Patent application title: MUTATED NUCLEOTIDE MOLECULE, AND TRANSFORMED PLANT CELLS AND PLANTS COMPRISING THE SAME
Inventors:
Swee-Suak Ko (Taipei, TW)
Min-Jeng Li (Taipei, TW)
Assignees:
Academia Sinica
IPC8 Class: AC12N1582FI
USPC Class:
800278
Class name: Multicellular living organisms and unmodified parts thereof and related processes method of introducing a polynucleotide molecule into or rearrangement of genetic material within a plant or plant part
Publication date: 2015-11-05
Patent application number: 20150315607
Abstract:
The present invention relates to a method for producing male sterile
plant, a mutated nucleotide molecule comprising a nucleotide sequence of
the transcription factor bHLH142 and an inserted T-DNA segment, and a
novel transformed plant cell and a male-sterile mutant plant comprising
the mutated nucleotide molecule, in which the transcription factor
bHLH142 is not expressed. The present invention also relates to a novel
reversible male sterile transgenic plant, wherein the transcription
factor bHLH142 is overexpressed, and its preparation method. The bHLH
gene is tissue specifically expresses in the anther and it plays a
pivotal role in pollen development. Both the male sterile and reversible
male sterile transgenic plants showed a completely male sterile
phenotype, but the fertility of the reversible male sterile transgenic
plant can be restored under low temperature.Claims:
1. A mutated nucleotide molecule, comprising a nucleotide sequence of the
transcription factor bHLH142 and an inserted T-DNA segment.
2. The mutated nucleotide molecule according to claim 1, wherein the nucleotide sequence of the transcription factor bHLH142 has a DNA sequence of SEQ ID No: 1 or a DNA sequence having at least 60% similarity to SEQ ID No: 1.
3. The mutated nucleotide molecule according to claim 1, wherein the T-DNA segment is inserted in the third intron of the nucleotide sequence of the transcription factor bHLH142.
4. The mutated nucleotide molecule according to claim 2, wherein the T-DNA segment is inserted at +1257 bp of the nucleotide sequence of the transcription factor bHLH142.
5. A transformed plant cell, which comprises the mutated nucleotide molecule according to claim 1.
6. The transformed plant cell according to claim 5, wherein the transcription factor bHLH142 has a DNA sequence of SEQ ID No: 1 or a DNA sequence having at least 60% similarity to SEQ ID No: 1.
7. The transformed plant cell according to claim 5, wherein the T-DNA segment is inserted in the third intron of the nucleotide sequence of the transcription factor bHLH142.
8. The transformed plant cell according to claim 6, wherein the T-DNA segment is inserted at +1257 bp of the nucleotide sequence of the transcription factor bHLH142.
9. A male sterile mutant plant, which comprises the mutated nucleotide molecule according to claim 1, and the transcription factor bHLH142 is not expressed.
10. The male sterile mutant plant according to claim 9, wherein the transcription factor bHLH142 has a DNA sequence of SEQ ID No: 1 or a DNA sequence having at least 60% similarity to SEQ ID No: 1.
11. The male sterile mutant plant according to claim 9, wherein the T-DNA segment is inserted in the third intron of the nucleotide sequence of the transcription factor bHLH142.
12. The male sterile mutant plant according to claim 10, wherein the T-DNA segment is inserted at +1257 bp of the nucleotide sequence of the transcription factor bHLH142.
13. The male sterile mutant plant according to claim 9, which is a homozygous mutant.
14. The male sterile mutant plant according to claim 9, wherein the plant is a monocot plant.
15. The male sterile mutant plant according to claim 14, wherein the monocot plant is rice, maize, wheat, millet, sorghum or Brachypodium distachyon.
16. The male sterile mutant plant according to claim 9, wherein the plant is a dicot plant.
17. The male sterile mutant plant according to claim 16, wherein the dicot plant is Arabidopsis or Brassica species.
18. A transformed plant cell, which comprises a plasmid comprising the sequence of the transcription factor bHLH142 and a strong promoter.
19. The transformed plant cell according to claim 18, wherein the transcription factor bHLH142 has a DNA sequence of SEQ ID No: 1 or a DNA sequence having at least 60% similarity to SEQ ID No: 1.
20. The transformed plant cell according to claim 18, wherein the strong promoter is Ubiquitin promoter, CaMV 35S promoter, Actin promoter, an anther tapetum-specific promoter or a pollen-specific promoter.
21. A reversible male sterile mutant plant, wherein the transcription factor bHLH142 is overexpressed.
22. The reversible male sterile transgenic plant according to claim 21, wherein the transcription factor bHLH142 has a DNA sequence of SEQ ID No: 1 or a DNA sequence having at least 60% similarity to SEQ ID No: 1.
23. The reversible male sterile transgenic plant according to claim 21, wherein the expression of the transcription factor bHLH142 is controlled by a strong promoter.
24. The reversible male sterile transgenic plant according to claim 21, wherein the fertility of the plant is recovered under low temperature.
25. The reversible male sterile transgenic plant according to claim 21, wherein the plant is a monocot plant.
26. The reversible male sterile transgenic plant according to claim 25, wherein the monocot plant is rice, maize, wheat, millet, sorghum or Brachypodium distachyon.
27. The reversible male sterile transgenic plant according to claim 21, wherein the plant is a dicot plant.
28. The reversible male sterile transgenic plant according to claim 27, wherein the dicot plant is Arabidopsis or Brassica species.
29. A method for preparing the reversible male sterile transgenic plant according to claim 21, comprising: (a) constructing a plasmid comprising the sequence of the transcription factor bHLH142 and a strong promoter; and (b) introducing the plasmid into a target plant.
30. The method according to claim 29, wherein the DNA sequence of bHLH142 is SEQ ID No: 1 or a DNA sequence having at least 60% similarity to SEQ ID No: 1.
31. The method according to claim 29, wherein the strong promoter is Ubiquitin promoter, CaMV 35S promoter, Actin promoter, an anther tapetum-specific promoter or a pollen-specific promoter.
32. The method according to claim 29, wherein the plasmid is introduced into calli of the target plant via Agrobacterium tumefaciens.
Description:
BACKGROUND OF THE INVENTION
[0001] Rice (Oryza sativa) is one of the most important staple crops in the world, feeding almost half of the world's population, and it serves as a model for monocots, which include many important agronomic crops (e.g. wheat, maize, sorghum, millet). Food and Agriculture Organization (FAO) predicts that rice yield will have to be increased 50-70% by 2050 to meet the demands. Several approaches are currently adopted to increase rice yields, such as heterosis breeding, population improvement, wide hybridization, genetic engineering, and molecular breeding1. Among these, hybrid rice is being considered the most promising one (15-20% increases in yield)2. Crops produced from F1 hybrid seeds offer significant benefits in terms of yield improvement, agronomic performance and consistency of end-use quality. This is due to the `hybrid vigor` generated by combining carefully selected parent lines. Hybrid crops are responsible for a dramatic increase in global crop yields in the past decades, and male sterility (MS) has played a significant role in this advancement. Male sterile traits can be divided into cytoplasmic male sterility (CMS), which is determined by cytoplasmic factors such as mitochondria, and genetic male sterility (GMS), which is determined by nuclear genes. CMS has long been used in hybrid corn production, while both CMS and GMS are currently used for hybrid rice production3, due to the convenience of controlling sterility expression by manipulating the gene-cytoplasm combinations in any selected genotype. Most importantly, it evades the need for emasculation in cross-pollinated species, thus encouraging cross breeding and producing pure hybrid seeds under natural conditions. However, commercial seed production must be simple and inexpensive, and the requirement for a maintainer line to produce the seed stocks of CMS line increases the production cost for this 3-line hybrid system.
[0002] On the other hand, genetic MS (GMS), controlled by nuclear genes, offers an alternative hybrid seed production system. For the two-line hybrid system, it is beneficial to use photoperiod- or temperature-inducible MS (PGMS or TGMS) mutants to maintain seed stocks for hybrid seed production. Currently, in China, PA64S is the most widely used maternal line in two-line hybrid rice breeding, and it is crossed with paternal line 93-11 to generate superhybrid rice, LYP94. PA64S, derived from a spontaneous PGMS japonica mutant NK58S (long day->13.5 h; Shi, 1985), is also a TGMS indica rice, whose MS is promoted by temperatures greater than 23.5° C., but recovers its fertility at temperatures between 21˜23° C. Recent mapping analyses demonstrate that the P/TGMS in these MS lines is regulated by a novel small RNA5. In the case of another rice genic MS mutant discovered recently, Carbon Starved Anther (CSA), the mutation on the R2R3 MYB transcription regulator defects pollen development6 and further study shows that csa is a new photoperiod-sensitive mutant, exhibiting MS under short-day conditions but male fertility under long-day conditions7. The molecular basis of its MS sensitivity to day length remains to be addressed.
[0003] Transgenic male sterility has been generated using a number of transgenes, but its application in commercial production of hybrid seeds is limited due to the lack of an efficient and economical means to maintain the MS lines, or the lack of suitable restorers8. Recently, a reversible MS system has been demonstrated in transgenic Arabidopsis plants by manipulating a R2R3 MYB domain protein (AtMYB103)8. Blocking the function of AtMYB103 using an insertion mutant or an AtMYB103EAR chimeric repressor construct under the control of the AtMYB103 promoter resulted in complete MS without seed setting8. A restorer containing the AtMYB103 gene driven by of a stronger anther-specific promoter was introduced into pollen donor plants and crossed into the MS transgenic plants for the repressor. The male fertility of F1 plants is restored. The chimeric repressor and the restorer constitute a reversible MS system for hybrid seed production. The successful application of this system for large scale hybrid seed production depends on whether the MS female parent lines can be multiplied efficiently and economically. Alternatively, an inducible promoter by chemicals or other factors (e.g. photoperiod or temperature) can be directly used to regulate the expression of a GMS gene (e.g., bHLH142) and control pollen development in transgenic plants, eliminating the costly need to maintain MS lines.
[0004] Rice anthers are composed of four lobes attached to a central core by connective and vascular tissue. When anther morphogenesis is completed, microsporocytes form in the middle, surrounded by four anther wall layers: an epidermal outer layer, endothecium, middle layer, and tapetum9. The tapetum is located in the innermost cell layer of the anther walls and plays an important role in supplying nutrients such as lipids, polysaccharides, proteins, and other nutrients for pollen development10. The tapetum undergoes programmed cell death (PCD) during the late stage of pollen development11; this PCD causes tapetal degeneration and is characterized by cellular condensation, mitochondria and cytoskeleton degeneration, nuclear condensation, and internucleosomal cleavage of chromosomal DNA. Tapetal PCD must occur at a specific stage of anther development for normal tapetum function and pollen development, and premature or delayed tapetal PCD and cellular degeneration can cause male sterility3,12-14.
[0005] Genetic and functional genomic studies of MS in Arabidopsis have shown that many transcription factors (TFs) play an essential role in pollen development and the regulation of tapetal PCD, such as mutations in DYSFUNCTIONAL TAPETUM 1 (DYT1), Defective in Tapetal Development and Function 1 (TDF1, AtMYB35), ABORTED MICROSPORES (AMS, homolog of TDR1 in rice), and MALE STERILITY 1 (MS1); and mutations in these factors all result in MS phenotype. The genetic regulatory pathway of pollen development suggests that DYT1, TDF115 and AMS16 function at early tapetum development, while MS18817 and MS115,18,19 play important roles in late tapetum development and pollen wall formation. Whilst, in rice, several TFs, such as Undeveloped Tapetum1 (UDT1, homolog of DYT1), are known to be key regulators of early tapetum development20. In addition, mutations in TAPETUM DEGENERATION RETARDATION (TDR1)14, GAMYB21,22, ETERNAL TAPETUM 1 (EAT1)23 and DELAYED TAPETUM DEGENERATION (DTD)24 all cause MS associated with tapetal PCD. TDR1, ortholog of the Arabidopsis AMS gene, plays an essential role in tapetal PCD in rice; and tdr1 shows delayed tapetal degeneration and nuclear DNA fragmentation as well as abortion of microspores after release from the tetrad. Molecular evidences indicate that TDR1 directly binds the promoter of CP1 and C6 for their transcription14. C6 encodes a lipid transfer protein that plays a crucial role in the development of lipidic orbicules and pollen exine during anther development17. CP1 is involved in intercellular protein degradation in biological system and its mutant shows defected pollen development25. EAT1 acts downstream of TDR1 and directly regulates the expression of AP25 and AP37, which encode aspartic proteases involved in tapetal PCD23.
[0006] The basic helix-loop-helix (bHLH) proteins are a superfamily of TFs and one of the largest TF families in plants. There are at least 177 bHLH genes in the rice genome26,27 and more than 167 bHLH genes in Arabidopsis genome28,29. Generally, eukaryotic TFs consist of at least two discrete domains, a DNA binding domain and an activation or repression domain that operate together to modulate the rate of transcriptional initiation from the promoter of target genes30. The bHLH TFs play many different roles in plant cell and tissue development as well as plant metabolism3. The HLH domain promotes protein-protein interaction, allowing the formation of homodimeric or heterodimeric complexes31. They bind as dimers to specific DNA target sites and are important regulatory components in diverse biological processes29. So far, three of the bHLH TFs have been shown to be involved in rice pollen development--UDT1 (bHLH164), TDR1 (bHLH5), and EAT1/DTD1 (bHLH141).
[0007] From a screening of T-DNA tagged rice mutant pool of TNG6732, we isolated a novel MS-related gene encoding for another member of the bHLH TFs (bHLH142). In this invention, the molecular mechanism of MS in this mutant is elucidated, and it suggests that bHLH142 is specifically expressed in the anther and bHLH142 coordinates with TDR1 in regulating EAT1 promoter activity in transcription of protease genes required for PCD during pollen development. That is to say, bHLH142 plays an essential role in rice pollen development by controlling tapetal PCD. Both null mutant and overexpression transgenic plants showed a completely male sterile phenotype. Most interestingly, the overexpression plants have restored the fertility under low temperature. Homologs of SEQ ID NO: 2 with high similarity are found in other major cereal crops, and its use may increase the productivity of cereal crops by manipulating the bHLH gene for development of male sterility and production of hybrid crops.
SUMMARY OF THE INVENTION
[0008] The object of the present invention is developing a mutated nucleotide molecule, and a transformed plant cell and a male sterile mutant plant comprising the mutated nucleotide molecule; in which the male sterile mutant plant can be used as a female parent to produce F1 hybrid seeds, thereby improving yield and quality of crops.
[0009] The present invention provides a mutated nucleotide molecule, comprising a nucleotide sequence of the transcription factor bHLH142 and an inserted T-DNA segment. Preferably, the T-DNA segment is inserted in the third intron of the nucleotide sequence of the transcription factor bHLH142; more preferably, the T-DNA segment is inserted at +1257 bp.
[0010] In one preferred embodiment of the mutated nucleotide molecule, the T-DNA segment has comprises a single copy of T-DNA.
[0011] In one preferred embodiment of the mutated nucleotide molecule, the nucleotide sequence of the transcription factor bHLH142 has a DNA sequence of SEQ ID No: 1 or a DNA sequence having at least 60% similarity to SEQ ID No: 1. Preferably, a DNA sequence having at least 80% similarity to SEQ ID No: 1; more preferably, a DNA sequence having at least 90% similarity to SEQ ID No: 1; even more preferably, a DNA sequence having at least 95% similarity to SEQ ID No: 1; and most preferably, a DNA sequence of SEQ ID No: 1. In addition, the transcription factor bHLH142 has a polypeptide sequence of SEQ ID No: 2 or a polypeptide sequence having at least 60% similarity to SEQ ID No: 2. Preferably, a polypeptide sequence having at least 80% similarity to SEQ ID No: 2; more preferably, a polypeptide sequence having at least 90% similarity to SEQ ID No: 2; even more preferably, a polypeptide sequence having at least 95% similarity to SEQ ID No: 2; and most preferably, a polypeptide sequence of SEQ ID No: 2.
[0012] The present invention provides a transformed plant cell, which comprises the above-mentioned mutated nucleotide molecule. Preferably, the T-DNA segment is inserted in the third intron of the nucleotide sequence of the transcription factor bHLH142; more preferably, the T-DNA segment is inserted at +1257 bp.
[0013] In one preferred embodiment of the transformed plant cell comprising the mutated nucleotide molecule, the nucleotide sequence of the transcription factor bHLH142 has a DNA sequence of SEQ ID No: 1 or a DNA sequence having at least 60% similarity to SEQ ID No: 1. Preferably, a DNA sequence having at least 80% similarity to SEQ ID No: 1; more preferably, a DNA sequence having at least 90% similarity to SEQ ID No: 1; even more preferably, a DNA sequence having at least 95% similarity to SEQ ID No: 1; and most preferably, a DNA sequence of SEQ ID No: 1. In addition, the transcription factor bHLH142 has a polypeptide sequence of SEQ ID No: 2 or a polypeptide sequence having at least 60% similarity to SEQ ID No: 2. Preferably, a polypeptide sequence having at least 80% similarity to SEQ ID No: 2; more preferably, a polypeptide sequence having at least 90% similarity to SEQ ID No: 2; even more preferably, a polypeptide sequence having at least 95% similarity to SEQ ID No: 2; and most preferably, a polypeptide sequence of SEQ ID No: 2.
[0014] The present invention also provides a male sterile mutant plant comprising the above-mentioned mutated nucleotide molecule, and the transcription factor bHLH142 is not expressed; particularly, not expressed in anthers. Preferably, the T-DNA segment is inserted in the third intron of the nucleotide sequence of the transcription factor bHLH142; more preferably, the T-DNA segment is inserted at +1257 bp.
[0015] In one preferred embodiment of the male sterile mutant plant, the nucleotide sequence of the transcription factor bHLH142 has a DNA sequence of SEQ ID No: 1 or a DNA sequence having at least 60% similarity to SEQ ID No: 1. Preferably, a DNA sequence having at least 80% similarity to SEQ ID No: 1; more preferably, a DNA sequence having at least 90% similarity to SEQ ID No: 1; even more preferably, a DNA sequence having at least 95% similarity to SEQ ID No: 1; and most preferably, a DNA sequence of SEQ ID No: 1. In addition, the transcription factor bHLH142 has a polypeptide sequence of SEQ ID No: 2 or a polypeptide sequence having at least 60% similarity to SEQ ID No: 2. Preferably, a polypeptide sequence having at least 80% similarity to SEQ ID No: 2; more preferably, a polypeptide sequence having at least 90% similarity to SEQ ID No: 2; even more preferably, a polypeptide sequence having at least 95% similarity to SEQ ID No: 2; and most preferably, a polypeptide sequence of SEQ ID No: 2.
[0016] In one preferred embodiment of the male sterile mutant plant, the male sterile mutant plant of the present invention is a homozygous mutant.
[0017] In one preferred embodiment of the male sterile mutant plant, the plant is a monocot; preferably, the monocot is rice, maize, wheat, millet, sorghum or Brachypodium distachyon.
[0018] In one preferred embodiment of the male sterile mutant plant, the plant is a dicot; preferably, the dicot is Arabidopsis or Brassica species.
[0019] The present invention also provides a transformed plant cell, which comprises a plasmid comprising the sequence of the transcription factor bHLH142 and a strong promoter.
[0020] In one preferred embodiment of the transformed plant cell comprising the sequence of the transcription factor bHLH142, the transcription factor bHLH142 has a DNA sequence of SEQ ID No: 1 or a DNA sequence having at least 60% similarity to SEQ ID No: 1. Preferably, a DNA sequence having at least 80% similarity to SEQ ID No: 1; more preferably, a DNA sequence having at least 90% similarity to SEQ ID No: 1; even more preferably, a DNA sequence having at least 95% similarity to SEQ ID No: 1; and most preferably, a DNA sequence of SEQ ID No: 1.
[0021] In one preferred embodiment of the transformed plant cell comprising the sequence of the transcription factor bHLH142, the strong promoter is Ubiquitin promoter, CaMV 35S promoter, Actin promoter, an anther tapetum-specific promoter or a pollen-specific promoter; preferably, the anther tapetum-specific promoter is Osg6B or TA29, and the pollen-specific promoter is LAT52 or LAT59.
[0022] The present invention also provides a reversible male sterile transgenic plant, wherein the transcription factor bHLH142 is overexpressed; particularly, overexpressed in anthers.
[0023] In one preferred embodiment of the reversible male sterile transgenic plant, the nucleotide sequence of the transcription factor bHLH142 has a DNA sequence of SEQ ID No: 1 or a DNA sequence having at least 60% similarity to SEQ ID No: 1. Preferably, a DNA sequence having at least 80% similarity to SEQ ID No: 1; more preferably, a DNA sequence having at least 90% similarity to SEQ ID No: 1; even more preferably, a DNA sequence having at least 95% similarity to SEQ ID No: 1; and most preferably, a DNA sequence of SEQ ID No: 1. In addition, the transcription factor bHLH142 has a polypeptide sequence of SEQ ID No: 2 or a polypeptide sequence having at least 60% similarity to SEQ ID No: 2. Preferably, a polypeptide sequence having at least 80% similarity to SEQ ID No: 2; more preferably, a polypeptide sequence having at least 90% similarity to SEQ ID No: 2; even more preferably, a polypeptide sequence having at least 95% similarity to SEQ ID No: 2; and most preferably, a polypeptide sequence of SEQ ID No: 2.
[0024] In one preferred embodiment of the reversible male sterile transgenic plant, the expression of the transcription factor bHLH142 is controlled by a strong promoter; preferably, by an Ubiquitin promoter, CaMV 35S promoter, Actin promoter, an anther tapetum-specific promoter or a pollen-specific promoter; preferably, the anther tapetum-specific promoter is Osg6B or TA29, and the pollen-specific promoter is LAT52 or LAT59.
[0025] In one preferred embodiment of the reversible male sterile transgenic plant, the pollen fertility of the plant is recovered under low temperature. Particularly, the pollen fertility of the plant is recovered at 21-23° C.
[0026] In one preferred embodiment of the reversible male sterile transgenic plant, the plant is a monocot; preferably, the monocot is rice, maize, wheat, millet, sorghum or Brachypodium distachyon.
[0027] In one preferred embodiment of the reversible male sterile transgenic plant, the plant is a dicot; preferably, the dicot is Arabidopsis or Brassica species.
[0028] The present invention also provides a method for preparing the above-mentioned reversible male sterile transgenic plant, comprising:
[0029] (a) constructing a plasmid comprising the DNA sequence of bHLH142 and a strong promoter, and
[0030] (b) introducing the plasmid into a target plant.
[0031] In one preferred embodiment of the preparation method, the DNA sequence of bHLH142 is SEQ ID No: 1 or a DNA sequence having at least 60% similarity to SEQ ID No: 1. Preferably, a DNA sequence having at least 80% similarity to SEQ ID No: 1; more preferably, a DNA sequence having at least 90% similarity to SEQ ID No: 1; even more preferably, a DNA sequence having at least 95% similarity to SEQ ID No: 1; and most preferably, a DNA sequence of SEQ ID No: 1.
[0032] In one preferred embodiment of the preparation method, the strong promoter is Ubiquitin promoter, CaMV 35S promoter, Actin promoter, an anther tapetum-specific promoter or a pollen-specific promoter; preferably, the anther tapetum-specific promoter is Osg6B or TA29, and the pollen-specific promoter is LAT52 or LAT59.
[0033] In one preferred embodiment of the preparation method, the plasmid is introduced into calli of the target plant via Agrobacterium tumefaciens.
BRIEF DESCRIPTION OF THE DRAWINGS
[0034] FIG. 1. Phenotypic Analyses of ms142 Mutant. (A) Comparison of the wild-type plant (left) and ms142 mutant (right) after bolting; (B) comparison of the wild-type panicle (left) and ms142 mutant (right) at heading stage; (C) comparison of the wild-type panicle (left) and ms142 mutant (right) at seed maturation stage; (D) phenotype of the wild-type spikelet (left) and ms142 mutant (right) one day before anthesis; (E) phenotype of the wild-type spikelet (left) and ms142 mutant (right) after anthesis; (F) comparison of the wild-type grain (left) and ms142 mutant (right) at harvest stage; (G) phenotype of the wild-type grain (left) and ms142 mutant (right) after removing rice husk; (H) phenotype of the wild-type anther (left) and ms142 mutant (right) one day before anthesis; (I) staining of anther by Sudan black in the wild-type (left) and ms142 mutant (right); (J) staining of wild-type pollen grains by I2/KI solution; and (K) staining of mutant pollen grains by I2/KI solution. Bars=20 cm in (A), 3 cm in (B) and (C), 0.5 cm in (D) to (G), 0.1 cm in (H) and (I), and 20 μm in (J) and (K).
[0035] FIG. 2. Evidences of T-DNA insertion in ms142 mutant. (A) Southern blotting to hptII probe confirmed single T-DNA insertion (marked with arrow) in ms142 mutant. (B) T-DNA tagged construction map of pTag4 vector. (C) Gene structure and T-DNA insertion site in the mutant at 3rd intron (+1257 bp from ATG). (D) Genotyping heterozygous T4 mutant progeny. Genotype: WT, wild-type like; He, heterozygous; and Ho, homozygous. Grain fertility: F, fertile; and S, sterile.
[0036] FIG. 3, Transverse anatomical comparison of the anther development in the wild type (TNG67) and ms142 mutant. E, epidermis; En, endothecium; ML, middle layer; T, tapetum; Ms, microsporocyte; Td, tetrads; Msp, microspore; MP, mature pollen; Arrow, degenerated microspore; and arrowhead, two tapetum layers. Bars=20 μm.
[0037] FIG. 4. TUNEL assay showed defect tapetal program cell death in ms142 mutant. DNA fragmentation signals (yellow fluorescence) started at tetrad stage and exhibited obvious positive signal at young microspore stage in wild type (TNG67). No DNA fragmentation signal was observed in ms142 anther. Red signal shows the propidium iodide (PI) staining, and yellow fluorescence is the merged signal from TUNEL (green) and PI. Scale bar=50 μm.
[0038] FIG. 5. Scheme of bHLH142 gene, multiple alignment, and subcellular localization of bHLH142 fused with GFP. (A) Scheme of the bHLH142 gene and T-DNA insertion position. Gray boxes represent exons and interventing lines represent introns. The ATG start codon and TGA stop codon are indicated. bHLH, basic helix-loop-helix domain (amino acids 182 to 228); NLS, two nuclear localization signals (amino acids 159 to 165 and 235 to 240, respectively). (B) Alignment of bHLH domains. The bHLH142 protein was aligned with the bHLH domains of the analogous proteins from other species. Asterisks indicate the conserved basic amino acid Arg that is important for binding DNA. Pound signs indicate conserved Leu residues important for forming the α-helix. (C) Schemes of fusion constructs. P35S, cauliflower mosaic virus 35S promoter; Trios, nopaline synthase gene terminator. The NLS domain of VirD2 fused with mRFP was used as the nuclear marker. (D) In vivo targeting of fusion protein. DIC (bright field, left column) and overlays (Merge, right column) are shown. Scale bar=20 μm.
[0039] FIG. 6. Phylogenetic analysis of bHLH142 related proteins. Aral, Arabidopsis lyrata; Aegt, Aegilops tauschii; At, Arabidopsis thaliana; Brad, Brachypodium distachyon; Os, Oryza sativa; Sei, Setaria italica; Sb, Sorghum bicolor; SeMa, Selaginella moellendorffii; Triu, Triticum urartu; and Zm, Zea may.
[0040] FIG. 7. Spatial and temporal gene expression of bHLH142 in various tissues of TNG67 (WT) by (A) RT-PCR and (B) qRT-PCR, and (C) in TNG67 spikelet at various developmental stages; and (D) ISH of bHLH142 antisense (left panel) and sense (right panel) probes in spikelet of TNG67 at meiosis stage. Error bars indicate SD (n=3), SC, sporogenous cell; MMC, meiocyte mother cell; Mei, meiosis; YM, young microspore; VP, vacuolated pollen; PM, pollen mitosis; and MP, mature pollen. Scale bar=1 mm in (C), 50 μm in (D).
[0041] FIG. 8. In situ hybridization analysis of bHLH142 expression in the anther of WT and ms142 mutant at various developmental stages. Transverse sections of anther were hybridized with antisense or sense dig-labeled probe of bHLH142. Scale bar=20 μm.
[0042] FIG. 9. Analysis of alternation in expression of key regulatory genes involved in pollen development in ms142 by qRT-PCR. Error bars indicate SD (n=3). MMC, meiocyte mother cell; Mei, meiosis; and VP, vacuolated pollen.
[0043] FIG. 10. Gene hierarchy of bHLH142, as determined by the expression profiles of the key regulatory genes involved in pollen development in ms142 and eat1 mutants. (A) Phenotype of spikelet in TNG67 (WT) and ms142. (B) Phenotype of spikelet in Hitomebore and H0530 (eat1 mutant). (C) Real-time RT-PCR of bHLH142 in ms142 and eat1 mutants and their respective wild-types. (D) Real-time RT-PCR of EAT1 (bHLH141) in ms142 and eat1 mutants and their respective wild types. qRT-PCR value presented are means±SE (n=3). MMC, meiocyte mother cell; Mei, meiosis; YM, young microspore; and VP, vacuolated pollen.
[0044] FIG. 11. Coordinated Regulation of EAT1 Promoter by bHLH142 and TDR1. (A) Schematic diagrams of the reporter, effector, and internal control plasmids used in the transient transactivation assay in rice leaf protoplasts. The reporter plasmid contains the CaMV35S minimal promoter and the EAT1 promoter sequence (2 Kb) fused to the firefly luciferase gene (Luc). In the effector plasmids, bHLH142, TDR1, and EAT1 genes were driven under the control of the CaMV35S promoter. Nos and t35s denote the terminators of nopaline synthase and CaMV35S, respectively. The pBI221 vector contains a CaMV35S promoter driving the expression of GUS as the internal control. (B) Transactivation of the Luc reporter gene by bHLH142 and TDR in rice protoplasts. Different effectors were co-transfected with the reporter and internal control plasmid (pBI221). The data represent means of three independent transient transformations. Error bars indicate SD. Transient transformation without the effector plastnid (mini35p) was used as a negative control.
[0045] FIG. 12. Analysis of protein interaction between bHLH142, TDR1, and EAT1 by (A) Yeast two-hybrid assay and (B) BiFC in rice leaf protoplasts expressing the indicated constructs. Scale bars=20 μm.
[0046] FIG. 13. The Interactions between bHLH142 Protein and TDR1, and between TDR1 and EAT1. (A) Yeast two-hybrid (Y2H) assays. Constructs expressing the full length bHLH142 were cloned into the prey vector pGADT7 (AD), and truncated forms of TDR and EAT1 were prepared in the bait vector pGBKT7 (BD). (B) BiFC in rice protoplasts expressing the indicated constructs. Bars represent 10 μm. (C) Co-IP assay of HA fused TDR and bHLH142 recombinant proteins expressed in E. coli using anti-HA antibody. (D) Co-IP assay of HA fused TDR and bHLH142 recombinant proteins expressed in E. coli using bHLH142 antibody.
[0047] FIG. 14. RNAi Knockdown (KD) of bHLH142 Inhibited Pollen Development. (A) Construct of RNAi vector. (B) RT-PCR showed down regulation of bHLH142 in the anthers of four RNAi knock down lines. (C) Phenotype of WT and KD line #3. Scale bars=20 cm in (C), 0.5 mm (D), 1 mm (E) and 20 μm (F).
[0048] FIG. 15. Overexpression rice bHLH142 driven by Ubiquitin promoter in transgenic rice. (A) Construction of bHLH142 (LOC_Os01g18870) driven by Ubiquitin promoter in pCAMBIA1301 vector, (B) Genomic PCR confirmed T-DNA insertion of target gene (upper panel) and selection marker hygromycin (lower panel) in the transgenic rice.
[0049] FIG. 16. Phenotype of TNG67 (WT) and Ubi::bHLH142 transgenic lines. (A) Plant type of WT (left) and transgenic line (right), (13) panicles of WT (bottom) and transgenic line (top panel), (C) panicles of WT (left) and different transgenic lines, (D) spikelet of WT (left) and different transgenic lines at one day before anthesis, (E) mature seed of WT (left) and different transgenic lines, and (F) removed husk rice seed of WT (left) and different transgenic lines. Scale bars=20 cm in (A), 3 cm in (B), 7 cm in (C) and 1 cm in (F).
[0050] FIG. 17. Overexpression bHLH142 prematurely up-regulated Udt1 and EAT1 before meiosis stage but significantly down-regulated MS2 that associated in pollen exine development. SC=sporogenous cell, MMC=meiocyte mother cell, Mei=meiosis, YM=young microspore, PM=pollen mitosis, MP=mature pollen.
[0051] FIG. 18. Gene expression pattern of bHLH142 homolog in various organs of maize by using RT-PCR. Floret size by measuring the length of floret. 10G, floret length at 10 mm with green color, 1 DBA, one day before anthesis.
[0052] FIG. 19. Heterologus overexpressing Ubi::bHLH142 caused male sterility in transgenic maize, (A) The transgenic line has smaller angle of tassel branch (right panel) than the WT (left panel). (B) Closed up tassel during anthesis stage, WT has large and anther opened during anthesis stage (left panel), while anther of transgenic maize were significantly smaller in size and anther no dehiscence (right panel). (C) Morphology of spikelet of WT (left panel) and transgenic line (right panel), (D) Stained of pollen grains by I2/KI solution in the fertile WT, but not stainable in the transgenic line and pollen was not viable showing transparent pollen. Scale bars=3 cm in (A), 2 mm in (C) and 50 um in (D).
[0053] FIG. 20. The male sterility of Overexpression Ubi::bHLH142 transgenic line is sensitive to environment. The bHLH142 overexpressed lines produced no pollen grains during summer season (Left panel) but had fertile pollens during winter seasons. Pollen grains were stained by I2/KI solution.
[0054] FIG. 21. A proposed model for the molecular function of bHLH142 in rice anther development, relative to other key regulators. bHLH142 is in the downstream of UDT1 but upstream of TDR1, and bHLH142 interacts with TDR1 protein and coordinately regulate the promoter of EAT1. In turn, EAT1 regulates AP37 and CP1 and promotes tapetal PCD. Evidences from previous works were indicated by black arrows, while data demonstrated in the present study were indicated by red arrows.
DETAILED DESCRIPTION OF THE INVENTION
[0055] The following examples are presented to demonstrate the present invention. These examples are in no way to be construed as a limitation on the invention. The disclosure would enable those skilled in the art to practice the present invention without engaging in undue experimentation. All recited publications are incorporated herein by reference in their entirety.
