Patent application title: Allergy Inhibitor Compositions And Kits And Methods Of Using The Same
Inventors:
IPC8 Class: AA61K3935FI
USPC Class:
1 1
Class name:
Publication date: 2017-04-27
Patent application number: 20170112918
Abstract:
Compositions, methods, and kits for inhibiting an allergic response
against an allergenic protein are disclosed. Compositions, methods and
kits for inhibiting an allergic response against a flea allergenic
protein; a feline allergenic protein; a canine allergenic protein; a dust
mite allergenic protein; a peanut allergenic protein; a Japanese cedar
allergenic protein; and a blomia tropicalis allergenic protein are
disclosed.Claims:
1.-31. (canceled)
32. A therapeutic composition for inhibiting an allergic response, the composition comprising: (a) an eukaryotic expression vector comprising a nucleotide sequence encoding an allergenic protein that comprises an antigen epitope; and (b) an allergenic protein that comprises an antigenic epitope, wherein the allergenic protein is selected from the group consisting of: feline allergenic protein, canine allergenic protein, dust mite allergenic protein, Japanese cedar allergenic protein, Blomia tropicalis allergenic protein, and flea salivary allergenic protein.
33. The therapeutic composition of claim 32, wherein the nucleotide sequence encodes an amino acid sequence selected from the group consisting of: SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8, SEQ ID NO:10, SEQ ID NO:12, SEQ ID NO:15, SEQ ID NO:17, SEQ ID NO:19, SEQ ID NO:21, SEQ ID NO:22, and SEQ ID NO:23.
34. The therapeutic composition of claim 32, wherein the nucleotide sequence comprises a nucleic acid sequence selected from the group consisting of: SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7, SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:14, SEQ ID NO:16, SEQ ID NO:18, SEQ ID NO:20, SEQ ID NO:24, and SEQ ID NO:25.
35. The therapeutic composition of claim 32, wherein the nucleotide sequence is operably linked to a promoter selected from the group consisting of: RSV promoter, CMV promoter, and SV40 promoter.
36. The therapeutic composition of claim 32, wherein a ratio of the eukaryotic expression vector by weight to the allergenic protein by weight is between 1:5 and 5:1.
37. The therapeutic composition of claim 36, wherein the ratio of the eukaryotic expression vector by weight to the allergenic protein by weight is 1:1.
38. The therapeutic composition of claim 32, wherein a molar ratio of the eukaryotic expression vector to the allergenic protein is between 1:100,000 to 20:100,000.
39. The therapeutic composition of claim 7, wherein the molar ratio of the eukaryotic expression vector to the allergenic protein is 15:100,000.
40. A method of inhibiting an allergenic reaction in a subject in need thereof, the method comprising administering the therapeutic composition of claim 32 to the subject.
41. The method of claim 40, wherein the eukaryotic expression vector and the allergenic protein are administered together.
42. The method of claim 40, wherein the eukaryotic expression vector and the allergenic protein are administered separately.
43. A method of inducing CD4.sup.+CD25.sup.- Tr cells in a subject in need thereof, the method comprising administering the therapeutic composition of claim 32 to the subject.
44. A kit for inhibiting an allergenic response in a subject in need thereof, the kit comprising: (a) a first container comprising an eukaryotic expression vector comprising a nucleotide sequence encoding an allergenic protein that comprises an antigenic epitope; (b) a second container comprising an allergenic protein that comprises an antigenic epitope, wherein the allergenic protein is selected from the group consisting of: feline allergenic protein, canine allergenic protein, dust mite allergenic protein, Japanese cedar allergenic protein, Blomia tropicalis allergenic protein, and flea salivary allergenic protein.
45. The kit of claim 44, wherein the nucleotide sequence encodes an amino acid sequence selected from the group consisting of: SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8, SEQ ID NO:10, SEQ ID NO:12, SEQ ID NO:15, SEQ ID NO:17, SEQ ID NO:19, SEQ ID NO:21, SEQ ID NO:22, and SEQ ID NO:23.
46. The kit of claim 44, wherein the nucleotide sequence comprises a nucleic acid sequence selected from the group consisting of: SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7, SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:14, SEQ ID NO:16, SEQ ID NO:18, SEQ ID NO:20, SEQ ID NO:24, and SEQ ID NO:25.
47. The kit of claim 44, wherein the nucleotide sequence is operably linked to a promoter selected from the group consisting of: RSV promoter, CMV promoter, and SV40 promoter.
48. The kit of claim 44, wherein a ratio of the eukaryotic expression vector by weight to the allergenic protein by weight is between 1:5 and 5:1.
49. The kit of claim 48, wherein the ratio of the eukaryotic expression vector by weight to the allergenic protein by weight is 1:1.
50. The kit of claim 44, wherein a molar ratio of the eukaryotic expression vector to the allergenic protein is between 1:100,000 to 20:100,000.
51. The kit of claim 50, wherein the molar ratio of the eukaryotic expression vector to the allergenic protein is 15:100,000.
Description:
CROSS REFERENCE TO RELATED APPLICATIONS
[0001] This application claims priority under 35 U.S.C. .sctn.120 as a continuation of U.S. Ser. No. 13/721,449, filed Dec. 20, 2012, allowed, which is a continuation of U.S. Ser. No. 11/644,435 filed Dec. 22, 2006, which issued as U.S. Pat. No. 8,349,333 on Jan. 8, 2013, which claims priority to Chinese application number 2005 10132381.X filed Dec. 23, 2005, each of which is incorporated herein by reference.
FIELD OF THE INVENTION
[0002] The present invention relates to compositions and kits which are useful to prevent and inhibit allergic reactions against allergens, and to methods of using such compositions and kits. The present invention provides compositions, kits and methods for preventing and inhibiting flea allergy dermatitis, allergic reactions to cat and canine fur or danders, dust mites, peanuts, Japanese cedar pollen and blomia tropicalis allergen.
BACKGROUND OF THE INVENTION
[0003] Allergic reactions to various allergens represent significant health concerns, particularly in instances in which the allergic reaction induces severe reactions and/or allergen induced immediate hypersensitivity (AIH).
[0004] Allergy is considered as the consequence of persistent T cell activation driving pathogenic inflammation against host dermis by specific allergens. Several approaches are used to ameliorate AIH and these include nonspecific immunosuppressive drugs or monoclonal antibodies targeted to T or B cells (A. J. Van Oosterhout et al., Am. J. Respir. Cell Mol. Biol. 17, 386 (Sep. 1, 1997); P. Proksch et al, J Immunol 174, 7075 (Jun. 1, 2005)). However, this situation is compromised as long term treated recipients can become generally compromised in their ability to fight infections. Redirecting immunity from Th2 type to Th1 type has also been demonstrated with limited success (S. Jilek, C. Barbey, F. Spertini, B. Corthesy, J Immunol 166, 3612 (Mar. 1, 2001)). A recent discovery of T regulatory cells, including the naturally occurring thymus derived CD4+CD25.sup.+ Treg cells (I. M. de Kleer et al, J Immunol 111, 6435 (May 15, 2004); D. Lundsgaard, T. L. Holm, L. Hornum, H. Markholst, Diabetes 54, 1040 (Apr. 1, 2005); M. J. McGeachy, L. A. Stephens, S. M. Anderton, J Immunol 175, 3025 (Sep. 1, 2005); I. Bellinghausen, B. Klostermann, J. Knop, J. Saloga, J Allergy Clin Immunol 111, 862 (Apr. 1, 2003); E. M. Ling et al, Lancet 363, 608 (Feb. 21, 2004); and J. Kearley, J. E. Barker, D. S. Robinson, C. M. Lloyd, J. Exp. Med., jem.20051166 (Nov. 28, 2005)), mucosal induced Th3 cells and antigen induced CD4.sup.+CD25'' Tr cells have been proposed to be use as immuno-regulators or suppressors or auto-reactive pathogenesis (H. Fukaura et al, J. Clin. Invest. 98, 70 (Jul. 1, 1996)). Various approaches have been explored to induce T regulatory cells to constrain the auto-reactive T cells. Preferentially, induction of antigen specific T regulatory cells targeted to allergy, asthma and autoimmune disease antigens are considered a promising strategy. Several lines of evidence have indicated that induction of antigen specific regulatory T cell 1 (TO) is possible via utility of immatured DCs, suboptimal immunogens or partial blocking the co-stimulatory molecules in DCs (A. Kumanogoh et al., J Immunol 166, 353 (Jan. 1, 2001); M. K. Levings et al, Blood 105, 1162 (Feb. 1, 2005); and S. K. Seo et al, Nat Med 10, 1088 (Oct. 1, 2004)). All these approaches are done either in vitro or in experimental conditions. Induction of Tr cells that can inhibit antigen specific T cells' function in vivo by co-inoculating antigen-matched DNA and protein antigens as co-administered vaccines (H. Jin et al, Virology 337, 1 83 (Jun. 20, 2005)).
[0005] The chief characteristic of the non-host flea is that it is a hematophagic parasite that may be found in the body of any mammalian or avian species of animal. Ctenocephalides felis is a parasite that occurs mainly in cats and dogs, while Ctenocephalides canis is limited to domestic dogs and feral dogs. Flea allergy dermatitis (FAD) is the most frequently seen skin ailment in cats and dogs. FAD results when a flea parasite bites and its saliva serves as an irritant and elicits an allergic reaction. The location of the bite appears red, swollen, irritated and itching. Often the animal will scratch at the bite with its paws, causing the wound to turn into a skin ulceration and eliciting further bacterial and fungal infections. This poses a great danger for the dog or cat and at present no effective pharmacotherapeutic or preventive methods exist for this disease.
[0006] In general, flea allergen refers to the various differently sized proteins from flea antigens that cause an allergic reaction. In some parts of the literature it is referred to as feline flea saliva allergenic protein FSA1 or Cte f 1. GeneBank AF102502, which is incorporated herein by reference, discloses the nucleotide sequences (SEQ ID NO:1) encoding the FSA1 or Cte f 1 protein derived from the flea salivary gland of the Ctenocephalides felis. The 653 nucleotide sequence includes coding sequences 1-531 which include coding sequences for the signal peptide (1-54) and mature protein sequence (55-528). GeneBank AAD17905, which is incorporated herein by reference, discloses the amino sequences (SEQ ID NO:2) of the FSA1 or Cte f 1 protein derived from the flea salivary gland of the Ctenocephalides felis. including the signal peptide (1-18) and mature protein sequences (19-176).
[0007] The chief feline allergenic protein is Fel dI. GeneBank M74953, which is incorporated herein by reference, discloses the amino acid of and nucleotide sequences (SEQ ID NO:3) encoding the Fel dI protein derived from the major allergen of the domestic cat. It possesses the secondary B secretion peptide sequence. This Fel d I sequence is 416 bp mRNA including the 5' untranslated region made up of sequences 1-25 and the coding sequence being sequences 26-292 encoding 88 amino acids (SEQ ID NO:4; GeneBank AAC41617, which is incorporated herein by reference). The signal peptide is encoded by 26-79 and the mature protein is encoded by 80-289. The 3' untranslated region is 293-416. GeneBank M74952, which is incorporated herein by reference, discloses the amino acid of and nucleotide sequences (SEQ ID NO:5) encoding the Fel dI protein derived from the major allergen of the domestic cat. This Fel dI sequence is 410 bp mRNA including the 5' untranslated region made up of sequences 1-7 and the coding sequence being sequences 8-286 encoding 92 amino acids (SEQ ID NO:6; GeneBank AAC37318, which is incorporated herein by reference). The signal peptide is encoded by 8-73 and the mature protein is encoded by 74-283. The 3' untranslated region is 287-410.
[0008] The chief canine allergenic proteins are the salivary lipid promoters Can f1 and Can f2. GeneBank AF027177, which is incorporated herein by reference, discloses the amino acid of and nucleotide sequences (SEQ ID NO:7) encoding the Can f1 protein derived from the salivary lipocalin proteins of the major allergen of the domestic dog. This Can f1 sequence is 525 bp mRNA encoding 174 amino acids (SEQ ID NO:8; GeneBank AAC48794, which is incorporated herein by reference).
[0009] GeneBank AF027178, which is incorporated herein by reference, discloses the amino acid of and nucleotide sequences (SEQ ID NO:9) encoding the Can f2 protein derived from the salivary lipocalin proteins of the major allergen of the domestic dog. This Can f2 sequence is 791 bp mRNA including a coding sequence of 195-737 encoding 180 amino acids (SEQ ID NO: 10; GeneBank AAC48795, which is incorporated herein by reference).
[0010] GeneBank Ul 1695, which is incorporated herein by reference, discloses the amino acid of and nucleotide sequences (SEQ ID NO:11) encoding the dust mite allergy source protein antigen Der P 1. This Der P 1 sequence is 1099 bp mRNA including a coding sequence of 50-1012 encoding 180 amino acids (SEQ ID NO: 12; GeneBank AAB60125, which is incorporated herein by reference). The coding sequence includes coding sequences 50-109 which encode a signal peptide and coding sequences 344-1009 which encodes the mature peptide. GeneBank AAB60125 discloses a signal peptide that includes amino acids 1-20 and a mature protein that includes amino acids 99-320.
[0011] GeneBank L77197, which is incorporated herein by reference, discloses the amino acid of and nucleotide sequences (SEQ ID NO: 13) encoding the peanut allergy source protein antigen Ara h II. This Ara h II sequence is 717 bp sequence encoding 110 amino acids and including a polyA signal 562-567.
[0012] GeneBank AF059616, which is incorporated herein by reference, discloses the amino acid of and nucleotide sequences (SEQ ID NO:14) encoding the peanut allergy source protein antigen Ara h II. This Ara h 5 sequence is 743 bp sequence including a coding sequence of 17-412. GeneBank AAD55587, which is incorporated herein by reference, discloses the 131 amino acid protein (SEQ ID NO: 15).
[0013] GeneBank AB081309, which is incorporated herein by reference, discloses the amino acid of and nucleotide sequences (SEQ ID NO: 16) encoding the Japanese cedar (cryptomeria japonicd) allergy source antigen Cry j 1.1. This Cryj 1.1 sequence is 1295 bp sequence including a coding sequence of 62-1186 in which a signal peptide is encoded by 62-124 and the mature protein is encoded by 125-1183 and a polyA site at 1295. GeneBank BAB86286, which is incorporated herein by reference, discloses the 374 amino acid protein (SEQ ID NO: 17) including a signal peptide of amino acids 1-21 and a mature protein of amino acids 22-374.
[0014] GeneBank AB081310, which is incorporated herein by reference, discloses the amino acid of and nucleotide sequences (SEQ ID NO: 18) encoding the Japanese cedar (cryptomeria japonicci) allergy source antigen Cry j 1.2. This Cry j 1.2 sequence is 1313 bp sequence including a coding sequence of 46-1170 in which a signal peptide is encoded by 46-108 and the mature protein is encoded by 109-1167 and a polyA site at 1313. GeneBank BAB86287, which is incorporated herein by reference, discloses the 374 amino acid protein (SEQ ID NO: 19) including a signal peptide of amino acids 1-21 and a mature protein of amino acids 22-374.
[0015] GeneBank U59102, which is incorporated herein by reference, discloses the amino acid of and nucleotide sequences (SEQ ID NO:20) encoding the blomia tropicalis allergy source protein antigen Blo t 5. This Blo t 5 sequence is 537 bp sequence including a coding sequence of 33-437. GeneBank AAD10850, which is incorporated herein by reference, discloses the 134 amino acid protein (SEQ ID NO:21).
[0016] There remains a need for compositions and methods of preventing and inhibiting the allergic reactions induced by these allergens.
SUMMARY OF THE INVENTION
[0017] The present invention relates to compositions for preventing and inhibiting an allergic response against an allergenic protein. The compositions comprise:
[0018] (a) an eukaryotic cell expression vector containing nucleotide sequences encoding an allergenic protein or a polypeptide that comprises an antigenic epitope of said allergenic protein; and,
[0019] (b) an allergenic protein or a polypeptide that comprises an antigenic epitope of said allergenic protein.
[0020] The present invention provides compositions for preventing and inhibiting an allergic response against an allergenic protein selected from the group consisting of: a flea allergenic protein; a feline allergenic protein; a canine allergenic protein; a dust mite allergenic protein; a peanut allergenic protein; a Japanese cedar allergenic protein; and a blomia tropicalis allergenic protein.
[0021] The present invention further relates to kits for preventing and inhibiting an allergic response against an allergenic protein. The kits comprise:
[0022] (a) a first container comprising a eukaryotic cell expression vector containing nucleotide sequences encoding an allergenic protein or a polypeptide that comprises an antigenic epitope of said allergenic protein; and
[0023] (b) a second container an allergenic protein or a polypeptide that comprises an antigenic epitope of said allergenic protein.
[0024] The present invention provides kits for preventing and inhibiting an allergic response against an allergenic protein selected from the group consisting of: a flea allergenic protein; a feline allergenic protein; a canine allergenic protein; a dust mite allergenic protein; a peanut allergenic protein; a Japanese cedar allergenic protein; and a blomia tropicalis allergenic protein.
[0025] The present invention further relates to methods of preventing and inhibiting an allergic reaction to an allergenic protein in an individual. The methods comprise the step or steps of administering to the individual
[0026] (a) an eukaryotic cell expression vector containing nucleotide sequences encoding an allergenic protein or a polypeptide that comprises an antigenic epitope of said allergenic protein; and
[0027] (b) an allergenic protein or a polypeptide that comprises an antigenic epitope of said allergenic protein.
[0028] The present invention provides of preventing and inhibiting an allergic reaction to an allergenic protein in an individual wherein said allergenic protein is selected from the group consisting of: a flea allergenic protein; a dust mite allergenic protein; a peanut allergenic protein; a Japanese cedar allergenic protein; and a blomia tropicalis allergenic protein.
BRIEF DESCRIPTION OF THE FIGURES
[0029] FIG. 1A, FIG. 1B, and FIG. 1C refer to a Flea allergy model in mice. C57b/6 mice were primed twice biweekly with flea antigens or saline as a negative control, challenged with the flea antigens, or PBS as the negative control, histamine as the positive control intradermally. In FIG. 1A, the local reactions after the skin test were measured at 30 min after the challenge. FIG. 1B shows anti-flea antigens of IgE production. In FIG. 1C, CD4+ T cell proliferation responses stimulated by flea antigens in vitro, were tested in mice. Results are representative of at least three experiments. * P<0.05, compared with naive control groups as indicated.
[0030] FIG. 2A, FIG. 2B, FIG. 2C, and FIG. 2D show that co-immunization of DNA and protein suppresses the development of immediate hyper-sensitivity reaction. FIG. 2A shows data from skin tests of mice after co-immunization with F or pcDF100+F. FIG. 2B shows that dose dependent skin responses in mice after co-immunization with pcDF100+F is displayed. FIG. 2C shows anti-flea antigen levels of IgE and IgG1 after induction and treatment. FIG. 2D shows CD4+ T cell proliferation responses stimulated by flea antigen-specific in vitro. Results are representative of at least three experiments. * P<0.05, compared with V+F and F vaccination groups as indicated.
[0031] FIG. 3A, FIG. 3B, and FIG. 3C show that CD4.sup.+CD25'' T cells are responsible for the observed suppression. In FIG. 3A, 5.times.10.sup.5 of CD3.sup.+ T cells were isolated from the spleens of flea antigen immunized mice and added to 96-well-plates. At the same time, 1.times.10.sup.5 of splenocytes from naive, F, V+F and pcDF100+F immunized mice were also added to the same plate. Similarly, 1.times.10.sup.5 non-T cells or T cells, purified CD8.sup.+, CD4.sup.+ or CD4.sup.+CD25'' T cells were isolated from the spleens of V+F or pcDF100+F immunized mice. These Co-cultures were stimulated with flea antigen (50 ug/ml) in the presence of 1.times.10.sup.5 bone marrow derived DCs for 48 h in vitro. Proliferation was examined by MTS-PMS (Promega) according to manufactors instructions and stimulation index (SI) was determined by the formula: counts of flea-antigen stimulated/counts of non-stimulated cultures). In FIG. 3B, 1.times.10.sup.6 of splenocytes from naive, F, V+F and pcDF100+F immunized mice were adoptively transferred into naive C57 mice. Similarly, 1.times.10.sup.6 non-T cells or T cells, 1.times.10.sup.6 purified CD8.sup.+, CD4.sup.+, 5.times.10.sup.5 CD4.sup.+CD25'' and CD4.sup.+CD25.sup.+T cells were isolated from spleens of pcDF100+F, V+F, F immunized or naive control mice and were adoptively transferred into syngeneic flea-antigen primed mice and skin test responses were examined. FIG. 3C shows that co-administration of DNA and protein induce antigen-specific suppression. I.times.10.sup.6CD4.sup.+CD25 T cells were isolated from spleens of pcDF100+F, V+F immunized or naive control mice and were adoptively transferred into naive mice, which were then immunized with specific Flea antigen or non-specific OVA protein 24 hours after transfer, then CD4.sup.+ T cells were isolated and their proliferation was analyzed. Results shown in the figure are representative of two experiments. * P<0.05 compared with V+F and F transfers as indicated.
