Patent application title: Directed Neural Differentiation
Inventors:
Sarah Elizabeth Lowell (Edinburgh, GB)
Austin Gerard Smith (Edinburgh, GB)
Barry Heavey (Athlone, IE)
IPC8 Class: AC12N508FI
USPC Class:
435368
Class name: Primate cell, per se human nervous system origin or derivative
Publication date: 2008-10-09
Patent application number: 20080248568
Inventors list |
Agents list |
Assignees list |
List by place |
Classification tree browser |
Top 100 Inventors |
Top 100 Agents |
Top 100 Assignees |
Usenet FAQ Index |
Documents |
Other FAQs |
Patent application title: Directed Neural Differentiation
Inventors:
Sarah Elizabeth Lowell
Austin Gerard Smith
Barry Heavey
Agents:
STERNE, KESSLER, GOLDSTEIN & FOX P.L.L.C.
Assignees:
Origin: WASHINGTON, DC US
IPC8 Class: AC12N508FI
USPC Class:
435368
Abstract:
Differentiation towards a neural fate, and away from a non-neural fate, is
promoted by activation of Notch signalling in ES cells and then
transferring the cells into neural differentiation protocols. Media for
neural differentiation comprises a Notch activator, e.g. a notch ligand
that can be clustered. Genetic manipulation is used as an alternative to
media additives for Notch activation.Claims:
1-6. (canceled)
7. A method of obtaining a culture enriched in neural cells, comprising:(a) providing a culture comprising multipotent or pluripotent cells which have the potential to form progeny committed to either a neural or a non-neural fate; and(b) activating Notch signalling in the cells.
8. A method according to claim 7, wherein the activating comprises expressing in the cells a Notch receptor or an activated form of a Notch receptor.
9. (canceled)
10. A method according to claim 7, wherein the activating comprises adding to culture medium a notch ligand.
11. (canceled)
12. A method according to claim 7, wherein the activating comprises expressing in the cells a downstream effector of Notch signalling.
13. A method according to claim 7, wherein the activating comprises adding to culture medium a composition comprising a downstream effector of Notch signalling and a transduction domain.
14. (canceled)
15. A method according to claim 7, wherein the activating comprises adding to culture medium a composition an activated form of a Notch receptor and a transduction domain.
16-18. (canceled)
16. A method according to claim 7 further comprising purifying the neural cells.
20-21. (canceled)
22. A method according to claim 7 comprising culturing the cells in medium non-permissive for the multipotent or the pluripotent cells.
23-27. (canceled)
28. A method of increasing the density of neural progenitor cells in culture, comprising activating Notch signalling in the cells.
29. A method according to claim 28, wherein the cells are human cells.
30. A composition comprising an activated form of a Notch receptor and a transduction domain.
31. A composition according to claim 30, wherein the activated form of a Notch receptor comprises an intracellular domain of a Notch receptor.
32. A composition comprising a downstream effector of Notch signalling and a transduction domain.
33. A composition according to claim 32, wherein the effector is a Hes transcription factor or a Hey transcription factor.
34-36. (canceled)
37. A fusion protein of an activated form of a Notch receptor and a transduction domain.
38. A fusion protein of a downstream effector of Notch signalling and a transduction domain.
39. A nucleotide sequence encoding the fusion protein of claim 37.
40. A vector comprising the nucleotide sequence of claim 39.
41. (canceled)
42. A medium for culture of neural cells, comprising a composition according to claim 30
43. A medium according to claim 42, wherein the medium is non-permissive for multipotent cells.
44. A medium according to claim 42, wherein the medium is non-permissive for pluripotent cells.
45. A pluripotent cell in which Notch signalling has been activated.
46. An ES cell according to claim 45.
47. (canceled)
48. (canceled)
49. A pluripotent cell engineered to express a peptide comprising a Notch intracellular domain.
50. An ES cell according to claim 49.
51. (canceled)
52. (canceled)
Description:
[0001]The present invention relates to directed neural differentiation. In
particular this invention relates to promoting differentiation of
pluripotent cells towards a neural fate, reducing differentiation towards
a non-neural fate and maintaining cultures of neural cells, and to
methods and compositions therefor.
[0002]An important goal in stem cell research is to reliably convert embryonic stem (ES) cells into the neural lineage. This is important for two reasons: to allow us to better understand the mechanism of neural specification and to provide neural cells and tissue, e.g. for regenerative therapies, with minimal contamination by non-neural cells.
[0003]Recent progress towards this goal came with the identification of culture conditions under which a large proportion of ES cells become neural progenitors (Ying et al., 2003b). The two central features of these culture conditions are that they support survival of both ES and neural cells, and that they lack a source of exogenous BMP, which is a critical inhibitor of neural specification for ES cells (and which can be provided by serum)(Ying et al., 2003a).
[0004]However, even under these optimised culture conditions, between 20 and 40% of cells resist neural specification: around half of these differentiate into non-neural lineages, whilst the others remain as undifferentiated ES cells.
[0005]Notch signalling is known in many different tissues to regulate differentiation decisions by mediating signalling between neighbouring cells (Lai, 2004). Notch receptors and ligands are expressed in ES cells, but their function in this context is hitherto unknown. Notch ligands include members of two closely related families called Delta (Delta 1, 3 and 4) and Jagged (Jagged is also sometimes referred to as Serrate and 2 forms, Jagged1 and Jagged2, have been identified). They are all transmembrane proteins that sit on the cell surface and bind and activate Notch receptors on neighbouring cells.
[0006]It is known that the effect of Notch on lineage committed cells is generally to inhibit differentiation. Thus, ES cells have been generated with a targeted deletion of RBPJk, the co-activator that is required for Notch to activate transcription of its target genes and is thought to mediate the activity of all 4 Notch receptors (Schroeder et al., 2003). These ES cells have been successfully differentiated into the mesodermal lineage--the focus of this study was to uncover a bias in the subsequent differentiation of ES-derived mesoderm towards cardiomyocyte and away from other mesodermal lineages.
[0007]In another study, RBPJk-deficient ES cells were able to generate neurospheres (Hitoshi et al., 2002). Unfortunately, neurosphere cultures are highly selective, making it difficult to draw conclusions about any quantitative effects on the overall rate of neural specification
[0008]Notch has also been studied in tissues; e.g. it is known to try to activate Notch signalling in neural cells by adding a Notch ligand-Fc fusion to cell culture medium and then adding an anti-Fc antibody to cluster the Notch ligands. But these studies are not relevant to the effect of Notch signalling on pluripotent cells.
[0009]It is desired to obtain pure populations of differentiated cells from ES cells, e.g. pure neural cell lines, and protocols exist for generating neural stem cells (NSCs) from ES cells. Even so, several days culture are required. It is desired to reduce this.
[0010]Even when NSCs are obtained, the cultures still contain high numbers of contaminating non-neural cells. In addition, many NSCs differentiate, leading to a less and less homogenous culture. Further culture control, e.g. to inhibit further differentiation until such is desired, would be an advantage.
[0011]Another problem with existing methods for culture of neural cells is that the cells are sensitive to small changes in culture conditions. Cell survival is often adversely affected by variation in, e.g., plating density.
[0012]A further problem with trying to derive neural progeny from ES cells, particularly human ES cells, is that culture at high density tends to suppress neural induction, so any neural induction can only be done at low cell density and as a result low yield, even though lower cell density can lead to reduced cell survival.
[0013]An object of the invention is to improve the bias of differentiating pluripotent cell, especially ES cell, cultures towards a neural fate, and preferably achieve conversion at a rate of as close to 100% as possible.
[0014]A further object of specific embodiments of the invention is to promote differentiation towards a neural stem cell fate without pushing the cells all the way to terminal differentiation, e.g. it is desired to maintain cells at the neural stem cell stage after initial differentiation.
[0015]Another object of specific embodiments of the invention is to allow conversion of ES cells to neural cells, both of mouse and human as well as other origin, at higher densities.
[0016]A still further object of specific embodiments of the invention is to make the neural cell cultures more robust, increasing the survival of cells and reducing the effects of changes in culture conditions.
[0017]In the present invention, neural specification is promoted through activation of Notch signalling. Increased Notch signalling can hence be used to convert pluripotent cell cultures into cultures of neural cells, for example after withdrawal of BMP and/or serum during culture of ES or other pluripotent cells.
[0018]A first aspect of the invention provides use of Notch activation in promoting differentiation of multipotent or pluripotent cells towards a neural fate. The cells are preferably pluripotent stem cells, especially embryonic stem (ES) cells.
[0019]We have found that ES cells can be biased by Notch signalling to differentiate with reduced production of non-neural cells. This yields a purer neural cell culture with reduced contaminating cells.
[0020]To activate Notch signalling, one option is, constitutively or reversibly, to express an activated form of Notch, for example the Notch intracellular domain. In examples described below we have missexpressed Notch, in an activated form, and found this promotes neural specification in culture, even at high cell densities.
[0021]A second aspect of the invention provides use of Notch activation in suppressing differentiation of neural progenitor cells or neural stem cells into terminally differentiated neural cells, i.e. neurons and glia. We have used activation of Notch in neural cell cultures to prevent or at least delay differentiation of neural progenitors into terminally differentiated cells. This maintains the neural cell culture at the progenitor stage, enabling further purification to be undertaken before, e.g., actively promoting differentiation into the desired terminally differentiated cells. Control of the culture and of its purity is as a result improved.
[0022]The invention is of application generally to animal cells, including rodent and primate cells. In particular embodiments the invention is carried out to culture mouse or human neural cells
[0023]The invention also provides methods of culturing cells to obtain cultures containing neural cells or neural progenitor cells, and preferably free or substantially free of non-neural cells.
[0024]One such method of obtaining a culture enriched in neural progenitor cells, comprises: [0025](a) providing a culture comprising multipotent or pluripotent cells which have the potential to form progeny committed to either a neural or a non-neural fate; and [0026](b) activating Notch signalling in the cells.
[0027]The step of activating Notch is found to lead to increased neural specification in the culture, either as the sole means of directing differentiation towards a neural fate or in combination with other means, such as addition of factors known to stimulate neural specification.
[0028]Reference to Notch is reference to a transmembrane protein of that name, currently known to be family of 4 members, which acts as a receptor for various identified ligands and which has been reported to mediate cell fate selection via lateral inhibition. The terms "Notch" and "Notch receptor" may, dependent upon context, have the same meaning. Binding of ligand leads to cleavage of the protein to yield an intracellular, activated from of Notch referred to variously as an activated form of Notch, a Notch intracellular domain and an activated form of the Notch receptor. The intracellular domain complexes with RBPjk (also known as CBP) and the complex binds DNA in the nucleus and activates transcription of Notch target genes. Reference to Notch signalling and activation of Notch and activation of Notch signalling refers to activation of one or more signalling pathways that are mediated in vivo by an activated form of Notch and thus refers to increased transcription of target genes and/or other downstream effectors.
[0029]Notch signalling can thus be activated by expressing, in multipotent or pluripotent cells, an activated form of a Notch receptor, such as an intracellular domain of a Notch receptor. In examples we have performed, this activating has been genetically obtained, using a vector encoding an activated form of the receptor.
[0030]Notch signalling can also be activated by adding a Notch ligand, i.e. a ligand that binds to cell surface Notch receptor, to the culture medium. This approach may be preferred to others as it avoids genetic manipulation of the cells and can easily and quickly be adjusted and controlled.
[0031]A preferred Notch ligand, used in examples below and described in more detail therein, further comprises a tag, either intrinsic to the ligand or added e.g. in a fusion protein, and being a tag which can be bound by a further medium component, namely a clustering molecule. The clustering molecule binds to two or more tags and hence holds two or more ligands in close proximity to each other. The tag can also be referred to as a clustering epitope, such as a peptide sequence that an antibody will bind to.
[0032]A preferred method of the invention thus further comprises adding tagged ligand and clustering molecule to the culture medium. In use, the clustering molecule binds to two or more tags and holds the ligands together. The ligands bind to Notch receptors on the surface of cells in the culture, and the Notch receptors are as a result held closely together or clustered on the cell surface.
[0033]This clustering is part of activation of Notch signalling, and it is believed that Notch ligands may need to be attached to the cell membrane in order to successfully activate Notch--some attempts using specially engineered soluble forms of Notch ligands either have no effect or they inhibit rather than activate Notch signalling. The ligands may only be active when they are clustered together, and that clustering is facilitated by their localisation in a cell membrane. To achieve Notch activation using soluble ligands that do not cross the cell plasma membrane, a soluble ligand can therefore be combined with a tag that is bound by a separate clustering molecule that binds to tags on two or more ligands, hence forming a clustering molecule(Notch ligand)2 complex and clustering the Notch ligands. For example, fusion proteins of Notch ligands with Fc sequences have been found effective as tagged ligands. Specifically, Carol Hicks in the laboratory of Gerry Weinmaster at UCLA has engineered fusion proteins with Delta or Jagged fused to an Fc molecule. Clustering is initiated by adding an anti-Fc antibody. The Delta-Fc or Jagged-Fc or other tagged ligand is added to the culture medium together with the Fc antibody. Another option is to provide the ligand in multivalent form--hence one ligand binds two or more receptors and clusters the receptors. Notch signaling was activated using a soluble ligand fused to a clustering molecule in Morrison et al., Cell, vol. 101, 26 May 2000, pp 499-510. Notch signaling was activated using F3 as ligand in Hu et al., Cell, vol. 115, 17 Oct. 2004, pp 163-175. Other known Notch ligands are Contactin, Lag2 and Serrate (another term for Jagged).
[0034]The soluble clustered Fc ligands can have different effects if too great an amount of ligand and/or antibody is used--as an excess can lead to antagonism of Notch signalling. The effect of the Fc-ligands can depend in particular upon the concentration of anti-Fc antibody used: too much antibody (thought to result in extra-large clusters) has the same effect as no antibody (no clustering) generating an antagonistic rather than agonistic effect. See: Hicks C, Ladi E, Lindsell C, Hsieh J J, Hayward S D, Collazo A, Weinmaster G. "A secreted Delta1-Fc fusion protein functions both as an activator and inhibitor of Notch1 signalling." J Neurosci Res. 2002 Jun 15;68(6):655-67. As this is well known and documented it is straightforward for the skilled person to avoid excess amounts of the clustering molecule to ensure that only Notch activation is obtained.
[0035]Notch signaling can also be activated by expressing one or more downstream effectors of Notch signalling. For example a product of a gene directly or indirectly targeted by the Notch intracellular domain can be misexpressed. Thus, as an example, Notch signalling can be achieved by expression of a Hes or Hey gene, leading to increased activity of the Hes or Hey transcription factors. Notch activation can be achieved by increased activity of downstream targets of Hes and Hey genes. Notch activation was carried out by expressing Hey genes by Sakamoto et al. as described in J. Biological Chemistry, vol. 278, no. 45, issue of 7 Nov. 2004, pp 44808-44815. Generally, this expression can be achieved using a range of different vectors, such as a transfection vector or with a viral vector, e.g. lentivirus, retrovirus or adenovirus.
[0036]Notch effectors can also comprise or be linked to a transduction domain that enables the Notch effector to cross the plasma membrane of the cells. In such embodiments, the transduction domain enables the effector to cross the plasma membrane and enter the cell from culture medium. Further details of the effectors and domains are given below.
[0037]Notch signalling can also be activated by co-culture with cells that express a Notch ligand, e.g. feeder cells expressing Delta or Jagged. Cells can be engineered to express a Notch ligand and then used for this purpose.
[0038]Notch signalling can be achieved by coating a portion of apparatus used in cell culture with a Notch ligand. For example a culture plate or dish, or a bead present in the culture can have a Notch ligand applied to its surface.
[0039]In embodiments of the invention in which a Notch activator is expressed in a cell, this expression can be constitutive or conditional. Expression can be controlled using recombinase proteins, the Cre-Lox system for example.
[0040]Activation of Notch can, as mentioned, be carried out in conjunction with other steps to promote neural specification. Prior to, at the same time as or subsequent to Notch activation, the cells can be induced to differentiate towards a neural fate.
[0041]This induction can be achieved by change in the medium in which cells are cultured. In one example, the inducing comprises removing serum from medium in which the cells are being cultured or transferring cells into medium free of serum.
[0042]The induction can be achieved by addition or removal of exogenous factors. In one example the inducing comprises adding a factor which promotes neural specification to medium in which the cells are being cultured or transferring the cells into medium containing such a factor. Suitable factors include retinoic acid and BMP antagonists e.g. noggin.
[0043]The induction can be achieved through alteration of the physical nature of the culture. For example, the inducing may comprise transferring the cells to a monolayer culture, transferring cells to serum free culture and/or plating on a PA6 feeder layer.
[0044]Once neural specification has been commenced it is preferred to take steps to purify the resulting culture, so as to increase further the proportion of neural cells. Another property of Notch activation can thus be exploited in the invention as Notch signalling has been found also to prevent or reduce differentiation of neural progenitors. By maintaining activation of Notch signalling, cells can be maintained as neural progenitors for longer periods. Maintenance of Notch activation can be achieved by continuing to maintain Notch ligands in the medium or continuing to express a Notch activator.
[0045]Further improvement of the proportion of neural cells is suitably carried out by removal of non-neural cells. In particular, multipotent or pluripotent cells which have not differentiated may need to be removed. One method is to culture the cells in, or transfer the cells to, medium or culture conditions which are non-permissive for the multipotent or pluripotent cells. In this way, Notch signalling promotes neural specification and non-permissive conditions remove the starting multi- or pluripotent cells.
[0046]Generally, absence of LIF is non-permissive for mouse pluripotent cells. Addition of a reducing agent into the culture is also generally non-permissive for pluripotent cells--for example mercaptoethanol can be added to the culture medium. If cells are replated at lower density this reduces the level of autocrine LIF and is also non-permissive for pluripotent cell survival.
[0047]The cultures may alternatively or additionally be improved using preferential expression in the desired neural cells of a selectable marker. A method of purifying the cells comprises expressing in the neural progenitor cells a selectable marker and selecting for cells expressing the marker. The selectable marker may encode resistance to antibiotic, in which case antibiotic can be used selectively to deplete the culture of non-neural cells, or a cell surface marker or a fluorescent protein, in which case antibodies to the marker or FACS can be used. Other selection strategies are known and described e.g. in EP 0695351.
[0048]As with other aspects of the invention, cells used in the methods are suitably pluripotent cells, especially ES cells, and separately can be rodent or primate cells, especially mouse or human cells. It is particularly preferred that the cells obtained are human neural or neural progenitor cells, obtained from human pluripotent cells.
[0049]In an example of operating within the invention, ES cells were differentiated in the presence of Notch signalling, leading to a mixed culture of ES (i.e. cells which have not differentiated) and NS cells, with a NS proportion of about 90%, of which 60% are radial glial cells and 30% are sox1 positive. This mixed culture was then transferred to medium which does not support ES cell growth or self renewal.
[0050]The invention thus uses Notch signalling to improve control of differentiation of pluripotent cells in culture. One approach is to express an activated Notch receptor. Another approach uses a ligand for cell surface receptors. A further approach is simply to add to the culture medium one or more ligands that bind on or outside and activate Notch inside the cells. Most currently known ligands do not normally function as secreted soluble proteins, so these ligands are preferably combined with a transduction domain, e.g. a soluble version of the Notch intracellular domain fused to that peptide, which can cross the plasma membrane of cells when added to culture medium. This circumvents the need for activation/cleavage of endogenous Notch receptor. This approach has been used successfully with cre recombinase and HoxB4 and is well suited to neural induction systems of the invention in which serum (which can inhibit that fusion protein translocation across membranes) is absent. Hes5 overexpression shows similar effects to Notch intracellular domain overexpression, and hence another specific embodiment of the invention is a that-Hes5 fusion, and this is another media additive. Hes5, when over-expressed, has a similar effect to Notch. Other ligands suitable for use in Notch activation include generally the Hes family, e.g. Hes1, Hes5, Hes6 and the Hey genes.
[0051]In examples set out in detail below we have used a gain of function approach to show that whilst Notch does not interfere with the self-renewal of embryonic stem cells under expansion conditions (LIF+serum) it does bias cells towards specification into the neural lineage after withdrawal of LIF+Serum. Constitutive expression of an activated form of Notch brought about rapid and synchronous neural specification whilst blocking differentiation into non-neural lineages. Furthermore, activated Notch did not bring about terminal differentiation of ES-derived cells, but rather allowed for their expansion as neural progenitors.
[0052]Our data suggest that Notch signalling may be a limiting requirement for neural specification, which is only received by a subset of cells during standard neural differentiation protocols. According to the invention, increasing the number of cells that receive a Notch signal improves the efficiency of neural specification protocols without compromising the ability to expand cells either as ES cells or as neural progenitors.
[0053]The invention has used both genetic and non-genetic means of activating Notch signalling pathways. Delta-Fc, Jagged-Fc and Contactin-Fc can also be used to activate Notch, mimicking the effects seen when miss-expressing activated Notch (NotchIC). There are advantages of avoiding resorting to genetic manipulation and we believe that non-genetic techniques will be favoured in future.
[0054]In examples set out in more detail below, we miss-expressed an activated form of the Notch receptor. Using constitutively active Notch, ES cells can be maintained in culture as ES cells, though with reduced conversion to non-neural cells. If the medium is then changed from LIF+BMP or LIF+serum to a N2B27-based medium, for differentiation, with Notch continuing to be expressed then the cells more rapidly and at a higher % than previously convert to neural cells. The cells become sox1 positive and then BLBP positive and can be kept as BLBP positive cells for several weeks. Some neurons are obtained which don't proliferate. When BMP is added there is no non-neural differentiation (whereas BMP would hitherto have been expected to drive non-neural differentiation).
[0055]The invention brings with it a number of advantages. There is an increased proportion of neural cells and a decreased proportion of non-neural. The contaminating cells tend to be just ES cells, which can be removed by transfer to non-ES supporting medium or adopting non-ES permissive conditions and/or media. We have additionally found activation of Notch signalling to be straightforward to do using both genetic manipulation and external ligands.
[0056]Notch activation has been found not to have a detrimental effect on ES cell self renewal and propagation. This is an advantage as pluripotent cells in which Notch is activated can be cultured as before and then when induced to differentiate Notch activation is used to reduce contamination of the desired neural culture by non-neural cells.
[0057]Increased cell density can be achieved and the cultures have been found to be more robust.
[0058]Further aspects of the invention provide medium additives, media and nucleotides and vectors encoding certain additives.
[0059]A nucleotide sequence of the invention encodes an activated form of a Notch receptor. The activated form of a Notch receptor preferably comprises an intracellular domain of a Notch receptor. This sequence can be used to provide Notch signalling without using external ligands and can be expressed reversibly or constitutively in cells so as to provide continuous Notch signalling. Also preferably, the activated form of a Notch receptor lacks an extracellular domain of a Notch receptor.
[0060]A vector of the invention comprises this nucleotide sequence. The vector of an embodiment of the invention used in examples below comprises a promoter that expresses the activated form of a Notch receptor in pluripotent cells and neural cells. The vector is used to transform cells, e.g. pluripotent cells so as to express the activated receptor.
[0061]A further nucleotide sequence of the invention encodes a downstream effector of Notch signalling. The effector is preferably selected from the Hes and Hey transcription factors. This sequence can also be used to provide Notch signalling without using external ligands and can be expressed reversibly or constitutively in cells so as to provide continuous Notch signalling.
[0062]A further vector of the invention comprises this nucleotide sequence. The vector of an embodiment of the invention used in examples below comprises a promoter that expresses the effector in pluripotent cells and neural cells. The vector is used to transform cells, e.g. pluripotent cells so as to express the effector.