Plant Materials and Growth Conditions
[0056] The seed of ms142 mutant was obtained from TRIM library. Seedlings of ms142 mutant and its WT (TNG67) were raised in half strength Kimura solution for 3 weeks and then transplanted into soil in AS-BCST GMO screen house located in Tainan, Taiwan.
Anther Anatomy
[0057] Spikelets and anthers of the WT and ms142 mutant were sampled at various stages of development and fixed overnight in phosphate buffer, pH 7.0, that contained 4% paraformaldehyde and 2.5% glutaraldehyde. They were then rinsed with the same buffer and post fixed for 30 min in phosphate buffer, pH 7.0, containing 1% osmium tetroxide. After dehydration, the specimens were embedded in Spurr's Resin (EMS). The processor, KOS Rapid Microwave Labstation, was chosen for post fixation, dehydration, resin infiltration, and embedding. For TEM, ultrathin sections (90 to 100 nm thick) collected on coated copper grids were stained with 6% uranyl acetated and 0.4% lead citrate and examine using transmission electron microscope.
Total RNA Isolation and PCR.
[0058] Total RNA was isolated from rice tissues using MaestroZol® RNA PLUS (Invitrogen) as described by the supplier. Various rice organs at different developmental stages were harvested for RNA isolation: root, shoot, flag leaf, internode, panicles of 0.5 cm, 1 cm, 5 cm, 9 cm, and 20 cm length, spikelet at 1 day before anthesis (1 DBA), lemma, palea, anthers, ovary, seed at 5 days after pollination (S1), 15 days after pollination (S3), 25 days after pollination (S5), and callus. The stages of anthers were classified into the following categories according to spikelet length: microspore mother cell (MMC) with spikelet length of approximately 2 mm, meiosis (4 mm), young microspore (YM, 6 mm), vacuolated pollen (VP, 8 mm), mitosis pollen (MP, 8 mm with light green lemma), and mature pollen at one day before anthesis (1 DBA). Total RNA was treated with DNase (Promega), and 1 μg RNA was used to synthesize the oligo(dT) primed first-strand cDNA using the M-MLV reverse transcriptase cDNA synthesis kit (Promega). One μL of the reverse transcription products was used as template in PCR reactions. Ubiquitin-like 5 and 18srRNA were used as normalizer control. Each sample has three biological repeats.
qRT-PCR Analysis.
[0059] Fifteen μL of RT-PCR reaction contained 4 μL, of 1/4 diluted cDNA, 3 μM of primers, and 7.5 μL of 2×KAPA SYBR FAST master mix (KAPA Biosystems, USA). Quantitative Real-Time PCR (qRT-PCR) was performed using a CFX96 Real-Time PCR detection system (Bio-Rad, USA). Quantification analysis was carried out using CFX Manager Software (Bio-Rad, USA). Primers used for qPCR are listed in Table 1.
TABLE-US-00001 TABLE 1 Primers Used in Examples RAP Acces- Primer's Names sion No. Sequence (5'→') Sequence ID Screening of ms142 Mutant T-DNA inserted S80qPCR-F3 Os01g0293100 GGAGCACGTACATCCAGCGG SEQ ID No. 3 S80-GT-R3 ACTCATCCACCACTTCAATCAGCC SEQ ID No. 4 RB-13 AACTCATGGCGATCTCTTACC SEQ ID No. 5 Hyg-F GATGTAGGAGGGCGTGGATA SEQ ID No. 6 Hyg-R CGTCT GCTGC TCCAT ACAAG SEQ ID No. 7 Quantitative Real-time PCR S80qPCR-F3 Os01g0293100 GGAGCACGTACATCCAGCGG SEQ ID No. 8 S80-GT-R3 ACTCATCCACCACTTCAATCAGCC SEQ ID No. 9 OsMS2-qRT-F Os03g0167600 TGGAGCAGTTCGCCAGCTACG SEQ ID No. 10 OsMS2-qRT-R CTTCTCCTCCTCCGACATCTCCC SEQ ID No. 11 OsTDR-F Os02g0120500 CGCTCGCTCGTCCCAAACAT SEQ ID No. 12 OsTDR-R CGGTCATTGCTGGGTCCTTGT SEQ ID No. 13 Os C6-F Os11g0582500 TCCTCCTCGTCCTGCTCGTC SEQ ID No. 14 Os C6-R GGTTCACGATGTGGCACAGG SEQ ID No. 15 18SrRNA-F2 TTAGG CCACG GAAGT TTGAG G SEQ ID No. 16 18SrRNA-R2 ACACT TCACC GGACC ATTCA A SEQ ID No. 17 Udt1-F Os07g0549600 GATCTTCTGGACCAAGAGGGCAG SEQ ID No. 18 Udt1-R GTCAGGAGTGTCTCAGATGCTTGG SEQ ID No. 19 OsCP1-F Os09g0381400 GGACCACCTGCTGCTGCAACT SEQ ID No. 20 OsCP1-R GAACACTTCGTGCCATCGCC SEQ ID No. 21 bHLH141-F Os04g0599300 TGGTGGAACAGAAGAGGCATGG SEQ ID No. 22 bHLH141-R GCATGAAGCAGAGAGTTGGCCTT SEQ ID No. 23 OsUBQ5-F Os01g0328400 GCGGAAGTAAGGAAGGAGGAGG SEQ ID No. 24 OsUBQ5-R GGCATCACAATCTTCACAGAGGTG SEQ ID No. 25 MSP1-F Os01g0917500 GAGAACTTCGAGCCGAGGGTCT SEQ ID No. 26 MSP1-R CCAGCCGACGAGGTTTCCAC SEQ ID No. 27 AP37-F Os04g0448500 AGGCGGGCAGCGTCTCCAT SEQ ID No. 28 AP37-R CCATAAGCCAGCCACGATGATGA SEQ ID No. 29 GAMyb-F Os01g0812000 CATCCTGGTCCATTCCTCAATGAC SEQ ID No. 30 GAMyb-R TTCAGGATGAGGTGAAGTGTCCC SEQ ID No. 31 Subcellular localization analysis S80cDNA-XbaI-F TATCTAGAGTGGTAGAGTGCGAGGAAG SEQ ID No. 32 KpnI-S80-nonstop-R GAGGTACCAGGTACTCATCCACCACTTCAA SEQ ID No. 33 In situ hybridization analysis S80qPCR-F2 CATGTTCAACACCAAGATTCATTCG SEQ ID No. 34 S80FLcds-R2 TGCAAACCATGACATACCAAAGATC SEQ ID No. 35 BiFC construction BiFC-TDR-BamHI-F AGGGATCCCACCACATGGGAAGAGGAGACC SEQ ID No. 36 BiFC-TDR-SalI-R ATGTCGACTCAAACGCGAGGTAATGCAGG SEQ ID No. 37 BiFC-UDT-BamHI-F GTGGATCCATGCCGCGGCGCGCGAGGGCGA SEQ ID No. 38 BiFC-UDT-Xho-R TGCTCGAGATGCTTGGAACCTCCACAATGCTGG SEQ ID No. 39 BiFC-bHLH141-Pst-F AGCTGCAGTTTGCCAAAATGATTGTTGGG SEQ ID No. 40 BiFC-bHLH141-Sal-R GCGTCGACTTGAATATGTCGAGGGCCTGG SEQ ID No. 41 S80-Y2H-AD-BamH-F GTGGATCCCGAGGAAGATGTATCACC SEQ ID No. 42 S80-Y2H-AD-Xho-R AGCTCGAGCTAGTTAGTACTCATCCACCAC SEQ ID No. 43 Yeast Two Hybrid S80-Y2H-AD-BamH-F GTGGATCCCGAGGAAGATGTATCACC SEQ ID No. 44 S80-Y2H-AD-Xho-R AGCTCGAGCTAGTTAGTACTCATCCACCAC SEQ ID No. 45 S80-Y2H-BD-Pst-R CGCTGCAGTTAGTACTCATCCACCACTT SEQ ID No. 46 141-Y2H-BD-Eco-F AGGAATTCTTTGCCAAAATGATTGTTGGG SEQ ID No. 47 141-Y2H-BD-Pst-R GCCTGCAGTTAGTTGAATATGTCGAGGGCCTG SEQ ID No. 48 TDR-Y2H-EcoR-F AAGAATTCATGGGAAGAGGAGACCACCTGC SEQ ID No. 49 TDR-Y2H-BamH-R CTGGATCCTCAATCAAACGCGAGGTAATGCA SEQ ID No. 50 Y2H-Deletion141-N-BD TAGAATTCATGAAGGGTGAGTTCGGAAAGGGC SEQ ID No. 51 Y2H-Deletion141-C-BD AGGTCGACTTACCCTCTCCTGCATTCAAGTACA SEQ ID No. 52 Y2H-DeletionTDR-N-BD ACGAATTCATGCATGTCCACCATAAGCCGC SEQ ID No. 53 Co-Immunoprecipitation CO-IP-HA-SacII-F AACCGCGGAAATGAGTTACCCATACGATGTTCCTG SEQ ID No. 54 CO-IP-HA-Not-R CAGCGGCCGCAGCGTAATCTGGAACGTCATAT SEQ ID No. 55 CO-IP-TDR-HA-Xba-F AATCTAGACATGGGAAGAGGAGACCACCTGC SEQ ID No. 56 TDR-cDNA-SalI-R CAGTCGACTCAATCAAACGCGAGGTAATGCA SEQ ID No. 57 CO-IP-S80-Flag-Xba-F CGTCTAGAGATGTATCACCCGCAGTG SEQ ID No. 58 S80-Y2H-AD-Xho-R AGCTCGAGCTAGTTAGTACTCATCCACCAC SEQ ID No. 59 Promoter transience assay BiFC-TDR-BamHI-F AGGGATCCCACCACATGGGAAGAGGAGACC SEQ ID No. 60 TDR-cDNA-SaII-R CAGTCGACTCAATCAAACGCGAGGTAATGCA SEQ ID No. 61 BiFC-bHLH141-Pst-F AGCTGCAGTTTGCCAAAATGATTGTTGGG SEQ ID No. 62 bHLH141-cDNA-Sal-R GCGTCGACTTAGTTGAATATGTCGAGGGCCT SEQ ID No. 63 141-2kb-P-Spe-F CCACTAGTTGCTTTGGTTTTGATTCCTGGAAG SEQ ID No. 64 141-2kb-P-Sal-R AAGTCGACAACAGTGCTAGGCACCTTCGC SEQ ID No. 65 S80-Y2H-AD-BamH-F GTGGATCCCGAGGAAGATGTATCACC SEQ ID No. 66 S80-Y2H-AD-Xho-R AGCTCGAGCTAGTTAGTACTCATCCACCAC SEQ ID No. 67 RNAi transgenic line PANDA-GUS-F-Spe acactagtATCTACCCGCTTCGCGTCGG SEQ ID No. 68 PANDA-GUS-R-Pst atctgcagCGAGTGAAGATCCCTTTCTTGTTACC SEQ ID No. 69 142RNAi-5'-F-Pst gcctgcagCAACAAACCTAGTTAATT SEQ ID No. 70 TAGCTCTAGTTGG 142RNAi-5'-R-Sal gcgtcgacAGGCTCTCAAGCGGCATCAG SEQ ID No. 71 142RNAi-5'-F-Spe gcactagtCAACAAACCTAGTTAATT SEQ ID No. 72 TAGCTCTAGTTGG 142RNAi-5'-R-Not tagcggccgcAGGCTCTCAAGCGGCATCAG SEQ ID No. 73 Overexpression bHLH142 in transgenic rice Hyg-F GATGTAGGAGGGCGTGGATA SEQ ID No. 74 Hyg-R CGTCT GCTGC TCCAT ACAAG SEQ ID No. 75 S80qPCR-GT-F GGAGCACGTACATCCAGCGG SEQ ID No. 76 Pzp200-NOS-R ATCGCAAGACCGGCAACAGGA SEQ ID No. 77 Overexpression bHLH142 in transgenic maize Zm142-F TGACCTCGTCCACCTCTCCG SEQ ID No 117 Zm142-R CAGTCTGTAACGAGCAAGCGGA SEQ ID No. 118
In-Situ Hybridization
[0060] Spikelets of TNG67 and ms142 at various developmental stages were fixed in PFA [4% paraformaldehyde, 4% dimethylsulfoxide 0.25% glutaraldehyde, 0.1% Tween 20, 0.1% Triton X-100 in diethyl pyrocarbonate (DEPC)-treated H2O] at 4° C. overnight immediately after collection, and the tissue processor, KOS Rapid Microwave Lab station, was used for dehydration and wax infiltration. After embedding, sections of 10 μm thickness were prepared by a rotary microtome (MICROM, 315R) and mounted on APS adhesive microscope slides (FINE FROST). Tissue sections were deparaffinized with xylene, rehydrated through an ethanol series, and pre-treated with proteinase K (2 mg/mL) in 1-phosphate buffered saline (PBS) at 37° C. for 30 min. Pre-hybridization (additionally including 25% RNAmate, BioChain) and hybridization were performed according to the previous protocols39. Hybridization was performed at 59° C. in hybridization solution: 50% formamide, 4×SSPE, 1×Denhardt's (Fluka), 250 μg/mL fish sperm DNA (Genemarker), 250 μg/mL yeast tRNA (Sigma), 10% dextran sulfate, 40 U/mL RNasin (Promega) and 40 ng of DIG-labeled RNA probe/per slide. RNA probes were synthesized by in vitro transcription of the RT-PCR fragment in pGEM-T easy vector using the DIG RNA labeling kit (SP6/T7, Roche). Antisense RNA probes were synthesized by SP6 RNA polymerase, while sense RNA probes were synthesized by T7 RNA polymerase and used as control. Sequence of fragment to synthesize RNA probe (SEQ ID No. 119):
TABLE-US-00002 5'-catgttcaacaccaagattcattcgggatctccagtgtttgcaa gtgcagtggccagcaggctgattgaagtggtggatgagtactaacta gatcgagctagctaattagccgaccgaccgatcgatatgatgaaagt ttctatgttgctagctagctagggttcttggatgcatgagtactgag tagctctttaattaatttccttttaattttagactgtttaatttgga ttggtaaagactcgtgttagcttttgggagatctttggtatgtcatg gtttgca-3'
Gene Hierarchy Analysis Using Knockout Mutants
[0061] We have some T-DNA/Tos17 knock out mutant lines in hands such as: in udt1 (TRIM), bHLH142 (ms142, TRIM), and eat1 (bHLH141) Tos17 mutant line H0530 (background of Hitomebore) was obtained from Rice Tos17 Insertion Mutant Database (http://tos.nias.affrc.go.jp/). Flanking sequences were confirmed by genotyping PCR amplification with specific primers (Table 1). We will verify their gene hierarchy using these mutants. Spikelet samples at various developmental stages were collected, isolated RNA, and performed qRT-PCR analysis.
TUNEL Assay
[0062] PCD is characterized by cellular condensation, mitochondria and cytoskeleton degeneration, nuclear condensation, and internucleosomal cleavage of chromosomal DNA33. To investigate the nature of the tapetal breakdown in ms142, the terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL) assay was performed using DeadEnd Fluorometric TUNEL system (Promega). This assay detects in situ DNA cleavage, a hallmark feature of apoptosis-like PCD, by enzymatically incorporating fluorescein-12-dUTP into the 3'-OH ends of fragmented DNA. Stage of anther development was based primarily on spikelet size and developmental stages.
Subcellular Localization of bHLH142
[0063] For subcellular localization of bHLH142, the coding sequences of the gene were subcloned into p2FGW7 (Invitrogen) to generate bHLH142-GFP fusion genes driven by the CaMV 35S promoter. Rice protoplasts were isolated and transformed using the polyethylene glycol (PEG) method following procedures described previously34. After incubation at room temperature for 16 h in light, protoplasts were observed with a Zeiss LSM 780 laser scanning confocal microscope.
Phylogenetic Analysis of the bHLH142 Subfamily
[0064] The bHLH142 protein sequence was used to search for the closest homologues from their plant species using BLASTP programs. Multiple sequence alignment of full-length protein sequences was performed using ClustalW online (http://www.ch.embnet.org/software/ClustalW.html), and the alignment was used to perform neighbor-joining analysis using Mega 5.0535. The numbers at the nodes represent percentage bootstrap values based on 1000 replications. The length of the branches is proportional to the expected numbers of amino acid substitutions per site. Gene identification numbers used to generate the phylogenetic trees and the alignment are listed in Table 2.
TABLE-US-00003 TABLE 2 Peptide Sequences Used for Phylogenetic Analysis Seq- uence Peptide ID. Peptide sequence (N' → C') ID BradXP_003567568 myhpqcellmphesldmdavvgqshlaasgvsaipaelnfhllhhsfvdtaaspqpptvdyffpgtdpppaa SEQ ID vqfeqlaatnhhamsmlrdyygqqypaetylrggprtttagssslvfgvahddesaaynmvgpfvesspttr No. 78 aagggrkrnrgsraaggpahggvekkekqrrlrltekytalmllipnrtkedratvisdaieyiqelgrtve eltllvgkkrrrngagehhlhqgdvvdaapavgaagelvlaaessegevqaplaalqpirstyiqrksket fvdvrivedevnikltkrrrdgclaaasralddlrldlvhlsggkigdchiymfntkihqgspvfasavas klievvdey TriuEMS50437 myhqqcellmphegldmdagqshhlaagsavpaelnfhllsyvdtavspqqptveyffggadqphaqfeqla SEQ ID anhqamtvlrdyygqyhpatadaylpgggprtgssslvfgaaeeesaymvggfqcspkprasgsrkrgrgag No. 79 ssfhgfpanggvekkekqrrqrlsekftalmllipnrtkedratviydaieyiqelgrtveeltllvekkrg rrehqgdvvdpaptlvagdgecsagevaaavmpampappqpirstyiqrrsketfvdvrivedevnikltkr rrdgclaaasralddlhldlvhlsggkigdchiymfntkihpgspvfasavasklievvdey SbXP_002457706 myhpqcellmaheaqdldaagqphhlavsgvagsipaelsfhllhsldataavnnsvtpqstidyflgvggad SEQ ID phqpaalqyeplpppgghhqhtmnmlrdycsnggggghyptaepylrgtrtgalvfgatdddesaaaympggp No. 80 fvetspppratggrkrgralgggfhaglangvekkekqrrqrltekytalmhlipnvtkpdratvisdaieyi qelgrtveeltllvekkrrrrelqgdvvdaaptavvvaaaatggeaessegevappppppaavqrqpirstyi qrrskdtsvdvriveedvnikltkrrrdgclaaasralddlrldlvhlsggkigdchiymfntkihkgssvfa savasrlmevvdey ZmLOC100283549 Myhpqcelltmahetpdldagqphltvsgvasipaelsfhllhsldaaaavnpvtappqstidyfiggadphq- q SEQ ID amqyeplppaagghhqytmdmfrdycdghyptaepyirgtmtgalvfgatddddsaaaympgghfetsppppr- atg No. 81 rgrkrgralgggfhavlangvekkekqrrirltekytalmhlipnvtktdratvisdaieyiqelgrtveelt- i lveklelqgdvvdaapaavvaaageaessegevappppavprqpirstyiqrrskdtsvdvri veedvnikltkrrrdgclaaasraldcllrldlvhlsggkigdcqiymfntkihkgssvfasavagrlmevvd- ey AegtEMT16792 myhqqcellmphedldmdagqshhlaaasavpaelnfhllsyvdaavspqqptveyffggadqphahsfhgfp- ang SEQ ID gvekkekqrrqrlsekftalmllipnrtkedratviydaieyiqelgrtveeltllvekkrgrrehqgdvvdp No. 82 aplvvagegecsagelrappppppqpirstyiqrrsketfvdvrivedevnikltkrrrdgclaaasra lddlhldlvhlsggkigdchiymfntkihpgspvfasavasklievvdey ZmEU974003 mvfskiqykvvsskiqtptlvrvetthetnmekkrrrrelqgdvvdaapaavvaaageaesse- gevapppp SEQ ID avprqpirstyiqrrskdtsvdvriveedvnikltkrrrdgclaaasralddlrldlvhlsggkigdcqiy No. 83 mfntkihkgssvfasavagrlmevvdey VitvCBI38213 mcrkitrprrmyvyeenacfdgtksvaegddegfsqsvappptnnsfedstnmrvsmedasatmeielhqq SEQ ID lafdmdqqcynsnndgndsnqvfsyemqemgfnhhqqqqedplllqqhqaemqnahqnfsaaypptpdllnlf- hlpr No. 84 ctpssllpnssisftnpdssataasgilydplfhlnlppqppvgggygdgddhrqfdngvlkftrdmacigkg- regk gtksfatekqrrehlndkynalrslvpnptksdrasvvgdaieyirellrtvnelkllvekkrcgrerskrhk- tede stgdvkssssikpepdqsyneslrsswlqrkskdtevdvriiddevtiklvqrkkincllfvskildelq ldlhhvagghvgdyysflfntkiyegssvyasaianklievvdrqyaaipipipipptssf VitvCAN77001 Mymyeenacfdgtksvaegddegfsqsvappptnnsfedstnmrvsmedasatmeielhqqlafdmdqqcyns- nn SEQ ID dgndsnqvfsyemqemgfnhhhqqqddplllqqhqaemqnaqqnfsaaypptpdllnlfhlprctpssllpns- s No. 85 isftnpdssataasgilydplfhlnlppqppvfrelfqslphgynlpasrvgslfgggmdereasgggygdgd- dhrq fdngvlkftrdmaciggregkgtksfatekqrrehlndkynalrslvpnptksdrasvvgdaieyirellrtv- nelkl lvekkrcgrerskrhktedestgdvkssssikpepdqsyneslrsswlqrkskdtevdvriiddevtiklvqr- kkin cllfvskildelqldlhhvagghvgdyysflfntkiyegssvyasaianklievvdrqyaaipipipipptss- f VitvXP_002265098.2 mgrgfshllgfcksenplndlsrlksrysgpnlrlvhsesnrnnienmvdafpvqidlnfsdsrndkatqt SEQ ID ishprvkhshqgarrrimrstnhaeqcqnkrgyrkitrprrmyvyeenacfdgtksvaegddegfsqsvappp- tnns No. 86 fedstnmrvsmedasatmeielhqqlafdmdqqcynsnndgndsnqvfsyemqemgfnhhqqqqedplllqqh- qae mqnahqnfsaaypptpdllnlfhlprctpssllpnssisftnpdssataasgilydplfhlnlppqppvfrel- fqslp hgynlpasrvgslfgggmdereasgggygdgddhrqfdngvlkftrdmacigkgregkgtksfatekqrrehl- ndkyn alrslvpnptksdrasvvgdaieyirellrtvnelkllvekkrcgrerskrhktedestgdvkssssikpepd- qs yneslrsswlqrkskdtevdvriiddevtiklvqrkkincllfvskildelqldlhhvagg hvgdyysflfntkiyegssvyasaianklievvdrqyaaipipipipptssf RiccXP_002534354.1 myeetgcfdpnsmvegaddglcqvlqippqpqplmagsttnshnsyeenlklsadqelsyhhsnnphhhhqed- das SEQ ID asaaaametqlqnhqmgfdthlmqdssnqmafnsstslqdatfaqtpdllnlfhlprgstssllpnssisft No. 87 npshtaplgfvgdlpmadtasassilydplfhlnlppqpplfrdlfqsIpphgyslpgsmvnslfgagvggdd- h vegsgdgggiyqdgdgeqqfdngvldftwdmpcmgkgrdagkktkpfaterqrrqhlndkykalqnlvpnp tkadrtsvvgdaidyikellrtvnelkllvekkrcarerskrqkteedsignghdsscitkplgdpdqsfnng slrsswierkskdtevdvriiddevtiklvqrkkincllfvskvldeIqIdlhhvagghi gdyysflfntkifegssvyasaianklievvdrhyastpstn PoptXP_002323376 myvetacfepnnsmvedvtddgfchaiplmagnsttnsfeehlklsmeefsshypqeesaaaasmeeiqlqhh- mafs SEQ ID nnntnhhlmqqyptqllsydhssnwdpniiqfqemhqvldqnssfdatantqsslppdllnlfnlprctstst- llpn No. 88 ssisftnpahkaplgfmgvdntsarfdpytlapqphlfrelvqslpphgytlptplfgggqgddhvdgqsggg- lsyq dgdhgdgvfeftdemacigkgikktgkvtkhfaterqrrehlngkytalrnlvpnpskndrasvvgeaidyik- ellr tvqelkllvekkrcgrerskwrkteddggvevldnsdikvepdqsaysngslrsswlqrkskdtevdvrlied- evti klvqrkrvncllyvskvldelqldlhhaaggligdyysflfntkinegscvyasaianrlievvdrqyasstt- tvpa agscy Zm_HLH_ miagggyfdgshdhilmegsmihdssqssiydntdveqqnfrfapfiiedhsnpanltseaarvid- qiqhqIgidie SEQ ID containing_ qdhsdhmmqevppaetenlvpavygvqdhilshqiegphnitveqqvlgydpasyrngtyaaahdllnslhiq- rcsl No. 89 protein ipefpstehifsdpaqnmvnrlditndlpgvanhesgmmfsdstvplgyhatqshmlkdlyhslpq- nyglftsdde rdgmvgvpgvsgnifgeidgrqfdspilgsrkqkggfgkgkgkanfatererrxqfnvkygalrslfpnptkn- dra sivgdaieyinelnrtvkelkillekkrnsadrrkilkldeeaaddgesssmqpvsddqxnqmngtirsswvq- rrs kecdvdvrivddeinikftekkransllcaakvleefhlelihvvggiigdhhifmfntkipkgssvyacava- kkl leaveikkqaynifn PoptXP_002308327 myeetacfetnnsiveggnddgfcqvspfmtgssttssfeesfklsmeelsnhyhqeesaaaasmeeiqlq SEQ ID hhmafnnnchhlmeqyptnhhqvlsydhpsnwdpntiqfqemhqvldqngnfnatantpssllpdllnlfn No. 90 lprctststllpnssisftnpahktpsgfmgvdstsvlfdsnplapqfrelvhslpphgyglpaplfgggq ggdhvdglsggglsyqdgghgdgvfeftaemacigkgirksgkvitkhfaterqrrehlngkytalnlvpn pskndrasvvgdainyikellrtveelkllvekkrngrerikrrkpeedggvdvlensntkveqdqstynn gslrsswlqrkskhtevdvrliedevtiklvqrkkvncllsvskvldelqIdlhhaaggligdyysflfnt kinegscvyasgiankllevvdrqyasstsvpaasc SbXP_002452697 miagggyfdgshdhilmegsmihdssqssiydntdveqqnfrlapfiiedhsnpanltseparvidgihh SEQ ID qlgidmeqdhsdhmiqgvppaetanlvpvvygvqdrilshqiegphnitveqqvldydpasygngtyaaa No. 91 hdllnslqiqrcslipefpstehifgdpaqnmvnplditndlqgvathesgmmfsdstlplgyhatqshm lkdlyhslpqnygiftsdderdgmvgvagvsgnifqeidgrqfdspvlgtrrqkggfgkgkgkanfater erreqlnvkygalrslfpnptkndrasivgdaidyinelnrtvkelkillekkrnstdrrkilklddeaa ddgesssmqpvsddqnnqmngairsswvqrrskecdvdvrivddeinikftekkransllca akvleefrlelihvvggiigdhhifmfntkipkgssvyacavakklleaveikkgalnifn MedtXP_003638306 mssgsgdkqnmheqngcfdpntkdegvenspndnntnnnnsleenfkpsveelpyhnhqnsqhlddvstytng- ftp SEQ ID ssvdieqlqnlglnigntynnmdnhlvqevyqnstwdpsvqdmdyvnhqehrqlseqqyqqfieaqnhnqsyn- pst No. 92 ildphypspdvlnllnlprcssslltnssticmtnptqnppnfhnsmtflgdlpigssdntsgssvlydplyp- lnlp pqppalrelfqslprgysmptnsrngslfgggdemegdgdmgvlefnrvtasvgkgrggkatkhfatekqrre- qlng kykilrdlipsptktdrasvvgdaieyirelirtvnelkllvekkrhgremckrlkteddaaescnikpfgdp- dgsi rtswlqrkskdsevdvriidddvtiklfqrkkvncllfvskvldelqlelhhvagghvgeycsflfnskvneg- ssvy asaianrvidvmdtqyaaglphisrl MedtXP_003638303 mssgsgdnqnmheqtgcfdpdtmaegvenspednnspqtmpnqvvagnsnnsieenfrpsveefsyhnhhspq- hl SEQ ID edvstytngftpsseniaqqnlglnignyyynnmdnlleqevyqnsswdpsaqdmdyanhqeyhqlhnhkqsy- nps No. 93 ttqaphypspdvlnlhfprssasslltnpsticitnptqkppnfhysmsflgdlpigsdnssgssvlydplfp- lnl paqspalrelpqslprvysmptnsrngspfgggdemegdggmgvsqfnkvtafvgkgkgkatehlttekqrre- glk grykilrslipnstkddrasvvgdaieylrelirtvnelkllvekkrheieickrhktedyaaeschmkpfgd- pdg sirtswlqrkskdsevdvriidddvtiklfqrkkvncllfvskvldelqlelnhvagghvgeycsflfnskvi- egs svhasaianrvidvldtqyaavvphnrm CucsXP_004173553 meldfqqaaaaptpgfdgeltsdsnpmlcldqsnwvgtqiqemgfnhnhvqsqfsdsaipptpytqppdllnf- lnmp SEQ ID ptarcsnnssisfsnlhtpamgaflgdlppgdapnssstslsilydplfhlnlppqpplfrelfhsIphgygm- paa No. 94 ssrgrggslfpegseiveregtagvyedgdgsgvlefsrdmadcigkrrdgkmtkhfaterqrrvqlndkyka- lrslv piptkndrasivgdainyiqellrevkelkllvekkrssrerskrvrtaeeieqgggsessnakggegvvedq- rynlr sswlqrktkdtevdvrivddevtvklvqrklnclllvsklledlqldlhhvagghigdyysflfntkiyegss vyasaiankvmeavdrqynntsispltnty BradXP_003580474 Miiggdyfegshdhslmagslihdssqapkcngntdielqkfkvpsfsseiltnstnlsseaarainhlqhql- g SEQ ID idleqdmqpvetatwdasicsiqdhiinnqisedpqnilveqqiqqydaaiypnssytpapdllnllhctvap- a No. 95 fptttsvfgdtslsstnyldlngeftgvaatpesglmftsdsalqlgyhatqshplkdichslpqnyglfpge- de revmigvgsvggdifqdiddrqfdtvlecrrgkgefgkgkgkanfatererreqlnvkyktlkdlfpnptksd- ra svvgdaieyidelnrtvkelkilveqkwhgnkrtkiikldeevaadgesssmkpmrddqdnqfdgtirsswvq- r rskechidvrivenevnikltekkkvnsllhaarvldefglelihavggiigdhhifmfntkv segssvyacavakrllqavdaqhqainifh CucsXP_004139000 myeetecsdpnsispetmphisafpnsfpppliaqqthpnfhhnnnlnlsidhisyhhhstalqpadameld SEQ ID fqqaaaaptpgfdqeltsdsnpmlcldqsnwvgtqiqemgfnhnhvqsqfsdsaipptpytqppdllnflnm No. 96 pptarcsnnssisfsnlhtpamgaflgdlppgdapnssstslsilydplfhlnlppqpplfrelfhslphgyg mpaassrgrggslfpegseiveregtagvyedgdgsgvlefsrdmadcigkrrdgkmtkhfaterqrrvqlnd- ky kalrslvpiptkndrasivgdainyiqellrevkelkllvekkrssrerskrvrtaeeieqgggsessnakgg- egv vedqrynlrsswlqrktkdtevdvrivddevtvklvqrklnclllvsklledlqldlhhvagghigdyysflf- ntki yegssvyasaiankvmeavdrqynntsispltnty AegtEMT05766 miaeggyfdgsrdailmagslihdsldsicdnteieqgnfhgpsffiedicnptnltsesartinhiqhraef- dmdq SEQ ID
