Patent application title: ISOLATION OF PROTEINS INVOLVED IN POSTTRANSCRIPTIONAL GENE SILENCING AND METHODS OF USE
Inventors:
T. Erik Mirkov (Harlingen, TX, US)
T. Erik Mirkov (Harlingen, TX, US)
Ivan L. Ingelbrecht (Jabbeke, BE)
IPC8 Class: AC12N1582FI
USPC Class:
800279
Class name: Multicellular living organisms and unmodified parts thereof and related processes method of introducing a polynucleotide molecule into or rearrangement of genetic material within a plant or plant part the polynucleotide confers pathogen or pest resistance
Publication date: 2012-11-01
Patent application number: 20120278946
Abstract:
The present disclosure relates, according to some embodiments, to methods
of increasing viral resistance in a plant, methods of increasing
transgene expression in a plant, and methods of suppressing expression of
a native gene in a plant. In some embodiments, the disclosure also
relates to polypeptides involved in post-translational gene silencing
(PTGS), for e.g., a protein active in PTGS or a suppressor of PTGS. The
disclosure further relates to transgenic plant cells in which PTGS is
altered (e.g., enhanced or suppressed) according to some embodiments.Claims:
1-23. (canceled)
24. A method of increasing viral resistance in a plant comprising the steps of: i) selecting a protein suppressive of PTGS in the plant; and ii) transforming the plant with a nucleic acid encoding the PTGS suppressive protein.
25. A method of increasing expression of a transgene in a plant comprising the steps of: i) selecting a protein active in PTGS in the plant; and ii) transforming the plant with a nucleic acid encoding the protein active in PTGS.
26. A method of suppressing expression of a native gene in a plant comprising: i) preparing a vector including a nucleic acid with a sequence of the coding portion of the gene wherein the nucleic acid, upon transcription, products an mRNA molecule double stranded in the region corresponding the to the coding portion of the gene; and ii) transforming the plant with the vector.
27. A polypeptide comprising an amino acid sequence having at least 78% identity with the amino acid sequence of SEQ.ID.NO. 2.
28. A polypeptide according to claim 27, wherein the polypeptide comprises the amino acid sequence of SEQ.ID.NO. 2.
29. A polypeptide comprising the amino acid sequence of SEQ.ID.NO. 4.
30. A polypeptide comprising an amino acid having at least 97% identity with the amino acid sequence of SEQ.ID.NO. 10.
31. A polypeptide according to claim 30, wherein the polypeptide comprises the amino acid sequence of SEQ.ID.NO. 10.
32. A transgenic plant cell in which PTGS is enhanced comprising a nucleic acid having a sequence with at least 40% identity with the nucleic acid sequence of SEQ.ID.NO. 3.
33. A transgenic plant cell in which PTGS is enhanced according to claim 32, wherein the nucleic acid comprises the nucleic acid sequence of SEQ.ID.NO. 3.
34. A transgenic plant cell comprising a nucleic acid having the nucleic acid sequence of SEQ.ID.NO. 3 or SEQ.ID.NO. 9.
Description:
CLAIM TO PRIOR APPLICATIONS
[0001] The present application is a divisional application of U.S. patent application Ser. No. 11/460,836 filed on Jul. 28, 2006, which is a divisional application of U.S. application Ser. No. 11/252,080 filed on Oct. 17, 2005, now abandoned, which is a divisional application of U.S. application Ser. No. 10/226,715 filed on Aug. 23, 2002, issued as U.S. Pat. No. 7,001,739 on Feb. 21, 2006, which claims priority to U.S. Provisional Patent Application Ser. No. 60/314,863 filed on Aug. 24, 2001, the contents of which are hereby incorporated by reference in their entirety.
TECHNICAL FIELD OF THE INVENTION
[0002] The present invention relates generally to sugarcane protein isolation and more particularly to isolation and characterization of proteins that are involved in posttranscriptional gene silencing. The invention also includes sugarcane and sorghum mosaic virus proteins involved in silencing and the cDNAs which encode them. Finally the invention includes use of these cDNAs and proteins to regulate silencing.
BACKGROUND OF THE INVENTION
[0003] In the course of evolution organisms have developed a spectrum of defense mechanisms that target alien, parasitic elements. The most specialized defense system is perhaps the vertebrate immune system, which provides effective protection against a wide range of infectious microbes. In the vertebrate immune system, it is essentially peptides that are recognized as non-self and eliminated. Transgenic plant studies have revealed the existence of another, more ancestral level of defense response. Homology-dependent gene silencing can be viewed as a novel, innate host defense system that is capable of recognizing foreign nucleic acids as non-self and inactivating or removing them from the cell. First recognized in plants and fungi, homology-dependent gene silencing mechanisms have now been shown to operate in a wide range of eukaryotic organisms. In plants, this type of gene silencing may occur at the level of DNA, by inhibition of transcription, or at the level of RNA, by enhanced RNA turnover. Both viral RNA and transgenes are subject to these host surveillance systems, which are only poorly understood at the molecular level. At the core of both silencing events resides a molecular mechanism that is able to recognize nucleic acid sequence homology.
[0004] Transgene-induced gene silencing in plants was originally described as the coordinated suppression of transgenes that share sequence similarity (Depicker and Van Montagu, 1997). This phenomenon is most often induced when multiple copies of a transgene are present at a single locus. Silencing not only affects all genes in that locus, i.e. in cis, but also acts in trans, and additionally down-regulates the expression of other, unlinked transgene(s). Silencing can also affect the expression of endogenous genes, provided they have sequence similarity to the silencing transgene, a phenomenon referred to as cosuppression (Napoli et al., 1990; Van der Krol et al., 1990).
[0005] In plants, cases of transgene-induced gene silencing belong to two different mechanistic classes: those that occur at the level of transcription and those that are due to enhanced RNA turnover. Transcriptional gene silencing (TGS) requires sequence identity in the promoter region and is associated with methylation and inactivation of the promoter sequences of the affected genes (Kumpatla et al., 1998). In posttranscriptional gene silencing (PTGS), the (trans)genes remain actively transcribed but the steady-state RNA levels are highly reduced due to sequence-specific RNA degradation. Some instances of PTGS are associated with DNA methylation located in the transcribed portion of the genes (Ingelbrecht et al., 1994; 1999).
[0006] The expression level, number and configuration of the integrated transgenes as well as developmental and environmental factors can all influence the occurrence of transgene-induced gene silencing. Importantly, transgene-induced gene silencing in plants is reversible, and in the absence of the silencer locus, expression of endogenous genes or other transgenes can be restored to normal. The changes in gene expression are therefore not due to irreversible changes in DNA but rather are epigenetic.
[0007] PTGS behaves as a non-clonal event and, in agreement with this, it has been shown that a sequence-specific signal is involved in the systemic spread of PTGS (Palauqui et al., 1997; Voinnet and Baulcombe, 1997). These experiments allow differentiation of separate initiation and maintenance phases in PTGS and further suggest that a molecular system amplifies the silencing signal during the course of long-distance movement of PTGS. Mutants that enhance (Dehio and Schell, 1994) or suppress (Elmayan et al., 1998; Mourrain et al., 2000; Dalmay et al, 2000) PTGS have been isolated in Arabidopsis thaliana but only two of the corresponding genes have been cloned (Mourrain et al., 2000; Dalmay et al, 2000). One of these has no significant similarity with any known or putative protein (Mourrain et al., 2000) and the other is similar to a RNA-dependent RNA polymerase (RdRp; Mourrain et al., 2000; Dalmay et al, 2000). Establishment of PTGS in plants requires separately identifiable initiation, spread, and maintenance phases, but the proteins involved in these pathways have not been characterized.
