttatatct attggcatgg 654cttgaatagt tgaaactcta atagtttctg tttggcataa
ggcaatgaac tggatttgat 714agcagaagta atctacatgt cacttttttt t
745127160PRTLycopersicon esculentum 127Met Ser Asp
Glu Glu His His Phe Glu Ser Lys Ala Asp Ala Gly Ala1 5
10 15Ser Lys Thr Tyr Pro Gln Gln Ala Gly
Thr Ile Arg Lys Asn Gly Tyr 20 25
30Ile Val Ile Lys Gly Arg Pro Cys Lys Val Val Glu Val Ser Thr Ser
35 40 45Lys Thr Gly Lys His Gly His
Ala Lys Cys His Phe Val Ala Ile Asp 50 55
60Ile Phe Asn Ala Lys Lys Leu Glu Asp Ile Val Pro Ser Ser His Asn65
70 75 80Cys Asp Val Pro
His Val Asn Arg Thr Asp Tyr Gln Leu Ile Asp Ile 85
90 95Ser Glu Asp Gly Phe Val Ser Leu Leu Thr
Glu Asn Gly Asn Thr Lys 100 105
110Asp Asp Leu Arg Leu Pro Thr Asp Asp Thr Leu Leu Asn Gln Val Lys
115 120 125Gly Gly Phe Glu Glu Gly Lys
Asp Leu Val Leu Ser Val Met Ser Ala 130 135
140Met Gly Glu Glu Gln Ile Cys Ala Val Lys Asp Ile Gly Thr Lys
Thr145 150 155
16012825DNAArtificial SequenceDescription of Artificial Sequence
Synthetic primer 128gaagctcgag gctgcaacca tgtcc
2512926DNAArtificial SequenceDescription of
Artificial Sequence Synthetic Primer 129ggggagctct tgttagtctc acttgg
26130906DNAArtificial
SequenceDescription of Artificial Sequence Synthetic construct
130cactgaatca aaggccatgg agtcaaagat tcaaatagag gacctaacag aactcgccgt
60aaagactggc gaacagttca tacagagtct cttacgactc aatgacaaga agaaaatctt
120cgtcaacatg gtggagcacg acacgcttgt ctacctccaa aaatatcaaa gatacagtct
180cagaagacca aagggaattg agacttttca acaaagggta atatccggaa acctcctcgg
240attccattgc ccagctatct gtcactttat tgtgaagata gtggaaaagg aaggtggctc
300ctacaaatgc catcattgcg ataaaggaaa ggccatcgtt gaagatgcct ctgccgacag
360tggtcccaaa gatggacccc cacccacgag gagcatcgtg gaaaaagaag acgttccaac
420cacgtcttca aagcaagtgg attgatgtga taacatggtg gagcaccaca cgcttgtcta
480cctccaaaaa tatcaaagat acagtctcag aagaccaaag ggaattgaga cttttcaaca
540aagggtaata tccggaaacc tcctcggatt ccattgccca gctatctgtc actttattgt
600gaagatagtg gaaaaggaag gtggctccta caaatgccat cattgcgata aaggaaaggc
660catcgttgaa gatgcctctg ccgacagtgg tcccaaagat ggacccccac ccacgaggag
720catcgtggaa aaagaagacg ttccaaccac gtcttcaaag caagtggatt gatgtgatat
780ctccactgac gtaagggatg acgcacaatc ccactatcct tcgcaagacc cttcctctat
840ataaggaagt tcatttcatt tggagaggac acgctgaaat caccagtctc tctctaagct
900tggatc
906131495DNAArtificial SequenceDescription of Artificial Sequence
Synthetic construct 131gctgcaacca tgtccgacga ggagcatcac tttgagtcca
gtgacgccgg agcgtccaaa 60acctaccctc aacaagctgg aaccatccgt aagaatggtt
acatcgtcat caaaaatcgt 120ccctgcaagg ttgttgaggt ttcaacctcg aagactggca
agcatggtca tgctaaatgt 180cattttgtag ctattgatat cttcaccagc aagaaactcg
aagatattgt tccttcttcc 240cacaattgtg atgttcctca tgtcaaccgt actgattatc
agctgattga catttctgaa 300gatggatatg tcagtttgtt gactgataac ggtagtacca
aggatgacct taagctccct 360aatgatgaca ctctgctcca acagatcaag agtgggtttg
atgatggaaa agatctagtg 420gtgagtgtaa tgtcagctat gggagaggaa cagatcaatg
ctcttaagga catcggtccc 480aagtgagact aacaa
495132662DNAArtificial SequenceDescription of
Artificial Sequence Synthetic construct 132gaattgatcc tctagagctt
tcgttcgtat catcggtttc gacaacgttc gtcaagttca 60atgcatcagt ttcattgcgc
acacaccaga atcctactga gttcgagtat tatggcattg 120ggaaaactgt ttttcttgta
ccatttgttg tgcttgtaat ttactgtgtt ttttattcgg 180ttttcgctat cgaactgtga
aatggaaatg gatggagaag agttaatgaa tgatatggtc 240cttttgttca ttctcaaatt
aatattattt gttttttctc ttatttgttg tgtgttgaat 300ttgaaattat aagagatatg
caaacatttt gttttgagta aaaatgtgtc aaatcgtggc 360ctctaatgac cgaagttaat
atgaggagta aaacacttgt agttgtacca ttatgcttat 420tcactaggca acaaatatat
tttcagacct agaaaagctg caaatgttac tgaatacaag 480tatgtcctct tgtgttttag
acatttatga actttccttt atgtaatttt ccagaatcct 540tgtcagattc taatcattgc
tttataatta tagttatact catggatttg tagttgagta 600tgaaaatatt ttttaatgca
ttttatgact tgccaattga ttgacaacat gcatcaatcg 660at
66213330DNAArtificial
SequenceDescription of Artificial Sequence Synthetic Primer
133gcgctcgagc tatgtctgat gaagaacacc
3013428DNAArtificial SequenceDescription of Artificial Sequence Synthetic
Primer 134tttgagctcc agaatgcaca caactagg
28135906DNAArtificial SequenceDescription of Artificial
Sequence Synthetic construct 135cactgaatca aaggccatgg agtcaaagat
tcaaatagag gacctaacag aactcgccgt 60aaagactggc gaacagttca tacagagtct
cttacgactc aatgacaaga agaaaatctt 120cgtcaacatg gtggagcacg acacgcttgt
ctacctccaa aaatatcaaa gatacagtct 180cagaagacca aagggaattg agacttttca
acaaagggta atatccggaa acctcctcgg 240attccattgc ccagctatct gtcactttat
tgtgaagata gtggaaaagg aaggtggctc 300ctacaaatgc catcattgcg ataaaggaaa
ggccatcgtt gaagatgcct ctgccgacag 360tggtcccaaa gatggacccc cacccacgag
gagcatcgtg gaaaaagaag acgttccaac 420cacgtcttca aagcaagtgg attgatgtga
taacatggtg gagcacgaca cgcttgtcta 480cctccaaaaa tatcaaagat acagtctcag
aagaccaaag ggaattgaga cttttcaaca 540aagggtaata tccggaaacc tcctcggatt
ccattgccca gctatctgtc actttattgt 600gaagatagtg gaaaaggaag gtggctccta
caaatgccat cattgcgata aaggaaaggc 660catcgttgaa gatgcctctg ccgacagtgg
tcccaaagat ggacccccac ccacgaggag 720catcgtggaa aaagaagacg ttccaaccac
gtcttcaaag caagtggatt gatgtgatat 780ctccactgac gtaagggatg acgcacaatc
ccactatcct tcgcaagacc cttcctctat 840ataaggaagt tcatttcatt tggagaggac
acgctgaaat caccagtctc tctctaagct 900tggatc
906136499DNAArtificial
SequenceDescription of Artificial Sequence Synthetic construct
136ctatgtctga tgaagaacac cattttgagt ccaaagctga tgctggtgcc tcaaaaactt
60accctcaaca agctggtact attcgcaaga atggttatat agttatcaaa ggcagacctt
120gcaaggttgt tgaggtctcc acttccaaaa ctggcaagca tggacatgca aaatgtcact
180ttgtggcaat cgacattttc aatgcaaaaa agcttgaaga tattgttcct tcatcccaca
240attgtgatgt gccacatgtc aatcgtactg actatcagct gattgacata tctgaagatg
300gttttgtgtc tcttcttact gaaaatggaa acaccaaaga cgacctcaga cttcccaccg
360atgacaccct gttgaaccag gttaaaggtg gatttgagga aggaaaggat ctcgttctgt
420ctgtgatgtc tgcaatgggt gaagagcaga tctgtgctgt gaaggacatt ggtaccaaga
480cctagttgtg tgcattctg
499137662DNAArtificial SequenceDescription of Artificial Sequence
Synthetic construct 137gaattgatcc tctagagctt tcgttcgtat catcggtttc
gacaacgttc gtcaagttca 60atgcatcagt ttcattgcgc acacaccaga atcctactga
gttcgagtat tatggcattg 120ggaaaactgt ttttcttgta ccatttgttg tgcttgtaat
ttactgtgtt ttttattcgg 180ttttcgctat cgaactgtga aatggaaatg gatggagaag
agttaatgaa tgatatggtc 240cttttgttca ttctcaaatt aatattattt gttttttctc
ttatttgttg tgtgttgaat 300ttgaaattat aagagatatg caaacatttt gttttgagta
aaaatgtgtc aaatcgtggc 360ctctaatgac cgaagttaat atgaggagta aaacacttgt
agttgtacca ttatgcttat 420tcactaggca acaaatatat tttcagacct agaaaagctg
caaatgttac tgaatacaag 480tatgtcctct tgtgttttag acatttatga actttccttt
atgtaatttt ccagaatcct 540tgtcagattc taatcattgc tttataatta tagttatact
catggatttg tagttgagta 600tgaaaatatt ttttaatgca ttttatgact tgccaattga
ttgacaacat gcatcaatcg 660at
66213829DNAArtificial SequenceDescription of
Artificial Sequence Synthetic Primer 138gcatgtcgac atgtctgacg
aggagcacc 29139906DNAArtificial
SequenceDescription of Artificial Sequence Synthetic construct
139cactgaatca aaggccatgg agtcaaagat tcaaatagag gacctaacag aactcgccgt
60aaagactggc gaacagttca tacagagtct cttacgactc aatgacaaga agaaaatctt
120cgtcaacatg gtggagcacg acacgcttgt ctacctccaa aaatatcaaa gatacagtct
180cagaagacca aagggaattg agacttttca acaaagggta atatccggaa acctcctcgg
240attccattgc ccagctatct gtcactttat tgtgaagata gtggaaaagg aaggtggctc
300ctacaaatgc catcattgcg ataaaggaaa ggccatcgtt gaagatgcct ctgccgacag
360tggtcccaaa gatggacccc cacccacgag gagcatcgtg gaaaaagaag acgttccaac
420cacgtcttca aagcaagtgg attgatgtga taacatggtg gagcacgaca cgcttgtcta
480cctccaaaaa tatcaaagat acagtctcag aagaccaaag ggaattgaga cttttcaaca
540aagggtaata tccggaaacc tcctcggatt ccattgccca gctatctgtc actttattgt
600gaagatagtg gaaaaggaag gtggctccta caaatgccat cattgcgata aaggaaaggc
660catcgttgaa gatgcctctg ccgacagtgg tcccaaagat ggacccccac ccacgaggag
720catcgtggaa aaagaagacg ttccaaccac gtcttcaaag caagtggatt gatgtgatat
780ctccactgac gtaagggatg acgcacaatc ccactatcct tcgcaagacc cttcctctat
840ataaggaagt tcatttcatt tggagaggac acgctgaaat caccagtctc tctctaagct
900tggatc
906140661DNAArtificial SequenceDescription of Artificial Sequence
Synthetic construct 140tcgacatgtc tgacgaggag caccacttcg agtccagcga
cgccggagct tccaaaacct 60accctcagca ggctggtaac atccgcaagg gtggtcacat
cgtcatcaag ggccgtccct 120gcaaggttgt tgaggtttcg acttcgaaga ctgggaagca
cggtcacgca aagtgtcact 180ttgttgctat tgacatcttc actgctaaga agctcgagga
tattgttccc tcttcccaca 240attgtgatgt tccccatgtg aaccgtattg actaccagtt
gattgatatc tctgagaatg 300gctttgttag ccttttgacc gacagtggtg gcaccaagga
cgacctcaag cttcccaccg 360atgataatct cagcgctctg atgaagagtg gattcgagga
gggaaaggat gtggtggtgt 420ctgtcatgtc ttccatggga gaggagcaga tctgtgccgt
caaggaagtt ggtggtggca 480agtaaaaccc attctctgag agaggataat cttattacca
gtggtcaatg ttataagaac 540aagaacttgt tttttttcct ttttctaatt tagatcattt
gtgttgtgtt tctttgttgc 600aagacaacca ttatctatta ttggttttgg attgtttaaa
aaaaaaaaaa aaaaaaaaaa 660a
661141662DNAArtificial SequenceDescription of
Artificial Sequence Synthetic construct 141gaattgatcc tctagagctt
tcgttcgtat catcggtttc gacaacgttc gtcaagttca 60atgcatcagt ttcattgcgc
acacaccaga atcctactga gttcgagtat tatggcattg 120ggaaaactgt ttttcttgta
ccatttgttg tgcttgtaat ttactgtgtt ttttattcgg 180ttttcgctat cgaactgtga
aatggaaatg gatggagaag agttaatgaa tgatatggtc 240cttttgttca ttctcaaatt