[0032] FIG. 4A, FIG. 4B, and FIG. 4C show that DCs from pcDF100+F co-immunized mice induce Trcells in vitro. FIG. 4A shows pcDF100+F co-immunization restricted MLR stimulatory activities on DCs. 48 h after immunization, DCs isolated from spleen of mice were used to stimulate T cell proliferation in MLR. T cell proliferation was measured by CFSE activity. Results are representative of one of two respective experiments. * P<0.05 compared with V+F and F vaccination groups as indicated. FIG. 4B shows data from DCs isolated from spleens of pcDF100+F.sub.7 V+F, F immunized or naive control mice co-cultured with naive CD4.sup.+ T cells. The T cells were restimulated once for two days for 3 cycles, using fresh DCs, and were then analyzed after each stimulation cycle for IL-10, IL-4 and IFN-y positive cell numbers, and for the ability to regulated MLR stimulatory activities. In FIG. 4C, MLR was performed with APC from C57 mice and T cells from Balb/c mice. T cell proliferation was measured by CFSE. Results are representative of two individual experiments. * P<0.05 compared with V+F and F vaccination groups as indicated.
[0033] FIG. 5A, FIG. 5B, and FIG. 5C show dermatological scores data.
[0034] FIG. 6 shows results of skin tests before co-immunization.
[0035] FIG. 7 shows results of skin tests after co-immunization.
[0036] FIG. 8 shows dermatological scores data after co-immunization.
[0037] FIG. 9A, FIG. 9B, FIG. 9C, FIG. 9D, FIG. 9E, and FIG. 9F show data demonstrating therapeutic effects of co-immunization on the FAD cats.
[0038] FIG. 10A, FIG. 10B, and FIG. 10C show data demonstrating therapeutic effects of co-immunization on the FAD cats.
[0039] FIG. 11 shows a map of plasmid pVAX1-K-FSA1.
DESCRIPTION OF PREFERRED EMBODIMENTS OF THE INVENTION
[0040] The present invention provides compositions, kits and methods which prevent and inhibit allergic reactions, and allergen induced immediate hypersensitivity. The present invention provides compositions, kits and methods which prevent and inhibit flea allergy dermatitis, compositions, kits and methods which prevent and inhibit feline allergy, compositions, kits and methods which prevent and inhibit canine allergy, compositions, kits and methods which prevent and inhibit mite allergy, compositions, kits and methods which prevent and inhibit peanut allergy, compositions, kits and methods which prevent and inhibit Japanese cedar allergy and compositions, kits and methods which prevent and inhibit blomia tropicalis allergy.
[0041] The compositions of the invention comprise an allergenic protein or a peptide or protein which comprises an antigenic epitope of the allergenic protein and an expression vector which encodes an allergenic protein or a peptide or protein which comprises an antigenic epitope of the allergenic protein.
[0042] The kits of the invention comprise a container that comprises an allergenic protein or a peptide or protein which comprises an antigenic epitope of the allergenic protein and container that comprises an expression vector which encodes an allergenic protein or a peptide or protein which comprises an antigenic epitope of the allergenic protein.
[0043] The methods of the invention comprise administering the compositions of the invention and/or the components of a kit of the invention in combination to an individual who has or is susceptible to allergic reactions, or allergen induced immediate hypersensitivity.
[0044] The allergenic protein or a peptide or protein which comprises an antigenic epitope of the allergenic protein present in the composition or kit and used in the method, and allergenic protein or a peptide or protein which comprises an antigenic epitope of the allergenic protein encoded by the expression vector present in the composition or kit and used in the method have amino acid sequence overlap such that they share epitopes, i.e at least one epitope of the allergenic protein or a peptide or protein which comprises an antigenic epitope of the allergenic protein present in the composition or kit and used in the method is the same as at least one epitope of the allergenic protein or a peptide or protein which comprises an antigenic epitope of the allergenic protein encoded by the expression vector present in the composition or kit and used in the method. In some embodiments, the allergenic protein or a peptide or protein which comprises an antigenic epitope of the allergenic protein present in the composition or kit and used in the method is the same as the allergenic protein or a peptide or protein which comprises an antigenic epitope of the allergenic protein encoded by the expression vector present in the composition or kit and used in the method. In some embodiments, the allergenic protein or a peptide or protein which comprises an antigenic epitope of the allergenic protein present in the composition or kit and used in the method is a fragment of the allergenic protein or a peptide or protein which comprises an antigenic epitope of the allergenic protein encoded by the expression vector present in the composition or kit and used in the method. In some embodiments, the allergenic protein or a peptide or protein which comprises an antigenic epitope of the allergenic protein encoded by the expression vector present in the composition or kit and used in the method is a fragment of the allergenic protein or a peptide or protein which comprises an antigenic epitope of the allergenic protein present in the composition or kit and used in the method. In some embodiments, the allergenic protein or a peptide or protein which comprises an antigenic epitope of the allergenic protein present in the composition or kit and used in the method is a fragment of the allergenic protein or a peptide or protein which comprises an antigenic epitope of the allergenic protein encoded by the expression vector present in the composition or kit and used in the method. In some embodiments, one or both of 1) the allergenic protein or a peptide or protein which comprises an antigenic epitope of the allergenic protein present in the composition or kit and used in the method and 2) the allergenic protein or a peptide or protein which comprises an antigenic epitope of the allergenic protein encoded by the expression vector present in the composition or kit and used in the method is identical to a naturally occurring protein which is an allergen. In some embodiments, both of 1) the allergenic protein or a peptide or protein which comprises an antigenic epitope of the allergenic protein present in the composition or kit and used in the method and 2) the allergenic protein or a peptide or protein which comprises an antigenic epitope of the allergenic protein encoded by the expression vector present in the composition or kit and used in the method are identical to a naturally occurring protein which is an allergen. In some embodiments, one or both of 1) the allergenic protein or a peptide or protein which comprises an antigenic epitope of the allergenic protein present in the composition or kit and used in the method and 2) the allergenic protein or a peptide or protein which comprises an antigenic epitope of the allergenic protein encoded by the expression vector present in the composition or kit and used in the method is identical to a fragment of a naturally occurring protein which is an allergen. In some embodiments, both of 1) the allergenic protein or a peptide or protein which comprises an antigenic epitope of the allergenic protein present in the composition or kit and used in the method and 2) the allergenic protein or a peptide or protein which comprises an antigenic epitope of the allergenic protein encoded by the expression vector present in the composition or kit and used in the method are identical to a fragment of a naturally occurring protein which is an allergen. In some embodiments, the allergenic protein or a peptide or protein which comprises an antigenic epitope of the allergenic protein present in the composition or kit and used in the method is identical to a fragment of a naturally occurring protein which is an allergen and the allergenic protein or a peptide or protein which comprises an antigenic epitope of the allergenic protein encoded by the expression vector present in the composition or kit and used in the method is identical to a naturally occurring protein which is an allergen. In some embodiments, the allergenic protein or a peptide or protein which comprises an antigenic epitope of the allergenic protein present in the composition or kit and used in the method is identical to a naturally occurring protein which is an allergen and the allergenic protein or a peptide or protein which comprises an antigenic epitope of the allergenic protein encoded by the expression vector present in the composition or kit and used in the method is identical to a fragment of naturally occurring protein which is an allergen.
[0045] In some embodiments, the composition or kit includes an allergenic protein such as a protein from a pathogen, food, environmental factor or irritant. In some embodiments, the composition or kit includes a peptide or protein which includes an antigenic epitope of an allergenic protein such as a protein from a pathogen, food, environmental factors or irritant. Similarly, in some embodiments, the composition or kit includes an expression vector which encodes an allergenic protein such as a protein from a pathogen, food or irritant and in some embodiments, the composition or kit includes an expression vector which encodes a peptide or protein which includes an antigenic epitope of an allergenic protein such as a protein from a pathogen, food, environmental factor or irritant.
[0046] In some embodiments, an allergenic protein or peptide or protein which includes an antigenic epitope of an allergenic protein that is encoded by the expression vector is identical to the allergenic protein or peptide or protein which includes an antigenic epitope of an allergenic protein included in the composition or kit. In some embodiments, an allergenic protein or peptide or protein which includes an antigenic epitope of an allergenic protein that is encoded by the expression vector is different from the allergenic protein or peptide or protein which includes an antigenic epitope of an allergenic protein included in the composition or kit. In some embodiments, the peptide or protein included in the composition is the allergenic protein. In some embodiments, the peptide or protein included in the composition is a fragment of the allergenic protein. In some embodiments, the peptide or protein encoded by the expression vector is the allergenic protein. In some embodiments, the peptide or protein encoded by the expression vector is a fragment of the allergenic protein. According to the invention, the methods comprise administering the compositions in amounts sufficient to suppress the allergic reaction against the allergenic protein when the individual is subsequently exposed to such protein.
[0047] In some embodiments, the present invention provides inhibitors for flea allergy dermatitis. The flea allergy dermatitis inhibitor of the present invention comprises a eukaryotic cell expression vector containing flea salivary allergenic protein (such as felis salivary antigen 1 (FSA1 or Cte f1)) or a peptide or protein that comprises an antigenic epitope of such allergenic protein, in combination with a flea salivary allergenic protein (such as felis salivary antigen 1 (FSA1 or Cte f1)) or a peptide or protein that comprises an antigenic epitope of such allergenic protein.
[0048] In some embodiments, the present invention provides inhibitors for feline allergy. The feline allergy inhibitor of the present invention comprises a eukaryotic cell expression vector containing feline allergenic protein (such as Fel dI) or a peptide or protein that comprises an antigenic epitope of such allergenic protein, in combination with a feline allergenic protein (such as Fel dI) or a peptide or protein that comprises an antigenic epitope of such allergenic protein.
[0049] In some embodiments, the present invention provides inhibitors for canine allergy. The canine allergy inhibitor of the present invention comprises a eukaryotic cell expression vector containing canine allergenic protein (such as Can f1 or Can f2) or a peptide or protein that comprises an antigenic epitope of such allergenic protein, in combination with a canine allergenic protein (such as Can f1 or Can f2) or a peptide or protein that comprises an antigenic epitope of such allergenic protein.
[0050] In some embodiments, the present invention provides inhibitors for dust mite allergy. The dust mite allergy inhibitor of the present invention comprises a eukaryotic cell expression vector containing a dust mite allergy allergenic protein (such as Der PI or Der F1) or a peptide or protein that comprises an antigenic epitope of such allergenic protein, in combination with a mite allergy allergenic protein (such as Der PI or Der F1) or a peptide or protein that comprises an antigenic epitope of such allergenic protein.
[0051] In some embodiments, the present invention provides inhibitors for peanut allergy. The peanut allergy inhibitor of the present invention comprises a eukaryotic cell expression vector containing a peanut allergy allergenic protein (such as Ara HII or Ara H5) or a peptide or protein that comprises an antigenic epitope of such allergenic protein, in combination with a peanut allergy allergenic protein (such as Ara HII or Ara H5) or a peptide or protein that comprises an antigenic epitope of such allergenic protein.
[0052] In some embodiments, the present invention provides inhibitors for Japanese cedar allergy. The Japanese cedar allergy inhibitor of the present invention comprises a eukaryotic cell expression vector containing a Japanese cedar allergy allergenic protein (such as Cry j 1.1 or Cry j 1.2) or a peptide or protein that comprises an antigenic epitope of such allergenic protein, in combination with a Japanese cedar allergy allergenic protein (such as Cry j 1.1 or Cry j 1.2) or a peptide or protein that comprises an antigenic epitope of such allergenic protein.
[0053] In some embodiments, the present invention provides inhibitors for blomia tropicalis allergy. The blomia tropicalis allergy inhibitor of the present invention comprises a eukaryotic cell expression vector containing a blomia tropicalis allergy allergenic protein (such as Blo t5) or a peptide or protein that comprises an antigenic epitope of such allergenic protein, in combination with a blomia tropicalis allergy allergenic protein (such as Blo t5) or a peptide or protein that comprises an antigenic epitope of such allergenic protein.
[0054] The allergenic protein may be expressed in Escherichia coli or eukaryotic cells (for example, yeast or CHO cells), for example, molecular cloning methodology is used to incorporate the allergenic protein coding sequence into the corresponding expression vector, causing the protein product to be expressed through the Escherichia coli, yeast or CHO cell systems. Purification is then used to obtain the allergenic protein. Similarly, peptides or proteins may be designed which include antigenic epitopes of allergenic proteins. Nucleic acid sequences encoding such peptides or proteins can be incorporated into expression vectors and produced in host cells where they express the peptide or protein which is then purified or peptides may be synthesized. Alternatively, the allergenic protein may be purified from natural sources.
[0055] The eukaryotic cell expression vector included in the compositions or kits of the invention may be an expression vector composed of a plasmid expression vector, a viral expression vector or bacteriophage expression vector. Plasmid DNA and chromosome DNA fragment-formed expression vector and other expression vectors are well known and commonly used in the field of genetic engineering. In some embodiments, the plasmid vector pVAX1 (Invitrogen) is used. In some embodiments, the plasmid vector provax which has the CMV promoter, an hCG leader and bovine growth hormone poly A is used. In some embodiments, the plasmid vector is a pcDNA3 plasmid (Invitrogen) which comprises a human cytomegalovirus immediate-early (CMV) promoter, bovine growth hormone polyadenylation signal (BGH polyA), T7 sequence, ColE1 origin of replication, and the JE virus signal sequence.
[0056] In the eukaryotic cell expression vectors, the coding sequence for the allergenic protein or peptide or protein that comprises an antigenic epitope of such allergenic protein is operably linked to regulatory sequences required for eukaryotic expression. Examples of suitable promoters include an RSV (Rous sarcoma virus) promoter, a CMV (cytomegalovirus) promoter such as the CMV immediate early promoter, an SV40 virus promoter, Mouse Mammary Tumor Virus (MMTV) promoter, Human Immunodeficiency Virus (HIV) such as the HIV Long Terminal Repeat (LTR) promoter, Moloney virus, ALV, Epstein Barr Virus (EBV), as well as promoters from human genes such as human Actin, human Myosin, human Hemoglobin, human muscle creatine and human metallothionein. Examples of polyadenylation signals useful to practice the present invention, include but are not limited to SV40 polyadenylation signals and LTR polyadenylation signals. In addition to the regulatory elements required for DNA expression, other elements may also be included in the DNA molecule. Such additional elements include enhancers. The enhancer may be selected from the group including but not limited to: human Actin, human Myosin, human Hemoglobin, human muscle creatine and viral enhancers such as those from CMV, RSV and EBV.
[0057] In some embodiments, the proportion of eukaryotic cell expression vector to allergenic protein or peptide or protein that comprises an antigenic epitope of such allergenic protein is 1:5-5:1; the preferred option is: 1:1 (the mole ratio is 1-20:100,000; the preferred molar ration is 15:100,000).
[0058] In some embodiments, the inhibitor composition or combination of kit components is introduced into the organism intramuscularly, intracutaneously/intradermally, transdermally, subcutaneously, intravenously and through mucosal tissue by means of injection, nebulizer/aerosol/spraying, nose drops, eye drops, orally, sublingual, buccal, vaginal, penetration, absorption, physical or chemical means; or it may be introduced into the organism through other physical mixture or package. The kit components do not have to be delivered together, nor do they have to be delivered at the same site or by the same route of administration.
[0059] The pharmaceutical composition may be introduced by various means including, for example, the needle injection, needleless injector, gene gun, electroporation, and microprojectile bombardment.
[0060] The composition and kit components may be formulated by one having ordinary skill in the art with compositions selected depending upon the chosen mode of administration. Suitable pharmaceutical carriers are described in the most recent edition of Remington's Pharmaceutical Sciences, A. Osol, a standard reference text in this field.
[0061] For parenteral administration, formulations may be provided as a solution, suspension, emulsion or lyophilized powder in association with a pharmaceutically acceptable parenteral vehicle. Examples of such vehicles are water, saline and dextrose solution. Liposomes and nonaqueous vehicles such as fixed oils may also be used. The vehicle or lyophilized powder may contain additives that maintain isotonicity (e.g., sodium chloride, mannitol) and chemical stability (e.g., buffers and preservatives). The formulation is sterilized by commonly used techniques. Injectable compositions may be sterile and pyrogen free.
[0062] The dosage administered varies depending upon factors such as: pharmacodynamic characteristics; its mode and route of administration; age, health, and weight of the recipient; nature and extent of symptoms; kind of concurrent treatment; and frequency of treatment. In some embodiments, the amount of composition used or the amount of combination of kit components is generally 1250 ug/kg body weight/administration; with the composition or kit components administered once every 1-30 days, preferably once every 7-14 days. In some embodiments, a single dose is administered. In some embodiments, multiple doses are administered. In some embodiments, a total of 2-3 administrations are administered.
[0063] In some embodiments, the compositions or kits are administered to individuals who are suffering from an allergic reaction. In some embodiments, the compositions or kits are administered to individuals who are not suffering from an allergic reaction but who have been exposed to the allergen or likely to have been exposed to the allergen. In some embodiments, the compositions or kits are administered to individuals who are not suffering from an allergic reaction but who are known to be allergic to the allergen, ie. who have previously had allergic responses to the allergen.
[0064] The methods of the present invention are useful in the fields of both human and veterinary medicine. Accordingly, the present invention relates to treatment and prevention of allergic reactions in mammals, birds and fish. The methods of the present invention can be particularly useful for mammalian species including human, bovine, ovine, porcine, equine, canine and feline species.
[0065] The Examples set out below include representative examples of aspects of the present invention. The Examples are not meant to limit the scope of the invention but rather serve exemplary purposes. In addition, various aspects of the invention can be summarized by the following description. However, this description is not meant to limit the scope of the invention but rather to highlight various aspects of the invention. One having ordinary skill in the art can readily appreciate additional aspects and embodiments of the invention.
EXAMPLES
Example 1
Polypeptide Synthesis of Flea Salivary Allergenic Peptides and Construction of a Eukaryotic Cell Expression Vectors for Expression of the Same
[0066] The described flea salivary allergenic protein (FSA) possesses the amino acid residue sequence as described in SEQ ID NO:2.
[0067] The synthesized polypeptide that comprises the amino acid residues described in SEQ ID NO: 22 is named pep66. The synthesized polypeptide that comprises amino acids described in SEQ ID NO: 23 is named pep 100.
[0068] Nucleotide sequences that encode pep66 have the nucleotide sequence described in SEQ ID NO 24 and are named FAD66.
[0069] Nucleotide sequences that encode pep 100 have the nucleotide sequence described in SEQ ID NO 25 and are named FAD 100.
[0070] Eukaryotic vectors that encode pep66 and pep 100 comprise at least one nucleotide sequence described in SEQ ID NO 24 or SEQ ID NO: 25 and are named pcDF66 or pcDF100, respectively.
[0071] Flea Salivary Allergenic Protein was purchased from the Greer Laboratory Company (Lenoir, N.C., United States) and was formulated by the systematic flea cultivation methods described by Lee, et al., in Parasite Immunology 19:13-19, 1997. At the end of one adult year, female fleas were obtained from isolated salivary glands of infected animals. Salivary gland cells were suspended in SDS-reduction buffer and agitated in an oscillator for 30 seconds. Cells were pulverized and crude protein was stored at -20.degree. C.
[0072] The described eukaryotic cell expression vectors comprising nucleotide sequences that encode FSA epitopes pep66 and pep 100 are named pcDF66 or pcDF100, respectively. 1. Synthesis of FSA polypeptides pep66, pep 100, and their encoded genes
[0073] We verified the amino acid sequences of MHC Class II epitopes of FSA and the chemically synthesized peptides pep66 and pep 100 using Epitlot software. Sequences of the newly synthesized genes and protein products are below:
[0074] Peptide 66-80: QEKEKCMKFCKKVCK (SEQ ID NO: 22), called pep66;
[0075] Peptide 100-114: GPDWKVSKECKDPNN (SEQ ID NO: 23), called pep 100.