[0063]The invention also provides a composition comprising a downstream effector of Notch signalling or an activated form of a Notch receptor and a transduction domain. This composition can be added to or included in culture medium so as to provide an activator of Notch signalling. The activated form of a Notch receptor typically comprises an intracellular domain of a Notch receptor.
[0064]The transduction domain enables the composition to enter cells. A number of suitable transduction domains are known in the art and reference to a transduction domain or a translation domain refers to a domain or fragment of a protein which effects transport of itself and/or other proteins and substances across a membrane or lipid bilayer and encompasses native domains and fragments, variants and derivatives that retain this binding function. The latter membrane may be that of an endosome where translocation will occur during the process of receptor-mediated endocytosis. Translocation domains can frequently be identified by the property of being able to form measurable pores in lipid membranes at low pH (Shone et al. (1987) Eur J. Biochem. 167, 175-180 describes a suitable test). The latter property of translocation domains may thus be used to identify other protein domains which could function as the translocation domain within the construct of the invention. Examples of translocation domains derived from bacterial neurotoxins are as follows: [0065]Botulinum type A neurotoxin--amino acid residues (449-871) [0066]Botulinum type B neurotoxin--amino acid residues (441-858) [0067]Botulinum type C neurotoxin--amino acid residues (442-866) [0068]Botulinum type D neurotoxin--amino acid residues (446-862) [0069]Botulinum type E neurotoxin--amino acid residues (423-845) [0070]Botulinum type F neurotoxin--amino acid residues (440-864) [0071]Botulinum type G neurotoxin--amino acid residues (442-863) [0072]Tetanus neurotoxin--amino acid residues (458-879)
[0073]Other suitable translocation domains are TAT (e.g. from HIV-1) and penetratin, short sequences of amino acids that internalize covalently linked peptides and convey them, or enable them to be conveyed, to the nucleus. Further suitable domains, referred to as protein transduction domains, such as VP22, derivatives of antennapedia and others, are described in Wadia et al, 2002. These domains can be linked to a Notch ligand or activated form of a Notch receptor chemically, e.g. via thiol functional groups or a fusion can be expressed comprising both components. The linked molecules, the fusions and compositions comprising the same form other aspects of the invention. These can be used e.g. as additives to culture medium as an alternative to transfecting cells with Notch ligands or activated forms of Notch receptors.
[0074]Translocation" in relation to translocation domain, means the internalization events which occur after binding to the cell surface. These events lead to the transport of substances into the cytosol of cells.
[0075]A composition for delivery of a Notch effector or an activated form of a Notch receptor 10 to an ES cell therefore comprises: [0076]the Notch effector or the activated form of a Notch receptor, and [0077]a translocation domain that translocates the Notch effector or the activated form of a Notch receptor into the ES cell.
[0078]The translocation domain can also be selected from (1) a HN domain of a diphtheria toxin, (2) a fragment or derivative of (1) that substantially retains the translocating activity of the HN domain of a diphtheria toxin, (3) a fusogenic peptide, (4) a membrane disrupting peptide, and (5) translocating fragments and derivatives of (3) and (4).
[0079]Further provided by the invention are isolated nucleotides encoding the fusion proteins of the invention and vectors comprising these nucleotides.
[0080]A medium of the invention, for culture of neural cells, comprises a component which activates Notch signalling in cells in the medium.
[0081]The medium may comprise a Notch ligand, which can be a multivalent Notch ligand and capable on its own of clustering Notch receptors. The medium may contain a Notch ligand-tag fusion protein and a clustering molecule which binds to two or more such fusion proteins. The medium may comprise an activated Notch receptor, such as an intracellular domain of a Notch receptor, or a Notch effector, optionally linked to a transduction domain as described above.
[0082]Further medium of the invention may be non-permissive for multipotent or pluripotent cells. This medium can be used to further deplete the culture of cells which have not differentiated into neural cells. Preferred medium is non-permissive for pluripotent cells.
[0083]A still further aspect of the invention provides a method of culture of neural cells to as to increase the density of cells in culture, the method comprising activating Notch signalling in the cells. Notch activation can be carried out as described for all other aspects of the invention. We have found that, in the absence of Notch activation, neural induction declines at cell densities 104 cells per cm2 but that in the presence of Notch activation in accordance with the invention neural induction can be successfully be achieved at densities of 5×104 cells per cm2 and that the resultant cultures are more resistant to small changes in culture conditions and are hence regarded as more robust cultures.
[0084]Another aspect of the invention provides a method of culture of pluripotent cells, preferably ES cells, so as to maintain the cells in a self-renewing state, comprising culturing the cells in medium comprising an agonist of a BMP receptor and in the presence of Notch activation. BMP agonists are suitably BMP 2 and BMP 4. Notch activation is preferably as described herein in respect of the other aspects of the invention.
[0085]A still further aspect of the invention provides a pluripotent cell in which Notch signalling has been activated by any of the embodiments of the invention. The cell is preferably a mouse or human cell and preferably an ES cell. A specific embodiment of this aspect of the invention is an ES cell engineered to express a peptide comprising a Notch intracellular domain.
[0086]The invention is now described in specific examples, illustrated by drawings in which:
[0087]FIG. 1 shows NotchIC ES cells or parental control 46C ES cells maintained in LIF+serum unless otherwise stated;
[0088]FIGS. 2-9 show NotchIC ES cells or parental control 46C cells cultured under monolayer differentiation conditions and the results of analysis of those cultures; and
[0089]FIG. 10 shows human ES cells cultured on feeder cells or under monolayer differentiation conditions and the results of analysis of those cultures.
[0090]In more detail:
[0091]FIG. 1 shows NotchIC ES cells or parental control 46C ES cells maintained in LIF+serum unless otherwise stated. (A-D) Colonies of NotchIC ES cells shown under phase contrast or stained for markers as indicated. E: Populations of NotchIC ES cells or parental control ES cells analysed by FACS passage 12 for expression of sox1-GFP. F: Populations of NotchIC ES cells or parental control ES cells analysed by FACS passage 12 for expression of NotchIC-CD2. G: RT-PCR analysis of Notch IC cells cultured in LIF+Serum (NotchIC ES cells) and of E13.5 embryo neural tissue as a positive control for neural markers.
[0092]FIG. 2 shows NotchIC ES cells or parental control 46C cells cultured under monolayer differentiation conditions. A: Typical FACS profile of sox1-GFP expression after 48h. B: Graph to indicate results of FACS analysis of the proportion of sox1-GFP positive cells at various time points from triplicate cultures. C-H: Intact cultures at 72h of monolayer differentiation, shown in phase contrast or stained for markers as indicated. I: Growth curve indicates the total number of cells at various time points in triplicate cultures. J: Typical FACS profile of sox1-GFP expression after 5 days.
[0093]FIG. 3 shows NotchIC ES cells or parental control 46C cells cultured under monolayer differentiation conditions. A: Graph to indicate results of FACS analysis of the proportion of sox1-GFP positive cells at various time points. B-G: Intact cultures at various time points shown in phase contrast or stained for markers as indicated. H: Quantitative PCR for BLBP during monolayer differentiation. I: Schematic diagram to illustrate the transition of ES cells into sox1-GFP positive neuroepithelial progenitors and then into BLBP+ radial glial neural progenitors.
[0094]FIG. 4 shows NotchIC ES cells or parental control 46C cells cultured under monolayer differentiation conditions and stained for Oct4 (red) to indicate ES cells together with a combination of BLBP and GFP (green) to indicate both types of neural progenitor together.
[0095]In FIG. 5, A, C, D are intact cultures at day 7 of monolayer differentiation, shown in phase contrast or stained for markers as indicated. B: Cells replated onto gelatin after 7 days monolayer differentiation, cultured for a further 7 days in the absence of serum then for the final 7 days in the presence of serum and 100 units/ml LIF, shown in phase contrast or stained for GFAP. F-K: Cells replated onto laminin after 7 days of monolayer differentiation and cultured for a further 5 days (F,G,H: total 12 days), 16 days (J: total 24 days) or 21 days (K: total 28 days) then fixed and stained for markers as indicated.
[0096]FIG. 6 shows the proportion of sox1-GFP cells (A) or GFP expression within intact cultures (B, C) after monolayer differentiation of 46C cells or NotchIC cells exposed to 4 uM gamma secretase inhibitor or to equivalent amounts of DMSO diluent.
[0097]FIG. 7 shows quantitative PCR for FGF5 during monolayer differentiation.
[0098]FIG. 8 shows FACS plots indicating the proportion of sox1-positive cells after monolayer culture of NotchIC cells or parental control cells at medium (104 cells/cm2) or higher (3×104 cells/cm2) densities.
[0099]FIG. 9 shows the results when NotchIC cells or parental control 46C cells were cultured for 3 days in the presence of 4 uM PD compound, SU compound, or in an equivalent concentration of DMSO diluent (`No inhibitor") and the proportion of sox1-GFP positive cells analysed by FACS on day3. Intact cultures were stained for Oct4 on day 5 to visualise undifferentiated ES cells.
[0100]FIG. 10 shows (A-I) Human ES cells plated on OP9 feeder cells expressing either GFP only (OP9 EV) of the Notch ligand Delta1 (OP9 D11) with γ-secretase inhibitor where indicated ("+inhibitor) were cultured for 7 d and stained for markers as indicated. (A) Higher magnification picture to indicate the cell morphology. (J,K) Quantification of Pax6 immunostaining (averages and standard eviations shown from four experiments). (L-T) Human ES cells grown under mono-layer differentiation conditions in the presence of γ-secretase inhibitor ("inhibitor") or DMSO vehicle and stained for Pax6,Sox1, or TRA1/81 as indicated. T shows quantification of Pax6 immunostaining (averages and standard deviations from four experiments,
[0101]Specific embodiments of the invention provide or use one or more of the following sequences, referred to herein by their SEQ ID NO:
TABLE-US-00001 SEQ ID NO Description 1 Notch full length DNA 2 Notch full length amino acid 3 Notch intracellular DNA 4 Notch intracellular amino acid 5 Hey1 DNA 6 Hey1 amino acid 7 Hey2 DNA 8 Hey2 amino acid 9 Hes1 DNA 10 Hes1 amino acid 11 Hes3 DNA 12 Hes3 amino acid 13 Hes5 DNA 14 Hes5 amino acid 15 Hes6 DNA 16 Hes6 amino acid 17 Fusion of NotchIC-tat 18 Fusion of NotchIC-protein transduction domain from antennapedia 19 Fusion of tat-Hey2 20 Fusion of tat-Hes5
EXAMPLES
[0102]Materials and Methods
[0103]Targeting NotchIC into 46C ES cells
[0104]Our targeting construct was based on similar constructs previously used for targeting into the ROSA locus. It contains a pgk-neo cassette flanked by loxP sites, followed by the coding sequence for the intracellular domain of NotchIC (Kopan et al., 1994) followed by an internal ribosomal entry site followed by the coding sequence for the human cell surface molecule CD2. This construct was transfected into 46C ES cells (Ying et al., 2003b) by electroporation, and clones were expanded under G418 selection. Correctly targeted clones were identified by Southern blotting after digestion with EcoRV. DNA gives an 11 kb band whilst untargeted wild type cells give a 3.8 kb fragment. A clone of targeted cells were transfected with a plasmid containing CRE under the control of the pCAG promoter.
[0105]ES cell culture
[0106]ES cells were maintained in GMEM supplemented with 2-mercaptoethanol, non-essential amino acids, sodium bicarbonate, 10% fetal calf serum (FCS) and 100 units/ml LIF on gelatinised tissue culture flasks (Smith, 1991).
[0107]Monolayer Differentiation
[0108]This is as described in detail in Ying et al., 2003b. Briefly, ES cells were washed to remove all traces of serum and then plated on gelatin-coated tissue culture plastic at a density of 1>104 cells/cm2 in N2B27 serum-free medium. N2B27 consists of a 1:1 ratio of DM/F12 and Neurobasal media supplemental with 0.5% N2 (made in house as described in (Ying et al., 2003b)), 0.5% B27 (Gibco) and 2-mercaptoethanol. Media was changed every second day.
[0109]In some experiments, the culture medium was supplemented with 100 units/ml LIF and with 10% FCS. The MAPK inhibitor PD184352 (gift of P. Cohen, Univ Dundee) was used at a concentration of 4 μM. The gamma secretase inhibitor (Calbiochem cat. 565771) was used at a concentration of 4 μM. Neither of these inhibitors had any obvious toxic effects over the time course of the experiments.
[0110]Immunofluorescence
[0111]Cells were fixed in 4% paraformaldehyde and incubated for 30 minutes in blocking buffer (PBS, 2% Goat serum and 0.1% Triton). Primary antibodies were diluted in blocking buffer and applied for 1 h at room temperature. After three washes in PBS, secondary antibodies conjugated to Alexa fluorophores (Molecular Probes) were diluted at 1:1000 in blocking buffer and applied for 1 h at room temperature. The cells were washed at least three times in PBS and visualised on a Olympus inverted fluorescence microscope.
[0112]In experiments where cells were counted, nuclei were counterstained with DAPI and at least 1000/cells per culture were counted from three separate cultures and an average taken.
[0113]Primary antibodies were obtained from the following sources:
[0114]Human CD2 (BD Biosciences); Oct4 (Santa Cruz); GFP (Molecular Probes); Nestin (DSHB); BLBP (Gift); Neuronal beta-III tubulin (Covance); GFAP (Sigma); O4 (DSHB); RC2 (DSHB). Antibodies obtained from the Developmental Studies Hybridoma Bank were developed under the auspices of the NICHD and maintained by The University of Iowa, Department of Biological Sciences, Iowa City, Iowa 52242.
[0115]Human ES Cell Culture and Differentiation
[0116]Undifferentiated human ES cells were maintained on a layer of human foreskin fibroblast (ATCC, Manassas, Va., United States) in the defined medium N2B27 supplemented with LIF (10 ng/ml), BMP-4 (3 ng/ml; R & D Systems, Minneapolis, Minn., United States), and bFGF (10 ng/ml; R & D Systems). Cells were passaged at a split ratio of 1:2 every week using collagenase IV (1 mg/ml). Feeder-free neural differentiation was performed following the monolayer protocol used for mouse cells, modified to suit human ES cells as follows: cells reaching 50% confluence were incubated in collagenase IV for 15 min, washed once in PBS and detached in N2B27 medium supplemented with bFGF (FGF medium, no LIF, no BMP4) using glass beads. Cells were then incubated for 4 h in a gelatinised flask in the same medium to allow the differential attachment of the feeder cells. Finally, the ES suspension was plated at a 1:1 ratio in culture dishes pre-coated with Matrigel (low growth factor Matrigel, 1:20; BD Biosciences PharMingen, San Diego, Calif., United States). When indicated, the γ-secretase inhibitor (4 μM) was added from the start of the feeder-removal step and then added every other day when the medium was changed. For coculture with OP-9 cells, hES cells were treated with collagenase as described above and then manually detached to avoid carry-over of human fibroblasts. Cells were then directly plated in FGF medium on a layer of γ-irradiated OP9-EV or OP9-D11 stromal cells (kindly provided by A. Cumano). Matrigel was used to promote the survival of the OP9 in the serum-free medium. The γ-secretase inhibitor was added at plating and then added every other day when the medium was changed.
[0117]Quantification of Neural Differentiation of Human ES Cells.
[0118]Cells were processed for immunocytochemistry and neural differentiation quantified as follows. For OP9 coculture experiments, the number of colonies with positive PAX-6 cells was counted and normalised to the total number of colonies in the well. For all experiments (feeder-free and OP9-dependent differentiation), Velocity image analysis software (Improvision, Lexington, Mass., United States) was used to quantify the extent of differentiation. Briefly, the software was used to calculate the area of the well (feeder-free experiments) or of each colony (OP9 experiments) covered by PAX-6-positive nuclei. The values were then normalised to the area covered by the cells or colonies, using DAPI staining. All experiments were repeated at least three times with four wells per condition.
Example 1-1
[0119]ES Cells and Early Neural Progenitors Express Notch Receptors and Ligands
[0120]We used RT-PCR to confirm that Notch1 and Jagged2 are expressed in ES cells and their earliest neural derivatives.
Example 1-2
[0121]Generation of 46C Cells Constitutively Expressing NotchIC
[0122]We generated a targeting construct in which a sequence encoding the intracellular domain of Notch (Kopan et al., 1994) was preceded by a floxed stop-pgk-neo cassette, and followed by a sequence encoding IRES-human CD2. Human CD2 is a cell surface molecule with no phenotypic effect on mouse cells, used here as tag to indicate NotchIC missexpression. This construct was targeted into the ROSA locus of 46C cells. 46C cells are a line of ES cells that contain the coding sequence of GFP targeted into one allele of the early neural marker sox1, and thus act as a convenient experimental system for monitoring neural induction [Stavridis, 2003 #221].
[0123]A clonal line of NotchIC-targeted ES cells was transfected with CRE in order to excise the stop codon and activate constitutive transcription of NotchIC-IRES-CD2. The successfully deleted population (designated "NotchIC cells") was separated from the undeleted population by FACS based on CD2 expression. The undeleted population was used as a control and these are referred to as "undeleted controls". Parental 46C cells (not targeted with NotchIC) were used as a second control population, and these are referred to as "parental 46C controls". Both control populations gave similar results in all experiments.
Example 1-3
[0124]NotchIC ES Cells are Indistinguishable from Control ES Cells
[0125]Notch IC cells showed no difference in growth rate or morphology compared to either undeleted controls or parental 46C control ES cells under standard culture conditions for maintaining undifferentiated proliferative ES cells (LIF and serum). They grew at a comparable rate over more than 20 passages (data not shown) expressed markers of undifferentiated ES cells, and lacked markers of differentiation (FIG. 1 and data not shown).
Example 2-1
[0126]Populations of NotchIC Cells Undergo Neural Specification More Rapidly and Homogenously than Controls
[0127]We next tested the effect of NotchIC on neural specification. We transferred NotchIC or control ES cells into a neural differentiation protocol that is based on adherent monolayer culture in the absence of exogenous growth factors (Ying and Smith, 2003). Cells were disaggregated and analysed by FACS for sox1-GFP expression every 12 hours during monolayer differentiation. Control ES cells generate sox1-GFP+ cells gradually: fewer than 2% could be detected after 24 h, and only 1111% after 48 h, In contrast, NotchIC cultures contained 9±1% sox1-GFP+ cells after only 24 h, increasing to 32±3% by the second day (FIG. 2A, B).
[0128]The distribution of sox1-GFP+ cells was also monitored in intact monolayer differentiation cultures, using an inverted fluorescence microscope. Cultures of control ES cells contain GFP-positive cells interspersed with GFP-negative cells in a "salt and pepper" pattern (FIG. 2C,E). In NotchIC cultures, distribution of GFP+ cells is more homogenous (FIG. 2D,F). There is also a difference in the variability of GFP intensity between cells: NotchIC cells have uniformly weak GFP expression whilst control cultures contain a mixture of bright and dim GFP cells (FIG. 2E, F: see also FACS profile J). Staining for another early neural marker, nestin, was consistent with entry of the NotchIC cells into the neural lineage (FIG. 2G, H).
[0129]FGF5 is a marker of primitive ectoderm, expressed transiently at an intermediate stage of differentiation of pluripotent ES cells towards neural tissue. Control ES cells acquire FGF5 at increasing levels over the first few days of monolayer differentiation, after which it declines as Sox1 expression increases (FIG. 7). In contrast, NotchIC-overexpressing ES cells have a maximal peak of high FGF5 expression after just 24 h differentiation, in keeping with the rapid induction of sox1in these cells (FIG. 7). This data also shows that NotchIC promotes the transition of ES cells into the neural lineage.
Example 2-2
[0130]NotchIC does not Increase Proliferation
[0131]The high proportion of sox1-GFP+ cells that emerge early during monolayer differentiation of NotchIC ES cells could be explained by an increase in the rate of conversion of ES cells to sox1+neural cells. Alternatively, it could be that NotchIC does not affect neural specification but instead increases the rate of proliferation in neural cells. We consider that the second possibility is unlikely because there are significant numbers of sox1-GFP+ cells even after only 24 h, before the onset of neural specification in control ES cell populations (FIG. 2B). Furthermore, the growth rate of NotchIC and control populations is indistinguishable during the course of the experiment (FIG. 2I).
Example 2-3
[0132]NotchIC Overcomes the Inhibitory Effects of High Cell Density on Neural Specification of ES Cells
[0133]Neural induction of normal ES cells during monolayer differentiation is strongly inhibited by even modest increases in cell density (FIG. 8). We found that NotchIC cells are resistant to the inhibitory effects of increasing cell density on neural differentiation.
Example 2-4
[0134]NotchIC does not Bypass the Requirement for FGF Signalling in Neural Specification
[0135]Although differentiation of ES cells into neural progenitors does not require exogenous growth factors, it is dependant upon FGF signalling, which is most likely provided by autocrine FGF4 (Ying and Smith, 2003). We found that NotchIC does not bypass this requirement: the MAPK inhibitor: PD184352 and the FGFR inhibitor SU5402 were both able to block induction of sox1 in NotchIC cells (FIG. 9).
Example 3-1
[0136]Sox1 Induction is Rapidly Followed by BLBP Induction
[0137]Sox1-GFP is expressed only transiently during monolayer differentiation of NotchIC cells. The intensity of sox1-GFP within each individual cell does not accumulate to high level in comparison to control cells (FIG. 2: compare E and F; also FIG. 2J), whilst expression within the NotchIC population as a whole reaches a plateau by day 3 and begins to decline soon afterwards (FIG. 3A).
[0138]Between day 4 and day 6, the majority of NotchIC cells undergo a striking morphological change to become bipolar (FIG. 3C). Control ES cells only rarely develop this bipolar morphology at this time (FIG. 3F). These bipolar cells express BLBP (FIG. 3 C,D) and RC2 (not shown). Furthermore, we found that expression of BLBP preceded any overt morphological change, appearing as early as the third day of monolayer differentiation (FIG. 3A). By the fifth day, at least 50% of NotchIC cells express BLBP, compared with fewer than 5% of control cells. At this time, there was little or no terminal differentiation into neurons (TUJ1 immunostaining), astrocytes (GFAP immunostaining) or oligodendrocytes (O4 immunostaining) (data not shown).
[0139]Quantitative PCR confirmed that BLBP was rapidly induced in NotchIC cells during the first few days of monolayer differentiation, whereas it remained undetectable in control cultures for the first five days (FIG. 3H)
[0140]BLBP-positive RC2-positive radial glia are descendants of sox1-positive neuroepithelial cells in vivo. The term glia is misleading: these cells are a major source of both neurons and glia in vivo and should be considered neural progenitors rather than differentiated glial cells. ES-derived neural cells mimic their in-vivo counterparts, progressing over time from an early sox1-postive progenitor to a later BLBP-positive progenitor (Conti, Pollard et al: (Bibel et al., 2004): see also FIG. 3, E, F, G). In keeping with these previous reports, we now find that NotchIC ES-derived early neural cells rapidly transform into BLBP+/neural progenitors, and this explains why numbers of sox1-GFP cells do not continue to increase beyond 3 days in monolayer differentiation.
[0141]Sox1-GFP expression is downregulated as BLBP is upregulated, such that the two markers are generally mutually exclusive (although the earliest BLBP+ cells do sometimes coexpress weak levels of GFP, possibly due to perdurance of GFP protein. data not shown). The total number of neural progenitors in day 5 monolayer can therefore be estimated by adding the number of sox1GFP+cells (around 30%) to the number of morphologically mature BLBP+ cells (more than 50%) to give a total of more than 80% neural cells.