aatattattt gttttttctc ttatttgttg tgtgttgaat 300ttgaaattat aagagatatg
caaacatttt gttttgagta aaaatgtgtc aaatcgtggc 360ctctaatgac cgaagttaat
atgaggagta aaacacttgt agttgtacca ttatgcttat 420tcactaggca acaaatatat
tttcagacct agaaaagctg caaatgttac tgaatacaag 480tatgtcctct tgtgttttag
acatttatga actttccttt atgtaatttt ccagaatcct 540tgtcagattc taatcattgc
tttataatta tagttatact catggatttg tagttgagta 600tgaaaatatt ttttaatgca
ttttatgact tgccaattga ttgacaacat gcatcaatcg 660at
662142325DNAArtificial
SequenceDescription of Artificial Sequence Synthetic construct
142ggtgcacgcc ctgatgaagc agtgtcttgg ggtaaaataa ggggatctgc taaaactgtc
60aaggtgtact gtgatgctac catagccttc cctttgttgg ttgctgaaac atttgcctcc
120aagagagaac aaagctgtga gcacaagacc taagcccaag aaagcttacg tctcttttat
180cggtttgttc ttccatcttg ttgttgtacc ctttgtcctg ctttacataa cattcatctc
240taaaacaata ctacctcctt ttgacaaaaa ataaaaaaaa ttggaaaaat ggtttcacaa
300gaataaaaaa aaaaaaaaaa aaaaa
325143906DNAArtificial SequenceDescription of Artificial Sequence
Synthetic construct 143cactgaatca aaggccatgg agtcaaagat tcaaatagag
gacctaacag aactcgccgt 60aaagactggc gaacagttca tacagagtct cttacgactc
aatgacaaga agaaaatctt 120cgtcaacatg gtggagcacg acacgcttgt ctacctccaa
aaatatcaaa gatacagtct 180cagaagacca aagggaattg agacttttca acaaagggta
atatccggaa acctcctcgg 240attccattgc ccagctatct gtcactttat tgtgaagata
gtggaaaagg aaggtggctc 300ctacaaatgc catcattgcg ataaaggaaa ggccatcgtt
gaagatgcct ctgccgacag 360tggtcccaaa gatggacccc cacccacgag gagcatcgtg
gaaaaagaag acgttccaac 420cacgtcttca aagcaagtgg attgatgtga taacatggtg
gagcacgaca cgcttgtcta 480cctccaaaaa tatcaaagat acagtctcag aagaccaaag
ggaattgaga cttttcaaca 540aagggtaata tccggaaacc tcctcggatt ccattgccca
gctatctgtc actttattgt 600gaagatagtg gaaaaggaag gtggctccta caaatgccat
cattgcgata aaggaaaggc 660catcgttgaa gatgcctctg ccgacagtgg tcccaaagat
ggacccccac ccacgaggag 720catcgtggaa aaagaagacg ttccaaccac gtcttcaaag
caagtggatt gatgtgatat 780ctccactgac gtaagggatg acgcacaatc ccactatcct
tcgcaagacc cttcctctat 840ataaggaagt tcatttcatt tggagaggac acgctgaaat
caccagtctc tctctaagct 900tggatc
906144325DNAArtificial SequenceDescription of
Artificial Sequence Synthetic construct 144tttttttttt tttttttttt
tattcttgtg aaaccatttt tccaattttt tttatttttt 60gtcaaaagga ggtagtattg
ttttagagat gaatgttatg taaagcagga caaagggtac 120aacaacaaga tggaagaaca
aaccgataaa agagacgtaa gctttcttgg gcttaggtct 180tgtgctcaca gctttgttct
ctcttggagg caaatgtttc agcaaccaac aaagggaagg 240ctatggtagc atcacagtac
accttgacag ttttagcaga tccccttatt ttaccccaag 300acactgcttc atcagggcgt
gcacc 325145662DNAArtificial
SequenceDescription of Artificial Sequence Synthetic construct
145gaattgatcc tctagagctt tcgttcgtat catcggtttc gacaacgttc gtcaagttca
60atgcatcagt ttcattgcgc acacaccaga atcctactga gttcgagtat tatggcattg
120ggaaaactgt ttttcttgta ccatttgttg tgcttgtaat ttactgtgtt ttttattcgg
180ttttcgctat cgaactgtgc aaatggaaat ggatggagaa gagttaatga atgatatggt
240ccttttgttc attctcaaat taatattatt tgttttttct cttatttgtt gtgtgttgaa
300tttgaaatta taagagatat gcaaacattt tgttttgagt aaaaatgtgt caaatcgtgg
360cctctaatga ccgaagttaa tatgaggagt aaaacacttg tagttgtacc attatgctta
420ttcactaggc aacaaatata ttttcagacc tagaaaagct gaaatgttac tgaatacaag
480tatgtcctct tgtgttttag acatttatga actttccttt atgtaatttt ccagaatcct
540tgtcagattc taatcattgc tttataatta tagttatact catggatttg tagttgagta
600tgaaaatatt ttttaatgca ttttatgact tgccaattga ttgacaacat gcatcaatcg
660at
66214628DNAArtificial SequenceDescription of Artificial Sequence
Synthetic Primer 146aagctcgaga tgtcggacga agagcacc
2814728DNAArtificial SequenceDescription of
Artificial Sequence Synthetic Primer 147gtagagctcc accaatacca
tctgcagc 28148906DNAArtificial
SequenceDescription of Artificial Sequence Synthetic construct
148cactgaatca aaggccatgg agtcaaagat tcaaatagag gacctaacag aactcgccgt
60aaagactggc gaacagttca tacagagtct cttacgactc aatgacaaga agaaaatctt
120cgtcaacatg gtggagcacg acacgcttgt ctacctccaa aaatatcaaa gatacagtct
180cagaagacca aagggaattg agacttttca acaaagggta atatccggaa acctcctcgg
240attccattgc ccagctatct gtcactttat tgtgaagata gtggaaaagg aaggtggctc
300ctacaaatgc catcattgcg ataaaggaaa ggccatcgtt gaagatgcct ctgccgacag
360tggtcccaaa gatggacccc cacccacgag gagcatcgtg gaaaaagaag acgttccaac
420cacgtcttca aagcaagtgg attgatgtga taacatggtg gagcacgaca cgcttgtcta
480cctccaaaaa tatcaaagat acagtctcag aagaccaaag ggaattgaga cttttcaaca
540aagggtaata tccggaaacc tcctcggatt ccattgccca gctatctgtc actttattgt
600gaagatagtg gaaaaggaag gtggctccta caaatgccat cattgcgata aaggaaaggc
660catcgttgaa gatgcctctg ccgacagtgg tcccaaagat ggacccccac ccacgaggag
720catcgtggaa aaagaagacg ttccaaccac gtcttcaaag caagtggatt gatgtgatat
780ctccactgac gtaagggatg acgcacaatc ccactatcct tcgcaagacc cttcctctat
840ataaggaagt tcatttcatt tggagaggac acgctgaaat caccagtctc tctctaagct
900tggatc
906149497DNAArtificial SequenceDescription of Artificial Sequence
Synthetic construct 149atgtcggacg aagagcacca cttcgaatcc aaggccgatg
ccggagcttc aaagacgtat 60cctcaacaag ctggtactat tcgtaaaggt ggtcacatcg
tcataaaaaa tcgtccttgc 120aaggtggttg aagtttcaac ttccaagaca ggcaagcacg
gtcatgctaa atgtcactcg 180tggcaattga cattttcact ggaaagaaac ttgaggatat
tgttccctct tctcacaatt 240gtgatgttcc tcatgtgaat aggactgatt atcaacttat
tgatatctct gaggatggct 300ttgtgagtct gttgactgaa aatggtaaca ccaaggatga
cttgagactc ccaactgatg 360atactcttct ggctcaggtc aaagatggtt ttgctgaggg
gaaagacctg gttctatcag 420tgatgtctgc catgggagag gagcagattt gtggtatcaa
ggacattggc cctaagtagc 480tgcagatggt attggtg
497150662DNAArtificial SequenceDescription of
Artificial Sequence Synthetic construct 150gaattgatcc tctagagctt
tcgttcgtat catcggtttc gacaacgttc gtcaagttca 60atgcatcagt ttcattgcgc
acacaccaga atcctactga gttcgagtat tatggcattg 120ggaaaactgt ttttcttgta
ccatttgttg tgcttgtaat ttactgtgtt ttttattcgg 180ttttcgctat cgaactgtga
aatggaaatg gatggagaag agttaatgaa tgatatggtc 240cttttgttca ttctcaaatt
aatattattt gttttttctc ttatttgttg tgtgttgaat 300ttgaaattat aagagatatg
caaacatttt gttttgagta aaaatgtgtc aaatcgtggc 360ctctaatgac cgaagttaat
atgaggagta aaacacttgt agttgtacca ttatgcttat 420tcactaggca acaaatatat
tttcagacct agaaaagctg caaatgttac tgaatacaag 480tatgtcctct tgtgttttag
acatttatga actttccttt atgtaatttt ccagaatcct 540tgtcagattc taatcattgc
tttataatta tagttatact catggatttg tagttgagta 600tgaaaatatt ttttaatgca
ttttatgact tgccaattga ttgacaacat gcatcaatcg 660at
66215127DNAArtificial
SequenceDescription of Artificial Sequence Synthetic Primer
151cgactcgagc agccatgtct gacgagg
2715229DNAArtificial SequenceDescription of Artificial Sequence Synthetic
Primer 152atcgagctca tcacttgggg ccaatatcc
29153906DNAArtificial SequenceDescription of Artificial
Sequence Synthetic construct 153cactgaatca aaggccatgg agtcaaagat
tcaaatagag gacctaacag aactcgccgt 60aaagactggc gaacagttca tacagagtct
cttacgactc aatgacaaga agaaaatctt 120cgtcaacatg gtggagcacg acacgcttgt
ctacctccaa aaatatcaaa gatacagtct 180cagaagacca aagggaattg agacttttca
acaaagggta atatccggaa acctcctcgg 240attccattgc ccagctatct gtcactttat
tgtgaagata gtggaaaagg aaggtggctc 300ctacaaatgc catcattgcg ataaaggaaa
ggccatcgtt gaagatgcct ctgccgacag 360tggtcccaaa gatggacccc cacccacgag
gagcatcgtg gaaaaagaag acgttccaac 420cacgtcttca aagcaagtgg attgatgtga
taacatggtg gagcacgaca cgcttgtcta 480cctccaaaaa tatcaaagat acagtctcag
aagaccaaag ggaattgaga cttttcaaca 540aagggtaata tccggaaacc tcctcggatt
ccattgccca gctatctgtc actttattgt 600gaagatagtg gaaaaggaag gtggctccta
caaatgccat cattgcgata aaggaaaggc 660catcgttgaa gatgcctctg ccgacagtgg
tcccaaagat ggacccccac ccacgaggag 720catcgtggaa aaagaagacg ttccaaccac
gtcttcaaag caagtggatt gatgtgatat 780ctccactgac gtaagggatg acgcacaatc
ccactatcct tcgcaagacc cttcctctat 840ataaggaagt tcatttcatt tggagaggac
acgctgaaat caccagtctc tctctaagct 900tggatc
906154486DNAArtificial
SequenceDescription of Artificial Sequence Synthetic construct
154cagccatgtc tgacgaggag catcaatttg agtctaaggc tgatgccgga gcatcaaaaa
60cttaccctca acaagctggt actattcgta agaacggtta tatcgtcatc aaaggccgtc
120catgcaaggt tgtggaagtc tctacatcca aaactggcaa gcacggtcac gccaaatgtc
180atttcgttgc tattgacatc ttcactggga agaagcttga ggatattgtc ccctcttcac
240acaattgtga tgtgccccat gttaatcgta cagattatca gcttattgac atctctgaag
300atggatttgt gagtctgctt actgacaatg gtaacaccaa ggatgacctc aggcttccta
360ctgatgaaaa tctgctttca ctgatcaagg acgggtttgc cgagggtaag gacctcgttg
420tgtctgttat gtcagctatg ggtgaggaac agattaatgc tttgaaggat attggcccca
480agtgat
486155662DNAArtificial SequenceDescription of Artificial Sequence
Synthetic construct 155gaattgatcc tctagagctt tcgttcgtat catcggtttc
gacaacgttc gtcaagttca 60atgcatcagt ttcattgcgc acacaccaga atcctactga
gttcgagtat tatggcattg 120ggaaaactgt ttttcttgta ccatttgttg tgcttgtaat
ttactgtgtt ttttattcgg 180ttttcgctat cgaactgtga aatggaaatg gatggagaag
agttaatgaa tgatatggtc 240cttttgttca ttctcaaatt aatattattt gttttttctc
ttatttgttg tgtgttgaat 300ttgaaattat aagagatatg caaacatttt gttttgagta
aaaatgtgtc aaatcgtggc 360ctctaatgac cgaagttaat atgaggagta aaacacttgt
agttgtacca ttatgcttat 420tcactaggca acaaatatat tttcagacct agaaaagctg
caaatgttac tgaatacaag 480tatgtcctct tgtgttttag acatttatga actttccttt
atgtaatttt ccagaatcct 540tgtcagattc taatcattgc tttataatta tagttatact
catggatttg tagttgagta 600tgaaaatatt ttttaatgca ttttatgact tgccaattga
ttgacaacat gcatcaatcg 660at
66215622DNAArtificial SequenceDescription of