[0076] Nucleotide sequences that encode pep66 and pep 100 have been named FAD66 and FAD 100, respectively. The nucleotide sequences comprise the following sequences:
[0077] FAD66: CAAGAGAAAG AAAAATGTAT GAAATTTTGC AAAAAAGTTT GCAAA (SEQ ID NO: 24
[0078] FAD 100: GGTCCTGATT GGAAAGTAAG CAAAGAATGC AAAGATCCCA ATAAC (SEQ ID NO: 25). 2. Construction of FAD66 and FAD 100 expression vectors
[0079] Expression vector pGFP (Clontech, Mountain View, Calif., U.S.A.) was purchased as a template. We conducted Polymerase chain reaction (PCR) amplification of FSA nucleotide sequences to label the FSA nucleotide sequence with the Green Fluorescent Protein (GFP) gene. Primer extension was completed by use of the P66F and PR primers as well as the PI OOF and PR primers. Sequences for the primers used in cloning method are as follows: P66F: 5'-AAGCTTGCCA TGCAAGAGAA AGAAAAATGT ATGAAATTTT GCAAAAAAGT TTGCAAAGGTACC GCCATGG TGAGCAAGGG CGAGGA-3' (SEQ ID NO: 26) (the 13.sup.th site-57.sup.th site basic group from the 5' terminal end of the amplification product said sequence is FAD66; the 58th site-63rd site basic group from the 5' terminal end of the amplification product is the Kpn 1 recognition site; the first through sixth basic nucleotide from the 5' terminal end of the amplification product is the Hind III recognition site); PR: 5'-TTA GGTACCTTAC TTGTAC AGCTCGTCCAT-3' (SEQ ID NO: 27) (the 4.sup.th site-9.sup.th site basic group from the 5' terminal end of the amplification product in said sequence is the Kpn I recognition site), PI OOF: 5'-AAGCTTGCCA TG GGTCCTGA TTGGAAAGTA AGCAAAGAAT GCAAAGATCC CAATAACGGT ACC GCCATGGTGAGCAAGGGCGAGGA-3' (SEQ ID NO: 28) (the 13.sup.th site-57.sup.th site basic group from the 5' terminal end of the amplification product in said sequence is FAD66; the 58.sup.th site-63.sup.rd site basic group from the 5' terminal end of the amplification product is the Kpn I recognition site; the 1.sup.st site-6.sup.th site basic group from the 5' terminal end of the amplification product in said sequence is the Hind III recognition site), PR: 5'-TTA GGTACCTTAC TTGTAC AGCTCGTCCAT-3' (SEQ ID NO: 27) (the 4.sup.th site-9.sup.th site basic group from the 5' terminal end of the amplification product is the Kpn I recognition site). PCR product and eukaryotic expression vector pcDNA3 (Invitrogen Corp., Carlsbad, Calif., U.S.A.) were digested using BamHX and Hind III. The amplification product was ligated into the plasmid using T4 DNA ligase, and then transformed into Escherichia coli Top 10. After E. coli were grown in incubators, plasmid DNA was extracted and restriction digestion was performed using Kpn I to obtain a positive clones. Positive clones included plasmid pcDF66-GFP containing FAD66 and GFP genes and plasmid pcDF100-GFP containing FAD 100 and GFP genes. After using pcDF66-GFP and pcDF100-GFP for Kpn I digestion, low melting point agarose gel electrophoresis was used to recover large fragments, and then self-binding is conducted. Finally, the product is transformed into Escherichia coli Top 10, the plasmid is extracted and restriction endonuclease Kpn/digestion assay is used to obtain a positive clone. Obtained clones included FAD66 expression vector pcDF66 and FAD 100 expression vector pcDF100.
[0080] Normal simian kidney cells (CV1 cells) (purchased from the Institute of Cell Biology, Shanghai) were cultured in DMEM containing 10% fetal calf serum under 5% CO2, and 37.degree. C. Transfections of pcDF66-GFP and pcDF100-GFP were performed on the CV1 cells in a 35 mm culture dish with 2.5.times.10.sup.5 cells per/ml and 2 ml per dish. Purification of the plasmids was performed in accordance with the methodology described in the Guidebook for Molecular Cloning Experimentation (3.sup.rd edition) (Chinese translation) (translated by Huang Peitang et al., Science Publishing Company, published September 2002). A positive ion liposome medium (Lipofectamine.TM. 2000, Invitrogen) was used to transfect and culture the CV1 cells according to the manufacturer's instructions (Invitrogen, CA, USA). After 24 hours of incubation, the cell culture is observed under fluorescence microscopy showing pcDF100-GFP and pcDF66-GFP was expressed in eukaryotic cells. The results demonstrate that pcDF100-GFP and pcDF66-GFP also may be expressed in eukaryotic cells.
Example 2
Flea Allergy Dermatitis Inhibitor as Therapy for Flea Allergy Dermatitis
[0081] Experiments in which Kunming white, BALB/c and C57BL/6 mice are immunized with a vector comprising FSA protein and nucleotide sequences that encode FSA proteins, or proteins thereof, demonstrate that immunization is an effective therapy for flea allergy dermatitis. Useful vectors for immunization can comprise a eukaryotic cell expression vector further comprising a nucleotide sequences encoding FSA protein (for example, pcDF66 or pcDF100), and either FSA synthesized peptides (pep66 or pep 100) or FSA protein (flea salivary allergenic protein). The immunization efficacy is superior to that of an immunization vector merely comprising a eukaryotic cell expression vector that included nucleotide sequences that encodes either FSA peptides or FSA protein. Inhibition studies demonstrate that the DNA sequences encoding different epitopes have different efficacies. For instance, FAD 100 appears to have a stronger therapeutic effect in suppressing Flea Allergy Dermatitis. Results in each strain of mouse were similar, indicating that immunosuppressive activity of the immunization is not limited to MHC genetic backgrounds. Therefore, we can directly deduce from the above results that through use of a FAD Inhibitor comprising a eukaryotic cell expression vector which further comprises a nucleotide sequence that encodes FSA protein and either a FSA protein or a FSA peptide, it is possible to effectively inhibit flea allergy dermatitis.
[0082] T-cell proliferation experiments and related cytokine expansion experiments demonstrate that a FAD Inhibitor, comprising a eukaryotic cell expression vector that encodes FSA protein and an FSA protein or FSA peptide, inhibit antigen-specific T-cell proliferation thereby suppressing an allergic reaction. Immunosuppression may be induced through IL-10, thus inhibiting IL-5, IL-13 and other cytokine expression levels. The FAD Inhibitor in the present invention may effectively prevent and/or treat flea allergy dermatitis, especially those cases caused by Ctenocephalides felis. 1. Kunming white mice experiments.
[0083] Three Hundred Sixty (360) female Kunming white mice were divided into a total of 12 groups of equal numbers. Each mouse in the first group was immunized with 100 microliters of 0.9% NaCl aqueous solution containing 100 micrograms of pcDF66. Each mouse in the second group was immunized with 100 microliters of 0.9% NaCl aqueous solution containing 100 micrograms pep66. Each mouse in the third group was immunized with 100 microliters of 0.9% NaCl aqueous solution containing 50 micrograms of pcDF66 and 50 micrograms of pep66. Each mouse in the fourth group was immunized with 100 microliters of 0.9% NaCl aqueous solution containing 100 micrograms of pcDF100. Each mouse in the fifth group was immunized with 100 microliters of 0.9% NaCl aqueous solution containing 100 micrograms of pep 100. Each mouse in the sixth group was immunized with 100 microliters of 0.9% NaCl aqueous solution containing 50 micrograms of pcDF100 and 50 micrograms of pep100. Each mouse in the seventh group was immunized with 100 microliters of 0.9% NaCl aqueous solution containing 50 micrograms of pcDF66 and 50 micrograms of pep 100. Each mouse in the eighth group was immunized with 100 microliters of 0.9% NaCl aqueous solution containing 50 micrograms of pcDF100 and 50 micrograms of pep66. Each mouse in the ninth group was immunized with 100 microliters of 0.9% NaCl aqueous solution containing 100 micrograms of inactivated flea antigen (purchased from the Greer Lab Company, North Carolina, United States). Each mouse in the tenth group was immunized with 100 microliters of 0.9% NaCl aqueous solution containing 50 micrograms of inactivated flea antigen (purchased from the Greer Lab Company, North Carolina, United States) and 50 micrograms of pcDF66. Each mouse in the eleventh group was immunized with 100 microliters of 0.9% NaCl aqueous solution containing 50 micrograms of inactivated flea antigen (purchased from the Greer Lab Company, North Carolina, United States) and 50 micrograms of pcDF100. Each mouse in the twelfth group was immunized with 100 microliters of 0.9% NaCl aqueous solution containing 100 micrograms of pcDNA3 to serve as a control. Fourteen days after the first immunization, a booster immunization was administered in the same dosage amount. Seven days after the booster immunization, skin tests were conducted using the following methods.
[0084] Hair was removed from the ventral murine chest cavity and an intracutaneous injection of 30 .PHI.l of inoculation of 1 ug/ul FSA protein was injected into 10 subjects. At the same time a histamine solution (with a concentration of 0.01% histamine) and PBS was injected in equal amounts to serve as positive controls and negative controls. There were 10 subjects for each control group. Twenty minutes after each injection, we measured blister diameters in micrometers. The t test results indicated that pcDF100 and pep 100 compound immunization (the sixth group) demonstrated a notable difference (P<0.05) when compared to pcDF100 single immunization (the fourth group) or pep 100 single immunization (the fifth group). There was a notable difference (P<0.05) between pcDF66 and pep66 compound immunization (the third group) and pcDF66 single immunization (the first group), while there was no notable difference with pep66 single immunization (the second group). Flea antigen and pcDF66 (the tenth group) or pcDF100 (the eleventh group) compound immunizations were notably different (P<0.05) than the results generated by flea antigen alone (the ninth group), The tenth and eleventh groups did not display any notable difference as compared to pcDF66 (the first group) or pcDF100 (the fourth group) immunizations. There was no difference in blister diameters measured in the sixth and seventh groups as compared to the expression vector or epitope polypeptide single immunity groups (Groups 1, 2, 3 and 4).
[0085] Based on the preceding skin test results, we can conclude that a eukaryotic cell expression vector comprising FSA protein or a nucleotide sequence encoding FSA peptide and said FSA peptide, or a eukaryotic cell expression vector comprising FSA protein or a nucleotide sequence that encodes FSA peptides and FSA protein reduces skin allergies in an antigen-specific way. These results indicate that reduction of allergic reactions on the skin may be induced through immunosuppression. 2. BALB/c and C57B/6 mice skin test results
[0086] In order to further verify the results obtained above with Kunming white mice, two pure strains of mice (BALB/c and C57B/6) were tested to determine whether immunosuppression of FAD was MHC restricted. Experimental methodologies were the same as those methodologies performed in the Kunming white mice. The t test results indicate that pcDF100 and pep 100 immunization (the sixth group) had no notable difference (P<0.05) as compared to pcDF100 single immunization (the fourth group) or pep 100 single immunization (the fifth group). Results from the flea antigen and pcDF100 compound immunization (the eleventh group) demonstrated a very significant difference (PO.01) compared to the results generated by immunizations of flea antigen (the ninth group) or pcDF100 (the fourth group). There was no significant difference among immunizations of Flea antigen and pcDF66 immunization (the tenth group), flea antigen immunization alone (the ninth group), or pcDF66 immunization alone (the first group). There was no significant difference among pcDF100 and pep66 immunization (the eighth group), pcDF100 immunization alone (the fourth group), or pep66 immunization alone (the second group).
[0087] The preceding results indicate that anti-allergic immunosuppression is antigen-specific. For example, immunization with pcDF100 and pep 100 was more effective in as compared to immunization with pcDF100 or pep100 alone. Immunization with the flea antigen and pcDF100 were more effective than any immunization vectors that included only one component.
[0088] Differences also exist in the level of immunosuppression among those groups that effectively treated FAD. For example, the flea antigen and pcDF100 compound immunity group had more of an effective immunosuppressive effect than compared any of the single immunity groups. The t test results indicate that pcDF100 and pep 100 compound immunity (the sixth group) are significantly different (P<0.05) when compared to results of immunization using pcDF100 alone (the fourth group) or pep100 alone (the fifth group). There are significant differences (P<0.05) in the immunosuppressive effect of the immunizations performed with pcDF100 and pep66 compound immunity (the eighth group) as compared to pcDF100 alone (the fourth group) or pep66 alone (the second group). There was an extremely significant difference (P<0.01) in immunizations using Flea antigen and pcDF100 (the eleventh group) as compared to flea antigen (the ninth group) or pcDF100 alone (the fourth group).
[0089] We interpret our results to conclude that anti-allergic immunosuppression is antigen-specific. For example, immunization was more effective in the pcDF100 and pep100 compound immunity group as compared to the pcDF100 single immunity group or the pep100 single immunity group. In addition, pep66 and pep100 peptides may have cross-reactivity. For example, immunization was more effective with pcDF100 and pep66 immunization as compared to pcDF100 single immunity group or the pep66 single immunity group. Immunization using the flea antigen and pcDF66 compound immunity did not produce clear immunosuppression. These results are consistent with those obtained in the BALB/c group. Although there are slight differences in the effectiveness of the immunization among each strain of mice studied, experimental results of the three different strains of mice same conclusion: immunization vectors comprising eukaryotic cell expression vectors further comprising FSA protein or nucleotide sequences that encode FSA peptides and FSA peptides; or eukaryotic cell expression vectors comprising FSA protein or nucleotide sequences that encode FSA peptides and FSA protein may mount effective anti-allergic immunosuppression.
Example 3
T-Cell Proliferation in Immunized Mice
[0090] Three Hundred Sixty (360) BALB/c mice and three hundred sixty C57B/6 mice were each divided into 12 groups of 30 mice per group. Immunization was performed in accordance with the methodology described in Example 2. At seven days after the booster immunization, splenic T-cells were isolated and T-cell expansion activity was tested. The specific methodology was: under aseptic conditions, splenic cells were taken from mice and used to form a single-cell suspension fluid. A hemolytic solution was used to remove red blood cells, which were then washed three times using PBS fluid. The cells were centrifuged and a cell count taken. Cell concentration was adjusted to 1.times.10.sup.6 cells/ml, and each cell suspension from each animal was divided into four experimental groups and plated into a 96-well culture plate. To one group, 100 ul Con-A (mitogen) was added to a final concentration of 5 fig/ml. Specific antigen (flea antigen) was added to serve as a stimulant to a final concentration of 5 ng/ml to the second group. No stimulant was added for a negative control group, and 100 ul BSA was added to a final concentration of 2 u.g/ml to serve as another unrelated antigen. After being incubated at 37.degree. C. in a CO2 culture for 48 h, 100 u.l MTS was added to each well at a final concentration of 5 mg/ml. After 4 hours of incubation, an enzyme labeler was used to read the OD value at 492 nm and calculate the stimulation index (SI=tested OD+non-stimulated OD). T-cell proliferation results for the BALB/c mice indicated that eukaryotic cell expression vectors comprising a nucleotide sequence that encodes FSA peptide flea salivary and said FSA peptide generate notable antigen-specific immunosuppression of FAD. For example, immunization was more effective from the vectors comprising pcDF100 and pep100 (the sixth group) and the vectors comprising flea antigen and pcDF100 (the eleventh group) as compared to immunization with vectors comprising single components. In addition, there was no clear immunosuppression demonstrated by immunization vectors comprising the pcDF66 and pep66 group (the third group), the flea antigen and pcDF66 group (the tenth group) as compared to the corresponding single immunity groups. Results using vectors comprising the pcDF100 and pep 100 compound immunity group and the flea antigen and pcDF100 compound immunity group are consistent with the immunosuppression effect shown by the skin tests. There was an extremely significant difference (P<0.01) in the immunosuppressive effect of the immunization vectors comprising the pcDF100 and pep66 (the eighth group) as compared to the immunosuppressive effect seen in the corresponding single immunity group.
[0091] C57B/6 T-cell proliferation results indicate that eukaryotic cell expression vectors containing nucleotide sequences that encode FSA peptide and said FSA peptide may produce clear antigen-specific immunosuppression. For example, there is a clear difference (PO.05) in the effect of immunization of pcDF66 and pep66 compound immunity (the third group) as compared to the corresponding single immunity groups (the first group and the second group). Additionally, there was an extremely significant difference (P<0.01) in the effect of immunization in the pcDF100 and pep100 compound immunity group (the sixth group) as compared to the corresponding single immunity groups (the fourth group and the fifth group). The immunization of mice using vector comprising the flea antigen and pcDF66 (the tenth group) is clearly more effective (P<0.05) than the corresponding single immunity groups (the ninth group and the first group). There was an extremely significant difference (P<0.01) in the effect of the immunization of the flea antigen and pcDF100 compound immunity group (the eleventh group) as compared to the corresponding single immunity groups (the ninth group and the fourth group). In addition, cross-reactivity exists between the two epitopes. For example, there is a clear difference (PO.05) in T-cell proliferation profiles with the pcDF66 and pep100 compound immunity group (the seventh group) and the corresponding single immunity groups (the first group and the fifth group). There is also a clear difference (P<0.05) in T-cell proliferation profiles of the pcDF100 and pep66 compound immunity group (the eighth group) and the corresponding single immunity groups (the fourth group and the second group). The T-cell proliferation profiles of in the pcDF100 and pep100 compound immunity group, the pcDF100 and pep66 compound immunity group and the flea antigen and pcDF100 compound immunity group results were all consistent with the skin test results.
Example 4
Changes in the Cytokine Levels of Immunized Mice
[0092] There were 360 Kunming white mice, 360 BALB/c mice, and 360 C57B/6 mice, and each set of subjects strain was divided into 12 groups of 30 mice. Immunization was performed in accordance with the method described in Example 2. Seven days after the booster immunization, the spleen was excised and total RNA (TRIZOL, Dingguo Biological Company) isolated. Reverse transcription for cDNA was performed in accordance with the Dalianbao Biological Company's RNA RT-PCR operating handbook. Briefly 1 jag of purified total RNA was placed in a 250 uL centrifuge tube. Then the following reagents were added in sequence: 4 ul MgCl.sub.2, 2 ul 10* buffer solution, 8.5 ul DEPC water, 2 ul dNTP mixture, 0.5 ul RNase inhibitor, 0.5 ul M-MLV reverse transcriptase (Promega), 0.5 uL Oligo (dT)i2 primer. The response conditions were 42.degree. C. for 30 min, 99.degree. C. for 5 min and 5.degree. C. for 5 minutes. The gene family hypoxanthine phosphoribosyltransferase (HPRT) was used as the internal source expression standard. The various groups' cDNA concentrations were adjusted to make all sample concentrations consistent. Then 2 .PHI.l of cDNA were used to conduct PCR amplification assessing the expression levels of the three cytokine genes: IFN-y, IL-4, and IL-10. The reaction's required primer and PCR reaction conditions are as indicated in Table 1. (Because the gene family HPRT has fixed expression in vivo, it is used as the template for the control's internal source expression standard).
TABLE-US-00001 TABLE 1 HPRT, IFN-y, IL-4 and IL-10 Primer Sequence and PCR Reaction Specifications. Target gene Primer Response conditions HPRT 5' GTTGGATACAGGCCAGACTTTGTTG (SEQ ID 94.degree. C. 30 sec, 60.degree. C. 30 sec NO: 29) 3' GAGGGTAGGCTGGCCTATGGCT (SEQ and 72.degree. C. 40 sec ID NO: 30) IFN-Y 5' CATTGAAAGCCTAGAAAGTCTG (SEQ ID 94.degree. C. 30 sec, 58.degree. C. 30 sec NO: 31) 3' CTCATGGAATGCATCCTTTTTCG (SEQ and 72.degree. C. 40 sec IDNO: 32) IL-4 5' GAAAGAGACCTTGACACAGCTG (SEQ 94.degree. C. 30 sec, 54.degree. C. 30 sec IDNO: 33) 3' GAACTCTTGCAGGTAATCCAGG (SEQ and 72.degree. C. 40 sec ID NO: 34) IL-10 5' CCAGTTTACCTGGTAGAAGTGATG (SEQ ID 94.degree. C. 30 sec, 56.degree. C. 30 sec NO: 35) and 72.degree. C. 40 sec 3'TGTCTAGGTCCTGGAGTCCAGCAGACTCAA (SEQIDNO.-36)
[0093] Bio-Rad Image software (Quantity One 4.2.0) was used to analyze images taken of the electrophoresis gels of the PCR products. Expression profile results obtained were generally consistent each strain of mouse. IL-IO expression levels in the pcDF66 and pep66 compound immunity (Group 3) were higher than pcDF66 single immunity (Group 1) and the pep66 single immunity (Group 2). IL-10 expression levels in the pcDF100 and pep 100 compound immunity (Group 6) were higher than pcDF100 single immunity group (Group 4) or pep 100 single immunity group (Group 5). IL-10 expression levels in the flea antigen and pcDF66 compound immunity group (Group 10) were higher than flea antigen (Group 9) or pcDF66 single immunity groups (Group 1). IL-10 expression levels in the flea antigen and pcDF66 compound immunity group (Group 11) were higher than flea antigen (Group 9) or pcDF100 single immunity groups (Group 4). There was no clear difference in the 1L-4 and IFN-y expression levels among the various groups. The results suggest that immunization conducted with eukaryotic cell expression vector comprising nucleotide sequences that encode FSA and said FSA protein or said FSA peptide compound enhanced IL-10 expression levels.