[0142]FIG. 4 shows typical cultures stained in green for both BLBP and Sox1-GFP in order to visualise all neural progenitors together, counterstained in red for Oct4 to indicate undifferentiated ES cells. In NotchIC cultures, the vast majority of the cells express neural markers by day 3 (FIG. 4A), with the only cells that resist neural differentiation being a minor subpopulation of undifferentiated ES cells (possibly due to autocrine LIF signalling). After three further days of differentiation, it is still the case that practically all cells can be accounted for by expression of either neural or ES markers (FIG. 4C). This contrasts with control cultures, which contain 15-30% of cells that lack both neural and ES cell markers, and which by day 6 have the morphology of non-neural differentiated cell types (FIG. 4B, D). These observations further confirm the rapid and relatively homogenous conversion of ES cells into neural progenitors. They also indicate that Notch biases differentiation in favour of neural and away from non-neural lineages.
Example 4
[0143]NotchIC Allows for the Maintenance of Neural Progenitors Rather than Promoting their Rapid Terminal Differentiation
[0144]The Notch signalling pathway has been reported to either promote self-renewal of neural stem cells (Ohtsuka et al., 2001), or to promote their differentiation into astrocytes (Tanigaki et al., 2001), depending upon context in which the gain of function experiments were carried out. In monolayer differentiation cultures, the majority of NotchIC cells persist as BLBP+nestin+RC2+neural progenitors for at least two to three weeks (FIG. 5F, G, H, I, J) (they do not survive much longer than this in the absence of exogenous growth factors). A subpopulation of cells differentiates into neurons during the second week of culture (FIG. 5A,H) but these neurons are outnumbered by undifferentiated BLBP+ cells (FIG. 5C, D, F-H). Astrocyte differentiation occurs only very rarely during the first two weeks (<1%: data not shown). Astrocytes begin to emerge during the third week of serum-free culture, but remain in the minority (FIG. 5K). Oligodendrocytes were never detected. NotchIC did not bring about rapid differentiation into astrocytes or into any other terminally differentiated cell types in our culture system, but rather allowed for the expansion of neural progenitors.
Example 5
[0145]NotchIC Neural Progenitors can Efficiently Differentiate into Astrocytes upon Exposure to Serum
[0146]The observation that most NotchIC neural progenitors either differentiate into neurons during the second week, or else remain undifferentiated for at least three weeks, raises the question of whether these cells have significant glial as well as neuronal potential.
[0147]In order to address this question, we treated the cells with LIF and serum, which are potent inducers of astrocyte differentiation from late neural progenitors. NotchIC ES-derived neural progenitors, like normal ES-derived neural cells (and similarly to or early embryonic neural cells) remain resistant to this effect of LIF or serum during their first 10-14 days. However, if transferred to serum-containing medium after 14 days than they efficiently (>70%) differentiate into GFAP-positive astrocytes over the subsequent 4 days (FIG. 5B)
[0148]The observation that the NotchIC-ES derived cells are able to generate significant numbers of neurons during the first 10 days of serum-free culture, and that they are also able to differentiate into astrocytes with high efficiency after three weeks culture and exposure to serum, confirms that they are neural progenitors.
Example 6
[0149]NotchIC and Neural Specification of ES Cells
[0150]A critical step in activating Notch in vivo is its cleavage by gamma secretase to release the intracellular domain. Gamma secretase inhibitors are effective inhibitors of Notch activity. One problem with these inhibitors is that they are not specific to Notch: they also inhibit cleavage by gamma secretase of other molecules. In our experiments we can use NotchIC cells as a negative control for any non-Notch-specific effects of the inhibitor; Since these cells already contain a pre-cleaved NotchIC fragment, they will be immune to effects of the gamma secretase inhibitor on Notch cleavage, whilst remaining vulnerable to any non-notch-specific effects of the inhibitor.
[0151]The gamma secretase inhibitor is able to significantly reduce induction of sox1during monolayer differentiation. This appears to be a specific effect on the Notch pathway because there is no significant effect on induction from sox1from cells expressing the constitutively active form of Notch (FIG. 6).
[0152]An alternative loss-of-function approach, using mutant ES cellswe have obtained from Tim Schroeder (GSF, Munich) which lack the critical downstream mediator of Notch signalling, RBPJk (Schroeder et al., 2003), is to test whether they fail to differentiate into neural cells and if so whether this can be rescued by transfection with RBPJk plasmid.
Example 7
[0153]Notch Promotes Neural Specification in Human ES Cells
[0154]We investigated whether the role of Notch in neural differentiation is conserved in human ES (hES) cells. We first confirmed that the Notch ligands Jagged1, Jagged2, Delta1, and Delta3 could all be readily detected in human ES cells by RT-PCR (FIG. S7). Mis-expression of Notch ligands in feeder cells has been shown to activate Notch in other cell types, so we decided to employ this strategy with hES cells.
[0155]We made use of OP9 cells that stably express the Notch ligand Delta1 together with GFP, through retroviral transduction (OP9-Delta1). Control OP9 feeder cells had been transduced with a GFP-only retrovirus (OP9-EV). When hES cells were plated onto OP9-EV feeder layers in serum-free medium containing bFGF but no LIF or BMP4, the majority of cells maintained an ES-like morphology after 1 wk. In contrast, when cells were plated onto OP9-D11 feeders in the same medium, the edges of the colonies, where they contact the GFP+ OP9-D11 feeder cells, underwent a morphological change within the first week (FIG. 10). They became compact and elongated with barely distinguishable nuclei (FIG. 10A, region between dotted lines). Antibody staining confirmed that these cells were negative for the ES markers TRA1/60 and TRA 1/81 (unpublished data) and positive for Sox1, Nestin, and Pax6 (FIG. 10B, 10C, and unpublished data). In human ES cells, Pax6 is the earliest known marker of neural differentiation, appearing several days before Sox1 begins to be expressed. We carried out quantification of this marker using image analysis software. This confirmed a significant increase in both the number of Pax6+ colonies (colonies containing more than ten Pax6+ cells) and in the area that is Pax6+ within each of these colonies in OP9-D11-supported cultures in comparison with OP9-EV colonies (FIG. 10D-10G, 10J, and 10K, p<0.01). The positive effect of OP9-D11 feeders on neural differentiation appeared to be specifically due to activation of Notch signalling, because it could be blocked by adding the gamma secretase inhibitor (FIG. 10H-10K, p<0.01). Exposure to neither Deltal nor the γ-secretase inhibitor had any discernible effect on cell number or viability.
[0156]We went on to test whether endogenous Notch signalling is required for neural differentiation in hES cells. For these experiments, we made use of a monolayer neural differentiation protocol adapted from that for mouse ES cells. Briefly, we removed the hES from feeders and from exogenous LIF and BMP4 and plated them onto Matrigel in FGF-only serum-free medium. Under these conditions, typically around 60% of the culture area loses expression of the ES cell marker TRA1/81, adopts a neural morphology, and becomes Sox1+ Pax6+ after 1 wk (FIG. 10L, 10M, 10P, 10R, and 10T). In contrast, in the presence of the γ-secretase inhibitor, there is a significant reduction in emergence of Pax6+ regions within the culture (FIGS. 10M, 10O, 10Q, 10S, and 10T, p<0.5), with a corresponding increased persistence of undifferentiated TRA1/81+ES cells (FIG. 10O). The γ-secretase inhibitor had no obvious effect on cell viability or cell number, and the majority of treated cells retained a healthy hES-like morphology (FIG. 10M).
[0157]These data show that Notch promotes neural differentiation in human ES cells.
Example 8
[0158]NotchIC Suppresses Nonneural Differentiation
[0159]We carried out quantitative RT-PCR to measure the expression of endoderm and mesoderm markers on the sixth day of monolayer differentiation. Several nonneural markers were readily detected in parental cell samples, in marked contrast to the barely detectable expression levels in R26NotchIC cell. These observations indicate that not only does Notch promote neural lineage entry but it also simultaneously suppresses nonneural commitment.
[0160]R26-NotchIC ES also showed a marked reduction in mesoderm differentiation when tested under an inductive differentiation protocol based on monolayer culture on collagen IV in the presence of batch-tested serum.
REFERENCES
[0161]Bain, G., Ray, W. J., Yao, M., and Gottlieb, D. I. (1996). Retinoic acid promotes neural and represses mesodermal gene expression in mouse embryonic stem cells in culture. Biochem Biophys Res Commun 223, 691-4. [0162]Bibel, M., Richter, J., Schrenk, K., Tucker, K. L., Staiger, V., Korte, M., Goetz, M., and Barde, Y. A. (2004). Differentiation of mouse embryonic stem cells into a defined neuronal lineage. Nat Neurosci 7, 1003-9. [0163]Gaiano, N., Nye, J. S., and Fisbell, G. (2000). Radial glial identity is promoted by Notch1 signalling in the murine forebrain. Neuron 26, 395-404. [0164]Hitoshi, S., Alexson, T., Tropepe, V., Donoviel, D., Elia, A. J., Nye, J. S., Conlon, R. A., Mak, T. W., Bernstein, A., and van der Kooy, D. (2002). Notch pathway molecules are essential for the maintenance, but not the generation, of mammalian neural stem cells. Genes Dev 16, 846-58. [0165]Kopan, R., Nye, J. S., and Weintraub, H. (1994). The intracellular domain of mouse Notch: a constitutively activated repressor of myogenesis directed at the basic helix-loop-helix region of MyoD. Development 120, 2385-96. [0166]Lai, E. C. (2004). Notch signalling: control of cell communication and cell fate. Development 131, 965-73. [0167]Li, Y., and Baker, N. E. (2001). Proneural enhancement by Notch overcomes Suppressor-of-Hairless repressor function in the developing Drosophila eye. Curr Biol 11, 330-8. [0168]Morrison, S. J., Perez, S. E., Qiao, Z., Verdi, J. M., Hicks, C., Weinmaster, G., and Anderson, D. J. (2000). Transient Notch activation initiates an irreversible switch from neurogenesis to gliogenesis by neural crest stem cells. Cell 101, 499-510. [0169]Nakamura, Y., Sakakibara, S., Miyata, T., Ogawa, M., Shimazaki, T., Weiss, S., Kageyama, R., and Okano, H. (2000). The bHLH gene hesI as a repressor of the neuronal commitment of CNS stem cells. J Neurosci 20, 283-93. [0170]Ohtsuka, T., Sakamoto, M., Guillemot, F., and Kageyama, R. (2001). Roles of the basic helix-loop-helix genes Hes1 and Hes5 in expansion of neural stem cells of the developing brain. J Biol Chem 276, 30467-74. [0171]Schroeder, T., Fraser, S. T., Ogawa, M., Nishikawa, S., Oka, C., Bornkamm, G. W., Honjo, T., and Just, U. (2003). Recombination signal sequence-binding protein Jkappa alters mesodermal cell fate decisions by suppressing cardiomyogenesis. Proc Natl Acad Sci USA 100, 4018-23. [0172]Smith, A. G. (1991). Culture and differentiation of embryonic stem cells. J. Tiss. Cult. Meth 13, 89-94. [0173]Tanigaki, K., Nogaki, F., Takahashi, J., Tashiro, K., Kurooka, H., and Honjo, T. (2001). Notch1 and Notch3 instructively restrict bFGF-responsive multipotent neural progenitor cells to an astroglial fate. Neuron 29, 45-55. [0174]Ying, Q. L., Nichols, J., Chambers, I., and Smith, A. (2003a). BMP induction of Id proteins suppresses differentiation and sustains embryonic stem cell self-renewal in collaboration with STAT3. Cell 115, 281-92. [0175]Ying, Q. L., and Smith, A. G. (2003). Defined conditions for neural commitment and differentiation. Methods Enzymol 365, 327-41. [0176]Ying, Q. L., Stavridis, M., Griffiths, D., Li, M., and Smith, A. (2003b). Conversion of embryonic stem cells into neuroectodermal precursors in adherent monoculture. Nat Biotechnol 21, 183-6.
[0177]The invention thus provides neural directed differentiation of pluripotent cell cultures, and methods and compositions therefor.
Sequence CWU
1
2017596DNAMus musculusCDS(1)..(7596) 1atg cca cgg ctc ctg acg ccc ttg ctc
tgc cta acg ctg ctg ccc gcg 48Met Pro Arg Leu Leu Thr Pro Leu Leu
Cys Leu Thr Leu Leu Pro Ala1 5 10
15ctc gcc gca aga ggc ttg aga tgc tcc cag cca agt ggg acc tgc
ctg 96Leu Ala Ala Arg Gly Leu Arg Cys Ser Gln Pro Ser Gly Thr Cys
Leu20 25 30aat gga ggt agg tgc gaa gtg
gcc agc ggc act gaa gcc tgt gtc tgc 144Asn Gly Gly Arg Cys Glu Val
Ala Ser Gly Thr Glu Ala Cys Val Cys35 40
45agc gga gcc ttt gtg ggc caa cga tgc cag gac tcc aat cct tgc ctc
192Ser Gly Ala Phe Val Gly Gln Arg Cys Gln Asp Ser Asn Pro Cys Leu50
55 60agc aca ccg tgt aag aat gct gga acg tgc
cac gtt gtg gac cat ggt 240Ser Thr Pro Cys Lys Asn Ala Gly Thr Cys
His Val Val Asp His Gly65 70 75
80ggc act gtg gat tat gcc tgc agc tgt ccc ctg ggt ttc tct ggg
ccc 288Gly Thr Val Asp Tyr Ala Cys Ser Cys Pro Leu Gly Phe Ser Gly
Pro85 90 95ctc tgc ctg aca cct ctg gac
aac gcc tgc ctg gcc aac ccc tgc cgc 336Leu Cys Leu Thr Pro Leu Asp
Asn Ala Cys Leu Ala Asn Pro Cys Arg100 105
110aat ggg ggc acc tgt gac ctg ctc act ctc aca gag tac aag tgc cgc
384Asn Gly Gly Thr Cys Asp Leu Leu Thr Leu Thr Glu Tyr Lys Cys Arg115
120 125tgc cca cca ggg tgg tca gga aaa tca
tgt cag cag gct gac ccc tgt 432Cys Pro Pro Gly Trp Ser Gly Lys Ser
Cys Gln Gln Ala Asp Pro Cys130 135 140gcc
tcc aac ccc tgt gcc aat ggt ggc cag tgc ctg ccc ttt gag tct 480Ala
Ser Asn Pro Cys Ala Asn Gly Gly Gln Cys Leu Pro Phe Glu Ser145
150 155 160tca tac atc tgt cgc tgc
ccg cct ggc ttc cat ggc ccc acc tgc agg 528Ser Tyr Ile Cys Arg Cys
Pro Pro Gly Phe His Gly Pro Thr Cys Arg165 170
175caa gat gtt aat gag tgc agc cag aac cct ggg ctg tgc cgc cat gga
576Gln Asp Val Asn Glu Cys Ser Gln Asn Pro Gly Leu Cys Arg His Gly180
185 190ggc acc tgc cac aat gag atc ggc tcc
tat cgc tgt gcc tgc cgt gcc 624Gly Thr Cys His Asn Glu Ile Gly Ser
Tyr Arg Cys Ala Cys Arg Ala195 200 205acc
cat act ggt ccc cac tgt gaa ctg ccc tat gtg ccc tgc agc ccc 672Thr
His Thr Gly Pro His Cys Glu Leu Pro Tyr Val Pro Cys Ser Pro210
215 220tca ccc tgc cag aat gga ggc acc tgc cgt cct
aca ggg gac acc acc 720Ser Pro Cys Gln Asn Gly Gly Thr Cys Arg Pro
Thr Gly Asp Thr Thr225 230 235
240cac gag tgt gcc tgc ttg cca ggt ttt gct gga cag aac tgt gaa gaa
768His Glu Cys Ala Cys Leu Pro Gly Phe Ala Gly Gln Asn Cys Glu Glu245
250 255aat gtg gat gac tgt cca gga aac aac
tgc aag aat ggg ggt gcc tgt 816Asn Val Asp Asp Cys Pro Gly Asn Asn
Cys Lys Asn Gly Gly Ala Cys260 265 270gtg
gac ggc gtg aat acc tac aat tgc cgc tgc cca ccg gag tgg acg 864Val