Artificial Sequence Synthetic Primer 156cactgctcac tagtttgatg gc
2215738DNAArtificial
SequenceDescription of Artificial Sequence Synthetic Primer
157gcgaagcggc catggctcga gttttttttt tttttttt
38158413DNALactuca sativa 158cactgctcac tagtttgatg gcagtgattc tggtgctcga
cctgatgaag ctgtctcctg 60ggggaaaata cgtggttctg ctaaatctgt caaggtgcac
tgtgatgcaa ctatcgcgtt 120ccctttactt gttgcagaaa catttgctgc aaagagagag
ggggagatga aaaatgttga 180gtcaaccaaa gctttggttt aaaaaggtgg aacagtgtag
gacagggact catttttgat 240attttgtttg ctaaaaaatg gtctttggaa gaatattgat
gcacacaaac aaggagacaa 300tgttactgat cttggagagt gtaatgtaaa atgtctaaat
aatttcaaag cttctcacaa 360caaatcaaac tttaaaaaaa aaaaaaaaaa aactcgagcc
atggccgctt cgc 413159388DNALactuca sativa 159ctagtttgat
ggcagtgatt ctggtgctcg acctgatgaa gctgtctcct gggggaaaat 60acgtggttct
gctaaatctg tcaaggtgca ctgtgatgca actatcgcgt tccctttact 120tgttgcagaa
acatttgctg caaagagaga gggggagatg aaaaatgttg agtcaaccaa 180agctttggtt
taaaaaggtg gaacagtgta ggacagggac tcatttttga tattttgttt 240gctaaaaaat
ggtctttgga agaatattga tgcacacaaa caaggagaca atgttactga 300tcttggagag
tgtacatgta aaatgtctaa ataatttcaa agcttctcac aacaaatcaa 360acttaaaaaa
aaaaaaaaaa aaactcga
38816023DNAArtificial SequenceDescription of Artificial Sequence
Synthetic Primer 160ggnttraayc gnathggnaa ytt
2316123DNAArtificial SequenceDescription of
Artificial Sequence Synthetic Primer 161tgrtcgganc crtcraaytc ngc
23162108DNAMycosphaerella
fijiensis 162cgccaagcta tttaggtgac actatagaat actcaagcta tgcatccaac
gcgttgggag 60ctctcccata tggtcgacct gcaggcggcc gcgaattcac tagtgatt
108163487DNAMycosphaerella fijiensisCDS(1)..(486) 163ggg tta
aat cgt att gga aac ttc tta gtg cca aac gac aat tac tgc 48Gly Leu
Asn Arg Ile Gly Asn Phe Leu Val Pro Asn Asp Asn Tyr Cys1 5
10 15cgc ttt gaa gac tgg gtg atg cca
atc ctc gac aca atg ctc gaa gaa 96Arg Phe Glu Asp Trp Val Met Pro
Ile Leu Asp Thr Met Leu Glu Glu 20 25
30cag gaa gca tgc aag ggt tcg ggc gaa gca atc cac tgg act ccc
agc 144Gln Glu Ala Cys Lys Gly Ser Gly Glu Ala Ile His Trp Thr Pro
Ser 35 40 45aaa atc atc aac cgg
ctt ggc aag gag gtc aac gac gaa tcg tcc gtg 192Lys Ile Ile Asn Arg
Leu Gly Lys Glu Val Asn Asp Glu Ser Ser Val 50 55
60tac tac tgg gca tgg aag aac gac att cca gtg ttc tgt ccg
gcg ctt 240Tyr Tyr Trp Ala Trp Lys Asn Asp Ile Pro Val Phe Cys Pro
Ala Leu65 70 75 80act
gat ggc agt ctc gga gac atg ctg tac ttc cac acg ttc aaa tcc 288Thr
Asp Gly Ser Leu Gly Asp Met Leu Tyr Phe His Thr Phe Lys Ser
85 90 95tca ccg cag cag ctt cga gtc
gac att gtg gaa gac atc cga aag atc 336Ser Pro Gln Gln Leu Arg Val
Asp Ile Val Glu Asp Ile Arg Lys Ile 100 105
110aac acc ctc gcc gtc cga gcc aag cgc act ggc atg atc att
ctc gga 384Asn Thr Leu Ala Val Arg Ala Lys Arg Thr Gly Met Ile Ile
Leu Gly 115 120 125ggc ggc att gtc
aag cac cac atc gca aat gcc aac ctg atg cgc aat 432Gly Gly Ile Val
Lys His His Ile Ala Asn Ala Asn Leu Met Arg Asn 130
135 140ggc gcg gaa agc gca gtg tac atc aat acc gcg ccg
aat tcg acg gat 480Gly Ala Glu Ser Ala Val Tyr Ile Asn Thr Ala Pro
Asn Ser Thr Asp145 150 155
160ccg acc a
487Pro Thr164162PRTMycosphaerella fijiensis 164Gly Leu Asn Arg Ile Gly
Asn Phe Leu Val Pro Asn Asp Asn Tyr Cys1 5
10 15Arg Phe Glu Asp Trp Val Met Pro Ile Leu Asp Thr
Met Leu Glu Glu 20 25 30Gln
Glu Ala Cys Lys Gly Ser Gly Glu Ala Ile His Trp Thr Pro Ser 35
40 45Lys Ile Ile Asn Arg Leu Gly Lys Glu
Val Asn Asp Glu Ser Ser Val 50 55
60Tyr Tyr Trp Ala Trp Lys Asn Asp Ile Pro Val Phe Cys Pro Ala Leu65
70 75 80Thr Asp Gly Ser Leu
Gly Asp Met Leu Tyr Phe His Thr Phe Lys Ser 85
90 95Ser Pro Gln Gln Leu Arg Val Asp Ile Val Glu
Asp Ile Arg Lys Ile 100 105
110Asn Thr Leu Ala Val Arg Ala Lys Arg Thr Gly Met Ile Ile Leu Gly
115 120 125Gly Gly Ile Val Lys His His
Ile Ala Asn Ala Asn Leu Met Arg Asn 130 135
140Gly Ala Glu Ser Ala Val Tyr Ile Asn Thr Ala Pro Asn Ser Thr
Asp145 150 155 160Pro
Thr165214DNAMycosphaerella fijiensis 165aatcgaattc ccgcggccgc catggcggcc
gggagcatgc gacgtcgggc ccaattcgcc 60ctatagtgag tcgtattaca attcactggc
cgtcgtttta caacgtcgtg actgggaaaa 120cctggcggta cccaacttaa tcgccttgca
gcacatcccc ctttcgccag ctggcgtaat 180agcgaagagg cccgcaccga tcgcccttcc
aaca 2141661490DNALactuca sativa
166gctcagcatt tcgtcgccgg tacctccccg ccgcctctcc ctaccggttc atcttcccca
60ccggcgctcc ctctccttca ccggctcgac aatatctttt tcgccctagt gttctacaga
120aacagcccct tagtagcgtg agagtgcagc agcccaagtt gcagatatgg gggaagtcgt
180taaagaggca ggggtgattg agaacttaag gtcggtggtg ttcaaggaat cagagagttt
240agaaggcagc tgtgctaaaa tccagggcta cgatttcaac actgggatta attattcgca
300gattctcaaa tccctaattt ccactggatt ccaagcttct aacctcggtg atgccattga
360aaccgttaat caaatgctag actggaggct ttctcatgag caagtaaccg aagattgcag
420tgaggaagag agtaatccag cttacagaga gtctgtcaag tgcaaaatct ttctagggtt
480cacttcaaac ctcatttctt ctggtgttcg agacattatt cgctatctaa ctcaacatca
540tatggttgat gtgattgtga cgacaaccgg tggcattgag gaagatctga taaaatgtct
600tgcagacaca tacagaggtg aattttctct acctggtgct gccttaaggt caaaaggtct
660taatcgtatt ggtaacttat tggtgccaaa tgataactac tgtaaatttg aggattggat
720tatcccaata ttcgaccaaa tgcttcaaga acaaaataca cagcatgtgt tatggactcc
780atcaaaagta atatcacgtt tggggaagga gattaataat gagagttctt atttatattg
840ggcatataag aacgacattc ctgtgttttg cccaggctta acagacgggt ctctagggga
900catgttatat ttccattcat ttcgcaatcc gggtcttgtt attgatgtag tacaagatat
960acgcgcgatt aacagcgagg ctgtgcatgc aaaccctaga aagacaggga tgataattct
1020aggaggaggg ttaccaaaac atcacatttg taatgcgaat atgatgagaa atggtgcgga
1080ttatgctgtt ttcatcaaca ctgctcagga gtttgatggc agtgattctg gtgctcgacc
1140tgatgaagct gtctcctggg ggaaaatacg tggttctgct aaatctgtca aggtgcactg
1200tgatgcaact atcgcgttcc ctttacttgt tgcagaaaca tttgctgcaa agagagaggg
1260ggagatgaaa aatgttgagt caaccaaagc tttggtttaa aaaggtggaa cagtgtagga
1320cagggactca tttttgatat tttgtttgct aaaaaatggt ctttggaaga atattgatgc
1380acacaaacaa ggagacaatg ttactgatct tggagagtgt acatgtaaaa tgtctaaata
1440atttcaaagc ttctcacaac aaatcaaact taaaaaaaaa aaaaaaaaaa
1490167377PRTLactuca sativa 167Met Gly Glu Val Val Lys Glu Ala Gly Val
Ile Glu Asn Leu Arg Ser1 5 10
15Val Val Phe Lys Glu Ser Glu Ser Leu Glu Gly Ser Cys Ala Lys Ile
20 25 30Gln Gly Tyr Asp Phe Asn
Thr Gly Ile Asn Tyr Ser Gln Ile Leu Lys 35 40
45Ser Leu Ile Ser Thr Gly Phe Gln Ala Ser Asn Leu Gly Asp
Ala Ile 50 55 60Glu Thr Val Asn Gln
Met Leu Asp Trp Arg Leu Ser His Glu Gln Val65 70
75 80Thr Glu Asp Cys Ser Glu Glu Glu Ser Asn
Pro Ala Tyr Arg Glu Ser 85 90
95Val Lys Cys Lys Ile Phe Leu Gly Phe Thr Ser Asn Leu Ile Ser Ser
100 105 110Gly Val Arg Asp Ile
Ile Arg Tyr Leu Thr Gln His His Met Val Asp 115
120 125Val Ile Val Thr Thr Thr Gly Gly Ile Glu Glu Asp
Leu Ile Lys Cys 130 135 140Leu Ala Asp
Thr Tyr Arg Gly Glu Phe Ser Leu Pro Gly Ala Ala Leu145
150 155 160Arg Ser Lys Gly Leu Asn Arg
Ile Gly Asn Leu Leu Val Pro Asn Asp 165
170 175Asn Tyr Cys Lys Phe Glu Asp Trp Ile Ile Pro Ile
Phe Asp Gln Met 180 185 190Leu
Gln Glu Gln Asn Thr Gln His Val Leu Trp Thr Pro Ser Lys Val 195
200 205Ile Ser Arg Leu Gly Lys Glu Ile Asn
Asn Glu Ser Ser Tyr Leu Tyr 210 215
220Trp Ala Tyr Lys Asn Asp Ile Pro Val Phe Cys Pro Gly Leu Thr Asp225
230 235 240Gly Ser Leu Gly
Asp Met Leu Tyr Phe His Ser Phe Arg Asn Pro Gly 245
250 255Leu Val Ile Asp Val Val Gln Asp Ile Arg
Ala Ile Asn Ser Glu Ala 260 265
270Val His Ala Asn Pro Arg Lys Thr Gly Met Ile Ile Leu Gly Gly Gly
275 280 285Leu Pro Lys His His Ile Cys
Asn Ala Asn Met Met Arg Asn Gly Ala 290 295
300Asp Tyr Ala Val Phe Ile Asn Thr Ala Gln Glu Phe Asp Gly Ser
Asp305 310 315 320Ser Gly
Ala Arg Pro Asp Glu Ala Val Ser Trp Gly Lys Ile Arg Gly
325 330 335Ser Ala Lys Ser Val Lys Val
His Cys Asp Ala Thr Ile Ala Phe Pro 340 345
350Leu Leu Val Ala Glu Thr Phe Ala Ala Lys Arg Glu Gly Glu
Met Lys 355 360 365Asn Val Glu Ser
Thr Lys Ala Leu Val 370 37516823DNAArtificial
SequenceDescription of Artificial Sequence Synthetic primer
168tgtgatgcna cnathgcntt ycc
2316935DNAArtificial SequenceDescription of Artificial Sequence Synthetic
primer 169gactgcagtc gacatcgatt tttttttttt ttttt
35170431DNAPetunia sp.CDS(1)..(87) 170tgt gat gca acg atc gcg
ttc ccc ata tta gta gct gag aca ttt gca 48Cys Asp Ala Thr Ile Ala
Phe Pro Ile Leu Val Ala Glu Thr Phe Ala1 5
10 15gct aag aaa aac gaa ttc tca cac ata agg tgc caa
gtt tgacaattga 97Ala Lys Lys Asn Glu Phe Ser His Ile Arg Cys Gln
Val 20 25gaagctgcgt ccaatcacat atctgaatgg
tctattcttg ttagttgcaa catcaattat 157tctgcgcaaa ctgactagaa agggtacatt
caagtgccct gagtttagag aacatttatg 217gttctaataa ttatttttcc cttcccatca
tgttatttgt gggaggagcc atatttagtt 277ctctccctgt gctcgagtgg agagtctctg
atgttgaatt tgagaataga gtagctggtc 337tggaggtgtg gccatagttc ttagtgtgca
taggttttga accttactgt attaatggct 397ctgggttttg gctcaaaaaa aaaaaaaaaa
aaaa 43117129PRTPetunia sp. 171Cys Asp Ala
Thr Ile Ala Phe Pro Ile Leu Val Ala Glu Thr Phe Ala1 5
10 15Ala Lys Lys Asn Glu Phe Ser His Ile
Arg Cys Gln Val 20 25172753DNAMedicago
sativaCDS(65)..(544) 172gccgaaccaa atctctgttc tcattatcgc tcttcggcac
cgccaccgcc actcctgaat 60caat atg tct gac gaa gag cac caa ttt gaa tct
aag gcc gat gcc ggt 109 Met Ser Asp Glu Glu His Gln Phe Glu Ser
Lys Ala Asp Ala Gly 1 5 10
15gct tcc aaa act tat cct caa caa gct ggt acc atc cgt aag aat gga
157Ala Ser Lys Thr Tyr Pro Gln Gln Ala Gly Thr Ile Arg Lys Asn Gly
20 25 30tac ata gtt atc aaa
aac cgt cca tgc aag gtg gtt gaa gtt tcg act 205Tyr Ile Val Ile Lys