[0008] Plant virus studies have greatly contributed to the current understanding of gene silencing in general and PTGS in particular. Applying the concept of pathogen-derived resistance, viral genes were introduced into plants and resulted in virus resistant phenotypes. Many resistance phenotypes do not require the expression of a functional protein but are mediated at the level of RNA. It is now an established fact that a mechanism similar to PTGS is the underlying molecular mechanism in most of these cases (van den Boogaart et al., 1998).
[0009] Posttranscriptional silencing of an endogenous plant gene or transgene can be triggered by replication of a recombinant virus that carries sequences homologous to these genes (Kumagai et al., 1995; Ruiz et al., 1998). This process involves sequence-specific RNA turnover, similar to PTGS induced by transgenes, hence the term virus-induced gene silencing. Moreover, natural virus infection of non-transgenic plants can induce a resistance mechanism that is strain-specific and targeted against RNA, similar to RNA-mediated resistance induced by (silenced) transgenes (Ratcliff et al., 1997; Covey et al., 1997). Transgene- and virus-induced gene silencing are collectively described as homology-dependent gene silencing because these mechanisms all target homologous nucleic acid sequences. It was proposed that homology-dependent gene silencing acts as a natural plant defense mechanism against invading DNA or RNA elements (Matzke and Matzke, 1998).
[0010] The demonstration that plant viral proteins can suppress PTGS provides direct evidence that PTGS functions as a host defense response in plants (Anandalakshmi et al., 1998; Brigneti et al., 1998; Kasschau and Carrington, 1998). At least 5 different proteins encoded by unrelated DNA and RNA viruses of plants have now been shown to act as suppressors of PTGS in Nicotiana benthamiana. Importantly, the suppression phenotypes induced by these viral proteins are distinct indicating that separate steps of the host PTGS defense system are targeted. For example, the potyviral helper-component proteinase (HC-Pro) can reverse the effects of PTGS in tissues that were previously silenced, whereas the 2b protein of Cucumber mosaic virus only affects initiation of PTGS (Voinnet et al., 1999). Although potyviral HC-Pro by itself is sufficient to suppress transgene-induced silencing, it appears that the potyviral P1 protein can enhance its ability to suppress virus-induced gene silencing (Anandalakshmi et al., 1998; V. Vance). The discovery of viral suppressors of silencing phenomena is unique to plants. So far, no animal or fungal viruses have been shown to suppress PTGS in these organisms.
[0011] It has been proposed that `aberrant` RNA molecules trigger PTGS in plants (Lindbo et al., 1993). The exact nature of this aberrant RNA is unknown but it could be double-stranded RNA (dsRNA) (Waterhouse et al., 1998), prematurely terminated transcripts, levels of RNA that exceed a certain threshold, or some other unusual characteristic. These RNA molecules would serve as templates for an RNA-dependent RNA polymerase (RdRp) and lead to the production of short complementary RNAs (cRNA). These cRNAs would then anneal with homologous mRNAs or viral RNAs and the resulting double-stranded RNA would be degraded by double strand-specific RNases. This model accounts for the sequence-specific RNA turnover and several aspects of it are supported by experimental data. For example, an RdRp that is induced during viral infection has been cloned in tomato (Schiebel et al., 1998) and small cRNAs have recently been identified in transgenic plants that display PTGS (Hamilton and Baulcombe, 1999). The identification of a double strand-specific RNase in Caenorhabditis elegans and a RdRp-like protein in Neurospora crassa, and recently in Arabadopsis, as essential components of PTGS-like mechanisms in these organisms (see below) provides further support for this hypothesis.
[0012] RNA-mediated genetic interference (RNAi) in C. elegans is a process that closely resembles PTGS in plants: both act at the posttranscriptional level and result in sequence-specific RNA turnover (Tabara et al., 1998; Montgomery and Fire, 1998). The trigger for RNAi in C. elegans is well characterized and consists of dsRNA (Sharp, 1999). RNA-specific silencing can be induced by locally injecting homologous dsRNA molecules in a few cells. Silencing then spreads from the site of injection into neighboring cells and tissues and is even transmitted to the F1 progeny. The ability of silencing to move both in space and over time strongly suggests that amplification of the silencing signal is taking place, similar to PTGS in plants.
[0013] Recently, several genes have been identified in C. elegans that are required for this interference process. The MUT-7 gene encodes a homolog of RNaseD, which is a double strand-specific RNase (Ketting et al., 1999). The RDE-1 gene belongs to a family of genes that are conserved from plants to vertebrates and several members of this family are required for gene silencing mechanisms in animal systems (Tabara et al., 1999). Interestingly, mutations in both these genes reactivate mobilization of endogenous transposons, suggesting that one function of RNAi is transposon silencing. Sequence-specific inhibition of gene function by dsRNA has also been demonstrated in trypanosomes, Drosophila and planaria and has been used in these organisms as a method to determine gene functions (Kennerdell and Carthew, 1998; Misquitta and Patterson, 1999; Sanchez Alvarado and Newmark, 1999).
[0014] Transgene-induced PTGS is termed `quelling` in the fungus N. crassa (Cogoni and Macino, 1997a). Quelling-defective (qde) mutants of N. crassa, in which transgene-induced gene silencing is impaired, have been isolated and could be classified in three qde complementation groups (Cogoni and Macino, 1997b). Two QDE genes that belong to two different complementation groups, have recently been cloned. The QDE-1 gene encodes a protein that contains an RdRp-motif (Cogoni and Macino, 1999a) and QDE-3 belongs to the RecQ DNA helicase family (Cogoni and Macino, 1999b).
[0015] As summarized above, there has been substantial progress in the general understanding of PTGS in plants and its importance as part of a general defense system is now fully appreciated. However, all of the biochemical pathways of PTGS and the enzymes that are involved have not yet been elucidated in plants. Insight into these mechanisms may come from analyzing mutants that are defective in PTGS. This approach has already been used with success in Neurospora and C. elegans and is currently also being followed for Arabidopsis. While this strategy is relatively straightforward and will surely result in the identification of genes that play a central role in this process, there are also limitations. For example, gene redundancies and possibly lethal, loss-of-function phenotypes might prevent identification of certain genes. There are also practical problems in generating and screening a sufficiently large number of mutants which limit this approach to model plants such as Arabidopsis.
[0016] An alternative or complementary approach involves directly identifying the host factors that mediate PTGS. The identification of viral proteins as suppressors of PTGS provides the necessary tools to pursue this strategy.
[0017] Identification and characterization of proteins that interact with a viral suppressor of PTGS will have an impact on understanding fundamentals of virus-host plant interactions, particularly on the mechanisms that plants employ to combat viral infection and on the counterdefensive strategies that viruses use to suppress or evade these responses. To date, viral suppression of PTGS is a process unique to plants. However, because PTGS is a defense mechanism that is conserved among various eukaryotic kingdoms, the identified protein interactions might also shed light on the molecular mechanisms of silencing phenomena in other organisms.
[0018] In addition to significance for basic (plant) molecular virology, establishing the biochemical pathways of host defense responses will facilitate the development of improved virus control strategies in plants. PTGS-based approaches for virus control are already in use but the lack of a solid understanding of the phenomenon necessitates a more empirical approach and has an uncertain outcome. Also, such approaches are currently limited because of their narrow range. Possible and realistic improvements involve enhanced and more predictable triggering and broadening the scope of the PTGS defense system.
[0019] Use of the method of the present invention will also contribute to plant genetic engineering in general. It is now clear that transgenes in plants (and other organisms) can be perceived as intrusive elements and consequently are inactivated. Developing procedures that allow stable and predictable transgene expression is one of the challenges of genetic engineering. The monocot crop plants provide the most important source of food worldwide and offer great potential for improvement through genetic transformation, not only for traits related to food production but also as recombinant expression systems for high value products.