Asp Gly Val Asn Thr Tyr Asn Cys Arg Cys Pro Pro Glu Trp Thr275
280 285ggt cag tac tgt aca gag gat gtg gac gaa tgt
cag ctc atg ccc aat 912Gly Gln Tyr Cys Thr Glu Asp Val Asp Glu Cys
Gln Leu Met Pro Asn290 295 300gcc tgc cag
aat ggc gga acc tgc cac aac aca cac ggc ggc tac aac 960Ala Cys Gln
Asn Gly Gly Thr Cys His Asn Thr His Gly Gly Tyr Asn305
310 315 320tgt gtg tgt gtc aat ggg tgg
act ggc gag gac tgc agt gag aac att 1008Cys Val Cys Val Asn Gly Trp
Thr Gly Glu Asp Cys Ser Glu Asn Ile325 330
335gat gac tgt gcc agt gcc gcc tgt ttc cag ggt gcc act tgc cac gac
1056Asp Asp Cys Ala Ser Ala Ala Cys Phe Gln Gly Ala Thr Cys His Asp340
345 350cgt gtg gct tcc ttc tac tgc gaa tgt
ccg cat ggg cgc aca ggt ctg 1104Arg Val Ala Ser Phe Tyr Cys Glu Cys
Pro His Gly Arg Thr Gly Leu355 360 365ctg
tgc cac ctc aac gat gcg tgc atc agc aac ccc tgc aac gag ggc 1152Leu
Cys His Leu Asn Asp Ala Cys Ile Ser Asn Pro Cys Asn Glu Gly370
375 380tcc aac tgt gac acc aac cct gtc aac ggc aaa
gcc atc tgc acc tgc 1200Ser Asn Cys Asp Thr Asn Pro Val Asn Gly Lys
Ala Ile Cys Thr Cys385 390 395
400ccc tcg ggg tac aca ggg cca gcc tgc agc cag gac gtg gat gag tgt
1248Pro Ser Gly Tyr Thr Gly Pro Ala Cys Ser Gln Asp Val Asp Glu Cys405
410 415gct ctg ggt gcc aac cct tgt gag cac
gca ggc aaa tgc ctc aac aca 1296Ala Leu Gly Ala Asn Pro Cys Glu His
Ala Gly Lys Cys Leu Asn Thr420 425 430ctg
ggt tct ttt gag tgc cag tgt cta cag ggc tac acg gga ccc cgc 1344Leu
Gly Ser Phe Glu Cys Gln Cys Leu Gln Gly Tyr Thr Gly Pro Arg435
440 445tgt gag att gat gtt aat gag tgc atc tcc aac
cca tgt cag aat gat 1392Cys Glu Ile Asp Val Asn Glu Cys Ile Ser Asn
Pro Cys Gln Asn Asp450 455 460gcc act tgc
ctg gac cag att ggg gag ttc caa tgc ata tgt atg cca 1440Ala Thr Cys
Leu Asp Gln Ile Gly Glu Phe Gln Cys Ile Cys Met Pro465
470 475 480ggt tat gaa ggt gta tac tgt
gaa atc aac acg gat gag tgc gcc agc 1488Gly Tyr Glu Gly Val Tyr Cys
Glu Ile Asn Thr Asp Glu Cys Ala Ser485 490
495agc ccc tgt ctg cac aat ggc cac tgc atg gac aag atc aat gag ttc
1536Ser Pro Cys Leu His Asn Gly His Cys Met Asp Lys Ile Asn Glu Phe500
505 510caa tgt cag tgc ccc aaa ggc ttc aac
ggg cac ctg tgc cag tat gat 1584Gln Cys Gln Cys Pro Lys Gly Phe Asn
Gly His Leu Cys Gln Tyr Asp515 520 525gtg
gat gag tgt gcc agc aca cca tgc aag aac ggt gcc aag tgc ctg 1632Val
Asp Glu Cys Ala Ser Thr Pro Cys Lys Asn Gly Ala Lys Cys Leu530
535 540gat ggg ccc aac acc tat acc tgc gtg tgt aca
gaa ggt tac aca ggg 1680Asp Gly Pro Asn Thr Tyr Thr Cys Val Cys Thr
Glu Gly Tyr Thr Gly545 550 555
560acc cac tgc gaa gtg gac att gac gag tgt gac cct gac ccc tgc cac
1728Thr His Cys Glu Val Asp Ile Asp Glu Cys Asp Pro Asp Pro Cys His565
570 575tat ggt tcc tgt aag gat ggt gtg gcc
acc ttt acc tgc ctg tgc cag 1776Tyr Gly Ser Cys Lys Asp Gly Val Ala
Thr Phe Thr Cys Leu Cys Gln580 585 590cca
ggc tac aca ggc cat cac tgt gag acc aac atc aat gag tgc cac 1824Pro
Gly Tyr Thr Gly His His Cys Glu Thr Asn Ile Asn Glu Cys His595
600 605agc caa ccg tgc cgc cat ggg ggc acc tgc cag
gac cgt gac aac tcc 1872Ser Gln Pro Cys Arg His Gly Gly Thr Cys Gln
Asp Arg Asp Asn Ser610 615 620tac ctc tgc
tta tgc ctc aag gga acc aca ggg ccc aac tgt gag atc 1920Tyr Leu Cys
Leu Cys Leu Lys Gly Thr Thr Gly Pro Asn Cys Glu Ile625
630 635 640aac ctg gat gac tgc gcc agc
aac ccc tgt gac tct ggc acc tgt ctg 1968Asn Leu Asp Asp Cys Ala Ser
Asn Pro Cys Asp Ser Gly Thr Cys Leu645 650
655gac aag att gat ggc tac gaa tgt gcc tgt gaa cca ggc tac aca gga
2016Asp Lys Ile Asp Gly Tyr Glu Cys Ala Cys Glu Pro Gly Tyr Thr Gly660
665 670agc atg tgt aac gtc aac att gac gaa
tgt gcg ggc agc ccc tgc cac 2064Ser Met Cys Asn Val Asn Ile Asp Glu
Cys Ala Gly Ser Pro Cys His675 680 685aac
ggg ggc act tgt gag gat ggc atc gcg ggc ttc act tgc cgc tgc 2112Asn
Gly Gly Thr Cys Glu Asp Gly Ile Ala Gly Phe Thr Cys Arg Cys690
695 700ccc gag ggc tac cat gac ccc acg tgc ctg tcc
gag gtc aac gag tgc 2160Pro Glu Gly Tyr His Asp Pro Thr Cys Leu Ser
Glu Val Asn Glu Cys705 710 715
720aac agt aac ccc tgc atc cac gga gct tgc cgg gat ggc ctc aat ggg
2208Asn Ser Asn Pro Cys Ile His Gly Ala Cys Arg Asp Gly Leu Asn Gly725
730 735tac aag tgt gac tgt gcc cct ggg tgg
agt gga aca aac tgt gac atc 2256Tyr Lys Cys Asp Cys Ala Pro Gly Trp
Ser Gly Thr Asn Cys Asp Ile740 745 750aac
aac aac gag tgt gag tcc aac cct tgt gtc aac ggt ggc acc tgc 2304Asn
Asn Asn Glu Cys Glu Ser Asn Pro Cys Val Asn Gly Gly Thr Cys755
760 765aag gac atg acc agt ggc tac gta tgc acc tgc
cga gaa ggc ttc agt 2352Lys Asp Met Thr Ser Gly Tyr Val Cys Thr Cys
Arg Glu Gly Phe Ser770 775 780ggc cct aat
tgc cag acc aac atc aac gaa tgt gcc tcc aac ccc tgc 2400Gly Pro Asn
Cys Gln Thr Asn Ile Asn Glu Cys Ala Ser Asn Pro Cys785
790 795 800ctg aac cag ggg acc tgc att
gat gat gtc gct gga tac aag tgc aac 2448Leu Asn Gln Gly Thr Cys Ile
Asp Asp Val Ala Gly Tyr Lys Cys Asn805 810
815tgt cct ctg cca tat aca gga gcc acg tgt gag gtg gtg ttg gcc cca
2496Cys Pro Leu Pro Tyr Thr Gly Ala Thr Cys Glu Val Val Leu Ala Pro820
825 830tgt gct acc agc ccc tgc aaa aac agc
ggg gta tgc aag gag tct gaa 2544Cys Ala Thr Ser Pro Cys Lys Asn Ser
Gly Val Cys Lys Glu Ser Glu835 840 845gac
tat gag agt ttt tcc tgt gtc tgt ccc aca ggc tgg caa ggt caa 2592Asp
Tyr Glu Ser Phe Ser Cys Val Cys Pro Thr Gly Trp Gln Gly Gln850
855 860acc tgc gag gtt gac atc aat gag tgt gtg aaa
agc cca tgt cgc cat 2640Thr Cys Glu Val Asp Ile Asn Glu Cys Val Lys
Ser Pro Cys Arg His865 870 875
880ggg gcc tcc tgc cag aac acc aat ggc agc tac cgc tgc ctc tgc cag
2688Gly Ala Ser Cys Gln Asn Thr Asn Gly Ser Tyr Arg Cys Leu Cys Gln885
890 895gcc ggc tat aca ggt cgc aac tgt gag
agt gac atc gat gac tgc cgc 2736Ala Gly Tyr Thr Gly Arg Asn Cys Glu
Ser Asp Ile Asp Asp Cys Arg900 905 910ccc
aac ccg tgt cac aat ggg ggt tcc tgc acc gat ggc atc aac aca 2784Pro
Asn Pro Cys His Asn Gly Gly Ser Cys Thr Asp Gly Ile Asn Thr915
920 925gcc ttc tgc gac tgc ctg ccc ggc ttc cag ggt
gcc ttc tgt gag gag 2832Ala Phe Cys Asp Cys Leu Pro Gly Phe Gln Gly
Ala Phe Cys Glu Glu930 935 940gac atc aat
gaa tgt gcc agc aat ccc tgc caa aat ggt gcc aat tgc 2880Asp Ile Asn
Glu Cys Ala Ser Asn Pro Cys Gln Asn Gly Ala Asn Cys945
950 955 960act gac tgt gtg gac agc tac
aca tgt acc tgc ccc gtg ggc ttc aat 2928Thr Asp Cys Val Asp Ser Tyr
Thr Cys Thr Cys Pro Val Gly Phe Asn965 970
975ggc atc cac tgc gag aac aac aca cct gac tgt act gag agc tcc tgc
2976Gly Ile His Cys Glu Asn Asn Thr Pro Asp Cys Thr Glu Ser Ser Cys980
985 990ttc aat ggt ggt acc tgt gtg gat ggt
atc aac tcc ttc acc tgt ctg 3024Phe Asn Gly Gly Thr Cys Val Asp Gly
Ile Asn Ser Phe Thr Cys Leu995 1000
1005tgt cca cct ggc ttc acg ggc agc tac tgt cag tat gat gtc aat
3069Cys Pro Pro Gly Phe Thr Gly Ser Tyr Cys Gln Tyr Asp Val Asn1010
1015 1020gag tgt gat tca cgg ccc tgt ctg
cac ggt ggt acc tgc caa gac 3114Glu Cys Asp Ser Arg Pro Cys Leu
His Gly Gly Thr Cys Gln Asp1025 1030
1035agc tat ggt act tat aag tgt acc tgc cca cag ggc tac act ggt
3159Ser Tyr Gly Thr Tyr Lys Cys Thr Cys Pro Gln Gly Tyr Thr Gly1040
1045 1050ctc aac tgc cag aac ctt gtg cgc
tgg tgc gac tcg gct ccc tgc 3204Leu Asn Cys Gln Asn Leu Val Arg
Trp Cys Asp Ser Ala Pro Cys1055 1060
1065aag aat ggt ggc agg tgc tgg cag acc aac acg cag tac cac tgt
3249Lys Asn Gly Gly Arg Cys Trp Gln Thr Asn Thr Gln Tyr His Cys1070
1075 1080gag tgc cgc agc ggc tgg act ggc
gtc aac tgc gac gtg ctc agt 3294Glu Cys Arg Ser Gly Trp Thr Gly
Val Asn Cys Asp Val Leu Ser1085 1090
1095gtg tcc tgt gag gtg gct gca cag aag cga ggc att gac gtc act
3339Val Ser Cys Glu Val Ala Ala Gln Lys Arg Gly Ile Asp Val Thr1100
1105 1110ctc ctg tgc cag cat gga ggg ctc
tgt gtg gat gag gga gat aaa 3384Leu Leu Cys Gln His Gly Gly Leu
Cys Val Asp Glu Gly Asp Lys1115 1120
1125cat tac tgc cac tgc cag gca ggc tac acg ggc agc tac tgt gag
3429His Tyr Cys His Cys Gln Ala Gly Tyr Thr Gly Ser Tyr Cys Glu1130
1135 1140gac gag gtg gac gag tgc tca cct
aac ccc tgc cag aat gga gct 3474Asp Glu Val Asp Glu Cys Ser Pro
Asn Pro Cys Gln Asn Gly Ala1145 1150
1155acc tgc act gac tat ctc ggc ggc ttt tcc tgc aag tgt gtg gct
3519Thr Cys Thr Asp Tyr Leu Gly Gly Phe Ser Cys Lys Cys Val Ala1160
1165 1170ggc tac cat ggg tct aac tgc tca
gag gag atc aac gag tgc ctg 3564Gly Tyr His Gly Ser Asn Cys Ser
Glu Glu Ile Asn Glu Cys Leu1175 1180
1185tcc cag ccc tgc cag aat ggg ggt acc tgc att gat ctg acc aac
3609Ser Gln Pro Cys Gln Asn Gly Gly Thr Cys Ile Asp Leu Thr Asn1190
1195 1200tcc tac aag tgt tcc tgc ccc cgg
ggg aca cag ggt gta cac tgt 3654Ser Tyr Lys Cys Ser Cys Pro Arg
Gly Thr Gln Gly Val His Cys1205 1210
1215gag atc aat gtt gat gac tgc cat ccc ccc ctt gac cct gcc tcc
3699Glu Ile Asn Val Asp Asp Cys His Pro Pro Leu Asp Pro Ala Ser1220
1225 1230cga agc ccc aag tgc ttc aac aat
ggc acc tgt gtg gac cag gtg 3744Arg Ser Pro Lys Cys Phe Asn Asn
Gly Thr Cys Val Asp Gln Val1235 1240
1245ggt ggc tat acc tgc acc tgc cca cca ggc ttc gtc ggg gag cgg
3789Gly Gly Tyr Thr Cys Thr Cys Pro Pro Gly Phe Val Gly Glu Arg1250
1255 1260tgt gag ggt gat gtc aat gaa tgt
ctc tcc aac ccc tgt gac cca 3834Cys Glu Gly Asp Val Asn Glu Cys
Leu Ser Asn Pro Cys Asp Pro1265 1270
1275cgt ggc acc cag aac tgt gtg cag cgt gtt aat gac ttc cac tgc
3879Arg Gly Thr Gln Asn Cys Val Gln Arg Val Asn Asp Phe His Cys1280
1285 1290gag tgc cgg gct ggc cac act gga
cgc cgc tgt gag tca gtc atc 3924Glu Cys Arg Ala Gly His Thr Gly
Arg Arg Cys Glu Ser Val Ile1295 1300
1305aat ggc tgc agg ggc aaa cct tgc aag aat ggg ggt gtc tgt gcc
3969Asn Gly Cys Arg Gly Lys Pro Cys Lys Asn Gly Gly Val Cys Ala1310
1315 1320gtg gcc tcc aac acc gcc cgt gga
ttc atc tgt agg tgc cct gcg 4014Val Ala Ser Asn Thr Ala Arg Gly
Phe Ile Cys Arg Cys Pro Ala1325 1330
1335ggc ttc gag ggt gcc aca tgt gag aat gat gcc cgc act tgt ggc
4059Gly Phe Glu Gly Ala Thr Cys Glu Asn Asp Ala Arg Thr Cys Gly1340
1345 1350agc tta cgc tgc ctc aac ggt ggt
aca tgc atc tcg ggc cca cgt 4104Ser Leu Arg Cys Leu Asn Gly Gly
Thr Cys Ile Ser Gly Pro Arg1355 1360
1365agt ccc acc tgc cta tgc ctg gga tcc ttc acc ggc cct gag tgc
4149Ser Pro Thr Cys Leu Cys Leu Gly Ser Phe Thr Gly Pro Glu Cys1370
1375 1380cag ttc cca gcc agc agc ccc tgt
gtg ggt agc aac ccc tgc tac 4194Gln Phe Pro Ala Ser Ser Pro Cys
Val Gly Ser Asn Pro Cys Tyr1385 1390
1395aat cag ggc acc tgt gag ccc aca tcc gag aac cct ttc tac cgc
4239Asn Gln Gly Thr Cys Glu Pro Thr Ser Glu Asn Pro Phe Tyr Arg1400
1405 1410tgt cta tgc cct gcc aaa ttc aac
ggg cta ctg tgc cac atc ctg 4284Cys Leu Cys Pro Ala Lys Phe Asn
Gly Leu Leu Cys His Ile Leu1415 1420
1425gac tac agc ttc aca ggt ggc gct ggg cgc gac att ccc cca ccg
4329Asp Tyr Ser Phe Thr Gly Gly Ala Gly Arg Asp Ile Pro Pro Pro1430
1435 1440cag att gag gag gcc tgt gag ctg
cct gag tgc cag gtg gat gca 4374Gln Ile Glu Glu Ala Cys Glu Leu
Pro Glu Cys Gln Val Asp Ala1445 1450
1455ggc aat aag gtc tgc aac ctg cag tgt aat aat cac gca tgt ggc
4419Gly Asn Lys Val Cys Asn Leu Gln Cys Asn Asn His Ala Cys Gly1460
1465 1470tgg gat ggt ggc gac tgc tcc ctc
aac ttc aat gac ccc tgg aag 4464Trp Asp Gly Gly Asp Cys Ser Leu
Asn Phe Asn Asp Pro Trp Lys1475 1480
1485aac tgc acg cag tct cta cag tgc tgg aag tat ttt agc gac ggc
4509Asn Cys Thr Gln Ser Leu Gln Cys Trp Lys Tyr Phe Ser Asp Gly1490
1495 1500cac tgt gac agc cag tgc aac tcg
gcc ggc tgc ctc ttt gat ggc 4554His Cys Asp Ser Gln Cys Asn Ser
Ala Gly Cys Leu Phe Asp Gly1505 1510
1515ttc gac tgc cag ctc acc gag gga cag tgc aac ccc ctg tat gac
4599Phe Asp Cys Gln Leu Thr Glu Gly Gln Cys Asn Pro Leu Tyr Asp1520
1525 1530cag tac tgc aag gac cac ttc agt
gat ggc cac tgc gac cag ggc 4644Gln Tyr Cys Lys Asp His Phe Ser
Asp Gly His Cys Asp Gln Gly1535 1540
1545tgt aac agt gcc gaa tgt gag tgg gat ggc cta gac tgt gct gag
4689Cys Asn Ser Ala Glu Cys Glu Trp Asp Gly Leu Asp Cys Ala Glu1550
1555 1560cat gta ccc gag cgg ctg gca gcc
ggc acc ctg gtg ctg gtg gtg 4734His Val Pro Glu Arg Leu Ala Ala
Gly Thr Leu Val Leu Val Val1565 1570
1575ctg ctt cca ccc gac cag cta cgg aac aac tcc ttc cac ttt ctg
4779Leu Leu Pro Pro Asp Gln Leu Arg Asn Asn Ser Phe His Phe Leu1580
1585 1590cgg gag ctc agc cac gtg ctg cac
acc aac gtg gtc ttc aag cgt 4824Arg Glu Leu Ser His Val Leu His
Thr Asn Val Val Phe Lys Arg1595 1600
1605gat gcg caa ggc cag cag atg atc ttc ccg tac tat ggc cac gag
4869Asp Ala Gln Gly Gln Gln Met Ile Phe Pro Tyr Tyr Gly His Glu1610
1615 1620gaa gag ctg cgc aag cac cca atc
aag cgc tct aca gtg ggt tgg 4914Glu Glu Leu Arg Lys His Pro Ile
Lys Arg Ser Thr Val Gly Trp1625 1630
1635gcc acc tct tca ctg ctt cct ggt acc agt ggt ggg cgc cag cgc
4959Ala Thr Ser Ser Leu Leu Pro Gly Thr Ser Gly Gly Arg Gln Arg1640
1645 1650agg gag ctg gac ccc atg gac atc
cgt ggc tcc att gtc tac ctg 5004Arg Glu Leu Asp Pro Met Asp Ile
Arg Gly Ser Ile Val Tyr Leu1655 1660
1665gag atc gac aac cgg caa tgt gtg cag tca tcc tcg cag tgc ttc
5049Glu Ile Asp Asn Arg Gln Cys Val Gln Ser Ser Ser Gln Cys Phe1670
1675 1680cag agt gcc acc gat gtg gct gcc
ttc cta ggt gct ctt gcg tca 5094Gln Ser Ala Thr Asp Val Ala Ala
Phe Leu Gly Ala Leu Ala Ser1685 1690
1695ctt ggc agc ctc aat att cct tac aag att gag gcc gtg aag agt
5139Leu Gly Ser Leu Asn Ile Pro Tyr Lys Ile Glu Ala Val Lys Ser1700
1705 1710gag ccg gtg gag cct ccg ctg ccc
tcg cag ctg cac ctc atg tac 5184Glu Pro Val Glu Pro Pro Leu Pro
Ser Gln Leu His Leu Met Tyr1715 1720
1725gtg gca gcg gcc gcc ttc gtg ctc ctg ttc ttt gtg ggc tgt ggg
5229Val Ala Ala Ala Ala Phe Val Leu Leu Phe Phe Val Gly Cys Gly1730
1735 1740gtg ctg ctg tcc cgc aag cgc cgg
cgg cag cat ggc cag ctc tgg 5274Val Leu Leu Ser Arg Lys Arg Arg
Arg Gln His Gly Gln Leu Trp1745 1750
1755ttc cct gag ggt ttc aaa gtg tca gag gcc agc aag aag aag cgg
5319Phe Pro Glu Gly Phe Lys Val Ser Glu Ala Ser Lys Lys Lys Arg1760
1765 1770aga gag ccc ctc ggc gag gac tca
gtc ggc ctc aag ccc ctg aag 5364Arg Glu Pro Leu Gly Glu Asp Ser
Val Gly Leu Lys Pro Leu Lys1775 1780
1785aat gcc tca gat ggt gct ctg atg gac gac aat cag aac gag tgg
5409Asn Ala Ser Asp Gly Ala Leu Met Asp Asp Asn Gln Asn Glu Trp1790
1795 1800gga gac gaa gac ctg gag acc aag
aag ttc cgg ttt gag gag cca 5454Gly Asp Glu Asp Leu Glu Thr Lys
Lys Phe Arg Phe Glu Glu Pro1805 1810
1815gta gtt ctc cct gac ctg agt gat cag act gac cac aga cag tgg
5499Val Val Leu Pro Asp Leu Ser Asp Gln Thr Asp His Arg Gln Trp1820
1825 1830acc cag cag cac ctg gac gct gct
gac ctg cgc atg tct gcc atg 5544Thr Gln Gln His Leu Asp Ala Ala
Asp Leu Arg Met Ser Ala Met1835 1840
1845gcc cca aca ccg cct cag ggg gag gtg gat gct gac tgc atg gat
5589Ala Pro Thr Pro Pro Gln Gly Glu Val Asp Ala Asp Cys Met Asp1850
1855 1860gtc aat gtt cga gga cca gat ggc
ttc aca ccc ctc atg att gcc 5634Val Asn Val Arg Gly Pro Asp Gly
Phe Thr Pro Leu Met Ile Ala1865 1870
1875tcc tgc agt gga ggg ggc ctt gag aca ggc aac agt gaa gaa gaa
5679Ser Cys Ser Gly Gly Gly Leu Glu Thr Gly Asn Ser Glu Glu Glu1880
1885 1890gaa gat gca cct gct gtc atc tct
gac ttc atc tac cag ggc gcc 5724Glu Asp Ala Pro Ala Val Ile Ser
Asp Phe Ile Tyr Gln Gly Ala1895 1900
1905agc ttg cac aac cag aca gac cgc acc ggg gag acc gcc ttg cac
5769Ser Leu His Asn Gln Thr Asp Arg Thr Gly Glu Thr Ala Leu His1910
1915 1920ttg gct gcc cga tac tct cgt tca
gat gct gca aag cgc ttg ctg 5814Leu Ala Ala Arg Tyr Ser Arg Ser
Asp Ala Ala Lys Arg Leu Leu1925 1930