[0094] In addition, expression profiles were generated on IL-5 and IL-13 in the three types of mice. Results indicated that for the eukaryotic cell expression vectors comprising nucleotide sequences that encode FSA peptides and said FSA protein or peptide (Group 3, Group 6, Group 10 and Group 11), IL-5 and IL-13 levels were clearly lower than those of the respective single immunity groups.
Example 5
Detection of Blood IgE Levels in Immunized Mice
[0095] Two groups of 360 BALB/c and 360 C57B/6 mice were each divided into 12 groups of 30 mice each and immunization was performed as described in Example 2. Blood was taken intravenously from the eye socket prior to immunization and 14 days after the booster immunization. The blood was separated by centrifugation and IgE levels were assessed using ELISA. The coated antigen used on the ELISA plates was flea antigen purchased from Greer Laboratory (Lenoir, N.C., United States). The first component on the ELISA plate was separated blood sera and the second component used for binding was the sheep anti-mice IgE antibody labeled with horseradish peroxidase antioxidant enzyme. The substrate was added to the system after antibody was bound to the antigens and the enzyme labeler was used to read the OD values at 492 nm. The IgE production in Kunming white mice, BALB/c mice, C57B/6 mice after immunization are generally consistent for all three groups of mice. Except for the flea antigen single immunity group (the ninth group), IgE levels for the groups were relatively low. The IgE levels produced after immunization with the flea antigen single immunity group (the ninth group) were the highest levels across all sets of mice. IgE levels were greatly reduced in the group immunized with flea antigen and pcDF66 (the tenth group) and the group immunized with the flea antigen and pcDF100 (the 11 group) as compared to the flea antigen single immunity group (the ninth group). This indicates that a eukaryotic cell expression vector comprising nucleotide sequences that encode FSA peptides and said FSA protein reduces IgE levels after immunization.
Example 6
Feline Allergenic Protein Antigen Fel d. I.I (Fel d I with the Minor B Leader) Encoded Genetic Clone and Eukaryotic Expression Assay
[0096] 1. The following sequences were used as Fel d I.1 primers. Primer were artificially synthesized:
[0097] (a) Fel d I.1 PI 5' primer: AAGCTTGGATGTTAGACGC (SEQ ID NO 37)
[0098] (b) Fel d I.1 P2 3' primer: GGTACCTTAACACAGAGGAC (SEQ ID NO 38)
[0099] 2. Fel d I.1 expression vector construction
[0100] Fel d I.1 cDNA was used as a template for PCR amplification of the Fel d I.1 gene using Fel d 1.1 PI and Fel d I.1 P2. The primers are further described below:
[0101] (a) Fel d I.1 PI 5'-AAGCTTGGATGTTAGACGC-3' (SEQ ID NO 37) (the 1.sup.st site-6.sup.th site basic group from the 5' terminal end of the primer in this sequence is the Hind\\\recognition site; the 9.sup.th site-11.sup.th site is the original initiator code);
[0102] (b) Fel d I.1 P2 5'-GGTACCTTAACACAGAGGAC-3' (SEQ ID NO 38) (the 1.sup.st site-6.sup.th site basic group from the 5' terminal end of the. primer in this sequence is the Kpn I recognition site; the 7.sup.th site-9.sup.th site is the original termination code).
[0103] Kpn I and Hind III, respectively, were used for digesting the PCR product and eukaryotic expression vector pVAX1 (Invitrogen Corp.). Digested nucleotide fragments were then ligated using T4 DNA ligase. The resultant plasmid product was transformed into Escherichia coli Top 10, grown for maximum copy number, and then the plasmid isolated using methods known by those of ordinary skill in the art. We performed a restriction digestion using the Kpn/and Hind III endonucleases. Selected plasmid clones named pVAX1-Fel d I.1 comprised the pVax plasmids sequence and the Fel d I.1 gene. Through sequential analysis (Augct Co. Ltd., Beijing China), further analysis was performed to obtain the final Fel d I.1 PI expression vector, pFeld 1.1.
[0104] 3. pFel d I.1 Eukaryotic Expression
[0105] Normal simian kidney cells (CV1 cells, purchased from the Shanghai Cell Institute) were cultured in DMEM containing 10% fetal calf serum under 5% CO.sub.2 and 37.degree. C. conditions. 2.5.times.10.sup.5 cells per/ml were pipeted in a 35 mm culture dishes in a 2 mL volume. Transfections were performed using standard methods. Briefly, purification of the plasmids was performed in accordance with the methodology in the Guidebook for Molecular Cloning Experimentation (third edition, Chinese translation) (translated by Huang Peitang et al., Science Publishing Company, published September 2002). A positive ion liposome medium (Lipofectamine.TM. 2000, Invitrogen) was used to transfect the cultured CV1 cells according to the manufacturer's instructions (Invitrogen, CA, USA). After 24 hours of transfection, the cells were collected and an RNA extraction reagent (TRIZOL, Dingguo Biological Company) was used to isolate total cellular RNA. In order to prevent contamination, total extracted RNA was separately loaded into several EP tubes. A micropipette was used to carefully suction 2 ul of total RNA extracted and RT-PCR reagent used to expand total cellular cDNA. After the specimen was added, reverse transcription was performed at 42.degree. C. for 30-60 minutes, denaturation at 99.degree. C. for 5 minutes, and The reaction tube was set aside at 5.degree. C. for 5 minutes prior to extraction for use. Using the isolated cDNA as a template for gene amplification PCR was performed using Feld1.1P1 and FeldI.1P2 as the primers. The PCR products were subject to low melting point agarose gel analysis to detect Feld1.1 positive bands. The results demonstrate that Fel dI.1 is expressed in eukaryotic cells.
Example 7
Feline Allergenic Protein Antigen Fel d. 1.2 Genetic Clone and Eukaryotic Expression Assay
[0106] 1. The following sequences were used as Fel d I.2 primers. Primers were artificially synthesized:
[0107] (a) Fel d I.2 P1 5' primer: AAGCTTGGATGAAGGGGGCTC (SEQ ID NO:39)
[0108] (b) Fel d I.2 P2 3' primer: GGTACCTTAACACAGAGGAC (SEQ ID NO:40)
[0109] 2. Fel d I.2 expression vector construction
[0110] Fel d I.2 cDNA was used as a template for PCR amplification using Fel d I.2 PI and Fel d I.2 P2 primers to generate copies of the Fel d I.2 gene. The primers are further described below: (a) Fel d I.2 PI 5'-AAGCTTGGATGAAGGGGGCTC-3' (SEQ ID NO:39) (the 1.sup.st site-6.sup.th site basic group from the 5' terminal end of the primer is the Hind III recognition site; the 9.sup.th site-11.sup.th site from the terminal end of the primer is the original initiator code); (b) Fel d I.2 P2 5'-GGTACCTTAACACAGAGGAC-3' (SEQ ID NO:40) (the 1.sup.st site-6.sup.th site basic group from the 5' terminal end of the primer is the Kpn I recognition site, the 7.sup.th site-9* site from the terminal end of the primer is the original termination code).
[0111] Kpn I and Hind III in succession were used for digesting the PCR product and eukaryotic expression vector, pVAX1 (Invitrogen Corp.). The digested plasmid and PCR product were ligated using T4 DNA ligase. The product was transformed into Escherichia coli Top 10, the plasmid extracted, and restriction endonuclease Kpn 1 and Hind 111 digestion assay used to obtain a positive clone, that is, plasmid pVAX1-Fel d I.2 containing the Can f 1 gene. Through sequential analysis (Augct Co. Ltd., Beijing China), further assay correction was performed to obtain the Fel d I.2 expression vector, pFeld I.2.
[0112] 3. pFeldI.2 Eukaryotic Expression
[0113] The pFeld I.2 expression vector was subjected to the digestion, transformation, isolation, and ligation protocols previously described in Example 6, Section 3. Isolated PCR products were subjected to low melting point agarose gel testing to confirm Feld1.2 positive bands. The results prove that Feld1.2 is expressed in eukaryotic cells.
Example 8
Canine Allergenic Protein Antigen Can f 1 Genetic Clone and Eukaryotic Expression Assay
[0114] 1. Can f 1 Primer Synthesis
[0115] The following sequences were used as Can f 1 primers. Primers were artificially synthesized:
[0116] Can f 1 PI 5' primer: AAGCTTATGAAGACCCTGCTCCTCAC (SEQ ID NO: 41) Can f 1 P2 3' primer: GGTACCCTACTGTCCTCCTGGAGAGC (SEQ ID NO: 42)
[0117] 2. Can f 1 Expression Vector Construction
[0118] Can f 1 cDNA was used as a template to perform PCR amplification using Can f 1 PI and Can f 1 P2 primers to generate copies of the Can f 1 gene. The primers are further described below: (a) Can f 1 PI 5'-AAGCTTATGAAGACCCTGCTCCTCAC-3' (SEQ ID NO:41) (the 1.sup.st site-6.sup.th site basic group from the 5' terminal end of the primer sequence is the Hind III recognition site; the 7.sup.th site-9.sup.th site from the terminal end of the primer sequence is the original initiator code); (b) Can f 1 P2 5.sup.r-GGTACCCTACTGTCCTCCTGGAGAGC-3' (SEQ ID NO:42) (the 1.sup.st site-6.sup.th site basic group from the 5' terminal end of the primer sequence is the Kpn I recognition site; the 7.sup.th site-9.sup.th site from the %' terminal end of the sequence is the original termination code).
[0119] PCR fragments and pVAX1 (Invitrogen, Inc.) were subjected to the digestion, transformation, isolation, and ligation methods as described in Example 7, Section 2. Positive clones were selected and analyzed using sequential analysis discussed in Example 7, Section 2 to obtain the Can f 1 expression vector, pCanf1.
[0120] 3. pCanf1 Eukaryotic Expression
[0121] The pCanf1 expression vector was subjected to transfection, total RNA isolation, and RT-PCR protocols as previously described in Example 6, Section 3. Isolated PCR products were subjected to low melting point agarose gel testing to confirm pCanf1 positive bands. The results prove that pCanf1 is expressed in eukaryotic cells.
Example 9
Canine Allergenic Protein Antigen Can f 2 Genetic Clone and Eukaryotic Expression Assay
[0122] 1. Can f 2 Primer Synthesis
[0123] The following sequences were used as Can f 2 primers. Primers were artificially synthesized:
[0124] Can f 2 PI 5' primer: AAGCTT ATGCAGCTCCTACTGCTG (SEQ ID NO: 43) Can f 2 P2 3' primer: GGTACCCTAGTCTCTGGAACCC (SEQ ID NO: 44)
[0125] 2. Can f 2 Expression Vector Construction
[0126] Can f 2 CDNA cDNA was used as a template to perform PCR amplification using Can f 2 PI and Can f 2 P2 primers to generate copies of the Can f 2 gene. The primers are further described below: (a) Can f 2 PI 5'-AAGCTT ATGCAGCTCCTACTGCTG-3' (SEQ ID NO:43) (the 1.sup.st site-6.sup.th site basic group from the 5' terminal end of the primer sequence is the Hind III recognition site; the 7.sup.th site-9.sup.th site is the original initiator code); (b) Can f 2 P2 5' GGTACCCTAGTCTCTGGAACCC-3' (SEQ ID NO:44) (the 1.sup.st site-6.sup.th site basic group from the 5' terminal end of this primer sequence is the Kpn I recognition site, the 1.sup.x site-9.sup.l site is the original termination code).
[0127] PCR fragments and pVAX1 (Invitrogen, Inc.) were subjected to the digestion, transformation, isolation, and ligation methods as described in Example 7, Section 2. Positive clones were selected and analyzed using sequential analysis discussed in Example 7, Section 2 to obtain the Can f 2 expression vector, pCanf2.
[0128] 3. pCanf2 Eukaryotic Expression
[0129] The pCanf2 expression vector was subjected to transfection, total RNA isolation, and RT-PCR protocols as previously described in Example 6, Section 3. Isolated PCR products were subjected to low melting point agarose gel testing to confirm pCanf2 positive bands. The results prove that pCanf2 is expressed in eukaryotic cells.
Example 10
Dust Mite Allergenic Protein Antigen Der p 1 Genetic Clone and Eukaryotic Expression Assay
[0130] 1. Der p 1 Primer Synthesis
[0131] The following sequences were used as Der p 1 primers. Primers were artificially synthesized:
[0132] Der p 1 PI 5' primer: AAGCTTAACATGAAAATTGTTTTGG (SEQ ID NO: 45) Der p 1 P2 3' primer: GGTACCGTTTAGAGAATGACAACAT (SEQ ID NO: 46)
[0133] 2. Der p 1 Expression Vector Construction
[0134] Der p 1 cDNA was used as a template to perform PCR amplification using Der pi PI and Der p 1 P2 primers to generate copies of the Der p 1 gene. The primers are further described below:
[0135] (a) Der p 1 PI 5'-AAGCTTAACATGAAAATTGGTTTTGG-3' (SEQ ID NO:45) (the 1.sup.st site-6.sup.th site basic group from the 5' terminal end of the primer sequence is the Hind III recognition site, the 10.sup.th site-12.sup.th site is the original initiator code);
[0136] (b) Der p 1 P2 5'-GGTACCGTTTAGAGAATGACAACAT-3' (SEQ ID NO:46) (the 1.sup.st site-6.sup.th site basic group from the 5' terminal end of the primer sequence is the Kpn I recognition site, the 9.sup.th site-11.sup.th site is the original termination code).
[0137] PCR fragments and pVAX1 (Invitrogen, Inc.) were subjected to the digestion, transformation, isolation, and ligation methods as described in Example 7, Section 2. Positive clones were selected and analyzed using sequential analysis discussed in Example 7, Section 2 to obtain the Der p 1 expression vector, pDerp1.
[0138] 3. Der p 1 Eukaryotic Expression
[0139] The pDerp1 expression vector was subjected to transfection, total RNA isolation, and RT-PCR protocols as previously described in Example 6, Section 3. Isolated PCR products were subjected to low melting point agarose gel testing to confirm pDerp1 positive bands. The results prove that pDerp1 is expressed in eukaryotic cells.
Example 11
Peanut Allergenic Protein Antigen Ara h II Genetic Clone and Eukaryotic Expression Assay
[0140] 1. Ara h II Primer Synthesis
[0141] The following sequences were used as Ara h II primers. Primers were artificially synthesized:
[0142] Ara h II PI 5' primer: AAGCTTCTCATGCAGAAGAT (SEQ ID NO: 47) Ara h II P2 3' primer: GGTACCTTAGTAT CTGTCTC (SEQ ID NO: 48)
[0143] 2. Ara h II Expression Vector Construction
[0144] Ara h II cDNA was used as a template to perform PCR amplification using Ara h II PI and Ara h II P2 primers to generate copies of the Ara h II gene. The primers are further described below: (a) Ara h II PI 5'-AAGCTTCTCATGCAGAAGAT-3.sup.f (SEQ ID NO:47) (the 1.sup.st site-6.sup.th site basic group from the 5' terminal end of the primer sequence is the HindlU recognition site, the 10.sup.th site-12.sup.th site is the original initiator code), Ara h II P2 5'-GGTACCTTAGTATCTGTCTC-3' (SEQ ID NO:48) (the 1.sup.st site-6.sup.th site basic group from the 5' terminal end of the primer sequence is the Kpn I recognition site, the 7.sup.th site-9.sup.th site from the 5' terminal end of the primer sequence is the original termination code).
[0145] PCR fragments and pVAX1 (Invitrogen, Inc.) were subjected to the digestion, transformation, isolation, and ligation methods as described in Example 7, Section 2. Positive clones were selected and analyzed using sequential analysis discussed in Example 7, Section 2 to obtain the Ara h II expression vector, pArahII.
[0146] 3. Ara h II Eukaryotic Expression
[0147] The pArahII expression vector was subjected to transfection, total RNA isolation, and RT-PCR protocols as previously described in Example 6, Section 3. Isolated PCR products were subjected to low melting point agarose gel testing to confirm pArahII positive bands. The results prove that pArahII is expressed in eukaryotic cells.
Example 12
Peanut Allergenic Protein Antigen Ara h 5 Genetic Clone and Eukaryotic Expression Assay
[0148] 1. Ara h 5 Primer Synthesis
[0149] The following sequences were used as Ara h 5 primers. Primers were artificially synthesized:
[0150] Ara h 5 PI 5' primer: AAGCTTATGTCGTGGCAAAC (SEQ ID NO: 49) Ara h 5 P2 3' primer: GGTACCTAAAGACCCGTATC (SEQ ID NO: 50)
[0151] 2. Ara h 5 Expression Vector Construction
[0152] Ara h 5 cDNA was used as a template to perform PCR amplification using Ara 5 PI and Ara h 5 P2 primers to generate copies of the Ara h 5 gene. The primers are further described below: (a) Ara h 5 PI 5'-AAGCTTATGTCGTGGCAAAC-3' (SEQ ID NO:49) (the 1.sup.st site-b 6.sup.th site basic group from the 5' terminal end of the primer sequence is the Hind III recognition site; the 7.sup.th site-9.sup.th site is the original initiator code); (b) Ara h 5 P2 5'-GGTACCTAAAGACCCGTATC-3' (SEQ ID NO:50) (the 1.sup.st site-6.sup.th site basic group from the 5' terminal end of the primer sequence is the Kpn I recognition site; the 7* site-9.sup.l site from the 5' terminal end of the primer sequence is the original termination code).
[0153] PCR fragments and pVAX1 (Invitrogen, Inc.) were subjected to the digestion, transformation, isolation, and ligation methods as described in Example 7, Section 2. Positive clones were selected and analyzed using sequential analysis discussed in Example 7, Section 2 to obtain the Ara h 5 expression vector, pArah5.
[0154] 3. pArah5 Eukaryotic Expression
[0155] The pArah5 expression vector was subjected to transfection, total RNA isolation, and RT-PCR protocols as previously described in Example 6, Section 3. Isolated PCR products were subjected to low melting point agarose gel testing to confirm pArah5 positive bands. The results prove that pArah5 is expressed in eukaryotic cells.
Example 13
Japanese Cedar (Cryptomeria japonica) Pollen Allergenic Protein Antigen Cry j 1.1 Genetic Clone and Eukaryotic Expression Assay
[0156] 1. Cry j 1.1 Primer Synthesis
[0157] The following sequences were used as Cry j 1.1 primers. Primers were artificially synthesized
[0158] Cry j 1. 1 PI 5' primer: AAGCTTATGGATTCCCCTTGCTTAT (SEQ ID NO:51) Cry j 1. 1 P2 3' primer: GGTACCATCAACAACGTTTAGAG (SEQ ID NO: 52)
[0159] 2. Cry j 1.1 Expression Vector Construction
[0160] Cry j 1.1 cDNA was used as a template to perform PCR amplification using Cry j 1.1 PI and Cry j 1.1 P2 primers to generate copies of the Cry j 1.1 gene. The primers are further described below:
[0161] (a) Cry j 1.1 PI 5'-AAGCTTATGGATTCCCCTTGCTTAT-3' (SEQ IDN0:51) (the 1.sup.st site-6.sup.th site basic group from the 5' terminal end of the primer sequence is the Hind III recognition site; the 7.sup.th site-9.sup.th site is the original initiator code);
[0162] (b) Cry j 1.1 P2 5'-GGTACCATCAACAACGTTTAGAG-3' (SEQ ID NO:52) (the 1.sup.st site-6.sup.th site basic group from the 5' terminal end of the primer sequence is the Kpn I recognition site; the 7 site-9 site from the 5' terminal end of the primer sequence is the original termination code).
[0163] PCR fragments and pVAX1 (Invitrogen, Inc.) were subjected to the digestion, transformation, isolation, and ligation methods as described in Example 7, Section 2. Positive clones were selected and analyzed using sequential analysis discussed in Example 7, Section 2 to obtain the Cry j 1.1 expression vector, pCryj1.1.
[0164] 3. pCryj1.1 Eukaryotic Expression
[0165] The pCryj1.1 expression vector was subjected to transfection, total RNA isolation, and RT-PCR protocols as previously described in Example 6, Section 3. Isolated PCR products were subjected to low melting point agarose gel testing to confirm pCryj1.1 positive bands. The results prove that Cry j 1.1 may be expressed in eukaryotic cells.