Asn Arg Pro Cys Lys Val Val Glu Val Ser Thr 35
40 45tcg aag acc ggg aag cat gga cat gcc aag tgt cat
ttt gtt gct att 253Ser Lys Thr Gly Lys His Gly His Ala Lys Cys His
Phe Val Ala Ile 50 55 60gat ata
ttc act tca aag aag ctt gag gat atc gtg ccg tct tca cat 301Asp Ile
Phe Thr Ser Lys Lys Leu Glu Asp Ile Val Pro Ser Ser His 65
70 75aat tgt gat gtt cct cac gtg aat cgt acc gat
tat cag ctt att gac 349Asn Cys Asp Val Pro His Val Asn Arg Thr Asp
Tyr Gln Leu Ile Asp80 85 90
95att tct gaa gat gga ttt gtg agc ttg ctc act gaa aat gga aac act
397Ile Ser Glu Asp Gly Phe Val Ser Leu Leu Thr Glu Asn Gly Asn Thr
100 105 110aag gat gac ctc aaa
ctt ccc acc gat gac agt ctg ctc act cag atc 445Lys Asp Asp Leu Lys
Leu Pro Thr Asp Asp Ser Leu Leu Thr Gln Ile 115
120 125aag gat gga ttt gct gag gga aag gac ctt gtt gta
tcc gtg atg tct 493Lys Asp Gly Phe Ala Glu Gly Lys Asp Leu Val Val
Ser Val Met Ser 130 135 140gct atg
ggt gag gag cag atc tgt gct ttg aag gat att ggt gga aag 541Ala Met
Gly Glu Glu Gln Ile Cys Ala Leu Lys Asp Ile Gly Gly Lys 145
150 155aac taacttattt ctagctattt gttttgtttg
ctatgatcat ttctctaagc 594Asn160tttcacattg ttttgtttaa gagatctgtc
ttatagattc ttgtgagact agtggtgttg 654gtggtgttta tggatttttg ttggtttatc
tgttactgtt tttctgttgt cgtgtgactt 714cttcaaacga gtatttcctt aaaaaaaaaa
aaaaaaaaa 753173160PRTMedicago sativa 173Met
Ser Asp Glu Glu His Gln Phe Glu Ser Lys Ala Asp Ala Gly Ala1
5 10 15Ser Lys Thr Tyr Pro Gln Gln
Ala Gly Thr Ile Arg Lys Asn Gly Tyr 20 25
30Ile Val Ile Lys Asn Arg Pro Cys Lys Val Val Glu Val Ser
Thr Ser 35 40 45Lys Thr Gly Lys
His Gly His Ala Lys Cys His Phe Val Ala Ile Asp 50 55
60Ile Phe Thr Ser Lys Lys Leu Glu Asp Ile Val Pro Ser
Ser His Asn65 70 75
80Cys Asp Val Pro His Val Asn Arg Thr Asp Tyr Gln Leu Ile Asp Ile
85 90 95Ser Glu Asp Gly Phe Val
Ser Leu Leu Thr Glu Asn Gly Asn Thr Lys 100
105 110Asp Asp Leu Lys Leu Pro Thr Asp Asp Ser Leu Leu
Thr Gln Ile Lys 115 120 125Asp Gly
Phe Ala Glu Gly Lys Asp Leu Val Val Ser Val Met Ser Ala 130
135 140Met Gly Glu Glu Gln Ile Cys Ala Leu Lys Asp
Ile Gly Gly Lys Asn145 150 155
160174743DNAMedicago sativaCDS(51)..(527) 174gctcacacat aacacattac
ccacgcgcgc agagaaagca tcaatcaatc atg tct 56
Met Ser
1gac gaa gaa cat cac ttt gaa ccc gct gcc gat gcc gga
gca tcc aaa 104Asp Glu Glu His His Phe Glu Pro Ala Ala Asp Ala Gly
Ala Ser Lys 5 10 15acc tac cct
caa cag gct ggt act att cgc aag aat ggt tac ata gtc 152Thr Tyr Pro
Gln Gln Ala Gly Thr Ile Arg Lys Asn Gly Tyr Ile Val 20
25 30atc aag tcc agg cct tgc aag gtt gtt gag gtt tct
act tca aaa aca 200Ile Lys Ser Arg Pro Cys Lys Val Val Glu Val Ser
Thr Ser Lys Thr35 40 45
50gga aaa cat gga cat gca aag tgt cac ttt gtt gct att gat att ttc
248Gly Lys His Gly His Ala Lys Cys His Phe Val Ala Ile Asp Ile Phe
55 60 65aat ggg aaa aaa ctt gaa
gat att gtt cct tca tcc cac aat tgt gat 296Asn Gly Lys Lys Leu Glu
Asp Ile Val Pro Ser Ser His Asn Cys Asp 70 75
80gtt cct cat gtg aat cgt act gat tat cag ttg att gat
att tct gaa 344Val Pro His Val Asn Arg Thr Asp Tyr Gln Leu Ile Asp
Ile Ser Glu 85 90 95gat gga ttt
gtg agt ttg ttg act gac aat gga agt acc aaa gat gat 392Asp Gly Phe
Val Ser Leu Leu Thr Asp Asn Gly Ser Thr Lys Asp Asp 100
105 110ctg aag ctt cct act gat gat tca cta ctt act cag
att aaa gat gga 440Leu Lys Leu Pro Thr Asp Asp Ser Leu Leu Thr Gln
Ile Lys Asp Gly115 120 125
130ttt gct gat gga aaa gat ctc gtg gtt tct gtt atg tct gcg atg ggt
488Phe Ala Asp Gly Lys Asp Leu Val Val Ser Val Met Ser Ala Met Gly
135 140 145gaa gag cag ata tgt
gcc ctg aag gac att ggt cct aag tagtaatttt 537Glu Glu Gln Ile Cys
Ala Leu Lys Asp Ile Gly Pro Lys 150
155tcgtatgatt gtgtgctcaa taaacttgtc aatgtggaaa agagcttgaa actcataaaa
597catggtgtgg agaaaaggac catatttatc ttgtctcttt aattattgaa ctctctggtt
657ttggccagat gaattcttta agtgtccctt gtaactttat atttttgttg agtttaatat
717tgaaccctaa aaaaaaaaaa aaaaaa
743175159PRTMedicago sativa 175Met Ser Asp Glu Glu His His Phe Glu Pro
Ala Ala Asp Ala Gly Ala1 5 10
15Ser Lys Thr Tyr Pro Gln Gln Ala Gly Thr Ile Arg Lys Asn Gly Tyr
20 25 30Ile Val Ile Lys Ser Arg
Pro Cys Lys Val Val Glu Val Ser Thr Ser 35 40
45Lys Thr Gly Lys His Gly His Ala Lys Cys His Phe Val Ala
Ile Asp 50 55 60Ile Phe Asn Gly Lys
Lys Leu Glu Asp Ile Val Pro Ser Ser His Asn65 70
75 80Cys Asp Val Pro His Val Asn Arg Thr Asp
Tyr Gln Leu Ile Asp Ile 85 90
95Ser Glu Asp Gly Phe Val Ser Leu Leu Thr Asp Asn Gly Ser Thr Lys
100 105 110Asp Asp Leu Lys Leu
Pro Thr Asp Asp Ser Leu Leu Thr Gln Ile Lys 115
120 125Asp Gly Phe Ala Asp Gly Lys Asp Leu Val Val Ser
Val Met Ser Ala 130 135 140Met Gly Glu
Glu Gln Ile Cys Ala Leu Lys Asp Ile Gly Pro Lys145 150
155176535DNAMedicago sativaCDS(2)..(313) 176c aag tgt cac
ttt gtt gga atc gat att ttc act tcc aaa aag ctc gaa 49Lys Cys His Phe
Val Gly Ile Asp Ile Phe Thr Ser Lys Lys Leu Glu1 5
10 15gat att gtg ccc tct tcc cac aat tgt gat
gtt ccc cat gtg aat cgt 97Asp Ile Val Pro Ser Ser His Asn Cys Asp
Val Pro His Val Asn Arg 20 25
30acc gac tat caa ttg atc gat att tct gaa gat gga ttt gtg agt ctg
145Thr Asp Tyr Gln Leu Ile Asp Ile Ser Glu Asp Gly Phe Val Ser Leu
35 40 45ctt act gaa aat ggt aac acc aag
gat gac ttg aag ctt cct acc gat 193Leu Thr Glu Asn Gly Asn Thr Lys
Asp Asp Leu Lys Leu Pro Thr Asp 50 55
60gac aat ctg ctt act cag atc aag gaa ggt ttt gct gag ggt aag gac
241Asp Asn Leu Leu Thr Gln Ile Lys Glu Gly Phe Ala Glu Gly Lys Asp65
70 75 80ctt gtt gtg tcc gtg
atg tct gct atg ggt gag gag cag att aat gcc 289Leu Val Val Ser Val
Met Ser Ala Met Gly Glu Glu Gln Ile Asn Ala 85
90 95ctg aaa gat att ggt cca aag aac tagcttcaac
tattactgcc tatttgttcc 343Leu Lys Asp Ile Gly Pro Lys Asn
100ttctatttaa gcaaaagata tttttgtatt gtcttattgt tcaacccttt ggttatggac
403actggtcaga ctagtcttac caactgtgtt tatggatgtg acacaaacta tttagcattt
463ttctgtttcc atgctgtccc gagttttttt tataaaaaat acaagtttgt taaaaaaaaa
523aaaaaaaaaa aa
535177104PRTMedicago sativa 177Lys Cys His Phe Val Gly Ile Asp Ile Phe
Thr Ser Lys Lys Leu Glu1 5 10
15Asp Ile Val Pro Ser Ser His Asn Cys Asp Val Pro His Val Asn Arg
20 25 30Thr Asp Tyr Gln Leu Ile
Asp Ile Ser Glu Asp Gly Phe Val Ser Leu 35 40
45Leu Thr Glu Asn Gly Asn Thr Lys Asp Asp Leu Lys Leu Pro
Thr Asp 50 55 60Asp Asn Leu Leu Thr
Gln Ile Lys Glu Gly Phe Ala Glu Gly Lys Asp65 70
75 80Leu Val Val Ser Val Met Ser Ala Met Gly
Glu Glu Gln Ile Asn Ala 85 90
95Leu Lys Asp Ile Gly Pro Lys Asn 100178774DNAMedicago
sativaCDS(1)..(477) 178atg tcg gat gac gag cac cat ttt gag tcc aaa gca
gat gct gga gca 48Met Ser Asp Asp Glu His His Phe Glu Ser Lys Ala
Asp Ala Gly Ala1 5 10
15tca aag aca tat cca caa caa gct ggt act atc cgc aaa aat ggt cac
96Ser Lys Thr Tyr Pro Gln Gln Ala Gly Thr Ile Arg Lys Asn Gly His
20 25 30att gtt atc aaa aac aga ccc
tgc aag gta gtt gaa gtt tca act tct 144Ile Val Ile Lys Asn Arg Pro
Cys Lys Val Val Glu Val Ser Thr Ser 35 40
45aaa act gga aag cat gga cat gca aag tgt cac ttt gtt ggt att
gat 192Lys Thr Gly Lys His Gly His Ala Lys Cys His Phe Val Gly Ile
Asp 50 55 60atc ttc act ggc aaa aaa
ctt gaa gat att gtt cct tct tcc cat aac 240Ile Phe Thr Gly Lys Lys
Leu Glu Asp Ile Val Pro Ser Ser His Asn65 70
75 80tgt gat gtt cct cat gtt aat cgc act gat tat
cag ctc att gat ata 288Cys Asp Val Pro His Val Asn Arg Thr Asp Tyr
Gln Leu Ile Asp Ile 85 90
95tct gag gat gga ttt gtg agt ttg ctg gct gag agt ggt ggt act aag
336Ser Glu Asp Gly Phe Val Ser Leu Leu Ala Glu Ser Gly Gly Thr Lys
100 105 110gat gac ctt aag ctt cca
acc gat gag agt ctg ctt tcg cag atc aag 384Asp Asp Leu Lys Leu Pro
Thr Asp Glu Ser Leu Leu Ser Gln Ile Lys 115 120
125gat gga ttt ggt gag ggt aaa gat ctg att gtg tct gtc atg
tct gct 432Asp Gly Phe Gly Glu Gly Lys Asp Leu Ile Val Ser Val Met
Ser Ala 130 135 140atg ggt gag gag cag
att tgt ggc ctc aag gac att ggc ccc aag 477Met Gly Glu Glu Gln
Ile Cys Gly Leu Lys Asp Ile Gly Pro Lys145 150
155taataaatcc atgaatgctt cttgaaaagg agatgatata tttaatattt gtgtgtgatt
537gttgtgtaaa aggaccaatc taagtttgtg caaattctta caagatttgg taatggttaa
597ctttattgag tctttgaccc cttatatttt tgtggtctaa gatcaatatt ggaagcatta
657agactgttat atgttgtgaa gaagttacat tggctactgg ccagtaacat ttaatcatgt
717gctaatataa gaccctctaa catttctact ataaaaaaaa aaaaaaaaaa aaaaaaa
774179159PRTMedicago sativa 179Met Ser Asp Asp Glu His His Phe Glu Ser
Lys Ala Asp Ala Gly Ala1 5 10
15Ser Lys Thr Tyr Pro Gln Gln Ala Gly Thr Ile Arg Lys Asn Gly His
20 25 30Ile Val Ile Lys Asn Arg
Pro Cys Lys Val Val Glu Val Ser Thr Ser 35 40
45Lys Thr Gly Lys His Gly His Ala Lys Cys His Phe Val Gly
Ile Asp 50 55 60Ile Phe Thr Gly Lys
Lys Leu Glu Asp Ile Val Pro Ser Ser His Asn65 70
75 80Cys Asp Val Pro His Val Asn Arg Thr Asp
Tyr Gln Leu Ile Asp Ile 85 90
95Ser Glu Asp Gly Phe Val Ser Leu Leu Ala Glu Ser Gly Gly Thr Lys
100 105 110Asp Asp Leu Lys Leu
Pro Thr Asp Glu Ser Leu Leu Ser Gln Ile Lys 115
120 125Asp Gly Phe Gly Glu Gly Lys Asp Leu Ile Val Ser
Val Met Ser Ala 130 135 140Met Gly Glu
Glu Gln Ile Cys Gly Leu Lys Asp Ile Gly Pro Lys145 150
155180598DNALactuca sativaCDS(1)..(477) 180atg tcg gac gag
gaa cac cag ttt gaa tca aag gcc gac gcc ggt gct 48Met Ser Asp Glu
Glu His Gln Phe Glu Ser Lys Ala Asp Ala Gly Ala1 5
10 15tcc aag acc tac cct cag caa gcc ggt act
atc cgc aag ggc ggc cac 96Ser Lys Thr Tyr Pro Gln Gln Ala Gly Thr
Ile Arg Lys Gly Gly His 20 25