1935gag gcc agt gca gat gcc aac atc cag gac aac atg ggc cgt act
5859Glu Ala Ser Ala Asp Ala Asn Ile Gln Asp Asn Met Gly Arg Thr1940
1945 1950ccg tta cat gca gca gtt tct gca
gat gct cag ggt gtc ttc cag 5904Pro Leu His Ala Ala Val Ser Ala
Asp Ala Gln Gly Val Phe Gln1955 1960
1965atc ctg ctc cgg aac agg gcc aca gat ctg gat gcc cga atg cat
5949Ile Leu Leu Arg Asn Arg Ala Thr Asp Leu Asp Ala Arg Met His1970
1975 1980gat ggc aca act cca ctg atc ctg
gct gcg cgc ctg gcc ctg gag 5994Asp Gly Thr Thr Pro Leu Ile Leu
Ala Ala Arg Leu Ala Leu Glu1985 1990
1995ggc atg ctg gag gac ctc atc aac tca cat gct gac gtc aat gcc
6039Gly Met Leu Glu Asp Leu Ile Asn Ser His Ala Asp Val Asn Ala2000
2005 2010gtg gat gac cta ggc aag tcg gct
ttg cat tgg gcg gcc gcg gtg 6084Val Asp Asp Leu Gly Lys Ser Ala
Leu His Trp Ala Ala Ala Val2015 2020
2025aac aat gtg gat gct gct gtt gtg ctc ctg aag aac gga gcc aac
6129Asn Asn Val Asp Ala Ala Val Val Leu Leu Lys Asn Gly Ala Asn2030
2035 2040aag gac atg cag aac aac aag gag
gag act ccc ctg ttc ctg gcc 6174Lys Asp Met Gln Asn Asn Lys Glu
Glu Thr Pro Leu Phe Leu Ala2045 2050
2055gcc cgt gag ggc agc tat gag act gcc aaa gtg ttg ctg gac cac
6219Ala Arg Glu Gly Ser Tyr Glu Thr Ala Lys Val Leu Leu Asp His2060
2065 2070ttt gcc aac cgg gac atc acg gat
cac atg gac cga ttg ccg cgg 6264Phe Ala Asn Arg Asp Ile Thr Asp
His Met Asp Arg Leu Pro Arg2075 2080
2085gac atc gca cag gag cgt atg cac cac gat atc gtg cgg ctt ttg
6309Asp Ile Ala Gln Glu Arg Met His His Asp Ile Val Arg Leu Leu2090
2095 2100gat gag tac aac ctg gtg cgc agc
cca cag ctg cat ggc act gcc 6354Asp Glu Tyr Asn Leu Val Arg Ser
Pro Gln Leu His Gly Thr Ala2105 2110
2115ctg ggt ggc aca ccc act ctg tct ccc aca ctc tgc tcg ccc aat
6399Leu Gly Gly Thr Pro Thr Leu Ser Pro Thr Leu Cys Ser Pro Asn2120
2125 2130ggc tac ctg ggc aat ctc aag tct
gcc aca cag ggc aag aag gcc 6444Gly Tyr Leu Gly Asn Leu Lys Ser
Ala Thr Gln Gly Lys Lys Ala2135 2140
2145cgc aag ccc agc acc aaa ggg ctg gct tgt ggt agc aag gaa gct
6489Arg Lys Pro Ser Thr Lys Gly Leu Ala Cys Gly Ser Lys Glu Ala2150
2155 2160aag gac ctc aag gca cgg agg aag
aag tcc cag gat ggc aag ggc 6534Lys Asp Leu Lys Ala Arg Arg Lys
Lys Ser Gln Asp Gly Lys Gly2165 2170
2175tgc ctg ttg gac agc tcg agc atg ctg tcg cct gtg gac tcc ctc
6579Cys Leu Leu Asp Ser Ser Ser Met Leu Ser Pro Val Asp Ser Leu2180
2185 2190gag tca ccc cat ggc tac ttg tca
gat gtg gcc tcg cca ccc ctc 6624Glu Ser Pro His Gly Tyr Leu Ser
Asp Val Ala Ser Pro Pro Leu2195 2200
2205ctc ccc tcc cca ttc cag cag tct cca tcc atg cct ctc agc cac
6669Leu Pro Ser Pro Phe Gln Gln Ser Pro Ser Met Pro Leu Ser His2210
2215 2220ctg cct ggt atg cct gac act cac
ctg ggc atc agc cac ttg aat 6714Leu Pro Gly Met Pro Asp Thr His
Leu Gly Ile Ser His Leu Asn2225 2230
2235gtg gca gcc aag cct gag atg gca gca ctg gct gga ggt agc cgg
6759Val Ala Ala Lys Pro Glu Met Ala Ala Leu Ala Gly Gly Ser Arg2240
2245 2250ttg gcc ttt gag cca ccc ccg cca
cgc ctc tcc cac ctg cct gta 6804Leu Ala Phe Glu Pro Pro Pro Pro
Arg Leu Ser His Leu Pro Val2255 2260
2265gcc tcc agt gcc agc aca gtg ctg agt acc aat ggc acg ggg gct
6849Ala Ser Ser Ala Ser Thr Val Leu Ser Thr Asn Gly Thr Gly Ala2270
2275 2280atg aat ttc acc gtg ggt gca ccg
gca agc ttg aat ggc cag tgt 6894Met Asn Phe Thr Val Gly Ala Pro
Ala Ser Leu Asn Gly Gln Cys2285 2290
2295gag tgg ctt ccc cgg ctc cag aat ggc atg gtg ccc agc cag tac
6939Glu Trp Leu Pro Arg Leu Gln Asn Gly Met Val Pro Ser Gln Tyr2300
2305 2310aac cca cta cgg ccg ggt gtg acg
ccg ggc aca ctg agc aca cag 6984Asn Pro Leu Arg Pro Gly Val Thr
Pro Gly Thr Leu Ser Thr Gln2315 2320
2325gca gct ggc ctc cag cat agc atg atg ggg cca cta cac agc agc
7029Ala Ala Gly Leu Gln His Ser Met Met Gly Pro Leu His Ser Ser2330
2335 2340ctc tcc acc aat acc ttg tcc ccg
att att tac cag ggc ctg ccc 7074Leu Ser Thr Asn Thr Leu Ser Pro
Ile Ile Tyr Gln Gly Leu Pro2345 2350
2355aac aca cgg ctg gca aca cag cct cac ctg gtg cag acc cag cag
7119Asn Thr Arg Leu Ala Thr Gln Pro His Leu Val Gln Thr Gln Gln2360
2365 2370gtg cag cca cag aac tta cag ctc
cag cct cag aac ctg cag cca 7164Val Gln Pro Gln Asn Leu Gln Leu
Gln Pro Gln Asn Leu Gln Pro2375 2380
2385cca tca cag cca cac ctc agt gtg agc tcg gca gcc aat ggg cac
7209Pro Ser Gln Pro His Leu Ser Val Ser Ser Ala Ala Asn Gly His2390
2395 2400ctg ggc cgg agc ttc ttg agt ggg
gag ccc agt cag gca gat gta 7254Leu Gly Arg Ser Phe Leu Ser Gly
Glu Pro Ser Gln Ala Asp Val2405 2410
2415caa ccg ctg ggc ccc agc agt ctg cct gtg cac acc att ctg ccc
7299Gln Pro Leu Gly Pro Ser Ser Leu Pro Val His Thr Ile Leu Pro2420
2425 2430cag gaa agc cag gcc ctg ccc aca
tca ctg cca tcc tcc atg gtc 7344Gln Glu Ser Gln Ala Leu Pro Thr
Ser Leu Pro Ser Ser Met Val2435 2440
2445cca ccc atg acc act acc cag ttc ctg acc cct cct tcc cag cac
7389Pro Pro Met Thr Thr Thr Gln Phe Leu Thr Pro Pro Ser Gln His2450
2455 2460agt tac tcc tcc tcc cct gtg gac
aac acc ccc agc cac cag ctg 7434Ser Tyr Ser Ser Ser Pro Val Asp
Asn Thr Pro Ser His Gln Leu2465 2470
2475cag gtg cca gag cac ccc ttc ctc acc cca tcc cct gag tcc cct
7479Gln Val Pro Glu His Pro Phe Leu Thr Pro Ser Pro Glu Ser Pro2480
2485 2490gac cag tgg tcc agc tcc tcc ccg
cat tcc aac atc tct gat tgg 7524Asp Gln Trp Ser Ser Ser Ser Pro
His Ser Asn Ile Ser Asp Trp2495 2500
2505tcc gag ggc atc tcc agc ccg ccc acc acc atg ccg tcc cag atc
7569Ser Glu Gly Ile Ser Ser Pro Pro Thr Thr Met Pro Ser Gln Ile2510
2515 2520acc cac att cca gag gca ttt aaa
taa 7596Thr His Ile Pro Glu Ala Phe
Lys2525 253022531PRTMus musculus 2Met Pro Arg Leu Leu Thr
Pro Leu Leu Cys Leu Thr Leu Leu Pro Ala1 5
10 15Leu Ala Ala Arg Gly Leu Arg Cys Ser Gln Pro Ser
Gly Thr Cys Leu20 25 30Asn Gly Gly Arg
Cys Glu Val Ala Ser Gly Thr Glu Ala Cys Val Cys35 40
45Ser Gly Ala Phe Val Gly Gln Arg Cys Gln Asp Ser Asn Pro
Cys Leu50 55 60Ser Thr Pro Cys Lys Asn
Ala Gly Thr Cys His Val Val Asp His Gly65 70
75 80Gly Thr Val Asp Tyr Ala Cys Ser Cys Pro Leu
Gly Phe Ser Gly Pro85 90 95Leu Cys Leu
Thr Pro Leu Asp Asn Ala Cys Leu Ala Asn Pro Cys Arg100
105 110Asn Gly Gly Thr Cys Asp Leu Leu Thr Leu Thr Glu
Tyr Lys Cys Arg115 120 125Cys Pro Pro Gly
Trp Ser Gly Lys Ser Cys Gln Gln Ala Asp Pro Cys130 135
140Ala Ser Asn Pro Cys Ala Asn Gly Gly Gln Cys Leu Pro Phe
Glu Ser145 150 155 160Ser
Tyr Ile Cys Arg Cys Pro Pro Gly Phe His Gly Pro Thr Cys Arg165
170 175Gln Asp Val Asn Glu Cys Ser Gln Asn Pro Gly
Leu Cys Arg His Gly180 185 190Gly Thr Cys
His Asn Glu Ile Gly Ser Tyr Arg Cys Ala Cys Arg Ala195
200 205Thr His Thr Gly Pro His Cys Glu Leu Pro Tyr Val
Pro Cys Ser Pro210 215 220Ser Pro Cys Gln
Asn Gly Gly Thr Cys Arg Pro Thr Gly Asp Thr Thr225 230
235 240His Glu Cys Ala Cys Leu Pro Gly Phe
Ala Gly Gln Asn Cys Glu Glu245 250 255Asn
Val Asp Asp Cys Pro Gly Asn Asn Cys Lys Asn Gly Gly Ala Cys260
265 270Val Asp Gly Val Asn Thr Tyr Asn Cys Arg Cys
Pro Pro Glu Trp Thr275 280 285Gly Gln Tyr
Cys Thr Glu Asp Val Asp Glu Cys Gln Leu Met Pro Asn290
295 300Ala Cys Gln Asn Gly Gly Thr Cys His Asn Thr His
Gly Gly Tyr Asn305 310 315
320Cys Val Cys Val Asn Gly Trp Thr Gly Glu Asp Cys Ser Glu Asn Ile325
330 335Asp Asp Cys Ala Ser Ala Ala Cys Phe
Gln Gly Ala Thr Cys His Asp340 345 350Arg
Val Ala Ser Phe Tyr Cys Glu Cys Pro His Gly Arg Thr Gly Leu355
360 365Leu Cys His Leu Asn Asp Ala Cys Ile Ser Asn
Pro Cys Asn Glu Gly370 375 380Ser Asn Cys
Asp Thr Asn Pro Val Asn Gly Lys Ala Ile Cys Thr Cys385
390 395 400Pro Ser Gly Tyr Thr Gly Pro
Ala Cys Ser Gln Asp Val Asp Glu Cys405 410
415Ala Leu Gly Ala Asn Pro Cys Glu His Ala Gly Lys Cys Leu Asn Thr420
425 430Leu Gly Ser Phe Glu Cys Gln Cys Leu
Gln Gly Tyr Thr Gly Pro Arg435 440 445Cys
Glu Ile Asp Val Asn Glu Cys Ile Ser Asn Pro Cys Gln Asn Asp450
455 460Ala Thr Cys Leu Asp Gln Ile Gly Glu Phe Gln
Cys Ile Cys Met Pro465 470 475
480Gly Tyr Glu Gly Val Tyr Cys Glu Ile Asn Thr Asp Glu Cys Ala
Ser485 490 495Ser Pro Cys Leu His Asn Gly
His Cys Met Asp Lys Ile Asn Glu Phe500 505
510Gln Cys Gln Cys Pro Lys Gly Phe Asn Gly His Leu Cys Gln Tyr Asp515
520 525Val Asp Glu Cys Ala Ser Thr Pro Cys
Lys Asn Gly Ala Lys Cys Leu530 535 540Asp
Gly Pro Asn Thr Tyr Thr Cys Val Cys Thr Glu Gly Tyr Thr Gly545
550 555 560Thr His Cys Glu Val Asp
Ile Asp Glu Cys Asp Pro Asp Pro Cys His565 570
575Tyr Gly Ser Cys Lys Asp Gly Val Ala Thr Phe Thr Cys Leu Cys
Gln580 585 590Pro Gly Tyr Thr Gly His His
Cys Glu Thr Asn Ile Asn Glu Cys His595 600
605Ser Gln Pro Cys Arg His Gly Gly Thr Cys Gln Asp Arg Asp Asn Ser610
615 620Tyr Leu Cys Leu Cys Leu Lys Gly Thr
Thr Gly Pro Asn Cys Glu Ile625 630 635
640Asn Leu Asp Asp Cys Ala Ser Asn Pro Cys Asp Ser Gly Thr
Cys Leu645 650 655Asp Lys Ile Asp Gly Tyr
Glu Cys Ala Cys Glu Pro Gly Tyr Thr Gly660 665
670Ser Met Cys Asn Val Asn Ile Asp Glu Cys Ala Gly Ser Pro Cys
His675 680 685Asn Gly Gly Thr Cys Glu Asp
Gly Ile Ala Gly Phe Thr Cys Arg Cys690 695
700Pro Glu Gly Tyr His Asp Pro Thr Cys Leu Ser Glu Val Asn Glu Cys705
710 715 720Asn Ser Asn Pro
Cys Ile His Gly Ala Cys Arg Asp Gly Leu Asn Gly725 730
735Tyr Lys Cys Asp Cys Ala Pro Gly Trp Ser Gly Thr Asn Cys
Asp Ile740 745 750Asn Asn Asn Glu Cys Glu
Ser Asn Pro Cys Val Asn Gly Gly Thr Cys755 760
765Lys Asp Met Thr Ser Gly Tyr Val Cys Thr Cys Arg Glu Gly Phe
Ser770 775 780Gly Pro Asn Cys Gln Thr Asn
Ile Asn Glu Cys Ala Ser Asn Pro Cys785 790
795 800Leu Asn Gln Gly Thr Cys Ile Asp Asp Val Ala Gly
Tyr Lys Cys Asn805 810 815Cys Pro Leu Pro
Tyr Thr Gly Ala Thr Cys Glu Val Val Leu Ala Pro820 825
830Cys Ala Thr Ser Pro Cys Lys Asn Ser Gly Val Cys Lys Glu
Ser Glu835 840 845Asp Tyr Glu Ser Phe Ser
Cys Val Cys Pro Thr Gly Trp Gln Gly Gln850 855
860Thr Cys Glu Val Asp Ile Asn Glu Cys Val Lys Ser Pro Cys Arg
His865 870 875 880Gly Ala
Ser Cys Gln Asn Thr Asn Gly Ser Tyr Arg Cys Leu Cys Gln885
890 895Ala Gly Tyr Thr Gly Arg Asn Cys Glu Ser Asp Ile
Asp Asp Cys Arg900 905 910Pro Asn Pro Cys
His Asn Gly Gly Ser Cys Thr Asp Gly Ile Asn Thr915 920
925Ala Phe Cys Asp Cys Leu Pro Gly Phe Gln Gly Ala Phe Cys
Glu Glu930 935 940Asp Ile Asn Glu Cys Ala
Ser Asn Pro Cys Gln Asn Gly Ala Asn Cys945 950
955 960Thr Asp Cys Val Asp Ser Tyr Thr Cys Thr Cys
Pro Val Gly Phe Asn965 970 975Gly Ile His
Cys Glu Asn Asn Thr Pro Asp Cys Thr Glu Ser Ser Cys980
985 990Phe Asn Gly Gly Thr Cys Val Asp Gly Ile Asn Ser
Phe Thr Cys Leu995 1000 1005Cys Pro Pro
Gly Phe Thr Gly Ser Tyr Cys Gln Tyr Asp Val Asn1010
1015 1020Glu Cys Asp Ser Arg Pro Cys Leu His Gly Gly
Thr Cys Gln Asp1025 1030 1035Ser Tyr
Gly Thr Tyr Lys Cys Thr Cys Pro Gln Gly Tyr Thr Gly1040
1045 1050Leu Asn Cys Gln Asn Leu Val Arg Trp Cys Asp
Ser Ala Pro Cys1055 1060 1065Lys Asn
Gly Gly Arg Cys Trp Gln Thr Asn Thr Gln Tyr His Cys1070
1075 1080Glu Cys Arg Ser Gly Trp Thr Gly Val Asn Cys
Asp Val Leu Ser1085 1090 1095Val Ser
Cys Glu Val Ala Ala Gln Lys Arg Gly Ile Asp Val Thr1100
1105 1110Leu Leu Cys Gln His Gly Gly Leu Cys Val Asp
Glu Gly Asp Lys1115 1120 1125His Tyr
Cys His Cys Gln Ala Gly Tyr Thr Gly Ser Tyr Cys Glu1130
1135 1140Asp Glu Val Asp Glu Cys Ser Pro Asn Pro Cys
Gln Asn Gly Ala1145 1150 1155Thr Cys
Thr Asp Tyr Leu Gly Gly Phe Ser Cys Lys Cys Val Ala1160
1165 1170Gly Tyr His Gly Ser Asn Cys Ser Glu Glu Ile
Asn Glu Cys Leu1175 1180 1185Ser Gln
Pro Cys Gln Asn Gly Gly Thr Cys Ile Asp Leu Thr Asn1190
1195 1200Ser Tyr Lys Cys Ser Cys Pro Arg Gly Thr Gln
Gly Val His Cys1205 1210 1215Glu Ile
Asn Val Asp Asp Cys His Pro Pro Leu Asp Pro Ala Ser1220
1225 1230Arg Ser Pro Lys Cys Phe Asn Asn Gly Thr Cys
Val Asp Gln Val1235 1240 1245Gly Gly
Tyr Thr Cys Thr Cys Pro Pro Gly Phe Val Gly Glu Arg1250
1255 1260Cys Glu Gly Asp Val Asn Glu Cys Leu Ser Asn
Pro Cys Asp Pro1265 1270 1275Arg Gly
Thr Gln Asn Cys Val Gln Arg Val Asn Asp Phe His Cys1280
1285 1290Glu Cys Arg Ala Gly His Thr Gly Arg Arg Cys
Glu Ser Val Ile1295 1300 1305Asn Gly
Cys Arg Gly Lys Pro Cys Lys Asn Gly Gly Val Cys Ala1310
1315 1320Val Ala Ser Asn Thr Ala Arg Gly Phe Ile Cys
Arg Cys Pro Ala1325 1330 1335Gly Phe
Glu Gly Ala Thr Cys Glu Asn Asp Ala Arg Thr Cys Gly1340
1345 1350Ser Leu Arg Cys Leu Asn Gly Gly Thr Cys Ile
Ser Gly Pro Arg1355 1360 1365Ser Pro
Thr Cys Leu Cys Leu Gly Ser Phe Thr Gly Pro Glu Cys1370
1375 1380Gln Phe Pro Ala Ser Ser Pro Cys Val Gly Ser
Asn Pro Cys Tyr1385 1390 1395Asn Gln
Gly Thr Cys Glu Pro Thr Ser Glu Asn Pro Phe Tyr Arg1400
1405 1410Cys Leu Cys Pro Ala Lys Phe Asn Gly Leu Leu
Cys His Ile Leu1415 1420 1425Asp Tyr
Ser Phe Thr Gly Gly Ala Gly Arg Asp Ile Pro Pro Pro1430
1435 1440Gln Ile Glu Glu Ala Cys Glu Leu Pro Glu Cys
Gln Val Asp Ala1445 1450 1455Gly Asn
Lys Val Cys Asn Leu Gln Cys Asn Asn His Ala Cys Gly1460
1465 1470Trp Asp Gly Gly Asp Cys Ser Leu Asn Phe Asn
Asp Pro Trp Lys1475 1480 1485Asn Cys
Thr Gln Ser Leu Gln Cys Trp Lys Tyr Phe Ser Asp Gly1490
1495 1500His Cys Asp Ser Gln Cys Asn Ser Ala Gly Cys
Leu Phe Asp Gly1505 1510 1515Phe Asp
Cys Gln Leu Thr Glu Gly Gln Cys Asn Pro Leu Tyr Asp1520
1525 1530Gln Tyr Cys Lys Asp His Phe Ser Asp Gly His
Cys Asp Gln Gly1535 1540 1545Cys Asn
Ser Ala Glu Cys Glu Trp Asp Gly Leu Asp Cys Ala Glu1550
1555 1560His Val Pro Glu Arg Leu Ala Ala Gly Thr Leu
Val Leu Val Val1565 1570 1575Leu Leu
Pro Pro Asp Gln Leu Arg Asn Asn Ser Phe His Phe Leu1580
1585 1590Arg Glu Leu Ser His Val Leu His Thr Asn Val
Val Phe Lys Arg1595 1600 1605Asp Ala
Gln Gly Gln Gln Met Ile Phe Pro Tyr Tyr Gly His Glu1610
1615 1620Glu Glu Leu Arg Lys His Pro Ile Lys Arg Ser
Thr Val Gly Trp1625 1630 1635Ala Thr
Ser Ser Leu Leu Pro Gly Thr Ser Gly Gly Arg Gln Arg1640
1645 1650Arg Glu Leu Asp Pro Met Asp Ile Arg Gly Ser
Ile Val Tyr Leu1655 1660 1665Glu Ile
Asp Asn Arg Gln Cys Val Gln Ser Ser Ser Gln Cys Phe1670
1675 1680Gln Ser Ala Thr Asp Val Ala Ala Phe Leu Gly
Ala Leu Ala Ser1685 1690 1695Leu Gly
Ser Leu Asn Ile Pro Tyr Lys Ile Glu Ala Val Lys Ser1700
1705 1710Glu Pro Val Glu Pro Pro Leu Pro Ser Gln Leu
His Leu Met Tyr1715 1720 1725Val Ala
Ala Ala Ala Phe Val Leu Leu Phe Phe Val Gly Cys Gly1730
1735 1740Val Leu Leu Ser Arg Lys Arg Arg Arg Gln His
Gly Gln Leu Trp1745 1750 1755Phe Pro
Glu Gly Phe Lys Val Ser Glu Ala Ser Lys Lys Lys Arg1760
1765 1770Arg Glu Pro Leu Gly Glu Asp Ser Val Gly Leu
Lys Pro Leu Lys1775 1780 1785Asn Ala
Ser Asp Gly Ala Leu Met Asp Asp Asn Gln Asn Glu Trp1790
1795 1800Gly Asp Glu Asp Leu Glu Thr Lys Lys Phe Arg
Phe Glu Glu Pro1805 1810 1815Val Val
Leu Pro Asp Leu Ser Asp Gln Thr Asp His Arg Gln Trp1820
1825 1830Thr Gln Gln His Leu Asp Ala Ala Asp Leu Arg
Met Ser Ala Met1835 1840 1845Ala Pro
Thr Pro Pro Gln Gly Glu Val Asp Ala Asp Cys Met Asp1850
1855 1860Val Asn Val Arg Gly Pro Asp Gly Phe Thr Pro
Leu Met Ile Ala1865 1870 1875Ser Cys
Ser Gly Gly Gly Leu Glu Thr Gly Asn Ser Glu Glu Glu1880
1885 1890Glu Asp Ala Pro Ala Val Ile Ser Asp Phe Ile
Tyr Gln Gly Ala1895 1900 1905Ser Leu
His Asn Gln Thr Asp Arg Thr Gly Glu Thr Ala Leu His1910
1915 1920Leu Ala Ala Arg Tyr Ser Arg Ser Asp Ala Ala
Lys Arg Leu Leu1925 1930 1935Glu Ala
Ser Ala Asp Ala Asn Ile Gln Asp Asn Met Gly Arg Thr1940
1945 1950Pro Leu His Ala Ala Val Ser Ala Asp Ala Gln
Gly Val Phe Gln1955 1960 1965Ile Leu
Leu Arg Asn Arg Ala Thr Asp Leu Asp Ala Arg Met His1970
1975 1980Asp Gly Thr Thr Pro Leu Ile Leu Ala Ala Arg
Leu Ala Leu Glu1985 1990 1995Gly Met
Leu Glu Asp Leu Ile Asn Ser His Ala Asp Val Asn Ala2000
2005 2010Val Asp Asp Leu Gly Lys Ser Ala Leu His Trp