[0020] Finally, gene silencing can be used as a way to produce `knock-out` phenotypes in reverse genetic studies. This has already been successfully applied in animal systems and its potential has been demonstrated in plants. With an increasing number of genes being discovered in sugarcane, many of which have no known function, it can be expected that these approaches will become even more important in the future.
[0021] Thus, the yeast two-hybrid method of the present invention has been used to unravel the pathway(s) of PTGS and plant defense responses and novel, key proteins involved in this process have been identified. In doing so, a cDNA library from silenced plant tissues rather than non-silence plant tissues has been used. These proteins and genes can be applied towards regulating PTGS of transgenes, endogenous plant genes, and viral genes. Specific applications of the present invention include but are not limited to, improved strategies for engineered virus resistance, increased expression of transgenes by inhibiting silencing, and modulation of silencing of native genes to obtain desirable traits or in functional genomic studies.
SUMMARY OF THE INVENTION
[0022] The present invention includes a method of isolating nucleic acid encoding a plant polypeptide active in PTGS. As used throughout the application, plant may mean a mature plant, an embryonic callus, or other stages of plant development. It may also mean a portion of a plant, a plant tissue, or a plant cell.
[0023] The first step of a method of the above method involves selecting a bait nucleic acid which encodes a bait protein active in PTGS in plants or suppressive of PTGS in plants. After bait selection, a cDNA prey library may be prepared from a plant that actively exhibits PTGS at the time of library generation. If an entire plant exhibiting PTGS is not available, tissues in which PTGS is exhibited may be selected.
[0024] After the bait and prey are selected, a yeast two-hybrid assay may be conducted with the bait and prey nucleic acids. Prey cDNA that yields a true positive yeast two-hybrid assay result encodes a polypeptide active in PTGS in the plant. True positive status may be verified using methods known in the art, such as null controls, reversal of bait and prey, and in vitro and in planta studies of interactions. Such in vitro assay may include farwestern blot assays and pull down assays. They may be performed under plant physiological conditions to eliminate false negatives and false positives. In planta studies may be performed in an embryonic callus or other plant tissue.
[0025] In an exemplary embodiment of the above method of the present invention, the bait nucleic acid comprises a sequence selected from SEQ.ID.NO. 1, SEQ.ID.NO. 3, SEQ.ID.NO. 5, SEQ.ID.NO. 7, SEQ.ID.NO. 9, or SEQ.ID.NO. 11. The entire nucleic acids of these sequences may be used or only portions thereof. Additionally, substituted nucleic acids and nucleic acids with similar identities may also be used.
[0026] In another exemplary embodiment the plant is a monocot, particularly sugarcane and more particularly Saccharum hybrid cultivar CP72-1210.
[0027] The present invention also includes several SrMV and sugarcane novel proteins and nucleic acids. Novel nucleic acids are provided in SEQ.ID.NOS. 1, 3, 5, 7, 9 and 11. Novel amino acid sequences are provided in SEQ.ID.NOS. 2, 4, 6, 8, 10 and 12. It will be apparent to one skilled in the art that portions of these nucleic acids and proteins or polypeptides may be used in various applications. Additionally, it will be apparent to one skilled in the art that nucleic acids and proteins or polypeptides with high similarity, particularly in regions related to functional domains, may also be substituted. The present invention includes such variations up to the point of disclosures already in the prior art. Each of these proteins is involved in PTGS either as an activator or suppressor. Some proteins may fail to function alone and may rather require or assist another protein for their PTGS-related functions.
[0028] The present invention additionally includes transgenic plants including any of the above novel nucleic acids, proteins or polypeptides. In particular, the invention includes a transgenic plant in which PTGS is suppressed that includes the nucleic acid of SEQ.ID.NO. 1 or the protein of SEQ.ID.NO. 2. It also includes a transgenic plant in which PTGS is enhanced that includes the nucleic acid of SEQ.ID.NO. 3 or the protein of SEQ.ID.NO. 4.
[0029] The invention additionally includes a method of increasing viral resistance in a plant in which a protein suppressive of PTGS in the plant is selected and the plant is transformed with a nucleic acid encoding the PTGS suppressive protein.
[0030] Another method of the invention involves a method of increasing expression of a transgene in a plant by selecting a protein active in PTGS in the plant and transforming the plant with a nucleic acid encoding the protein active in PTGS.
[0031] Yet another method of the present invention involves suppressing expression of a native gene in a plant by preparing a vector including a nucleic acid with a sequence of the coding portion of the gene wherein the nucleic acid, upon transcription, products an mRNA molecule double stranded in the region corresponding the to the coding portion of the gene. A plant is then transformed with the vector.
BRIEF DESCRIPTION OF THE DRAWINGS
[0032] For a more complete understanding of the present invention and the advantages thereof, reference is now made to the following description taken in conjunction with the accompanying drawings, wherein like reference numbers represent like parts, and which:
[0033] FIG. 1A provides the cDNA (SEQ.ID.NO. 1) and FIG. 1B provides the amino acid (SEQ.ID.NO. 2) sequences for Sorghum Mosaic Virus (SrMV) P1/HC-Pro according to an embodiment of the present invention;
[0034] FIG. 2 is a Northern analysis of SrMV (coat protein) CP mRNA in which the lane labeled "S" includes mRNA derived from a plant posttranscriptionally silenced for SrMV CP; lanes labeled "Retransformed S" include mRNA from samples of the silenced plant which were retransformed with either nptII alone or with nptII and SrMV P1/HC-Pro; the lane including mRNA from a plant that is not silenced is labeled "NS"; and the empty lane is labeled "b";
[0035] FIG. 3 is a β-galactosidase filter lift assay on L40 yeast transformed with various DNA binding and activation domain constructs and grown on media that does not select for interactions (-leu +zeo) or media that does select for transformation and interaction (-leu, -his +zeo); where the region labeled "A" represents yeast transformed with SrMV HC-Pro bait+SrMV HC-Pro prey; the region labeled "B" represents yeast transformed with SrMV HC-Pro bait+empty prey; the region labeled "C" represents yeast transformed with lamin bait+SrMV HC-Pro prey; and the region labeled "D" represents yeast transformed with empty bait+SrMV HC-Pro prey; although the figure is presented in black and white, growth in the regions marked A on both plants appears blue while, on the -leu +zeo plate, growth in regions B-D appears red;
[0036] FIG. 4 is a gel analysis of the cDNA library cloned in pIVING1154; where Lane 1 contains a λPstI size marker; Lane 2 contains a lower band of 0.8 kb which is the CP coding region amplified from the plasmid cDNA library and a top band of 6.6-kb which is the plasmid vector; and Lane 3 contains a SfiI digest of plasmid library showing the 6.6-kb vector and inserts visible as a smear with size range between 0.5 and 2.0 kb;
[0037] FIG. 5 is a β-galactosidase filter lift assay on L40 yeast transformed with various DNA binding and activation domain constructs and grown on media that does not select for interactions (-leu +zeo) or media that does select for transformation and interaction (-leu, -his +zeo); where the region labeled "A" represents yeast transformed with SrMV HC-Pro bait+SrMV HC-Pro prey; the region labeled "B" represents yeast transformed with empty bait+RNase H-like protein prey; the region labeled "C" represents yeast transformed with lamin bait+RNase H-like protein prey; and the region labeled "D" represents yeast transformed with SrMV HC-Pro bait+RNase H-like protein prey; although the figure is not presented in color, the regions marked A and D on both plates appear blue, while the regions marked B and C on the -leu +zeo plate appear red;
[0038] FIG. 6A is the cDNA sequence of the RNase H-like protein (SEQ.ID.NO. 3) and FIG. 6B is the encoded amino acid sequence (SEQ.ID.NO. 4), according to an embodiment of the present invention;