30atc gtc atc aaa aat cga gct tgc aag gtt gtt gaa gtt tct acc tcc
144Ile Val Ile Lys Asn Arg Ala Cys Lys Val Val Glu Val Ser Thr Ser
35 40 45aag act ggg aag cat ggg cat gct
aag tgt cac ttt gtc gcc att gat 192Lys Thr Gly Lys His Gly His Ala
Lys Cys His Phe Val Ala Ile Asp 50 55
60atc ttc aat ggt aaa aag ctt gaa gat att gtt ccc tca tcc cat aac
240Ile Phe Asn Gly Lys Lys Leu Glu Asp Ile Val Pro Ser Ser His Asn65
70 75 80tgt gat gtt cct cat
gtc aac cgc act gac tac cag cta att gat att 288Cys Asp Val Pro His
Val Asn Arg Thr Asp Tyr Gln Leu Ile Asp Ile 85
90 95tct gag gat gga ttt gtg agt ttg ctg act gaa
aat gga aac act aaa 336Ser Glu Asp Gly Phe Val Ser Leu Leu Thr Glu
Asn Gly Asn Thr Lys 100 105
110gat gac ctc aag ctc cca act gat gat gct ctc ctg act cag atc aag
384Asp Asp Leu Lys Leu Pro Thr Asp Asp Ala Leu Leu Thr Gln Ile Lys
115 120 125gat gga ttt gca gag gga aag
gac ttg gtg gtg agt gtg atg tca gca 432Asp Gly Phe Ala Glu Gly Lys
Asp Leu Val Val Ser Val Met Ser Ala 130 135
140atg gga gag gag cag atc tgt gct ctt aag gat atc ggc cca aag
477Met Gly Glu Glu Gln Ile Cys Ala Leu Lys Asp Ile Gly Pro Lys145
150 155tagtaatcta cttctactaa gtctataaat
gaatccataa acacttgttg cttgcattat 537cagcagaatg ttttctgtta taatagtagc
aatctactct accaaaaaaa aaaaaaaaaa 597a
598181159PRTLactuca sativa 181Met Ser
Asp Glu Glu His Gln Phe Glu Ser Lys Ala Asp Ala Gly Ala1 5
10 15Ser Lys Thr Tyr Pro Gln Gln Ala
Gly Thr Ile Arg Lys Gly Gly His 20 25
30Ile Val Ile Lys Asn Arg Ala Cys Lys Val Val Glu Val Ser Thr
Ser 35 40 45Lys Thr Gly Lys His
Gly His Ala Lys Cys His Phe Val Ala Ile Asp 50 55
60Ile Phe Asn Gly Lys Lys Leu Glu Asp Ile Val Pro Ser Ser
His Asn65 70 75 80Cys
Asp Val Pro His Val Asn Arg Thr Asp Tyr Gln Leu Ile Asp Ile
85 90 95Ser Glu Asp Gly Phe Val Ser
Leu Leu Thr Glu Asn Gly Asn Thr Lys 100 105
110Asp Asp Leu Lys Leu Pro Thr Asp Asp Ala Leu Leu Thr Gln
Ile Lys 115 120 125Asp Gly Phe Ala
Glu Gly Lys Asp Leu Val Val Ser Val Met Ser Ala 130
135 140Met Gly Glu Glu Gln Ile Cys Ala Leu Lys Asp Ile
Gly Pro Lys145 150 155182614DNALactuca
sativaCDS(1)..(327) 182ggg aag cac gga cac gcc aag tgt cat ttc gtt gcc
att gat atc ttc 48Gly Lys His Gly His Ala Lys Cys His Phe Val Ala
Ile Asp Ile Phe1 5 10
15aat gga aaa aag ctt gaa gat atc gtt ccc tct tcc cac aat tgt gat
96Asn Gly Lys Lys Leu Glu Asp Ile Val Pro Ser Ser His Asn Cys Asp
20 25 30gtt cca cat gtc aac cga act
gac tac cag ctc att gac att tct gaa 144Val Pro His Val Asn Arg Thr
Asp Tyr Gln Leu Ile Asp Ile Ser Glu 35 40
45gat gga ttt gtg agt ctg ttg act gaa aat ggt ggc act aag gat
gat 192Asp Gly Phe Val Ser Leu Leu Thr Glu Asn Gly Gly Thr Lys Asp
Asp 50 55 60ctc aag ctt cca acc gat
gat agt ctg ctt aca cag att aag gat gga 240Leu Lys Leu Pro Thr Asp
Asp Ser Leu Leu Thr Gln Ile Lys Asp Gly65 70
75 80ttt gct gaa gga aag gat ctt gtg gtg aca gtg
atg tct gcg atg gga 288Phe Ala Glu Gly Lys Asp Leu Val Val Thr Val
Met Ser Ala Met Gly 85 90
95gag gag cag atc tgt gct ctt aag gat att ggt ccc aag taactttacc
337Glu Glu Gln Ile Cys Ala Leu Lys Asp Ile Gly Pro Lys 100
105ttgtgtggat tgagagagat tttatatgaa gaactgcgtt tgctagattt
gtggtgatgg 397ttatttaaga aaaaaaaact ttattaggtt tcttttttct attttacaaa
tttttgtttt 457gtttttggtt attgttgatg tgctaaaaaa gtcttgaaat ttgggcttgg
ggagaggatg 517ggcaatatgt acgtacactt tgtatctcga tatgtttggg atgcgacttg
tgaacttatg 577gagtggttat tctgttaaaa aaaaaaaaaa aaaaaaa
614183109PRTLactuca sativa 183Gly Lys His Gly His Ala Lys Cys
His Phe Val Ala Ile Asp Ile Phe1 5 10
15Asn Gly Lys Lys Leu Glu Asp Ile Val Pro Ser Ser His Asn
Cys Asp 20 25 30Val Pro His
Val Asn Arg Thr Asp Tyr Gln Leu Ile Asp Ile Ser Glu 35
40 45Asp Gly Phe Val Ser Leu Leu Thr Glu Asn Gly
Gly Thr Lys Asp Asp 50 55 60Leu Lys
Leu Pro Thr Asp Asp Ser Leu Leu Thr Gln Ile Lys Asp Gly65
70 75 80Phe Ala Glu Gly Lys Asp Leu
Val Val Thr Val Met Ser Ala Met Gly 85 90
95Glu Glu Gln Ile Cys Ala Leu Lys Asp Ile Gly Pro Lys
100 105184536DNALactuca sativaCDS(1)..(339)
184tcg aag act ggt aag cac ggt cat gcg aag tgt cac ttt gtt gca atc
48Ser Lys Thr Gly Lys His Gly His Ala Lys Cys His Phe Val Ala Ile1
5 10 15gac atc ttc aat gga aaa
aag ctt gaa gat atc gtc cca tct tcc cat 96Asp Ile Phe Asn Gly Lys
Lys Leu Glu Asp Ile Val Pro Ser Ser His 20 25
30aac tgt gat gtc cct cat gtc aac cgc act gac tac cag
cta att gac 144Asn Cys Asp Val Pro His Val Asn Arg Thr Asp Tyr Gln
Leu Ile Asp 35 40 45atc tct gaa
gat gga ttt gtg agt ttg ttg act gaa aat ggc aac acc 192Ile Ser Glu
Asp Gly Phe Val Ser Leu Leu Thr Glu Asn Gly Asn Thr 50
55 60aag gat gac ctg aaa ctg cca acc gat gat gct ctt
ttg act cag atc 240Lys Asp Asp Leu Lys Leu Pro Thr Asp Asp Ala Leu
Leu Thr Gln Ile65 70 75
80aag gag gga ttt gga gag ggg aag gat ttg gtt gtg agt gtg atg tct
288Lys Glu Gly Phe Gly Glu Gly Lys Asp Leu Val Val Ser Val Met Ser
85 90 95gcc atg gga gag gag cag
ata tgt gct gtt aag gac att tct ggt ccg 336Ala Met Gly Glu Glu Gln
Ile Cys Ala Val Lys Asp Ile Ser Gly Pro 100
105 110aag tagactgatc tttttatatc aacagaaagg tatgttcttg
ttcttgttgt 389Lystgttgttgtt gtaatagtag cagcatttta ctttgtgcac
tactatgact agctcatttg 449ggctacgaac agtgtatttt attggtttgt gtactttgaa
atttgaattc ctccatgttg 509tcaagtgtgc aaaaaaaaaa aaaaaaa
536185113PRTLactuca sativa 185Ser Lys Thr Gly Lys
His Gly His Ala Lys Cys His Phe Val Ala Ile1 5
10 15Asp Ile Phe Asn Gly Lys Lys Leu Glu Asp Ile
Val Pro Ser Ser His 20 25
30Asn Cys Asp Val Pro His Val Asn Arg Thr Asp Tyr Gln Leu Ile Asp
35 40 45Ile Ser Glu Asp Gly Phe Val Ser
Leu Leu Thr Glu Asn Gly Asn Thr 50 55
60Lys Asp Asp Leu Lys Leu Pro Thr Asp Asp Ala Leu Leu Thr Gln Ile65
70 75 80Lys Glu Gly Phe Gly
Glu Gly Lys Asp Leu Val Val Ser Val Met Ser 85
90 95Ala Met Gly Glu Glu Gln Ile Cys Ala Val Lys
Asp Ile Ser Gly Pro 100 105
110Lys186471DNALactuca sativa 186tcgaagacag ggaagcacgg tcatgccaag
tgtcactttg ttgcaatcga catttttaac 60gggaaaaagc ttgaagatat tgttccatca
tcccataatt gtgatgtccc ccatgtcaat 120cgcaccgatt accagctaat tgatatttct
gaagatggat ttgtgagttt gctgactgaa 180aatggtaaca ccaaagatga cctcaaactc
cccaccgatg atgctctcct cactcagata 240aaggatggat ttggagaggg gaagggcctg
gttgtgactg ttatgtcagc aatgggagag 300gaacagatat gtgctctcaa ggacattggc
cccaaataac ttttcaaaaa tccacaaaaa 360cacacactat aatattatta taacatttgg
attaacagaa aacaagttgt gtttttattt 420gttttatcat gtacagatgt tatggattaa
aaaaaaaaaa aaaaaaaaaa a 471187112PRTLactuca sativa 187Ser Lys
Thr Gly Lys His Gly His Ala Lys Cys His Phe Val Ala Ile1 5
10 15Asp Ile Phe Asn Gly Lys Lys Leu
Glu Asp Ile Val Pro Ser Ser His 20 25
30Asn Cys Asp Val Pro His Val Asn Arg Thr Asp Tyr Gln Leu Ile
Asp 35 40 45Ile Ser Glu Asp Gly
Phe Val Ser Leu Leu Thr Glu Asn Gly Asn Thr 50 55
60Lys Asp Asp Leu Lys Leu Pro Thr Asp Asp Ala Leu Leu Thr
Gln Ile65 70 75 80Lys
Asp Gly Phe Gly Glu Gly Lys Gly Leu Val Val Thr Val Met Ser
85 90 95Ala Met Gly Glu Glu Gln Ile
Cys Ala Leu Lys Asp Ile Gly Pro Lys 100 105
110188700DNALycopersicon esculentumCDS(56)..(532)
188aaatttctcc ttctccttaa tcctctccac cggcgaaccg gcgaagatca aaacg atg
58 Met
1tcg gac gaa gag cac
cac ttc gaa tcc aag gcc gat gcc gga gct tca 106Ser Asp Glu Glu His
His Phe Glu Ser Lys Ala Asp Ala Gly Ala Ser 5
10 15aag acg tat cct caa caa gct ggt act att cgt aaa
ggt ggt cac atc 154Lys Thr Tyr Pro Gln Gln Ala Gly Thr Ile Arg Lys
Gly Gly His Ile 20 25 30gtc ata
aaa aat cgt cct tgc aag gtg gtt gaa gtt tca act tcc aag 202Val Ile
Lys Asn Arg Pro Cys Lys Val Val Glu Val Ser Thr Ser Lys 35
40 45aca ggc aag cac ggt cat gct aaa tgt cac ttc
gtg gca att gac att 250Thr Gly Lys His Gly His Ala Lys Cys His Phe
Val Ala Ile Asp Ile50 55 60
65ttc act gga aag aaa ctt gag gat att gtt ccc tct tct cac aat tgt
298Phe Thr Gly Lys Lys Leu Glu Asp Ile Val Pro Ser Ser His Asn Cys
70 75 80gat gtt cct cat gtg
aat agg act gat tat caa ctt att gat atc tct 346Asp Val Pro His Val
Asn Arg Thr Asp Tyr Gln Leu Ile Asp Ile Ser 85
90 95gag gat ggc ttt gtg agt ctg ttg act gaa aat ggt
aac acc aag gat 394Glu Asp Gly Phe Val Ser Leu Leu Thr Glu Asn Gly
Asn Thr Lys Asp 100 105 110gac ttg
aga ctc cca act gat gat act ctt ctg gct cag gtc aaa gat 442Asp Leu
Arg Leu Pro Thr Asp Asp Thr Leu Leu Ala Gln Val Lys Asp 115
120 125ggt ttt gct gag ggg aaa gac ctg gtt cta tca
gtg atg tct gcc atg 490Gly Phe Ala Glu Gly Lys Asp Leu Val Leu Ser
Val Met Ser Ala Met130 135 140
145gga gag gag cag att tgt ggt atc aag gac att ggc cct aag
532Gly Glu Glu Gln Ile Cys Gly Ile Lys Asp Ile Gly Pro Lys
150 155tagctgcaga tggtattggt gtatgtttac agagtttcta
taaaagatgt attaagaacc 592aaaacttctt tactttctct tgcagttgct ctatataact
gccatttaac tattattata 652tgtgttgtga ttagattctt gtctcactac agtatttcct
ttactctg 700189159PRTLycopersicon esculentum 189Met Ser
Asp Glu Glu His His Phe Glu Ser Lys Ala Asp Ala Gly Ala1 5
10 15Ser Lys Thr Tyr Pro Gln Gln Ala
Gly Thr Ile Arg Lys Gly Gly His 20 25
30Ile Val Ile Lys Asn Arg Pro Cys Lys Val Val Glu Val Ser Thr
Ser 35 40 45Lys Thr Gly Lys His
Gly His Ala Lys Cys His Phe Val Ala Ile Asp 50 55
60Ile Phe Thr Gly Lys Lys Leu Glu Asp Ile Val Pro Ser Ser
His Asn65 70 75 80Cys
Asp Val Pro His Val Asn Arg Thr Asp Tyr Gln Leu Ile Asp Ile
85 90 95Ser Glu Asp Gly Phe Val Ser
Leu Leu Thr Glu Asn Gly Asn Thr Lys 100 105
110Asp Asp Leu Arg Leu Pro Thr Asp Asp Thr Leu Leu Ala Gln
Val Lys 115 120 125Asp Gly Phe Ala
Glu Gly Lys Asp Leu Val Leu Ser Val Met Ser Ala 130