Example 14
Japanese Cedar (Cryptomeria japonica) Pollen Allergenic Protein Antigen Cry j 1.2 Genetic Clone and Eukaryotic Expression Assay
[0166] 1. Cry j 1.2 Primer Synthesis
[0167] The following sequences were used as Cry j 1.2 primers. Primers were artificially synthesized
[0168] Cry j 1.2 PI 5' primer: AAGCTTATGGATTCCCCTTGCTTAG (SEQ ID NO: 53) Cryj 1.2 P2 3' primer: GGTACCTCAACAACGTTTAGAGAGAG (SEQ IDNO:54)
[0169] 2. Cry j 1.2 Expression Vector Construction
[0170] Cryj 1.2 cDNA was used as a template to perform PCR amplification using Cryj 1.2 PI and Cryj 1.2 P2 primers to generate copies of the Cryj 1.1 gene. The primers are further described below:
[0171] (a) Cry j 1.2 PI 5'-AAGCTTATGGATTCCCCTTGCTTAG-3' (SEQ ID NO:53) (the 1.sup.st site-6.sup.th site basic group from the 5' terminal end of the primer sequence is the Hind III recognition site; the 7.sup.th site-9.sup.th site from the 5' terminal end of the primer sequence is the original initiator code);
[0172] (b) Cry j 1.2 P2 5'-GGTACCTCAACAACGTTTAGAGAGAG-3' (SEQ ID NO:54) (the 1.sup.st site-6.sup.th site basic group from the 5' terminal end of the primer sequence is the Kpn I recognition site; the 7.sup.th site-9.sup.th site from the 5' terminal end of the primer sequence is the original termination code).
[0173] PCR fragments and pVAX1 (Invitrogen, Inc.) were subjected to the digestion, transformation, isolation, and ligation methods as described in Example 7, Section 2. Positive clones were selected and analyzed using sequential analysis discussed in Example 7, Section 2.to obtain the Cry j 1.2 expression vector, pCryj1.2.
[0174] 3. pCryj1.2 Eukaryotic Expression
[0175] The pCryj1.2 expression vector was subjected to the transfection, total RNA isolation, and RT-PCR protocols previously described in Example 6, Section 3. Isolated PCR products were subjected to low melting point agarose gel testing to confirm pCryj1.2 positive bands. The results prove that Cry j 1.2 may be expressed in eukaryotic cells.
Example 15
Blomia tropicalis Allergenic Protein Antigen Blo t 5 Genetic Clone and Eukaryotic Expression Assay
[0176] 1. Blo t 5 Primer Synthesis
[0177] The following sequences were used as Blo t 5 primers. Primers were artificially synthesized Blo t 5 PI 5' primer: AAGCTTACAATGAAGTTCGC (SEQ ID NO: 55) Blo t 5 P2 3' primer: GGTACCAATTTTTATTGGGT (SEQ ID NO: 56)
[0178] 2. Blo t 5 Expression Vector Construction
[0179] Blo t 5 cDNA was used as a template to perform PCR amplification using Blo t 5 PI and Blo t 5 P2 primers to generate copies of the Blo t 5 gene. The primers are further described below:
[0180] (a) Blo t 5 PI 5'-AAGCTTACAATGAAGTTCGC-3' (SEQ ID NO:55) (the 1.sup.st site-6.sup.th site basic group from the 5' terminal end of the primer sequence is the Hind III recognition site; the 10.sup.th-12.sup.th site from the 5' terminal end of the primer sequence is the original initiator code);
[0181] (b) Blo t 5 P2 5'-GGTACCAATTTTTATTGGGT-3' (SEQ ID NO:56) (the 1.sup.st site-6.sup.th site basic group from the 5' terminal end of the primer sequence is the Kpn I recognition site; the 12.sup.th site-14.sup.th site from the 5' terminal end of the primer sequence is the original termination code).
[0182] PCR fragments and pVAX1 (Invitrogen, Inc.) were subjected to the digestion, transformation, isolation, and ligation methods as described in Example 7, Section 2. Positive clones were selected and analyzed using sequential analysis discussed in Example 7, Section 2 to obtain the Blo t 5 expression vector, pBlot5.
[0183] 3. pBlotSEukaryotic Expression
[0184] The pBlot5 expression vector was subjected to transfection, total RNA isolation, and RT-PCR protocols as previously described in Example 6, Section 3. Isolated PCR products were subjected to low melting point agarose gel testing to confirm pBlot5 positive bands. The results prove that pBlot5 may be expressed in eukaryotic cells.
Example 16
Induction of Adaptive T Regulatory Cells that Suppress the Allergic Response: Conversion of T Regulatory Cells by Suboptimal DCs that are Induced by Co-Immunization of DNA and Protein Vaccines
[0185] Total flea extracts induce immediate intradermal allergic reactions in mice, therefore this system can be used to evaluate immunotherapeutic methods aimed at amelioration of AIH. Using this rodent model of AIH induced by flea allergens, co-immunization of DNA and protein vaccines encoding the flea salivary specific antigen (FSA1) ameliorates experimental AIH, including antigen induced wheel formation, elevated T cell proliferation, infiltration of lymphocytes and mast cells to the site of challenge. The amelioration of AIH was directly related to the induction of a specific population of flea antigenic specific T cells exhibiting a CD4.sup.+/CD257FoxP3.sup.+ phenotype, a characteristic of Tr. These Tr also express IL-10, IFN-y and the transcriptional factor T-bet after antigen stimulation. These Tr are driven by MHC-II.sup.+/CD40.sup.low DC populations that are induced by the co-immunization of DNA and protein vaccines. These studies identify important cellular players in the control of AIH. Exploitation of these cellular regulations and their induction will provide novel direction to develop therapies for allergic and related disorders.
Methods
[0186] Histology Analysis.
[0187] On day 14 following the last immunization, skin samples from mice were collected and fixed in 4% of paraformaldehyde, embedded in paraffin blocks from every group of mice. Four to five-micrometer sections were cut and placed on sylan-coated glass slides prior to rehydrated in xylene and washed with decreasing concentrations of alcohol solutions. The endogenous peroxidase activity was blocked by 3% hydrogen peroxide at room temperature for 10 min and the antigen retrieval was accomplished by boiling the slides in 0.01 M citrate buffer (pH 6.0). Lastly, slides were stained with hematoxylin and eosin (H&E) or toluidine blue for mast cells and analyzed under a light microscope for histology changes.
[0188] Skin Test.
[0189] On day 14 after the last immunization, the mice were challenged with 1 Hg/ul of flea-saliva-antigen on nonlesional lateral thorax skin intradermally, PBS is used as a negative control and histamine is used as a positive control. The diameter of the skin reaction was measured within 20 min after challenge by using a calibrated micrometer. Reaction was considered as a positive when the injection site was larger than half the size of the sums of diameters injected comparing the positive and negative control challenges.
[0190] T Cell Recall Responses.
[0191] The T cells isolated from immunized mice on day 14 were cultured at 5.times.10.sup.4 cells/well in triplicate in 96-well plates containing RPMI-10/5% FCS and then stimulated with 20 ug/ml of Flea-saliva-antigen for 48 h. Following the stimulation, cell proliferation was assessed by a colorimetric reaction after the addition of 20 pi of an MTS-PMS (Pormaga, USA) solution for 2-4 hrs and its color density was read at 570 nm by plate reader (Magellan, Tecan Austria GmbH).
[0192] Measurement of Flea Antigen-Specific Antibodies.
[0193] Serum concentration of anti-flea IgG1, IgG2a, IgG2b, IgM and IgE isotypes were measured using flea antigen coated plates by ELISA and detection with specific horseradish peroxidase-conjugated rat anti-IgG1, IgG2a, IgG2b, IgM and IgE antibodies (SouthernBiotech, Birmingham, USA), absorbance (450 nm) was measured using an ELISA plate reader (Magellan, Tecan Austria GmbH).
[0194] RT-PCR.
[0195] Total RNA was isolated from spleen and skin tissue 14 d after immunization using TRIzol reagent (Promega). cDNA was synthesized and PCR was performed in a 50 pi reaction mixture with 5 u, 1 cDNA and 1.0 M of each of the following primers: HPRT, IL-2, IFN-y, IL-4, IL-5, IL-13, IL-10 and 1L-12Q8). For Gata3 and T-bet analysis, CD4.sup.+CD25'' T cells were isolated and total RNA was prepared. RT-PCR was done as described using specific primer sequences as follows for Gata3, 5'-GGAGGCATCCAGACCCGAAAC-3.sup.1 (forward) (SEQ ID NO: 57) and 5'-ACCATGGCGGTGACC-ATGC-3.sup.1 (reverse) (SEQ ID NO: 58); for T-bet, 5'-TGAAGCCCACACTCCTACCC-3.sup.1 (forward) (SEQ ID NO: 59); and 5'-GCGGCATTTTCTCAGTTGGG-3.sup.1 (reverse) (SEQ ID NO: 60).
[0196] Isolation of CD4.sup.+CD25'' T Cells and Adoptive Transfer.
[0197] Single splenocyte suspensions were prepared from mouse spleen and CD4.sup.+CD25'' T cells were isolated and purified by using the MagCellect Mouse CD4.sup.+CD25.sup.+ Regulatory T Cell Isolation Kit according to the manufacturer's protocol (R&D Systems, Inc., USA). The purity of the selected cell populations was 96-98%. The purified cells (1.times.10.sup.6 per mouse) were adoptively transferred intravenously into C571BL6 mice.
[0198] CFSE Labeling and Co-Culture of Cells.
[0199] Naive CD4.sup.+ T cells isolated from C57/BL6 mice were labeled with CFSE (Molecular Probes). For assay of regulatory activity, 1.times.104 pcDF100+F induced regulatory or control T cells were co-cultured with 4.times.104 purified and CFSE labeled naive CD4.sup.+ T cells in the presence of flea antigen (100 u.g/ml) and 1.times.104 bone marrow derived DCs. For some cultures, Tr cells were co-cultured with anti-IL-10, anti-IFN-y or an isotype control antibody at 100 g/ml. After 72 h, cells were collected and labeled then CFSE+ cells were selected for analysis by flow cytometry.
[0200] Flow Cytometry.
[0201] CD4.sup.+CD25'' T cells were isolated and incubated on ice with PE-conjugated antibodies to CD44, CD69, CD62L (eBioscience, CA, USA). Flow cytometry of cytokine production and FoxP3 expression in T cells were performed, single cell suspensions were prepared from the animals spleens and Fc receptors were blocked with excess anti-Fc (BD PharMingen, USA). Cells were washed with ice-cold PBS. For intracellular cytokine staining, T cells were stimulated overnight with Con A (Sigma-Aldrich) in the presence of anti-CD28 mAb (BD PharMingen, USA). Collected cells were fixed with 4% paraformaldehyde and permeabilized with 0.1% saponin (Sigma-Aldrich). For staining of surface of CD4 or cytoplasmic IL-10, IL-4, IFN-y or FoxP3, the appropriate concentrations of phycoerythrin-labeled antibodies (eBioscience, CA, USA) were added to premeabilized cells for 30 min on ice followed by washing twice with cold PBS. Samples were processed and screened using FACSCalibur and data were analyzed with Cell Questpro software (BD).
Results
[0202] A flea salivary allergen, FSA1, has been identified and implicated as one of the causes for allergy dermatitis observed in cats and dogs. The degree of skin reaction or intradermal test (TDT) to assess the immediate intradermal flea-antigen reactivity can be achieved by flea allergy challenge intradermally. As expected, administration of mice with flea antigen induced significant skin reactions (FIG. 1A), induced mast cells activation, induced coincident IgE production (FIG. 1B), and induced strong CD4+ T cell proliferation responses (FIG. 1C) in C57 mice when compared with naive control animals after intradermal challenge. This data demonstrates the utility of the flea antigen allergic model for evaluation of novel therapeutic strategies against AIH.
[0203] This model was used to examine the ability of the co-inoculation strategy to protect animals from the immediate hypersensitivity after flea allergen challenge. C57/BL6 mice were pre-sensitized with various test vaccines and animals were intradermal challenged with the flea extracts, or with histamine as the positive or using PBS as the negative control. The immediate hypersensitivity reaction was blocked in group immunized 14 days earlier of co-inoculation of a plasmid DNA, pcDF100, encoding an epitope of FSA1 (aa100-1 14) mixed with the total flea proteins (designated as pcDF100+F) (FIG. 2A). To determine if the inhibition observed was due to the plasmid backbone or rather was related to a DNA construct encoding another region of FSA1. Robust inflammation of the challenged sites were observed in the mice immunized with either vector control plus flea proteins (designated as V+F), or a DNA construct encoding another region of FSA1 at aa66-80, pD66, again mixed with the flea proteins (designated as pcDF66+F, FIG. 2A). We also examined the influence of host immune balance toward Th1 type on the inhibition of allergic reaction, the pcDF100 primed and the flea protein boosted animals (designated as pcDF100.fwdarw.F; FIG. 2A). The severe reaction following challenge was observed in mice primed with pcDF100 and boosted with F (FIG. 2A), suggesting that induction of a strong immune response worsens the allergic reaction.
[0204] Histological analysis revealed infiltrations by leukocytes and mast cells in the skin lesions of mice immunized with F or V+F at the challenge sites; whereas, mice immunized with pcDF100+F showed normal intradermal structure which was free of inflammatory cells.
[0205] We next analyzed if the blocking of the immune reaction induced by the co-inoculation was a dose dependent. pcDF100 at dose of 25, 50, 100 and 200 (ig was co-immunized with 100 ug of flea proteins, respectively. Dosage at 50 jig of pcDF100 showed significant inhibition of the intradermal reaction which reached maximal inhibition at 100 (ig. A dosage of 25 ug exhibited only a minimal effect on lesion formation, as the animals developed severe reactions similar to those observed in animals inoculated with either F or V+F (FIG. 2B) or positive controls.
[0206] High levels of IL-4, IL-5 and IL-13 are the characteristics of allergic reactions and these immune modulators are implicated in allergy severity. Different profiles of the cytokines associated with the co-inoculated regimens were examined. Mice co-inoculated with F or V+F produced higher level of mRNA expressions for IL-4, IL-5 and IL-13; whereas, mice co-immunized with pcDF100+F produced relative low levels of these cytokines, suggesting that an anti-inflammatory immune regulatory function was derived from the co-inoculation of pcDF100+F. No significant differences were found in the levels of IL-2 or IFN-y among the co-inoculated regimens, but IFN-y was slightly higher in mice primed with pcDF100 and boosted by F. This result again suggests that the induced inhibition of the allergic reaction by co-immunization is not due to an un-balanced Th2 to Th1 response by the allergenic-specific T-helper cells.
[0207] As flea antigen triggered IgE-mediated allergy is well characterized, the ability of co-inoculation of pcDF100+F to inhibit anti-flea induced IgE production was examined. The levels of anti-flea IgE and IgG in sera were measured on days 14, 28 and 42 and were reduced slightly in mice co-immunized with pcDF100+F compared with groups immunized with V+F or F alone (FIG. 2C), suggesting that the co-inoculation does not influence the IgE production.
[0208] Proliferative CD4.sup.+ T cells are known to be involved in the development of immediate hypersensitivity. I isolated CD4.sup.+ T cells from the spleen of mice co-immunized with F, V+F or pcDF100+F was examined on day 14 after the second immunization for their recall proliferative responses to flea antigens in vitro. Immunizations of F and V+F resulted in strong proliferation of CD4.sup.+ T cells; whereas the CD4.sup.+ T cells isolated from pcDF100+F immunized mice showed little, if any, proliferation in response to the flea antigen stimulation (FIG. 2D). These results suggest that the inhibition of hypersensitivity observed by co-immunization of pcDF100+F is related to the non-responsive antigen specific CD4.sup.+ T cells.
[0209] The results indicate that the co-immunization with pcDF100 and flea proteins antigen induces an inhibition of the allergic reaction via down-regulating the levels of inflammatory cytokines and CD4.sup.+ cells induced by flea intradermal challenge. This suggests that the prevention of allergy is likely antigen specific since the antigen mismatched combinations did not produce the same effect.
[0210] To test if antigen-specific Tr cells have been induced by the co-immunization of flea DNA and protein vaccines, splenocytes were collected from co-immunized C57B/6 mice and mixed with the flea-antigen-specific effector T cells of syngeneic mice to exam their ability to inhibit recall proliferative responses in vitro. As shown in FIG. 3A, splenocytes from mice co-immunized with pcDF100+F significantly inhibited T cell recall immune responses. In contrast, splenocytes from mice immunized with F or V+F as well as from naive mice failed to inhibit the antigen specific T cell proliferative responses (FIG. 3A). This indicates that cells within the splenocyte population likely generated during the co-immunization of animals can suppress this antigen specific T cell proliferation. Purified non-T cells, T cells or subsets of T cells from the spleen of mice co-immunized with pcDF100+F were identified and tested individually for suppression of recall proliferation. Significant inhibition was observed from reactions of either purified T cells, purified CD4.sup.+ cells, or purified CD4.sup.+CD25'' cells from the mice co-immunized with pcDF100+F, but not from other subsets of cells (FIG. 3A). However, the inhibitions from CD4.sup.+CD25.sup.+ cells are in general thought to be independent of antigen sensitization, whereas the inhibition of CD4.sup.+CD25'' cells of this reaction is in an antigen dependent manner (FIG. 3A).
[0211] To further examine this issue in vivo, adoptive transfer was utilized. Antigen naive syngeneic recipient mice were adoptively transferred with either total splenocytes, T, CD4.sup.+ or CD8.sup.+ cells isolated from C57B/6 mice co-immunized with pcDF100+F, F or V+F, respectively. All recipient mice were next challenged intradermally by flea extract to induce the hypersensitivity. Splenocytes, T and CD4.sup.+ T cells from mice immunized with the pcDF100+F, but not with the F or V+F, were all able to suppress the development of the immediate hypersensitivity reaction (FIG. 3B). In contrast, CD8.sup.+ T cells isolated from all three experimental groups and naive control mice did not suppress this reaction. Both in vitro and in vivo results indicate that CD4.sup.+CD25'' Tr cells can mediate this suppression.
[0212] To investigate the observed role of CD4.sup.+CD25'Tr antigen specificity, the CD4.sup.+C25'' Trcells taken from C57B/6 mice co-immunized with pcDF100+F were adoptively transferred into the syngeneic recipient mice that were subsequently immunized twice at a biweekly interval with flea-antigen or OVA in Freunds' complete adjuvant (FCA). On day 14 after the last immunization, T cells were isolated and tested for their ability to proliferate to either flea antigen or OVA in vitro. T cells from recipient mice immunized with flea antigen did not respond the flea antigen stimulation in vitro; whereas, the T cells from recipient mice immunized with OVA-FCA responded well with OVA stimulation, but not to the flea antigens stimulation in vitro. As the controls, the naive mice immunized with flea antigen respond well to the flea antigen stimulation, but not to OVA stimulation in vitro and vice versa (FIG. 3C). This result indicates that adoptive transferred CD4.sup.+C25* Tr cells only inhibit the flea antigen specific T cell priming and proliferation in vivo; while the responses from the irrelevant antigen specific T cells were not affected.
[0213] Taken together, these data demonstrate that CD4.sup.+CD25'' Tr cells were induced by the co-immunization of DNA and protein vaccines. These appear to be unique CD4+ Tr cells as they function in an antigen-specific manner.
[0214] To determine if Tr cells induced by co-inoculation express certain types of cytokines and unique markers associated with Tr cells as previously documented (J. D. Fontenot, M. A. Gavin, A. Y. Rudensky, Nat Immunol 4, 330 (Apr. 1, 2003); M. G. Roncarolo, R. Bacchetta, C. Bordignon, S. Narula, M. K. Levings, Immunol Rev 182, 68 (Aug. 1, 2001); and P. Stock et al, Nat Immunol 5, 1149 (Nov. 1, 2004)), CD4.sup.+CD25'' cells were isolated from the mice immunized with F, V+F or pcDF100+F on days 1, 3, 7 and 14 post immunization and T cell profiles were followed by intracellular staining with specific fluorescent labeled antibodies. After co-immunization, on days 1, 3, 7 and 14, CD4.sup.+CD25'' T cells were isolated from F, V+F and pcDF100+F immunized mice and intracellular cytokine production for IL-10, IFN-y and IL-4 expression was assessed by flow cytometry. CD4.sup.+CD25'' Tr cells isolated from F, V+F, pcDF100+F and naive mice as controls were analyzed for expression of CD69, CD44 and CD62L, and for their expression of FoxP3. Tr cells express T-bet but not gata-3. On day 14 after immunization, total RNA was extracted from CD4.sup.+CD25'' T cells from all three groups and RT-PCR was used to test the expression of HPRT, T-bet and gata-3. Over the course of 14 days, the CD4.sup.+CD25'' T cells isolated from mice co-immunized with pcDF100+F expressed high levels of IL-10, IFN-y, FoxP3, and a minimal amount of IL-4. In contrast, CD4.sup.+CD25'' T cells produced a higher level of IL-4 and no expressions of FoxP3, IL-10, or IFN-y from the mice immunized with F or V+F. Since the transcriptional factor Foxp3, has been demonstrated to be a hallmark of the Tr cells, the co-vaccination induced CD4.sup.+CD25''Tr cells can be categorized into the regulatory class of T cells but they have a unique phenotype. T cell activation markers are expressed equally high including CD44 and CD69 and low for CD62L among the immunized groups, suggesting that the induced CD4.sup.+CD25' Tr cells are fully activated by the immunization.