dlhghmiqetqvetsnwvpamfgtqnhiisqqsieqqmddydaasypdgahtaapdllnllqiprysmttafp- stehi No. 97 fgdpgqnagnqldinndvlgraihdsgmmlgdstlplqyndnqshlfkdlyhslpqsfglfssdderdramgv- vgaa gnilqeidgrqfgspklgrskkggfgkakanfatekerreqinvkygalrsllpsptkndrasivgdaieyin- elnr tlkeltslvegdtkhrmkrlkldddaacdngessslqqvkddqdsqlngairsswiqrrskechvdvrivgne- in ikftekkktnsllcaakvidefrlelihvvggvigdqrifmfntkisegssvyasalaskllramemehlavd- ifs FravXP_004308623 myvdsstaaagacnfdpntdtnpmsesapevvlhhqnnpttfasthdenlrslsmeeelsnyhhnhnaameie- q SEQ ID qlqtemgfgtmdqntnntnphlinpfdthqatnwdnndemqqqqqlppvaptpdllslfhlpnssvlphssit- ft No. 98 npktpggcfpgsfgyetlpetpsgavasnsvmydpmfhlnqlppqpqpplfrellqslphgykrngsslfsng- gde vddgsrqlfengvlefskemkpfgrgrggnkgtkhfaterqrrvqlndkfsalrelvpnptkpdrasvvgdai- dyi qelkrtvselkllvekkrcgrerskrhkteqdigardddescnmkplgdpdhdhsynngslrsswlqrksk dtevdvriiddevtiklvqrkkinlllsvsklldelqlelhhaagghignsysflfntkmyegsslyas aianklidtvdrqyaaaipptnsy AegtEMT07628 miwiegarhcfvkmivggdyfegshdhnlmtgslthdsslapkcndntnielgrfkvqsfsadilsdstnlss- eaar SEQ ID ainhlqhqlgigleqdmppvetatwdtsictiqdqiinhqlsedpqnilvqqqiqqydaalypnsgytpapdl- lnllh No. 99 ctvapvfpatasvfgdtalsggtnyldlngeftgvaaipdsglmytsdpalqlgyhaapshalkdichslpqn- yglfp sederdvmlgvgsvggdlfqdmddrqfetvlegrrgkgefgkgkgkanfatererreqlnvkyktlrmlfpnp- tkndr asvvgdaieyidelnrtvkelkilveqkwhgtnrrrirkldeeaaadgesssmrpmrdeqdnqldgairsswv- qrrsr echvdvriveneinikltekkkanssllhvakvldefhleiihvvggiigdhyifmfntkvtegssvyacava- kri lqavdaqhqaldifn AralXP_002879311 Myeesscfdpnpmvdnngsfcaaettfpvshqfqppvgsttnsfnddlklptmeefsafpsvislpnsetqn SEQ ID qnisnnnhlinqmiqepnwgvsedntgffmntshpnttttpipdllsllhlprcsmalpssnlsdimagscf No. 100 tydplchlnlppqpplipsndysgyllgidtntttqgdesnvgdennnaqfdsgiiefskeirrkgrgkrkn kpfttererrchlneryealkllipnpskgdrasilqdgidyinelrrrvselkylverkreggrhknneld nninnnnsndhdndeddiddenmekkpesdvvdqcssnnslrcswlqrkskvtevdvrivddevtikvvqkk kinclllvskvldqlqldlyhvaggqigehysflfntkiyegstiyasaianrvievvdkhymaalpiny AralXP_002892324 megggmfeeigcfdpnapaemtaessfspaeppptitvigsnsnsncsledlsefhlspqdsslpasasay SEQ ID vhqlhvnatpncdhqfqssmhqtlqgpsypqqsnnwdngyqdfvnlvpnhttpdllsllqlprsslppfan No. 101 pslqdiimttsssvaaydplfhlnfplqppngtfigvdqdqteienqgvnlmydeennnldnglnrkgrgs rkrkvfptererrvhfkdrfgdlknlipnptkndrasivgeaidyikellrtidefkllvekkrtkqrnre gddvidenfkaqsevveqclinkkrtnahtswlkrkskftevdvriidddvtikivqkkkinclvfvskvv dqlqldlhhvagaqigehhsflfnakicegssvyasaiadrvmevlekqymealstnngyhcyssd AtbNP_180679 myeesscfdpnsmvdnnggfcaaettvshqfqpplgsttnsfdddlklptmdefsvfpsvislpnsetqnqni- sn SEQ ID nnhlinqmiqesnwgvsednsnffmntshpnttttpipdllsllhlprcsmslpssdimagscftydplfhln- lp No. 102 pqpplipsndysgyllgidtntttqrdesnvgdennnaqfdsgiiefskeirrkgrgkrknkpfttererrch- lner yealkllipspskpskgdrasilqdgidyinelrrrvselkylverkrcggrhknnevddnnnnknlddhgne- ddd dddenmekkpesdvidqcssnnslrcswlqrkskvtevdvrivddevtikvvqkkkinclllvskvldqlqld- lhh vaggqigehysflfntkiyegstiyasaianrvievvdkhymaslpnsny AtbCAD58593 myeesscfdpnsmvdnnggfcaaettvshqfqpplgsttnsfdddlklptmdefsvfpsvislpnsetqnqni- snn SEQ ID nhlinqmiqesnwgvsednsnffmntshpnttttpipdllsllhlprcsmslpssdimagscftydplfhlnl- ppq No. 103 pplipsndysgyllgidtntttqrdesnvgdennnaqfdsgiiefskeirrkgrgkrknkpfttererrchln- ery ealkllipspskpskgdrasilqdgidyinelrrrvselkylverkrcggrhknnevddnnnnknlddhgned- ddd ddenmekkpesdvidqcssnnslrcswlqrkskvtevdvrivddevtikvvqkkkinclllvskvldqlqldl- hhv aggqigehysflfntkiyegstiyasaianrvievvdkhymaslpnsny AtbNP_172107 mggggmfeeigcfdpnapaemtaessfspaeppptitvigsnsnsncsledlsafhlspqdsslpasasaya SEQ ID hqlhinatpncdhqfqssmhqtlqgpsypqqsnhwdngyqdfvnlgpnhttpdllsllqlprsslppfanpsi No. 104 qdiimttsssvaaydplfhlnfplqppngsfmgvdqdqtetnqgvnlmydeennnlddglnrkgrgskkrki fptererrvhfkdrfgdlknlipnptkndrasivgeaidyikellrtidefkllvekkrvkqrnregddvvde nfkaqsevveqclinkknnalrcswlkrkskftdvdvriiddevtikivqkkkincllfvskvvdqleldlh hvagaqigehhsflfnakisegssvyasaiadrvmevlkkgymealsanngyhcyssd GlymXP_003548659 myeesscydpdammaegaedcfpqmvseseavmsatptqththntfaysyscgedaanangpiamehpqqnpy- n SEQ ID ysntqfveelysnqqftyhtptpdlldllhlpnpipgdnrtnvssysydpylhlnlqqqqqptlrellphmpa- lrn No. 105 dfpfggaaggddiqdfgnglvdftlqqevgkrrggkrtkqftsttterqrrvdlsskfdalkelipnpsksdr- asv vgdainyirelkrtveelkllvelkkrlekqrvmmrhkvetegessnldpaeyseslrsswiqrktkdtevdv- ri vdnevtiklvqrkkidclvhvshlldqlnldlqhvagghigdfcsylfntkicegssiyasaiankliqvmdt- s laaasla GlymXP_003524131 Mheqtgefdpntmgesvpflkdnfpqtlppspivvgnttnsnnnmdnhlvqevidaypyqlstwdpatvqelq- diay SEQ ID anhteqqqqqqqneqqfqqietqncsqsynnpssildppypspdllnllhmprcsasslltnpsicltnptqn- tpn No. 106 fqnpmaflgdltigsentsassvlydplfhlnlppqppalrelfqslprgyslptnsrngslfaggdemegdg- sq ldmgvlefnrvtpsvgkgrggkatkhfatekqrreqlngkykilrnlipsptkligwvwfntddrasvvgdai- dyi relirtvnelkllvekkryakerykrpkteedaaescnikpfgdpdggirtswlqrkskdsevdvriidddvt- ikl fqrkkincllfvskvldelqlelhhvagghvgeycsflfnskglvslrximegssvyasaianrvidvldsqy- ta avphtnsy GlymXP_003532668 mstgdrpkmhdqtgcfdpnttgesvpslkdnfpqtlppsspmvvgntttnsnnnmdnhlvqevidafpyqqst- wdpti SEQ ID vqelqdmayanhteqtqqqqqneqqfqqfetqncsqsynnpssildppypspdllnllhmprcsasslltnps- icltn No. 107 ptqntpnfqnpmaflgdlpigsentsassvlydplfhlnlppqppalrelfqslprgyslptnsrngslfggg- demeg dgsqldmgvlefnrvtltpsvgkgrrgkatkhfatekqrreqlngkykilrnlipsptklvgfvltqtdrasv- vgda idyirelirtvnelkllvekkryakdrckrpkteedaaescnikpfgdpdggirtswlqrkskdsevdvriid- ddvt iklfqrkkincllfvskvldelqlelhhvagghvgeycsflfnskglvslrximegssvyasaian rvidvldsqyaaavphtnsy LotjACN21644 maegvssqkdsfpqtlldpqpqslmvtenttnsnnimdnhlvqevidaplyqqstwdpnvqevqdmsyanhpe- qqf SEQ ID qhidaqnycqsytpsildpsypspdllnflhlptcsasslltnppnicisnptqrtpnfqnsmtflgdlpmgp- dnts No. 108 assvlydplfhlnlppqppalrelfqslprgyrlptssrddslfgggdemegdgsqldmgvldfnrdtasvgk- gre gkgakpfatekdrreqlngkykilrslipnptkligwvlfkpdrasvvgdaieyirelirtvnelkllvekkr- her erckrpkneedaeescnikpfgdpdgyirtswlqrkskdsevdvriidddvtikffqrkkincllfvskvlde- lq lelhhlagghvgeywsflfnskrpvsltqviegssvyasaianrvidvldsqyaaavpqtssy AtbAAL55717 mgcfdpntpaevtvessfsqaeppppppqvlvagstsnsncsveveelsefhlspqdcpqasstplqfhinpp- pp SEQ ID ppppcdqlhnnlihqmashqqqhsnwdngyqdfvnlgpnsattpdllsllhlprcslppnhhpssmlptsfsd- im No. 109 ssssaaavmydplfhlnfpmqprdqnqlrngscllgvedqiqmdanggmnvlyfegannnnggfeneilefnn- gv trkgrgsrksrtsptererrvhfndrffdlknlipnptkidrasivgeaidyikellrtieefkmlvekkrcg- rf rskkrarvgeggggedqeeeedtvnykpqsevdqscfnknnnnslrcswlkrkskvtevdvriiddevtiklv- qk kkincllfttkvldqlqldlhhvaggqigehysflfntkicegscvyasgiadtlmevvekqymeavpsngy AtbNP_180680 meeereslyeemgcfdpntpaevtvessfsqaeppppppqvlvagstsnsncsveveelsefhlspqdepqas- stp SEQ ID lqfhinppppppppcdqlhnnlihqmashqqqhsnwdngyqdfvnlgpnsattpdllsllhlprcslppnhhp- ssm No. 110 lptsfsdimssssaaavmydplfhlnfpmqprdqnqlrngscllgvedqiqmdanggmnvlyfegannnnggf- ene ilefnngvtrkgrgsrksrtsptererrvhfndrffdlknlipnptkidrasivgeaidyikellrtieefkm- lve kkrcgrfrskkrarvgeggggedqeeeedtvnykpqsevdqscfnkunnnslrcswlkrkskvtevdvriidd- evt iklvqkkkincllfttkvldqlqldlhhvaggqigehysflfntkicegscvyasgiadtlmevvekqymeav- psngy OsBAD67851 Mdeqrgrggfdelvllhqqqeqrrrreqqqeeeeeevrrqmfgavvgglaafpaaaaalgcqq- qvdcggelggfcdse SEQ ID aggssepeaaagarprggsgskrsaaevhnlsekrrrskinekmkalqslipnsnktdkasmldeaieylkql- qlqvq No. 111 mlsmrngvylnpsylsgalepaqasgmfaalggnnvtvvhpgtvmppvnqssgahhlfdlnsppqnqpqslil- psv pstaipeppfhlessqshlrqfqlpgssefhkilflhvllsvkdgvswrdnakappiitsrksarkrde lhqeriihvehq OsbHLH141_LOC_ mivgagyfedshdqslmagslihdsnqapassentsidlqkfkvhpystealsntanlaeaarainhlqhqle- i SEQ ID Os04g51070 dleqevppvetanwdpaictipdhiinhqfsedpqnilveqqiqqydsalypngvytpapdll- nlmqctmapafpa No. 112 ttsvfgdttlngtnyldlngeltgvaavpdsgsglmfasdsalqlgyhgtqshlikdichslpqnyglfpsed- er dviigvgsgdlfqpiddrqfdsvlecrrgkgefgkgkgkanfatererreqlnvkfrtlrmlfpnptkndras- i vgdaieyidelnrtvkelkilveqkrhgnnrrkvlkldqeaaadgesssmrpvrddqdnqlhgairsswvqrr skechvdvrivddevnikltekkkansllhaakvldefqlelihvvggiigdhhifmfntkvsegsavyaca vakkllqavdvqhqaldifn OsbHLH142_LOC_ myhpqcellmpleslemdvgqshlaaavaaampgelnfhllhsldaaaaaasstaasassqptvdyffggadq- qpppp SEQ ID Os01g18870 aamqydqlaaphhhqtvamlrdyygghyppaaaaaaateayfrggprtagssslvfgpaddes- afmvgpfessptprs No. 113 gggrkrsratagfhgggpangvekkekqrrlrltekynalmllipnrtkedratvisdaieyiqelgrtveel- tllv ekkrrrremqgdvvdaatssvvagmdqaaessegevmaaaamgavappprqapirstyiqrrsketfvdvriv- edd vnikltkrrrdgclaaasralddlrldlvhlsggkigdchiymfntkihsgspvfasavasrlievvdey OsUDT_LOC_ mprrarargggggggeevkveddfidsvlnfggggggeedgddgeeeqqqqqaaaaamgkefk- sknleaerrrrg SEQ ID Os07g36460 rlngnifalravvpkitkmskeatlsdaiehiknlqnevlelqrqlgdspgeawekqcsascs- esfvptenahyq No. 114 gqvelislgsckynlkifwtkraglftkvlealcsykvqvlslntisfygyaesfftievkgeqdvvmvelrs- ll ssivevpsi OsTDR_LOC_ mgrgdhllmknsnaaaaaaavngggtsldaalrplvgsdgwdyciywrlspdqrflemtgfcc- sseleaqvsal SEQ ID Os02g02820 ldlpssipldsssigmhaqallsnqpiwqssseeeeadggggaktrllvpvagglvelfasry- maeeqqmaelv No. 115 maqcggggagddgggqawpppetpsfqwdggadaqrlmyggsslnlfdaaaadddpflgggggdavgdeaaaa- g awpyagmavsepsvavaqeqmqhaagggvaesgsegrklhggdpeddgdgegrsggakrqqcknleaerkrrk- k lnghlyklrslvpnitkmdrasilgdaidyivglqkqvkelqdelednhvhhkppdvlidhpppaslvgldnd- d asppnshqqqpplavsgsssrrsnkdpamtddkvggggggghrmepqlevrqvqgnelfvqvlwehkpggfvr- l mdamnalglevinvnvttyktlvlnvfrvmvrdsevavgadrvrdsllevtretypgvwpspqeeddakfdgg- d ggqaaaaaaaaggehyhdevgggyhqhlhylafd
Yeast Two Hybrid (Y2H) Assay
[0065] The MATCHMAKER GAL4 Two-Hybrid System (Clontech, USA) was used for Y2H assays. Since both full-length EAT1 and TDR1 proteins were reported having self-activation (Ji et al., 2013), we made a truncated EAT1 (EAT1.sup.Δ, amino acids 1-254) and a truncated TDR1 (TDR.sup.Δ, amino acids 1-344) to reduce self activation. The full length cDNA of bHLH142 was cloned into pGAD-T7 (Clontech, USA), and full length bHLH142, EAT1, TDR, EAT1.sup.Δ, and TDR.sup.Δ were cloned into pGBK-T7 (Clontech, USA), respectively. The pairs of constructs to be tested were co-transformed into AH109 yeast cells and selected on plates containing Leu (for pGADT7 plasmid) and Trp (for pGBKT7 plasmid) dropout medium for 3-4 days at 30° C. Transformants were tested for specific protein interactions by growing on SD/-Leu/-Trp/-His plates with 30 mM 3-amino-1,2,4 triazole (3AT), and tested after X-α-Gal induction to confirm positive interaction. This system provides a transcriptional assay for detecting and confirming protein interactions in vivo in yeast.
Bimolecular Fluorescence Complementation (BiFC) Assay
[0066] BiFC assay allows visualization of protein-protein interactions in living cells and the direct detection of the protein complexes in subcellular compartments, providing insights into their functions. Full-length cDNAs of bHLH142, UDT1, TDR1, and EAT1 were independently introduced into pJET1.2 (Thermo Scientific). The sequence for the N-terminal amino acid residues 1-174 of YFP was then in-frame fused to the sequence of the C-terminal region of the tested proteins, while the sequence of the C-terminal amino acid residues 175-239 of YIP was in-frame fused to the sequence of the N-terminal end of the proteins. Next, the tested genes were introduced into pSAT5-DEST_CYN1 and pSAT4(A)-DEST_NYN1. Ballistic bombardment-mediated transient transformation in rice protoplasts was carried out following a previously published protocol36. Florescence images were photographed on a LSM 780 Plus ELYRA S.1 confocal microscope with Plan-Apochromat 40×/1.4 oil objective lens (Zeiss, Germany).
Co-Immunoprecipitation Assay
[0067] Recombinant proteins of bHLH142 and TDR1 fused with hemagglutinin (HA) tag were expressed in bacteria harboring pET-53-DEST (HIS-tag), and cell extracts after lysis were cleared by centrifugation at 12,000 rpm for 15 min, suspended in binding buffer (20 mM Tris-HCl, pH 7.9, 500 mM NaCl), and sonicated on ice for 30 s using an ultrasonic homogenizer (Misonix XL Sonicator Ultrasonic Cell Processor). The supernatants were purified using Ni2+ resin. For immunoprecipitation, extracts were pre-cleared by 30 min incubation with 20 μl of Pure Proteome Protein G Magnetic Beads (Millipore Co., Billerica, Mass.) at 4° C. with rotation. The antibodies (anti-bHLH142 or anti-HA) were then added to the pre-cleared extracts. After incubation for 4 h at 4° C., 40 μL of PureProteome Protein G Magnetic Beads was added, and the extracts were further incubated for 10 min at room temperature with rotation. After extensive washing, bound proteins were analyzed by western blotting. Rabbit antiserum against rice bHLH142 was produced using a synthetic peptide (CSPTPRSGGGRKRSR, SEQ ID No. 116) as antigen (GenScript Co).
RNAi-Mediated Gene Silencing of bHLH142
[0068] To generate an RNA intereference (RNAi) construct for suppressing the expression of bHLH142, a 149 bp fragment from 5'UTR region of bHLH142 was amplified by PCR with specific primers (Table 1) and cloned into pENTR (Invitrogen) to yield an entry vector pPZP200 hph-Ubi-bHLHI42 RNAi-NOS (12,483 bp). The RNAi construct was transformed into WT (TNG67) rice calli via Agrobacterium tumefaciens-mediated transformation system37. Transgenic plants were regenerated from transformed calli by selection on hygromycin-containing medium.
Examples
Identification of a New Male Sterility Rice Mutant
[0069] From the T2 population of Taiwan Rice Insertional Mutants (TRIM) (http://trim.sinica.edu.tw) lines we identified a T-DNA-tagged rice mutant (denoted ms142) with a completely MS phenotype. In the field, this mutant produced no viable seeds but maintained a normal vegetative growth (FIG. 1A), with panicles and spikelet developing similarly to those of the wild-type (WT) (FIGS. 1B to 1E). The ms142 mutant exhibits normal opening of spikelets and elongation of anther filaments, and its anthers are exerted completely in the husk (FIG. 1E). However, the anthers of ms142 were significantly smaller in size and appeared yellowish white (FIG. 1H), and there were no pollen dehiscence (FIGS. 1B and 1E) and pollen grain filling (FIGS. 1F and 1G). The anthers of the ms142 mutant could not be stained by Sudan black due to the lack of lipid accumulation (FIG. 1I) and showed no pollen grain development (FIG. 1K). As revealed later, ms142 with a full MS is a null mutant.
Sequence Analysis of the T-DNA-Tagged Gene in ms142 Mutant
[0070] To determine T-DNA insertion copy number, Southern blot analysis of T2 mutant lines using hptII as a probe was conducted, and only a single band was detected in the mutant lines (FIG. 2A). Thus, the mutation in ms142 is due to a single T-DNA insertion. Analysis of flanking sequence tag (FST) in the TRIM database suggests that ms142 is a putative mutant with T-DNA inserted at 1257 bp from the ATG start codon in the 3rd intron of bHLH142 (RAP Locus Os01g0293100, MSU Locus Os01g18870). The protein encoded by the gene is annotated as a basic helix-loop-helix dimerization region bHLH domain containing protein (RiceXPro Version 3.0). The bHLH142 gene comprises of four exons and three introns. Furthermore, genotyping by PCR using specific primers acrossing the T-DNA insertion site verified its FST (FIG. 2). The primers are designed for the mutant allele (R+RB, amplicon=340 bp) and wild-type allele (F+R, amplicon=458 bp). Three primers sets were incorporated in one PCR reaction for genotyping.
Agronomic Traits of ms142 Mutant and Genetic Study
[0071] The agronomic traits of the mutant were examined in the selfed progenies of heterozygous mutant grown in the outdoor GMO net house. Heterozygous plants behaved similarly to WT in terms of vegetative and reproductive growth and produced fertile seeds. However, homozygous ms142 mutant plants exhibited similar plant height, panicle number, and panicle length to the WT, but produced no viable seeds.
[0072] To understand whether the sterility in ms142 is due to male sterility or female sterility, homozygous mutant was backcrossed with WT pollen; and all F1 plants displayed WT-like phenotype in growth and fertility (data not shown). These results imply that the female organs of ms142 develop normally. When the ms142 BCF1 was selfed, the BCF2 progenies segregated into fertile and sterile plants in a ratio of 3:1 (Table 3), suggesting the MS trait is controlled by a recessive gene. Consistent with mutant phenotype, backcross segregants showed MS only in the homozygous plants, indicating that the MS phenotype co-segregated with the genotype. Moreover, when the selfed seeds derived from heterozygous plants of T2, T3, T4, and BCF2 generations were planted in different years and different cropping seasons, the scoring of phenotype indicated that MS in ms142 is stable and not affected by cropping season or year. Again, the fertile and sterile plants segregated approximately in a 3:1 ratio, supported by Chi-square analysis (data for T4 and BCF2 shown in Table 4). Taken together, these genetic analyses validate that the MS in ms142 is controlled by a single recessive locus.
TABLE-US-00004 TABLE 3 Genotype Trait WT like Heterozygote Homozygote Plant height (cm) 118.4 ± 7.9 116.7 ± 8 118.4 ± 7 Tiller number 8 ± 2.7 9 ± 2.4 13.8 ± 3.8 Panicle number 7.8 ± 2.7 8.7 ± 2.5 8.7 ± 2.1 Ave. panicle length (cm) 20.1 ± 1.4 20.1 ± 1.5 20.2 ± 1.6 Panicle Fwt (g) 30.8 ± 12.6 34 ± 12.5 8.8 ± 3.sup. Grain No./panicle 168.3 ± 28.1 164.3 ± 30.2 180 ± 31.2 Fertile grain number 1137 ± 466 1248 ± 464 .sup. 6 ± 4.2 Grain fertility (%) 84.9 ± 7.5 86.6 ± 6.sup. 0.4 ± 0.2 Grain yield/plant (g) 28.3 ± 11.3 31.3 ± 11.3 0.2 ± 0.1 1000 grain wt. (g) .sup. 25 ± 0.8 25.2 ± 1.sup. 6.8 ± 1.2 n = 27 62 22
TABLE-US-00005 TABLE 4 Cropping season Wild type Heterozygous Homozygous X2 BCF2 (n = 111) 1st Crop No. of plant 28 61 22 1.59 Grain fertility Fertile Fertile Sterile ms142 T4 (n = 119)a 1st Crop No. of plant 30 58 31 0.07 Grain fertility Fertile Fertile Sterile ms142 T4 (n = 120)a 2nd Crop No. of plant 29 57 34 0.71 Grain fertility Fertile Fertile Sterile
Defects in Anther Wall and Pollen Development in the ms142 Mutant
[0073] To determine the defects in the anthers of ms142, we examined the anatomy of anther in WT and homozygous mutant. At the microspore mother cell (MMC) stage, the WT anther walls contained epidermal cell layer, endothecial cell layer, middle layer and tapetal cell layer (FIG. 3A). During the early meiosis stage, the microspore mother cells underwent meiosis to form tetrads of haploid microspores; the tapetal cells differentiated to form large vacuole; and the middle layer cells began to degenerate (FIG. 3B). At tetrad stage, the meiocytes formed tetrads (FIG. 3C). During the young microspore stage, free microspores were released into the anther locule, and the microspores developed and exine was deposited on pollen grain wall. The middle layers shrinked and the tapetal cell layers became very dense (FIG. 3D). At the mature pollen stage, the uninucleate pollen developed to trinucleate pollen with starch, protein, lipid, and other nutrients enriched in the pollen cytoplasm. At maturity, the tapetal cells were completely degenerated and the endothecial cell layers were thickening, ready for anther dehiscence (FIG. 3E).
[0074] At the MMC stage, there were no visible differences in the anthers between WT and ms142. The ms142 anther consisted of normal epidermis, endothecium, middle layer and tapetum (FIG. 3F). During early meiosis stage, however, ms142 microspore mother cells did not enter meiosis and formed abnormal organelles (FIG. 3G, indicated by arrows). Abnormal endoplasmic reticulum structure and apoptosis was also observed by transmission electron microscopy. The ms142 tapetal cells continuously became vacuolated and elongated, with some cells divided into two tapetum layers (FIG. 3G). The mid-layers of the mutant tapetum maintained their initial shapes, but failed to divide into four cells at tetrad stage (FIG. 3H). The ms142 mutant's microspores finally degenerated during the vacuolated pollen stage. The tapetal and middle layer cells contained a large vacuole, and the middle layer cells did not degenerate (FIG. 3I). Consequently, there were no mature pollen grains formed in the locules at the mature stage. The mutant anther wall still retained four to five layers of cells, i.e. epidermis, endothecium, middle layer, and one or two layers of tapetum cells. By contrast, the endothecial cell layer did not become thickened in the mutant even at the latter stage of anther development (FIG. 3J).
Mutated bHLH142 Causes Defects in Tapetal PCD
[0075] Histological analysis indicated that ms142 has abnormal anther morphology and aborted degradation of tapetal cells (FIG. 3). Thus, we suspect that mutation of bHLH142 might have altered tapetal PCD, which is responsible for tapetal degeneration3,12-14. Tapetal PCD is characterized by cellular condensation, mitochondria and cytoskeleton degeneration, nuclear condensation, and internucleosomal cleavage of chromosomal DNA33. Therefore, we performed the TUNEL assay to detect DNA fragmentation in the anthers of WT and ms142. A TUNEL positive signal began to appear in the tapetal cells of WT during meiosis and a strong TUNEL signal was detected during the young microspore stages (FIG. 4). In contrast, no DNA fragmentation was observed in the tapetal layer in ms142 throughout anther development (FIG. 4).
bHLH142 is a Nuclear Protein
[0076] The gene structure of bHLH142, shown in FIG. 5A, indicates that bHLH domain contains bipartite nuclear localization signal (NLS), and the gene is predicted to encode a protein of 379 amino acids with a theoretical molecular mass of 40.7 kDa and pI of 6.2.
[0077] The nucleotide sequence of bHLH142 is shown below:
TABLE-US-00006 SEQ ID NO: 1 AAGAAACCAACTGCTTTCTCCTACCCAATATCACCCTTGCCCCTTTTAT ATACTCTTCCTCTCATCACCTTCTCGATCGGCCTCTCTCCTCTCCTCTC ATCAGCTCACACCCCCAACCAACAAACCTAGTTAATTTAGCTCTAGTTG GTTCATCCCTGCTGCACTGCGAGCTCAAGTAATCGATCTGAGCTCTGAA GAAAAAGGTGGTAGAGTGCGAGGAAGATGTATCACCCGCAGTGCGAGCT CCTGATGCCGCTTGAGAGCCTGGAGATGGACGTCGGCCAGTCGCACCTC GCCGCCGCCGTCGCAGCAGCCATGCCGGGGGAGCTCAACTTCCACCTCC TCCACTCGCTCGACGCCGCCGCGGCGGCTGCCTCCTCCACCGCCGCCTC GGCCTCCTCCCAGCCCACCGTCGACTACTTCTTCGGCGGCGCCGACCAG CAGCCGCCGCCGCCGGCGGCGATGCAGTACGACCAGCTGGCGGCGCCGC ACCACCACCAGACGGTGGCCATGCTGCGCGACTACTACGGCGGCCACTA CCCGCCGGCGGCGGCGGCGGCGGCGGCCACGGAGGCCTACTTCCGCGGC GGGCCAAGGACGGCCGGGTCGTCGTCGCTCGTGTTGGGGCCGGCCGACG ACGAGTCGGCCTTCATGGTCGGACCCTTCGAGAGCTCCCCGACGCCGCG GTCCGGCGGCGGCAGGAAGCGTAGCCGCGCCACCGCCGGCTTCCACGGC GGCGGGCCGGCCAACGGCGTCGAGAAGAAGGAGAAGCAGCGCCGCCTGC GGCTCACCGAGAAGTACAACGCCCTCATGCTCCTCATCCCCAACCGCAC CAAGGAGGATAGAGCGACGGTGATCTCAGACGCGATCGAGTACATCCAG GAGCTAGGGAGGACGGTGGAGGAGCTGACGCTGCTGGTGGAGAAGAAGC GGCGGCGGAGGGAGATGCAGGGGGACGTGGTGGACGCGGCGACCTCGTC GGTGGTGGCGGGGATGGATCAGGCGGCGGAGAGCTCGGAGGGCGAGGTG ATGGCGGCGGCGGCGATGGGCGCGGTGGCACCGCCGCCGCGGCAGGCGC CGATCCGGAGCACGTACATCCAGCGGCGGAGCAAGGAGACGTTCGTGGA CGTGCGGATCGTGGAGGACGACGTGAACATCAAGCTCACCAAGCGCCGC CGCGACGGCTGTCTCGCCGCCGCGTCGCGCGCGCTGGACGACCTCCGCC TCGACCTCGTCCACCTCTCCGGCGGCAAGATCGGCGACTGCCACATCTA CATGTTCAACACCAAGATTCATTCGGGATCTCCAGTGTTTGCAAGTGCA GTGGCCAGCAGGCTGATTGAAGTGGTGGATGAGTACTAACTAGCTCGAG CTAGCTAATTAGCCGACCGACCGATCGATATGATGAAAGTTTCTATGTT GCTAGCTAGCTAGGGTTCTTGGATGCATGAGTACTGAGTAGCTCTTTAA TTAATTTCCTTTTAATTTTAGACTGTTTAATTTGGATTGGTAAAGACTC GTGTTAGCTTTTGGGAGATCTTTGGTATGTCATGGTTTGCATGTATTAT TTTGGTCTACTTGGATAAATAATTGATGCTCTTTGAGACGTTAATTAAT.
[0078] The amino acid sequence of bHLH142 is shown below:
TABLE-US-00007 SEQ ID No: 2 MYHPQCELLMPLESLEMDVGQSHLAAAVAAAMPGELNFHLLHSL DAAAAAASSTAASASSQPTVDYFFGGADQQPPPPAAMQYDQLAA PHHHQTVAMLRDYYGGHYPPAAAAAAATEAYFRGGPRTAGSSSL VFGPADDESAFMVGPFESSPTPRSGGGRKRSRATAGFHGGGPAN GVEKKEKQRRLRLTEKYNALMLLIPNRTKEDRATVISDAIEYIQ ELGRTVEELTLLVEKKRRRREMQGDVVDAATSSVVAGMDQAAES SGEVMAAAAMGAVAPPPRQAPIRSTYIQRRSKETFVDVRIVEDD VNIKLTKRRRDGCLAAASRALDDLRLDLVHLSGGKIGDCHIYMF NTKIHSGSPVFASAVASRLIEVVDEY
[0079] Since the bHLH proteins are characterized as TFs, we assumed that bHLH142 is localized in the nucleus. To verify its subcellular localization, we constructed a fusion gene of the green fluorescent protein gene (GFP) and bHLH142 under the control of the 35S promoter and the nos terminator for transient expression in rice leaf mesophyll protoplasts (FIG. 5C). As a positive control, NLS sequence was also fused to red fluorescent protein (RFP) gene using the same regulatory elements. These constructs were introduced into rice protoplasts by particle bombardment. As expected, with the GFP construct alone, free GFP was found in the nucleoplasm as well as in the cytoplasm. However, bHLH142:GFP fusion protein and the positive control of NLS:RFP were exclusively located in the nucleus (FIG. 5D). These results confirm that, as a TF, bHLH142 protein is localized in the nucleus.
Phylogenetic Analysis
[0080] To understand the evolutionary relationship of bHLH142 among various organisms, we used full-length bHLH142 protein sequence to NOM BLAST database and retrieved 21 homologs containing bHLH domain from 10 diverse terrestrial plants. The phylogenetic tree shows that UDT1 (bHLH164) and TDR1 (bHLH5) are in the same cluster, while bHLH142 and EAT1 (bHLH141) evolved and diversified into two separate clades. Phylogenetic analysis also suggests that bHLH142 is descended from a common ancestor of monocots. Rice bHLH142 shares a high similarity with the related proteins from Brachypodium distachyon, millet (Setaria italica), Triticum urartu, maize (Zea may), Sorghum (Sorghum bicolor) and Aegilops tauschii (FIG. 6). The conserved homologs of bHLH142 from the important cereal crops, such as maize, millet, sorghum and wheat, share 84.1%, 79.2%, 72.2%, and 78% similarity in amino acid sequence to the rice counterpart (Table 5). The maize homolog, GRMZM2G021276, is highly expressed in immature tassel and meiotic tassel and anther. In accordance, our RT-PCR data also verified that maize homolog is tissue specifically expressed in meiotic anther (FIG. 14). This result implies that the maize bHLH142 homolog may also play a similar role in anther and pollen development.
TABLE-US-00008 TABLE 5 Crop Accession No. Amino acid similarity Sorghum XP 002457706 72.2% Maize ZmLOC100283549 84.1% Wheat EMS50437 78.0% Millet XP_004967599 79.2% Brachypodium XP_003567568 80.7%
Expression Pattern of bHLH142
[0081] Both RT-PCR and qRT-PCR analyses with WT showed that the bHLH142 mRNA is accumulated in young rice panicle and anther only, but not in other tissues (e.g. root, shoot, leaf, lemma, palea, ovary, and seed). In particular, high levels of transcripts were found in developing panicles (FIGS. 7A and 7B). Specifically, bHLH142 transcripts were highly expressed in meiocyte mother cells (MMC) and extremely highly expressed in the anther at meiosis stage (FIG. 7C). Also, in-situ hybridization (ISH) clearly demonstrated the specific expression of bHLH142 in the anther but not in lemma and palea of WT spikelet at early meiosis stage (FIG. 7D). ISH with the cross sections of WT anther at various developmental stages showed positive signals in the tapetal layers at early meiosis stage and in the tapetal layers and meiocytes during meiosis stage, with decreasing signals at young microspore stage and negligible signals after vacuolated pollen stage. Interestingly, ISH signal was also detected in the vascular cells (FIG. 8), suggesting that the target genes of bHLH142 might also be associated with nutrient acquisition in the anther. On the contrary, there was no ISH signal detected in the anthers of the null mutant of ms142 (FIG. 8).