135 140Met Gly Glu Glu Gln Ile Cys Gly Ile Lys Asp Ile
Gly Pro Lys145 150
155190764DNALycopersicon esculentumCDS(43)..(522) 190aaagaatcct
agagagagaa agggaatcct agagagagaa gc atg tcg gac gaa 54
Met Ser Asp Glu
1gaa cac cat ttt gag tca aag gca gat gct ggt gcc
tca aaa act ttc 102Glu His His Phe Glu Ser Lys Ala Asp Ala Gly Ala
Ser Lys Thr Phe5 10 15
20cca cag caa gct gga acc atc cgt aag aat ggt tac atc gtt atc aaa
150Pro Gln Gln Ala Gly Thr Ile Arg Lys Asn Gly Tyr Ile Val Ile Lys
25 30 35ggc cgt ccc tgc aag gtt
gtt gag gtc tcc act tca aaa act gga aaa 198Gly Arg Pro Cys Lys Val
Val Glu Val Ser Thr Ser Lys Thr Gly Lys 40 45
50cac gga cat gct aaa tgt cac ttt gtg gca att gac att
ttc aat gga 246His Gly His Ala Lys Cys His Phe Val Ala Ile Asp Ile
Phe Asn Gly 55 60 65aag aaa ctg
gaa gat atc gtt ccg tcc tcc cac aat tgt gat gtg cca 294Lys Lys Leu
Glu Asp Ile Val Pro Ser Ser His Asn Cys Asp Val Pro 70
75 80cat gtt aac cgt acc gac tat cag ctg att gat atc
tct gaa gat ggt 342His Val Asn Arg Thr Asp Tyr Gln Leu Ile Asp Ile
Ser Glu Asp Gly85 90 95
100ttt gtc tca ctt ctt act gaa agt gga aac acc aag gat gac ctc agg
390Phe Val Ser Leu Leu Thr Glu Ser Gly Asn Thr Lys Asp Asp Leu Arg
105 110 115ctt ccc acc gat gaa
aat ctg ctg aag cag gtt aaa gat ggg ttc cag 438Leu Pro Thr Asp Glu
Asn Leu Leu Lys Gln Val Lys Asp Gly Phe Gln 120
125 130gaa gga aag gat ctt gtg gtg tct gtt atg tct gcg
atg ggc gaa gag 486Glu Gly Lys Asp Leu Val Val Ser Val Met Ser Ala
Met Gly Glu Glu 135 140 145cag att
aac gcc gtt aag gat gtt ggt acc aag aat tagttatgtc 532Gln Ile
Asn Ala Val Lys Asp Val Gly Thr Lys Asn 150 155
160atggcagcat ataatcactg ccaaagcttt aagacattat catatcctaa
tgtggtactt 592tgatatcact agattataaa ctgtgttatt tgcactgttc aaaacaaaag
aaagaaaact 652gctgttatgg ctagagaaag tattggcttt gagcttttga cagcacagtt
gaactatgtg 712aaaattctac tttttttttt ttgggtaaaa tactgctcgt ttaatgtttt
gc 764191160PRTLycopersicon esculentum 191Met Ser Asp Glu Glu
His His Phe Glu Ser Lys Ala Asp Ala Gly Ala1 5
10 15Ser Lys Thr Phe Pro Gln Gln Ala Gly Thr Ile
Arg Lys Asn Gly Tyr 20 25
30Ile Val Ile Lys Gly Arg Pro Cys Lys Val Val Glu Val Ser Thr Ser
35 40 45Lys Thr Gly Lys His Gly His Ala
Lys Cys His Phe Val Ala Ile Asp 50 55
60Ile Phe Asn Gly Lys Lys Leu Glu Asp Ile Val Pro Ser Ser His Asn65
70 75 80Cys Asp Val Pro His
Val Asn Arg Thr Asp Tyr Gln Leu Ile Asp Ile 85
90 95Ser Glu Asp Gly Phe Val Ser Leu Leu Thr Glu
Ser Gly Asn Thr Lys 100 105
110Asp Asp Leu Arg Leu Pro Thr Asp Glu Asn Leu Leu Lys Gln Val Lys
115 120 125Asp Gly Phe Gln Glu Gly Lys
Asp Leu Val Val Ser Val Met Ser Ala 130 135
140Met Gly Glu Glu Gln Ile Asn Ala Val Lys Asp Val Gly Thr Lys
Asn145 150 155
160192825DNAPopulus deltoidesCDS(60)..(536) 192ggtactctct gtgttttttt
attgttagtg ttttctagtt ctcttgatca atcgccgcc 59atg tct gac gag gag cat
cac ttc gag tca aag gcc gat gct gga gct 107Met Ser Asp Glu Glu His
His Phe Glu Ser Lys Ala Asp Ala Gly Ala1 5
10 15tca aaa act ttt cct cag caa gct ggt acc att cgc
aag aat ggt tac 155Ser Lys Thr Phe Pro Gln Gln Ala Gly Thr Ile Arg
Lys Asn Gly Tyr 20 25 30ata
gtc atc aag aat cgc cct tgc aag gtc gtg gag gtt tca acc tct 203Ile
Val Ile Lys Asn Arg Pro Cys Lys Val Val Glu Val Ser Thr Ser 35
40 45aaa act ggc aag cat ggc cat gct aaa
tgt cac ttt gtt gcg att gat 251Lys Thr Gly Lys His Gly His Ala Lys
Cys His Phe Val Ala Ile Asp 50 55
60att ttc aat gga aaa aag ctt gaa gat att gtt cct tct tcc cac aac
299Ile Phe Asn Gly Lys Lys Leu Glu Asp Ile Val Pro Ser Ser His Asn65
70 75 80tgt gat gtt ccc cac
gtc acc cgt act gac tac cag ctg att gat atc 347Cys Asp Val Pro His
Val Thr Arg Thr Asp Tyr Gln Leu Ile Asp Ile 85
90 95tct gag gat gga ttt gtg agc tta ctg act gag
aat ggc aat act aag 395Ser Glu Asp Gly Phe Val Ser Leu Leu Thr Glu
Asn Gly Asn Thr Lys 100 105
110gat gac ctg aaa ctc cca act gat gac agt ctc ctc act cag att aag
443Asp Asp Leu Lys Leu Pro Thr Asp Asp Ser Leu Leu Thr Gln Ile Lys
115 120 125gat gga ttt ggt gag ggg aaa
gat ctt gtt gtg acg gtg atg tct tcc 491Asp Gly Phe Gly Glu Gly Lys
Asp Leu Val Val Thr Val Met Ser Ser 130 135
140atg gga gag gag cag atc tgc gcc ctc aag gac att ggt ccc aag
536Met Gly Glu Glu Gln Ile Cys Ala Leu Lys Asp Ile Gly Pro Lys145
150 155taatctgcgc ctggctgcaa gcagcgcatg
cccgaggttg actgctgata gttgtagttt 596tagcatgggg ttttatgtgg gtttgagaac
tgggcccatt taagtaacgt tggcggcctt 656atatgaagac ttgccattat ttaagtggtt
ttttatttgt aatgaaggat attcttgtgg 716ttcttgttgt ggagaaagtt tgttctgcca
ttaagactgt atccaccttc ttgccccctt 776gggtggtttc aaccaatttc cagtttttat
taaaaaaaaa aaaaaaaaa 825193159PRTPopulus deltoides 193Met
Ser Asp Glu Glu His His Phe Glu Ser Lys Ala Asp Ala Gly Ala1
5 10 15Ser Lys Thr Phe Pro Gln Gln
Ala Gly Thr Ile Arg Lys Asn Gly Tyr 20 25
30Ile Val Ile Lys Asn Arg Pro Cys Lys Val Val Glu Val Ser
Thr Ser 35 40 45Lys Thr Gly Lys
His Gly His Ala Lys Cys His Phe Val Ala Ile Asp 50 55
60Ile Phe Asn Gly Lys Lys Leu Glu Asp Ile Val Pro Ser
Ser His Asn65 70 75
80Cys Asp Val Pro His Val Thr Arg Thr Asp Tyr Gln Leu Ile Asp Ile
85 90 95Ser Glu Asp Gly Phe Val
Ser Leu Leu Thr Glu Asn Gly Asn Thr Lys 100
105 110Asp Asp Leu Lys Leu Pro Thr Asp Asp Ser Leu Leu
Thr Gln Ile Lys 115 120 125Asp Gly
Phe Gly Glu Gly Lys Asp Leu Val Val Thr Val Met Ser Ser 130
135 140Met Gly Glu Glu Gln Ile Cys Ala Leu Lys Asp
Ile Gly Pro Lys145 150
155194846DNAPopulus deltoidesCDS(101)..(580)modified_base(729)a, c, g, t,
unknown or other 194gtgttggtgt cctcttgtgt cttcagagtc atcgtcctct
ttcacttgtt cttctttttt 60ctgcttgaga gagaatcgat ccaagcctaa ctaaaccatc
atg tcg gac gag gag 115
Met Ser Asp Glu Glu 1
5cac cac ttt gaa tcc aag gct gat gca ggc gcc tcc aag act tac cct
163His His Phe Glu Ser Lys Ala Asp Ala Gly Ala Ser Lys Thr Tyr Pro
10 15 20cag caa gct ggt acc
atc cgc aag aat ggt tac ata gtc atc aag aac 211Gln Gln Ala Gly Thr
Ile Arg Lys Asn Gly Tyr Ile Val Ile Lys Asn 25
30 35cgc cct tgc aag gtt gtt gag gtt tcc acc tca aag
aca ggc aag cac 259Arg Pro Cys Lys Val Val Glu Val Ser Thr Ser Lys
Thr Gly Lys His 40 45 50gga cat
gct aag tgc cac ttt gtg gga att gat ata ttt aat ggg aaa 307Gly His
Ala Lys Cys His Phe Val Gly Ile Asp Ile Phe Asn Gly Lys 55
60 65aag ctt gaa gat att gtt ccc tca tct cac aac
tgt gat gtt ccc cat 355Lys Leu Glu Asp Ile Val Pro Ser Ser His Asn
Cys Asp Val Pro His70 75 80
85gtt aat cgt gtt gat tat caa ttg att gat atc tct gaa gat ggt ttt
403Val Asn Arg Val Asp Tyr Gln Leu Ile Asp Ile Ser Glu Asp Gly Phe
90 95 100gtg agt ctt ttg act
gaa aat gga aat acc aag gat gat ctg agg ctt 451Val Ser Leu Leu Thr
Glu Asn Gly Asn Thr Lys Asp Asp Leu Arg Leu 105
110 115ccc act gat gac agt cta ctt acc cag att aaa gat
gga ttt ggc gaa 499Pro Thr Asp Asp Ser Leu Leu Thr Gln Ile Lys Asp
Gly Phe Gly Glu 120 125 130ggg aag
gac ctt gtg gtg tct gtc atg tct gcc atg gga gag gag cag 547Gly Lys
Asp Leu Val Val Ser Val Met Ser Ala Met Gly Glu Glu Gln 135
140 145atc tgt gcc ctt aag gac att ggc cca aaa aat
tgaatatgat gctatagctg 600Ile Cys Ala Leu Lys Asp Ile Gly Pro Lys
Asn150 155 160tagctgtagc tgatggcaaa
acgatttctt ttatgttgct aacaaggatt tttatccact 660atgcatgtgt acaacaaact
tataagacgg tattttagct atggaattga aactcgcaag 720gcatgtaang gaggaggtgg
gaaggacccc ggaaatgctg attccctgat gaagggaact 780tgtgttatca gcaggtggga
atttatatat atttctattt cgctaaaaaa aaaaaaaaaa 840aaaaaa
846195160PRTPopulus
deltoides 195Met Ser Asp Glu Glu His His Phe Glu Ser Lys Ala Asp Ala Gly
Ala1 5 10 15Ser Lys Thr
Tyr Pro Gln Gln Ala Gly Thr Ile Arg Lys Asn Gly Tyr 20
25 30Ile Val Ile Lys Asn Arg Pro Cys Lys Val
Val Glu Val Ser Thr Ser 35 40
45Lys Thr Gly Lys His Gly His Ala Lys Cys His Phe Val Gly Ile Asp 50
55 60Ile Phe Asn Gly Lys Lys Leu Glu Asp
Ile Val Pro Ser Ser His Asn65 70 75
80Cys Asp Val Pro His Val Asn Arg Val Asp Tyr Gln Leu Ile
Asp Ile 85 90 95Ser Glu
Asp Gly Phe Val Ser Leu Leu Thr Glu Asn Gly Asn Thr Lys 100
105 110Asp Asp Leu Arg Leu Pro Thr Asp Asp
Ser Leu Leu Thr Gln Ile Lys 115 120
125Asp Gly Phe Gly Glu Gly Lys Asp Leu Val Val Ser Val Met Ser Ala
130 135 140Met Gly Glu Glu Gln Ile Cys
Ala Leu Lys Asp Ile Gly Pro Lys Asn145 150
155 160196824DNAPopulus deltoidesCDS(48)..(524)
196ggtattttgt ttgttttgtg attgcgactc tcttatcaat cgcggcc atg tct gac
56 Met Ser Asp
1gag gag cag cac ttc gag tca
aaa gct gat gcg gga gct tcg aaa act 104Glu Glu Gln His Phe Glu Ser
Lys Ala Asp Ala Gly Ala Ser Lys Thr 5 10
15tac cct caa caa gct ggt acc att cgc aag agc ggt tac att gtc atc
152Tyr Pro Gln Gln Ala Gly Thr Ile Arg Lys Ser Gly Tyr Ile Val Ile20
25 30 35aag aat cgc cct
tgc aag gtt gtg gag gtt tct acc tct aaa act ggc 200Lys Asn Arg Pro
Cys Lys Val Val Glu Val Ser Thr Ser Lys Thr Gly 40
45 50aag cac ggc cat gcc aaa tgt cac ttt gtt
gca att gat atc ttc aat 248Lys His Gly His Ala Lys Cys His Phe Val
Ala Ile Asp Ile Phe Asn 55 60
65gga aaa aag ctt gaa gat att gtt cct tct tcc cac aac tgt gat gtt
296Gly Lys Lys Leu Glu Asp Ile Val Pro Ser Ser His Asn Cys Asp Val
70 75 80ccc cat gtc acc cgt act gac tat
cag ctg att gat atc tca gag gat 344Pro His Val Thr Arg Thr Asp Tyr
Gln Leu Ile Asp Ile Ser Glu Asp 85 90
95gga ttt gtg agc ttg ctg act gag aat ggc aat acc aag gat gac ctg
392Gly Phe Val Ser Leu Leu Thr Glu Asn Gly Asn Thr Lys Asp Asp Leu100
105 110 115agg ctc cca act
gat gag agt ctc ctc tct cag atc aag gat gga ttt 440Arg Leu Pro Thr