[0215] To analyze the Th phenotype for the induced CD4.sup.+CD25'' Tr cells based on the observed cytokine expression patterns as described above, the expression of both T-bet and gata-3 genes of the CD4.sup.+CD25'' T cells from mice immunized with F, V+F or pcDF100+F were analyzed by the RT-PCR method. The results show that the CD4.sup.+CD25'' T cells from the pcDF100+F immunized mice, but not from the F or V+F immunized mice, expressed higher level of T-bet, a hallmark for Th1 cells. In the contrast, the CD4.sup.+CD25'' T cells from the F and V+F immunized mice expressed higher levels of gata-3, a characteristic Th2 cells.
[0216] Collectively, these data demonstrate that CD4.sup.+CD25'' T cell induced by the co-immunization of DNA and protein vaccines has as an adaptive Th1 phenotypic Tr cell which can suppress the antigen-specific CD4.sup.+ T cell proliferative function.
[0217] Since antigen presenting cell (APC) activates T cells to promote adaptive immunity, the induction of CD4.sup.+CD25'' Tr cell is apparently through specific APC activation by the co-inoculation of DNA+protein vaccines. To investigate this question, a similar experiments described as above was set up to assess dendritic cell (DC) function and phenotype and their influence on naive T cells. The effects of co-inoculation on the maturation of DC was analyzed. Expression of costimulatory molecules on DCs from co-immunized mice was examined. Splenocytes were isolated and stained for expression of surface markers on DCs by gating on CD11 positive cells 48 h after co-immunization of pcDF100+F, V+F or F. Co-immunization of pcDF100+F did not affect maturation of DCs as DCs isolated from the spleen 48 h after the co-immunization expressed high and similar levels of CD80, CD86, MHC II, IL-12, IL-6, IL-la/f3, IFN-a/8 and TNF-a which are the characteristics of matured DCs; whereas these molecules on the immature DCs from naive control mice remain expressed at relatively low levels, suggesting the co-inoculation enables the immature DC be induced to mature. However, we observed that the level of CD40 expression was dramatically reduced in pcDF100+F co-immunized mice compared to all other groups, suggesting a unique phenotype of DC may be involved in the induction of the observed Tr. DCs from mice immunized with V+F or F were observed to have the ability to activate heterogeneous T cells to proliferate; whereas the Immature DCs of naive mice have no such ability as expected (FIG. 4A). Interestingly, DCs from mice co-immunized with pcDF100+F had a restricted capacity to activate T cells to proliferate (FIG. 4B), suggesting that an alternative mechanism of Tr induction is induced in the co-immunization group. To explore if the DC of co-immunized mice are able to convert naive T cells to a Tr phenotype, DCs obtained from mice after their being co-immunized with pcDF100+F were co-cultured with syngeneic naive CD4.sup.+ T cells in vitro and subsequently characterized the resulting CD4.sup.+ T cells by FACS. These T cells upregulated higher levels of CD44, and CD69, but lower levels of CD62L, suggesting that the DC again has the capacity to activate T cells. Similar results were obtained from analysis of DCs from V+F or F co-immunized mice. Furthermore, the activated CD4.sup.+T cells in the pcDF100+F group produced significant higher level of IL-10 and IFN-y, but reduced IL-4 (FIG. 4B). However, activated T cells from V+F or F immunized groups produce significant level of IL-4, but little IL-10 and IFN-y (FIG. 4B). The cytokine production were analyzed after three rounds of re-stimulation with fresh DCs isolated from the D+F immunized mice. These studies demonstrated an increase in the production of IL-10 in T-cells, which is one of characteristic of regulatory T cells. To further characterize their regulatory function, IL-10.sup.+ T cells were isolated after DC stimulation and subjected to MLR to see if they block responder T cell proliferation in the MLR. As shown, the proliferation of responder T cells were inhibited by the presence of IL-10.sup.+ T cells, but not from T cells isolated from co-culture with DCs of mice immunized with V+F and F, or the control animals (FIG. 4C). These data demonstrate that DCs from the D+F co-immunized mice develop large populations of T regulatory cells in vitro.
[0218] To demonstrate the same conversion in vivo, DCs collected from BALB/c mice after co-immunization with pcDFlOO+F were co-transferred with syngenic naive CD4.sup.+T cells into nude mice (nu/nu) and subsequently analyzed these transferred T cells by FACS for intracellular staining for IL-lO and T reg markers. DCs from pcDFlOO+F co-immunized mice induced Tr cells in vivo. DCs isolated from spleens of pcDFlOO+F, V+F, F immunized or naive control mice were co-adoptive-transfer with naive CD4.sup.+CD25.sup.-T cells. The T cells were analyzed for IL-10, IFN-.gamma., IL-4, FoxP3 and CD25 on day 3 and 7. Co-transferred T cells expressed more IL-10, FoxP3 and IFN-.gamma., but little IL-4. However, co-transferred T cells with DCs from V+F or F immunized mice expressed higher levels of IL-4, but little IL-10 and IFN-.gamma., supporting the in vitro data. Specific IL-10 cytokine production was elevated after two rounds of re-stimulation with the DCs freshly isolated from D+F immunized mice. These data demonstrate that DCs from D+F co-immunized mice drive likely naive T cells into Tr cells in vivo. Consistent with previous results, such conversion was only made within the CD4.sup.+cD25'' population since the frequency of CD25.sup.+ was not observed to be influenced testing vivo.
[0219] Finally, experiments were performed to identify what molecules in the interaction between DC and T cells play a role in the development of Tr. Since the Tr is within the activated T cell compartment as demonstrated above, the signals should include classic activation pathway signals. Important among these is the up-regulation of major histocompatibility complex (MHC) class II and co-stimulatory molecules (CD80/CD86), which provide the two requisite signals for naive T cell activation. To study this issue, DCs were isolated from spleen of mice 48 h after pcDF100+F co-immunization and co-cultured with naive CD4.sup.+ T cells from naive syngeneic mice in the presence or absence of reagents to block signaling molecules including anti-CD80, anti-CD86, anti-CD40 and anti-MHC-II. After seven days of re-stimulation, T cells were isolated and added into MLR system to examine their regulatory functions. The results showed that both anti-CD40 and anti-MHC II mAbs can partially reverse the suppressive effects on MLR by the Tr cells; whereas mAbs against CD80, CD86 and had no ability to block the induction of the suppression phenotype. These results demonstrate that both CD40 and MHC II signals play roles in the induction of Tr in the flea allergen Induced immediate hypersensitivity model.
[0220] Adaptive Trl cells have been observed and induced by DCs processing a suboptimal immunogen or subimmunogen to inhibit normal T cells' function mediated via secretion of IL-10 or TGF-b or both. It has been further demonstrated that immature DCs drive the differentiation of IL-10-producing Trl cells with producing IL-10, TGF-R, and IFN-y, but they do not produce IL-4 or IL-2, they are hyporesponsive to antigenspecific and polyclonal activation. In additional evidence has shown that suboptimal activation of DC by a minute antigen stimulation can induces the Trl conversion. This stimulation of DCs lacks the co-stimulation signaling and thus presents a tolerance signal to T cells.
[0221] Co-inoculation of DNA encoding a flea antigen with flea protein induces adaptive Tr cells which inhibit the allergic reaction induced by flea allergen challenge. The cells exhibit a phenotype of CD4.sup.+CD25''Foxp3.sup.+ and suppress in vivo and in vitro antigen specific as well as MLR immune responses through the production of IL-10 and IFN-y. MHC-ll.sup.+/CD40.sup.low DC populations are induced by such co-immunization and in turn to convert naive T cells into Tr cells.
Example 17
Testing the Prevention and/or Therapeutic Approaches Against FAD Allergen Through Co-Immunization with Vaccines in a Feline FAD Model Establishing the Feline Model and Testing Co-Immunization
[0222] There are more than 15 kinds of allergen in the flea saliva. Among them, an 18 kDa protein, flea saliva antigen 1 (FSA1), has been determined a main allergen which can cause FAD. This gene has been cloned and expressed it in the E. coli system. In addition, the pVAX-FSA1 eukaryotic expressing construct has also been prepared.
[0223] A cat FAD model was established and subsequently used to demonstrate that the therapeutic effect of co-immunization of FSA1+pVAX-FSA1 on the established FAD in cats.
Determination of Number of Fleas, Duration of a Cycle
Animal and Parasite
[0224] Ten pathogen-free cats were purchased from North China Pharmaceutical Group (Shijiazhuang, Hebei) and housed animal facility at the Center for Disease Control and Prevention of China (CDC) in the course of experiment. All cats were over one year old, which is an important factor since the younger animals may be tolerable to the flea allergen in this experiment. The cats were grouped with breed and sex randomly. The sterile fleas were supplied by the CDC.
[0225] To determine relationship of infestation and FAD symptom, 10 cats were separated into three groups, including an experimental group with six cats and two control groups with two cats each. One control group was treated nitenpyram, and another was not treated with the nitenpyram. Each experimental cat was lived separated on day 0 and infestated with 100 fleas. After two days, all cats in this group were given the nitenpyram to remove the flea from their bodies. Two of the control cats were also given this medicine on day 0. This challenge cycle was repeated every other week for 7 times
Immunization Scheme for the FAD Induced or Control Cats with FSA1 and pVAX-FSA1 as Co-Immunogens
[0226] The 6 FAD cats were separated into three groups. Two of them were co-immunized with 400 u.g FSA1 i.p. and 400 u,g pVAX-FSA1 plasmid at double lateral abdomen subcutaneously. Two were co-immunized with 400 u.g FSA1 i.p. and 400 u.g pVAX vector subcutaneously. The last two were not immunized and used as the positive control group. The 4 un-FAD cats were separated into two groups. Two were co-immunized with 400 u.g FSA1 i.p. and 400( g pVAX-FSA1 plasmid at double lateral abdomen subcutaneously and the other two were immunized with 400 u.g FSA1 i.p. and 400 u.g pVAX vector subcutaneously. The two control cats with nitenpyram were sent into different groups. The cats were immunized three times: at days 0, 9, and 16. After that, the cats were challenged for six cycles as above. During the second immunization, a cycle of challenge was done at the same time, because we wanted to keep positive cats at the susceptible state for next therapy. The program of immunization was as Table 2.
TABLE-US-00002 TABLE 2 Immunization Cat PSA1 + pVAX- PSA1 + pVAX Treat with No. FSA1 vector nitenpyram prophylactive 1 - + + 2 + - - 4 + - + 5 - + - therapeutic 8 + - + 9 - + + 10 - + + 11 + - + Positive control 3 - - + 7 - - +
Dermatological Scores.
Method:
[0227] Cats were scored by the dermatological assessments two days after ceasing each infestation. The assessments were included erythema, papules, crusts, scale, alopecia, excoriation. The body of each cat was divided into three portions to assess which part might have the most severe clinical outcome. According the previous documented report, three parts were consisted of: (1) back, from the head to the tail of dorsal surface; (2) double lateral abdomen, from the scapula to the tail; (3) chest and underside, from laryngeal to the caudomedial thighs.
Smearing Counts of Peripheral Blood Cells from the Cats
Method:
[0228] The anti-coagulant peripheral blood at 2 ml was collected on days 0, 2, 16, 30, 44, 58, 62 from the experimental group and day 0, 14, 28, 42, 56, 60 from the control groups. The same samples were collected after the immunization. A drop of blood was smeared on a clean glass slide. After the smear was dried, the cells were stained with Wright-Giemsa stain for 15 min. After a rinse in deionized water, the smear was gently dried with Kimwipe paper and dehydrated by 96% and 100% ethanol with each 10 seconds treatment. The slide was then treated with xylene for 30 min.
[0229] After the staining, nucleus and cytoplasm of blood cell was distinguished by a blue and pink staining. The percentage of each type of cells was counted by a total number of 200 cells in a view under a light microscope.
Therapeutic Approaches:
[0230] The same methods as above were used except that total RNA was extracted from mononuclear cells which were isolated from peripheral blood on day 7 post the third immunization from each group.
Statistical Analysis
[0231] Analysis of variance (ANOVA) was used to detect the differences which included dermatology scores from each cycle, scores of various lesions and scores on different sites among three groups. The differences of skin test and IgE level were determined also by ANOVA. Statistical analyses of other ones were performed using the Student's/-test. In these analyses, data were converted into logarithmic plot. If the P<0.05, indicates a significant difference.
[0232] Statistical analyses of were also performed using the Student's/-test. In these analyses, the data was converted into log. If the PO.05, the data indicated significant differences.
Results:
Observation and Dermatological Scores of the Cats
[0233] Lesions were scored according to their types, locations, sizes and numbers.
[0234] The total scores were analyzed from the groups, and the lesions and sites for the experimental group were assessed. The FAD group registered more scores than in other groups. The scores were increased at the fourth infestation cycle, and then stayed at the level of 5.0. The scores in the FAD group were significant higher than in other control groups (P<0.01). These results support the conclusion that the cat FAD model was valid and feasible to evaluate a therapeutic or prophylactic treatment against the allergic reaction.
[0235] To assess where the lesions occur most, what type, and when to occur, statistic analysis was done and it was found that papules and fur loss were the most common factors to contribute to the dermatology scores in FAD group. The reading of erythema was not a contributing factor since the erythema reached to the peak on day 44, but it fallen at the end of this experiment, indicating no persistency. On the other hand, the papules were increased, and remained at a high level after 44 days and reached to the peak on the day 86 (P<0.05), suggesting its persistency. Similarly, fur loss was maintained at high level after day 44 (P<0.05). These two readouts are indicated as a good onset to reflect FAD. The scores from other lesions were randomly distributed and inconsistent in the two control groups, in which no significant difference of lesions were observed.
[0236] Although, most of dermal lesions tended to be located on the backs and heads compared other sites on day 44 (P<0.05), the lesions were extended all over the body at the end of the experiment. This observation was different from the previous reports in dog, of which the chest and underside were tended to have the most lesions. That may owe to the habitual differences of the cats and dogs.
[0237] To eliminate the interference by the nitenpyram treatment, any differences between these treated with nitenpyram and the ones without treatment were assessed. From the dermatology score, no significant difference was seen in the control groups. Thus, the nitenpyram treatment did not influence the results obtained from our experiments (FIG. 5).
Cell Percentage in Peripheral Blood by Blood Smear Analysis
[0238] A comparison of the number of each type of cells in peripheral blood from the different groups showed that the number of eosinophils was higher in the experimental group than those in control groups. Blood smears were performed for each infestation cycle, but the percent of each type of cells became constant after the third cycle. As the result from day 58 show in Table 3, the significant changes occurred in the number of eosinophils among the three groups. Since the increase of eosinophils is related to allergic diseases as previously documented, these results indicate that the cats in the experimental group were more susceptible to the infestation of fleas. There was no obvious difference between the two control groups, the nitenpyram seems not to interfere the cell numbers in the cats' peripheral blood.
TABLE-US-00003 TABLE 3 Cell types and their percentage in peripheral blood Lympho- Mono- Neutro- Baso- Eosino- cytes cytes phils phils phils (%) (%) <%) (%) (%) Experimental 23.4 2.6 61.5 1.0 11.5 group Control group 21.8 3.6 70.5 0.7 3.4 with nitenpyram Control group 23.1 3.6 68.3 0.6 4.4 without nitenpyram
Histopathological Examination
[0239] First, the differences between the normal skins and those with lesions were analyzed. Skin biopsies were chosen from all groups on day 58. However, the skin biopsies from one cat may also display differences in some degree, especially those with lesions. That was not enough to indicate whether the cats infestated with fleas were induced to have FAD. For this reason, the skin biopsies from each group were collected after the IDT with the flea extracts since IDT with flea provides an antigen specific recalled immune responses.
Skin Test to Evaluate Whether the Cats were Allergic to the Flea Extracts.
[0240] The diameter of wheal or bleb was measured 15 min after the IDT injection. Since each cat had a different level of sensitivity to the IDT, the results from each individual animal were recorded. FIG. 6 shows the skin reactions in all groups. An allergic reaction was considered to have occurred if an IDT value was above the average of threshold (the sum of saline and histamine is divided by 2), or not occurred if an IDT value was below the average of the threshold. The results are shown in Table 4. Cats with dermatology scores at 5.0 or above were identified as the positive for FAD.
TABLE-US-00004 TABLE 4 Comparison of dermatological assessment with the intradermal skin test (IDT). Control Control group Positive Positive Positive skin Positive Positive Cat Experimental group with without Dermatology clinical skin test test with skin test skin test No. group nitenpyram nitenpyram scores scores with BSA flea extract with FS with FSA1 I No Yes No 0 - - - - - 2 No No Yes 1 - - - - - 4 No Yes No 1 - - - - - 5 No No Yes 0 - - - - - 3 Yes No No 7 + - + - + 7 Yes No No 6 + - + - - 8 Yes No No 8 + - + - + 9 Yes No No 2 - - + - + 10 Yes No No 6 + - + - + 11 Yes No No 3 - - + - +
Skin Test to Determine the Effect of Co-Immunization of FSA1 and pVAX-FSA1
[0241] The cats with FAD were co-immunized with FSA1+pVAX-FSA1 or vector+FSA1 as described in experimental design B in Table 2. Before and after the immunizations, the cats were tested by IDT with various flea antigens or control antigens. The IDT readings were recorded on 7 days after the last immunization as summarized in FIG. 7 and Table 5.
[0242] The results showed that FAD cats co-immunized with the FSA1+pVAX-FSA1 had less skin-reactions to the flea extracts or flea specific antigens (such as FSA1 protein) challenges; whereas the cats immunized with vector+FSA1 had more skin-reactions to the same challenges.
[0243] Comparing the differences before and after immunization, we observed that the skin reaction was much smaller after the immunization than before in the co-immunized group as shown FIG. 7, suggesting the co-immunization significant decreased sensitivity to the flea challenge. On contrary, no significant effect was seen in groups immunized with FSA1 and pVAX vector, indicating that the cats were remained their allergic status. The status of all cats was listed in Table 5.
TABLE-US-00005 TABLE 5 FSA1 FSAl + IDT IDT IDT The state for Cat Positive and pVAX - with with flea with FE before No. control pVAX FSAl BSA extracts FSA1 immunization I No Yes No - 2 No No Yes - 4 No No Yes - 5 No Yes No - 3. Yes No No - + - + 7 Yes No No - + - + 8 No No Yes - - - + 9 No Yes No - + - + 10 No Yes No - + - + II No No Yes - - - +
Therapeutic Effects of Co-Immunization on the FAD Cats
[0244] To determine the therapeutic effect of co-immunization on lesions of the FAD cats, the lesions were recorded over period of 53 days after the first immunization and shown in FIG. 8. The lesion scores in the co-immunized group were dramatically reduced from 5.5 to 2 (FIG. 8, square points on line). Whereas, the effects on the group immunized with FSA1 and pVAX vector was reduced but to a lesser extent (FIG. 8, triangle points on line). The result suggested the co-immunization had a therapeutic effect on the established FAD in cats.
Correlation of the Type of Lesions Affected by the Co-Immunization on the FAD Cats
[0245] To correlate which type of lesions reduced with the co-immunization, the scores on each type of lesions in every group were analyzed and the results are shown in FIG. 9A-9F. Only the score of the papules were reduced from 4.0 to 0.5 and coincident with the co-immunization of FSA1+pVAX-FSA1 (FIG. 9B, square points on line). Other type of lesions in experimental group was remained unchanged.
Correlation of the Location of Lesions Affected by the Co-Immunization on the FAD Cats Therapeutic Effects of Co-Immunization on the FAD Cats
[0246] After analysis the correlation, the data showed that the scores of double lateral abdomen in therapeutic group were reduced from 1.5 to 0 (FIG. 10B) after the co-immunization of FSA1+pVAX-FSA1, but no effect in the other places as shown in FIGS. 10A-10C. The therapeutic group immunized with FSA1 and pVAX vector showed a lingering response (FIG. 10B, triangle points). The dermatologic scores of FAD cats immunized with FSA1 and pVAX vector were reduced from 1.5 to 0.5 after 7 days of the last immunization. In contrast, those in the co-immunized with FSA1+pVAX-FSA1 were reduced promptly after its first immunization and remained at low level thorough (FIG. 10B, square points).
SUMMARY
[0247] These data demonstrate the successful induction of a FAD model in feline by flea infestations. Physiological or pathogenic parameters in the FAD feline have been characterized which can be used as a model to evaluate treatment of immunotherapeutic or prophylactic approaches.