Ala Ala Ala Val2015 2020 2025Asn Asn
Val Asp Ala Ala Val Val Leu Leu Lys Asn Gly Ala Asn2030
2035 2040Lys Asp Met Gln Asn Asn Lys Glu Glu Thr Pro
Leu Phe Leu Ala2045 2050 2055Ala Arg
Glu Gly Ser Tyr Glu Thr Ala Lys Val Leu Leu Asp His2060
2065 2070Phe Ala Asn Arg Asp Ile Thr Asp His Met Asp
Arg Leu Pro Arg2075 2080 2085Asp Ile
Ala Gln Glu Arg Met His His Asp Ile Val Arg Leu Leu2090
2095 2100Asp Glu Tyr Asn Leu Val Arg Ser Pro Gln Leu
His Gly Thr Ala2105 2110 2115Leu Gly
Gly Thr Pro Thr Leu Ser Pro Thr Leu Cys Ser Pro Asn2120
2125 2130Gly Tyr Leu Gly Asn Leu Lys Ser Ala Thr Gln
Gly Lys Lys Ala2135 2140 2145Arg Lys
Pro Ser Thr Lys Gly Leu Ala Cys Gly Ser Lys Glu Ala2150
2155 2160Lys Asp Leu Lys Ala Arg Arg Lys Lys Ser Gln
Asp Gly Lys Gly2165 2170 2175Cys Leu
Leu Asp Ser Ser Ser Met Leu Ser Pro Val Asp Ser Leu2180
2185 2190Glu Ser Pro His Gly Tyr Leu Ser Asp Val Ala
Ser Pro Pro Leu2195 2200 2205Leu Pro
Ser Pro Phe Gln Gln Ser Pro Ser Met Pro Leu Ser His2210
2215 2220Leu Pro Gly Met Pro Asp Thr His Leu Gly Ile
Ser His Leu Asn2225 2230 2235Val Ala
Ala Lys Pro Glu Met Ala Ala Leu Ala Gly Gly Ser Arg2240
2245 2250Leu Ala Phe Glu Pro Pro Pro Pro Arg Leu Ser
His Leu Pro Val2255 2260 2265Ala Ser
Ser Ala Ser Thr Val Leu Ser Thr Asn Gly Thr Gly Ala2270
2275 2280Met Asn Phe Thr Val Gly Ala Pro Ala Ser Leu
Asn Gly Gln Cys2285 2290 2295Glu Trp
Leu Pro Arg Leu Gln Asn Gly Met Val Pro Ser Gln Tyr2300
2305 2310Asn Pro Leu Arg Pro Gly Val Thr Pro Gly Thr
Leu Ser Thr Gln2315 2320 2325Ala Ala
Gly Leu Gln His Ser Met Met Gly Pro Leu His Ser Ser2330
2335 2340Leu Ser Thr Asn Thr Leu Ser Pro Ile Ile Tyr
Gln Gly Leu Pro2345 2350 2355Asn Thr
Arg Leu Ala Thr Gln Pro His Leu Val Gln Thr Gln Gln2360
2365 2370Val Gln Pro Gln Asn Leu Gln Leu Gln Pro Gln
Asn Leu Gln Pro2375 2380 2385Pro Ser
Gln Pro His Leu Ser Val Ser Ser Ala Ala Asn Gly His2390
2395 2400Leu Gly Arg Ser Phe Leu Ser Gly Glu Pro Ser
Gln Ala Asp Val2405 2410 2415Gln Pro
Leu Gly Pro Ser Ser Leu Pro Val His Thr Ile Leu Pro2420
2425 2430Gln Glu Ser Gln Ala Leu Pro Thr Ser Leu Pro
Ser Ser Met Val2435 2440 2445Pro Pro
Met Thr Thr Thr Gln Phe Leu Thr Pro Pro Ser Gln His2450
2455 2460Ser Tyr Ser Ser Ser Pro Val Asp Asn Thr Pro
Ser His Gln Leu2465 2470 2475Gln Val
Pro Glu His Pro Phe Leu Thr Pro Ser Pro Glu Ser Pro2480
2485 2490Asp Gln Trp Ser Ser Ser Ser Pro His Ser Asn
Ile Ser Asp Trp2495 2500 2505Ser Glu
Gly Ile Ser Ser Pro Pro Thr Thr Met Pro Ser Gln Ile2510
2515 2520Thr His Ile Pro Glu Ala Phe Lys2525
253032346DNAMus musculusCDS(1)..(2346) 3cgg cgg cag cat ggc cag ctc
tgg ttc cct gag ggt ttc aaa gtg tca 48Arg Arg Gln His Gly Gln Leu
Trp Phe Pro Glu Gly Phe Lys Val Ser1 5 10
15gag gcc agc aag aag aag cgg aga gag ccc ctc ggc gag
gac tca gtc 96Glu Ala Ser Lys Lys Lys Arg Arg Glu Pro Leu Gly Glu
Asp Ser Val20 25 30ggc ctc aag ccc ctg
aag aat gcc tca gat ggt gct ctg atg gac gac 144Gly Leu Lys Pro Leu
Lys Asn Ala Ser Asp Gly Ala Leu Met Asp Asp35 40
45aat cag aac gag tgg gga gac gaa gac ctg gag acc aag aag ttc
cgg 192Asn Gln Asn Glu Trp Gly Asp Glu Asp Leu Glu Thr Lys Lys Phe
Arg50 55 60ttt gag gag cca gta gtt ctc
cct gac ctg agt gat cag act gac cac 240Phe Glu Glu Pro Val Val Leu
Pro Asp Leu Ser Asp Gln Thr Asp His65 70
75 80aga cag tgg acc cag cag cac ctg gac gct gct gac
ctg cgc atg tct 288Arg Gln Trp Thr Gln Gln His Leu Asp Ala Ala Asp
Leu Arg Met Ser85 90 95gcc atg gcc cca
aca ccg cct cag ggg gag gtg gat gct gac tgc atg 336Ala Met Ala Pro
Thr Pro Pro Gln Gly Glu Val Asp Ala Asp Cys Met100 105
110gat gtc aat gtt cga gga cca gat ggc ttc aca ccc ctc atg
att gcc 384Asp Val Asn Val Arg Gly Pro Asp Gly Phe Thr Pro Leu Met
Ile Ala115 120 125tcc tgc agt gga ggg ggc
ctt gag aca ggc aac agt gaa gaa gaa gaa 432Ser Cys Ser Gly Gly Gly
Leu Glu Thr Gly Asn Ser Glu Glu Glu Glu130 135
140gat gca cct gct gtc atc tct gac ttc atc tac cag ggc gcc agc ttg
480Asp Ala Pro Ala Val Ile Ser Asp Phe Ile Tyr Gln Gly Ala Ser Leu145
150 155 160cac aac cag aca
gac cgc acc ggg gag acc gcc ttg cac ttg gct gcc 528His Asn Gln Thr
Asp Arg Thr Gly Glu Thr Ala Leu His Leu Ala Ala165 170
175cga tac tct cgt tca gat gct gca aag cgc ttg ctg gag gcc
agt gca 576Arg Tyr Ser Arg Ser Asp Ala Ala Lys Arg Leu Leu Glu Ala
Ser Ala180 185 190gat gcc aac atc cag gac
aac atg ggc cgt act ccg tta cat gca gca 624Asp Ala Asn Ile Gln Asp
Asn Met Gly Arg Thr Pro Leu His Ala Ala195 200
205gtt tct gca gat gct cag ggt gtc ttc cag atc ctg ctc cgg aac agg
672Val Ser Ala Asp Ala Gln Gly Val Phe Gln Ile Leu Leu Arg Asn Arg210
215 220gcc aca gat ctg gat gcc cga atg cat
gat ggc aca act cca ctg atc 720Ala Thr Asp Leu Asp Ala Arg Met His
Asp Gly Thr Thr Pro Leu Ile225 230 235
240ctg gct gcg cgc ctg gcc ctg gag ggc atg ctg gag gac ctc
atc aac 768Leu Ala Ala Arg Leu Ala Leu Glu Gly Met Leu Glu Asp Leu
Ile Asn245 250 255tca cat gct gac gtc aat
gcc gtg gat gac cta ggc aag tcg gct ttg 816Ser His Ala Asp Val Asn
Ala Val Asp Asp Leu Gly Lys Ser Ala Leu260 265
270cat tgg gcg gcc gcg gtg aac aat gtg gat gct gct gtt gtg ctc ctg
864His Trp Ala Ala Ala Val Asn Asn Val Asp Ala Ala Val Val Leu Leu275
280 285aag aac gga gcc aac aag gac atg cag
aac aac aag gag gag act ccc 912Lys Asn Gly Ala Asn Lys Asp Met Gln
Asn Asn Lys Glu Glu Thr Pro290 295 300ctg
ttc ctg gcc gcc cgt gag ggc agc tat gag act gcc aaa gtg ttg 960Leu
Phe Leu Ala Ala Arg Glu Gly Ser Tyr Glu Thr Ala Lys Val Leu305
310 315 320ctg gac cac ttt gcc aac
cgg gac atc acg gat cac atg gac cga ttg 1008Leu Asp His Phe Ala Asn
Arg Asp Ile Thr Asp His Met Asp Arg Leu325 330
335ccg cgg gac atc gca cag gag cgt atg cac cac gat atc gtg cgg ctt
1056Pro Arg Asp Ile Ala Gln Glu Arg Met His His Asp Ile Val Arg Leu340
345 350ttg gat gag tac aac ctg gtg cgc agc
cca cag ctg cat ggc act gcc 1104Leu Asp Glu Tyr Asn Leu Val Arg Ser
Pro Gln Leu His Gly Thr Ala355 360 365ctg
ggt ggc aca ccc act ctg tct ccc aca ctc tgc tcg ccc aat ggc 1152Leu
Gly Gly Thr Pro Thr Leu Ser Pro Thr Leu Cys Ser Pro Asn Gly370
375 380tac ctg ggc aat ctc aag tct gcc aca cag ggc
aag aag gcc cgc aag 1200Tyr Leu Gly Asn Leu Lys Ser Ala Thr Gln Gly
Lys Lys Ala Arg Lys385 390 395
400ccc agc acc aaa ggg ctg gct tgt ggt agc aag gaa gct aag gac ctc
1248Pro Ser Thr Lys Gly Leu Ala Cys Gly Ser Lys Glu Ala Lys Asp Leu405
410 415aag gca cgg agg aag aag tcc cag gat
ggc aag ggc tgc ctg ttg gac 1296Lys Ala Arg Arg Lys Lys Ser Gln Asp
Gly Lys Gly Cys Leu Leu Asp420 425 430agc
tcg agc atg ctg tcg cct gtg gac tcc ctc gag tca ccc cat ggc 1344Ser
Ser Ser Met Leu Ser Pro Val Asp Ser Leu Glu Ser Pro His Gly435
440 445tac ttg tca gat gtg gcc tcg cca ccc ctc ctc
ccc tcc cca ttc cag 1392Tyr Leu Ser Asp Val Ala Ser Pro Pro Leu Leu
Pro Ser Pro Phe Gln450 455 460cag tct cca
tcc atg cct ctc agc cac ctg cct ggt atg cct gac act 1440Gln Ser Pro
Ser Met Pro Leu Ser His Leu Pro Gly Met Pro Asp Thr465
470 475 480cac ctg ggc atc agc cac ttg
aat gtg gca gcc aag cct gag atg gca 1488His Leu Gly Ile Ser His Leu
Asn Val Ala Ala Lys Pro Glu Met Ala485 490
495gca ctg gct gga ggt agc cgg ttg gcc ttt gag cca ccc ccg cca cgc
1536Ala Leu Ala Gly Gly Ser Arg Leu Ala Phe Glu Pro Pro Pro Pro Arg500
505 510ctc tcc cac ctg cct gta gcc tcc agt
gcc agc aca gtg ctg agt acc 1584Leu Ser His Leu Pro Val Ala Ser Ser
Ala Ser Thr Val Leu Ser Thr515 520 525aat
ggc acg ggg gct atg aat ttc acc gtg ggt gca ccg gca agc ttg 1632Asn
Gly Thr Gly Ala Met Asn Phe Thr Val Gly Ala Pro Ala Ser Leu530
535 540aat ggc cag tgt gag tgg ctt ccc cgg ctc cag
aat ggc atg gtg ccc 1680Asn Gly Gln Cys Glu Trp Leu Pro Arg Leu Gln
Asn Gly Met Val Pro545 550 555
560agc cag tac aac cca cta cgg ccg ggt gtg acg ccg ggc aca ctg agc
1728Ser Gln Tyr Asn Pro Leu Arg Pro Gly Val Thr Pro Gly Thr Leu Ser565
570 575aca cag gca gct ggc ctc cag cat agc
atg atg ggg cca cta cac agc 1776Thr Gln Ala Ala Gly Leu Gln His Ser
Met Met Gly Pro Leu His Ser580 585 590agc
ctc tcc acc aat acc ttg tcc ccg att att tac cag ggc ctg ccc 1824Ser
Leu Ser Thr Asn Thr Leu Ser Pro Ile Ile Tyr Gln Gly Leu Pro595
600 605aac aca cgg ctg gca aca cag cct cac ctg gtg
cag acc cag cag gtg 1872Asn Thr Arg Leu Ala Thr Gln Pro His Leu Val
Gln Thr Gln Gln Val610 615 620cag cca cag
aac tta cag ctc cag cct cag aac ctg cag cca cca tca 1920Gln Pro Gln
Asn Leu Gln Leu Gln Pro Gln Asn Leu Gln Pro Pro Ser625
630 635 640cag cca cac ctc agt gtg agc
tcg gca gcc aat ggg cac ctg ggc cgg 1968Gln Pro His Leu Ser Val Ser
Ser Ala Ala Asn Gly His Leu Gly Arg645 650
655agc ttc ttg agt ggg gag ccc agt cag gca gat gta caa ccg ctg ggc
2016Ser Phe Leu Ser Gly Glu Pro Ser Gln Ala Asp Val Gln Pro Leu Gly660
665 670ccc agc agt ctg cct gtg cac acc att
ctg ccc cag gaa agc cag gcc 2064Pro Ser Ser Leu Pro Val His Thr Ile
Leu Pro Gln Glu Ser Gln Ala675 680 685ctg
ccc aca tca ctg cca tcc tcc atg gtc cca ccc atg acc act acc 2112Leu
Pro Thr Ser Leu Pro Ser Ser Met Val Pro Pro Met Thr Thr Thr690
695 700cag ttc ctg acc cct cct tcc cag cac agt tac
tcc tcc tcc cct gtg 2160Gln Phe Leu Thr Pro Pro Ser Gln His Ser Tyr
Ser Ser Ser Pro Val705 710 715
720gac aac acc ccc agc cac cag ctg cag gtg cca gag cac ccc ttc ctc
2208Asp Asn Thr Pro Ser His Gln Leu Gln Val Pro Glu His Pro Phe Leu725
730 735acc cca tcc cct gag tcc cct gac cag
tgg tcc agc tcc tcc ccg cat 2256Thr Pro Ser Pro Glu Ser Pro Asp Gln
Trp Ser Ser Ser Ser Pro His740 745 750tcc
aac atc tct gat tgg tcc gag ggc atc tcc agc ccg ccc acc acc 2304Ser
Asn Ile Ser Asp Trp Ser Glu Gly Ile Ser Ser Pro Pro Thr Thr755
760 765atg ccg tcc cag atc acc cac att cca gag gca
ttt aaa taa 2346Met Pro Ser Gln Ile Thr His Ile Pro Glu Ala
Phe Lys770 775 7804781PRTMus musculus
4Arg Arg Gln His Gly Gln Leu Trp Phe Pro Glu Gly Phe Lys Val Ser1
5 10 15Glu Ala Ser Lys Lys Lys
Arg Arg Glu Pro Leu Gly Glu Asp Ser Val20 25
30Gly Leu Lys Pro Leu Lys Asn Ala Ser Asp Gly Ala Leu Met Asp Asp35
40 45Asn Gln Asn Glu Trp Gly Asp Glu Asp
Leu Glu Thr Lys Lys Phe Arg50 55 60Phe
Glu Glu Pro Val Val Leu Pro Asp Leu Ser Asp Gln Thr Asp His65
70 75 80Arg Gln Trp Thr Gln Gln
His Leu Asp Ala Ala Asp Leu Arg Met Ser85 90
95Ala Met Ala Pro Thr Pro Pro Gln Gly Glu Val Asp Ala Asp Cys Met100
105 110Asp Val Asn Val Arg Gly Pro Asp
Gly Phe Thr Pro Leu Met Ile Ala115 120
125Ser Cys Ser Gly Gly Gly Leu Glu Thr Gly Asn Ser Glu Glu Glu Glu130
135 140Asp Ala Pro Ala Val Ile Ser Asp Phe
Ile Tyr Gln Gly Ala Ser Leu145 150 155
160His Asn Gln Thr Asp Arg Thr Gly Glu Thr Ala Leu His Leu
Ala Ala165 170 175Arg Tyr Ser Arg Ser Asp
Ala Ala Lys Arg Leu Leu Glu Ala Ser Ala180 185
190Asp Ala Asn Ile Gln Asp Asn Met Gly Arg Thr Pro Leu His Ala
Ala195 200 205Val Ser Ala Asp Ala Gln Gly
Val Phe Gln Ile Leu Leu Arg Asn Arg210 215
220Ala Thr Asp Leu Asp Ala Arg Met His Asp Gly Thr Thr Pro Leu Ile225
230 235 240Leu Ala Ala Arg
Leu Ala Leu Glu Gly Met Leu Glu Asp Leu Ile Asn245 250
255Ser His Ala Asp Val Asn Ala Val Asp Asp Leu Gly Lys Ser
Ala Leu260 265 270His Trp Ala Ala Ala Val
Asn Asn Val Asp Ala Ala Val Val Leu Leu275 280
285Lys Asn Gly Ala Asn Lys Asp Met Gln Asn Asn Lys Glu Glu Thr
Pro290 295 300Leu Phe Leu Ala Ala Arg Glu
Gly Ser Tyr Glu Thr Ala Lys Val Leu305 310
315 320Leu Asp His Phe Ala Asn Arg Asp Ile Thr Asp His
Met Asp Arg Leu325 330 335Pro Arg Asp Ile
Ala Gln Glu Arg Met His His Asp Ile Val Arg Leu340 345
350Leu Asp Glu Tyr Asn Leu Val Arg Ser Pro Gln Leu His Gly
Thr Ala355 360 365Leu Gly Gly Thr Pro Thr
Leu Ser Pro Thr Leu Cys Ser Pro Asn Gly370 375
380Tyr Leu Gly Asn Leu Lys Ser Ala Thr Gln Gly Lys Lys Ala Arg
Lys385 390 395 400Pro Ser
Thr Lys Gly Leu Ala Cys Gly Ser Lys Glu Ala Lys Asp Leu405
410 415Lys Ala Arg Arg Lys Lys Ser Gln Asp Gly Lys Gly
Cys Leu Leu Asp420 425 430Ser Ser Ser Met
Leu Ser Pro Val Asp Ser Leu Glu Ser Pro His Gly435 440
445Tyr Leu Ser Asp Val Ala Ser Pro Pro Leu Leu Pro Ser Pro
Phe Gln450 455 460Gln Ser Pro Ser Met Pro
Leu Ser His Leu Pro Gly Met Pro Asp Thr465 470
475 480His Leu Gly Ile Ser His Leu Asn Val Ala Ala
Lys Pro Glu Met Ala485 490 495Ala Leu Ala
Gly Gly Ser Arg Leu Ala Phe Glu Pro Pro Pro Pro Arg500
505 510Leu Ser His Leu Pro Val Ala Ser Ser Ala Ser Thr
Val Leu Ser Thr515 520 525Asn Gly Thr Gly
Ala Met Asn Phe Thr Val Gly Ala Pro Ala Ser Leu530 535
540Asn Gly Gln Cys Glu Trp Leu Pro Arg Leu Gln Asn Gly Met
Val Pro545 550 555 560Ser
Gln Tyr Asn Pro Leu Arg Pro Gly Val Thr Pro Gly Thr Leu Ser565
570 575Thr Gln Ala Ala Gly Leu Gln His Ser Met Met
Gly Pro Leu His Ser580 585 590Ser Leu Ser
Thr Asn Thr Leu Ser Pro Ile Ile Tyr Gln Gly Leu Pro595
600 605Asn Thr Arg Leu Ala Thr Gln Pro His Leu Val Gln
Thr Gln Gln Val610 615 620Gln Pro Gln Asn
Leu Gln Leu Gln Pro Gln Asn Leu Gln Pro Pro Ser625 630
635 640Gln Pro His Leu Ser Val Ser Ser Ala
Ala Asn Gly His Leu Gly Arg645 650 655Ser
Phe Leu Ser Gly Glu Pro Ser Gln Ala Asp Val Gln Pro Leu Gly660
665 670Pro Ser Ser Leu Pro Val His Thr Ile Leu Pro
Gln Glu Ser Gln Ala675 680 685Leu Pro Thr
Ser Leu Pro Ser Ser Met Val Pro Pro Met Thr Thr Thr690
695 700Gln Phe Leu Thr Pro Pro Ser Gln His Ser Tyr Ser
Ser Ser Pro Val705 710 715
720Asp Asn Thr Pro Ser His Gln Leu Gln Val Pro Glu His Pro Phe Leu725
730 735Thr Pro Ser Pro Glu Ser Pro Asp Gln
Trp Ser Ser Ser Ser Pro His740 745 750Ser
Asn Ile Ser Asp Trp Ser Glu Gly Ile Ser Ser Pro Pro Thr Thr755
760 765Met Pro Ser Gln Ile Thr His Ile Pro Glu Ala
Phe Lys770 775 7805900DNAMus
musculusCDS(1)..(900) 5atg aag aga gct cac cca gac tac agc tcc tca gat
agt gag ctg gac 48Met Lys Arg Ala His Pro Asp Tyr Ser Ser Ser Asp
Ser Glu Leu Asp1 5 10
15gag acc atc gag gtg gaa aag gag agc gcg gac gag aat gga aac ttg
96Glu Thr Ile Glu Val Glu Lys Glu Ser Ala Asp Glu Asn Gly Asn Leu20
25 30agt tcg gcg ctg tgt tcc atg tcc cca acg
aca tcg tcc cag gtt ttg 144Ser Ser Ala Leu Cys Ser Met Ser Pro Thr
Thr Ser Ser Gln Val Leu35 40 45gcc agg
aaa aga cgg aga ggc atc atc gag aag cgc cga cga gac cga 192Ala Arg
Lys Arg Arg Arg Gly Ile Ile Glu Lys Arg Arg Arg Asp Arg50
55 60atc aat aac agt ttg tct gag ctg aga agg ctg gta
ccc agt gcc ttt 240Ile Asn Asn Ser Leu Ser Glu Leu Arg Arg Leu Val
Pro Ser Ala Phe65 70 75
80gag aag cag gga tct gct aag cta gaa aaa gct gag atc ttg cag atg
288Glu Lys Gln Gly Ser Ala Lys Leu Glu Lys Ala Glu Ile Leu Gln Met85
90 95act gtg gat cac ctg aaa atg ctg cac act
gca gga ggg aaa ggt tat 336Thr Val Asp His Leu Lys Met Leu His Thr
Ala Gly Gly Lys Gly Tyr100 105 110ttt gac
gcg cac gcc ctg gct atg gac tat cgg agt ttg ggg ttt cgg 384Phe Asp
Ala His Ala Leu Ala Met Asp Tyr Arg Ser Leu Gly Phe Arg115
120 125gaa tgc ctg gcc gaa gtt gcc cgt tat ctg agc atc
att gaa gga ctc 432Glu Cys Leu Ala Glu Val Ala Arg Tyr Leu Ser Ile
Ile Glu Gly Leu130 135 140gat gcc tcc gac
ccg ctt cgc gtt cgc ctg gtc tcc cat ctc aac aac 480Asp Ala Ser Asp
Pro Leu Arg Val Arg Leu Val Ser His Leu Asn Asn145 150
155 160tac gca tcc cag cgg gaa gcc gcg agc
ggc gct cac ggt ggc ctc gga 528Tyr Ala Ser Gln Arg Glu Ala Ala Ser
Gly Ala His Gly Gly Leu Gly165 170 175cac
att ccc tgg gga agt gcc ttc gga cat cac cca cac atc gca cac 576His
Ile Pro Trp Gly Ser Ala Phe Gly His His Pro His Ile Ala His180
185 190cct ctg ctg ctg ccc cag aat ggc cac ggg aac
gct ggc acc gcg gcg 624Pro Leu Leu Leu Pro Gln Asn Gly His Gly Asn
Ala Gly Thr Ala Ala195 200 205tca ccc acg
gag ccg cat cac cag ggc agg ctg gct tct gcc cat ccg 672Ser Pro Thr