Asp Glu Ser Leu Leu Ser Gln Ile Lys Asp Gly Phe 120
125 130ggc gag ggt aaa gat ctt gtt gtg act gtg
atg tcc tcc atg gga gag 488Gly Glu Gly Lys Asp Leu Val Val Thr Val
Met Ser Ser Met Gly Glu 135 140
145gag cag atc tgc gcc ctc aag gac gtt ggc ccg aag taaattgtac
534Glu Gln Ile Cys Ala Leu Lys Asp Val Gly Pro Lys 150
155cctgcttcat gcaggcatgc ccaaggttga cggctgatag ttgtagctta gcaagtggtt
594ctatgtgggt ttgagaattg gtcccgtttc aggaacattc cgtactagat gcggccttcc
654cttatatgaa gacttgccat tatttgagtg gcgttttatc cgtgatgata tttttgtggt
714tcttggtgtg gggaagttta ttctgccagt aagactgcat acacttcttg gcacctgtgg
774tggtttcaaa caattacagt ttttatttcc ttaaaaaaaa aaaaaaaaaa
824197159PRTPopulus deltoides 197Met Ser Asp Glu Glu Gln His Phe Glu Ser
Lys Ala Asp Ala Gly Ala1 5 10
15Ser Lys Thr Tyr Pro Gln Gln Ala Gly Thr Ile Arg Lys Ser Gly Tyr
20 25 30Ile Val Ile Lys Asn Arg
Pro Cys Lys Val Val Glu Val Ser Thr Ser 35 40
45Lys Thr Gly Lys His Gly His Ala Lys Cys His Phe Val Ala
Ile Asp 50 55 60Ile Phe Asn Gly Lys
Lys Leu Glu Asp Ile Val Pro Ser Ser His Asn65 70
75 80Cys Asp Val Pro His Val Thr Arg Thr Asp
Tyr Gln Leu Ile Asp Ile 85 90
95Ser Glu Asp Gly Phe Val Ser Leu Leu Thr Glu Asn Gly Asn Thr Lys
100 105 110Asp Asp Leu Arg Leu
Pro Thr Asp Glu Ser Leu Leu Ser Gln Ile Lys 115
120 125Asp Gly Phe Gly Glu Gly Lys Asp Leu Val Val Thr
Val Met Ser Ser 130 135 140Met Gly Glu
Glu Gln Ile Cys Ala Leu Lys Asp Val Gly Pro Lys145 150
155198898DNAPopulus deltoidesCDS(85)..(564) 198gctgtcctct
tctgtctcga gaagtcatca tcctcttcca ttattttttc cttttctgct 60cgagagagaa
ccgaaccaac caac atg tcg gac gag gag cac cac ttt gaa 111
Met Ser Asp Glu Glu His His Phe Glu
1 5tcg aag gct gat gca gga gcc tcc aag act tac cct cag
caa gct ggt 159Ser Lys Ala Asp Ala Gly Ala Ser Lys Thr Tyr Pro Gln
Gln Ala Gly10 15 20
25act atc cgc aag aat ggc tat ata gtc atc aag aac cgt cct tgc aag
207Thr Ile Arg Lys Asn Gly Tyr Ile Val Ile Lys Asn Arg Pro Cys Lys
30 35 40gtt gtt gag gtt tcc acc
tca aag aca ggc aag cat gga cat gct aag 255Val Val Glu Val Ser Thr
Ser Lys Thr Gly Lys His Gly His Ala Lys 45 50
55tgc cac ttt gtg ggg att gat ata ttt aat ggg aaa aag
ctt gaa gat 303Cys His Phe Val Gly Ile Asp Ile Phe Asn Gly Lys Lys
Leu Glu Asp 60 65 70att gtt cct
tca tct cac aac tgt gat gtt ccc cat gtt aat cgt act 351Ile Val Pro
Ser Ser His Asn Cys Asp Val Pro His Val Asn Arg Thr 75
80 85gat tat cag ctg att gat atc tct gaa gat ggc ttt
gtg agt ctt ttg 399Asp Tyr Gln Leu Ile Asp Ile Ser Glu Asp Gly Phe
Val Ser Leu Leu90 95 100
105act gaa aat gga aat acc aag gat gat ctg agg ctt ccc act gat gat
447Thr Glu Asn Gly Asn Thr Lys Asp Asp Leu Arg Leu Pro Thr Asp Asp
110 115 120agt ctg ctt act ccg
att aaa gat ggg ttt ggt gaa gga aag gac ctt 495Ser Leu Leu Thr Pro
Ile Lys Asp Gly Phe Gly Glu Gly Lys Asp Leu 125
130 135gtg gtg act gtc atg tct gcc atg gga gag gag cag
atc tgt gcc ctt 543Val Val Thr Val Met Ser Ala Met Gly Glu Glu Gln
Ile Cys Ala Leu 140 145 150aag gac
att ggt cca aaa aat tgaatatgat gctgtagcag cagctatagc 594Lys Asp
Ile Gly Pro Lys Asn 155 160tgatggcgaa gcgagtttta
tgttgctaac aaggattttt atgcactatg cgttgtgtac 654aacaaactta taagacagta
ttttagatat gtcattaaaa ctcgcaaggc atgtcatgga 714ggtggaagag gacacaggag
atgcttattc cctgatgaag ggaactaggt tttcagcagg 774atggaattta tatatatata
tctacttcac atttgtctgt gctacttaaa gataaataaa 834ggtttgtgct caaccctttt
cctttcaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 894aaaa
898199160PRTPopulus
deltoides 199Met Ser Asp Glu Glu His His Phe Glu Ser Lys Ala Asp Ala Gly
Ala1 5 10 15Ser Lys Thr
Tyr Pro Gln Gln Ala Gly Thr Ile Arg Lys Asn Gly Tyr 20
25 30Ile Val Ile Lys Asn Arg Pro Cys Lys Val
Val Glu Val Ser Thr Ser 35 40
45Lys Thr Gly Lys His Gly His Ala Lys Cys His Phe Val Gly Ile Asp 50
55 60Ile Phe Asn Gly Lys Lys Leu Glu Asp
Ile Val Pro Ser Ser His Asn65 70 75
80Cys Asp Val Pro His Val Asn Arg Thr Asp Tyr Gln Leu Ile
Asp Ile 85 90 95Ser Glu
Asp Gly Phe Val Ser Leu Leu Thr Glu Asn Gly Asn Thr Lys 100
105 110Asp Asp Leu Arg Leu Pro Thr Asp Asp
Ser Leu Leu Thr Pro Ile Lys 115 120
125Asp Gly Phe Gly Glu Gly Lys Asp Leu Val Val Thr Val Met Ser Ala
130 135 140Met Gly Glu Glu Gln Ile Cys
Ala Leu Lys Asp Ile Gly Pro Lys Asn145 150
155 160200379PRTNicotiana tabacum 200Met Gly Glu Ala Leu
Asn Val Met Glu Ser Val Arg Ser Ile Val Phe1 5
10 15Lys Glu Ser Glu Asn Leu Glu Gly Ser Ala Thr
Lys Ile Glu Gly Tyr 20 25
30Asp Phe Asn Lys Gly Val Asn Tyr Ala Glu Leu Phe Lys Ser Met Ala
35 40 45Ser Thr Gly Phe Gln Ala Ala Asn
Leu Gly Asp Ala Ile Gln Ile Val 50 55
60Asn Gln Met Leu Asp Trp Arg Leu Ser His Glu Gln Pro Met Glu Asp65
70 75 80Cys Ser Glu Glu Glu
Arg Asp Val Ala Tyr Arg Glu Ser Val Thr Cys 85
90 95Lys Ile Phe Leu Gly Phe Thr Ser Asn Leu Val
Ser Ser Gly Val Arg 100 105
110Asp Thr Ile Arg Tyr Leu Val Gln His Arg Met Val Asp Val Val Val
115 120 125Thr Thr Ala Gly Gly Ile Glu
Glu Asp Leu Ile Lys Cys Leu Ala Pro 130 135
140Thr Tyr Lys Gly Asp Phe Ser Leu Pro Gly Ala Val Leu Arg Ser
Lys145 150 155 160Gly Leu
Asn Arg Ile Gly Asn Leu Leu Val Pro Asn Asp Asn Tyr Cys
165 170 175Lys Phe Glu Asn Trp Ile Ile
Pro Ile Phe Asp Gln Met Tyr Glu Glu 180 185
190Gln Ile Lys Glu Lys Val Leu Trp Thr Pro Ser Lys Val Ile
Ala Arg 195 200 205Leu Ala Lys Glu
Ile Asn Asp Glu Thr Ser Tyr Leu Tyr Trp Ala Tyr 210
215 220Lys Asn Arg Ile Pro Val Phe Cys Pro Gly Leu Thr
Asp Gly Ser Leu225 230 235
240Gly Asp Met Leu Tyr Phe His Ser Phe Lys Lys Gly Asp Pro Asp Asn
245 250 255Pro Asp Leu Asn Pro
Gly Leu Ile Ile Asp Ile Val Gly Asp Ile Arg 260
265 270Ala Met Asn Ser Glu Ala Val His Ala Gly Ser Arg
Lys Thr Gly Met 275 280 285Ile Ile
Leu Gly Gly Gly Leu Pro Lys His His Val Cys Asn Ala Asn 290
295 300Met Met Arg Asn Gly Ala Asp Phe Ala Val Tyr
Ile Asn Thr Ala Gln305 310 315
320Glu Phe Asp Gly Ser Asp Ser Gly Ala Arg Pro Asp Glu Ala Val Ser
325 330 335Trp Gly Lys Ile
Arg Gly Gly Ala Lys Thr Val Lys Val His Cys Asp 340
345 350Ala Thr Ile Ala Phe Pro Ile Leu Val Ala Glu
Thr Phe Ala Ala Lys 355 360 365Arg
Lys Glu Leu Ser His Ile Arg Cys Gln Val 370
375201368PRTArabidopsis sp. 201Met Glu Asp Asp Arg Val Phe Ser Ser Val
His Ser Thr Val Phe Lys1 5 10
15Glu Ser Glu Ser Leu Glu Gly Lys Cys Asp Lys Ile Gln Gly Tyr Asp
20 25 30Phe Asn Gln Gly Val Asp
Tyr Pro Lys Leu Met Arg Ser Met Leu Thr 35 40
45Thr Gly Phe Gln Ala Ser Asn Leu Gly Glu Ala Ile Asp Val
Val Asn 50 55 60Gln Met Leu Asp Trp
Arg Leu Ala Asp Glu Thr Thr Val Ala Glu Asp65 70
75 80Cys Ser Glu Glu Glu Lys Asn Pro Ser Phe
Arg Glu Ser Val Lys Cys 85 90
95Lys Ile Phe Leu Gly Phe Thr Ser Asn Leu Val Ser Ser Gly Val Arg
100 105 110Asp Thr Ile Arg Tyr
Leu Val Gln His His Met Val Asp Val Ile Val 115
120 125Thr Thr Thr Gly Gly Val Glu Glu Asp Leu Ile Lys
Cys Leu Ala Pro 130 135 140Thr Phe Lys
Gly Asp Phe Ser Leu Pro Gly Ala Tyr Leu Arg Ser Lys145
150 155 160Gly Leu Asn Arg Ile Gly Asn
Leu Leu Val Pro Asn Asp Asn Tyr Cys 165
170 175Lys Phe Glu Asp Trp Ile Ile Pro Ile Phe Asp Glu
Met Leu Lys Glu 180 185 190Gln
Lys Glu Glu Asn Val Leu Trp Thr Pro Ser Lys Leu Leu Ala Arg 195
200 205Leu Gly Lys Glu Ile Asn Asn Glu Ser
Ser Tyr Leu Tyr Trp Ala Tyr 210 215
220Lys Met Asn Ile Pro Val Phe Cys Pro Gly Leu Thr Asp Gly Ser Leu225
230 235 240Gly Asp Met Leu
Tyr Phe His Ser Phe Arg Thr Ser Gly Leu Ile Ile 245
250 255Asp Val Val Gln Asp Ile Arg Ala Met Asn
Gly Glu Ala Val His Ala 260 265
270Asn Pro Lys Lys Thr Gly Met Ile Ile Leu Gly Gly Gly Leu Pro Lys
275 280 285His His Ile Cys Asn Ala Asn
Met Met Arg Asn Gly Ala Asp Tyr Ala 290 295
300Val Phe Ile Asn Pro Gly Gln Glu Phe Asp Gly Ser Asp Ser Gly
Ala305 310 315 320Arg Pro
Asp Glu Ala Val Ser Trp Gly Lys Ile Arg Gly Ser Ala Lys
325 330 335Thr Val Lys Val Tyr Cys Asp
Ala Thr Ile Ala Phe Pro Leu Leu Val 340 345
350Ala Glu Thr Phe Ala Thr Lys Arg Asp Gln Thr Cys Glu Ser
Lys Thr 355 360 365202379PRTZea
mays 202Met Gly Ala Glu Leu Gly Glu Ala Gly Val Pro Ala Arg Asp Met Val1
5 10 15Asp Ala Leu Glu
Gly Val Arg Ser Ile Leu Leu Lys Pro Ser Glu Arg 20
25 30Leu Asp Asp Lys Arg Phe Thr Arg Ile Ala Gly
Ala Asp Phe Asp Asp 35 40 45Ala
Gly Leu Gly Leu Ala Gly Leu Leu Gly Ser Leu Ala Ser Thr Gly 50
55 60Phe Gln Ala Ser Asn Leu Gly Asp Ala Ile
Asp Val Val Asn Gln Met65 70 75
80Leu Asp Trp Arg Leu Ser His Glu Lys Pro Ser Asp Asp Cys Ala
Glu 85 90 95Ala Glu Leu
Asp Pro Ala Tyr Arg Glu Ser Val Lys Cys Lys Ile Phe 100
105 110Leu Gly Phe Thr Ser Asn Leu Val Ser Ser
Gly Val Arg Glu Ile Ile 115 120
125Arg Phe Leu Val Gln His Arg Met Val Asp Val Ile Val Thr Thr Ala 130
135 140Gly Gly Ile Glu Glu Asp Leu Ile
Lys Cys Leu Ala Pro Thr Tyr Arg145 150
155 160Gly Asp Phe Thr Leu Pro Gly Ala Leu Leu Arg Ser
Arg Gly Leu Asn 165 170
175Arg Ile Gly Asn Leu Leu Val Pro Asn Asp Asn Tyr Cys Lys Phe Glu
180 185 190His Trp Ile Met Pro Ile
Leu Asp Lys Met Leu Leu Glu Gln Ser Thr 195 200
205Gln Asn Val Trp Thr Pro Ser Lys Val Ile Ala Arg Leu Gly
Lys Glu 210 215 220Ile Asn Asp Glu Ser
Ser Tyr Leu Tyr Trp Ala Tyr Lys Asn Asn Ile225 230
235 240Pro Val Phe Cys Pro Ala Leu Thr Asp Gly
Ser Ile Gly Asp Met Leu 245 250
255Phe Cys His Ser Val His Ser Pro Gly Leu Val Val Asp Ile Val Gln
260 265 270Asp Val Arg Leu Met
Asn Ala Glu Thr Ile His Ala Ser Pro Arg Lys 275
280 285Thr Gly Ile Ile Leu Leu Gly Gly Gly Leu Pro Lys
His His Ile Cys 290 295 300Asn Ala Asn
Met Leu Arg Asp Gly Ala Asp Tyr Ala Val Tyr Val Asn305