[0039] FIG. 7A is the cDNA sequence of the sugarcane RING zinc finger protein that interacts with SrMV HC-Pro (SEQ.ID.NO. 5); FIG. 7B is the encoded amino acid sequence (SEQ.ID.NO. 6) according to an embodiment of the present invention;
[0040] FIG. 8A is the cDNA sequence of the LRR (leucine-rich repeat) transmembrane protein kinase that interacts with SrMV HC-Pro (SEQ.ID.NO. 7); FIG. 8B is the encoded amino acid sequence (SEQ.ID.NO. 8) according to an embodiment of the present invention;
[0041] FIG. 9A is the cDNA sequence of a nucleic acid encoding the sugarcane 14-3-3 protein that interacts with the RNase H-like protein (SEQ.ID.NO. 9); FIG. 9B is the encoded amino acid (SEQ.ID.NO. 10), according to an embodiment of the invention;
[0042] FIG. 10A is the cDNA sequence of the sugarcane RING zinc finger protein that interacts with RNase H-like protein (SEQ.ID.NO. 11) and FIG. 10B provides the encoded amino acid sequence (SEQ.ID.NO. 12) according to an embodiment of the present invention;
[0043] FIG. 11A shows a 12% PA, CB stained gel; FIG. 11B shows a Large S-AP probed blot; in parts of FIG. 10, the molecular weight lane is indicated as "MW", lane 1 contains the E. coli expression product of Bugbuster® (Novagen, Madison, Wis., affiliate of Merck KgaA, Darmstadt, Germany) Insoluble pET30 with no insert; lane 2 contains the E. coli expression product of Bugbuster® Insoluble pET30 with an SrMV HC-Pro insert; lane 3 contains one preparation of Ni column purified E. coli expression product of Bugbuster® Insoluble pET30 with an SrMV HC-Pro insert; and lane 4 contains a second preparation of Ni column purified E. coli expression product of Bugbuster® Insoluble pET30 with an SrMV HC-Pro insert;
[0044] FIG. 12 shows a two hour exposure of a 15% SDS PAGE gel containing RNase H-like protein (lane labeled "RNase H") labeled with 35S Cysteine according to the present invention; control lanes are provided in which no DNA was used in the preparation procedure (lane labeled "No DNA") and in which Luciferase DNA was used (lane labeled "Luciferase"); molecular weight markers are indicated;
[0045] FIG. 13 shows a 32 hour exposure of a farwestern blot probed under in vitro plant cell physiological conditions with a 35S labeled RNase H-like protein transcription and translation (TNT) product; the lane labeled "His-S/HC-Pro" contains Ni column purified, His-S tagged SrMV HC-Pro protein produced in E. coli; the lane labeled "His-S" contains the HIS-S tag only; the lane labeled "BSA" contains untagged bovine serum albumen; the lane labeled "HEWL" contains untagged hen egg white lysozyme and the lane labeled "RNase A" contains untagged RNase A;
[0046] FIG. 14 shows a 6 hour exposure of a farwestern blot probed under in vitro plant cell physiological conditions with a 35S labeled RNase H-like protein transcription and translation (TNT) product; the lane labeled "His-S/HC-Pro" contains Ni column purified, His-S tagged SrMV HC-Pro protein produced in E. coli; the lane labeled "His-S/RNase H" contains Ni column purified, His-S tagged RNase H-like protein produced in E. coli; the lane labeled "His-S" contains the product in E. coli of the His-S tag only; the unlabeled lanes contain plant extracts which are from left to right, from: healthy sugarcane plant, SrMV infected sugarcane plant, sugarcane plant transgenic for SrMV P1/HC-Pro, sugarcane plant transgenic for delta N 12 SrMV HC-Pro.; and the lane labeled MWM contains molecular weight markers, which were used to produce the size indicators on the side of the blot;
[0047] FIG. 15 shows a 32 hour exposure of a 15% SDS PAGE gel on which Ni column pull down products of a TNT 35S labeled protein/His-S tagged protein physiological incubation were run; the top lane marker indicates the source of DNA used in a TNT procedure in to obtain 35S labeled protein, where "RNase" indicates the use of RNase H-like protein cDNA, "No DNA" indicates a control in which no DNA template was provided, and "Luciferase" indicates a control in which Luciferase DNA was provided; the bottom lane marker indicates the His-S tagged product produced in E. coli, where "His-S/HC-Pro" or "HS/HC-Pro" indicates Ni column purified His-S tagged SrMV HC-Pro protein, "His-S" indicates the His-S tag only, and "BSA" indicates untagged bovine serum albumen;
[0048] FIG. 16 shows a 3 hour exposure of a farwestern blot probed under in vitro plant cell physiological conditions with a 35S labeled RNase H-like protein transcription and translation (TNT) product; the lane labeled "His-S/14-3-3" contains Ni column purified, His-S tagged sugarcane 14-3-3 protein produced in E. coli; the lane labeled "His-S" contains the His-S tag only; the lane labeled "BSA" contains untagged bovine serum albumen; the lane labeled "HEWL" contains untagged hen egg white lysozyme; and the lane labeled "RNase A" contains untagged RNase A.
[0049] FIG. 17A depicts a DNA construct containing sense and antisense sequences in opposite directions and separated by an intron which, following transcription to mRNA, form a double strand due to sense/antisense sequence complementation shown in FIG. 17B;
[0050] FIG. 18 depicts the pMCG161 plasmid expression vector, according to one embodiment of the present invention;
[0051] FIG. 19 depicts the results of transient expression of constructs inserted in the pMGC161 plasmid in embryonic sugarcane calli where the callus 1 is transformed with GUS under control of a Ubi promoter; callus 2 is transformed with GUS under control of a Ubi promoter and DNA that produces double stranded GUS mRNA under control of a 35Sint promoter; callus 3 is transformed with the same constructs as callus 2 and additionally with DNA that produces double stranded RNase H-like protein mRNA under control of the 35Sint promoter; callus 4 is transformed with the same constructs as callus 2 and additionally with SrMV P1/HC-Pro under the control of the Ubi promoter; callus 5 is transformed with the same constructs as callus 2 and additionally with SrMV HC-Pro-delta N 12 under control of the Ubi promoter for ease of plasmid construction to include a start ATG, the first N-terminal amino acids of the full length SrMV HC-Pro are deleted via deletion of 36 cDNA base pairs); and callus 6 is transformed with the same constructs as callus 2 and additionally with DNA that produces double stranded GFP under control of the 35Sint promoter; although the figure is not provided in color, all darker regions in the calli of the figure are blue while ligher regions are pinkish tan, as will be apparent to one skilled in the art;
[0052] FIG. 20 is a graphical representation of three experiments such as that of FIG. 19; vertical bars represent the average of these three independent experiments while vertical line represent standard errors; treatment 1 represents transformation with GUS under control of a Ubi promoter; treatment 2 represents transformation with GUS under control of a Ubi promoter and DNA that produces double stranded GUS mRNA under control of a 35Sint promoter; treatment 3 represents transformation with the same constructs as treatment 2 and additionally with DNA that produces double stranded RNase H-like protein mRNA under control of the 35Sint promoter; treatment 4 represents transformation with the same constructs as treatment 2 and additionally with RNase H-like protein under control of the 35Sint promoter; treatment 5 represents transformation with the same constructs as treatment 2 and additionally with SrMV P1 and HC-Pro under the control of the Ubi promoter; treatment 6 represents transformation with the same constructs as callus 2 and additionally with SrMV HC-Pro-delta N 12 under control of the Ubi promoter; and treatment 7 represents transformation with the same constructs as callus 2 and additionally with DNA that produces double stranded GFP mRNA under control of the 35Sint promoter;
[0053] FIG. 21 depicts 4 embryonic calli transiently transformed with constructs including, in callus 1, GUS under control of the 35Sint promoter; in callus 2, GUS under control of the 35Sint promoter and DNA that produces double stranded GUS mRNA also under control of the 35Sint promoter; in callus 3, the same constructs as callus 2 and additionally RNase H-like protein under control of the 35Sint promoter; in callus 4, the same constructs as callus 2 and additionally DNA that produces RNase H-like protein mRNA under control of the 35Sint promoter; although the figures is not presented in color, the darker colored calli, calli 1 and 4 are blue while the lighter colored calli, calli 2 and 3 are pinkish tan was will apparent to one skilled in the art.