[0082] In addition, the expression patterns of various known pollen regulatory genes in the anther of WT versus ms142, as examined by qRT-PCR, confirmed the knockout of bHLH142 transcript in the ms142 null mutant (FIG. 9A). Also, expression of TDR1, bHLH141 (EAT1), AP37, AP25, CP1, CYP703A3, CYP704B2, MS2 and C6 was significantly down-regulated in the ms142 anther, relative to the WT anthers (FIG. 9E, 9G-9M). However, MSP1 and UDT1 transcripts were up-regulated in the mutant at MMC and meiosis stages (FIG. 9B-9C). There was no significant change in the GAMYB transcripts in ms142 (FIG. 9D). Interestingly, the suppression of TDR1 expression in ms142 was less, compared to other downstream genes (FIG. 9E-9M). In WT anther, bHLH142 transcript was more abundant during meiosis stage, but it was negligible in the ms142 null mutant. Interestingly, EAT1 mutant also showed a lower amount of bHLH142 mRNA during meiosis stage (FIG. 10). We also compared EAT1 mRNA expression in these mutants to gain more insights into the regulatory interplay of these TFs in pollen development. In WT, EAT1 was expressed slightly later (at young microspore stage) than bHLH142 (at meiosis stage) (FIG. 10). Interestingly, ms142 anther exhibited a similar amount of EAT1 mRNA to WT anthers at MMC, but tended to decline after young microspore stage (FIG. 10). Taken together, these data suggest that bHLH142 plays a role in the downstream of UDT1, but upstream of EAT1.
bHLH142 and TDR1 Coordinately Regulate EAT1 Promoter Activity
[0083] Based on the alternations in expression of known pollen regulatory genes in the different mutants (FIG. 9 and FIG. 10), we assumed that bHLH142 might regulate EAT1 promoter activity and carried out transient promoter assays with EAT1.sub.pro-Luc construct. Our results demonstrated that bHLH142 or TDR1 protein alone cannot drive the expression of EAT1.sub.pro-Luc independently. However, when combined, these two TF proteins together significantly increased Luc expression from the EAT1 promoter by up to 30 fold. However, additional expression of EAT1 in the same cells reduced Luc expression from a 30-fold down to an 18-fold increase, presumably due to the competition between EAT1 and bHLH142 in binding to TDR1 (FIG. 11). Apparently, TDR1 and bHLH142 co-regulate the activity of EAT1 promoter.
Protein Interactions Among bHLH142, TDR1 (bHLH5) and EAT1 (bHLH141)
[0084] We performed yeast two-hybrid analysis to determine whether bHLH142, as bait, interacts with the prey, TDR1 or EAT1. As previously reported that full-length EAT1 and TDR1 proteins possess self-activation activity in nature23,24, our Y2H study also confirmed this phenomenon (FIG. 12A). Therefore, we also constructed a truncated EAT1.sup.Δan(1-254) (truncated EAT1 at amino acids 1-254) and TDR.sup.Δna(1-344) (truncated TDR amino acids at 1-344) to eliminate self activation (FIG. 12A). Our results showed bHLH142 is not self-activated (FIG. 12A); only the yeast strains co-expressing both bHLH142 and TDR.sup.Δan(1-344) grew normally on stringent selection media (FIG. 13A) and there was no direct interaction between bHLH142 and EAT1.sup.Δaa(1-254). Thus, bHLH142 was not directly interacting with EAT1 as demonstrated in the yeast cells (FIG. 10A), and the retention of the C-terminal sequences of TDR1 is sufficient to confer the interaction of the two proteins. Clearly, the amino acid sequences in TDR.sup.Δaa(1-344) and EAT1.sup.Δaa(1-254) contain the interacting sites, consistent with the previous study24. These results are further supported by our results of EAT1 promoter assays in that both bHLH142 and TDR1 are required for the transcription of EAT1 (FIG. 11). Moreover, bimolecular fluorescent complementation (BiFC) assay showed that yellow fluorescent protein (YFP) signals are detected only in the nucleus of the rice cells co-expressing both NYN1-bHLH142 and CYN1-TDR1 and in the cells co-expressing both NYN1-TDR1 and CYN1-EAT1, but not in the cells co-expressing both NYN1-bHLH142 and CYN1-EAT1 (FIG. 13B). In vitro interaction of bHLH142 and TDR1 proteins was further validated by co-immunopreciptation (Co-IP) experiment, where interaction between HA fused TDR1 and bHLH142 was confirmed (FIGS. 13C and 13D). Taken together, all of these molecular data provide solid evidences of the physical interaction between TDR1 and bHLH142.
RNAi Transgenic Rice Lines Validate the Role of bHLH142 in Pollen Development
[0085] To further validate the biological function of bHLH142, we generated an RNA interference (RNAi) construct to suppress the expression of bHLH142 in rice. The gene specific region from the 5'UTR of bHLH142 was, amplified, fused with β-glucuronidase (GUS) intron and introduced into WT calli via Agrobacterium tumefaciens. All 16 TO RNAi transgenic lines obtained had a MS phenotype similar to the T-DNA mutant ms142. These RNAi lines showed reduced expression of bHLH142, as examined by RT-PCR, and produced poorly developed anthers without pollen grains (FIG. 14). This result further supports the notion that bHLH142 plays a key role in rice anther and pollen development. The RNAi fragment (SEQ ID No. 120):
TABLE-US-00009 caacaaacctagttaatttagctctagttggttcatccctgctgca ctgcgagctcaagtaatcgatctgagctctgaagaaaaaggtggta gagtgcgaggaagatgtatcacccgcagtgcgagctcctgatgccg cttgagagcct
Overexpression bHLH142 Caused Male Sterility
[0086] For functional genomic study, we constructed overexpressing bHLH142 driven by constitutively express Ubiquitin promoter (FIG. 15A) and introduced into wild-type (TNG67 background) calli via Agrobacterium tumefaciens38 Genomic PCR confirmed T-DNA insertion of target gene and selection marker hygromycin (lower panel) PCR has band in the 23 tested TO transgenic lines (FIG. 15B).
[0087] Interesting to observe that overexpressing bHLH142 TO transgenic plants all showed grain sterility (FIG. 16). Ubi::bHLH142 transgenic lines have similar plant type and panicle length (FIGS. 168 and 16C) with the WT except no grain filling after anthesis stage (FIG. 16A). It was observed anthers of overexpression transgenic lines were shorter and showed light yellow color than the WT (FIG. 16D). After seed maturation stage, grain of WT was filled with starch but there was no viable seed in those overexpressing lines (FIGS. 16E and 16F).
[0088] By using RT-PCR, we detected some regulatory genes associated with pollen development in rice. As expected, overexpression line constitutively express abundant of bHLH142 transcripts during various stages of anther development. Interestingly, Udt1 was simultaneously upregulated in the overexpression line. EAT1 mRNA also prematurely upregulated before meiosis stage but decrease its expression at latter stage of anther development. However, MS2 was significantly downregulated in the overexpressing line (FIG. 17). Defect of grain fertility in overexpressing line presumably due to prematurely upregulation of tapetal program cell death genes such as UDT120 and EAT123. Moreover, downregulation of MS2 that reported contribution to pollen exine development might associate with the defect of pollen development in the overexpression line.
Heterologuos Overexpression bHLH142 Confers Male Sterility in Maize
[0089] Since bHLH142 shares high identity with maize38, and gene specific primer sets were designed from homolog of maize ZmLOC100283549 (denoted Zm-142). RT-PCR indicated that Zm-142 was not expressed in vegetative organs of maize such as leaf, root, shoot, and stamen. Interestingly, Zm-142 specifically expressed in floret of 1 mm to 7 mm length but not detectable at later stage and in the mature pollen (FIG. 18). The expression pattern of Zm-142 was similar to bHLH14238.
[0090] Therefore, we use the similar construct of overexpression bHLH142 in FIG. 15A was transformed into maize (in cultivar Crystal White background) using agrobacterium-mediated pollen transformation method. One transgenic maize showed obvious male sterility phenotype with smaller angle of tassel branch (FIG. 19A, right panel) than the WT (FIG. 19A, left panel). Closed up tassel during anthesis stage, WT has larger anther than transgenic line and it has normal opening of spikelets and elongation of anther filaments during anthesis stage (FIG. 19B, left panel). Whilst, anther of transgenic maize were significantly smaller in size and anther was completely no elongation of anther filaments (FIG. 19B, right panel). Morphology of spikelet of WT at one day before anthesis was shown in FIG. 19C (left panel) with long and fat anther, but anther of transgenic line was short and shrinkage (FIG. 19C, right panel). Stained of maize mature pollen grains with I2/KI solution, and the fertile WT pollen grains stained with dark red color. However, pollens of transgenic line could not be stained and transparent due to no starch accumulation. That implied transgenic line was male sterile (FIG. 19D).
Overexpression bHLH142 Induces Reversible Male Sterility in Low Temperature
[0091] bHLH142-overexpressed plants also showed a completely male sterile phenotype during summer season (FIG. 20, left panel). By contrast, during winter low temperature conditions, anthers of overexpression transgenic line produce many pollen grains inside the locules and their pollen grains can be stained by I2/KI solution, indicating that the plants have restored the fertility (FIG. 20, right panel). Therefore, this novel functionality nature of our target bHLH142 has a big advantage over other genetic MS (GMS) genes for hybrid crop production. This reversible pollen fertility trait makes it more desirable in producing hybrid crop seeds just in one cross without the need to maintain the seed stocks of the MS lines as with cytoplasmic MS (CMS). In addition, biotech companies are known to prefer adopting overexpression over suppression approach in generating transgenic lines because overexpression lines are more stable than RNAi or antisense knock-down lines.
[0092] Rice bHLH142 have homologous in maize, sorghum and wheat, and they share more than 70% similarity in amino acid sequence to the rice counterpart (Table 3). This will benefit to genetic engineering male sterile for F1 hybrid seed production and generating hybrid vigor (heterosis) in terms of growth and grain yield in cereal crops.
bHLH142 is a New Major Regulator of Rice Anther Development
[0093] So far, three of the bHLH TFs have been shown to be involved in pollen development in rice and mutations of these TF genes all lead to complete MS, including UDT1 (bHLH164)20, TDR1 (bHLH514, and EAT1/DTD1 (bHLH141)23,24. They all play an important role in pollen development by regulating tapetal PCD. In this invention, we identified a novel rice MS mutant, ms142 (FIG. 1), with T-DNA inserted in the intron of bHLH142, which encodes another basic helix-loop-helix dimerization region bHLH domain containing TF protein. The phenotype is characterized by having small anthers without pollen grain development (FIG. 1). Genetic analyses suggest that the mutation is due to a single T-DNA insertion event. We further show that this TF is located in the nuclei (FIG. 5) and plays an essential role in regulating rice pollen development. Close anatomical examination of anther development, in parallel with TUNEL assay of DNA degradation and ISH of key gene transcripts, in the null mutant demonstrate that defects in microspore development is associated with defects in tapetal PCD. Timely degradation of tapetum cells is essential for viable pollen development. Furthermore, suppressed expression of bHLH142 in WT rice by RNAi confers the MS phenotype (FIG. 21). Thus, this invention identifies the involvement of another bHLH TF in the dynamic regulation of pollen development in rice and likely in other plants as well.
[0094] Our analysis of expression profile of known regulatory genes involved in pollen development demonstrates the down-regulation of several genes, such as TDR1, EAT1, AP37, CP1, C6, MS2, etc. in ms142 during pollen development (FIG. 9). Therefore, we suggest that bHLH142 participates in the same regulatory pathway of anther development, with a higher gene hierarchy (FIG. 10). An earlier study reports that TDR1 positively regulates CP1 and C614, while a recent study suggests that EAT1 interacts with TDR1 in regulating the expression of two aspartic protease genes, AP25 and AP3723. Another study also reported that mutation in DTD (same as EAT1, bHLH141) in rice results in severe MS24. In consistent with this report, we also find that the NIAS Tos17 mutant H0530, a knockout of EAT1 (bHLH141) gene, also fails to produce pollens (FIG. 10B). However, the eat1/dtd mutant exhibits normal meiosis process, but ms142 cannot exceed beyond meiosis cell division, which may be due to the extra role played by bHLH142. Consistent with this notion, our ISH analysis revealed that, besides tapetal layer, bHLH142 is also expressed in meiocytes as well as in the vascular bundle (FIG. 8). This result suggests that bHLH142 might also play an additional role in nutrient acquisition for cell plate formation during microspore development.
[0095] This invention uncovers bHLH142 as another critical factor in the bHLH TF family for pollen development, besides UDT1 (bLHL164), TDR1 (bLHL5) and EAT1 (bHLH141). Our mutagenesis analysis suggests that the gene hierarchy of bHLH142 is in the downstream of UDT1 (bHLH164) but upstream of TDR1 (bHLH5) and EAT1 (bHLH141) (FIG. 10 and FIG. 9). Interestingly, all these 4 bHLH TFs are tissue specifically expressed in the anther and participate in the important process of sequential pollen development events, particularly in tapetal PCD. Thus, it is rather unique in that several of the bHLH TFs coordinate in regulating the anther development; and it is likely that more TFs might be involved in controlling the regulatory network to ensure normal pollen development.
[0096] Also, we noticed a lower suppression in expression of TDR1 in ms142, compared to other downstream genes in the regulatory network, which may be attributed to the fact that TDR1 is also known to be regulated by another TF GAMYB22. In agreement, we also found that the expression of GAMYB is not altered in ms142 (FIG. 9). Taken together, these results suggest that two parallel pathways may exist in the regulatory circuit leading to TDR1 during pollen development.
bHLH142 Functions Coordinately with TDR1 to Regulate EAT1 Promoter
[0097] Since TDR1 and EAT1 mRNA are both down-regulated in ms142, we hypothesize that TDR1 interacts with bHLH142 and positively regulate EAT1 promoter for transcriptional activities of AP25 and AP37, encoding aspartate proteases for tapetal PCD. Our promoter transient assay provides solid evidence that bHLH142 and TDR1 work coordinately in regulating EAT1 promoter (FIG. 11). We also demonstrates that additional expression of EAT1 protein significantly reduced EAT1-Luc promoter strength from a 30 fold down to 18 fold increase (FIG. 11), which may be attributed to the competition between bHLH142 and EAT to interact with TDR1. Presumably, more EAT1 favors TDR1-EAT1 interaction and might consequently reduce the interaction between bHLH142 and TDR1, therefore reducing EAT1 transcriptional activation (FIG. 11). It is likely that bHLH142 interacts with TDR1 and TDR1 in turn interacts with EAT1 and bHLH142 does not directly interact with EAT1 (FIG. 13). Whether some other TFs may be required to regulate the transcription of bHLH142 is worth further investigation to unravel the entire regulatory gene hierarchy.
[0098] Our molecular studies provide solid in vivo (Y2H, BiFC) and in vitro (co-IP) evidences that both bHLH142 and TDR1 can form protein interaction (FIG. 13). The co-IP provides the most convincing evidence that the two proteins physically interact in vitro. Subcellular localization also demonstrates that bHLH142 protein is localized in the nucleus (FIGS. 5D and 13B) and its protein is not self activated (FIG. 13 and FIG. 12). Since we also found self activation of full length TDR1 and EAT1 in our Y2H experiment (FIG. 12), N-terminal truncated forms of TDR.sup.Δaa(1-344) and EAT1.sup.Δaa(1-254), were used in our experiment to reduce self activation. These two N-terminal truncated protein forms did not exhibit self activation in yeast cells (FIG. 12A). Therefore, we are confident that bHLH142 interacts with TDR1 by using these truncated proteins to eliminate the bias (FIG. 13). Our data indicate that bHLH142 interacts with TDR1 in the C-terminal (FIG. 13), and support the conclusion of the previous study in that DTD/EAT1 (bHLH141) interacts with TDR1 in the C-terminal region24. In other words, both bHLH142 and EAT1 (bHLH141) can interact with TDR1 in the C' terminal of TDR1. This finding also supports the result of our EAT1 promoter assay, where additional EAT1 protein reduces EAT1 promoter activity, presumably due to the competition between bHLH142 and EAT1 proteins in the C' terminal of TDR1. Based on this and previous works, the current regulatory network for rice pollen development is presented in FIG. 21. Previous works with various rice MS mutants suggest that UDT1 and GAMYB may positively regulate the transcription of TDR122 and TDR1 in turn controls the transcription of C6 and CP114. A recent study presents evidence that TDR1 interacts with EAT1 for its direct regulation of the expression of two aspartate proteinase genes for initiation of tapetal PCD23. In this invention, we demonstrate that bHLH142 acts downstream of UDT1 but upstream of TDR1 and EAT1, and then bHLH142 interact with TDR1 in activating EAT1 transcription (indicated by red arrows in FIG. 21). Furthermore, we showed that EAT1 also positively regulate the transcription of AP37 and CP1 directly, two proteins involved in tapetal PCD at late pollen development stage.
REFERENCES
[0099] 1. Khush, (2000), Rice Germplasm enhancement at IRK Phillipp. J. Crop Sci. 25, 45-51,
[0100] 2. Gao et al., (2013), Dissecting yield-associated loci in super hybrid rice by resequencing recombinant inbred lines and improving parental genome sequences. Proc Natl Acad Sci USA 110, 14492-14497.
[0101] 3. Lao et al., (2013), A detrimental mitochondrial-nuclear interaction causes cytoplasmic male sterility in rice. Nat Genet 45, 573-577.
[0102] 4. Luo, et al., (1992), Pei'ai 64S, a dual purpose sterile line whose sterility is induced by low critical temperature. Hybrid Rice 1:27-29.
[0103] 5. Zhou et al., (2012), Photoperiod- and thermo-sensitive genic male sterility in rice are caused by a point mutation in a novel noncoding RNA that produces a small RNA. Cell Res 22, 649-660.
[0104] 6. Zhang et al., (2010), Carbon starved anther encodes a MYB domain protein that regulates sugar partitioning required for rice pollen development. Plant Cell 22, 672-689.
[0105] 7. Zhang et al., (2013), Mutation in CSA creates a new photoperiod-sensitive genic male sterile line applicable for hybrid rice seed production. Proc Natl Acad Sci USA 110, 76-81,
[0106] 8. Li et al., (2007), Suppression and restoration of male fertility using a transcription factor. Plant Biotechnol J 5, 297-312.
[0107] 9. Goldberg et al., (1993). Anther development: basic principles and practical applications. Plant Cell 5: 1217-1229.
[0108] 10. Zhu et al., (2008). Defective in Tapetal development and function 1 is essential for anther development and tapetal function for microspore maturation in Arabidopsis. Plant J. 55: 266-277.
[0109] 11. Wu and Cheun, (2000). Programmed cell death in plant reproduction. Plant Mol. Biol. 44: 267-281.
[0110] 12. Papini et al., (1999), Programmed-cell-death events during tapetum development of angiosperms. Protoplasma 207: 213-221.
[0111] 13. Kawanabe et al., (2006). Abolition of the tapetum suicide program ruins microsporogenesis. Plant Cell Physiol. 47: 784-787.
[0112] 14. Li et al., (2006a), The rice tapetum degeneration retardation gene is required for tapetum degradation and anther development. Plant Cell 18, 2999-3014.
[0113] 15. Ito and Shinozaki, (2002), The male sterility1 gene of Arabidopsis, encoding a nuclear protein with a PHD-finger motif, is expressed in tapetal cells and is required for pollen maturation. Plant Cell Physiol 43, 1285-1292,
[0114] 16. Sorensen et al., (2003), The Arabidopsis ABORTED MICROSPORES (AMS) gene encodes a MYC class transcription factor. Plant J 33, 413-423.
[0115] 17. Zhang et al., (2007), Transcription factor AtMYB 103 is required for anther development by regulating tapetum development, callose dissolution and exine formation in Arabidopsis. Plant J 52, 528-538,
[0116] 18. Wilson et al., (2001), The Arabidopsis MALE STERILITY1 (MS1) gene is a transcriptional regulator of male gametogenesis, with homology to the PHD-finger family of transcription factors. Plant J 28, 27-39.
[0117] 19. Ito et al., (2007), Arabidopsis MALE STERILITY1 encodes a PHD-type transcription factor and regulates pollen and tapetum development. Plant Cell 19, 3549-3562.
[0118] 20. Jung et al., (2005), Rice Undeveloped Tapetum1 is a major regulator of early tapetum development. Plant Cell 17, 2705-2722.
[0119] 21. Aya et al., (2009), Gibberellin modulates anther development in rice via the transcriptional regulation of GAMYB. Plant Cell 21, 1453-1472.
[0120] 22. Liu et al., (2010), Identification of gamyb-4 and analysis of the regulatory role of GAMYB in rice anther development. J Integr Plant Biol 52, 670-678.
[0121] 23. Niu et al., (2013), EAT1 promotes tapetal cell death by regulating aspartic proteases during male reproductive development in rice. Nat Commun 4, 1445,
[0122] 24. Ji et al., (2013), A Novel Rice bHLH Transcription Factor, DTD, Acts Coordinately with TDR in Controlling Tapetum Function and Pollen Development. Mol Plant.
[0123] 25. Lee et al., (2004), Isolation and characterization of a rice cysteine protease gene, OsCP1, using T-DNA gene-trap system. Plant Mol Biol 54, 755-765.
[0124] 26. Li et al., (2006b), Genome-wide analysis of basic/helix-loop-helix transcription factor family in rice and Arabidopsis. Plant Physiol 141, 1167-1184.
[0125] 27. Carretero-Paulet et al., (2010), Genome-wide classification and evolutionary analysis of the bHLH family of transcription factors in Arabidopsis, poplar, rice, moss, and algae. Plant Physiol 153, 1398-1412.
[0126] 28. Bailey et al., (2003), Update on the basic helix-loop-helix transcription factor gene family in Arabidopsis thaliana. Plant Cell 15, 2497-2502.
[0127] 29. Toledo-Ortiz et al., (2003), The Arabidopsis basic/helix-loop-helix transcription factor family. Plant Cell 15, 1749-1770.
[0128] 30. Ptashne, (1988), How eukaryotic transcriptional activators work. Nature 335, 683-689.