310 315 320Thr Ala Gln Glu Phe Asp Gly
Ser Asp Ser Gly Ala Arg Pro Asp Glu 325
330 335Ala Val Ser Trp Gly Lys Ile Lys Ser Ser Ala Arg
Thr Val Lys Val 340 345 350His
Cys Asp Ala Thr Ile Ala Phe Pro Leu Leu Val Ala Ala Thr Phe 355
360 365Ala Arg Lys Val His Gly Thr Lys Ser
Thr Asn 370 375203820DNAPopulus deltoides
203gaattcggca cgagggtatt ttgtttgttt tgtgattgcg actctcttat caatcgcggc
60catgtctgac gaggagcagc acttcgagtc aaaagctgat gcgggagctt cgaaaactta
120ccctcaacaa gctggtacca ttcgcaagag cggttacatt gtcatcaaga atcgcccttg
180caaggttgtg gaggtttcta cctctaaaac tggcaagcac ggccatgcca aatgtcactt
240tgttgcaatt gatatcttca atggaaaaaa gcttgaagat attgttcctt cttcccacaa
300ctgtgatgtt ccccatgtca cccgtactga ctatcagctg attgatatct cagaggatgg
360atttgtgagc ttgctgactg agaatggcaa taccaaggat gacctgaggc tcccaactga
420tgagagtctc ctctctcaga tcaaggatgg atttggcgag ggtaaagatc ttgttgtgac
480tgtgatgtcc tccatgggag aggagcagat ctgcgccctc aaggacgttg gcccgaagta
540aattgtaccc tgcttcatgc aggcatgccc aaggttgacg gctgatagtt gtagcttagc
600aagtggttct atgtgggttt gagaattggt cccgtttcag gaacattccg tactagatgc
660ggccttccct tatatgaaga cttgccatta tttgagtggc gttttatccg tgatgatatt
720tttgtggttc ttggtgtggg gaagtttatt ctgccagtaa gactgcatac acttcttggc
780acctgtggtg gtttcaaaca attacagttt ttatttcctt
820204159PRTPopulus deltoides 204Met Ser Asp Glu Glu Gln His Phe Glu Ser
Lys Ala Asp Ala Gly Ala1 5 10
15Ser Lys Thr Tyr Pro Gln Gln Ala Gly Thr Ile Arg Lys Ser Gly Tyr
20 25 30Ile Val Ile Lys Asn Arg
Pro Cys Lys Val Val Glu Val Ser Thr Ser 35 40
45Lys Thr Gly Lys His Gly His Ala Lys Cys His Phe Val Ala
Ile Asp 50 55 60Ile Phe Asn Gly Lys
Lys Leu Glu Asp Ile Val Pro Ser Ser His Asn65 70
75 80Cys Asp Val Pro His Val Thr Arg Thr Asp
Tyr Gln Leu Ile Asp Ile 85 90
95Ser Glu Asp Gly Phe Val Ser Leu Leu Thr Glu Asn Gly Asn Thr Lys
100 105 110Asp Asp Leu Arg Leu
Pro Thr Asp Glu Ser Leu Leu Ser Gln Ile Lys 115
120 125Asp Gly Phe Gly Glu Gly Lys Asp Leu Val Val Thr
Val Met Ser Ser 130 135 140Met Gly Glu
Glu Gln Ile Cys Ala Leu Lys Asp Val Gly Pro Lys145 150
15520528DNAArtificial SequenceDescription of Artificial
Sequence Synthetic primer 205gaagctcgag gactctctta tcaatcgc
2820631DNAArtificial SequenceDescription of
Artificial Sequence Synthetic primer 206ggggagctca gggtacaatt
tacttcgggc c 31207906DNAPopulus
deltoides 207cactgaatca aaggccatgg agtcaaagat tcaaatagag gacctaacag
aactcgccgt 60aaagactggc gaacagttca tacagagtct cttacgactc aatgacaaga
agaaaatctt 120cgtcaacatg gtggagcacg acacgcttgt ctacctccaa aaatatcaaa
gatacagtct 180cagaagacca aagggaattg agacttttca acaaagggta atatccggaa
acctcctcgg 240attccattgc ccagctatct gtcactttat tgtgaagata gtggaaaagg
aaggtggctc 300ctacaaatgc catcattgcg ataaaggaaa ggccatcgtt gaagatgcct
ctgccgacag 360tggtcccaaa gatggacccc cacccacgag gagcatcgtg gaaaaagaag
acgttccaac 420cacgtcttca aagcaagtgg attgatgtga taacatggtg gagcacgaca
cgcttgtcta 480cctccaaaaa tatcaaagat acagtctcag aagaccaaag ggaattgaga
cttttcaaca 540aagggtaata tccggaaacc tcctcggatt ccattgccca gctatctgtc
actttattgt 600gaagatagtg gaaaaggaag gtggctccta caaatgccat cattgcgata
aaggaaaggc 660catcgttgaa gatgcctctg ccgacagtgg tcccaaagat ggacccccac
ccacgaggag 720catcgtggaa aaagaagacg ttccaaccac gtcttcaaag caagtggatt
gatgtgatat 780ctccactgac gtaagggatg acgcacaatc ccactatcct tcgcaagacc
cttcctctat 840ataaggaagt tcatttcatt tggagaggac acgctgaaat caccagtctc
tctctaagct 900tggatc
906208512DNAPopulus deltoides 208gactctctta tcaatcgcgg
ccatgtctga cgaggagcag cacttcgagt caaaagctga 60tgcgggagct tcgaaaactt
accctcaaca agctggtacc attcgcaaga gcggttacat 120tgtcatcaag aatcgccctt
gcaaggttgt ggaggtttct acctctaaaa ctggcaagca 180cggccatgcc aaatgtcact
ttgttgcaat tgatatcttc aatggaaaaa agcttgaaga 240tattgttcct tcttcccaca
actgtgatgt tccccatgtc acccgtactg actatcagct 300gattgatatc tcagaggatg
gatttgtgag cttgctgact gagaatggca ataccaagga 360tgacctgagg ctcccaactg
atgagagtct cctctctcag atcaaggatg gatttggcga 420gggtaaagat cttgttgtga
ctgtgatgtc ctccatggga gaggagcaga tctgcgccct 480caaggacgtt ggcccgaagt
aaattgtacc ct 512209662DNAPopulus
deltoides 209gaattgatcc tctagagctt tcgttcgtat catcggtttc gacaacgttc
gtcaagttca 60atgcatcagt ttcattgcgc acacaccaga atcctactga gttcgagtat
tatggcattg 120ggaaaactgt ttttcttgta ccatttgttg tgcttgtaat ttactgtgtt
ttttattcgg 180ttttcgctat cgaactgtga aatggaaatg gatggagaag agttaatgaa
tgatatggtc 240cttttgttca ttctcaaatt aatattattt gttttttctc ttatttgttg
tgtgttgaat 300ttgaaattat aagagatatg caaacatttt gttttgagta aaaatgtgtc
aaatcgtggc 360ctctaatgac cgaagttaat atgaggagta aaacacttgt agttgtacca
ttatgcttat 420tcactaggca acaaatatat tttcagacct agaaaagctg caaatgttac
tgaatacaag 480tatgtcctct tgtgttttag acatttatga actttccttt atgtaatttt
ccagaatcct 540tgtcagattc taatcattgc tttataatta tagttatact catggatttg
tagttgagta 600tgaaaatatt ttttaatgca ttttatgact tgccaattga ttgacaacat
gcatcaatcg 660at
662210466DNALactuca sativaCDS(3)..(329) 210cg cat ttg cgt cgt
cgt ggg ata ttt cga ttt ggg gaa tcg aca ttt 47 His Leu Arg Arg
Arg Gly Ile Phe Arg Phe Gly Glu Ser Thr Phe 1 5
10 15tta atc tgg aaa aag ctt gaa gat att gtt cca
tca tcc cat aat tgt 95Leu Ile Trp Lys Lys Leu Glu Asp Ile Val Pro
Ser Ser His Asn Cys 20 25
30gat gtc ccc cat gtc aat cgc acc gat tac cag cta att gat att tct
143Asp Val Pro His Val Asn Arg Thr Asp Tyr Gln Leu Ile Asp Ile Ser
35 40 45gaa gat gga ttt gtg agt ttg ctg
act gaa aat ggt aac acc aaa gat 191Glu Asp Gly Phe Val Ser Leu Leu
Thr Glu Asn Gly Asn Thr Lys Asp 50 55
60gac ctc aaa ctc ccc acc gat gat gct ctc ctc act cag ata aag gat
239Asp Leu Lys Leu Pro Thr Asp Asp Ala Leu Leu Thr Gln Ile Lys Asp65
70 75 80gga ttt gga gag ggg
aag ggc ctg gtt gtg act gtt atg tca gca atg 287Gly Phe Gly Glu Gly
Lys Gly Leu Val Val Thr Val Met Ser Ala Met 85
90 95gga gag gaa cag ata tgt gct ctc aag gac att
ggc ccc aaa 329Gly Glu Glu Gln Ile Cys Ala Leu Lys Asp Ile
Gly Pro Lys 100 105
110taacttttca aaaatccaca aaaacacaca ctataatatt attataacat ttggattaac
389agaaaacaag ttgtgttttt atttgtttta tcatgtacag atgttatgga ttaaaaaaaa
449aaaaaaaaaa aaaaaaa
466211109PRTLactuca sativa 211His Leu Arg Arg Arg Gly Ile Phe Arg Phe Gly
Glu Ser Thr Phe Leu1 5 10
15Ile Trp Lys Lys Leu Glu Asp Ile Val Pro Ser Ser His Asn Cys Asp
20 25 30Val Pro His Val Asn Arg Thr
Asp Tyr Gln Leu Ile Asp Ile Ser Glu 35 40
45Asp Gly Phe Val Ser Leu Leu Thr Glu Asn Gly Asn Thr Lys Asp
Asp 50 55 60Leu Lys Leu Pro Thr Asp
Asp Ala Leu Leu Thr Gln Ile Lys Asp Gly65 70
75 80Phe Gly Glu Gly Lys Gly Leu Val Val Thr Val
Met Ser Ala Met Gly 85 90
95Glu Glu Gln Ile Cys Ala Leu Lys Asp Ile Gly Pro Lys 100
105
User Contributions:
comments("1"); ?> comment_form("1"); ?>Inventors list |
Agents list |
Assignees list |
List by place |
Classification tree browser |
Top 100 Inventors |
Top 100 Agents |
Top 100 Assignees |
Usenet FAQ Index |
Documents |
Other FAQs |
User Contributions:
Comment about this patent or add new information about this topic:
| People who visited this patent also read: | |
| Patent application number | Title |
|---|---|
| 20110092848 | MRI Biopsy Targeting Cube with Eccentric Lock |
| 20110092847 | MRI Biopsy Targeting Cube with Snap Corners |
| 20110092846 | Cerebral Compliance Monitoring |
| 20110092844 | MICRO-INSULT TEST AND USE THEREFOR |
| 20110092843 | AERODYNAMIC TISSUE DRIVER |
 |  |
 |  |
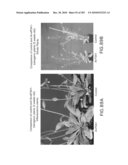 |  |
 |  |
 |  |
 |  |
 | 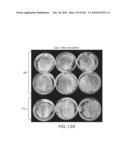 |
 |  |
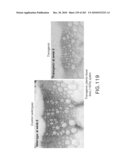 |  |
 | 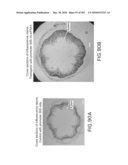 |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 | 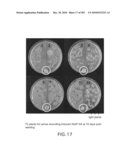 |
 | 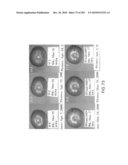 |
 |  |
 | 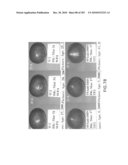 |
 |  |
 |  |
 |  |
 |  |
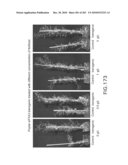 | 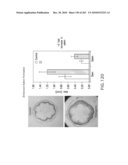 |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 | 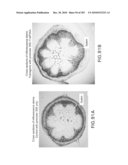 |
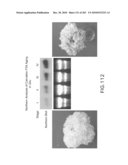 |  |
 |  |
 |  |
 |
| New patent applications in this class: | |
| Date | Title |
|---|---|
| 2016-06-23 | Plants having one or more enhanced yield-related traits and a method for making the same |
| 2016-06-09 | Transgenic maize |
| 2016-05-19 | Methods and compositions for improvement in seed yield |
| 2016-05-12 | Means and methods for yield performance in plants |
| 2016-04-21 | Plants having one or more enhanced yield-related traits and a method for making the same |
| New patent applications from these inventors: | |
| Date | Title |
|---|---|
| 2015-05-07 | Combination treatment of multiple myeloma |
| 2014-12-18 | Use of apoptosis-specific elf-5a sirna to down regulate expression of proinflammatory cytokines to treat sepsis |
| 2014-10-23 | Treatment of b cell lymphomas |
| 2014-10-02 | Use of sirna to achieve down regulation of an endogenous gene in combination with the use of a sense construct to achieve expression of a polynucleotide |
| 2014-07-31 | Nucleic acids, polypetides, and methods for modulating apoptosis |
| Top Inventors for class "Multicellular living organisms and unmodified parts thereof and related processes" | |
| Rank | Inventor's name |
|---|---|
| 1 | Gregory J. Holland |
| 2 | William H. Eby |
| 3 | Richard G. Stelpflug |
| 4 | Laron L. Peters |
| 5 | Justin T. Mason |