DETAILED DESCRIPTION OF EXAMPLE EMBODIMENTS OF THE INVENTION
[0054] The present invention includes a novel method for determination of plant proteins active in PTGS or suppressive of PTGS. The invention may be used in both monocot and dicot plants, including sugarcane.
[0055] The methods of the present invention may be used, inter alia: (i) to identify, isolate and characterize cellular proteins that interact with these PTGS suppressive viral proteins or plant proteins involved in PTGS using a yeast two-hybrid system; and (ii) to evaluate these plant/viral protein interactions in vitro under physiological conditions and in vivo in either transient studies or in transgenic plants, such as sugarcane, expressing these viral proteins. Because many viral proteins such as P1 and HC-Pro are multifunctional proteins involved in various aspects of plant/potyvirus interactions, the methods may also be used to assess the role of interacting host proteins in gene silencing. The methods may be carried out using a combination of molecular genetics, immunological studies, transient antisense suppression studies, and plant transformation.
[0056] The overall method includes using a yeast two-hybrid system to search for plant proteins that interact with viral suppressors of PTGS to identify proteins involved in PTGS. These proteins or proteins identified as having a role in PTGS through other methods may then serve as bait to locate other proteins involved in PTGS. Because proteins that interact with other proteins involved in PTGS may be either suppressive of or active in PTGS, the yeast two-hybird assays may identify both types of proteins. Whether the proteins involved in PTGS are active in or suppressive of PTGS may be determined through a variety of methods, including identification of motifs with particular functions, comparison with known proteins, and in planta studies.
[0057] In the present invention the bait protein may be either active in PTGS or may function as a suppressor. The prey used is derived from an expression library of the plant of interest. In an exemplary embodiment of the invention, the prey library is derived from a plant in which PTGS is occurring. This facilitates identification of proteins involved in PTGS because such proteins are actively being produced in the silenced plant and may therefore be better represented in the mRNA pool of a silenced plant than in a non-silenced plant.
[0058] After identification of an interacting prey, the bait and prey portions of the two-hybrid screen of the present invention may be reversed to help identify false positives. Comparison with controls designed to look for activation of the reporter system absent either the biat or prey may also be used to identify false positives.
[0059] Such yeast two-hybrid screens may then be followed by assays such a farwestern blots or pull down assays to further determine whether identified proteins are false positives and also to characterize their function. In planta physiological conditions may be used in such assays.
[0060] In planta studies may also be conducted using transiently or permanently transformed embryonic calli transformed with either a test protein or a DNA encoding a double stranded mRNA of the test protein (which induces PTGS of that protein) in order to further evaluate the suppressive or effective role of the test protein in PTGS.
[0061] Proteins identified in one or more of the above screens may also be further characterized by identifying putative functional domains and also be searching for overall cDNA and amino acid sequence similarity with other proteins.
[0062] Often proteins and cDNAs identified using the above methodologies may be novel and patentable. In the present invention, five such novel proteins and cDNAs have been identified: the sugarcane 14-3-3 protein and cDNA, the RNase H-like protein and cDNA, the sugarcane LRR transmembrane protein kinase and cDNA, and the two sugarcane RING zinc finger proteins and cDNAs. Additionally, the SrMV P1/HC-Pro protein and cDNA used in some embodiments of the method of the present invention are also novel.
[0063] The above cDNAs as well as other identified using the methodologies of the present invention may also be used to construct transgenic plants, plant cells and plant tissues (collectively "plant entities") in which PTGS is either enhanced or suppressed. The methodologies used in the assay methods to generate embryonic calli and other methods know to the art may be used to construct these transgenic plant entities. Transformation may be transient or permanent, depending upon the intended use of the transgenic plant entity.
[0064] Permanently transformed plant entities with increased PTGS may be virus resistant. PTGS may be suppressed in other plants to allow increased expression of a transgene for any of a variety of reasons, including improvement of plant health, adaptation to certain growing conditions, producing of a novel nutrient or vaccine, and production of a protein later purified for medical or industrial uses. Finally, the PTGS-regulatory methods of the present invention may be used to induce PTGS of a particular gene, for instance by introducing a construct encoding dsRNA for a portion of the gene, thereby offering a novel method of producing knock-out plants.
[0065] The following examples are provided only to illustrate certain aspects of the invention and are not intended to embody the total scope of the invention or any aspect thereof. Variations of the exemplary embodiments of the invention below will be apparent to one skilled in the art and are intended to be included within the scope of the invention.
EXAMPLES
Example 1
Nucleotide Sequence of Sorghum Mosaic Virus
[0066] SrMV is a member of the genus Potyvirus and can cause mosaic disease and yield loss in poaceous plants such as sugarcane and sorghum (Shukla et al., 1994). A 2.0-kb region located at the 3' end of the SrMV strain H genomic RNA which encompasses the 3' untranslated region, the complete open reading frame for the coat protein and part of the NIb ORF has been previously sequenced (Yang and Mirkov, 1997). The remaining part of the SrMV genomic RNA has been sequenced in the present invention from overlapping RT-PCR products. From the combined sequences, a 9,581 bp consensus was derived that contains a continuous ORF encoding a 3079 amino acid putative polyprotein with ten gene products typical for members of the genus Potyvirus (Ingelbrecht et al., in preparation). The SrMV polyprotein consensus sequence was compared to that of Tobacco etch virus (TEV-HAT), Maize dwarf mosaic virus (MDMV-Bu), Plum pox virus (PPV-D) and Pea seedborne mosaic virus (PSbMV-D) in a multiple sequence alignment using Clustal X. Based on this alignment, putative proteolytic cleavage sites were positioned in the SrMV polyprotein. (See FIG. 1.) A novel nucleic acid (SEQ.ID.NO. 1) encoding a novel SrMV P1/HC-Pro protein (SEQ.ID.NO. 2) was developed.
Example 2
SrMV P1/HC-Pro Reverses PTGS
[0067] The potyviral HC-Pro is a multifunctional protein and harbors at least 3 functional domains. The N-terminal part contains a highly conserved KITC motif and is involved in aphid transmission (Thornbury et al., 1985; Atreya and Pirone, 1991). The central part is required for long distance movement and virus amplification (Cronin et al., 1995; Kasschau et al., 1997) while the C-terminal part represents a papain-like proteinase involved in polyprotein processing (Carrington et al., 1990). In addition, potyviral HC-Pro is the determinant of potyviral synergistic interactions with other viruses (Pruss et al., 1997).
[0068] Potyviral HC-Pro can reverse the effects of PTGS in tissues that were previously silenced. This suppression phenotype has been observed for the potyviral HC-Pro protein of three different potyviruses, TEV, PVY and PSbMV, suggesting that this particular function of potyviral HC-Pro may be conserved among different potyviruses (Voinnet et al., 1999). From this combined with the presence of conserved amino-acid sequence motifs in the SrMV HC-Pro, it was postulated that the SrMV HC-Pro might also act as a suppressor of PTGS in sugarcane. However, the suppressor ability of SrMV HC-Pro could not be assumed based on sequence similarity alone and experiments were required to determine if suppressor activity was exhibited in sugarcane and the extent of such suppression.