[0129] 31. Massari and Murre, (2000), Helix-loop-helix proteins: regulators of transcription in eucaryotic organisms, Mol Cell Biol 20, 429-440.
[0130] 32. Hsing et al., (2007), A rice gene activation/knockout mutant resource for high throughput functional genomics. Plant Mol Biol 63, 351-364,
[0131] 33. Phan et al., (2011). The MYB80 transcription factor is required for pollen development and the regulation of tapetal programmed cell death in Arabidopsis thaliana. Plant Cell 23: 2209-2224.
[0132] 34. Bart et al., (2006), A novel system for gene silencing using siRNAs in rice leaf and stem-derived protoplasts. Plant Methods 2, 13.
[0133] 35. Tamura et al., (2011), MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol 28, 2731-2739.
[0134] 36. Hsu et at, (2011), Integration of molecular biology tools for identifying promoters and genes abundantly expressed in flowers of Oncidium Gower Ramsey. BMC Plant Biol 11, 60.
[0135] 37. Chan et al., (1993), Agrobacterium-mediated production of transgenic rice plants expressing a chimeric alpha-amylase promoter/beta-glucuronidase gene. Plant Mol Biol 22, 491-506.
[0136] 38. Ko et al., (2014). The bHLH142 Transcription Factor Coordinates with TDR1 to Modulate the Expression of EAT1 and Regulate Pollen Development in Rice. Plant Cell 26, 2486-2504.
[0137] 39. Bi et al. (2005). The rice nucellin gene ortholog OsAsp1 encodes an active aspartic protease without a plant-specific insert and is strongly expressed in early embryo. Plant Cell Physiol 46, 87-98.
Sequence CWU
1
1
12011617DNAOryza sativa 1aagaaaccaa ctgctttctc ctacccaata tcacccttgc
cccttttata tactcttcct 60ctcatcacct tctcgatcgg cctctctcct ctcctctcat
cagctcacac ccccaaccaa 120caaacctagt taatttagct ctagttggtt catccctgct
gcactgcgag ctcaagtaat 180cgatctgagc tctgaagaaa aaggtggtag agtgcgagga
agatgtatca cccgcagtgc 240gagctcctga tgccgcttga gagcctggag atggacgtcg
gccagtcgca cctcgccgcc 300gccgtcgcag cagccatgcc gggggagctc aacttccacc
tcctccactc gctcgacgcc 360gccgcggcgg ctgcctcctc caccgccgcc tcggcctcct
cccagcccac cgtcgactac 420ttcttcggcg gcgccgacca gcagccgccg ccgccggcgg
cgatgcagta cgaccagctg 480gcggcgccgc accaccacca gacggtggcc atgctgcgcg
actactacgg cggccactac 540ccgccggcgg cggcggcggc ggcggccacc gaggcgtact
tccgcggcgg gccaaggacg 600gccgggtcgt cgtcgctcgt gttcggcccg gccgacgacg
agtcggcctt catggtcgga 660cccttcgaga gctccccgac gccgcggtcc ggcggcggca
ggaagcgtag ccgcgccacc 720gccggcttcc acggcggcgg gccggccaac ggcgtcgaga
agaaggagaa gcagcgccgc 780ctgcggctca ccgagaagta caacgccctc atgctcctca
tccccaaccg caccaaggag 840gatagagcga cggtgatctc agacgcgatc gagtacatcc
aggagctagg gaggacggtg 900gaggagctga cgctgctggt ggagaagaag cggcggcgga
gggagatgca gggggacgtg 960gtggacgcgg cgacgtcgtc ggtggtggcg gggatggatc
aggcggcgga gagctcggag 1020ggcgaggtga tggcggcggc ggcgatgggc gcggtggcac
cgccgccgcg gcaggcgccg 1080atccggagca cgtacatcca gcggcggagc aaggagacgt
tcgtggacgt gcggatcgtg 1140gaggacgacg tgaacatcaa gctcaccaag cgccgccgcg
acggctgtct cgccgccgcg 1200tcgcgcgcgc tggacgacct ccgcctcgac ctcgtccacc
tctccggcgg caagatcggc 1260gactgccaca tctacatgtt caacaccaag attcattcgg
gatctccagt gtttgcaagt 1320gcagtggcca gcaggctgat tgaagtggtg gatgagtact
aactagctcg agctagctaa 1380ttagccgacc gaccgatcga tatgatgaaa gtttctatgt
tgctagctag ctagggttct 1440tggatgcatg agtactgagt agctctttaa ttaatttcct
tttaatttta gactgtttaa 1500tttggattgg taaagactcg tgttagcttt tgggagatct
ttggtatgtc atggtttgca 1560tgtattattt tggtctactt ggataaataa ttgatgctct
ttgagacgtt aattaat 16172379PRTOryza sativa 2Met Tyr His Pro Gln Cys
Glu Leu Leu Met Pro Leu Glu Ser Leu Glu 1 5
10 15 Met Asp Val Gly Gln Ser His Leu Ala Ala Ala
Val Ala Ala Ala Met 20 25
30 Pro Gly Glu Leu Asn Phe His Leu Leu His Ser Leu Asp Ala Ala
Ala 35 40 45 Ala
Ala Ala Ser Ser Thr Ala Ala Ser Ala Ser Ser Gln Pro Thr Val 50
55 60 Asp Tyr Phe Phe Gly Gly
Ala Asp Gln Gln Pro Pro Pro Pro Ala Ala 65 70
75 80 Met Gln Tyr Asp Gln Leu Ala Ala Pro His His
His Gln Thr Val Ala 85 90
95 Met Leu Arg Asp Tyr Tyr Gly Gly His Tyr Pro Pro Ala Ala Ala Ala
100 105 110 Ala Ala
Ala Thr Glu Ala Tyr Phe Arg Gly Gly Pro Arg Thr Ala Gly 115
120 125 Ser Ser Ser Leu Val Phe Gly
Pro Ala Asp Asp Glu Ser Ala Phe Met 130 135
140 Val Gly Pro Phe Glu Ser Ser Pro Thr Pro Arg Ser
Gly Gly Gly Arg 145 150 155
160 Lys Arg Ser Arg Ala Thr Ala Gly Phe His Gly Gly Gly Pro Ala Asn
165 170 175 Gly Val Glu
Lys Lys Glu Lys Gln Arg Arg Leu Arg Leu Thr Glu Lys 180
185 190 Tyr Asn Ala Leu Met Leu Leu Ile
Pro Asn Arg Thr Lys Glu Asp Arg 195 200
205 Ala Thr Val Ile Ser Asp Ala Ile Glu Tyr Ile Gln Glu
Leu Gly Arg 210 215 220
Thr Val Glu Glu Leu Thr Leu Leu Val Glu Lys Lys Arg Arg Arg Arg 225
230 235 240 Glu Met Gln Gly
Asp Val Val Asp Ala Ala Thr Ser Ser Val Val Ala 245
250 255 Gly Met Asp Gln Ala Ala Glu Ser Ser
Glu Gly Glu Val Met Ala Ala 260 265
270 Ala Ala Met Gly Ala Val Ala Pro Pro Pro Arg Gln Ala Pro
Ile Arg 275 280 285
Ser Thr Tyr Ile Gln Arg Arg Ser Lys Glu Thr Phe Val Asp Val Arg 290
295 300 Ile Val Glu Asp Asp
Val Asn Ile Lys Leu Thr Lys Arg Arg Arg Asp 305 310
315 320 Gly Cys Leu Ala Ala Ala Ser Arg Ala Leu
Asp Asp Leu Arg Leu Asp 325 330
335 Leu Val His Leu Ser Gly Gly Lys Ile Gly Asp Cys His Ile Tyr
Met 340 345 350 Phe
Asn Thr Lys Ile His Ser Gly Ser Pro Val Phe Ala Ser Ala Val 355
360 365 Ala Ser Arg Leu Ile Glu
Val Val Asp Glu Tyr 370 375
320DNAArtificial Sequenceprimer 3ggagcacgta catccagcgg
20424DNAArtificial Sequenceprimer
4actcatccac cacttcaatc agcc
24521DNAArtificial Sequenceprimer 5aactcatggc gatctcttac c
21620DNAArtificial Sequenceprimer
6gatgtaggag ggcgtggata
20720DNAArtificial Sequenceprimer 7cgtctgctgc tccatacaag
20820DNAArtificial Sequenceprimer
8ggagcacgta catccagcgg
20924DNAArtificial Sequenceprimer 9actcatccac cacttcaatc agcc
241021DNAArtificial Sequenceprimer
10tggagcagtt cgccagctac g
211123DNAArtificial Sequenceprimer 11cttctcctcc tccgacatct ccc
231220DNAArtificial Sequenceprimer
12cgctcgctcg tcccaaacat
201321DNAArtificial Sequenceprimer 13cggtcattgc tgggtccttg t
211420DNAArtificial Sequenceprimer
14tcctcctcgt cctgctcgtc
201520DNAArtificial Sequenceprimer 15ggttcacgat gtggcacagg
201621DNAArtificial Sequenceprimer
16ttaggccacg gaagtttgag g
211721DNAArtificial Sequenceprimer 17acacttcacc ggaccattca a
211823DNAArtificial Sequenceprimer
18gatcttctgg accaagaggg cag
231924DNAArtificial Sequenceprimer 19gtcaggagtg tctcagatgc ttgg
242021DNAArtificial Sequenceprimer
20ggaccacctg ctgctgcaac t
212120DNAArtificial Sequenceprimer 21gaacacttcg tgccatcgcc
202222DNAArtificial Sequenceprimer
22tggtggaaca gaagaggcat gg
222323DNAArtificial Sequenceprimer 23gcatgaagca gagagttggc ctt
232422DNAArtificial Sequenceprimer
24gcggaagtaa ggaaggagga gg
222524DNAArtificial Sequenceprimer 25ggcatcacaa tcttcacaga ggtg
242622DNAArtificial Sequenceprimer
26gagaacttcg agccgagggt ct
222720DNAArtificial Sequenceprimer 27ccagccgacg aggtttccac
202819DNAArtificial Sequenceprimer
28aggcgggcag cgtctccat
192923DNAArtificial Sequenceprimer 29ccataagcca gccacgatga tga
233024DNAArtificial Sequenceprimer
30catcctggtc cattcctcaa tgac
243123DNAArtificial Sequenceprimer 31ttcaggatga ggtgaagtgt ccc
233227DNAArtificial Sequenceprimer
32tatctagagt ggtagagtgc gaggaag
273330DNAArtificial Sequenceprimer 33gaggtaccag gtactcatcc accacttcaa
303425DNAArtificial Sequenceprimer
34catgttcaac accaagattc attcg
253525DNAArtificial Sequenceprimer 35tgcaaaccat gacataccaa agatc
253630DNAArtificial Sequenceprimer
36agggatccca ccacatggga agaggagacc
303729DNAArtificial Sequenceprimer 37atgtcgactc aaacgcgagg taatgcagg
293830DNAArtificial Sequenceprimer
38gtggatccat gccgcggcgc gcgagggcga
303933DNAArtificial Sequenceprimer 39tgctcgagat gcttggaacc tccacaatgc tgg
334029DNAArtificial Sequenceprimer
40agctgcagtt tgccaaaatg attgttggg
294129DNAArtificial Sequenceprimer 41gcgtcgactt gaatatgtcg agggcctgg
294226DNAArtificial Sequenceprimer
42gtggatcccg aggaagatgt atcacc
264330DNAArtificial Sequenceprimer 43agctcgagct agttagtact catccaccac
304426DNAArtificial Sequenceprimer
44gtggatcccg aggaagatgt atcacc
264530DNAArtificial Sequenceprimer 45agctcgagct agttagtact catccaccac
304628DNAArtificial Sequenceprimer
46cgctgcagtt agtactcatc caccactt
284729DNAArtificial Sequenceprimer 47aggaattctt tgccaaaatg attgttggg
294832DNAArtificial Sequenceprimer
48gcctgcagtt agttgaatat gtcgagggcc tg
324930DNAArtificial Sequenceprimer 49aagaattcat gggaagagga gaccacctgc
305031DNAArtificial Sequenceprimer
50ctggatcctc aatcaaacgc gaggtaatgc a
315132DNAArtificial Sequenceprimer 51tagaattcat gaagggtgag ttcggaaagg gc
325233DNAArtificial Sequenceprimer
52aggtcgactt accctctcct gcattcaagt aca
335330DNAArtificial Sequenceprimer 53acgaattcat gcatgtccac cataagccgc
305435DNAArtificial Sequenceprimer
54aaccgcggaa atgagttacc catacgatgt tcctg
355532DNAArtificial Sequenceprimer 55cagcggccgc agcgtaatct ggaacgtcat at
325631DNAArtificial Sequenceprimer
56aatctagaca tgggaagagg agaccacctg c
315731DNAArtificial Sequenceprimer 57cagtcgactc aatcaaacgc gaggtaatgc a
315826DNAArtificial Sequenceprimer
58cgtctagaga tgtatcaccc gcagtg
265930DNAArtificial Sequenceprimer 59agctcgagct agttagtact catccaccac
306030DNAArtificial Sequenceprimer
60agggatccca ccacatggga agaggagacc
306131DNAArtificial Sequenceprimer 61cagtcgactc aatcaaacgc gaggtaatgc a
316229DNAArtificial Sequenceprimer
62agctgcagtt tgccaaaatg attgttggg
296331DNAArtificial Sequenceprimer 63gcgtcgactt agttgaatat gtcgagggcc t
316431DNAArtificial Sequenceprimer
64ccactagttg ctttggtttg attcctggaa g
316529DNAArtificial Sequenceprimer 65aagtcgacaa cagtgctagg caccttcgc
296626DNAArtificial Sequenceprimer
66gtggatcccg aggaagatgt atcacc
266730DNAArtificial Sequenceprimer 67agctcgagct agttagtact catccaccac
306828DNAArtificial Sequenceprimer
68acactagtat ctacccgctt cgcgtcgg
286934DNAArtificial Sequenceprimer 69atctgcagcg agtgaagatc cctttcttgt
tacc 347039DNAArtificial Sequenceprimer
70gcctgcagca acaaacctag ttaatttagc tctagttgg
397128DNAArtificial Sequenceprimer 71gcgtcgacag gctctcaagc ggcatcag
287239DNAArtificial Sequenceprimer
72gcactagtca acaaacctag ttaatttagc tctagttgg
397330DNAArtificial Sequenceprimer 73tagcggccgc aggctctcaa gcggcatcag
307420DNAArtificial Sequenceprimer
74gatgtaggag ggcgtggata
207520DNAArtificial Sequenceprimer 75cgtctgctgc tccatacaag
207620DNAArtificial Sequenceprimer
76ggagcacgta catccagcgg
207721DNAArtificial Sequenceprimer 77atcgcaagac cggcaacagg a
2178366PRTBrachypodium distachyon 78Met
Tyr His Pro Gln Cys Glu Leu Leu Met Pro His Glu Ser Leu Asp 1
5 10 15 Met Asp Ala Val Val Gly
Gln Ser His Leu Ala Ala Ser Gly Val Ser 20
25 30 Ala Ile Pro Ala Glu Leu Asn Phe His Leu
Leu His His Ser Phe Val 35 40
45 Asp Thr Ala Ala Ser Pro Gln Pro Pro Thr Val Asp Tyr Phe
Phe Pro 50 55 60
Gly Thr Asp Pro Pro Pro Ala Ala Val Gln Phe Glu Gln Leu Ala Ala 65
70 75 80 Thr Asn His His Ala
Met Ser Met Leu Arg Asp Tyr Tyr Gly Gln Gln 85
90 95 Tyr Pro Ala Glu Thr Tyr Leu Arg Gly Gly
Pro Arg Thr Thr Thr Gly 100 105
110 Ser Ser Ser Leu Val Phe Gly Val Ala His Asp Asp Glu Ser Ala
Ala 115 120 125 Tyr
Asn Met Val Gly Pro Phe Val Glu Ser Ser Pro Thr Thr Arg Ala 130
135 140 Ala Gly Gly Gly Arg Lys
Arg Asn Arg Gly Ser Arg Ala Ala Gly Gly 145 150
155 160 Pro Ala His Gly Gly Val Glu Lys Lys Glu Lys
Gln Arg Arg Leu Arg 165 170
175 Leu Thr Glu Lys Tyr Thr Ala Leu Met Leu Leu Ile Pro Asn Arg Thr
180 185 190 Lys Glu
Asp Arg Ala Thr Val Ile Ser Asp Ala Ile Glu Tyr Ile Gln 195
200 205 Glu Leu Gly Arg Thr Val Glu
Glu Leu Thr Leu Leu Val Gly Lys Lys 210 215
220 Arg Arg Arg Asn Gly Ala Gly Glu His His Leu His
Gln Gly Asp Val 225 230 235
240 Val Asp Ala Ala Pro Ala Val Gly Ala Ala Gly Glu Leu Val Leu Ala
245 250 255 Ala Glu Ser
Ser Glu Gly Glu Val Gln Ala Pro Leu Ala Ala Leu Gln 260
265 270 Pro Ile Arg Ser Thr Tyr Ile Gln
Arg Lys Ser Lys Glu Thr Phe Val 275 280
285 Asp Val Arg Ile Val Glu Asp Glu Val Asn Ile Lys Leu
Thr Lys Arg 290 295 300
Arg Arg Asp Gly Cys Leu Ala Ala Ala Ser Arg Ala Leu Asp Asp Leu 305
310 315 320 Arg Leu Asp Leu
Val His Leu Ser Gly Gly Lys Ile Gly Asp Cys His 325
330 335 Ile Tyr Met Phe Asn Thr Lys Ile His
Gln Gly Ser Pro Val Phe Ala 340 345
350 Ser Ala Val Ala Ser Lys Leu Ile Glu Val Val Asp Glu Tyr
355 360 365 79350PRTTriticum
urartu 79Met Tyr His Gln Gln Cys Glu Leu Leu Met Pro His Glu Gly Leu Asp
1 5 10 15 Met Asp
Ala Gly Gln Ser His His Leu Ala Ala Gly Ser Ala Val Pro 20
25 30 Ala Glu Leu Asn Phe His Leu
Leu Ser Tyr Val Asp Thr Ala Val Ser 35 40
45 Pro Gln Gln Pro Thr Val Glu Tyr Phe Phe Gly Gly
Ala Asp Gln Pro 50 55 60
His Ala Gln Phe Glu Gln Leu Ala Ala Asn His Gln Ala Met Thr Val 65
70 75 80 Leu Arg Asp
Tyr Tyr Gly Gln Tyr His Pro Ala Thr Ala Asp Ala Tyr 85
90 95 Leu Pro Gly Gly Gly Pro Arg Thr
Gly Ser Ser Ser Leu Val Phe Gly 100 105
110 Ala Ala Glu Glu Glu Ser Ala Tyr Met Val Gly Gly Phe
Gln Cys Ser 115 120 125
Pro Lys Pro Arg Ala Ser Gly Ser Arg Lys Arg Gly Arg Gly Ala Gly 130
135 140 Ser Ser Phe His
Gly Phe Pro Ala Asn Gly Gly Val Glu Lys Lys Glu 145 150
155 160 Lys Gln Arg Arg Gln Arg Leu Ser Glu
Lys Phe Thr Ala Leu Met Leu 165 170
175 Leu Ile Pro Asn Arg Thr Lys Glu Asp Arg Ala Thr Val Ile
Tyr Asp 180 185 190
Ala Ile Glu Tyr Ile Gln Glu Leu Gly Arg Thr Val Glu Glu Leu Thr
195 200 205 Leu Leu Val Glu
Lys Lys Arg Gly Arg Arg Glu His Gln Gly Asp Val 210
215 220 Val Asp Pro Ala Pro Thr Leu Val
Ala Gly Asp Gly Glu Cys Ser Ala 225 230
235 240 Gly Glu Val Ala Ala Ala Val Met Pro Ala Met Pro
Ala Pro Pro Gln 245 250
255 Pro Ile Arg Ser Thr Tyr Ile Gln Arg Arg Ser Lys Glu Thr Phe Val
260 265 270 Asp Val Arg
Ile Val Glu Asp Glu Val Asn Ile Lys Leu Thr Lys Arg 275
280 285 Arg Arg Asp Gly Cys Leu Ala Ala
Ala Ser Arg Ala Leu Asp Asp Leu 290 295
300 His Leu Asp Leu Val His Leu Ser Gly Gly Lys Ile Gly
Asp Cys His 305 310 315
320 Ile Tyr Met Phe Asn Thr Lys Ile His Pro Gly Ser Pro Val Phe Ala
325 330 335 Ser Ala Val Ala
Ser Lys Leu Ile Glu Val Val Asp Glu Tyr 340
345 350 80379PRTSorghum bicolor 80Met Tyr His Pro Gln Cys
Glu Leu Leu Met Ala His Glu Ala Gln Asp 1 5
10 15 Leu Asp Ala Ala Gly Gln Pro His His Leu Ala
Val Ser Gly Val Ala 20 25
30 Gly Ser Ile Pro Ala Glu Leu Ser Phe His Leu Leu His Ser Leu
Asp 35 40 45 Ala
Thr Ala Ala Val Asn Asn Ser Val Thr Pro Gln Ser Thr Ile Asp 50
55 60 Tyr Phe Leu Gly Val Gly
Gly Ala Asp Pro His Gln Pro Ala Ala Leu 65 70
75 80 Gln Tyr Glu Pro Leu Pro Pro Pro Gly Gly His
His Gln His Thr Met 85 90
95 Asn Met Leu Arg Asp Tyr Cys Ser Asn Gly Gly Gly Gly Gly His Tyr
100 105 110 Pro Thr
Ala Glu Pro Tyr Leu Arg Gly Thr Arg Thr Gly Ala Leu Val 115
120 125 Phe Gly Ala Thr Asp Asp Asp
Glu Ser Ala Ala Ala Tyr Met Pro Gly 130 135
140 Gly Pro Phe Val Glu Thr Ser Pro Pro Pro Arg Ala
Thr Gly Gly Arg 145 150 155
160 Lys Arg Gly Arg Ala Leu Gly Gly Gly Phe His Ala Gly Leu Ala Asn
165 170 175 Gly Val Glu
Lys Lys Glu Lys Gln Arg Arg Gln Arg Leu Thr Glu Lys 180
185 190 Tyr Thr Ala Leu Met His Leu Ile
Pro Asn Val Thr Lys Pro Asp Arg 195 200
205 Ala Thr Val Ile Ser Asp Ala Ile Glu Tyr Ile Gln Glu
Leu Gly Arg 210 215 220
Thr Val Glu Glu Leu Thr Leu Leu Val Glu Lys Lys Arg Arg Arg Arg 225
230 235 240 Glu Leu Gln Gly
Asp Val Val Asp Ala Ala Pro Thr Ala Val Val Val 245
250 255 Ala Ala Ala Ala Thr Gly Gly Glu Ala
Glu Ser Ser Glu Gly Glu Val 260 265
270 Ala Pro Pro Pro Pro Pro Pro Ala Ala Val Gln Arg Gln Pro
Ile Arg 275 280 285
Ser Thr Tyr Ile Gln Arg Arg Ser Lys Asp Thr Ser Val Asp Val Arg 290
295 300 Ile Val Glu Glu Asp
Val Asn Ile Lys Leu Thr Lys Arg Arg Arg Asp 305 310
315 320 Gly Cys Leu Ala Ala Ala Ser Arg Ala Leu
Asp Asp Leu Arg Leu Asp 325 330
335 Leu Val His Leu Ser Gly Gly Lys Ile Gly Asp Cys His Ile Tyr
Met 340 345 350 Phe
Asn Thr Lys Ile His Lys Gly Ser Ser Val Phe Ala Ser Ala Val 355
360 365 Ala Ser Arg Leu Met Glu
Val Val Asp Glu Tyr 370 375
81365PRTZea mays 81Met Tyr His Pro Gln Cys Glu Leu Leu Thr Met Ala His
Glu Thr Pro 1 5 10 15
Asp Leu Asp Ala Gly Gln Pro His Leu Thr Val Ser Gly Val Ala Ser
20 25 30 Ile Pro Ala Glu
Leu Ser Phe His Leu Leu His Ser Leu Asp Ala Ala 35
40 45 Ala Ala Val Asn Pro Val Thr Ala Pro
Pro Gln Ser Thr Ile Asp Tyr 50 55
60 Phe Leu Gly Gly Ala Asp Pro His Gln Gln Ala Met Gln
Tyr Glu Pro 65 70 75
80 Leu Pro Pro Ala Ala Gly Gly His His Gln Tyr Thr Met Asp Met Phe
85 90 95 Arg Asp Tyr Cys
Asp Gly His Tyr Pro Thr Ala Glu Pro Tyr Ile Arg 100
105 110 Gly Thr Met Thr Gly Ala Leu Val Phe
Gly Ala Thr Asp Asp Asp Asp 115 120
125 Ser Ala Ala Ala Tyr Met Pro Gly Gly His Phe Glu Thr Ser
Pro Pro 130 135 140
Pro Pro Arg Ala Thr Gly Arg Gly Arg Lys Arg Gly Arg Ala Leu Gly 145
150 155 160 Gly Gly Phe His Ala
Val Leu Ala Asn Gly Val Glu Lys Lys Glu Lys 165
170 175 Gln Arg Arg Leu Arg Leu Thr Glu Lys Tyr
Thr Ala Leu Met His Leu 180 185
190 Ile Pro Asn Val Thr Lys Thr Asp Arg Ala Thr Val Ile Ser Asp
Ala 195 200 205 Ile
Glu Tyr Ile Gln Glu Leu Gly Arg Thr Val Glu Glu Leu Thr Leu 210
215 220 Leu Val Glu Lys Lys Arg
Arg Arg Arg Glu Leu Gln Gly Asp Val Val 225 230
235 240 Asp Ala Ala Pro Ala Ala Val Val Ala Ala Ala
Gly Glu Ala Glu Ser 245 250
255 Ser Glu Gly Glu Val Ala Pro Pro Pro Pro Ala Val Pro Arg Gln Pro
260 265 270 Ile Arg
Ser Thr Tyr Ile Gln Arg Arg Ser Lys Asp Thr Ser Val Asp 275
280 285 Val Arg Ile Val Glu Glu Asp
Val Asn Ile Lys Leu Thr Lys Arg Arg 290 295
300 Arg Asp Gly Cys Leu Ala Ala Ala Ser Arg Ala Leu
Asp Asp Leu Arg 305 310 315
320 Leu Asp Leu Val His Leu Ser Gly Gly Lys Ile Gly Asp Cys Gln Ile
325 330 335 Tyr Met Phe
Asn Thr Lys Ile His Lys Gly Ser Ser Val Phe Ala Ser 340
345 350 Ala Val Ala Gly Arg Leu Met Glu
Val Val Asp Glu Tyr 355 360 365
82268PRTAegilops tauschii 82Met Tyr His Gln Gln Cys Glu Leu Leu Met Pro
His Glu Asp Leu Asp 1 5 10
15 Met Asp Ala Gly Gln Ser His His Leu Ala Ala Ala Ser Ala Val Pro
20 25 30 Ala Glu
Leu Asn Phe His Leu Leu Ser Tyr Val Asp Ala Ala Val Ser 35
40 45 Pro Gln Gln Pro Thr Val Glu
Tyr Phe Phe Gly Gly Ala Asp Gln Pro 50 55
60 His Ala His Ser Phe His Gly Phe Pro Ala Asn Gly
Gly Val Glu Lys 65 70 75
80 Lys Glu Lys Gln Arg Arg Gln Arg Leu Ser Glu Lys Phe Thr Ala Leu
85 90 95 Met Leu Leu
Ile Pro Asn Arg Thr Lys Glu Asp Arg Ala Thr Val Ile 100
105 110 Tyr Asp Ala Ile Glu Tyr Ile Gln
Glu Leu Gly Arg Thr Val Glu Glu 115 120
125 Leu Thr Leu Leu Val Glu Lys Lys Arg Gly Arg Arg Glu
His Gln Gly 130 135 140
Asp Val Val Asp Pro Ala Pro Leu Val Val Ala Gly Glu Gly Glu Cys 145
150 155 160 Ser Ala Gly Glu
Leu Arg Ala Pro Pro Pro Pro Pro Pro Gln Pro Ile 165
170 175 Arg Ser Thr Tyr Ile Gln Arg Arg Ser
Lys Glu Thr Phe Val Asp Val 180 185
190 Arg Ile Val Glu Asp Glu Val Asn Ile Lys Leu Thr Lys Arg
Arg Arg 195 200 205
Asp Gly Cys Leu Ala Ala Ala Ser Arg Ala Leu Asp Asp Leu His Leu 210
215 220 Asp Leu Val His Leu
Ser Gly Gly Lys Ile Gly Asp Cys His Ile Tyr 225 230
235 240 Met Phe Asn Thr Lys Ile His Pro Gly Ser
Pro Val Phe Ala Ser Ala 245 250
255 Val Ala Ser Lys Leu Ile Glu Val Val Asp Glu Tyr
260 265 83170PRTZea mays 83Met Val Phe Ser
Lys Ile Gln Tyr Lys Val Val Ser Ser Lys Ile Gln 1 5
10 15 Thr Pro Thr Leu Val Arg Val Glu Thr
Thr His Glu Thr Asn Met Glu 20 25
30 Lys Lys Arg Arg Arg Arg Glu Leu Gln Gly Asp Val Val Asp
Ala Ala 35 40 45
Pro Ala Ala Val Val Ala Ala Ala Gly Glu Ala Glu Ser Ser Glu Gly 50
55 60 Glu Val Ala Pro Pro
Pro Pro Ala Val Pro Arg Gln Pro Ile Arg Ser 65 70
75 80 Thr Tyr Ile Gln Arg Arg Ser Lys Asp Thr
Ser Val Asp Val Arg Ile 85 90
95 Val Glu Glu Asp Val Asn Ile Lys Leu Thr Lys Arg Arg Arg Asp
Gly 100 105 110 Cys
Leu Ala Ala Ala Ser Arg Ala Leu Asp Asp Leu Arg Leu Asp Leu 115
120 125 Val His Leu Ser Gly Gly
Lys Ile Gly Asp Cys Gln Ile Tyr Met Phe 130 135
140 Asn Thr Lys Ile His Lys Gly Ser Ser Val Phe
Ala Ser Ala Val Ala 145 150 155
160 Gly Arg Leu Met Glu Val Val Asp Glu Tyr 165
170 84433PRTVitis vinifera 84Met Cys Arg Lys Ile Thr Arg Pro
Arg Arg Met Tyr Val Tyr Glu Glu 1 5 10
15 Asn Ala Cys Phe Asp Gly Thr Lys Ser Val Ala Glu Gly
Asp Asp Glu 20 25 30
Gly Phe Ser Gln Ser Val Ala Pro Pro Pro Thr Asn Asn Ser Phe Glu
35 40 45 Asp Ser Thr Asn
Met Arg Val Ser Met Glu Asp Ala Ser Ala Thr Met 50
55 60 Glu Ile Glu Leu His Gln Gln Leu
Ala Phe Asp Met Asp Gln Gln Cys 65 70
75 80 Tyr Asn Ser Asn Asn Asp Gly Asn Asp Ser Asn Gln
Val Phe Ser Tyr 85 90
95 Glu Met Gln Glu Met Gly Phe Asn His His Gln Gln Gln Gln Glu Asp
100 105 110 Pro Leu Leu
Leu Gln Gln His Gln Ala Glu Met Gln Asn Ala His Gln 115
120 125 Asn Phe Ser Ala Ala Tyr Pro Pro
Thr Pro Asp Leu Leu Asn Leu Phe 130 135
140 His Leu Pro Arg Cys Thr Pro Ser Ser Leu Leu Pro Asn
Ser Ser Ile 145 150 155
160 Ser Phe Thr Asn Pro Asp Ser Ser Ala Thr Ala Ala Ser Gly Ile Leu
165 170 175 Tyr Asp Pro Leu
Phe His Leu Asn Leu Pro Pro Gln Pro Pro Val Gly 180
185 190 Gly Gly Tyr Gly Asp Gly Asp Asp His
Arg Gln Phe Asp Asn Gly Val 195 200
205 Leu Lys Phe Thr Arg Asp Met Ala Cys Ile Gly Lys Gly Arg
Glu Gly 210 215 220
Lys Gly Thr Lys Ser Phe Ala Thr Glu Lys Gln Arg Arg Glu His Leu 225
230 235 240 Asn Asp Lys Tyr Asn
Ala Leu Arg Ser Leu Val Pro Asn Pro Thr Lys 245
250 255 Ser Asp Arg Ala Ser Val Val Gly Asp Ala
Ile Glu Tyr Ile Arg Glu 260 265
270 Leu Leu Arg Thr Val Asn Glu Leu Lys Leu Leu Val Glu Lys Lys
Arg 275 280 285 Cys
Gly Arg Glu Arg Ser Lys Arg His Lys Thr Glu Asp Glu Ser Thr 290
295 300 Gly Asp Val Lys Ser Ser
Ser Ser Ile Lys Pro Glu Pro Asp Gln Ser 305 310
315 320 Tyr Asn Glu Ser Leu Arg Ser Ser Trp Leu Gln
Arg Lys Ser Lys Asp 325 330
335 Thr Glu Val Asp Val Arg Ile Ile Asp Asp Glu Val Thr Ile Lys Leu
340 345 350 Val Gln
Arg Lys Lys Ile Asn Cys Leu Leu Phe Val Ser Lys Ile Leu 355
360 365 Asp Glu Leu Gln Leu Asp Leu
His His Val Ala Gly Gly His Val Gly 370 375
380 Asp Tyr Tyr Ser Phe Leu Phe Asn Thr Lys Ile Tyr
Glu Gly Ser Ser 385 390 395
400 Val Tyr Ala Ser Ala Ile Ala Asn Lys Leu Ile Glu Val Val Asp Arg
405 410 415 Gln Tyr Ala
Ala Ile Pro Ile Pro Ile Pro Ile Pro Pro Thr Ser Ser 420
425 430 Phe 85456PRTVitis vinifera
85Met Tyr Met Tyr Glu Glu Asn Ala Cys Phe Asp Gly Thr Lys Ser Val 1
5 10 15 Ala Glu Gly Asp
Asp Glu Gly Phe Ser Gln Ser Val Ala Pro Pro Pro 20
25 30 Thr Asn Asn Ser Phe Glu Asp Ser Thr
Asn Met Arg Val Ser Met Glu 35 40
45 Asp Ala Ser Ala Thr Met Glu Ile Glu Leu His Gln Gln Leu
Ala Phe 50 55 60
Asp Met Asp Gln Gln Cys Tyr Asn Ser Asn Asn Asp Gly Asn Asp Ser 65
70 75 80 Asn Gln Val Phe Ser
Tyr Glu Met Gln Glu Met Gly Phe Asn His His 85
90 95 His Gln Gln Gln Asp Asp Pro Leu Leu Leu
Gln Gln His Gln Ala Glu 100 105
110 Met Gln Asn Ala Gln Gln Asn Phe Ser Ala Ala Tyr Pro Pro Thr
Pro 115 120 125 Asp
Leu Leu Asn Leu Phe His Leu Pro Arg Cys Thr Pro Ser Ser Leu 130
135 140 Leu Pro Asn Ser Ser Ile
Ser Phe Thr Asn Pro Asp Ser Ser Ala Thr 145 150
155 160 Ala Ala Ser Gly Ile Leu Tyr Asp Pro Leu Phe
His Leu Asn Leu Pro 165 170
175 Pro Gln Pro Pro Val Phe Arg Glu Leu Phe Gln Ser Leu Pro His Gly
180 185 190 Tyr Asn
Leu Pro Ala Ser Arg Val Gly Ser Leu Phe Gly Gly Gly Met 195
200 205 Asp Glu Arg Glu Ala Ser Gly
Gly Gly Tyr Gly Asp Gly Asp Asp His 210 215
220 Arg Gln Phe Asp Asn Gly Val Leu Lys Phe Thr Arg
Asp Met Ala Cys 225 230 235
240 Ile Gly Arg Gly Arg Glu Gly Lys Gly Thr Lys Ser Phe Ala Thr Glu
245 250 255 Lys Gln Arg
Arg Glu His Leu Asn Asp Lys Tyr Asn Ala Leu Arg Ser 260
265 270 Leu Val Pro Asn Pro Thr Lys Ser