[0248] The co-immunization of FSA1+pVAX-FSA1 vaccines was demonstrated to suppress the established FAD in cats. Such suppression seems to be an antigenic specific, which supports the results observed in mouse studies.
Example 18
pVAXI-K-FSA1
[0249] Plasmid pVAX1-K-FSA1 comprises the FSA1 coding sequence linked to a Kozak sequence in plasmid backbone pVAX (Invitrogen). The sequence of the K-FSA1 insert is SEQ ID NO:61. Nucleotides 1-9 correspond to the Kozak sequence, the open reading from of FSA1 plus missed 8 amino acids begins at nucleotide 10. A map of the plasmid pVAX1-K-FSA1 is shown in FIG. 11.
Sequence CWU
1
1
611640DNACtenocephalides felis 1atgaattatt gttttttagt atttttagta
tatttagtat ttgcagttaa tggggaagat 60atttggaaag ttaataaaaa atgtacatca
ggtggaaaaa atcaagatag aaaactcgat 120caaataattc aaaaaggcca acaagttaaa
atccaaaata tttgcaaatt aatacgagat 180aaaccacata caaatcaaga gaaagaaaaa
tgtatgaaat tttgcaaaaa agtttgcaaa 240ggttatagag gagcttgtga tggcaatatt
tgctactgca gcaggccaag taatttaggt 300cctgattgga aagtaagcaa agaatgcaaa
gatcccaata acaaagattc tcgtcctacg 360gaaatagttc catatcgaca acaattagca
attccaaata tttgcaaact aaaaaattca 420gagaccaatg aagattccaa atgcaaaaaa
cattgcaaag aaaaatgtcg tggtggaaat 480gatgctggat gtgatggaaa cttttgttat
tgtcgaccaa aaaataaata ataattataa 540taaataaatt gttatagtta ttagttatcc
catcacatat tagaaaagtg gcttataatt 600tatgaacaat ataacacata aattagttgt
gtaaaaaaaa 6402176PRTCtenocephalides felis 2Met
Asn Tyr Cys Phe Leu Val Phe Leu Val Tyr Leu Val Phe Ala Val1
5 10 15Asn Gly Glu Asp Ile Trp Lys
Val Asn Lys Lys Cys Thr Ser Gly Gly 20 25
30Lys Asn Gln Asp Arg Lys Leu Asp Gln Ile Ile Gln Lys Gly
Gln Gln 35 40 45Val Lys Ile Gln
Asn Ile Cys Lys Leu Ile Arg Asp Lys Pro His Thr 50 55
60Asn Gln Glu Lys Glu Lys Cys Met Lys Phe Cys Lys Lys
Val Cys Lys65 70 75
80Gly Tyr Arg Gly Ala Cys Asp Gly Asn Ile Cys Tyr Cys Ser Arg Pro
85 90 95Ser Asn Leu Gly Pro Asp
Trp Lys Val Ser Lys Glu Cys Lys Asp Pro 100
105 110Asn Asn Lys Asp Ser Arg Pro Thr Glu Ile Val Pro
Tyr Arg Gln Gln 115 120 125Leu Ala
Ile Pro Asn Ile Cys Lys Leu Lys Asn Ser Glu Thr Asn Glu 130
135 140Asp Ser Lys Cys Lys Lys His Cys Lys Glu Lys
Cys Arg Gly Gly Asn145 150 155
160Asp Ala Gly Cys Asp Gly Asn Phe Cys Tyr Cys Arg Pro Lys Asn Lys
165 170
1753416DNACtenocephalides felis 3ggcctggcgg tgctcctgga aaaggatgtt
agacgcagcc ctcccaccct gccctactgt 60tgcggccaca gcagattgtg aaatttgccc
agccgtgaag agggatgttg acctattcct 120gacgggaacc cccgacgaat atgttgagca
agtggcacaa tacaaagcac tacctgtagt 180attggaaaat gccagaatac tgaagaactg
cgttgatgca aaaatgacag aagaggataa 240ggagaatgct ctcagcttgc tggacaaaat
atacacaagt cctctgtgtt aaaggagcca 300tcactgccag gagccctaag gaagccactg
aactgatcac taagtagtct cagcagcctg 360ccatgtccag gtgtcttact agaggattcc
agcaataaaa gccttgcaat tcaaac 416488PRTCtenocephalides felis 4Met
Leu Asp Ala Ala Leu Pro Pro Cys Pro Thr Val Ala Ala Thr Ala1
5 10 15Asp Cys Glu Ile Cys Pro Ala
Val Lys Arg Asp Val Asp Leu Phe Leu 20 25
30Thr Gly Thr Pro Asp Glu Tyr Val Glu Gln Val Ala Gln Tyr
Lys Ala 35 40 45Leu Pro Val Val
Leu Glu Asn Ala Arg Ile Leu Lys Asn Cys Val Asp 50 55
60Ala Lys Met Thr Glu Glu Asp Lys Glu Asn Ala Leu Ser
Leu Leu Asp65 70 75
80Lys Ile Tyr Thr Ser Pro Leu Cys 855410DNACtenocephalides
felis 5ctgcatcatg aagggggctc gtgttctcgt gcttctctgg gctgccttgc tcttgatctg
60gggtggaaat tgtgaaattt gcccagccgt gaagagggat gttgacctat tcctgacggg
120aacccccgac gaatatgttg agcaagtggc acaatacaaa gcactacctg tagtattgga
180aaatgccaga atactgaaga actgcgttga tgcaaaaatg acagaagagg ataaggagaa
240tgctctcagc ttgctggaca aaatatacac aagtcctctg tgttaaagga gccatcactg
300ccaggagccc taaggaagcc actgaactga tcactaagta gtctcagcag cctgccatgt
360ccaggtgtct tactagagga ttccagcaat aaaagccttg caattcaaac
410692PRTCtenocephalides felis 6Met Lys Gly Ala Arg Val Leu Val Leu Leu
Trp Ala Ala Leu Leu Leu1 5 10
15Ile Trp Gly Gly Asn Cys Glu Ile Cys Pro Ala Val Lys Arg Asp Val
20 25 30Asp Leu Phe Leu Thr Gly
Thr Pro Asp Glu Tyr Val Glu Gln Val Ala 35 40
45Gln Tyr Lys Ala Leu Pro Val Val Leu Glu Asn Ala Arg Ile
Leu Lys 50 55 60Asn Cys Val Asp Ala
Lys Met Thr Glu Glu Asp Lys Glu Asn Ala Leu65 70
75 80Ser Leu Leu Asp Lys Ile Tyr Thr Ser Pro
Leu Cys 85 907525DNACtenocephalides canis
7atgaagaccc tgctcctcac catcggcttc agcctcattg cgatcctgca ggcccaggat
60accccagcct tgggaaagga cactgtggct gtgtcaggga aatggtatct gaaggccatg
120acagcagacc aggaggtgcc tgagaagcct gactcagtga ctcccatgat cctcaaagcc
180cagaaggggg gcaacctgga agccaagatc accatgctga caaatggtca gtgccagaac
240atcacggtgg tcctgcacaa aacctctgag cctggcaaat acacggcata cgagggccag
300cgtgtcgtgt tcatccagcc gtccccggtg agggaccact acattctcta ctgcgagggc
360gagctccatg ggaggcagat ccgaatggcc aagcttctgg gaagggatcc tgagcagagc
420caagaggcct tggaggattt tcgggaattc tcaagagcca aaggattgaa ccaggagatt
480ttggaactcg cgcagagcga aacctgctct ccaggaggac agtag
5258174PRTCtenocephalides canis 8Met Lys Thr Leu Leu Leu Thr Ile Gly Phe
Ser Leu Ile Ala Ile Leu1 5 10
15Gln Ala Gln Asp Thr Pro Ala Leu Gly Lys Asp Thr Val Ala Val Ser
20 25 30Gly Lys Trp Tyr Leu Lys
Ala Met Thr Ala Asp Gln Glu Val Pro Glu 35 40
45Lys Pro Asp Ser Val Thr Pro Met Ile Leu Lys Ala Gln Lys
Gly Gly 50 55 60Asn Leu Glu Ala Lys
Ile Thr Met Leu Thr Asn Gly Gln Cys Gln Asn65 70
75 80Ile Thr Val Val Leu His Lys Thr Ser Glu
Pro Gly Lys Tyr Thr Ala 85 90
95Tyr Glu Gly Gln Arg Val Val Phe Ile Gln Pro Ser Pro Val Arg Asp
100 105 110His Tyr Ile Leu Tyr
Cys Glu Gly Glu Leu His Gly Arg Gln Ile Arg 115
120 125Met Ala Lys Leu Leu Gly Arg Asp Pro Glu Gln Ser
Gln Glu Ala Leu 130 135 140Glu Asp Phe
Arg Glu Phe Ser Arg Ala Lys Gly Leu Asn Gln Glu Ile145
150 155 160Leu Glu Leu Ala Gln Ser Glu
Thr Cys Ser Pro Gly Gly Gln 165
1709791DNACtenocephalides canis 9agagctggac ccgtgtgtgt gctggccaat
gagccctgga gggtccggct ccagagtacc 60ctcttggcac agggccgagt ccatcgggac
agatgaacct agaggactcc actgccctcc 120catccacggg gccgggtcac cagactctgc
aagtctccag ctgtcgccaa acccagacag 180aaggtgctgt ggacatgcag ctcctactgc
tgaccgtggg cctggcactg atctgtggcc 240tccaggctca ggagggaaac catgaggagc
cccagggagg cctagaggag ctgtctggga 300ggtggcactc cgttgccctg gcctccaaca
agtccgatct gatcaaaccc tgggggcact 360tcagggtttt catccacagc atgagcgcaa
aggacggcaa cctgcacggg gatatcctta 420taccgcagga cggccagtgc gagaaagtct
ccctcactgc gttcaagact gccaccagca 480acaaatttga cctggagtac tggggacaca
atgacctgta cctggcagag gtagacccca 540agagctacct gattctctac atgatcaacc
agtacaacga tgacaccagc ctggtggctc 600acttgatggt ccgggacctc agcaggcagc
aggacttcct gccggcattc gaatctgtat 660gtgaagacat cggtctgcac aaggaccaga
ttgtggttct gagcgatgac gatcgctgcc 720agggttccag agactagggc ctcagccacg
cagagagcca agcagcagga tctcacctgc 780ctgagtacgg t
79110180PRTCtenocephalides canis 10Met
Gln Leu Leu Leu Leu Thr Val Gly Leu Ala Leu Ile Cys Gly Leu1
5 10 15Gln Ala Gln Glu Gly Asn His
Glu Glu Pro Gln Gly Gly Leu Glu Glu 20 25
30Leu Ser Gly Arg Trp His Ser Val Ala Leu Ala Ser Asn Lys
Ser Asp 35 40 45Leu Ile Lys Pro
Trp Gly His Phe Arg Val Phe Ile His Ser Met Ser 50 55
60Ala Lys Asp Gly Asn Leu His Gly Asp Ile Leu Ile Pro
Gln Asp Gly65 70 75
80Gln Cys Glu Lys Val Ser Leu Thr Ala Phe Lys Thr Ala Thr Ser Asn
85 90 95Lys Phe Asp Leu Glu Tyr
Trp Gly His Asn Asp Leu Tyr Leu Ala Glu 100
105 110Val Asp Pro Lys Ser Tyr Leu Ile Leu Tyr Met Ile
Asn Gln Tyr Asn 115 120 125Asp Asp
Thr Ser Leu Val Ala His Leu Met Val Arg Asp Leu Ser Arg 130
135 140Gln Gln Asp Phe Leu Pro Ala Phe Glu Ser Val
Cys Glu Asp Ile Gly145 150 155
160Leu His Lys Asp Gln Ile Val Val Leu Ser Asp Asp Asp Arg Cys Gln
165 170 175Gly Ser Arg Asp
180111099DNADermatophagoides pteronyssinus 11gaattccttt
ttttttcttt ctctctctaa aatctaaaat ccatccaaca tgaaaattgt 60tttggccatc
gcctcattgt tggcattgag cgctgtttat gctcgtccat catcgatcaa 120aacttttgaa
gaatacaaaa aagccttcaa caaaagttat gctaccttcg aagatgaaga 180agctgcccgt
aaaaactttt tggaatcagt aaaatatgtt caatcaaatg gaggtgccat 240caaccatttg
tccgatttgt cgttggatga attcaaaaac cgatttttga tgagtgcaga 300agcttttgaa
cacctcaaaa ctcaattcga tttgaatgct gaaactaacg cctgcagtat 360caatggaaat
gctccagctg aaatcgattt gcgacaaatg cgaactgtca ctcccattcg 420tatgcaagga
ggctgtggtt catgttgggc tttctctggt gttgccgcaa ctgaatcagc 480ttatttggct
taccgtaatc aatcattgga tcttgctgaa caagaattag tcgattgtgc 540ttcccaacac
ggttgtcatg gtgataccat tccacgtggt attgaataca tccaacataa 600tggtgtcgtc
caagaaagct actatcgata cgttgcacga gaacaatcat gccgacgacc 660aaatgcacaa
cgtttcggta tctcaaacta ttgccaaatt tacccaccaa atgtaaacaa 720aattcgtgaa
gctttggctc aaacccacag cgctattgcc gtcattattg gcatcaaaga 780tttagacgca
ttccgtcatt atgatggccg aacaatcatt caacgcgata atggttacca 840accaaactat
cacgctgtca acattgttgg ttacagtaac gcacaaggtg tcgattattg 900gatcgtacga
aacagttggg ataccaattg gggtgataat ggttacggtt attttgctgc 960caacatcgat
ttgatgatga ttgaagaata tccatatgtt gtcattctct aaacaaaaag 1020acaatttctt
atatgattgt cactaattta tttaaaatca aaatttttag aaaatgaata 1080aattcattca
caaaaatta
109912320PRTEscherichia coli 12Met Lys Ile Val Leu Ala Ile Ala Ser Leu
Leu Ala Leu Ser Ala Val1 5 10
15Tyr Ala Arg Pro Ser Ser Ile Lys Thr Phe Glu Glu Tyr Lys Lys Ala
20 25 30Phe Asn Lys Ser Tyr Ala
Thr Phe Glu Asp Glu Glu Ala Ala Arg Lys 35 40
45Asn Phe Leu Glu Ser Val Lys Tyr Val Gln Ser Asn Gly Gly
Ala Ile 50 55 60Asn His Leu Ser Asp
Leu Ser Leu Asp Glu Phe Lys Asn Arg Phe Leu65 70
75 80Met Ser Ala Glu Ala Phe Glu His Leu Lys
Thr Gln Phe Asp Leu Asn 85 90
95Ala Glu Thr Asn Ala Cys Ser Ile Asn Gly Asn Ala Pro Ala Glu Ile
100 105 110Asp Leu Arg Gln Met
Arg Thr Val Thr Pro Ile Arg Met Gln Gly Gly 115
120 125Cys Gly Ser Cys Trp Ala Phe Ser Gly Val Ala Ala
Thr Glu Ser Ala 130 135 140Tyr Leu Ala
Tyr Arg Asn Gln Ser Leu Asp Leu Ala Glu Gln Glu Leu145
150 155 160Val Asp Cys Ala Ser Gln His
Gly Cys His Gly Asp Thr Ile Pro Arg 165
170 175Gly Ile Glu Tyr Ile Gln His Asn Gly Val Val Gln
Glu Ser Tyr Tyr 180 185 190Arg
Tyr Val Ala Arg Glu Gln Ser Cys Arg Arg Pro Asn Ala Gln Arg 195
200 205Phe Gly Ile Ser Asn Tyr Cys Gln Ile
Tyr Pro Pro Asn Val Asn Lys 210 215
220Ile Arg Glu Ala Leu Ala Gln Thr His Ser Ala Ile Ala Val Ile Ile225
230 235 240Gly Ile Lys Asp
Leu Asp Ala Phe Arg His Tyr Asp Gly Arg Thr Ile 245
250 255Ile Gln Arg Asp Asn Gly Tyr Gln Pro Asn
Tyr His Ala Val Asn Ile 260 265
270Val Gly Tyr Ser Asn Ala Gln Gly Val Asp Tyr Trp Ile Val Arg Asn
275 280 285Ser Trp Asp Thr Asn Trp Gly
Asp Asn Gly Tyr Gly Tyr Phe Ala Ala 290 295
300Asn Ile Asp Leu Met Met Ile Glu Glu Tyr Pro Tyr Val Val Ile
Leu305 310 315
32013717DNAArachis hypogaea 13gctcaccata ctagtagccc tcgccctttt cctcctcgct
gcccacgcat ctgcgaggca 60gcagtgggaa ctccaaggag acagaagatg ccagagccag
ctcgagaggg cgaacctgag 120gccctgcgag caacatctca tgcagaagat ccaacgtgac
gaggattcat atgaacggga 180cccgtacagc cctagtcagg atccgtacag ccctagtcca
tatgatcgga gaggcgctgg 240atcctctcag caccaagaga ggtgttgcaa tgagctgaac
gagtttgaga acaaccaaag 300gtgcatgtgc gaggcattgc aacagatcat ggagaaccag
agcgataggt tgcaggggag 360gcaacaggag caacagttca agagggagct caggaacttg
cctcaacagt gcggccttag 420ggcaccacag cgttgcgact tggacgtcga aagtggcggc
agagacagat actaaacacc 480tatctcaaaa aaagaaaaga aaagaaaaga aaatagctta
tatataagct attatctatg 540gttatgttta gttttggtaa taataaagat catcactata
tgaatgtgtt gatcgtgtta 600actaaggcaa gcttaggtta tatgagcacc tttagagtgc
ttttatggcg ttgtctatgt 660tttgttgctg cagagttgta accatcttga aataatataa
aaagatcatg ttttgtt 71714766DNAArachis hypogaea 14agaaagagaa
gacaagatgt cgtggcaaac ctacgtcgat aaccaccttc tctgcgaaat 60tgaaggcgac
cacctctcct ccgccgcaat cctcggccaa gacggcggtg tttgggctca 120gagctctcat
ttccctcagt tcaagcctga ggaaattact gctatcatga acgactttgc 180tgagcctgga
tcgctcgccc ctaccgggtt gtacctcggt ggcaccaaat acatggttat 240ccaaggtgaa
cccggagcta tcattccagg gaagaagggt cctggtggtg ttaccattga 300gaagacgaat
caggcgttaa tcatcggaat ctacgataag ccaatgactc cggggcagtg 360caacatgatt
gttgaaaggc tgggtgatta tctcattgat acgggtcttt aagtcctctt 420tgttatttct
tgttatctgc ttgcttattt cactggctcc tatacgaggc ttcgcatcga 480tgtgccaaga
gaatgctcga ttgtagtgta ataatattaa ttgatgggta ttcaaaagtc 540atgggatctg
cgtctaggga agaagttatg gtgcttgaga agtgaatgat aactatcatc 600tctgttgttg
tgctttttag cgggtatctg tatacaattt acaagtggtt ttaatgctgt 660gggcataaat
gggcattaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 720aaaaaaaaaa
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaa
76615131PRTArachis hypogaea 15Met Ser Trp Gln Thr Tyr Val Asp Asn His Leu
Leu Cys Glu Ile Glu1 5 10
15Gly Asp His Leu Ser Ser Ala Ala Ile Leu Gly Gln Asp Gly Gly Val
20 25 30Trp Ala Gln Ser Ser His Phe
Pro Gln Phe Lys Pro Glu Glu Ile Thr 35 40
45Ala Ile Met Asn Asp Phe Ala Glu Pro Gly Ser Leu Ala Pro Thr
Gly 50 55 60Leu Tyr Leu Gly Gly Thr
Lys Tyr Met Val Ile Gln Gly Glu Pro Gly65 70
75 80Ala Ile Ile Pro Gly Lys Lys Gly Pro Gly Gly
Val Thr Ile Glu Lys 85 90
95Thr Asn Gln Ala Leu Ile Ile Gly Ile Tyr Asp Lys Pro Met Thr Pro
100 105 110Gly Gln Cys Asn Met Ile
Val Glu Arg Leu Gly Asp Tyr Leu Ile Asp 115 120
125Thr Gly Leu 130161295DNACryptomeria japonica
16aatctgctca taatcatagc atagccgtat agaaagaaat tctacactct gctaccaaaa
60aatggattcc ccttgcttag tagcattact ggttttctct tttgtaattg gatcttgctt
120ttctgataat cccatagaca gctgctggag aggagactca aactgggcac aaaacagaat
180gaagctcgca gattgtgcag tgggcttcgg aagctccacc atgggaggca agggaggaga
240tctttatacg gtcacgaact cagatgacga ccctgtgaat cctgcaccag gaactctgcg
300ctatggagca acccgagata ggcccctgtg gataattttc agtgggaata tgaatataaa
360gctcaaaatg cctatgtaca ttgctgggta taagactttt gatggcaggg gagcacaagt
420ttatattggc aatggcggtc cctgtgtgtt tatcaagaga gttagcaatg ttatcataca
480cggtttgtat ctgtacggct gtagtactag tgttttgggg aatgttttga taaacgagag
540ttttggggtg gagcctgttc atcctcagga tggcgatgct cttactctgc gcactgctac
600aaatatttgg attgatcata attctttctc caattcttct gatggtctgg tcgatgtcac
660tcttacttcg actggagtta ctatttcaaa caatcttttt ttcaaccatc ataaagtgat
720gttgttaggg catgatgatg catatagtga tgacaaatcc atgaaggtga cagtggcgtt
780caatcaattt ggacctaact gtggacaaag aatgcccagg gcacgatatg gacttgtaca
840tgttgcaaac aataattatg acccatggac tatatatgca attggtggga gttcaaatcc
900aaccattcta agtgaaggga atagtttcac tgcaccaaat gagagctaca agaagcaagt