[0069] Reversal of PTGS by SrMV P1/HC-Pro was demonstrated by retransforming a plant which is posttranscriptionally silenced for the CP transgene (Ingelbrecht et al., 1999, plant #16). The silenced plant #16 had originally been generated by selection on bialaphos. Embryogenic callus generated from plant #16 was therefore retransformed with either nptII alone or with nptII and SrMV P1/HC-Pro, and transgenic plants were selected on geneticin. As shown in the Northern blot analysis of the CP gene in FIG. 2, in 5 of the 6 plants retransformed with nptII and SrMV P1/HC-Pro, PTGS has been suppressed and the steady state levels of the CP mRNA are equivalent to the levels seen in a plant that is not silenced (in FIG. 2, NS=plant #463 of Ingelbrecht et al., 1999). No such reversal was seen in plants retransformed with nptII alone. Southern and Northern analysis of nptII and SrMV P1/HC-Pro on these plants revealed that the one plant retransformed with nptII and SrMV P1/HC-Pro in which PTGS was not reversed was only transgenic for nptII (data not shown).
Example 3
The Yeast Two-Hybrid System
[0070] The yeast two-hybrid system has become a powerful tool for the study of protein-protein interactions in vivo. It can also be used to search an expression library for proteins that interact with a target protein. This latter application is being increasingly used with success in both animal (Kleiman and Manley, 1999) and plant systems (Kohalmi et al., 1998; Gindullis et al., 1999). Currently, high throughput two-hybrid procedures are being developed to catalogue protein-protein interactions on a genome-wide scale (Walkout et al., 2000).
[0071] Before embarking on a screen, it is useful to study the behavior of the target protein, termed bait, in the yeast cells. For example, one should demonstrate that the bait protein by itself does not activate transcription of the marker genes that are used in the selection and screening procedures. False positives can still be encountered following this precaution, but the great majority of these can be eliminated in subsequent steps. Finally, protein-protein interactions identified in the yeast cells may be verified via independent experimental procedures, preferably in the original biological system or a physiologically equivalent environment.
[0072] A yeast two-hybrid screening procedure typically consists of the following steps:
[0073] 1) Subclone the coding region of the target protein into a LexA or GAL4 binding domain vector to create a bait. In an example of the present method this bait is the SrMV HC-Pro sequence cloned into pHYBLexZeo to generate pIVING1281 or SrMV P1 cloned into pAS2 to generate pIVING1088-6.
[0074] 2) Transform the bait into the yeast reporter strain to verify that the expressed protein does not activate the reporter gene(s) by itself and is not toxic to the yeast cells. In an example of the present method the yeast strain L40 is transformed with either pIVING1168, pIVING1281, or pIVING1088-6 alone, and also co-transformed with either a pGAD424 derivative (pIVING1154) that lacks an insert, empty pHYBLexZeo, or pHYBLexZeo containing a non-specific bait (lamin). These sets of transformed yeast strains function as negative controls for the screening procedures as known in the art.
[0075] 3) Subclone a cDNA library of sequences into a GAL4 activator domain vector to create a library of prey. In an example of the present system, the cDNA library is derived from a transgenic sugarcane plant that displays PTGS (Ingelbrecht et al., 1999, plant #16) and is cloned into a modified pGAD424 derivative (pIVING1154), which constitutes the prey.
[0076] 4) Transform the yeast reporter strain carrying the bait plasmid with the cDNA/prey library.
[0077] 5) Screen yeast bait and prey co-transformed colonies for expression of a reporter gene that is fused to a GAL4 activated promoter. In an example of the present system, the yeast strain L40 contains reporter constructs for expression of a β-galactosidase gene, as well as a reporter construct for the HIS gene. Cells in which bait and prey fusion proteins interact in the yeast nucleus will grow on minimal media in the absence of histidine and will produce β-galactosidase enzyme activity at levels elevated from the negative control cells.
[0078] 6) Verify positive interactions of co-transformants and eliminate false positives by reestablishing and retesting yeast strains, testing yeast strains which only contain the previously identified prey construct and testing the prey construct with empty bait or bait that contains a non-specific protein (such as lamin). If the prey construct activates transcription of a reporter gene that is under regulation of a GAL4-inducible promoter under any of the above conditions, the prey may be considered a false positive.
[0079] This method, as used in the examples of the present system, may be used to isolate host proteins from sugarcane and other plants that are involved in PTGS using the HC-Pro and P1 proteins from SrMV or other proteins involved in PTGS as bait. Experiments using a library-scale yeast two-hybrid screen with SrMV HC-Pro as bait have identified 441 interacting proteins. At least 12 of these (including a protein with high similarity to a viral RNase H referred to herein as the "RNase H-like protein") have been confirmed to be true interactors. Additional proteins involved in PTGS, such as a novel sugarcane 14-3-3 protein, have also been identified using the above method based on their ability to interact with the RNase H-like protein used as bait.
Example 4
Self-Interaction with HC-Pro or P1 in a Yeast Two-Hybrid System
[0080] The yeast two-hybrid method of this invention may be used to identify proteins from sugarcane that interact with HC-Pro or P1 of SrMV. The two-hybrid procedure is based on the reconstruction of a functional transcriptional activator such as GAL4 or LexA, whose DNA binding domain (DBD) and transcription activation domain (AD) are expressed on two different vectors (Fields and Song, 1989). In the present example, the bait protein, i.e. the SrMV HC-Pro, was fused to the DNA-binding domain and the cDNA library was constructed in the activation domain vector, which produces the prey. These vectors were introduced into the yeast strain L40, which has an endogenous β-galactosidase (lacZ) reporter gene and the nutritional marker HIS3 for selection.
[0081] A 1.4-kb RT-PCR fragment encompassing the complete open reading frame of HC-Pro was amplified from SrMV virion particles and subcloned in pCR4-TOPO by T/A cloning (Invitrogen, Carlsbad, Calif.), yielding pIVING1148-1. The complete HC-Pro ORF was cloned as a fusion protein into the two-hybrid vectors pHYBLexZeo (bait) and pGAD424 (prey), yielding the plasmids pIVING1281 and pIVING1168, respectively. The P1 ORF was subcloned as a 0.78-kb fragment in pAS2 yielding pIVING1088-6. The vector-insert junctions were confirmed by sequencing for all constructs.
[0082] Recent studies have shown that potyviral HC-Pro proteins can interact with themselves in a yeast two-hybrid system (Urcuqui-Inchima et al., 1999; Guo et al., 1999), in agreement with an earlier proposal that the potyviral HC-Pro is biologically active as a homodimer (Thornbury et al., 1985). The ability of the SrMV HC-Pro for self-interaction was tested in the present example. It was also verified that the SrMV HC-Pro bait construct does not activate transcription of the lacZ gene, either by itself or in combination with empty bait or prey vectors. These latter experiments were also conducted for the P1 bait construct pIVING1088-6. As shown in FIG. 3, the SrMV HC-Pro does interact with itself (A) in the two-hybrid system demonstrating the functionality of this construct. Secondly, no lacZ expression can be observed in combination with either empty prey (B) or empty bait (D) vectors or a bait vector containing a non-specific protein (lamin; C). Therefore, SrMV HC-Pro does not activate transcription by itself and can be used as bait to screen for interacting proteins. Similar results were obtained for the SrMV P1 bait construct (data not shown).
Example 5
Development of a cDNA Library from Silenced Sugarcane
[0083] The development of SrMV-resistant sugarcane plants via transformation with an untranslatable form of the capsid protein sequence has been previously described and it has been demonstrated that the underlying resistance mechanism is related to PTGS (Ingelbrecht et al., 1999). As described in Ingelbrecht et al., 1999, plant #16 is a recovery plant that is immune to infection with SrMV after recovery from the initial infection and is posttranscriptionally silenced for the CP transgene. Although the CP transgenes are actively transcribed, the CP steady-state mRNA level is below the detection limit on an RNA gel blot (See Example 2 and FIG. 2).