Asp Arg Ala Ser Val Val Gly Asp 275 280
285 Ala Ile Glu Tyr Ile Arg Glu Leu Leu Arg Thr Val Asn
Glu Leu Lys 290 295 300
Leu Leu Val Glu Lys Lys Arg Cys Gly Arg Glu Arg Ser Lys Arg His 305
310 315 320 Lys Thr Glu Asp
Glu Ser Thr Gly Asp Val Lys Ser Ser Ser Ser Ile 325
330 335 Lys Pro Glu Pro Asp Gln Ser Tyr Asn
Glu Ser Leu Arg Ser Ser Trp 340 345
350 Leu Gln Arg Lys Ser Lys Asp Thr Glu Val Asp Val Arg Ile
Ile Asp 355 360 365
Asp Glu Val Thr Ile Lys Leu Val Gln Arg Lys Lys Ile Asn Cys Leu 370
375 380 Leu Phe Val Ser Lys
Ile Leu Asp Glu Leu Gln Leu Asp Leu His His 385 390
395 400 Val Ala Gly Gly His Val Gly Asp Tyr Tyr
Ser Phe Leu Phe Asn Thr 405 410
415 Lys Ile Tyr Glu Gly Ser Ser Val Tyr Ala Ser Ala Ile Ala Asn
Lys 420 425 430 Leu
Ile Glu Val Val Asp Arg Gln Tyr Ala Ala Ile Pro Ile Pro Ile 435
440 445 Pro Ile Pro Pro Thr Ser
Ser Phe 450 455 86568PRTVitis vinifera 86Met Gly
Arg Gly Phe Ser His Leu Leu Gly Phe Cys Lys Ser Glu Asn 1 5
10 15 Pro Leu Asn Asp Leu Ser Arg
Leu Lys Ser Arg Tyr Ser Gly Pro Asn 20 25
30 Leu Arg Leu Val His Ser Glu Ser Asn Arg Asn Asn
Ile Glu Asn Met 35 40 45
Val Asp Ala Phe Pro Val Gln Ile Asp Leu Asn Phe Ser Asp Ser Arg
50 55 60 Asn Asp Lys
Ala Thr Gln Thr Ile Ser His Pro Arg Val Lys His Ser 65
70 75 80 His Gln Gly Ala Arg Arg Arg
Ile Met Arg Ser Thr Asn His Ala Glu 85
90 95 Gln Cys Gln Asn Lys Arg Gly Tyr Arg Lys Ile
Thr Arg Pro Arg Arg 100 105
110 Met Tyr Val Tyr Glu Glu Asn Ala Cys Phe Asp Gly Thr Lys Ser
Val 115 120 125 Ala
Glu Gly Asp Asp Glu Gly Phe Ser Gln Ser Val Ala Pro Pro Pro 130
135 140 Thr Asn Asn Ser Phe Glu
Asp Ser Thr Asn Met Arg Val Ser Met Glu 145 150
155 160 Asp Ala Ser Ala Thr Met Glu Ile Glu Leu His
Gln Gln Leu Ala Phe 165 170
175 Asp Met Asp Gln Gln Cys Tyr Asn Ser Asn Asn Asp Gly Asn Asp Ser
180 185 190 Asn Gln
Val Phe Ser Tyr Glu Met Gln Glu Met Gly Phe Asn His His 195
200 205 Gln Gln Gln Gln Glu Asp Pro
Leu Leu Leu Gln Gln His Gln Ala Glu 210 215
220 Met Gln Asn Ala His Gln Asn Phe Ser Ala Ala Tyr
Pro Pro Thr Pro 225 230 235
240 Asp Leu Leu Asn Leu Phe His Leu Pro Arg Cys Thr Pro Ser Ser Leu
245 250 255 Leu Pro Asn
Ser Ser Ile Ser Phe Thr Asn Pro Asp Ser Ser Ala Thr 260
265 270 Ala Ala Ser Gly Ile Leu Tyr Asp
Pro Leu Phe His Leu Asn Leu Pro 275 280
285 Pro Gln Pro Pro Val Phe Arg Glu Leu Phe Gln Ser Leu
Pro His Gly 290 295 300
Tyr Asn Leu Pro Ala Ser Arg Val Gly Ser Leu Phe Gly Gly Gly Met 305
310 315 320 Asp Glu Arg Glu
Ala Ser Gly Gly Gly Tyr Gly Asp Gly Asp Asp His 325
330 335 Arg Gln Phe Asp Asn Gly Val Leu Lys
Phe Thr Arg Asp Met Ala Cys 340 345
350 Ile Gly Lys Gly Arg Glu Gly Lys Gly Thr Lys Ser Phe Ala
Thr Glu 355 360 365
Lys Gln Arg Arg Glu His Leu Asn Asp Lys Tyr Asn Ala Leu Arg Ser 370
375 380 Leu Val Pro Asn Pro
Thr Lys Ser Asp Arg Ala Ser Val Val Gly Asp 385 390
395 400 Ala Ile Glu Tyr Ile Arg Glu Leu Leu Arg
Thr Val Asn Glu Leu Lys 405 410
415 Leu Leu Val Glu Lys Lys Arg Cys Gly Arg Glu Arg Ser Lys Arg
His 420 425 430 Lys
Thr Glu Asp Glu Ser Thr Gly Asp Val Lys Ser Ser Ser Ser Ile 435
440 445 Lys Pro Glu Pro Asp Gln
Ser Tyr Asn Glu Ser Leu Arg Ser Ser Trp 450 455
460 Leu Gln Arg Lys Ser Lys Asp Thr Glu Val Asp
Val Arg Ile Ile Asp 465 470 475
480 Asp Glu Val Thr Ile Lys Leu Val Gln Arg Lys Lys Ile Asn Cys Leu
485 490 495 Leu Phe
Val Ser Lys Ile Leu Asp Glu Leu Gln Leu Asp Leu His His 500
505 510 Val Ala Gly Gly His Val Gly
Asp Tyr Tyr Ser Phe Leu Phe Asn Thr 515 520
525 Lys Ile Tyr Glu Gly Ser Ser Val Tyr Ala Ser Ala
Ile Ala Asn Lys 530 535 540
Leu Ile Glu Val Val Asp Arg Gln Tyr Ala Ala Ile Pro Ile Pro Ile 545
550 555 560 Pro Ile Pro
Pro Thr Ser Ser Phe 565 87469PRTRicinus
communis 87Met Tyr Glu Glu Thr Gly Cys Phe Asp Pro Asn Ser Met Val Glu
Gly 1 5 10 15 Ala
Asp Asp Gly Leu Cys Gln Val Leu Gln Ile Pro Pro Gln Pro Gln
20 25 30 Pro Leu Met Ala Gly
Ser Thr Thr Asn Ser His Asn Ser Tyr Glu Glu 35
40 45 Asn Leu Lys Leu Ser Ala Asp Gln Glu
Leu Ser Tyr His His Ser Asn 50 55
60 Asn Pro His His His His Gln Glu Asp Asp Ala Ser Ala
Ser Ala Ala 65 70 75
80 Ala Ala Met Glu Thr Gln Leu Gln Asn His Gln Met Gly Phe Asp Thr
85 90 95 His Leu Met Gln
Asp Ser Ser Asn Gln Val Met Ala Phe Asn Ser Ser 100
105 110 Thr Ser Leu Gln Asp Ala Thr Phe Ala
Gln Thr Pro Asp Leu Leu Asn 115 120
125 Leu Phe His Leu Pro Arg Gly Ser Thr Ser Ser Leu Leu Pro
Asn Ser 130 135 140
Ser Ile Ser Phe Thr Asn Pro Ser His Thr Ala Pro Leu Gly Phe Val 145
150 155 160 Gly Asp Leu Pro Met
Ala Asp Thr Ala Ser Ala Ser Ser Ile Leu Tyr 165
170 175 Asp Pro Leu Phe His Leu Asn Leu Pro Pro
Gln Pro Pro Leu Phe Arg 180 185
190 Asp Leu Phe Gln Ser Leu Pro Pro His Gly Tyr Ser Leu Pro Gly
Ser 195 200 205 Met
Val Asn Ser Leu Phe Gly Ala Gly Val Gly Gly Asp Asp His Val 210
215 220 Glu Gly Ser Gly Asp Gly
Gly Gly Ile Tyr Gln Asp Gly Asp Gly Glu 225 230
235 240 Gln Gln Phe Asp Asn Gly Val Leu Asp Phe Thr
Trp Asp Met Pro Cys 245 250
255 Met Gly Lys Gly Arg Asp Ala Gly Lys Lys Thr Lys Pro Phe Ala Thr
260 265 270 Glu Arg
Gln Arg Arg Gln His Leu Asn Asp Lys Tyr Lys Ala Leu Gln 275
280 285 Asn Leu Val Pro Asn Pro Thr
Lys Ala Asp Arg Thr Ser Val Val Gly 290 295
300 Asp Ala Ile Asp Tyr Ile Lys Glu Leu Leu Arg Thr
Val Asn Glu Leu 305 310 315
320 Lys Leu Leu Val Glu Lys Lys Arg Cys Ala Arg Glu Arg Ser Lys Arg
325 330 335 Gln Lys Thr
Glu Glu Asp Ser Ile Gly Asn Gly His Asp Ser Ser Cys 340
345 350 Ile Thr Lys Pro Leu Gly Asp Pro
Asp Gln Ser Phe Asn Asn Gly Ser 355 360
365 Leu Arg Ser Ser Trp Ile Glu Arg Lys Ser Lys Asp Thr
Glu Val Asp 370 375 380
Val Arg Ile Ile Asp Asp Glu Val Thr Ile Lys Leu Val Gln Arg Lys 385
390 395 400 Lys Ile Asn Cys
Leu Leu Phe Val Ser Lys Val Leu Asp Glu Leu Gln 405
410 415 Leu Asp Leu His His Val Ala Gly Gly
His Ile Gly Asp Tyr Tyr Ser 420 425
430 Phe Leu Phe Asn Thr Lys Ile Phe Glu Gly Ser Ser Val Tyr
Ala Ser 435 440 445
Ala Ile Ala Asn Lys Leu Ile Glu Val Val Asp Arg His Tyr Ala Ser 450
455 460 Thr Pro Ser Thr Asn
465 88467PRTPopulus trichocarpa 88Met Tyr Val Glu Thr Ala
Cys Phe Glu Pro Asn Asn Ser Met Val Glu 1 5
10 15 Asp Val Thr Asp Asp Gly Phe Cys His Ala Ile
Pro Leu Met Ala Gly 20 25
30 Asn Ser Thr Thr Asn Ser Phe Glu Glu His Leu Lys Leu Ser Met
Glu 35 40 45 Glu
Phe Ser Ser His Tyr Pro Gln Glu Glu Ser Ala Ala Ala Ala Ser 50
55 60 Met Glu Glu Ile Gln Leu
Gln His His Met Ala Phe Ser Asn Asn Asn 65 70
75 80 Thr Asn His His Leu Met Gln Gln Tyr Pro Thr
Gln Leu Leu Ser Tyr 85 90
95 Asp His Ser Ser Asn Trp Asp Pro Asn Ile Ile Gln Phe Gln Glu Met
100 105 110 His Gln
Val Leu Asp Gln Asn Ser Ser Phe Asp Ala Thr Ala Asn Thr 115
120 125 Gln Ser Ser Leu Pro Pro Asp
Leu Leu Asn Leu Phe Asn Leu Pro Arg 130 135
140 Cys Thr Ser Thr Ser Thr Leu Leu Pro Asn Ser Ser
Ile Ser Phe Thr 145 150 155
160 Asn Pro Ala His Lys Ala Pro Leu Gly Phe Met Gly Val Asp Asn Thr
165 170 175 Ser Ala Arg
Phe Asp Pro Tyr Thr Leu Ala Pro Gln Pro His Leu Phe 180
185 190 Arg Glu Leu Val Gln Ser Leu Pro
Pro His Gly Tyr Thr Leu Pro Thr 195 200
205 Pro Leu Phe Gly Gly Gly Gln Gly Asp Asp His Val Asp
Gly Gln Ser 210 215 220
Gly Gly Gly Leu Ser Tyr Gln Asp Gly Asp His Gly Asp Gly Val Phe 225
230 235 240 Glu Phe Thr Asp
Glu Met Ala Cys Ile Gly Lys Gly Ile Lys Lys Thr 245
250 255 Gly Lys Val Thr Lys His Phe Ala Thr
Glu Arg Gln Arg Arg Glu His 260 265
270 Leu Asn Gly Lys Tyr Thr Ala Leu Arg Asn Leu Val Pro Asn
Pro Ser 275 280 285
Lys Asn Asp Arg Ala Ser Val Val Gly Glu Ala Ile Asp Tyr Ile Lys 290
295 300 Glu Leu Leu Arg Thr
Val Gln Glu Leu Lys Leu Leu Val Glu Lys Lys 305 310
315 320 Arg Cys Gly Arg Glu Arg Ser Lys Trp Arg
Lys Thr Glu Asp Asp Gly 325 330
335 Gly Val Glu Val Leu Asp Asn Ser Asp Ile Lys Val Glu Pro Asp
Gln 340 345 350 Ser
Ala Tyr Ser Asn Gly Ser Leu Arg Ser Ser Trp Leu Gln Arg Lys 355
360 365 Ser Lys Asp Thr Glu Val
Asp Val Arg Leu Ile Glu Asp Glu Val Thr 370 375
380 Ile Lys Leu Val Gln Arg Lys Arg Val Asn Cys
Leu Leu Tyr Val Ser 385 390 395
400 Lys Val Leu Asp Glu Leu Gln Leu Asp Leu His His Ala Ala Gly Gly
405 410 415 Leu Ile
Gly Asp Tyr Tyr Ser Phe Leu Phe Asn Thr Lys Ile Asn Glu 420
425 430 Gly Ser Cys Val Tyr Ala Ser
Ala Ile Ala Asn Arg Leu Ile Glu Val 435 440
445 Val Asp Arg Gln Tyr Ala Ser Ser Thr Thr Thr Val
Pro Ala Ala Gly 450 455 460
Ser Cys Tyr 465 89473PRTZea maysmisc_feature(284)..(284)Xaa
can be any naturally occurring amino acid 89Met Ile Ala Gly Gly Gly Tyr
Phe Asp Gly Ser His Asp His Ile Leu 1 5
10 15 Met Glu Gly Ser Met Ile His Asp Ser Ser Gln
Ser Ser Ile Tyr Asp 20 25
30 Asn Thr Asp Val Glu Gln Gln Asn Phe Arg Phe Ala Pro Phe Ile
Ile 35 40 45 Glu
Asp His Ser Asn Pro Ala Asn Leu Thr Ser Glu Ala Ala Arg Val 50
55 60 Ile Asp Gln Ile Gln His
Gln Leu Gly Ile Asp Ile Glu Gln Asp His 65 70
75 80 Ser Asp His Met Met Gln Glu Val Pro Pro Ala
Glu Thr Glu Asn Leu 85 90
95 Val Pro Ala Val Tyr Gly Val Gln Asp His Ile Leu Ser His Gln Ile
100 105 110 Glu Gly
Pro His Asn Ile Thr Val Glu Gln Gln Val Leu Gly Tyr Asp 115
120 125 Pro Ala Ser Tyr Arg Asn Gly
Thr Tyr Ala Ala Ala His Asp Leu Leu 130 135
140 Asn Ser Leu His Ile Gln Arg Cys Ser Leu Ile Pro
Glu Phe Pro Ser 145 150 155
160 Thr Glu His Ile Phe Ser Asp Pro Ala Gln Asn Met Val Asn Arg Leu
165 170 175 Asp Ile Thr
Asn Asp Leu Pro Gly Val Ala Asn His Glu Ser Gly Met 180
185 190 Met Phe Ser Asp Ser Thr Val Pro
Leu Gly Tyr His Ala Thr Gln Ser 195 200
205 His Met Leu Lys Asp Leu Tyr His Ser Leu Pro Gln Asn
Tyr Gly Leu 210 215 220
Phe Thr Ser Asp Asp Glu Arg Asp Gly Met Val Gly Val Pro Gly Val 225
230 235 240 Ser Gly Asn Ile
Phe Gln Glu Ile Asp Gly Arg Gln Phe Asp Ser Pro 245
250 255 Ile Leu Gly Ser Arg Lys Gln Lys Gly
Gly Phe Gly Lys Gly Lys Gly 260 265
270 Lys Ala Asn Phe Ala Thr Glu Arg Glu Arg Arg Xaa Gln Phe
Asn Val 275 280 285
Lys Tyr Gly Ala Leu Arg Ser Leu Phe Pro Asn Pro Thr Lys Asn Asp 290
295 300 Arg Ala Ser Ile Val
Gly Asp Ala Ile Glu Tyr Ile Asn Glu Leu Asn 305 310
315 320 Arg Thr Val Lys Glu Leu Lys Ile Leu Leu
Glu Lys Lys Arg Asn Ser 325 330
335 Ala Asp Arg Arg Lys Ile Leu Lys Leu Asp Glu Glu Ala Ala Asp
Asp 340 345 350 Gly
Glu Ser Ser Ser Met Gln Pro Val Ser Asp Asp Gln Xaa Asn Gln 355
360 365 Met Asn Gly Thr Ile Arg
Ser Ser Trp Val Gln Arg Arg Ser Lys Glu 370 375
380 Cys Asp Val Asp Val Arg Ile Val Asp Asp Glu
Ile Asn Ile Lys Phe 385 390 395
400 Thr Glu Lys Lys Arg Ala Asn Ser Leu Leu Cys Ala Ala Lys Val Leu
405 410 415 Glu Glu
Phe His Leu Glu Leu Ile His Val Val Gly Gly Ile Ile Gly 420
425 430 Asp His His Ile Phe Met Phe
Asn Thr Lys Ile Pro Lys Gly Ser Ser 435 440
445 Val Tyr Ala Cys Ala Val Ala Lys Lys Leu Leu Glu
Ala Val Glu Ile 450 455 460
Lys Lys Gln Ala Tyr Asn Ile Phe Asn 465 470
90463PRTPopulus trichocarpa 90Met Tyr Glu Glu Thr Ala Cys Phe Glu Thr
Asn Asn Ser Ile Val Glu 1 5 10
15 Gly Gly Asn Asp Asp Gly Phe Cys Gln Val Ser Pro Phe Met Thr
Gly 20 25 30 Ser
Ser Thr Thr Ser Ser Phe Glu Glu Ser Phe Lys Leu Ser Met Glu 35
40 45 Glu Leu Ser Asn His Tyr
His Gln Glu Glu Ser Ala Ala Ala Ala Ser 50 55
60 Met Glu Glu Ile Gln Leu Gln His His Met Ala
Phe Asn Asn Asn Cys 65 70 75
80 His His Leu Met Glu Gln Tyr Pro Thr Asn His His Gln Val Leu Ser
85 90 95 Tyr Asp
His Pro Ser Asn Trp Asp Pro Asn Thr Ile Gln Phe Gln Glu 100
105 110 Met His Gln Val Leu Asp Gln
Asn Gly Asn Phe Asn Ala Thr Ala Asn 115 120
125 Thr Pro Ser Ser Leu Leu Pro Asp Leu Leu Asn Leu
Phe Asn Leu Pro 130 135 140
Arg Cys Thr Ser Thr Ser Thr Leu Leu Pro Asn Ser Ser Ile Ser Phe 145
150 155 160 Thr Asn Pro
Ala His Lys Thr Pro Ser Gly Phe Met Gly Val Asp Ser 165
170 175 Thr Ser Val Leu Phe Asp Ser Asn
Pro Leu Ala Pro Gln Phe Arg Glu 180 185
190 Leu Val His Ser Leu Pro Pro His Gly Tyr Gly Leu Pro
Ala Pro Leu 195 200 205
Phe Gly Gly Gly Gln Gly Gly Asp His Val Asp Gly Leu Ser Gly Gly 210
215 220 Gly Leu Ser Tyr
Gln Asp Gly Gly His Gly Asp Gly Val Phe Glu Phe 225 230
235 240 Thr Ala Glu Met Ala Cys Ile Gly Lys
Gly Ile Arg Lys Ser Gly Lys 245 250
255 Val Ile Thr Lys His Phe Ala Thr Glu Arg Gln Arg Arg Glu
His Leu 260 265 270
Asn Gly Lys Tyr Thr Ala Leu Arg Asn Leu Val Pro Asn Pro Ser Lys
275 280 285 Asn Asp Arg Ala
Ser Val Val Gly Asp Ala Ile Asn Tyr Ile Lys Glu 290
295 300 Leu Leu Arg Thr Val Glu Glu Leu
Lys Leu Leu Val Glu Lys Lys Arg 305 310
315 320 Asn Gly Arg Glu Arg Ile Lys Arg Arg Lys Pro Glu
Glu Asp Gly Gly 325 330
335 Val Asp Val Leu Glu Asn Ser Asn Thr Lys Val Glu Gln Asp Gln Ser
340 345 350 Thr Tyr Asn
Asn Gly Ser Leu Arg Ser Ser Trp Leu Gln Arg Lys Ser 355
360 365 Lys His Thr Glu Val Asp Val Arg
Leu Ile Glu Asp Glu Val Thr Ile 370 375
380 Lys Leu Val Gln Arg Lys Lys Val Asn Cys Leu Leu Ser
Val Ser Lys 385 390 395
400 Val Leu Asp Glu Leu Gln Leu Asp Leu His His Ala Ala Gly Gly Leu
405 410 415 Ile Gly Asp Tyr
Tyr Ser Phe Leu Phe Asn Thr Lys Ile Asn Glu Gly 420
425 430 Ser Cys Val Tyr Ala Ser Gly Ile Ala
Asn Lys Leu Leu Glu Val Val 435 440
445 Asp Arg Gln Tyr Ala Ser Ser Thr Ser Val Pro Ala Ala Ser
Cys 450 455 460
91473PRTSorghum bicolor 91Met Ile Ala Gly Gly Gly Tyr Phe Asp Gly Ser His
Asp His Ile Leu 1 5 10
15 Met Glu Gly Ser Met Ile His Asp Ser Ser Gln Ser Ser Ile Tyr Asp
20 25 30 Asn Thr Asp
Val Glu Gln Gln Asn Phe Arg Leu Ala Pro Phe Ile Ile 35
40 45 Glu Asp His Ser Asn Pro Ala Asn
Leu Thr Ser Glu Pro Ala Arg Val 50 55
60 Ile Asp Gln Ile His His Gln Leu Gly Ile Asp Met Glu
Gln Asp His 65 70 75
80 Ser Asp His Met Ile Gln Gly Val Pro Pro Ala Glu Thr Ala Asn Leu
85 90 95 Val Pro Val Val
Tyr Gly Val Gln Asp Arg Ile Leu Ser His Gln Ile 100
105 110 Glu Gly Pro His Asn Ile Thr Val Glu
Gln Gln Val Leu Asp Tyr Asp 115 120
125 Pro Ala Ser Tyr Gly Asn Gly Thr Tyr Ala Ala Ala His Asp
Leu Leu 130 135 140
Asn Ser Leu Gln Ile Gln Arg Cys Ser Leu Ile Pro Glu Phe Pro Ser 145
150 155 160 Thr Glu His Ile Phe
Gly Asp Pro Ala Gln Asn Met Val Asn Pro Leu 165
170 175 Asp Ile Thr Asn Asp Leu Gln Gly Val Ala
Thr His Glu Ser Gly Met 180 185
190 Met Phe Ser Asp Ser Thr Leu Pro Leu Gly Tyr His Ala Thr Gln
Ser 195 200 205 His
Met Leu Lys Asp Leu Tyr His Ser Leu Pro Gln Asn Tyr Gly Ile 210
215 220 Phe Thr Ser Asp Asp Glu
Arg Asp Gly Met Val Gly Val Ala Gly Val 225 230
235 240 Ser Gly Asn Ile Phe Gln Glu Ile Asp Gly Arg
Gln Phe Asp Ser Pro 245 250
255 Val Leu Gly Thr Arg Arg Gln Lys Gly Gly Phe Gly Lys Gly Lys Gly
260 265 270 Lys Ala
Asn Phe Ala Thr Glu Arg Glu Arg Arg Glu Gln Leu Asn Val 275
280 285 Lys Tyr Gly Ala Leu Arg Ser
Leu Phe Pro Asn Pro Thr Lys Asn Asp 290 295
300 Arg Ala Ser Ile Val Gly Asp Ala Ile Asp Tyr Ile
Asn Glu Leu Asn 305 310 315
320 Arg Thr Val Lys Glu Leu Lys Ile Leu Leu Glu Lys Lys Arg Asn Ser
325 330 335 Thr Asp Arg
Arg Lys Ile Leu Lys Leu Asp Asp Glu Ala Ala Asp Asp 340
345 350 Gly Glu Ser Ser Ser Met Gln Pro
Val Ser Asp Asp Gln Asn Asn Gln 355 360
365 Met Asn Gly Ala Ile Arg Ser Ser Trp Val Gln Arg Arg
Ser Lys Glu 370 375 380
Cys Asp Val Asp Val Arg Ile Val Asp Asp Glu Ile Asn Ile Lys Phe 385
390 395 400 Thr Glu Lys Lys
Arg Ala Asn Ser Leu Leu Cys Ala Ala Lys Val Leu 405
410 415 Glu Glu Phe Arg Leu Glu Leu Ile His
Val Val Gly Gly Ile Ile Gly 420 425
430 Asp His His Ile Phe Met Phe Asn Thr Lys Ile Pro Lys Gly
Ser Ser 435 440 445
Val Tyr Ala Cys Ala Val Ala Lys Lys Leu Leu Glu Ala Val Glu Ile 450
455 460 Lys Lys Gln Ala Leu
Asn Ile Phe Asn 465 470 92486PRTMedicago
truncatula 92Met Ser Ser Gly Ser Gly Asp Lys Gln Asn Met His Glu Gln Asn
Gly 1 5 10 15 Cys
Phe Asp Pro Asn Thr Lys Asp Glu Gly Val Glu Asn Ser Pro Asn
20 25 30 Asp Asn Asn Thr Asn
Asn Asn Asn Ser Leu Glu Glu Asn Phe Lys Pro 35
40 45 Ser Val Glu Glu Leu Pro Tyr His Asn
His Gln Asn Ser Gln His Leu 50 55
60 Asp Asp Val Ser Thr Tyr Thr Asn Gly Phe Thr Pro Ser
Ser Val Asp 65 70 75
80 Ile Glu Gln Leu Gln Asn Leu Gly Leu Asn Ile Gly Asn Thr Tyr Asn
85 90 95 Asn Met Asp Asn
His Leu Val Gln Glu Val Tyr Gln Asn Ser Thr Trp 100
105 110 Asp Pro Ser Val Gln Asp Met Asp Tyr
Val Asn His Gln Glu His Arg 115 120
125 Gln Leu Ser Glu Gln Gln Tyr Gln Gln Phe Ile Glu Ala Gln
Asn His 130 135 140
Asn Gln Ser Tyr Asn Pro Ser Thr Ile Leu Asp Pro His Tyr Pro Ser 145
150 155 160 Pro Asp Val Leu Asn
Leu Leu Asn Leu Pro Arg Cys Ser Ser Ser Leu 165
170 175 Leu Thr Asn Ser Ser Thr Ile Cys Met Thr
Asn Pro Thr Gln Asn Pro 180 185
190 Pro Asn Phe His Asn Ser Met Thr Phe Leu Gly Asp Leu Pro Ile
Gly 195 200 205 Ser
Ser Asp Asn Thr Ser Gly Ser Ser Val Leu Tyr Asp Pro Leu Tyr 210
215 220 Pro Leu Asn Leu Pro Pro
Gln Pro Pro Ala Leu Arg Glu Leu Phe Gln 225 230
235 240 Ser Leu Pro Arg Gly Tyr Ser Met Pro Thr Asn
Ser Arg Asn Gly Ser 245 250
255 Leu Phe Gly Gly Gly Asp Glu Met Glu Gly Asp Gly Asp Met Gly Val
260 265 270 Leu Glu
Phe Asn Arg Val Thr Ala Ser Val Gly Lys Gly Arg Gly Gly 275
280 285 Lys Ala Thr Lys His Phe Ala
Thr Glu Lys Gln Arg Arg Glu Gln Leu 290 295
300 Asn Gly Lys Tyr Lys Ile Leu Arg Asp Leu Ile Pro
Ser Pro Thr Lys 305 310 315
320 Thr Asp Arg Ala Ser Val Val Gly Asp Ala Ile Glu Tyr Ile Arg Glu
325 330 335 Leu Ile Arg
Thr Val Asn Glu Leu Lys Leu Leu Val Glu Lys Lys Arg 340
345 350 His Gly Arg Glu Met Cys Lys Arg
Leu Lys Thr Glu Asp Asp Ala Ala 355 360
365 Glu Ser Cys Asn Ile Lys Pro Phe Gly Asp Pro Asp Gly
Ser Ile Arg 370 375 380
Thr Ser Trp Leu Gln Arg Lys Ser Lys Asp Ser Glu Val Asp Val Arg 385
390 395 400 Ile Ile Asp Asp
Asp Val Thr Ile Lys Leu Phe Gln Arg Lys Lys Val 405
410 415 Asn Cys Leu Leu Phe Val Ser Lys Val
Leu Asp Glu Leu Gln Leu Glu 420 425
430 Leu His His Val Ala Gly Gly His Val Gly Glu Tyr Cys Ser
Phe Leu 435 440 445
Phe Asn Ser Lys Val Asn Glu Gly Ser Ser Val Tyr Ala Ser Ala Ile 450
455 460 Ala Asn Arg Val Ile
Asp Val Met Asp Thr Gln Tyr Ala Ala Gly Leu 465 470
475 480 Pro His Ile Ser Arg Leu
485 93483PRTMedicago truncatula 93Met Ser Ser Gly Ser Gly Asp Asn Gln
Asn Met His Glu Gln Thr Gly 1 5 10
15 Cys Phe Asp Pro Asp Thr Met Ala Glu Gly Val Glu Asn Ser
Pro Glu 20 25 30
Asp Asn Asn Ser Pro Gln Thr Met Pro Asn Gln Val Val Ala Gly Asn
35 40 45 Ser Asn Asn Ser
Ile Glu Glu Asn Phe Arg Pro Ser Val Glu Glu Phe 50
55 60 Ser Tyr His Asn His His Ser Pro
Gln His Leu Glu Asp Val Ser Thr 65 70
75 80 Tyr Thr Asn Gly Phe Thr Pro Ser Ser Glu Asn Ile
Ala Gln Gln Asn 85 90
95 Leu Gly Leu Asn Ile Gly Asn Tyr Tyr Tyr Asn Asn Met Asp Asn Leu
100 105 110 Leu Glu Gln
Glu Val Tyr Gln Asn Ser Ser Trp Asp Pro Ser Ala Gln 115
120 125 Asp Met Asp Tyr Ala Asn His Gln
Glu Tyr His Gln Leu His Asn His 130 135
140 Lys Gln Ser Tyr Asn Pro Ser Thr Thr Gln Ala Pro His
Tyr Pro Ser 145 150 155
160 Pro Asp Val Leu Asn Leu His Phe Pro Arg Ser Ser Ala Ser Ser Leu
165 170 175 Leu Thr Asn Pro
Ser Thr Ile Cys Ile Thr Asn Pro Thr Gln Lys Pro 180
185 190 Pro Asn Phe His Tyr Ser Met Ser Phe
Leu Gly Asp Leu Pro Ile Gly 195 200
205 Ser Asp Asn Ser Ser Gly Ser Ser Val Leu Tyr Asp Pro Leu
Phe Pro 210 215 220
Leu Asn Leu Pro Ala Gln Ser Pro Ala Leu Arg Glu Leu Pro Gln Ser 225
230 235 240 Leu Pro Arg Val Tyr
Ser Met Pro Thr Asn Ser Arg Asn Gly Ser Pro 245
250 255 Phe Gly Gly Gly Asp Glu Met Glu Gly Asp
Gly Gly Met Gly Val Ser 260 265
270 Gln Phe Asn Lys Val Thr Ala Phe Val Gly Lys Gly Lys Gly Lys
Ala 275 280 285 Thr
Glu His Leu Thr Thr Glu Lys Gln Arg Arg Glu Gln Leu Lys Gly 290
295 300 Arg Tyr Lys Ile Leu Arg
Ser Leu Ile Pro Asn Ser Thr Lys Asp Asp 305 310
315 320 Arg Ala Ser Val Val Gly Asp Ala Ile Glu Tyr
Leu Arg Glu Leu Ile 325 330
335 Arg Thr Val Asn Glu Leu Lys Leu Leu Val Glu Lys Lys Arg His Glu
340 345 350 Ile Glu
Ile Cys Lys Arg His Lys Thr Glu Asp Tyr Ala Ala Glu Ser 355
360 365 Cys His Met Lys Pro Phe Gly
Asp Pro Asp Gly Ser Ile Arg Thr Ser 370 375
380 Trp Leu Gln Arg Lys Ser Lys Asp Ser Glu Val Asp
Val Arg Ile Ile 385 390 395
400 Asp Asp Asp Val Thr Ile Lys Leu Phe Gln Arg Lys Lys Val Asn Cys
405 410 415 Leu Leu Phe
Val Ser Lys Val Leu Asp Glu Leu Gln Leu Glu Leu Asn 420
425 430 His Val Ala Gly Gly His Val Gly
Glu Tyr Cys Ser Phe Leu Phe Asn 435 440
445 Ser Lys Val Ile Glu Gly Ser Ser Val His Ala Ser Ala
Ile Ala Asn 450 455 460
Arg Val Ile Asp Val Leu Asp Thr Gln Tyr Ala Ala Val Val Pro His 465
470 475 480 Asn Arg Met
94412PRTCucumis sativus 94Met Glu Leu Asp Phe Gln Gln Ala Ala Ala Ala Pro
Thr Pro Gly Phe 1 5 10
15 Asp Gln Glu Leu Thr Ser Asp Ser Asn Pro Met Leu Cys Leu Asp Gln
20 25 30 Ser Asn Trp
Val Gly Thr Gln Ile Gln Glu Met Gly Phe Asn His Asn 35
40 45 His Val Gln Ser Gln Phe Ser Asp
Ser Ala Ile Pro Pro Thr Pro Tyr 50 55
60 Thr Gln Pro Pro Asp Leu Leu Asn Phe Leu Asn Met Pro
Pro Thr Ala 65 70 75
80 Arg Cys Ser Asn Asn Ser Ser Ile Ser Phe Ser Asn Leu His Thr Pro
85 90 95 Ala Met Gly Ala
Phe Leu Gly Asp Leu Pro Pro Gly Asp Ala Pro Asn 100
105 110 Ser Ser Ser Thr Ser Leu Ser Ile Leu
Tyr Asp Pro Leu Phe His Leu 115 120
125 Asn Leu Pro Pro Gln Pro Pro Leu Phe Arg Glu Leu Phe His
Ser Leu 130 135 140
Pro His Gly Tyr Gly Met Pro Ala Ala Ser Ser Arg Gly Arg Gly Gly 145
150 155 160 Ser Leu Phe Pro Glu
Gly Ser Glu Ile Val Glu Arg Glu Gly Thr Ala 165
170 175 Gly Val Tyr Glu Asp Gly Asp Gly Ser Gly
Val Leu Glu Phe Ser Arg 180 185
190 Asp Met Ala Asp Cys Ile Gly Lys Arg Arg Asp Gly Lys Met Thr
Lys 195 200 205 His
Phe Ala Thr Glu Arg Gln Arg Arg Val Gln Leu Asn Asp Lys Tyr 210
215 220 Lys Ala Leu Arg Ser Leu
Val Pro Ile Pro Thr Lys Asn Asp Arg Ala 225 230
235 240 Ser Ile Val Gly Asp Ala Ile Asn Tyr Ile Gln
Glu Leu Leu Arg Glu 245 250
255 Val Lys Glu Leu Lys Leu Leu Val Glu Lys Lys Arg Ser Ser Arg Glu
260 265 270 Arg Ser
Lys Arg Val Arg Thr Ala Glu Glu Ile Glu Gln Gly Gly Gly 275
280 285 Ser Glu Ser Ser Asn Ala Lys
Gly Gly Glu Gly Val Val Glu Asp Gln 290 295
300 Arg Tyr Asn Leu Arg Ser Ser Trp Leu Gln Arg Lys
Thr Lys Asp Thr 305 310 315
320 Glu Val Asp Val Arg Ile Val Asp Asp Glu Val Thr Val Lys Leu Val
325 330 335 Gln Arg Lys
Leu Asn Cys Leu Leu Leu Val Ser Lys Leu Leu Glu Asp 340
345 350 Leu Gln Leu Asp Leu His His Val
Ala Gly Gly His Ile Gly Asp Tyr 355 360
365 Tyr Ser Phe Leu Phe Asn Thr Lys Ile Tyr Glu Gly Ser
Ser Val Tyr 370 375 380
Ala Ser Ala Ile Ala Asn Lys Val Met Glu Ala Val Asp Arg Gln Tyr 385
390 395 400 Asn Asn Thr Ser
Ile Ser Pro Leu Thr Asn Thr Tyr 405 410
95465PRTBrachypodium distachyon 95Met Ile Ile Gly Gly Asp Tyr Phe
Glu Gly Ser His Asp His Ser Leu 1 5 10
15 Met Ala Gly Ser Leu Ile His Asp Ser Ser Gln Ala Pro
Lys Cys Asn 20 25 30
Gly Asn Thr Asp Ile Glu Leu Gln Lys Phe Lys Val Pro Ser Phe Ser
35 40 45 Ser Glu Ile Leu
Thr Asn Ser Thr Asn Leu Ser Ser Glu Ala Ala Arg 50
55 60 Ala Ile Asn His Leu Gln His Gln
Leu Gly Ile Asp Leu Glu Gln Asp 65 70
75 80 Met Gln Pro Val Glu Thr Ala Thr Trp Asp Ala Ser
Ile Cys Ser Ile 85 90
95 Gln Asp His Ile Ile Asn Asn Gln Ile Ser Glu Asp Pro Gln Asn Ile
100 105 110 Leu Val Glu
Gln Gln Ile Gln Gln Tyr Asp Ala Ala Ile Tyr Pro Asn 115
120 125 Ser Ser Tyr Thr Pro Ala Pro Asp
Leu Leu Asn Leu Leu His Cys Thr 130 135
140 Val Ala Pro Ala Phe Pro Thr Thr Thr Ser Val Phe Gly
Asp Thr Ser 145 150 155
160 Leu Ser Ser Thr Asn Tyr Leu Asp Leu Asn Gly Glu Phe Thr Gly Val
165 170 175 Ala Ala Thr Pro
Glu Ser Gly Leu Met Phe Thr Ser Asp Ser Ala Leu 180
185 190 Gln Leu Gly Tyr His Ala Thr Gln Ser
His Pro Leu Lys Asp Ile Cys 195 200
205 His Ser Leu Pro Gln Asn Tyr Gly Leu Phe Pro Gly Glu Asp
Glu Arg 210 215 220