Glu Pro His His Gln Gly Arg Leu Ala Ser Ala His Pro210
215 220gag gcg ccg gcc ttg cga gcg ccc cct agt ggc ggc
ttg gga ccg gtg 720Glu Ala Pro Ala Leu Arg Ala Pro Pro Ser Gly Gly
Leu Gly Pro Val225 230 235
240ctt ccc gtg gtg acc tcg gcc tcc aaa ctg tct cca ccg ctg ctc tcc
768Leu Pro Val Val Thr Ser Ala Ser Lys Leu Ser Pro Pro Leu Leu Ser245
250 255tct gtg gcc tcg ctc tca gcc ttc ccc
ttt tcc ttc agc tcc ttc cac 816Ser Val Ala Ser Leu Ser Ala Phe Pro
Phe Ser Phe Ser Ser Phe His260 265 270cta
ctg agc cct tcg aca ccc acg cag gca gca aac ctt ggc aag ccc 864Leu
Leu Ser Pro Ser Thr Pro Thr Gln Ala Ala Asn Leu Gly Lys Pro275
280 285tat aga cct tgg ggg aca gag atc gga gct ttc
taa 900Tyr Arg Pro Trp Gly Thr Glu Ile Gly Ala
Phe290 2956299PRTMus musculus 6Met Lys Arg Ala His Pro
Asp Tyr Ser Ser Ser Asp Ser Glu Leu Asp1 5
10 15Glu Thr Ile Glu Val Glu Lys Glu Ser Ala Asp Glu
Asn Gly Asn Leu20 25 30Ser Ser Ala Leu
Cys Ser Met Ser Pro Thr Thr Ser Ser Gln Val Leu35 40
45Ala Arg Lys Arg Arg Arg Gly Ile Ile Glu Lys Arg Arg Arg
Asp Arg50 55 60Ile Asn Asn Ser Leu Ser
Glu Leu Arg Arg Leu Val Pro Ser Ala Phe65 70
75 80Glu Lys Gln Gly Ser Ala Lys Leu Glu Lys Ala
Glu Ile Leu Gln Met85 90 95Thr Val Asp
His Leu Lys Met Leu His Thr Ala Gly Gly Lys Gly Tyr100
105 110Phe Asp Ala His Ala Leu Ala Met Asp Tyr Arg Ser
Leu Gly Phe Arg115 120 125Glu Cys Leu Ala
Glu Val Ala Arg Tyr Leu Ser Ile Ile Glu Gly Leu130 135
140Asp Ala Ser Asp Pro Leu Arg Val Arg Leu Val Ser His Leu
Asn Asn145 150 155 160Tyr
Ala Ser Gln Arg Glu Ala Ala Ser Gly Ala His Gly Gly Leu Gly165
170 175His Ile Pro Trp Gly Ser Ala Phe Gly His His
Pro His Ile Ala His180 185 190Pro Leu Leu
Leu Pro Gln Asn Gly His Gly Asn Ala Gly Thr Ala Ala195
200 205Ser Pro Thr Glu Pro His His Gln Gly Arg Leu Ala
Ser Ala His Pro210 215 220Glu Ala Pro Ala
Leu Arg Ala Pro Pro Ser Gly Gly Leu Gly Pro Val225 230
235 240Leu Pro Val Val Thr Ser Ala Ser Lys
Leu Ser Pro Pro Leu Leu Ser245 250 255Ser
Val Ala Ser Leu Ser Ala Phe Pro Phe Ser Phe Ser Ser Phe His260
265 270Leu Leu Ser Pro Ser Thr Pro Thr Gln Ala Ala
Asn Leu Gly Lys Pro275 280 285Tyr Arg Pro
Trp Gly Thr Glu Ile Gly Ala Phe290 29571020DNAMus
musculusCDS(1)..(1020) 7atg aag cgc cct tgt gag gaa acg acc tcc gaa agc
gac ctg gac gag 48Met Lys Arg Pro Cys Glu Glu Thr Thr Ser Glu Ser
Asp Leu Asp Glu1 5 10
15acc atc gac gtg ggg agc gag aac aat tac cct ggg cac gct aca agc
96Thr Ile Asp Val Gly Ser Glu Asn Asn Tyr Pro Gly His Ala Thr Ser20
25 30tca gtg atg agg tcc aat tca ccg aca act
acc tct cag att atg gca 144Ser Val Met Arg Ser Asn Ser Pro Thr Thr
Thr Ser Gln Ile Met Ala35 40 45aga aag
aaa agg aga ggg atc ata gaa aaa agg cgt cgg gat cga ata 192Arg Lys
Lys Arg Arg Gly Ile Ile Glu Lys Arg Arg Arg Asp Arg Ile50
55 60aat aac agt tta tct gaa ttg aga aga cta gtg cca
aca gct ttt gaa 240Asn Asn Ser Leu Ser Glu Leu Arg Arg Leu Val Pro
Thr Ala Phe Glu65 70 75
80aaa caa gga tct gcc aag tta gaa aag gct gaa ata ttg caa atg aca
288Lys Gln Gly Ser Ala Lys Leu Glu Lys Ala Glu Ile Leu Gln Met Thr85
90 95gtg gat cat ttg aag atg ctc cag gct aca
ggg ggt aaa ggc tac ttt 336Val Asp His Leu Lys Met Leu Gln Ala Thr
Gly Gly Lys Gly Tyr Phe100 105 110gat gcc
cat gct ctt gcc aca gac ttc atg agc att gga ttc cga gag 384Asp Ala
His Ala Leu Ala Thr Asp Phe Met Ser Ile Gly Phe Arg Glu115
120 125tgc ttg aca gaa gtg gct agg tac cta agc tca gtg
gaa ggc ctt gac 432Cys Leu Thr Glu Val Ala Arg Tyr Leu Ser Ser Val
Glu Gly Leu Asp130 135 140ccg tcg gac cca
cta cgc gtg cgc ctt gtc tct cat ctc agc acc tgt 480Pro Ser Asp Pro
Leu Arg Val Arg Leu Val Ser His Leu Ser Thr Cys145 150
155 160gcc tcc cag cgg gag gca gca gtg atg
aca tcc tcc atg gcc cac cac 528Ala Ser Gln Arg Glu Ala Ala Val Met
Thr Ser Ser Met Ala His His165 170 175cat
cac ccc ttg cac cct cac cac tgg gca gct gct ttc cac cat ctc 576His
His Pro Leu His Pro His His Trp Ala Ala Ala Phe His His Leu180
185 190ccc aca gcc ctg ctc cag ccc aat gga ctc cac
aca tca gag tca acc 624Pro Thr Ala Leu Leu Gln Pro Asn Gly Leu His
Thr Ser Glu Ser Thr195 200 205cca tgt cgc
cta tcc aca tct tca gaa gtg cct tct gct cat ggc tct 672Pro Cys Arg
Leu Ser Thr Ser Ser Glu Val Pro Ser Ala His Gly Ser210
215 220gct ctc ctc aca gca acg ttt gcc cat gca gat tct
gct ctt cgg atg 720Ala Leu Leu Thr Ala Thr Phe Ala His Ala Asp Ser
Ala Leu Arg Met225 230 235
240cca tca ggg ggc acc gtt gca ccc tgc gtg cca cct ctc tcc acc tct
768Pro Ser Gly Gly Thr Val Ala Pro Cys Val Pro Pro Leu Ser Thr Ser245
250 255ctt ctg tct ctt tcg gcc act gtg cat
gcc gca gct gca gca gcc act 816Leu Leu Ser Leu Ser Ala Thr Val His
Ala Ala Ala Ala Ala Ala Thr260 265 270gca
gct gca cac agc ttc cct ctg tcc ttc gca ggg gct ttt ccc atg 864Ala
Ala Ala His Ser Phe Pro Leu Ser Phe Ala Gly Ala Phe Pro Met275
280 285ctc ccg tcc aat gca gcg gca gca gcc gct gtt
gct gct gca aca gca 912Leu Pro Ser Asn Ala Ala Ala Ala Ala Ala Val
Ala Ala Ala Thr Ala290 295 300atc agc cca
ccc ttg tcg gta tcc gca gcc tcc agt cct cag cag aca 960Ile Ser Pro
Pro Leu Ser Val Ser Ala Ala Ser Ser Pro Gln Gln Thr305
310 315 320agc act ggg aca aac aat aaa
cct tac caa ccc tgg ggg aca gaa gtt 1008Ser Thr Gly Thr Asn Asn Lys
Pro Tyr Gln Pro Trp Gly Thr Glu Val325 330
335gga gcc ttt taa
1020Gly Ala Phe8339PRTMus musculus 8Met Lys Arg Pro Cys Glu Glu Thr Thr
Ser Glu Ser Asp Leu Asp Glu1 5 10
15Thr Ile Asp Val Gly Ser Glu Asn Asn Tyr Pro Gly His Ala Thr
Ser20 25 30Ser Val Met Arg Ser Asn Ser
Pro Thr Thr Thr Ser Gln Ile Met Ala35 40
45Arg Lys Lys Arg Arg Gly Ile Ile Glu Lys Arg Arg Arg Asp Arg Ile50
55 60Asn Asn Ser Leu Ser Glu Leu Arg Arg Leu
Val Pro Thr Ala Phe Glu65 70 75
80Lys Gln Gly Ser Ala Lys Leu Glu Lys Ala Glu Ile Leu Gln Met
Thr85 90 95Val Asp His Leu Lys Met Leu
Gln Ala Thr Gly Gly Lys Gly Tyr Phe100 105
110Asp Ala His Ala Leu Ala Thr Asp Phe Met Ser Ile Gly Phe Arg Glu115
120 125Cys Leu Thr Glu Val Ala Arg Tyr Leu
Ser Ser Val Glu Gly Leu Asp130 135 140Pro
Ser Asp Pro Leu Arg Val Arg Leu Val Ser His Leu Ser Thr Cys145
150 155 160Ala Ser Gln Arg Glu Ala
Ala Val Met Thr Ser Ser Met Ala His His165 170
175His His Pro Leu His Pro His His Trp Ala Ala Ala Phe His His
Leu180 185 190Pro Thr Ala Leu Leu Gln Pro
Asn Gly Leu His Thr Ser Glu Ser Thr195 200
205Pro Cys Arg Leu Ser Thr Ser Ser Glu Val Pro Ser Ala His Gly Ser210
215 220Ala Leu Leu Thr Ala Thr Phe Ala His
Ala Asp Ser Ala Leu Arg Met225 230 235
240Pro Ser Gly Gly Thr Val Ala Pro Cys Val Pro Pro Leu Ser
Thr Ser245 250 255Leu Leu Ser Leu Ser Ala
Thr Val His Ala Ala Ala Ala Ala Ala Thr260 265
270Ala Ala Ala His Ser Phe Pro Leu Ser Phe Ala Gly Ala Phe Pro
Met275 280 285Leu Pro Ser Asn Ala Ala Ala
Ala Ala Ala Val Ala Ala Ala Thr Ala290 295
300Ile Ser Pro Pro Leu Ser Val Ser Ala Ala Ser Ser Pro Gln Gln Thr305
310 315 320Ser Thr Gly Thr
Asn Asn Lys Pro Tyr Gln Pro Trp Gly Thr Glu Val325 330
335Gly Ala Phe9849DNAMus musculusCDS(1)..(849) 9atg cca gct
gat ata atg gag aaa aat tcc tcc tcc ccg gtg gct gct 48Met Pro Ala
Asp Ile Met Glu Lys Asn Ser Ser Ser Pro Val Ala Ala1 5
10 15acc cca gcc agt gtc aac acg aca ccg
gac aaa cca aag acg gcc tct 96Thr Pro Ala Ser Val Asn Thr Thr Pro
Asp Lys Pro Lys Thr Ala Ser20 25 30gag
cac aga aag tca tca aag cct atc atg gag aag agg cga agg gca 144Glu
His Arg Lys Ser Ser Lys Pro Ile Met Glu Lys Arg Arg Arg Ala35
40 45aga ata aat gaa agt cta agc caa ctg aaa aca
ctg att ttg gat gca 192Arg Ile Asn Glu Ser Leu Ser Gln Leu Lys Thr
Leu Ile Leu Asp Ala50 55 60ctt aag aaa
gat agc tcc cgg cat tcc aag cta gag aag gca gac att 240Leu Lys Lys
Asp Ser Ser Arg His Ser Lys Leu Glu Lys Ala Asp Ile65 70
75 80ctg gaa atg act gtg aag cac ctc
cgg aac ctg cag cgg gcg cag atg 288Leu Glu Met Thr Val Lys His Leu
Arg Asn Leu Gln Arg Ala Gln Met85 90
95acc gcc gcg ctc agc aca gac ccg agc gtg ttg ggg aaa tac cgc gcc
336Thr Ala Ala Leu Ser Thr Asp Pro Ser Val Leu Gly Lys Tyr Arg Ala100
105 110ggc ttc agc gaa gag tgc atg agc gag
gtg acg cgt ttc ctg tcc acg 384Gly Phe Ser Glu Glu Cys Met Ser Glu
Val Thr Arg Phe Leu Ser Thr115 120 125tgt
gag ggc gtt acc acc gag gtg ctc act cgg cta atg ggc cac ctg 432Cys
Glu Gly Val Thr Thr Glu Val Leu Thr Arg Leu Met Gly His Leu130
135 140gcc agt tgc atg acc cag atc atc gcc atg tac
ccc ggg cag gcg cac 480Ala Ser Cys Met Thr Gln Ile Ile Ala Met Tyr
Pro Gly Gln Ala His145 150 155
160ccc gcc ttg cag gcg ccg ccg ccg ccg ccc ccg tca gga cct gcc ggt
528Pro Ala Leu Gln Ala Pro Pro Pro Pro Pro Pro Ser Gly Pro Ala Gly165
170 175ccc cag cac gcg cca ttc gcg ccg ccg
ccg ccg ccg ctt gtg ccc atc 576Pro Gln His Ala Pro Phe Ala Pro Pro
Pro Pro Pro Leu Val Pro Ile180 185 190ccc
ggg ggc gcg gcg ccc cct ccc ggc agc gca ccc tgc aag ttg ggc 624Pro
Gly Gly Ala Ala Pro Pro Pro Gly Ser Ala Pro Cys Lys Leu Gly195
200 205agc cag gct gga gag gct gcc aag gtt ttt ggc
ggc ttc caa gtg gtg 672Ser Gln Ala Gly Glu Ala Ala Lys Val Phe Gly
Gly Phe Gln Val Val210 215 220ccg gct cct
gac ggc caa ttt gcc ttt ctc atc ccc aac ggg gcc ttc 720Pro Ala Pro
Asp Gly Gln Phe Ala Phe Leu Ile Pro Asn Gly Ala Phe225
230 235 240gct cac agc ggc ccg gtc atc
ccg gtc tac acc agc aac agt ggg acc 768Ala His Ser Gly Pro Val Ile
Pro Val Tyr Thr Ser Asn Ser Gly Thr245 250
255tcg gtg ggt cct aac gca gtg tca cct tcc agt ggc tcc tcg ctc act
816Ser Val Gly Pro Asn Ala Val Ser Pro Ser Ser Gly Ser Ser Leu Thr260
265 270tcg gac tcc atg tgg aga ccg tgg cgg
aac tga 849Ser Asp Ser Met Trp Arg Pro Trp Arg
Asn275 28010282PRTMus musculus 10Met Pro Ala Asp Ile Met
Glu Lys Asn Ser Ser Ser Pro Val Ala Ala1 5
10 15Thr Pro Ala Ser Val Asn Thr Thr Pro Asp Lys Pro
Lys Thr Ala Ser20 25 30Glu His Arg Lys
Ser Ser Lys Pro Ile Met Glu Lys Arg Arg Arg Ala35 40
45Arg Ile Asn Glu Ser Leu Ser Gln Leu Lys Thr Leu Ile Leu
Asp Ala50 55 60Leu Lys Lys Asp Ser Ser
Arg His Ser Lys Leu Glu Lys Ala Asp Ile65 70
75 80Leu Glu Met Thr Val Lys His Leu Arg Asn Leu
Gln Arg Ala Gln Met85 90 95Thr Ala Ala
Leu Ser Thr Asp Pro Ser Val Leu Gly Lys Tyr Arg Ala100
105 110Gly Phe Ser Glu Glu Cys Met Ser Glu Val Thr Arg
Phe Leu Ser Thr115 120 125Cys Glu Gly Val
Thr Thr Glu Val Leu Thr Arg Leu Met Gly His Leu130 135
140Ala Ser Cys Met Thr Gln Ile Ile Ala Met Tyr Pro Gly Gln
Ala His145 150 155 160Pro
Ala Leu Gln Ala Pro Pro Pro Pro Pro Pro Ser Gly Pro Ala Gly165
170 175Pro Gln His Ala Pro Phe Ala Pro Pro Pro Pro
Pro Leu Val Pro Ile180 185 190Pro Gly Gly
Ala Ala Pro Pro Pro Gly Ser Ala Pro Cys Lys Leu Gly195
200 205Ser Gln Ala Gly Glu Ala Ala Lys Val Phe Gly Gly
Phe Gln Val Val210 215 220Pro Ala Pro Asp
Gly Gln Phe Ala Phe Leu Ile Pro Asn Gly Ala Phe225 230
235 240Ala His Ser Gly Pro Val Ile Pro Val
Tyr Thr Ser Asn Ser Gly Thr245 250 255Ser
Val Gly Pro Asn Ala Val Ser Pro Ser Ser Gly Ser Ser Leu Thr260
265 270Ser Asp Ser Met Trp Arg Pro Trp Arg Asn275
28011528DNAMus musculusCDS(1)..(528) 11atg gag aag aag cgc
cgt gca cgc atc aac gtg tca ctg gag cag ctg 48Met Glu Lys Lys Arg
Arg Ala Arg Ile Asn Val Ser Leu Glu Gln Leu1 5
10 15agg tct ctt ctg gag aga cac tac tca cat cag
ata cgg aaa cga aag 96Arg Ser Leu Leu Glu Arg His Tyr Ser His Gln
Ile Arg Lys Arg Lys20 25 30ctg gag aag
gcc gat atc ctg gag ctg agt gtt aag tac atg aga agc 144Leu Glu Lys
Ala Asp Ile Leu Glu Leu Ser Val Lys Tyr Met Arg Ser35 40
45ctc cag aac tca ctg caa gga ctc tgg cca gta ccc agt
ggg gtg gac 192Leu Gln Asn Ser Leu Gln Gly Leu Trp Pro Val Pro Ser
Gly Val Asp50 55 60tac ccg tcc ggc ttc
caa ggc ggc ttg cgt ggc gtg agc cag agg ctt 240Tyr Pro Ser Gly Phe
Gln Gly Gly Leu Arg Gly Val Ser Gln Arg Leu65 70
75 80cgg ccc gga gag gga gac agc ggc ctg cgc
tgc ccc ttg ctg ctc cag 288Arg Pro Gly Glu Gly Asp Ser Gly Leu Arg
Cys Pro Leu Leu Leu Gln85 90 95cgc agg
gaa ggc agc acc acg gac agc gcc aac cca cag gcg acc tct 336Arg Arg
Glu Gly Ser Thr Thr Asp Ser Ala Asn Pro Gln Ala Thr Ser100
105 110gtt ctc aac cct tgc ctc ccg gcc atc tgg gcc ccc
agt cgc gct gca 384Val Leu Asn Pro Cys Leu Pro Ala Ile Trp Ala Pro
Ser Arg Ala Ala115 120 125ggt ggc tcc cac
tcc ccg cag tcc ccg ctc cct ctc cct gga ggt ctc 432Gly Gly Ser His
Ser Pro Gln Ser Pro Leu Pro Leu Pro Gly Gly Leu130 135
140ctt gag tct tcc acc gat gtt gtg gca cca cat cct gca tca
aac tgc 480Leu Glu Ser Ser Thr Asp Val Val Ala Pro His Pro Ala Ser
Asn Cys145 150 155 160cag
gcc gag agt acg aga ccc ggg ttt cgc gtg tgg cga ccg tgg tga 528Gln
Ala Glu Ser Thr Arg Pro Gly Phe Arg Val Trp Arg Pro Trp165
170 17512175PRTMus musculus 12Met Glu Lys Lys Arg Arg
Ala Arg Ile Asn Val Ser Leu Glu Gln Leu1 5
10 15Arg Ser Leu Leu Glu Arg His Tyr Ser His Gln Ile
Arg Lys Arg Lys20 25 30Leu Glu Lys Ala
Asp Ile Leu Glu Leu Ser Val Lys Tyr Met Arg Ser35 40
45Leu Gln Asn Ser Leu Gln Gly Leu Trp Pro Val Pro Ser Gly
Val Asp50 55 60Tyr Pro Ser Gly Phe Gln
Gly Gly Leu Arg Gly Val Ser Gln Arg Leu65 70
75 80Arg Pro Gly Glu Gly Asp Ser Gly Leu Arg Cys
Pro Leu Leu Leu Gln85 90 95Arg Arg Glu
Gly Ser Thr Thr Asp Ser Ala Asn Pro Gln Ala Thr Ser100
105 110Val Leu Asn Pro Cys Leu Pro Ala Ile Trp Ala Pro
Ser Arg Ala Ala115 120 125Gly Gly Ser His
Ser Pro Gln Ser Pro Leu Pro Leu Pro Gly Gly Leu130 135
140Leu Glu Ser Ser Thr Asp Val Val Ala Pro His Pro Ala Ser
Asn Cys145 150 155 160Gln
Ala Glu Ser Thr Arg Pro Gly Phe Arg Val Trp Arg Pro Trp165
170 17513504DNAMus musculusCDS(1)..(504) 13atg gcc cca
agt acc gtg gcg gtg gag atg ctc agt ccc aag gag aaa 48Met Ala Pro
Ser Thr Val Ala Val Glu Met Leu Ser Pro Lys Glu Lys1 5
10 15aac cga ctg cgg aag ccg gtg gtg gag
aag atg cgt cgg gac cgc atc 96Asn Arg Leu Arg Lys Pro Val Val Glu
Lys Met Arg Arg Asp Arg Ile20 25 30aac
agc agc ata gag cag ctg aag ctg ctg ctg gag cag gag ttc gcg 144Asn
Ser Ser Ile Glu Gln Leu Lys Leu Leu Leu Glu Gln Glu Phe Ala35
40 45cgg cac cag ccc aac tcc aag ctg gag aag gcc
gac atc ctg gag atg 192Arg His Gln Pro Asn Ser Lys Leu Glu Lys Ala
Asp Ile Leu Glu Met50 55 60gcc gtc agc
tac ctg aaa cac agc aaa gcc ttc gcc gcg gcc gcc ggc 240Ala Val Ser
Tyr Leu Lys His Ser Lys Ala Phe Ala Ala Ala Ala Gly65 70
75 80ccc aag agc ctg cac cag gac tac
agc gag ggc tac tcc tgg tgc ctg 288Pro Lys Ser Leu His Gln Asp Tyr
Ser Glu Gly Tyr Ser Trp Cys Leu85 90
95cag gag gcg gta cag ttc ctg acc ctg cac gcc gcc agc gac acg cag
336Gln Glu Ala Val Gln Phe Leu Thr Leu His Ala Ala Ser Asp Thr Gln100
105 110atg aag ctg ctt tac cac ttc cag cgg
ccc cca gct ccc gcc gct cct 384Met Lys Leu Leu Tyr His Phe Gln Arg
Pro Pro Ala Pro Ala Ala Pro115 120 125gcc
aag gag ccc ccg gcg ccc gga gct gcg ccc caa ccg gcc cgc tcc 432Ala
Lys Glu Pro Pro Ala Pro Gly Ala Ala Pro Gln Pro Ala Arg Ser130
135 140tcc gct aag gct gct gct gct gcg gtc tcc acc
tcg cgc caa ccc gcc 480Ser Ala Lys Ala Ala Ala Ala Ala Val Ser Thr
Ser Arg Gln Pro Ala145 150 155
160tgc ggc ctc tgg cgg ccc tgg tga
504Cys Gly Leu Trp Arg Pro Trp16514167PRTMus musculus 14Met Ala Pro
Ser Thr Val Ala Val Glu Met Leu Ser Pro Lys Glu Lys1 5
10 15Asn Arg Leu Arg Lys Pro Val Val Glu
Lys Met Arg Arg Asp Arg Ile20 25 30Asn
Ser Ser Ile Glu Gln Leu Lys Leu Leu Leu Glu Gln Glu Phe Ala35
40 45Arg His Gln Pro Asn Ser Lys Leu Glu Lys Ala
Asp Ile Leu Glu Met50 55 60Ala Val Ser
Tyr Leu Lys His Ser Lys Ala Phe Ala Ala Ala Ala Gly65 70
75 80Pro Lys Ser Leu His Gln Asp Tyr
Ser Glu Gly Tyr Ser Trp Cys Leu85 90
95Gln Glu Ala Val Gln Phe Leu Thr Leu His Ala Ala Ser Asp Thr Gln100