[0084] Using poly(A)+ RNA from silenced leaf tissue of this plant a cDNA fusion library was constructed in the prey vector pIVING 1154 using the SMART® PCR cDNA library construction kit (offered by Clontech, Palo Alto Calif., a part of Beckton Dickinson, Franklin Lakes, N.J.). To create the vector p1VING1154, pGAD424 was modified to allow directional cloning of the cDNA into asymmetric SfiI sites. The library has an estimated 1.6×106 primary clones. As shown in FIG. 4, the inserts range between 0.5 to 2.0 kb with an average size of about 0.8 kb. Also, a 0.8-kb fragment containing the CP coding sequence could be readily amplified from the plasmid cDNA library indicating that low abundance mRNAs are represented.
Example 6
Library Scale Two-Hybrid Screen with HC-Pro Bait
[0085] The yeast strain L40 containing the SrMV HC-Pro bait construct was transformed with the prey cDNA library representing the equivalent of approximately 1.6×106 primary E. coli clones (see Example 4). About 77 million primary yeast transformants were plated on selective media (-leucine, -histidine+zeocin) and 776 yeast colonies were recovered by the fourth day of growth. Of these, 441 showed a range of light to heavy blue staining in the standard β-galactosidase colony-lift filter assay. To date, all of these double transformants have been colony purified, and all of the prey plasmids have been isolated. A RNase H-like protein, a RING zinc finger protein and LRR transmembrane protein kinase have been identified among the true positives. Some results of this screen and additional screens with SrMV P1 or RNase H like-protein as the bait are depicted in Table 1.
TABLE-US-00001 TABLE 1 Bait (DBD) Prey (AD) Interaction SrMV HC-Pro SrMV HC-Pro ++ SrMV P1 SrMV P1 -/+ SrMV HC-Pro SrMV P1 - SrMV P1 SrMV HC-Pro - SrMV HC-Pro RNase H-like prot. +++ RNase H-like prot. SrMV HC-Pro +++ SrMV P1 RNase H-like prot. - RNase H-like prot. RNase H-like prot. +++ A -/+ plus designates that very light blue staining was observed only after a 24 hour incubation. +++ designates that strong blue staining was seen in less than 30 minutes, and ++ designates strong blue staining in less than 3 hours. The RNase H-like protein self interaction may indicate that the protein functions as a homodimer.
Example 7
RNase H-Like Protein
[0086] One of the prey plasmids that was recovered during the experiments of Example 5 was retransformed into the SrMV HC-Pro bait yeast strain to reconfirm activation of the reporter genes. The plasmid encodes a protein with approximately 40% identity to a viral RNase H. This protein has been recovered several times using the methods of these examples. As shown in FIG. 5, the interaction with HC-Pro is very strong (D) and there is no interaction in combination with an empty bait plasmid or with a lamin bait plasmid (B and C, respectively). Furthermore, the RNase H-like gene is in the correct reading frame with respect to the fusion protein, so this is a true interactor. The cDNA sequence (SEQ.ID.NO. 3) for a nucleic acid encoding the RNase H-like protein and its amino acid sequence (SEQ.ID.NO. 4) are provided in FIG. 6.
Example 8
RING Zinc Finger Protein that Interacts with HC-Pro
[0087] Another protein identified as a true positive in HC-Pro interaction screens is a RING zinc finger protein. A partial cDNA sequence of a nucleic acid encoding this protein is provided in FIG. 7A (SEQ.ID.NO. 5). The encoded amino acid sequence is included as FIG. 7B (SEQ.ID.NO. 6). The nucleic acid sequence is sufficient to identify nucleic acids encoding the protein, but likely lacks approximately 300 bp at the 5' end in the figure. However, because a protein encoded by the possibly truncated nucleic acid sequence was identified in a two-hybrid screen, a nucleic acid with the sequence provided must encode a sufficient portion of the protein to allow interaction with SrMV HC-Pro. With the information disclosed in this invention, it is possible for a person skilled in the art to isolate the full length cDNA and protein sequence.
Example 9
LRR Transmembrane Protein Kinase
[0088] Another protein identified as a true positive in SrMV HC-Pro yeast two-hybrid screens is an LRR transmembrane protein kinase. A partial cDNA sequence of a nucleic acid encoding this protein is provided in FIG. 8A (SEQ.ID.NO. 7). The encoded amino acid sequence is included in FIG. 8B (SEQ.ID.NO. 8). The sequence is sufficient to identify nucleic acids encoding the protein, but likely lacks approximately 1 kb at the 5' end in the figure. However, because a protein encoded by the possibly truncated nucleic acid sequence was identified in a two-hybrid screen, a nucleic acid with the sequence provided must encode a sufficient portion of the protein to allow interaction with SrMV HC-Pro. With the information disclosed in this invention, it is possible for a person skilled in the art to isolate the full length cDNA and protein sequence.
Example 10
Library Scale Two-Hybrid Screen with Rnase H-Like Protein as Bait
[0089] A library scale two-hybrid screen similar to that of Example 5 was conduced using the same prey library (described in Example 4) with the RNase H-like protein as bait. Using the RNase H-like protein as bait, at least two true positives for prey/host proteins were identified and further characterized. The results of some of these assays as well as assays with SrMV HC-Pro and SrMV P1 as bait are depicted in Table 2.
TABLE-US-00002 TABLE 2 Bait (DBD) Prey (AD) Interaction SrMV HC-Pro SrMV HC-Pro ++ RNase H-like prot. SrMV HC-Pro +++ SrMV P1 RNase H-like prot. - RNase H-like prot. RNase H-like prot. +++ RNase H-like prot. Sugarcane 14-3-3 +++ SrMV HC-Pro Sugarcane 14-3-3 - SrMV P1 Sugarcane 14-3-3 - A -/+ designates that very light blue staining was observed only after a 24 hour incubation. +++ designates that strong blue staining was seen in less than 30 minutes, and ++ designates strong blue staining in less than 3 hours.
Example 11
Sugarcane 14-3-3 Protein
[0090] One protein identified through its ability to interact with the RNase H-like protein is a sugarcane 14-3-3 protein. The sequence of a nucleic acid encoding the sugarcane 14-3-3 protein is provided in FIG. 9A (SEQ.ID.NO. 9). The encoded amino acid sequence is provided in FIG. 9B (SEQ.ID.NO. 10). The role of another 14-3-3 protein in signal transduction in the dicot Arabidopsis has recently been described in Sehnke et al., 2002. A similar role in monocots is suggested by the disclosed in this invention. Specifically, 14-3-3 proteins exhibit a phosphorylation-dependent association with proteins in dicots. The RNase H-like protein of the present invention contains a serine with a 99.1% chance of being phosphorylated. This serine is also located in a good consensus 14-3-3 protein client-binding site. More specifically, the putative phosphorylation/binding site is VIQNpSPPDL (wherein "pS" designates phosphoserine) (SEQ. ID. NO. 13) beginning at amino acid 48 of the protein in FIG. 9B. Binding of sugarcane 14-3-3 and the RNase H-like protein in order to achieve a functional dimer (or as part of a functional complex containing other proteins) is also suggested by the absence of DNase or RNase activity of either protein in isolation. As shown in Table 2, 14-3-3 does not interact noticeably with SrMV HC-Pro or SrMV P1.