Glu Val Met Ile Gly Val Gly Ser Val Gly Gly Asp Ile Phe Gln Asp 225
230 235 240 Ile Asp Asp Arg Gln
Phe Asp Thr Val Leu Glu Cys Arg Arg Gly Lys 245
250 255 Gly Glu Phe Gly Lys Gly Lys Gly Lys Ala
Asn Phe Ala Thr Glu Arg 260 265
270 Glu Arg Arg Glu Gln Leu Asn Val Lys Tyr Lys Thr Leu Lys Asp
Leu 275 280 285 Phe
Pro Asn Pro Thr Lys Ser Asp Arg Ala Ser Val Val Gly Asp Ala 290
295 300 Ile Glu Tyr Ile Asp Glu
Leu Asn Arg Thr Val Lys Glu Leu Lys Ile 305 310
315 320 Leu Val Glu Gln Lys Trp His Gly Asn Lys Arg
Thr Lys Ile Ile Lys 325 330
335 Leu Asp Glu Glu Val Ala Ala Asp Gly Glu Ser Ser Ser Met Lys Pro
340 345 350 Met Arg
Asp Asp Gln Asp Asn Gln Phe Asp Gly Thr Ile Arg Ser Ser 355
360 365 Trp Val Gln Arg Arg Ser Lys
Glu Cys His Ile Asp Val Arg Ile Val 370 375
380 Glu Asn Glu Val Asn Ile Lys Leu Thr Glu Lys Lys
Lys Val Asn Ser 385 390 395
400 Leu Leu His Ala Ala Arg Val Leu Asp Glu Phe Gln Leu Glu Leu Ile
405 410 415 His Ala Val
Gly Gly Ile Ile Gly Asp His His Ile Phe Met Phe Asn 420
425 430 Thr Lys Val Ser Glu Gly Ser Ser
Val Tyr Ala Cys Ala Val Ala Lys 435 440
445 Arg Leu Leu Gln Ala Val Asp Ala Gln His Gln Ala Ile
Asn Ile Phe 450 455 460
His 465 96480PRTCucumis sativus 96Met Tyr Glu Glu Thr Glu Cys Ser Asp
Pro Asn Ser Ile Ser Pro Glu 1 5 10
15 Thr Met Pro His Ile Ser Ala Phe Pro Asn Ser Phe Pro Pro
Pro Leu 20 25 30
Ile Ala Gln Gln Thr His Pro Asn Phe His His Asn Asn Asn Leu Asn
35 40 45 Leu Ser Ile Asp
His Ile Ser Tyr His His His Ser Thr Ala Leu Gln 50
55 60 Pro Ala Asp Ala Met Glu Leu Asp
Phe Gln Gln Ala Ala Ala Ala Pro 65 70
75 80 Thr Pro Gly Phe Asp Gln Glu Leu Thr Ser Asp Ser
Asn Pro Met Leu 85 90
95 Cys Leu Asp Gln Ser Asn Trp Val Gly Thr Gln Ile Gln Glu Met Gly
100 105 110 Phe Asn His
Asn His Val Gln Ser Gln Phe Ser Asp Ser Ala Ile Pro 115
120 125 Pro Thr Pro Tyr Thr Gln Pro Pro
Asp Leu Leu Asn Phe Leu Asn Met 130 135
140 Pro Pro Thr Ala Arg Cys Ser Asn Asn Ser Ser Ile Ser
Phe Ser Asn 145 150 155
160 Leu His Thr Pro Ala Met Gly Ala Phe Leu Gly Asp Leu Pro Pro Gly
165 170 175 Asp Ala Pro Asn
Ser Ser Ser Thr Ser Leu Ser Ile Leu Tyr Asp Pro 180
185 190 Leu Phe His Leu Asn Leu Pro Pro Gln
Pro Pro Leu Phe Arg Glu Leu 195 200
205 Phe His Ser Leu Pro His Gly Tyr Gly Met Pro Ala Ala Ser
Ser Arg 210 215 220
Gly Arg Gly Gly Ser Leu Phe Pro Glu Gly Ser Glu Ile Val Glu Arg 225
230 235 240 Glu Gly Thr Ala Gly
Val Tyr Glu Asp Gly Asp Gly Ser Gly Val Leu 245
250 255 Glu Phe Ser Arg Asp Met Ala Asp Cys Ile
Gly Lys Arg Arg Asp Gly 260 265
270 Lys Met Thr Lys His Phe Ala Thr Glu Arg Gln Arg Arg Val Gln
Leu 275 280 285 Asn
Asp Lys Tyr Lys Ala Leu Arg Ser Leu Val Pro Ile Pro Thr Lys 290
295 300 Asn Asp Arg Ala Ser Ile
Val Gly Asp Ala Ile Asn Tyr Ile Gln Glu 305 310
315 320 Leu Leu Arg Glu Val Lys Glu Leu Lys Leu Leu
Val Glu Lys Lys Arg 325 330
335 Ser Ser Arg Glu Arg Ser Lys Arg Val Arg Thr Ala Glu Glu Ile Glu
340 345 350 Gln Gly
Gly Gly Ser Glu Ser Ser Asn Ala Lys Gly Gly Glu Gly Val 355
360 365 Val Glu Asp Gln Arg Tyr Asn
Leu Arg Ser Ser Trp Leu Gln Arg Lys 370 375
380 Thr Lys Asp Thr Glu Val Asp Val Arg Ile Val Asp
Asp Glu Val Thr 385 390 395
400 Val Lys Leu Val Gln Arg Lys Leu Asn Cys Leu Leu Leu Val Ser Lys
405 410 415 Leu Leu Glu
Asp Leu Gln Leu Asp Leu His His Val Ala Gly Gly His 420
425 430 Ile Gly Asp Tyr Tyr Ser Phe Leu
Phe Asn Thr Lys Ile Tyr Glu Gly 435 440
445 Ser Ser Val Tyr Ala Ser Ala Ile Ala Asn Lys Val Met
Glu Ala Val 450 455 460
Asp Arg Gln Tyr Asn Asn Thr Ser Ile Ser Pro Leu Thr Asn Thr Tyr 465
470 475 480 97459PRTAegilops
tauschii 97Met Ile Ala Glu Gly Gly Tyr Phe Asp Gly Ser Arg Asp Ala Ile
Leu 1 5 10 15 Met
Ala Gly Ser Leu Ile His Asp Ser Leu Asp Ser Ile Cys Asp Asn
20 25 30 Thr Glu Ile Glu Gln
Gly Asn Phe His Gly Pro Ser Phe Phe Ile Glu 35
40 45 Asp Ile Cys Asn Pro Thr Asn Leu Thr
Ser Glu Ser Ala Arg Thr Ile 50 55
60 Asn His Ile Gln His Arg Ala Glu Phe Asp Met Asp Gln
Asp Leu His 65 70 75
80 Gly His Met Ile Gln Glu Thr Gln Val Glu Thr Ser Asn Trp Val Pro
85 90 95 Ala Met Phe Gly
Thr Gln Asn His Ile Ile Ser Gln Gln Ser Ile Glu 100
105 110 Gln Gln Met Asp Asp Tyr Asp Ala Ala
Ser Tyr Pro Asp Gly Ala His 115 120
125 Thr Ala Ala Pro Asp Leu Leu Asn Leu Leu Gln Ile Pro Arg
Tyr Ser 130 135 140
Met Thr Thr Ala Phe Pro Ser Thr Glu His Ile Phe Gly Asp Pro Gly 145
150 155 160 Gln Asn Ala Gly Asn
Gln Leu Asp Ile Asn Asn Asp Val Leu Gly Arg 165
170 175 Ala Ile His Asp Ser Gly Met Met Leu Gly
Asp Ser Thr Leu Pro Leu 180 185
190 Gln Tyr Asn Asp Asn Gln Ser His Leu Phe Lys Asp Leu Tyr His
Ser 195 200 205 Leu
Pro Gln Ser Phe Gly Leu Phe Ser Ser Asp Asp Glu Arg Asp Arg 210
215 220 Ala Met Gly Val Val Gly
Ala Ala Gly Asn Ile Leu Gln Glu Ile Asp 225 230
235 240 Gly Arg Gln Phe Gly Ser Pro Lys Leu Gly Arg
Ser Lys Lys Gly Gly 245 250
255 Phe Gly Lys Ala Lys Ala Asn Phe Ala Thr Glu Lys Glu Arg Arg Glu
260 265 270 Gln Ile
Asn Val Lys Tyr Gly Ala Leu Arg Ser Leu Leu Pro Ser Pro 275
280 285 Thr Lys Asn Asp Arg Ala Ser
Ile Val Gly Asp Ala Ile Glu Tyr Ile 290 295
300 Asn Glu Leu Asn Arg Thr Leu Lys Glu Leu Thr Ser
Leu Val Glu Gly 305 310 315
320 Asp Thr Lys His Arg Met Lys Arg Leu Lys Leu Asp Asp Ala Ala Cys
325 330 335 Asp Asn Gly
Glu Ser Ser Ser Leu Gln Gln Val Lys Asp Asp Gln Asp 340
345 350 Ser Gln Leu Asn Gly Ala Ile Arg
Ser Ser Trp Ile Gln Arg Arg Ser 355 360
365 Lys Glu Cys His Val Asp Val Arg Ile Val Gly Asn Glu
Ile Asn Ile 370 375 380
Lys Phe Thr Glu Lys Lys Lys Thr Asn Ser Leu Leu Cys Ala Ala Lys 385
390 395 400 Val Ile Asp Glu
Phe Arg Leu Glu Leu Ile His Val Val Gly Gly Val 405
410 415 Ile Gly Asp Gln Arg Ile Phe Met Phe
Asn Thr Lys Ile Ser Glu Gly 420 425
430 Ser Ser Val Tyr Ala Ser Ala Leu Ala Ser Lys Leu Leu Arg
Ala Met 435 440 445
Glu Met Glu His Leu Ala Val Asp Ile Phe Ser 450 455
98465PRTFragaria vesca 98Met Tyr Val Asp Ser Ser Thr Ala
Ala Ala Gly Ala Cys Asn Phe Asp 1 5 10
15 Pro Asn Thr Asp Thr Asn Pro Met Ser Glu Ser Ala Pro
Glu Val Val 20 25 30
Leu His His Gln Asn Asn Pro Thr Thr Phe Ala Ser Thr His Asp Glu
35 40 45 Asn Leu Arg Ser
Leu Ser Met Glu Glu Glu Leu Ser Asn Tyr His His 50
55 60 Asn His Asn Ala Ala Met Glu Ile
Glu Gln Gln Leu Gln Thr Glu Met 65 70
75 80 Gly Phe Gly Thr Met Asp Gln Asn Thr Asn Asn Thr
Asn Pro His Leu 85 90
95 Ile Asn Pro Phe Asp Thr His Gln Ala Thr Asn Trp Asp Asn Asn Asp
100 105 110 Glu Met Gln
Gln Gln Gln Gln Leu Pro Pro Val Ala Pro Thr Pro Asp 115
120 125 Leu Leu Ser Leu Phe His Leu Pro
Asn Ser Ser Val Leu Pro His Ser 130 135
140 Ser Ile Thr Phe Thr Asn Pro Lys Thr Pro Gly Gly Cys
Phe Pro Gly 145 150 155
160 Ser Phe Gly Tyr Glu Thr Leu Pro Glu Thr Pro Ser Gly Ala Val Ala
165 170 175 Ser Asn Ser Val
Met Tyr Asp Pro Met Phe His Leu Asn Gln Leu Pro 180
185 190 Pro Gln Pro Gln Pro Pro Leu Phe Arg
Glu Leu Leu Gln Ser Leu Pro 195 200
205 His Gly Tyr Lys Arg Asn Gly Ser Ser Leu Phe Ser Asn Gly
Gly Asp 210 215 220
Glu Val Asp Asp Gly Ser Arg Gln Leu Phe Glu Asn Gly Val Leu Glu 225
230 235 240 Phe Ser Lys Glu Met
Lys Pro Phe Gly Arg Gly Arg Gly Gly Asn Lys 245
250 255 Gly Thr Lys His Phe Ala Thr Glu Arg Gln
Arg Arg Val Gln Leu Asn 260 265
270 Asp Lys Phe Ser Ala Leu Arg Glu Leu Val Pro Asn Pro Thr Lys
Pro 275 280 285 Asp
Arg Ala Ser Val Val Gly Asp Ala Ile Asp Tyr Ile Gln Glu Leu 290
295 300 Lys Arg Thr Val Ser Glu
Leu Lys Leu Leu Val Glu Lys Lys Arg Cys 305 310
315 320 Gly Arg Glu Arg Ser Lys Arg His Lys Thr Glu
Gln Asp Ile Gly Ala 325 330
335 Arg Asp Asp Asp Glu Ser Cys Asn Met Lys Pro Leu Gly Asp Pro Asp
340 345 350 His Asp
His Ser Tyr Asn Asn Gly Ser Leu Arg Ser Ser Trp Leu Gln 355
360 365 Arg Lys Ser Lys Asp Thr Glu
Val Asp Val Arg Ile Ile Asp Asp Glu 370 375
380 Val Thr Ile Lys Leu Val Gln Arg Lys Lys Ile Asn
Leu Leu Leu Ser 385 390 395
400 Val Ser Lys Leu Leu Asp Glu Leu Gln Leu Glu Leu His His Ala Ala
405 410 415 Gly Gly His
Ile Gly Asn Ser Tyr Ser Phe Leu Phe Asn Thr Lys Met 420
425 430 Tyr Glu Gly Ser Ser Leu Tyr Ala
Ser Ala Ile Ala Asn Lys Leu Ile 435 440
445 Asp Thr Val Asp Arg Gln Tyr Ala Ala Ala Ile Pro Pro
Thr Asn Ser 450 455 460
Tyr 465 99480PRTAegilops tauschii 99Met Ile Trp Ile Glu Gly Ala Arg His
Cys Phe Val Lys Met Ile Val 1 5 10
15 Gly Gly Asp Tyr Phe Glu Gly Ser His Asp His Asn Leu Met
Thr Gly 20 25 30
Ser Leu Thr His Asp Ser Ser Leu Ala Pro Lys Cys Asn Asp Asn Thr
35 40 45 Asn Ile Glu Leu
Gln Arg Phe Lys Val Gln Ser Phe Ser Ala Asp Ile 50
55 60 Leu Ser Asp Ser Thr Asn Leu Ser
Ser Glu Ala Ala Arg Ala Ile Asn 65 70
75 80 His Leu Gln His Gln Leu Gly Ile Gly Leu Glu Gln
Asp Met Pro Pro 85 90
95 Val Glu Thr Ala Thr Trp Asp Thr Ser Ile Cys Thr Ile Gln Asp Gln
100 105 110 Ile Ile Asn
His Gln Leu Ser Glu Asp Pro Gln Asn Ile Leu Val Gln 115
120 125 Gln Gln Ile Gln Gln Tyr Asp Ala
Ala Leu Tyr Pro Asn Ser Gly Tyr 130 135
140 Thr Pro Ala Pro Asp Leu Leu Asn Leu Leu His Cys Thr
Val Ala Pro 145 150 155
160 Val Phe Pro Ala Thr Ala Ser Val Phe Gly Asp Thr Ala Leu Ser Gly
165 170 175 Gly Thr Asn Tyr
Leu Asp Leu Asn Gly Glu Phe Thr Gly Val Ala Ala 180
185 190 Ile Pro Asp Ser Gly Leu Met Tyr Thr
Ser Asp Pro Ala Leu Gln Leu 195 200
205 Gly Tyr His Ala Ala Pro Ser His Ala Leu Lys Asp Ile Cys
His Ser 210 215 220
Leu Pro Gln Asn Tyr Gly Leu Phe Pro Ser Glu Asp Glu Arg Asp Val 225
230 235 240 Met Leu Gly Val Gly
Ser Val Gly Gly Asp Leu Phe Gln Asp Met Asp 245
250 255 Asp Arg Gln Phe Glu Thr Val Leu Glu Gly
Arg Arg Gly Lys Gly Glu 260 265
270 Phe Gly Lys Gly Lys Gly Lys Ala Asn Phe Ala Thr Glu Arg Glu
Arg 275 280 285 Arg
Glu Gln Leu Asn Val Lys Tyr Lys Thr Leu Arg Met Leu Phe Pro 290
295 300 Asn Pro Thr Lys Asn Asp
Arg Ala Ser Val Val Gly Asp Ala Ile Glu 305 310
315 320 Tyr Ile Asp Glu Leu Asn Arg Thr Val Lys Glu
Leu Lys Ile Leu Val 325 330
335 Glu Gln Lys Trp His Gly Thr Asn Arg Arg Arg Ile Arg Lys Leu Asp
340 345 350 Glu Glu
Ala Ala Ala Asp Gly Glu Ser Ser Ser Met Arg Pro Met Arg 355
360 365 Asp Glu Gln Asp Asn Gln Leu
Asp Gly Ala Ile Arg Ser Ser Trp Val 370 375
380 Gln Arg Arg Ser Arg Glu Cys His Val Asp Val Arg
Ile Val Glu Asn 385 390 395
400 Glu Ile Asn Ile Lys Leu Thr Glu Lys Lys Lys Ala Asn Ser Ser Leu
405 410 415 Leu His Val
Ala Lys Val Leu Asp Glu Phe His Leu Glu Ile Ile His 420
425 430 Val Val Gly Gly Ile Ile Gly Asp
His Tyr Ile Phe Met Phe Asn Thr 435 440
445 Lys Val Thr Glu Gly Ser Ser Val Tyr Ala Cys Ala Val
Ala Lys Arg 450 455 460
Ile Leu Gln Ala Val Asp Ala Gln His Gln Ala Leu Asp Ile Phe Asn 465
470 475 480
100430PRTArabidopsis lyrata 100Met Tyr Glu Glu Ser Ser Cys Phe Asp Pro
Asn Pro Met Val Asp Asn 1 5 10
15 Asn Gly Ser Phe Cys Ala Ala Glu Thr Thr Phe Pro Val Ser His
Gln 20 25 30 Phe
Gln Pro Pro Val Gly Ser Thr Thr Asn Ser Phe Asn Asp Asp Leu 35
40 45 Lys Leu Pro Thr Met Glu
Glu Phe Ser Ala Phe Pro Ser Val Ile Ser 50 55
60 Leu Pro Asn Ser Glu Thr Gln Asn Gln Asn Ile
Ser Asn Asn Asn His 65 70 75
80 Leu Ile Asn Gln Met Ile Gln Glu Pro Asn Trp Gly Val Ser Glu Asp
85 90 95 Asn Thr
Gly Phe Phe Met Asn Thr Ser His Pro Asn Thr Thr Thr Thr 100
105 110 Pro Ile Pro Asp Leu Leu Ser
Leu Leu His Leu Pro Arg Cys Ser Met 115 120
125 Ala Leu Pro Ser Ser Asn Leu Ser Asp Ile Met Ala
Gly Ser Cys Phe 130 135 140
Thr Tyr Asp Pro Leu Cys His Leu Asn Leu Pro Pro Gln Pro Pro Leu 145
150 155 160 Ile Pro Ser
Asn Asp Tyr Ser Gly Tyr Leu Leu Gly Ile Asp Thr Asn 165
170 175 Thr Thr Thr Gln Gly Asp Glu Ser
Asn Val Gly Asp Glu Asn Asn Asn 180 185
190 Ala Gln Phe Asp Ser Gly Ile Ile Glu Phe Ser Lys Glu
Ile Arg Arg 195 200 205
Lys Gly Arg Gly Lys Arg Lys Asn Lys Pro Phe Thr Thr Glu Arg Glu 210
215 220 Arg Arg Cys His
Leu Asn Glu Arg Tyr Glu Ala Leu Lys Leu Leu Ile 225 230
235 240 Pro Asn Pro Ser Lys Gly Asp Arg Ala
Ser Ile Leu Gln Asp Gly Ile 245 250
255 Asp Tyr Ile Asn Glu Leu Arg Arg Arg Val Ser Glu Leu Lys
Tyr Leu 260 265 270
Val Glu Arg Lys Arg Cys Gly Gly Arg His Lys Asn Asn Glu Leu Asp
275 280 285 Asn Asn Ile Asn
Asn Asn Asn Ser Asn Asp His Asp Asn Asp Glu Asp 290
295 300 Asp Ile Asp Asp Glu Asn Met Glu
Lys Lys Pro Glu Ser Asp Val Val 305 310
315 320 Asp Gln Cys Ser Ser Asn Asn Ser Leu Arg Cys Ser
Trp Leu Gln Arg 325 330
335 Lys Ser Lys Val Thr Glu Val Asp Val Arg Ile Val Asp Asp Glu Val
340 345 350 Thr Ile Lys
Val Val Gln Lys Lys Lys Ile Asn Cys Leu Leu Leu Val 355
360 365 Ser Lys Val Leu Asp Gln Leu Gln
Leu Asp Leu Tyr His Val Ala Gly 370 375
380 Gly Gln Ile Gly Glu His Tyr Ser Phe Leu Phe Asn Thr
Lys Ile Tyr 385 390 395
400 Glu Gly Ser Thr Ile Tyr Ala Ser Ala Ile Ala Asn Arg Val Ile Glu
405 410 415 Val Val Asp Lys
His Tyr Met Ala Ala Leu Pro Ile Asn Tyr 420
425 430 101421PRTArabidopsis lyrata 101Met Glu Gly Gly
Gly Met Phe Glu Glu Ile Gly Cys Phe Asp Pro Asn 1 5
10 15 Ala Pro Ala Glu Met Thr Ala Glu Ser
Ser Phe Ser Pro Ala Glu Pro 20 25
30 Pro Pro Thr Ile Thr Val Ile Gly Ser Asn Ser Asn Ser Asn
Cys Ser 35 40 45
Leu Glu Asp Leu Ser Glu Phe His Leu Ser Pro Gln Asp Ser Ser Leu 50
55 60 Pro Ala Ser Ala Ser
Ala Tyr Val His Gln Leu His Val Asn Ala Thr 65 70
75 80 Pro Asn Cys Asp His Gln Phe Gln Ser Ser
Met His Gln Thr Leu Gln 85 90
95 Gly Pro Ser Tyr Pro Gln Gln Ser Asn Asn Trp Asp Asn Gly Tyr
Gln 100 105 110 Asp
Phe Val Asn Leu Val Pro Asn His Thr Thr Pro Asp Leu Leu Ser 115
120 125 Leu Leu Gln Leu Pro Arg
Ser Ser Leu Pro Pro Phe Ala Asn Pro Ser 130 135
140 Leu Gln Asp Ile Ile Met Thr Thr Ser Ser Ser
Val Ala Ala Tyr Asp 145 150 155
160 Pro Leu Phe His Leu Asn Phe Pro Leu Gln Pro Pro Asn Gly Thr Phe
165 170 175 Ile Gly
Val Asp Gln Asp Gln Thr Glu Ile Glu Asn Gln Gly Val Asn 180
185 190 Leu Met Tyr Asp Glu Glu Asn
Asn Asn Leu Asp Asn Gly Leu Asn Arg 195 200
205 Lys Gly Arg Gly Ser Arg Lys Arg Lys Val Phe Pro
Thr Glu Arg Glu 210 215 220
Arg Arg Val His Phe Lys Asp Arg Phe Gly Asp Leu Lys Asn Leu Ile 225
230 235 240 Pro Asn Pro
Thr Lys Asn Asp Arg Ala Ser Ile Val Gly Glu Ala Ile 245
250 255 Asp Tyr Ile Lys Glu Leu Leu Arg
Thr Ile Asp Glu Phe Lys Leu Leu 260 265
270 Val Glu Lys Lys Arg Thr Lys Gln Arg Asn Arg Glu Gly
Asp Asp Val 275 280 285
Ile Asp Glu Asn Phe Lys Ala Gln Ser Glu Val Val Glu Gln Cys Leu 290
295 300 Ile Asn Lys Lys
Asn Asn Ala Leu Arg Cys Ser Trp Leu Lys Arg Lys 305 310
315 320 Ser Lys Phe Thr Glu Val Asp Val Arg
Ile Ile Asp Asp Asp Val Thr 325 330
335 Ile Lys Ile Val Gln Lys Lys Lys Ile Asn Cys Leu Val Phe
Val Ser 340 345 350
Lys Val Val Asp Gln Leu Gln Leu Asp Leu His His Val Ala Gly Ala
355 360 365 Gln Ile Gly Glu
His His Ser Phe Leu Phe Asn Ala Lys Ile Cys Glu 370
375 380 Gly Ser Ser Val Tyr Ala Ser Ala
Ile Ala Asp Arg Val Met Glu Val 385 390
395 400 Leu Glu Lys Gln Tyr Met Glu Ala Leu Ser Thr Asn
Asn Gly Tyr His 405 410
415 Cys Tyr Ser Ser Asp 420 102428PRTArabidopsis
thaliana 102Met Tyr Glu Glu Ser Ser Cys Phe Asp Pro Asn Ser Met Val Asp
Asn 1 5 10 15 Asn
Gly Gly Phe Cys Ala Ala Glu Thr Thr Phe Thr Val Ser His Gln
20 25 30 Phe Gln Pro Pro Leu
Gly Ser Thr Thr Asn Ser Phe Asp Asp Asp Leu 35
40 45 Lys Leu Pro Thr Met Asp Glu Phe Ser
Val Phe Pro Ser Val Ile Ser 50 55
60 Leu Pro Asn Ser Glu Thr Gln Asn Gln Asn Ile Ser Asn
Asn Asn His 65 70 75
80 Leu Ile Asn Gln Met Ile Gln Glu Ser Asn Trp Gly Val Ser Glu Asp
85 90 95 Asn Ser Asn Phe
Phe Met Asn Thr Ser His Pro Asn Thr Thr Thr Thr 100
105 110 Pro Ile Pro Asp Leu Leu Ser Leu Leu
His Leu Pro Arg Cys Ser Met 115 120
125 Ser Leu Pro Ser Ser Asp Ile Met Ala Gly Ser Cys Phe Thr
Tyr Asp 130 135 140
Pro Leu Phe His Leu Asn Leu Pro Pro Gln Pro Pro Leu Ile Pro Ser 145
150 155 160 Asn Asp Tyr Ser Gly
Tyr Leu Leu Gly Ile Asp Thr Asn Thr Thr Thr 165
170 175 Gln Arg Asp Glu Ser Asn Val Gly Asp Glu
Asn Asn Asn Ala Gln Phe 180 185
190 Asp Ser Gly Ile Ile Glu Phe Ser Lys Glu Ile Arg Arg Lys Gly
Arg 195 200 205 Gly
Lys Arg Lys Asn Lys Pro Phe Thr Thr Glu Arg Glu Arg Arg Cys 210
215 220 His Leu Asn Glu Arg Tyr
Glu Ala Leu Lys Leu Leu Ile Pro Ser Pro 225 230
235 240 Ser Lys Gly Asp Arg Ala Ser Ile Leu Gln Asp
Gly Ile Asp Tyr Ile 245 250
255 Asn Glu Leu Arg Arg Arg Val Ser Glu Leu Lys Tyr Leu Val Glu Arg
260 265 270 Lys Arg
Cys Gly Gly Arg His Lys Asn Asn Glu Val Asp Asp Asn Asn 275
280 285 Asn Asn Lys Asn Leu Asp Asp
His Gly Asn Glu Asp Asp Asp Asp Asp 290 295
300 Asp Glu Asn Met Glu Lys Lys Pro Glu Ser Asp Val
Ile Asp Gln Cys 305 310 315
320 Ser Ser Asn Asn Ser Leu Arg Cys Ser Trp Leu Gln Arg Lys Ser Lys
325 330 335 Val Thr Glu
Val Asp Val Arg Ile Val Asp Asp Glu Val Thr Ile Lys 340
345 350 Val Val Gln Lys Lys Lys Ile Asn
Cys Leu Leu Leu Val Ser Lys Val 355 360
365 Leu Asp Gln Leu Gln Leu Asp Leu His His Val Ala Gly
Gly Gln Ile 370 375 380
Gly Glu His Tyr Ser Phe Leu Phe Asn Thr Lys Ile Tyr Glu Gly Ser 385
390 395 400 Thr Ile Tyr Ala
Ser Ala Ile Ala Asn Arg Val Ile Glu Val Val Asp 405
410 415 Lys His Tyr Met Ala Ser Leu Pro Asn
Ser Asn Tyr 420 425
103428PRTArabidopsis thaliana 103Met Tyr Glu Glu Ser Ser Cys Phe Asp Pro
Asn Ser Met Val Asp Asn 1 5 10
15 Asn Gly Gly Phe Cys Ala Ala Glu Thr Thr Phe Thr Val Ser His
Gln 20 25 30 Phe
Gln Pro Pro Leu Gly Ser Thr Thr Asn Ser Phe Asp Asp Asp Leu 35
40 45 Lys Leu Pro Thr Met Asp
Glu Phe Ser Val Phe Pro Ser Val Ile Ser 50 55
60 Leu Pro Asn Ser Glu Thr Gln Asn Gln Asn Ile
Ser Asn Asn Asn His 65 70 75
80 Leu Ile Asn Gln Met Ile Gln Glu Ser Asn Trp Gly Val Ser Glu Asp
85 90 95 Asn Ser
Asn Phe Phe Met Asn Thr Ser His Pro Asn Thr Thr Thr Thr 100
105 110 Pro Ile Pro Asp Leu Leu Ser
Leu Leu His Leu Pro Arg Cys Ser Met 115 120
125 Ser Leu Pro Ser Ser Asp Ile Met Ala Gly Ser Cys
Phe Thr Tyr Asp 130 135 140
Pro Leu Phe His Leu Asn Leu Pro Pro Gln Pro Pro Leu Ile Pro Ser 145
150 155 160 Asn Asp Tyr
Ser Gly Tyr Leu Leu Gly Ile Asp Thr Asn Thr Thr Thr 165
170 175 Gln Arg Asp Glu Ser Asn Val Gly
Asp Glu Asn Asn Asn Ala Gln Phe 180 185
190 Asp Ser Gly Ile Ile Glu Phe Ser Lys Glu Ile Arg Arg
Lys Gly Arg 195 200 205
Gly Lys Arg Lys Asn Lys Pro Phe Thr Thr Glu Arg Glu Arg Arg Cys 210
215 220 His Leu Asn Glu
Arg Tyr Glu Ala Leu Lys Leu Leu Ile Pro Ser Pro 225 230
235 240 Ser Lys Gly Asp Arg Ala Ser Ile Leu
Gln Asp Gly Ile Asp Tyr Ile 245 250
255 Asn Glu Leu Arg Arg Arg Val Ser Glu Leu Lys Tyr Leu Val
Glu Arg 260 265 270
Lys Arg Cys Gly Gly Arg His Lys Asn Asn Glu Val Asp Asp Asn Asn
275 280 285 Asn Asn Lys Asn
Leu Asp Asp His Gly Asn Glu Asp Asp Asp Asp Asp 290
295 300 Asp Glu Asn Met Glu Lys Lys Pro
Glu Ser Asp Val Ile Asp Gln Cys 305 310
315 320 Ser Ser Asn Asn Ser Leu Arg Cys Ser Trp Leu Gln
Arg Lys Ser Lys 325 330
335 Val Thr Glu Val Asp Val Arg Ile Val Asp Asp Glu Val Thr Ile Lys
340 345 350 Val Val Gln
Lys Lys Lys Ile Asn Cys Leu Leu Leu Val Ser Lys Val 355
360 365 Leu Asp Gln Leu Gln Leu Asp Leu
His His Val Ala Gly Gly Gln Ile 370 375
380 Gly Glu His Tyr Ser Phe Leu Phe Asn Thr Lys Ile Tyr
Glu Gly Ser 385 390 395
400 Thr Ile Tyr Ala Ser Ala Ile Ala Asn Arg Val Ile Glu Val Val Asp
405 410 415 Lys His Tyr Thr
Ala Ser Leu Pro Asn Ser Asn Tyr 420 425
104420PRTArabidopsis thaliana 104Met Gly Gly Gly Gly Met Phe Glu
Glu Ile Gly Cys Phe Asp Pro Asn 1 5 10
15 Ala Pro Ala Glu Met Thr Ala Glu Ser Ser Phe Ser Pro
Ser Glu Pro 20 25 30
Pro Pro Thr Ile Thr Val Ile Gly Ser Asn Ser Asn Ser Asn Cys Ser
35 40 45 Leu Glu Asp Leu
Ser Ala Phe His Leu Ser Pro Gln Asp Ser Ser Leu 50
55 60 Pro Ala Ser Ala Ser Ala Tyr Ala
His Gln Leu His Ile Asn Ala Thr 65 70
75 80 Pro Asn Cys Asp His Gln Phe Gln Ser Ser Met His
Gln Thr Leu Gln 85 90
95 Asp Pro Ser Tyr Ala Gln Gln Ser Asn His Trp Asp Asn Gly Tyr Gln
100 105 110 Asp Phe Val
Asn Leu Gly Pro Asn His Thr Thr Pro Asp Leu Leu Ser 115
120 125 Leu Leu Gln Leu Pro Arg Ser Ser
Leu Pro Pro Phe Ala Asn Pro Ser 130 135
140 Ile Gln Asp Ile Ile Met Thr Thr Ser Ser Ser Val Ala
Ala Tyr Asp 145 150 155
160 Pro Leu Phe His Leu Asn Phe Pro Leu Gln Pro Pro Asn Gly Ser Phe
165 170 175 Met Gly Val Asp
Gln Asp Gln Thr Glu Thr Asn Gln Gly Val Asn Leu 180
185 190 Met Tyr Asp Glu Glu Asn Asn Asn Leu
Asp Asp Gly Leu Asn Arg Lys 195 200
205 Gly Arg Gly Ser Lys Lys Arg Lys Ile Phe Pro Thr Glu Arg
Glu Arg 210 215 220
Arg Val His Phe Lys Asp Arg Phe Gly Asp Leu Lys Asn Leu Ile Pro 225
230 235 240 Asn Pro Thr Lys Asn
Asp Arg Ala Ser Ile Val Gly Glu Ala Ile Asp 245
250 255 Tyr Ile Lys Glu Leu Leu Arg Thr Ile Asp
Glu Phe Lys Leu Leu Val 260 265
270 Glu Lys Lys Arg Val Lys Gln Arg Asn Arg Glu Gly Asp Asp Val
Val 275 280 285 Asp
Glu Asn Phe Lys Ala Gln Ser Glu Val Val Glu Gln Cys Leu Ile 290
295 300 Asn Lys Lys Asn Asn Ala
Leu Arg Cys Ser Trp Leu Lys Arg Lys Ser 305 310
315 320 Lys Phe Thr Asp Val Asp Val Arg Ile Ile Asp
Asp Glu Val Thr Ile 325 330
335 Lys Ile Val Gln Lys Lys Lys Ile Asn Cys Leu Leu Phe Val Ser Lys
340 345 350 Val Val
Asp Gln Leu Glu Leu Asp Leu His His Val Ala Gly Ala Gln 355
360 365 Ile Gly Glu His His Ser Phe
Leu Phe Asn Ala Lys Ile Ser Glu Gly 370 375
380 Ser Ser Val Tyr Ala Ser Ala Ile Ala Asp Arg Val
Met Glu Val Leu 385 390 395
400 Lys Lys Gln Tyr Met Glu Ala Leu Ser Ala Asn Asn Gly Tyr His Cys
405 410 415 Tyr Ser Ser
Asp 420 105380PRTGlycine max 105Met Tyr Glu Glu Ser Ser Cys
Tyr Asp Pro Asp Ala Met Met Ala Glu 1 5
10 15 Gly Ala Glu Asp Cys Phe Pro Gln Met Val Ser
Glu Ser Glu Ala Val 20 25
30 Met Ser Ala Thr Pro Thr Gln Thr His Thr His Asn Thr Phe Ala
Tyr 35 40 45 Ser
Tyr Ser Cys Gly Glu Asp Ala Ala Asn Ala Asn Gly Pro Ile Ala 50
55 60 Met Glu His Pro Gln Gln
Asn Pro Tyr Asn Tyr Ser Asn Thr Gln Phe 65 70
75 80 Val Glu Glu Leu Tyr Ser Asn Gln Gln Phe Thr
Tyr His Thr Pro Thr 85 90
95 Pro Asp Leu Leu Asp Leu Leu His Leu Pro Asn Pro Ile Pro Gly Asp
100 105 110 Asn Arg
Thr Asn Val Ser Ser Val Ser Tyr Asp Pro Tyr Leu His Leu 115
120 125 Asn Leu Gln Gln Gln Gln Gln
Pro Thr Leu Arg Glu Leu Leu Pro His 130 135
140 Met Pro Ala Leu Arg Asn Asp Phe Pro Phe Gly Gly
Ala Ala Gly Gly 145 150 155
160 Asp Asp Ile Gln Asp Phe Gly Asn Gly Leu Val Asp Phe Thr Gln Gln
165 170 175 Glu Val Gly
Lys Arg Arg Gly Gly Lys Arg Thr Lys Gln Phe Thr Ser 180
185 190 Thr Thr Thr Glu Arg Gln Arg Arg
Val Asp Leu Ser Ser Lys Phe Asp 195 200
205 Ala Leu Lys Glu Leu Ile Pro Asn Pro Ser Lys Ser Asp
Arg Ala Ser 210 215 220
Val Val Gly Asp Ala Ile Asn Tyr Ile Arg Glu Leu Lys Arg Thr Val 225
230 235 240 Glu Glu Leu Lys
Leu Leu Val Glu Lys Lys Arg Leu Glu Lys Gln Arg 245
250 255 Val Met Met Arg His Lys Val Glu Thr
Glu Gly Glu Ser Ser Asn Leu 260 265
270 Asp Pro Ala Glu Tyr Ser Glu Ser Leu Arg Ser Ser Trp Ile
Gln Arg 275 280 285
Lys Thr Lys Asp Thr Glu Val Asp Val Arg Ile Val Asp Asn Glu Val 290
295 300 Thr Ile Lys Leu Val
Gln Arg Lys Lys Ile Asp Cys Leu Val His Val 305 310
315 320 Ser His Leu Leu Asp Gln Leu Asn Leu Asp
Leu Gln His Val Ala Gly 325 330
335 Gly His Ile Gly Asp Phe Cys Ser Tyr Leu Phe Asn Thr Lys Ile
Cys 340 345 350 Glu
Gly Ser Ser Ile Tyr Ala Ser Ala Ile Ala Asn Lys Leu Ile Gln 355
360 365 Val Met Asp Thr Ser Leu
Ala Ala Ala Ser Leu Ala 370 375 380
106463PRTGlycine maxmisc_feature(429)..(429)Xaa can be any naturally
occurring amino acid 106Met His Glu Gln Thr Gly Cys Phe Asp Pro Asn Thr
Met Gly Glu Ser 1 5 10
15 Val Pro Phe Leu Lys Asp Asn Phe Pro Gln Thr Leu Pro Pro Ser Pro
20 25 30 Ile Val Val
Gly Asn Thr Thr Asn Ser Asn Asn Asn Met Asp Asn His 35
40 45 Leu Val Gln Glu Val Ile Asp Ala
Tyr Pro Tyr Gln Leu Ser Thr Trp 50 55