960aaccatacgt attggatgca aaacatcatc atcttgttca aattgggtgt ggcaatctac
1020acaagatgtt ttttataatg gagcttattt tgtatcatca gggaaatatg aagggggtaa
1080tatatacaca aagaaagaag ctttcaatgt tgagaatggg aatgcaactc ctcaattgac
1140aaaaaatgct ggggttttaa catgctctct ctctaaacgt tgttgatgat gcatatattc
1200tagcatgttg tactatctaa attaacatca acaagaaata tatcatgatg tatattgttg
1260tattgatgtc aaaataaaaa tgttctttta ctatt
129517374PRTCryptomeria japonica 17Met Asp Ser Pro Cys Leu Val Ala Leu
Leu Val Phe Ser Phe Val Ile1 5 10
15Gly Ser Cys Phe Ser Asp Asn Pro Ile Asp Ser Cys Trp Arg Gly
Asp 20 25 30Ser Asn Trp Ala
Gln Asn Arg Met Lys Leu Ala Asp Cys Ala Val Gly 35
40 45Phe Gly Ser Ser Thr Met Gly Gly Lys Gly Gly Asp
Leu Tyr Thr Val 50 55 60Thr Asn Ser
Asp Asp Asp Pro Val Asn Pro Ala Pro Gly Thr Leu Arg65 70
75 80Tyr Gly Ala Thr Arg Asp Arg Pro
Leu Trp Ile Ile Phe Ser Gly Asn 85 90
95Met Asn Ile Lys Leu Lys Met Pro Met Tyr Ile Ala Gly Tyr
Lys Thr 100 105 110Phe Asp Gly
Arg Gly Ala Gln Val Tyr Ile Gly Asn Gly Gly Pro Cys 115
120 125Val Phe Ile Lys Arg Val Ser Asn Val Ile Ile
His Gly Leu Tyr Leu 130 135 140Tyr Gly
Cys Ser Thr Ser Val Leu Gly Asn Val Leu Ile Asn Glu Ser145
150 155 160Phe Gly Val Glu Pro Val His
Pro Gln Asp Gly Asp Ala Leu Thr Leu 165
170 175Arg Thr Ala Thr Asn Ile Trp Ile Asp His Asn Ser
Phe Ser Asn Ser 180 185 190Ser
Asp Gly Leu Val Asp Val Thr Leu Thr Ser Thr Gly Val Thr Ile 195
200 205Ser Asn Asn Leu Phe Phe Asn His His
Lys Val Met Leu Leu Gly His 210 215
220Asp Asp Ala Tyr Ser Asp Asp Lys Ser Met Lys Val Thr Val Ala Phe225
230 235 240Asn Gln Phe Gly
Pro Asn Cys Gly Gln Arg Met Pro Arg Ala Arg Tyr 245
250 255Gly Leu Val His Val Ala Asn Asn Asn Tyr
Asp Pro Trp Thr Ile Tyr 260 265
270Ala Ile Gly Gly Ser Ser Asn Pro Thr Ile Leu Ser Glu Gly Asn Ser
275 280 285Phe Thr Ala Pro Asn Glu Ser
Tyr Lys Lys Gln Val Thr Ile Arg Ile 290 295
300Gly Cys Lys Thr Ser Ser Ser Cys Ser Asn Trp Val Trp Gln Ser
Thr305 310 315 320Gln Asp
Val Phe Tyr Asn Gly Ala Tyr Phe Val Ser Ser Gly Lys Tyr
325 330 335Glu Gly Gly Asn Ile Tyr Thr
Lys Lys Glu Ala Phe Asn Val Glu Asn 340 345
350Gly Asn Ala Thr Pro Gln Leu Thr Lys Asn Ala Gly Val Leu
Thr Cys 355 360 365Ser Leu Ser Lys
Arg Cys 370181313DNACryptomeria japonica 18tagcatagcc gtatagaaag
aaattctaca ctctgctacc aaaaaatgga ttccccttgc 60ttagtagcat tactggtttt
ctcttttgta attggatctt gcttttctga taatcccata 120gacagctgct ggagaggaga
ctcaaactgg gcacaaaaca gaatgaagct cgcagattgt 180gcagtgggct tcggaagctc
caccatggga ggcaagggag gagatcttta tacggtcacg 240aactcagatg acgaccctgt
gaatcctgca ccaggaactc tgcgctatgg agcaacccga 300gataggcccc tgtggataat
tttcagtggg aatatgaata taaagctcaa aatgcctatg 360tacattgctg ggtataagac
ttttgatggc aggggagcac aagtttatat tggcaatggc 420ggtccctgtg tgtttatcaa
gagagttagc aatgttatca tacacggttt gtatctgtac 480ggctgtagta ctagtgtttt
ggggaatgtt ttgataaacg agagttttgg ggtggagcct 540gttcatcctc aggatggcga
tgctcttact ctgcgcactg ctacaaatat ttggattgat 600cataattctt tctccaattc
ttctgatggt ctggtcgatg tcactcttac ttcgactgga 660gttactattt caaacaatct
ttttttcaac catcataaag tgatgttgtt agggcatgat 720gatgcatata gtgatgacaa
atccatgaag gtgacagtgg cgttcaatca atttggacct 780aactgtggac aaagaatgcc
cagggcacga tatggacttg tacatgttgc aaacaataat 840tatgacccat ggactatata
tgcaattggt gggagttcaa atccaaccat tctaagtgaa 900gggaatagtt tcactgcacc
aaatgagagc tacaagaagc aagtaaccat acgtattgga 960tgcaaaacat catcatcttg
ttcaaattgg gtgtggcaat ctacacaaga tgttttttat 1020aatggagctt attttgtatc
atcagggaaa tatgaagggg gtaatatata cacaaagaaa 1080gaagctttca atgttgagaa
tgggaatgca actcctcaat tgacaaaaaa tgctggggtt 1140ttaacatgct ctctctctaa
acgttgttga tgatgcatat attctagcat gttgtactat 1200ctaaattaac atcaacaaga
aatatatcat gatgtatatt gttgtattga tgtcaaaata 1260aaaatgtatc ttttactatt
tatcaacatg ttatctttga tgtgcaagtt aat 131319374PRTCryptomeria
japonica 19Met Asp Ser Pro Cys Leu Val Ala Leu Leu Val Phe Ser Phe Val
Ile1 5 10 15Gly Ser Cys
Phe Ser Asp Asn Pro Ile Asp Ser Cys Trp Arg Gly Asp 20
25 30Ser Asn Trp Ala Gln Asn Arg Met Lys Leu
Ala Asp Cys Ala Val Gly 35 40
45Phe Gly Ser Ser Thr Met Gly Gly Lys Gly Gly Asp Leu Tyr Thr Val 50
55 60Thr Asn Ser Asp Asp Asp Pro Val Asn
Pro Ala Pro Gly Thr Leu Arg65 70 75
80Tyr Gly Ala Thr Arg Asp Arg Pro Leu Trp Ile Ile Phe Ser
Gly Asn 85 90 95Met Asn
Ile Lys Leu Lys Met Pro Met Tyr Ile Ala Gly Tyr Lys Thr 100
105 110Phe Asp Gly Arg Gly Ala Gln Val Tyr
Ile Gly Asn Gly Gly Pro Cys 115 120
125Val Phe Ile Lys Arg Val Ser Asn Val Ile Ile His Gly Leu Tyr Leu
130 135 140Tyr Gly Cys Ser Thr Ser Val
Leu Gly Asn Val Leu Ile Asn Glu Ser145 150
155 160Phe Gly Val Glu Pro Val His Pro Gln Asp Gly Asp
Ala Leu Thr Leu 165 170
175Arg Thr Ala Thr Asn Ile Trp Ile Asp His Asn Ser Phe Ser Asn Ser
180 185 190Ser Asp Gly Leu Val Asp
Val Thr Leu Thr Ser Thr Gly Val Thr Ile 195 200
205Ser Asn Asn Leu Phe Phe Asn His His Lys Val Met Leu Leu
Gly His 210 215 220Asp Asp Ala Tyr Ser
Asp Asp Lys Ser Met Lys Val Thr Val Ala Phe225 230
235 240Asn Gln Phe Gly Pro Asn Cys Gly Gln Arg
Met Pro Arg Ala Arg Tyr 245 250
255Gly Leu Val His Val Ala Asn Asn Asn Tyr Asp Pro Trp Thr Ile Tyr
260 265 270Ala Ile Gly Gly Ser
Ser Asn Pro Thr Ile Leu Ser Glu Gly Asn Ser 275
280 285Phe Thr Ala Pro Asn Glu Ser Tyr Lys Lys Gln Val
Thr Ile Arg Ile 290 295 300Gly Cys Lys
Thr Ser Ser Ser Cys Ser Asn Trp Val Trp Gln Ser Thr305
310 315 320Gln Asp Val Phe Tyr Asn Gly
Ala Tyr Phe Val Ser Ser Gly Lys Tyr 325
330 335Glu Gly Gly Asn Ile Tyr Thr Lys Lys Glu Ala Phe
Asn Val Glu Asn 340 345 350Gly
Asn Ala Thr Pro Gln Leu Thr Lys Asn Ala Gly Val Leu Thr Cys 355
360 365Ser Leu Ser Lys Arg Cys
37020537DNABlomia tropicalis 20aaaacactca caatccacaa actcaaacaa
caatgaagtt cgccatcgtt cttattgcct 60gctttgccgc ttcggttttg gctcaagagc
acaagccaaa gaaggatgat ttccgaaacg 120aattcgatca cttgttgatc gaacaggcaa
accatgctat cgaaaaggga gaacatcaat 180tgctttactt gcaacaccaa ctcgacgaat
tgaatgaaaa caagagcaag gaattgcaag 240agaaaatcat tcgagaactt gatgttgttt
gcgccatgat cgaaggagcc caaggagctt 300tggaacgtga attgaagcga actgatctta
acattttgga acgattcaac tacgaagagg 360ctcaaactct cagcaagatc ttgcttaagg
atttgaagga aaccgaacaa aaagtgaagg 420atattcaaac ccaataaaaa tttagaattg
tacaatttta catttttgat atgattaaat 480gtcaataaat gttcaataaa taaattcaat
ttttaactat aaaaaaaaaa aaaaaaa 53721134PRTBlomia tropicalis 21Met
Lys Phe Ala Ile Val Leu Ile Ala Cys Phe Ala Ala Ser Val Leu1
5 10 15Ala Gln Glu His Lys Pro Lys
Lys Asp Asp Phe Arg Asn Glu Phe Asp 20 25
30His Leu Leu Ile Glu Gln Ala Asn His Ala Ile Glu Lys Gly
Glu His 35 40 45Gln Leu Leu Tyr
Leu Gln His Gln Leu Asp Glu Leu Asn Glu Asn Lys 50 55
60Ser Lys Glu Leu Gln Glu Lys Ile Ile Arg Glu Leu Asp
Val Val Cys65 70 75
80Ala Met Ile Glu Gly Ala Gln Gly Ala Leu Glu Arg Glu Leu Lys Arg
85 90 95Thr Asp Leu Asn Ile Leu
Glu Arg Phe Asn Tyr Glu Glu Ala Gln Thr 100
105 110Leu Ser Lys Ile Leu Leu Lys Asp Leu Lys Glu Thr
Glu Gln Lys Val 115 120 125Lys Asp
Ile Gln Thr Gln 1302215PRTArtificial Sequenceartificial pep66 peptide;
engineered de novo 22Gln Glu Lys Glu Lys Cys Met Lys Phe Cys Lys Lys Val
Cys Lys1 5 10
152315PRTArtificial Sequenceartificial peptide named pep100; engineered
de novo 23Gly Pro Asp Trp Lys Val Ser Lys Glu Cys Lys Asp Pro Asn Asn1
5 10 152445DNAArtificial
Sequenceartificial primer sequence named FAD66 primer; engineered de
novo 24caagagaaag aaaaatgtat gaaattttgc aaaaaagttt gcaaa
452545DNAArtificial Sequenceartificial primer sequence named FAD100
primer; engineered de novo 25ggtcctgatt ggaaagtaag caaagaatgc
aaagatccca ataac 452686DNAArtificial
Sequenceartificial primer sequence named P66F Primer; engineered de
novo 26aagcttgcca tgcaagagaa agaaaaatgt atgaaatttt gcaaaaaagt ttgcaaaggt
60accgccatgg tgagcaaggg cgagga
862730DNAArtificial Sequenceartificial primer sequence named PR Primer;
engineered de novo 27ttaggtacct tacttgtaca gctcgtccat
302886DNAArtificial Sequenceartificial primer
sequence named P100F primer; engineered de novo 28aagcttgcca
tgggtcctga ttggaaagta agcaaagaat gcaaagatcc caataacggt 60accgccatgg
tgagcaaggg cgagga
862925DNAArtificial Sequenceartificial primer sequence named HPRT 5'
Primer; engineered de novo 29gttggataca ggccagactt tgttg
253022DNAArtificial Sequenceartificial primer
sequence named HPRT 3' primer; engineered de novo 30gagggtaggc
tggcctatgg ct
223122DNAArtificial Sequenceartificial primer sequence named IFN-gamma 5'
primer; engineered de novo 31cattgaaagc ctagaaagtc tg
223223DNAArtificial Sequenceartificial
primer sequence named IFN-gamma 3' primer; engineered de novo
32ctcatggaat gcatcctttt tcg
233322DNAArtificial Sequenceartificial primer sequence named IL-4 5'
primer; engineered de novo 33gaaagagacc ttgacacagc tg
223422DNAArtificial Sequenceartificial primer
sequence named IL-4 3' primer; engineered de novo 34gaactcttgc
aggtaatcca gg
223524DNAArtificial Sequenceartificial primer sequence named IL-10 5'
primer; engineered de novo 35ccagtttacc tggtagaagt gatg
243630DNAArtificial Sequenceartificial primer
sequence named IL-10 3' primer; engineered de novo 36tgtctaggtc
ctggagtcca gcagactcaa
303719DNAArtificial Sequenceartificial primer sequence named Fe1 d I.1
P1 5' primer; engineered de novo 37aagcttggat gttagacgc
193820DNAArtificial Sequenceartificial
primer sequence named Fe1 d I.1 P2 3' primer; engineered de novo
38ggtaccttaa cacagaggac
203921DNAArtificial Sequenceartificial primer sequence named Fe1 d I.2
P1 5' primer; engineered de novo 39aagcttggat gaagggggct c
214020DNAArtificial Sequenceartificial
primer sequence named Fe1 d I.2 P2 3' primer; engineered de novo
40ggtaccttaa cacagaggac
204126DNAArtificial Sequenceartificial primer sequence named Can f 1 P1
5' primer; engineered de novo 41aagcttatga agaccctgct cctcac
264226DNAArtificial Sequenceartificial
primer sequence named Can f 1 P2 3' primer; engineered de novo
42ggtaccctac tgtcctcctg gagagc
264323DNAArtificial Sequenceartificial primer sequence named Can f 2 P1
5' primer; engineered de novo 43aagcttatgc agctcctact gct
234422DNAArtificial Sequenceartificial
primer sequence named Can f 2 P2 3' primer; engineered de novo
44ggtaccctag tctctggaac cc
224525DNAArtificial Sequenceartificial primer sequence named Der p 1 P1
5' primer; engineered de novo 45aagcttaaca tgaaaattgt tttgg
254625DNAArtificial Sequenceartificial
primer sequence named Der p 1 P2 3' primer; engineered de novo
46ggtaccgttt agagaatgac aacat
254720DNAArtificial Sequenceartificial primer sequence named Ara h II P1
5' primer; engineered de novo 47aagcttctca tgcagaagat
204820DNAArtificial Sequenceartificial
primer sequence named Ara h II P2 3' primer; engineered de novo
48ggtaccttag tatctgtctc
204920DNAArtificial Sequenceartificial primer sequence named Ara h 5 P1
5' primer; engineered de novo 49aagcttatgt cgtggcaaac
205020DNAArtificial Sequenceartificial
primer sequence named Ara h 5 P2 3' primer; engineered de novo
50ggtacctaaa gacccgtatc
205125DNAArtificial Sequenceartificial primer sequence named Cry j l. 1
P1 5' primer; engineered de novo 51aagcttatgg attccccttg cttat
255223DNAArtificial Sequenceartificial
primer sequence named Cry j l. 1 P2 3' primer; engineered de novo
52ggtaccatca acaacgttta gag
235325DNAArtificial Sequenceartificial primer sequence named Cry j l.2
P1 5' primer; engineered de novo 53aagcttatgg attccccttg cttag
255426DNAArtificial Sequenceartificial
primer sequence named Cry j l.2 P2 3' primer; engineered de novo
54ggtacctcaa caacgtttag agagag
265520DNAArtificial Sequenceartificial primer sequence named Blo t 5 P1
5' primer; engineered de novo 55aagcttacaa tgaagttcgc
205620DNAArtificial Sequenceartificial
primer sequence named Blo t 5 P2 3' primer; engineered de novo
56ggtaccaatt tttattgggt
205721DNAArtificial Sequenceartificial primer sequence named Gata 3 5'
primer; engineered de novo 57ggaggcatcc agacccgaaa c
215819DNAArtificial Sequenceartificial primer
sequence named GATA 3 3' primer; engineered de novo 58accatggcgg
tgaccatgc
195920DNAArtificial Sequenceartificial primer sequence named T-bet 5'
primer; engineered de novo 59tgaagcccac actcctaccc
206020DNAArtificial Sequenceartificial primer
sequence named T-bet 3' primer; engineered de novo 60gcggcatttt
ctcagttggg
2061489DNAArtificial SequenceK-FSA1 insert which includes FSA coding
sequence linked to a Kozak sequence 61gccgccacca tggaagatat ttggaaagtt
aataaaaaat gtacatcagg tggaaaaaat 60caagatagaa aactcgatca aataattcaa
aaaggccaac aagttaaaat ccaaaatatt 120tgcaaattaa tacgggataa accacataca
aatcaagaga aagaaaaatg tatgaaattt 180agcaaaaaag tttgcaaagg ttatagagga
gcttgtgatg gcaatatttg ctactgcagc 240aggccaagta atttaggtcc tgattggaaa
gtaagcaaag aatgcaaaga tcccaataac 300aaagattctc ggcctacgga aatagttcca
tatcggcagc aattagcaat tccaaatatt 360tgcaaactaa aaaattcagg gaccaatgaa
gattccaaat gcaaaaaaca ttgcaaagaa 420aaatgtcgtg gtggaaatga tgctggatgt
gatggaaact tttgttattg tcggccaaaa 480aataaataa
489
User Contributions:
Comment about this patent or add new information about this topic:
| People who visited this patent also read: | |
| Patent application number | Title |
|---|---|
| 20200011000 | A LIQUID DISPENSING DEVICE, PARTICULARLY FOR DISPENSING WASHING AGENTS IN A WASHING MACHINE |
| 20200010999 | METHOD FOR INSPECTING UNBALANCE ERROR OF WASHING MACHINE AND WASHING MACHINE |
| 20200010998 | DIRECT DRIVING APPARATUS FOR WASHING MACHINE |
| 20200010997 | LAUNDRY TREATING APPARATUS |
| 20200010996 | TRACK MECHANISM AND FRONT-LOADING WASHING MACHINE WITH SAME |
 |  |
 |  |
 |  |
 | 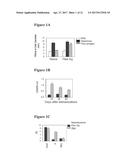 |
 |  |
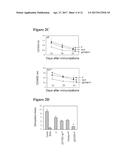 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |
| New patent applications in this class: | |
| Date | Title |
|---|---|
| 2022-09-22 | Electronic device |
| 2022-09-22 | Front-facing proximity detection using capacitive sensor |
| 2022-09-22 | Touch-control panel and touch-control display apparatus |
| 2022-09-22 | Sensing circuit with signal compensation |
| 2022-09-22 | Reduced-size interfaces for managing alerts |