105 110Met Lys Leu Leu Tyr His Phe Gln Arg Pro
Pro Ala Pro Ala Ala Pro115 120 125Ala Lys
Glu Pro Pro Ala Pro Gly Ala Ala Pro Gln Pro Ala Arg Ser130
135 140Ser Ala Lys Ala Ala Ala Ala Ala Val Ser Thr Ser
Arg Gln Pro Ala145 150 155
160Cys Gly Leu Trp Arg Pro Trp16515675DNAMus musculusCDS(1)..(675) 15atg
gct ccg tcc cag gcg ccc agc cgg gac cgt gca ggc cag gag gat 48Met
Ala Pro Ser Gln Ala Pro Ser Arg Asp Arg Ala Gly Gln Glu Asp1
5 10 15gag gac cgc tgg gaa gca cgg
ggg gac cgc aag gcc cgg aag ccc ctg 96Glu Asp Arg Trp Glu Ala Arg
Gly Asp Arg Lys Ala Arg Lys Pro Leu20 25
30gtg gag aag aag cga cgc gca cgg atc aac gag agt ctt cag gag ctg
144Val Glu Lys Lys Arg Arg Ala Arg Ile Asn Glu Ser Leu Gln Glu Leu35
40 45cgg ctg ctg ctg gcc ggt acc gag gtg cag
gcc aag cta gag aac gcc 192Arg Leu Leu Leu Ala Gly Thr Glu Val Gln
Ala Lys Leu Glu Asn Ala50 55 60gag gtt
ctg gag ctg acc gtg agg cgc gtg cag ggc gcg ctg cga ggc 240Glu Val
Leu Glu Leu Thr Val Arg Arg Val Gln Gly Ala Leu Arg Gly65
70 75 80cgg gcg cgc gag cgg gag cag
ctg cag gct gaa gca agc gag cgc ttc 288Arg Ala Arg Glu Arg Glu Gln
Leu Gln Ala Glu Ala Ser Glu Arg Phe85 90
95gct gct ggc tac atc cag tgc atg cat gag gtg cac acg ttc gtg tcc
336Ala Ala Gly Tyr Ile Gln Cys Met His Glu Val His Thr Phe Val Ser100
105 110acg tgc caa gcc atc gat gcc act gtc
tca gct gaa ctc ctg aac cac 384Thr Cys Gln Ala Ile Asp Ala Thr Val
Ser Ala Glu Leu Leu Asn His115 120 125ctg
cta gaa tcc atg ccg ctg cgt gag ggt agc agc ttt cag gat ctg 432Leu
Leu Glu Ser Met Pro Leu Arg Glu Gly Ser Ser Phe Gln Asp Leu130
135 140ctg ggg gac tcc cta gct ggg ctg cct gga ggc
tct ggg agg agc agc 480Leu Gly Asp Ser Leu Ala Gly Leu Pro Gly Gly
Ser Gly Arg Ser Ser145 150 155
160tgg cct cca gga ggg tcc cca gaa tcc cca ttg tcc agt ccc cca ggt
528Trp Pro Pro Gly Gly Ser Pro Glu Ser Pro Leu Ser Ser Pro Pro Gly165
170 175ccc ggg gac gac ctg tgt tct gac cta
gag gag atc cca gag gct gaa 576Pro Gly Asp Asp Leu Cys Ser Asp Leu
Glu Glu Ile Pro Glu Ala Glu180 185 190cta
aac cgg gtg cct gct gag ggg ccg gat ttg gtg tct aca tcc tta 624Leu
Asn Arg Val Pro Ala Glu Gly Pro Asp Leu Val Ser Thr Ser Leu195
200 205ggc agc ctg act gca gct cgc cgg gct cag agt
gtg tgg agg cct tgg 672Gly Ser Leu Thr Ala Ala Arg Arg Ala Gln Ser
Val Trp Arg Pro Trp210 215 220tga
67516224PRTMus
musculus 16Met Ala Pro Ser Gln Ala Pro Ser Arg Asp Arg Ala Gly Gln Glu
Asp1 5 10 15Glu Asp Arg
Trp Glu Ala Arg Gly Asp Arg Lys Ala Arg Lys Pro Leu20 25
30Val Glu Lys Lys Arg Arg Ala Arg Ile Asn Glu Ser Leu
Gln Glu Leu35 40 45Arg Leu Leu Leu Ala
Gly Thr Glu Val Gln Ala Lys Leu Glu Asn Ala50 55
60Glu Val Leu Glu Leu Thr Val Arg Arg Val Gln Gly Ala Leu Arg
Gly65 70 75 80Arg Ala
Arg Glu Arg Glu Gln Leu Gln Ala Glu Ala Ser Glu Arg Phe85
90 95Ala Ala Gly Tyr Ile Gln Cys Met His Glu Val His
Thr Phe Val Ser100 105 110Thr Cys Gln Ala
Ile Asp Ala Thr Val Ser Ala Glu Leu Leu Asn His115 120
125Leu Leu Glu Ser Met Pro Leu Arg Glu Gly Ser Ser Phe Gln
Asp Leu130 135 140Leu Gly Asp Ser Leu Ala
Gly Leu Pro Gly Gly Ser Gly Arg Ser Ser145 150
155 160Trp Pro Pro Gly Gly Ser Pro Glu Ser Pro Leu
Ser Ser Pro Pro Gly165 170 175Pro Gly Asp
Asp Leu Cys Ser Asp Leu Glu Glu Ile Pro Glu Ala Glu180
185 190Leu Asn Arg Val Pro Ala Glu Gly Pro Asp Leu Val
Ser Thr Ser Leu195 200 205Gly Ser Leu Thr
Ala Ala Arg Arg Ala Gln Ser Val Trp Arg Pro Trp210 215
22017792PRTArtificial sequenceSynthetic 17Arg Arg Gln His
Gly Gln Leu Trp Phe Pro Glu Gly Phe Lys Val Ser1 5
10 15Glu Ala Ser Lys Lys Lys Arg Arg Glu Pro
Leu Gly Glu Asp Ser Val20 25 30Gly Leu
Lys Pro Leu Lys Asn Ala Ser Asp Gly Ala Leu Met Asp Asp35
40 45Asn Gln Asn Glu Trp Gly Asp Glu Asp Leu Glu Thr
Lys Lys Phe Arg50 55 60Phe Glu Glu Pro
Val Val Leu Pro Asp Leu Ser Asp Gln Thr Asp His65 70
75 80Arg Gln Trp Thr Gln Gln His Leu Asp
Ala Ala Asp Leu Arg Met Ser85 90 95Ala
Met Ala Pro Thr Pro Pro Gln Gly Glu Val Asp Ala Asp Cys Met100
105 110Asp Val Asn Val Arg Gly Pro Asp Gly Phe Thr
Pro Leu Met Ile Ala115 120 125Ser Cys Ser
Gly Gly Gly Leu Glu Thr Gly Asn Ser Glu Glu Glu Glu130
135 140Asp Ala Pro Ala Val Ile Ser Asp Phe Ile Tyr Gln
Gly Ala Ser Leu145 150 155
160His Asn Gln Thr Asp Arg Thr Gly Glu Thr Ala Leu His Leu Ala Ala165
170 175Arg Tyr Ser Arg Ser Asp Ala Ala Lys
Arg Leu Leu Glu Ala Ser Ala180 185 190Asp
Ala Asn Ile Gln Asp Asn Met Gly Arg Thr Pro Leu His Ala Ala195
200 205Val Ser Ala Asp Ala Gln Gly Val Phe Gln Ile
Leu Leu Arg Asn Arg210 215 220Ala Thr Asp
Leu Asp Ala Arg Met His Asp Gly Thr Thr Pro Leu Ile225
230 235 240Leu Ala Ala Arg Leu Ala Leu
Glu Gly Met Leu Glu Asp Leu Ile Asn245 250
255Ser His Ala Asp Val Asn Ala Val Asp Asp Leu Gly Lys Ser Ala Leu260
265 270His Trp Ala Ala Ala Val Asn Asn Val
Asp Ala Ala Val Val Leu Leu275 280 285Lys
Asn Gly Ala Asn Lys Asp Met Gln Asn Asn Lys Glu Glu Thr Pro290
295 300Leu Phe Leu Ala Ala Arg Glu Gly Ser Tyr Glu
Thr Ala Lys Val Leu305 310 315
320Leu Asp His Phe Ala Asn Arg Asp Ile Thr Asp His Met Asp Arg
Leu325 330 335Pro Arg Asp Ile Ala Gln Glu
Arg Met His His Asp Ile Val Arg Leu340 345
350Leu Asp Glu Tyr Asn Leu Val Arg Ser Pro Gln Leu His Gly Thr Ala355
360 365Leu Gly Gly Thr Pro Thr Leu Ser Pro
Thr Leu Cys Ser Pro Asn Gly370 375 380Tyr
Leu Gly Asn Leu Lys Ser Ala Thr Gln Gly Lys Lys Ala Arg Lys385
390 395 400Pro Ser Thr Lys Gly Leu
Ala Cys Gly Ser Lys Glu Ala Lys Asp Leu405 410
415Lys Ala Arg Arg Lys Lys Ser Gln Asp Gly Lys Gly Cys Leu Leu
Asp420 425 430Ser Ser Ser Met Leu Ser Pro
Val Asp Ser Leu Glu Ser Pro His Gly435 440
445Tyr Leu Ser Asp Val Ala Ser Pro Pro Leu Leu Pro Ser Pro Phe Gln450
455 460Gln Ser Pro Ser Met Pro Leu Ser His
Leu Pro Gly Met Pro Asp Thr465 470 475
480His Leu Gly Ile Ser His Leu Asn Val Ala Ala Lys Pro Glu
Met Ala485 490 495Ala Leu Ala Gly Gly Ser
Arg Leu Ala Phe Glu Pro Pro Pro Pro Arg500 505
510Leu Ser His Leu Pro Val Ala Ser Ser Ala Ser Thr Val Leu Ser
Thr515 520 525Asn Gly Thr Gly Ala Met Asn
Phe Thr Val Gly Ala Pro Ala Ser Leu530 535
540Asn Gly Gln Cys Glu Trp Leu Pro Arg Leu Gln Asn Gly Met Val Pro545
550 555 560Ser Gln Tyr Asn
Pro Leu Arg Pro Gly Val Thr Pro Gly Thr Leu Ser565 570
575Thr Gln Ala Ala Gly Leu Gln His Ser Met Met Gly Pro Leu
His Ser580 585 590Ser Leu Ser Thr Asn Thr
Leu Ser Pro Ile Ile Tyr Gln Gly Leu Pro595 600
605Asn Thr Arg Leu Ala Thr Gln Pro His Leu Val Gln Thr Gln Gln
Val610 615 620Gln Pro Gln Asn Leu Gln Leu
Gln Pro Gln Asn Leu Gln Pro Pro Ser625 630
635 640Gln Pro His Leu Ser Val Ser Ser Ala Ala Asn Gly
His Leu Gly Arg645 650 655Ser Phe Leu Ser
Gly Glu Pro Ser Gln Ala Asp Val Gln Pro Leu Gly660 665
670Pro Ser Ser Leu Pro Val His Thr Ile Leu Pro Gln Glu Ser
Gln Ala675 680 685Leu Pro Thr Ser Leu Pro
Ser Ser Met Val Pro Pro Met Thr Thr Thr690 695
700Gln Phe Leu Thr Pro Pro Ser Gln His Ser Tyr Ser Ser Ser Pro
Val705 710 715 720Asp Asn
Thr Pro Ser His Gln Leu Gln Val Pro Glu His Pro Phe Leu725
730 735Thr Pro Ser Pro Glu Ser Pro Asp Gln Trp Ser Ser
Ser Ser Pro His740 745 750Ser Asn Ile Ser
Asp Trp Ser Glu Gly Ile Ser Ser Pro Pro Thr Thr755 760
765Met Pro Ser Gln Ile Thr His Ile Pro Glu Ala Phe Lys Tyr
Gly Arg770 775 780Lys Lys Arg Arg Gln Arg
Arg Arg785 79018797PRTArtificial sequenceSynthetic 18Arg
Arg Gln His Gly Gln Leu Trp Phe Pro Glu Gly Phe Lys Val Ser1
5 10 15Glu Ala Ser Lys Lys Lys Arg
Arg Glu Pro Leu Gly Glu Asp Ser Val20 25
30Gly Leu Lys Pro Leu Lys Asn Ala Ser Asp Gly Ala Leu Met Asp Asp35
40 45Asn Gln Asn Glu Trp Gly Asp Glu Asp Leu
Glu Thr Lys Lys Phe Arg50 55 60Phe Glu
Glu Pro Val Val Leu Pro Asp Leu Ser Asp Gln Thr Asp His65
70 75 80Arg Gln Trp Thr Gln Gln His
Leu Asp Ala Ala Asp Leu Arg Met Ser85 90
95Ala Met Ala Pro Thr Pro Pro Gln Gly Glu Val Asp Ala Asp Cys Met100
105 110Asp Val Asn Val Arg Gly Pro Asp Gly
Phe Thr Pro Leu Met Ile Ala115 120 125Ser
Cys Ser Gly Gly Gly Leu Glu Thr Gly Asn Ser Glu Glu Glu Glu130
135 140Asp Ala Pro Ala Val Ile Ser Asp Phe Ile Tyr
Gln Gly Ala Ser Leu145 150 155
160His Asn Gln Thr Asp Arg Thr Gly Glu Thr Ala Leu His Leu Ala
Ala165 170 175Arg Tyr Ser Arg Ser Asp Ala
Ala Lys Arg Leu Leu Glu Ala Ser Ala180 185
190Asp Ala Asn Ile Gln Asp Asn Met Gly Arg Thr Pro Leu His Ala Ala195
200 205Val Ser Ala Asp Ala Gln Gly Val Phe
Gln Ile Leu Leu Arg Asn Arg210 215 220Ala
Thr Asp Leu Asp Ala Arg Met His Asp Gly Thr Thr Pro Leu Ile225
230 235 240Leu Ala Ala Arg Leu Ala
Leu Glu Gly Met Leu Glu Asp Leu Ile Asn245 250
255Ser His Ala Asp Val Asn Ala Val Asp Asp Leu Gly Lys Ser Ala
Leu260 265 270His Trp Ala Ala Ala Val Asn
Asn Val Asp Ala Ala Val Val Leu Leu275 280
285Lys Asn Gly Ala Asn Lys Asp Met Gln Asn Asn Lys Glu Glu Thr Pro290
295 300Leu Phe Leu Ala Ala Arg Glu Gly Ser
Tyr Glu Thr Ala Lys Val Leu305 310 315
320Leu Asp His Phe Ala Asn Arg Asp Ile Thr Asp His Met Asp
Arg Leu325 330 335Pro Arg Asp Ile Ala Gln
Glu Arg Met His His Asp Ile Val Arg Leu340 345
350Leu Asp Glu Tyr Asn Leu Val Arg Ser Pro Gln Leu His Gly Thr
Ala355 360 365Leu Gly Gly Thr Pro Thr Leu
Ser Pro Thr Leu Cys Ser Pro Asn Gly370 375
380Tyr Leu Gly Asn Leu Lys Ser Ala Thr Gln Gly Lys Lys Ala Arg Lys385
390 395 400Pro Ser Thr Lys
Gly Leu Ala Cys Gly Ser Lys Glu Ala Lys Asp Leu405 410
415Lys Ala Arg Arg Lys Lys Ser Gln Asp Gly Lys Gly Cys Leu
Leu Asp420 425 430Ser Ser Ser Met Leu Ser
Pro Val Asp Ser Leu Glu Ser Pro His Gly435 440
445Tyr Leu Ser Asp Val Ala Ser Pro Pro Leu Leu Pro Ser Pro Phe
Gln450 455 460Gln Ser Pro Ser Met Pro Leu
Ser His Leu Pro Gly Met Pro Asp Thr465 470
475 480His Leu Gly Ile Ser His Leu Asn Val Ala Ala Lys
Pro Glu Met Ala485 490 495Ala Leu Ala Gly
Gly Ser Arg Leu Ala Phe Glu Pro Pro Pro Pro Arg500 505
510Leu Ser His Leu Pro Val Ala Ser Ser Ala Ser Thr Val Leu
Ser Thr515 520 525Asn Gly Thr Gly Ala Met
Asn Phe Thr Val Gly Ala Pro Ala Ser Leu530 535
540Asn Gly Gln Cys Glu Trp Leu Pro Arg Leu Gln Asn Gly Met Val
Pro545 550 555 560Ser Gln
Tyr Asn Pro Leu Arg Pro Gly Val Thr Pro Gly Thr Leu Ser565
570 575Thr Gln Ala Ala Gly Leu Gln His Ser Met Met Gly
Pro Leu His Ser580 585 590Ser Leu Ser Thr
Asn Thr Leu Ser Pro Ile Ile Tyr Gln Gly Leu Pro595 600
605Asn Thr Arg Leu Ala Thr Gln Pro His Leu Val Gln Thr Gln
Gln Val610 615 620Gln Pro Gln Asn Leu Gln
Leu Gln Pro Gln Asn Leu Gln Pro Pro Ser625 630
635 640Gln Pro His Leu Ser Val Ser Ser Ala Ala Asn
Gly His Leu Gly Arg645 650 655Ser Phe Leu
Ser Gly Glu Pro Ser Gln Ala Asp Val Gln Pro Leu Gly660
665 670Pro Ser Ser Leu Pro Val His Thr Ile Leu Pro Gln
Glu Ser Gln Ala675 680 685Leu Pro Thr Ser
Leu Pro Ser Ser Met Val Pro Pro Met Thr Thr Thr690 695
700Gln Phe Leu Thr Pro Pro Ser Gln His Ser Tyr Ser Ser Ser
Pro Val705 710 715 720Asp
Asn Thr Pro Ser His Gln Leu Gln Val Pro Glu His Pro Phe Leu725
730 735Thr Pro Ser Pro Glu Ser Pro Asp Gln Trp Ser
Ser Ser Ser Pro His740 745 750Ser Asn Ile
Ser Asp Trp Ser Glu Gly Ile Ser Ser Pro Pro Thr Thr755
760 765Met Pro Ser Gln Ile Thr His Ile Pro Glu Ala Phe
Lys Arg Gln Ile770 775 780Lys Ile Trp Phe
Gln Asn Arg Arg Met Lys Trp Lys Lys785 790
79519350PRTArtificial sequenceSynthetic 19Tyr Gly Arg Lys Lys Arg Arg
Gln Arg Arg Arg Met Lys Arg Pro Cys1 5 10
15Glu Glu Thr Thr Ser Glu Ser Asp Leu Asp Glu Thr Ile
Asp Val Gly20 25 30Ser Glu Asn Asn Tyr
Pro Gly His Ala Thr Ser Ser Val Met Arg Ser35 40
45Asn Ser Pro Thr Thr Thr Ser Gln Ile Met Ala Arg Lys Lys Arg
Arg50 55 60Gly Ile Ile Glu Lys Arg Arg
Arg Asp Arg Ile Asn Asn Ser Leu Ser65 70
75 80Glu Leu Arg Arg Leu Val Pro Thr Ala Phe Glu Lys
Gln Gly Ser Ala85 90 95Lys Leu Glu Lys
Ala Glu Ile Leu Gln Met Thr Val Asp His Leu Lys100 105
110Met Leu Gln Ala Thr Gly Gly Lys Gly Tyr Phe Asp Ala His
Ala Leu115 120 125Ala Thr Asp Phe Met Ser
Ile Gly Phe Arg Glu Cys Leu Thr Glu Val130 135
140Ala Arg Tyr Leu Ser Ser Val Glu Gly Leu Asp Pro Ser Asp Pro
Leu145 150 155 160Arg Val
Arg Leu Val Ser His Leu Ser Thr Cys Ala Ser Gln Arg Glu165
170 175Ala Ala Val Met Thr Ser Ser Met Ala His His His
His Pro Leu His180 185 190Pro His His Trp
Ala Ala Ala Phe His His Leu Pro Thr Ala Leu Leu195 200
205Gln Pro Asn Gly Leu His Thr Ser Glu Ser Thr Pro Cys Arg
Leu Ser210 215 220Thr Ser Ser Glu Val Pro
Ser Ala His Gly Ser Ala Leu Leu Thr Ala225 230
235 240Thr Phe Ala His Ala Asp Ser Ala Leu Arg Met
Pro Ser Gly Gly Thr245 250 255Val Ala Pro
Cys Val Pro Pro Leu Ser Thr Ser Leu Leu Ser Leu Ser260
265 270Ala Thr Val His Ala Ala Ala Ala Ala Ala Thr Ala
Ala Ala His Ser275 280 285Phe Pro Leu Ser
Phe Ala Gly Ala Phe Pro Met Leu Pro Ser Asn Ala290 295
300Ala Ala Ala Ala Ala Val Ala Ala Ala Thr Ala Ile Ser Pro
Pro Leu305 310 315 320Ser
Val Ser Ala Ala Ser Ser Pro Gln Gln Thr Ser Thr Gly Thr Asn325
330 335Asn Lys Pro Tyr Gln Pro Trp Gly Thr Glu Val
Gly Ala Phe340 345 35020178PRTArtificial
SequenceSynthetic 20Tyr Gly Arg Lys Lys Arg Arg Gln Arg Arg Arg Met Ala
Pro Ser Thr1 5 10 15Val
Ala Val Glu Met Leu Ser Pro Lys Glu Lys Asn Arg Leu Arg Lys20
25 30Pro Val Val Glu Lys Met Arg Arg Asp Arg Ile
Asn Ser Ser Ile Glu35 40 45Gln Leu Lys
Leu Leu Leu Glu Gln Glu Phe Ala Arg His Gln Pro Asn50 55
60Ser Lys Leu Glu Lys Ala Asp Ile Leu Glu Met Ala Val
Ser Tyr Leu65 70 75
80Lys His Ser Lys Ala Phe Ala Ala Ala Ala Gly Pro Lys Ser Leu His85
90 95Gln Asp Tyr Ser Glu Gly Tyr Ser Trp Cys
Leu Gln Glu Ala Val Gln100 105 110Phe Leu
Thr Leu His Ala Ala Ser Asp Thr Gln Met Lys Leu Leu Tyr115
120 125His Phe Gln Arg Pro Pro Ala Pro Ala Ala Pro Ala
Lys Glu Pro Pro130 135 140Ala Pro Gly Ala
Ala Pro Gln Pro Ala Arg Ser Ser Ala Lys Ala Ala145 150
155 160Ala Ala Ala Val Ser Thr Ser Arg Gln
Pro Ala Cys Gly Leu Trp Arg165 170 175Pro
Trp
User Contributions:
comments("1"); ?> comment_form("1"); ?>Inventors list |
Agents list |
Assignees list |
List by place |
Classification tree browser |
Top 100 Inventors |
Top 100 Agents |
Top 100 Assignees |
Usenet FAQ Index |
Documents |
Other FAQs |
User Contributions:
Comment about this patent or add new information about this topic:
| People who visited this patent also read: | |
| Patent application number | Title |
|---|---|
| 20100035260 | Compositions, devices, systems, for using a Nanopore |
| 20100035259 | BIOMARKERS AND METHODS FOR DETERMINING SENSITIVITY TO VASCULAR ENDOTHELIAL GROWTH FACTOR RECEPTOR-2 MODULATORS |
| 20100035258 | ASSAY AND METHOD |
| 20100035257 | METHODS TO DETERMINE THE RESPONSIVENESS TO CISPLATIN TREATMENT |
| 20100035256 | ENDURACIDIN BIOSYNTHETIC GENE CLUSTER FROM STREPTOMYCES FUNGICIDICUS |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
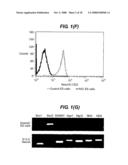 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
| Similar patent applications: | |
| Date | Title |
|---|---|
| 2010-06-24 | Small molecule compounds for stem cell differentiation |
| 2009-10-15 | Cell cycle regulation and differentiation |
| 2010-04-22 | Recovery of volatile products from fermentation broth |
| 2010-05-13 | Method for removing bacteria from a fermentation broth |
| 2010-06-03 | Cytokines and genes differentially affected by tnf blockers |
| New patent applications from these inventors: | |
| Date | Title |
|---|---|
| 2014-12-18 | Neural stem cells |
| Top Inventors for class "Chemistry: molecular biology and microbiology" | |
| Rank | Inventor's name |
|---|---|
| 1 | Marshall Medoff |
| 2 | Anthony P. Burgard |
| 3 | Mark J. Burk |
| 4 | Robin E. Osterhout |
| 5 | Rangarajan Sampath |