60 Asp Pro Ala Thr Val Gln Glu Leu Gln Asp Ile Ala Tyr
Ala Asn His 65 70 75
80 Thr Glu Gln Gln Gln Gln Gln Gln Gln Asn Glu Gln Gln Phe Gln Gln
85 90 95 Ile Glu Thr Gln
Asn Cys Ser Gln Ser Tyr Asn Asn Pro Ser Ser Ile 100
105 110 Leu Asp Pro Pro Tyr Pro Ser Pro Asp
Leu Leu Asn Leu Leu His Met 115 120
125 Pro Arg Cys Ser Ala Ser Ser Leu Leu Thr Asn Pro Ser Ile
Cys Leu 130 135 140
Thr Asn Pro Thr Gln Asn Thr Pro Asn Phe Gln Asn Pro Met Ala Phe 145
150 155 160 Leu Gly Asp Leu Thr
Ile Gly Ser Glu Asn Thr Ser Ala Ser Ser Val 165
170 175 Leu Tyr Asp Pro Leu Phe His Leu Asn Leu
Pro Pro Gln Pro Pro Ala 180 185
190 Leu Arg Glu Leu Phe Gln Ser Leu Pro Arg Gly Tyr Ser Leu Pro
Thr 195 200 205 Asn
Ser Arg Asn Gly Ser Leu Phe Ala Gly Gly Asp Glu Met Glu Gly 210
215 220 Asp Gly Ser Gln Leu Asp
Met Gly Val Leu Glu Phe Asn Arg Val Thr 225 230
235 240 Pro Ser Val Gly Lys Gly Arg Gly Gly Lys Ala
Thr Lys His Phe Ala 245 250
255 Thr Glu Lys Gln Arg Arg Glu Gln Leu Asn Gly Lys Tyr Lys Ile Leu
260 265 270 Arg Asn
Leu Ile Pro Ser Pro Thr Lys Leu Ile Gly Trp Val Trp Phe 275
280 285 Asn Thr Asp Asp Arg Ala Ser
Val Val Gly Asp Ala Ile Asp Tyr Ile 290 295
300 Arg Glu Leu Ile Arg Thr Val Asn Glu Leu Lys Leu
Leu Val Glu Lys 305 310 315
320 Lys Arg Tyr Ala Lys Glu Arg Tyr Lys Arg Pro Lys Thr Glu Glu Asp
325 330 335 Ala Ala Glu
Ser Cys Asn Ile Lys Pro Phe Gly Asp Pro Asp Gly Gly 340
345 350 Ile Arg Thr Ser Trp Leu Gln Arg
Lys Ser Lys Asp Ser Glu Val Asp 355 360
365 Val Arg Ile Ile Asp Asp Asp Val Thr Ile Lys Leu Phe
Gln Arg Lys 370 375 380
Lys Ile Asn Cys Leu Leu Phe Val Ser Lys Val Leu Asp Glu Leu Gln 385
390 395 400 Leu Glu Leu His
His Val Ala Gly Gly His Val Gly Glu Tyr Cys Ser 405
410 415 Phe Leu Phe Asn Ser Lys Gly Leu Val
Ser Leu Arg Xaa Ile Met Glu 420 425
430 Gly Ser Ser Val Tyr Ala Ser Ala Ile Ala Asn Arg Val Ile
Asp Val 435 440 445
Leu Asp Ser Gln Tyr Thr Ala Ala Val Pro His Thr Asn Ser Tyr 450
455 460 107475PRTGlycine
maxmisc_feature(441)..(441)Xaa can be any naturally occurring amino acid
107Met Ser Thr Gly Asp Arg Pro Lys Met His Asp Gln Thr Gly Cys Phe 1
5 10 15 Asp Pro Asn Thr
Thr Gly Glu Ser Val Pro Ser Leu Lys Asp Asn Phe 20
25 30 Pro Gln Thr Leu Pro Pro Ser Ser Pro
Met Val Val Val Gly Asn Thr 35 40
45 Thr Thr Asn Ser Asn Asn Asn Met Asp Asn His Leu Val Gln
Glu Val 50 55 60
Ile Asp Ala Phe Pro Tyr Gln Gln Ser Thr Trp Asp Pro Thr Ile Val 65
70 75 80 Gln Glu Leu Gln Asp
Met Ala Tyr Ala Asn His Thr Glu Gln Thr Gln 85
90 95 Gln Gln Gln Gln Asn Glu Gln Gln Phe Gln
Gln Phe Glu Thr Gln Asn 100 105
110 Cys Ser Gln Ser Tyr Asn Asn Pro Ser Ser Ile Leu Asp Pro Pro
Tyr 115 120 125 Pro
Ser Pro Asp Leu Leu Asn Leu Leu His Met Pro Arg Cys Ser Ala 130
135 140 Ser Ser Leu Leu Thr Asn
Pro Ser Ile Cys Leu Thr Asn Pro Thr Gln 145 150
155 160 Asn Thr Pro Asn Phe Gln Asn Pro Met Ala Phe
Leu Gly Asp Leu Pro 165 170
175 Ile Gly Ser Glu Asn Thr Ser Ala Ser Ser Val Leu Tyr Asp Pro Leu
180 185 190 Phe His
Leu Asn Leu Pro Pro Gln Pro Pro Ala Leu Arg Glu Leu Phe 195
200 205 Gln Ser Leu Pro Arg Gly Tyr
Ser Leu Pro Thr Asn Ser Arg Asn Gly 210 215
220 Ser Leu Phe Gly Gly Gly Asp Glu Met Glu Gly Asp
Gly Ser Gln Leu 225 230 235
240 Asp Met Gly Val Leu Glu Phe Asn Arg Val Thr Leu Thr Pro Ser Val
245 250 255 Gly Lys Gly
Arg Arg Gly Lys Ala Thr Lys His Phe Ala Thr Glu Lys 260
265 270 Gln Arg Arg Glu Gln Leu Asn Gly
Lys Tyr Lys Ile Leu Arg Asn Leu 275 280
285 Ile Pro Ser Pro Thr Lys Leu Val Gly Phe Val Leu Thr
Gln Thr Asp 290 295 300
Arg Ala Ser Val Val Gly Asp Ala Ile Asp Tyr Ile Arg Glu Leu Ile 305
310 315 320 Arg Thr Val Asn
Glu Leu Lys Leu Leu Val Glu Lys Lys Arg Tyr Ala 325
330 335 Lys Asp Arg Cys Lys Arg Pro Lys Thr
Glu Glu Asp Ala Ala Glu Ser 340 345
350 Cys Asn Ile Lys Pro Phe Gly Asp Pro Asp Gly Gly Ile Arg
Thr Ser 355 360 365
Trp Leu Gln Arg Lys Ser Lys Asp Ser Glu Val Asp Val Arg Ile Ile 370
375 380 Asp Asp Asp Val Thr
Ile Lys Leu Phe Gln Arg Lys Lys Ile Asn Cys 385 390
395 400 Leu Leu Phe Val Ser Lys Val Leu Asp Glu
Leu Gln Leu Glu Leu His 405 410
415 His Val Ala Gly Gly His Val Gly Glu Tyr Cys Ser Phe Leu Phe
Asn 420 425 430 Ser
Lys Gly Leu Val Ser Leu Arg Xaa Ile Met Glu Gly Ser Ser Val 435
440 445 Tyr Ala Ser Ala Ile Ala
Asn Arg Val Ile Asp Val Leu Asp Ser Gln 450 455
460 Tyr Ala Ala Ala Val Pro His Thr Asn Ser Tyr
465 470 475 108443PRTLotus japonicus
108Met Ala Glu Gly Val Ser Ser Gln Lys Asp Ser Phe Pro Gln Thr Leu 1
5 10 15 Leu Asp Pro Gln
Pro Gln Ser Leu Met Val Thr Glu Asn Thr Thr Asn 20
25 30 Ser Asn Asn Ile Met Asp Asn His Leu
Val Gln Glu Val Ile Asp Ala 35 40
45 Pro Leu Tyr Gln Gln Ser Thr Trp Asp Pro Asn Val Gln Glu
Val Gln 50 55 60
Asp Met Ser Tyr Ala Asn His Pro Glu Gln Gln Phe Gln His Ile Asp 65
70 75 80 Ala Gln Asn Tyr Cys
Gln Ser Tyr Thr Pro Ser Ile Leu Asp Pro Ser 85
90 95 Tyr Pro Ser Pro Asp Leu Leu Asn Phe Leu
His Leu Pro Thr Cys Ser 100 105
110 Ala Ser Ser Leu Leu Thr Asn Pro Pro Asn Ile Cys Ile Ser Asn
Pro 115 120 125 Thr
Gln Arg Thr Pro Asn Phe Gln Asn Ser Met Thr Phe Leu Gly Asp 130
135 140 Leu Pro Met Gly Pro Asp
Asn Thr Ser Ala Ser Ser Val Leu Tyr Asp 145 150
155 160 Pro Leu Phe His Leu Asn Leu Pro Pro Gln Pro
Pro Ala Leu Arg Glu 165 170
175 Leu Phe Gln Ser Leu Pro Arg Gly Tyr Arg Leu Pro Thr Ser Ser Arg
180 185 190 Asp Asp
Ser Leu Phe Gly Gly Gly Asp Glu Met Glu Gly Asp Gly Ser 195
200 205 Gln Leu Asp Met Gly Val Leu
Asp Phe Asn Arg Asp Thr Ala Ser Val 210 215
220 Gly Lys Gly Arg Glu Gly Lys Gly Ala Lys Pro Phe
Ala Thr Glu Lys 225 230 235
240 Asp Arg Arg Glu Gln Leu Asn Gly Lys Tyr Lys Ile Leu Arg Ser Leu
245 250 255 Ile Pro Asn
Pro Thr Lys Leu Ile Gly Trp Val Leu Phe Lys Pro Asp 260
265 270 Arg Ala Ser Val Val Gly Asp Ala
Ile Glu Tyr Ile Arg Glu Leu Ile 275 280
285 Arg Thr Val Asn Glu Leu Lys Leu Leu Val Glu Lys Lys
Arg His Glu 290 295 300
Arg Glu Arg Cys Lys Arg Pro Lys Asn Glu Glu Asp Ala Glu Glu Ser 305
310 315 320 Cys Asn Ile Lys
Pro Phe Gly Asp Pro Asp Gly Tyr Ile Arg Thr Ser 325
330 335 Trp Leu Gln Arg Lys Ser Lys Asp Ser
Glu Val Asp Val Arg Ile Ile 340 345
350 Asp Asp Asp Val Thr Ile Lys Phe Phe Gln Arg Lys Lys Ile
Asn Cys 355 360 365
Leu Leu Phe Val Ser Lys Val Leu Asp Glu Leu Gln Leu Glu Leu His 370
375 380 His Leu Ala Gly Gly
His Val Gly Glu Tyr Trp Ser Phe Leu Phe Asn 385 390
395 400 Ser Lys Arg Pro Val Ser Leu Thr Gln Val
Ile Glu Gly Ser Ser Val 405 410
415 Tyr Ala Ser Ala Ile Ala Asn Arg Val Ile Asp Val Leu Asp Ser
Gln 420 425 430 Tyr
Ala Ala Ala Val Pro Gln Thr Ser Ser Tyr 435 440
109447PRTArabidopsis thaliana 109Met Gly Cys Phe Asp Pro Asn
Thr Pro Ala Glu Val Thr Val Glu Ser 1 5
10 15 Ser Phe Ser Gln Ala Glu Pro Pro Pro Pro Pro
Pro Gln Val Leu Val 20 25
30 Ala Gly Ser Thr Ser Asn Ser Asn Cys Ser Val Glu Val Glu Glu
Leu 35 40 45 Ser
Glu Phe His Leu Ser Pro Gln Asp Cys Pro Gln Ala Ser Ser Thr 50
55 60 Pro Leu Gln Phe His Ile
Asn Pro Pro Pro Pro Pro Pro Pro Pro Cys 65 70
75 80 Asp Gln Leu His Asn Asn Leu Ile His Gln Met
Ala Ser His Gln Gln 85 90
95 Gln His Ser Asn Trp Asp Asn Gly Tyr Gln Asp Phe Val Asn Leu Gly
100 105 110 Pro Asn
Ser Ala Thr Thr Pro Asp Leu Leu Ser Leu Leu His Leu Pro 115
120 125 Arg Cys Ser Leu Pro Pro Asn
His His Pro Ser Ser Met Leu Pro Thr 130 135
140 Ser Phe Ser Asp Ile Met Ser Ser Ser Ser Ala Ala
Ala Val Met Tyr 145 150 155
160 Asp Pro Leu Phe His Leu Asn Phe Pro Met Gln Pro Arg Asp Gln Asn
165 170 175 Gln Leu Arg
Asn Gly Ser Cys Leu Leu Gly Val Glu Asp Gln Ile Gln 180
185 190 Met Asp Ala Asn Gly Gly Met Asn
Val Leu Tyr Phe Glu Gly Ala Asn 195 200
205 Asn Asn Asn Gly Gly Phe Glu Asn Glu Ile Leu Glu Phe
Asn Asn Gly 210 215 220
Val Thr Arg Lys Gly Arg Gly Ser Arg Lys Ser Arg Thr Ser Pro Thr 225
230 235 240 Glu Arg Glu Arg
Arg Val His Phe Asn Asp Arg Phe Phe Asp Leu Lys 245
250 255 Asn Leu Ile Pro Asn Pro Thr Lys Ile
Asp Arg Ala Ser Ile Val Gly 260 265
270 Glu Ala Ile Asp Tyr Ile Lys Glu Leu Leu Arg Thr Ile Glu
Glu Phe 275 280 285
Lys Met Leu Val Glu Lys Lys Arg Cys Gly Arg Phe Arg Ser Lys Lys 290
295 300 Arg Ala Arg Val Gly
Glu Gly Gly Gly Gly Glu Asp Gln Glu Glu Glu 305 310
315 320 Glu Asp Thr Val Asn Tyr Lys Pro Gln Ser
Glu Val Asp Gln Ser Cys 325 330
335 Phe Asn Lys Asn Asn Asn Asn Ser Leu Arg Cys Ser Trp Leu Lys
Arg 340 345 350 Lys
Ser Lys Val Thr Glu Val Asp Val Arg Ile Ile Asp Asp Glu Val 355
360 365 Thr Ile Lys Leu Val Gln
Lys Lys Lys Ile Asn Cys Leu Leu Phe Thr 370 375
380 Thr Lys Val Leu Asp Gln Leu Gln Leu Asp Leu
His His Val Ala Gly 385 390 395
400 Gly Gln Ile Gly Glu His Tyr Ser Phe Leu Phe Asn Thr Lys Ile Cys
405 410 415 Glu Gly
Ser Cys Val Tyr Ala Ser Gly Ile Ala Asp Thr Leu Met Glu 420
425 430 Val Val Glu Lys Gln Tyr Met
Glu Ala Val Pro Ser Asn Gly Tyr 435 440
445 110458PRTArabidopsis thaliana 110Met Glu Glu Glu Arg
Glu Ser Leu Tyr Glu Glu Met Gly Cys Phe Asp 1 5
10 15 Pro Asn Thr Pro Ala Glu Val Thr Val Glu
Ser Ser Phe Ser Gln Ala 20 25
30 Glu Pro Pro Pro Pro Pro Pro Gln Val Leu Val Ala Gly Ser Thr
Ser 35 40 45 Asn
Ser Asn Cys Ser Val Glu Val Glu Glu Leu Ser Glu Phe His Leu 50
55 60 Ser Pro Gln Asp Cys Pro
Gln Ala Ser Ser Thr Pro Leu Gln Phe His 65 70
75 80 Ile Asn Pro Pro Pro Pro Pro Pro Pro Pro Cys
Asp Gln Leu His Asn 85 90
95 Asn Leu Ile His Gln Met Ala Ser His Gln Gln Gln His Ser Asn Trp
100 105 110 Asp Asn
Gly Tyr Gln Asp Phe Val Asn Leu Gly Pro Asn Ser Ala Thr 115
120 125 Thr Pro Asp Leu Leu Ser Leu
Leu His Leu Pro Arg Cys Ser Leu Pro 130 135
140 Pro Asn His His Pro Ser Ser Met Leu Pro Thr Ser
Phe Ser Asp Ile 145 150 155
160 Met Ser Ser Ser Ser Ala Ala Ala Val Met Tyr Asp Pro Leu Phe His
165 170 175 Leu Asn Phe
Pro Met Gln Pro Arg Asp Gln Asn Gln Leu Arg Asn Gly 180
185 190 Ser Cys Leu Leu Gly Val Glu Asp
Gln Ile Gln Met Asp Ala Asn Gly 195 200
205 Gly Met Asn Val Leu Tyr Phe Glu Gly Ala Asn Asn Asn
Asn Gly Gly 210 215 220
Phe Glu Asn Glu Ile Leu Glu Phe Asn Asn Gly Val Thr Arg Lys Gly 225
230 235 240 Arg Gly Ser Arg
Lys Ser Arg Thr Ser Pro Thr Glu Arg Glu Arg Arg 245
250 255 Val His Phe Asn Asp Arg Phe Phe Asp
Leu Lys Asn Leu Ile Pro Asn 260 265
270 Pro Thr Lys Ile Asp Arg Ala Ser Ile Val Gly Glu Ala Ile
Asp Tyr 275 280 285
Ile Lys Glu Leu Leu Arg Thr Ile Glu Glu Phe Lys Met Leu Val Glu 290
295 300 Lys Lys Arg Cys Gly
Arg Phe Arg Ser Lys Lys Arg Ala Arg Val Gly 305 310
315 320 Glu Gly Gly Gly Gly Glu Asp Gln Glu Glu
Glu Glu Asp Thr Val Asn 325 330
335 Tyr Lys Pro Gln Ser Glu Val Asp Gln Ser Cys Phe Asn Lys Asn
Asn 340 345 350 Asn
Asn Ser Leu Arg Cys Ser Trp Leu Lys Arg Lys Ser Lys Val Thr 355
360 365 Glu Val Asp Val Arg Ile
Ile Asp Asp Glu Val Thr Ile Lys Leu Val 370 375
380 Gln Lys Lys Lys Ile Asn Cys Leu Leu Phe Thr
Thr Lys Val Leu Asp 385 390 395
400 Gln Leu Gln Leu Asp Leu His His Val Ala Gly Gly Gln Ile Gly Glu
405 410 415 His Tyr
Ser Phe Leu Phe Asn Thr Lys Ile Cys Glu Gly Ser Cys Val 420
425 430 Tyr Ala Ser Gly Ile Ala Asp
Thr Leu Met Glu Val Val Glu Lys Gln 435 440
445 Tyr Met Glu Ala Val Pro Ser Asn Gly Tyr 450
455 111315PRTOryza sativa 111Met Asp Glu Gln
Arg Gly Arg Gly Gly Phe Asp Glu Leu Val Leu Leu 1 5
10 15 His Gln Gln Gln Glu Gln Arg Arg Arg
Arg Glu Gln Gln Gln Glu Glu 20 25
30 Glu Glu Glu Glu Glu Val Arg Arg Gln Met Phe Gly Ala Val
Val Gly 35 40 45
Gly Leu Ala Ala Phe Pro Ala Ala Ala Ala Ala Leu Gly Gln Gln Gln 50
55 60 Val Asp Cys Gly Gly
Glu Leu Gly Gly Phe Cys Asp Ser Glu Ala Gly 65 70
75 80 Gly Ser Ser Glu Pro Glu Ala Ala Ala Gly
Ala Arg Pro Arg Gly Gly 85 90
95 Ser Gly Ser Lys Arg Ser Arg Ala Ala Glu Val His Asn Leu Ser
Glu 100 105 110 Lys
Arg Arg Arg Ser Lys Ile Asn Glu Lys Met Lys Ala Leu Gln Ser 115
120 125 Leu Ile Pro Asn Ser Asn
Lys Thr Asp Lys Ala Ser Met Leu Asp Glu 130 135
140 Ala Ile Glu Tyr Leu Lys Gln Leu Gln Leu Gln
Val Gln Met Leu Ser 145 150 155
160 Met Arg Asn Gly Val Tyr Leu Asn Pro Ser Tyr Leu Ser Gly Ala Leu
165 170 175 Glu Pro
Ala Gln Ala Ser Gln Met Phe Ala Ala Leu Gly Gly Asn Asn 180
185 190 Val Thr Val Val His Pro Gly
Thr Val Met Pro Pro Val Asn Gln Ser 195 200
205 Ser Gly Ala His His Leu Phe Asp Pro Leu Asn Ser
Pro Pro Gln Asn 210 215 220
Gln Pro Gln Ser Leu Ile Leu Pro Ser Val Pro Ser Thr Ala Ile Pro 225
230 235 240 Glu Pro Pro
Phe His Leu Glu Ser Ser Gln Ser His Leu Arg Gln Phe 245
250 255 Gln Leu Pro Gly Ser Ser Glu Phe
His Lys Ile Leu Phe Leu His Val 260 265
270 Leu Leu Ser Val Lys Asp Gly Val Ser Trp Arg Asp Asn
Ala Lys Ala 275 280 285
Pro Pro Ile Ile Thr Ser Arg Lys Ser Ala Arg Lys Arg Asp Glu Leu 290
295 300 His Gln Glu Arg
Ile Ile His Val Glu His Gln 305 310 315
112464PRTOryza sativa 112Met Ile Val Gly Ala Gly Tyr Phe Glu Asp Ser His
Asp Gln Ser Leu 1 5 10
15 Met Ala Gly Ser Leu Ile His Asp Ser Asn Gln Ala Pro Ala Ser Ser
20 25 30 Glu Asn Thr
Ser Ile Asp Leu Gln Lys Phe Lys Val His Pro Tyr Ser 35
40 45 Thr Glu Ala Leu Ser Asn Thr Ala
Asn Leu Ala Glu Ala Ala Arg Ala 50 55
60 Ile Asn His Leu Gln His Gln Leu Glu Ile Asp Leu Glu
Gln Glu Val 65 70 75
80 Pro Pro Val Glu Thr Ala Asn Trp Asp Pro Ala Ile Cys Thr Ile Pro
85 90 95 Asp His Ile Ile
Asn His Gln Phe Ser Glu Asp Pro Gln Asn Ile Leu 100
105 110 Val Glu Gln Gln Ile Gln Gln Tyr Asp
Ser Ala Leu Tyr Pro Asn Gly 115 120
125 Val Tyr Thr Pro Ala Pro Asp Leu Leu Asn Leu Met Gln Cys
Thr Met 130 135 140
Ala Pro Ala Phe Pro Ala Thr Thr Ser Val Phe Gly Asp Thr Thr Leu 145
150 155 160 Asn Gly Thr Asn Tyr
Leu Asp Leu Asn Gly Glu Leu Thr Gly Val Ala 165
170 175 Ala Val Pro Asp Ser Gly Ser Gly Leu Met
Phe Ala Ser Asp Ser Ala 180 185
190 Leu Gln Leu Gly Tyr His Gly Thr Gln Ser His Leu Ile Lys Asp
Ile 195 200 205 Cys
His Ser Leu Pro Gln Asn Tyr Gly Leu Phe Pro Ser Glu Asp Glu 210
215 220 Arg Asp Val Ile Ile Gly
Val Gly Ser Gly Asp Leu Phe Gln Glu Ile 225 230
235 240 Asp Asp Arg Gln Phe Asp Ser Val Leu Glu Cys
Arg Arg Gly Lys Gly 245 250
255 Glu Phe Gly Lys Gly Lys Gly Lys Ala Asn Phe Ala Thr Glu Arg Glu
260 265 270 Arg Arg
Glu Gln Leu Asn Val Lys Phe Arg Thr Leu Arg Met Leu Phe 275
280 285 Pro Asn Pro Thr Lys Asn Asp
Arg Ala Ser Ile Val Gly Asp Ala Ile 290 295
300 Glu Tyr Ile Asp Glu Leu Asn Arg Thr Val Lys Glu
Leu Lys Ile Leu 305 310 315
320 Val Glu Gln Lys Arg His Gly Asn Asn Arg Arg Lys Val Leu Lys Leu
325 330 335 Asp Gln Glu
Ala Ala Ala Asp Gly Glu Ser Ser Ser Met Arg Pro Val 340
345 350 Arg Asp Asp Gln Asp Asn Gln Leu
His Gly Ala Ile Arg Ser Ser Trp 355 360
365 Val Gln Arg Arg Ser Lys Glu Cys His Val Asp Val Arg
Ile Val Asp 370 375 380
Asp Glu Val Asn Ile Lys Leu Thr Glu Lys Lys Lys Ala Asn Ser Leu 385
390 395 400 Leu His Ala Ala
Lys Val Leu Asp Glu Phe Gln Leu Glu Leu Ile His 405
410 415 Val Val Gly Gly Ile Ile Gly Asp His
His Ile Phe Met Phe Asn Thr 420 425
430 Lys Val Ser Glu Gly Ser Ala Val Tyr Ala Cys Ala Val Ala
Lys Lys 435 440 445
Leu Leu Gln Ala Val Asp Val Gln His Gln Ala Leu Asp Ile Phe Asn 450
455 460 113379PRTOryza
sativa 113Met Tyr His Pro Gln Cys Glu Leu Leu Met Pro Leu Glu Ser Leu Glu
1 5 10 15 Met Asp
Val Gly Gln Ser His Leu Ala Ala Ala Val Ala Ala Ala Met 20
25 30 Pro Gly Glu Leu Asn Phe His
Leu Leu His Ser Leu Asp Ala Ala Ala 35 40
45 Ala Ala Ala Ser Ser Thr Ala Ala Ser Ala Ser Ser
Gln Pro Thr Val 50 55 60
Asp Tyr Phe Phe Gly Gly Ala Asp Gln Gln Pro Pro Pro Pro Ala Ala 65
70 75 80 Met Gln Tyr
Asp Gln Leu Ala Ala Pro His His His Gln Thr Val Ala 85
90 95 Met Leu Arg Asp Tyr Tyr Gly Gly
His Tyr Pro Pro Ala Ala Ala Ala 100 105
110 Ala Ala Ala Thr Glu Ala Tyr Phe Arg Gly Gly Pro Arg
Thr Ala Gly 115 120 125
Ser Ser Ser Leu Val Phe Gly Pro Ala Asp Asp Glu Ser Ala Phe Met 130
135 140 Val Gly Pro Phe
Glu Ser Ser Pro Thr Pro Arg Ser Gly Gly Gly Arg 145 150
155 160 Lys Arg Ser Arg Ala Thr Ala Gly Phe
His Gly Gly Gly Pro Ala Asn 165 170
175 Gly Val Glu Lys Lys Glu Lys Gln Arg Arg Leu Arg Leu Thr
Glu Lys 180 185 190
Tyr Asn Ala Leu Met Leu Leu Ile Pro Asn Arg Thr Lys Glu Asp Arg
195 200 205 Ala Thr Val Ile
Ser Asp Ala Ile Glu Tyr Ile Gln Glu Leu Gly Arg 210
215 220 Thr Val Glu Glu Leu Thr Leu Leu
Val Glu Lys Lys Arg Arg Arg Arg 225 230
235 240 Glu Met Gln Gly Asp Val Val Asp Ala Ala Thr Ser
Ser Val Val Ala 245 250
255 Gly Met Asp Gln Ala Ala Glu Ser Ser Glu Gly Glu Val Met Ala Ala
260 265 270 Ala Ala Met
Gly Ala Val Ala Pro Pro Pro Arg Gln Ala Pro Ile Arg 275
280 285 Ser Thr Tyr Ile Gln Arg Arg Ser
Lys Glu Thr Phe Val Asp Val Arg 290 295
300 Ile Val Glu Asp Asp Val Asn Ile Lys Leu Thr Lys Arg
Arg Arg Asp 305 310 315
320 Gly Cys Leu Ala Ala Ala Ser Arg Ala Leu Asp Asp Leu Arg Leu Asp
325 330 335 Leu Val His Leu
Ser Gly Gly Lys Ile Gly Asp Cys His Ile Tyr Met 340
345 350 Phe Asn Thr Lys Ile His Ser Gly Ser
Pro Val Phe Ala Ser Ala Val 355 360
365 Ala Ser Arg Leu Ile Glu Val Val Asp Glu Tyr 370
375 114234PRTOryza sativa 114Met Pro Arg Arg
Ala Arg Ala Arg Gly Gly Gly Gly Gly Gly Gly Glu 1 5
10 15 Glu Val Lys Val Glu Asp Asp Phe Ile
Asp Ser Val Leu Asn Phe Gly 20 25
30 Gly Gly Gly Gly Gly Glu Glu Asp Gly Asp Asp Gly Glu Glu
Glu Gln 35 40 45
Gln Gln Gln Gln Ala Ala Ala Ala Ala Met Gly Lys Glu Phe Lys Ser 50
55 60 Lys Asn Leu Glu Ala
Glu Arg Arg Arg Arg Gly Arg Leu Asn Gly Asn 65 70
75 80 Ile Phe Ala Leu Arg Ala Val Val Pro Lys
Ile Thr Lys Met Ser Lys 85 90
95 Glu Ala Thr Leu Ser Asp Ala Ile Glu His Ile Lys Asn Leu Gln
Asn 100 105 110 Glu
Val Leu Glu Leu Gln Arg Gln Leu Gly Asp Ser Pro Gly Glu Ala 115
120 125 Trp Glu Lys Gln Cys Ser
Ala Ser Cys Ser Glu Ser Phe Val Pro Thr 130 135
140 Glu Asn Ala His Tyr Gln Gly Gln Val Glu Leu
Ile Ser Leu Gly Ser 145 150 155
160 Cys Lys Tyr Asn Leu Lys Ile Phe Trp Thr Lys Arg Ala Gly Leu Phe
165 170 175 Thr Lys
Val Leu Glu Ala Leu Cys Ser Tyr Lys Val Gln Val Leu Ser 180
185 190 Leu Asn Thr Ile Ser Phe Tyr
Gly Tyr Ala Glu Ser Phe Phe Thr Ile 195 200
205 Glu Val Lys Gly Glu Gln Asp Val Val Met Val Glu
Leu Arg Ser Leu 210 215 220
Leu Ser Ser Ile Val Glu Val Pro Ser Ile 225 230
115552PRTOryza sativa 115Met Gly Arg Gly Asp His Leu Leu Met
Lys Asn Ser Asn Ala Ala Ala 1 5 10
15 Ala Ala Ala Ala Val Asn Gly Gly Gly Thr Ser Leu Asp Ala
Ala Leu 20 25 30
Arg Pro Leu Val Gly Ser Asp Gly Trp Asp Tyr Cys Ile Tyr Trp Arg
35 40 45 Leu Ser Pro Asp
Gln Arg Phe Leu Glu Met Thr Gly Phe Cys Cys Ser 50
55 60 Ser Glu Leu Glu Ala Gln Val Ser
Ala Leu Leu Asp Leu Pro Ser Ser 65 70
75 80 Ile Pro Leu Asp Ser Ser Ser Ile Gly Met His Ala
Gln Ala Leu Leu 85 90
95 Ser Asn Gln Pro Ile Trp Gln Ser Ser Ser Glu Glu Glu Glu Ala Asp
100 105 110 Gly Gly Gly
Gly Ala Lys Thr Arg Leu Leu Val Pro Val Ala Gly Gly 115
120 125 Leu Val Glu Leu Phe Ala Ser Arg
Tyr Met Ala Glu Glu Gln Gln Met 130 135
140 Ala Glu Leu Val Met Ala Gln Cys Gly Gly Gly Gly Ala
Gly Asp Asp 145 150 155
160 Gly Gly Gly Gln Ala Trp Pro Pro Pro Glu Thr Pro Ser Phe Gln Trp
165 170 175 Asp Gly Gly Ala
Asp Ala Gln Arg Leu Met Tyr Gly Gly Ser Ser Leu 180
185 190 Asn Leu Phe Asp Ala Ala Ala Ala Asp
Asp Asp Pro Phe Leu Gly Gly 195 200
205 Gly Gly Gly Asp Ala Val Gly Asp Glu Ala Ala Ala Ala Gly
Ala Trp 210 215 220
Pro Tyr Ala Gly Met Ala Val Ser Glu Pro Ser Val Ala Val Ala Gln 225
230 235 240 Glu Gln Met Gln His
Ala Ala Gly Gly Gly Val Ala Glu Ser Gly Ser 245
250 255 Glu Gly Arg Lys Leu His Gly Gly Asp Pro
Glu Asp Asp Gly Asp Gly 260 265
270 Glu Gly Arg Ser Gly Gly Ala Lys Arg Gln Gln Cys Lys Asn Leu
Glu 275 280 285 Ala
Glu Arg Lys Arg Arg Lys Lys Leu Asn Gly His Leu Tyr Lys Leu 290
295 300 Arg Ser Leu Val Pro Asn
Ile Thr Lys Met Asp Arg Ala Ser Ile Leu 305 310
315 320 Gly Asp Ala Ile Asp Tyr Ile Val Gly Leu Gln
Lys Gln Val Lys Glu 325 330
335 Leu Gln Asp Glu Leu Glu Asp Asn His Val His His Lys Pro Pro Asp
340 345 350 Val Leu
Ile Asp His Pro Pro Pro Ala Ser Leu Val Gly Leu Asp Asn 355
360 365 Asp Asp Ala Ser Pro Pro Asn
Ser His Gln Gln Gln Pro Pro Leu Ala 370 375
380 Val Ser Gly Ser Ser Ser Arg Arg Ser Asn Lys Asp
Pro Ala Met Thr 385 390 395
400 Asp Asp Lys Val Gly Gly Gly Gly Gly Gly Gly His Arg Met Glu Pro
405 410 415 Gln Leu Glu
Val Arg Gln Val Gln Gly Asn Glu Leu Phe Val Gln Val 420
425 430 Leu Trp Glu His Lys Pro Gly Gly
Phe Val Arg Leu Met Asp Ala Met 435 440
445 Asn Ala Leu Gly Leu Glu Val Ile Asn Val Asn Val Thr
Thr Tyr Lys 450 455 460
Thr Leu Val Leu Asn Val Phe Arg Val Met Val Arg Asp Ser Glu Val 465
470 475 480 Ala Val Gln Ala
Asp Arg Val Arg Asp Ser Leu Leu Glu Val Thr Arg 485
490 495 Glu Thr Tyr Pro Gly Val Trp Pro Ser
Pro Gln Glu Glu Asp Asp Ala 500 505
510 Lys Phe Asp Gly Gly Asp Gly Gly Gln Ala Ala Ala Ala Ala
Ala Ala 515 520 525
Ala Gly Gly Glu His Tyr His Asp Glu Val Gly Gly Gly Tyr His Gln 530
535 540 His Leu His Tyr Leu
Ala Phe Asp 545 550 11615PRTArtificial
Sequenceantigen 116Cys Ser Pro Thr Pro Arg Ser Gly Gly Gly Arg Lys Arg
Ser Arg 1 5 10 15
11720DNAArtificial Sequenceprimer 117tgacctcgtc cacctctccg
2011822DNAArtificial Sequenceprimer
118cagtctgtaa cgagcaagcg ga
22119286DNAOryza sativa 119catgttcaac accaagattc attcgggatc tccagtgttt
gcaagtgcag tggccagcag 60gctgattgaa gtggtggatg agtactaact agctcgagct
agctaattag ccgaccgacc 120gatcgatatg atgaaagttt ctatgttgct agctagctag
ggttcttgga tgcatgagta 180ctgagtagct ctttaattaa tttcctttta attttagact
gtttaatttg gattggtaaa 240gactcgtgtt agcttttggg agatctttgg tatgtcatgg
tttgca 286120149DNAOryza sativa 120caacaaacct
agttaattta gctctagttg gttcatccct gctgcactgc gagctcaagt 60aatcgatctg
agctctgaag aaaaaggtgg tagagtgcga ggaagatgta tcacccgcag 120tgcgagctcc
tgatgccgct tgagagcct 149
User Contributions:
Comment about this patent or add new information about this topic:
 |  |
 |  |
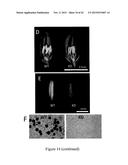 |  |
 |  |
 |  |
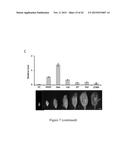 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
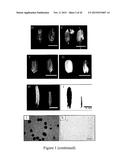 |  |
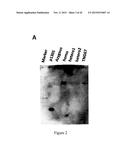 |  |
 |  |
 |  |
 | 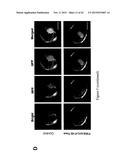 |
 |  |
 |  |
 | 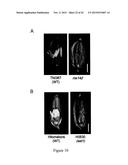 |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
| Similar patent applications: | |
| Date | Title |
|---|---|
| 2016-02-04 | Novel gene ibenod93 and transgenic plants using the same |
| 2016-03-31 | Low lignin non-transgenic alfalfa varieties and methods for producing the same |
| 2016-01-07 | Light-activated chimeric opsins and methods of using the same |
| 2016-01-07 | Hahb11 provides improved plant yield and tolerance to abiotic stress |
| 2015-12-03 | Humanized mouse and methods of using the same |
| New patent applications in this class: | |
| Date | Title |
|---|---|
| 2022-05-05 | Suppression of target gene expression through genome editing of native mirnas |
| 2019-05-16 | Plants having altered agronomic characteristics under nitrogen limiting conditions and related constructs and methods involving low nitrogen tolerance genes |
| 2017-08-17 | Genes and proteins for aromatic polyketide synthesis |
| 2017-08-17 | Insecticidal proteins and methods for their use |
| 2016-09-01 | Bg1 compositions and methods to increase agronomic performance of plants |
| Top Inventors for class "Multicellular living organisms and unmodified parts thereof and related processes" | |
| Rank | Inventor's name |
|---|---|
| 1 | Gregory J. Holland |
| 2 | William H. Eby |
| 3 | Richard G. Stelpflug |
| 4 | Laron L. Peters |
| 5 | Justin T. Mason |