Example 12
RING Zinc Finger Protein that Interacts with RNase H-Like Protein
[0091] Another protein identified as a true positive in RNase H-like protein interaction screens is a RING zinc finger protein. This is not the same RING zinc finger protein identified as interacting with SrMV HC-Pro. A partial cDNA sequence of a nucleic acid encoding this protein is provided in FIG. 10A (SEQ.ID.NO. 11). The amino acid sequence encoded by the nucleic acid is provided in FIG. 10B (SEQ.ID.NO. 12)
Example 13
DNA Sequence Analyses of the Prey Genes Identified in the Two-Hybrid Screen
[0092] Prey sequences that have been verified by the above screening procedures may be DNA sequenced utilizing standard fluorescence-based thermocycle sequencing methods, and restriction maps created. This sequence information may be utilized to search the genetic databases with BLASTx and BLASTp to determine sequence similarity with known genes or proteins. In cases where similarity is clear, one may verify that the sequence under investigation is inserted in the appropriate reading frame in the prey vector. If the sequence is not in the appropriate reading frame, the prey can be considered a false positive.
Example 14
Characterization of the Prey Nucleic Acids Identified in the Two-Hybrid Screen for the Ability of their Corresponding Proteins to Interact with SrMV HC-Pro Protein or Other Bait Protein In Vitro Under Physiological Conditions
[0093] Although prey and bait combinations identified in the two-hybrid screen of the present invention may represent proteins which interact in yeast cells, several caveats of the assay may produce results which are not indicative of interactions that occur in planta. The yeast two-hybrid system assay requires that the bait and prey GAL4 fusion proteins both be imported into the yeast nucleus, which biochemically is a much more reducing environment than the yeast cytoplasm. This may lead to different patterns of protein folding than might otherwise occur for proteins whose operating environment is a more oxidative cytoplasm. Additionally, protein fragments placed in an unnatural position in a yeast two-hybird assay (e.g. an N term region in the C term of the fusion protein) may fail to fold or function properly because of positional effects.
[0094] Given that in vivo studies often require specialized reagents in the present method, an initial in vitro screening procedure in which potential protein interactions are verified under physiological conditions may be used to save time and expense associated with plant transformations in the case of false positives. However, it does remain possible that in vitro conditions may lack certain requirements for interaction found only in living cells and thus generate false negatives. Therefore, in some instances it may be desirable to proceed with in planta studies without or despite results in vitro. Such an approach is within the scope of the present invention.
[0095] If in vitro studies are preformed, host/prey proteins identified in the two-hybrid screen may be further tested for their ability to interact with SrMV P1 and SrMV HC-Pro or other proteins involved in PTGS in vitro under physiological conditions (i.e. pH 6.8 and 0.1 M ionic strength) as either polyhistidine, S-tag, or glutathione-s-transferase (GST) fusion-proteins. A series of vectors from Novagen, Inc. (Madison, Wis., affiliate of Merck KgaA, Darmstadt, Germany) including pET15b, pET29a,b,c and pET30a,b,c, allow the construction of N-terminal or C-terminal polyhistidine fusion proteins, which in the case of pET29 and pET30 derivatives may also contain S-tags. Pharmacia Corp. (Peapack, N.J.) also produces pGEX2, pGEX3 and pGEX4 derivatives in which GST fusion proteins may be produced. These vectors allow the specific purification of "tagged" proteins that have either polyhistidine tags (using Ni+-chelated resin), S-tags (using S-agarose), or GST-tags (using reduced glutathione-conjugated resins). Bait and prey inserts identified from the two-hybrid screen may be subcloned into one of these vectors. Other vectors encoding tag regions may also be used.
[0096] One approach is to construct two vector types, if possible, for the bait as well as for each prey that has been identified and to produce GST- and polyhistidine tagged proteins in E. coli. These tagged proteins may be purified with kits available from Pharmacia, Novagen, or other sources, or using methods known to the art and appropriate for the selected tag. His-S tagged SrMV HC-Pro produced in the pET30 vector purified with a Ni column is shown in FIG. 11 in lanes 3 and 4. As FIG. 11 indicates, tagged protein is produced only when an insert is present in the vector and may be largely purified using a Ni column.
[0097] Radiolabelled SrMV HC-Pro, SrMV P1, RNase H-like protein or other bait proteins of interest may be produced by translation of their corresponding transcripts in rabbit reticulocyte lysate. Using the TNS method, transcripts may be generated in a T7 in vitro transcription system or other system known to the art. Subsequent translation in the presence of S-35-labeled methionine or cysteine allows radiolabeling of the bait protein. FIG. 12 shows that RNase H-like protein produced using the above methodology is, in fact, radiolabeled. A small amount of this translated lysate may then be incubated with the tagged prey protein bound to its corresponding resin, and retention of radiolabeled bait protein may be assayed with SDS-PAGE gels and autoradiograms. Positive controls can include tagged bait protein, and negative controls can include the tags themselves, as well as the resins without any other proteins. These procedures or various modifications thereof readily determined by one skilled in the art allow one to determine whether prospective prey proteins are capable of interacting with either one or both viral proteins in vitro under physiological conditions.
Example 15
Farwestern Blots and Other Assays Indicate Interaction of Various Proteins
[0098] FIG. 13 shows a farwestern blot of His-S tagged SrMV HC-Pro probed with 35S labeled RNase H-like protein. Both proteins were produced according to the methods of the above examples. A comparison of binding of labeled RNase H-like protein to the His-S tagged SrMV HC-Pro to the various controls indicates that the interaction is specific between the two proteins and also that it occurs in conditions approximating plant physiological conditions.
[0099] FIG. 14 shows a farwestern blot of His-S tagged SrMV HC-Pro and RNase H-like protein and a plant extracts probed with 35S labeled RNase H-like protein. A comparison of the lanes indicates that the RNase H interacts with itself and SrMV HCPro and with other sugarcane proteins--from left to right--healthy sugarcane plant, SrMV infected sugarcane plant, sugarcane plant transgenic for SrMV P1/HC-Pro, sugarcane plant transgenic for SrMV delta N 12 HC-Pro. An extra band in the three plants expressing SrMV HC-Pro that is slightly smaller than the tagged SrMV HC-Pro may be seen in FIG. 14.
[0100] FIG. 15 shows the results of a Ni column pull down of His-S tagged protein and its binding partner. The proteins attached to the Ni column were removed and run on an SDS PAGE gel. The results indicate that His-S tagged SrMV HC-Pro and not control proteins pulled down labeled RNase H-like protein. Label controls were not pulled down. This indicates that the SrMV HC-Pro/RNase H-like protein interaction is specific and viable under physiological conditions.
[0101] FIG. 16 shows a farwestern blot of His-S tagged sugarcane 14-3-3 probed with 35S labeled RNase H-like protein. A comparison of binding of labeled RNase H-like protein to the His-S tagged sugarcane 14-3-3 to the various controls indicates that the interaction is specific between the two proteins and also that it occurs in conditions approximating physiological conditions.
Example 16
HC-Pro Deletion Mutant and Interaction with Rnase H-Like Protein
[0102] In order to better determine the region of SrMV HC-Pro responsible for interaction with the RNase H-like protein, a series of deletion mutants were created and their interaction was tested in yeast two-hybrid and farwestern blot assays as described above. The results of these experiments are summarized in Table 3 and indicate that the C terminal portion, including the last 10 kDa of the HC-Pro protein are required for interaction with the RNase H-like protein while the N terminal region up to 7 kDa is not necessary.
TABLE-US-00003 TABLE 3 HC-Pro segment Assay Interaction Full length Yeast two-hybrid +++ --------------→ Full length Farwestern +++ --------------→ N 1.3 kDa deletion Yeast two-hybrid +++ -------------→ N 7.0 kDa deletion Yeast two-hybrid +++ ---------→ C 10 kDa deletion Farwestern - ---------→ C 20 kDa deletion Farwestern - ----→ +++ in yeast two-hybrid assays indicates that strong blue staining was seen in less than 30 minutes. +++ in farwestern assays indicates that binding was apparent after a short exposure time.
Example 17
Complete Cloning of Partial cDNA Prey Clones
[0103] Candidate clones whose corresponding protein sequences interact with SrMV HC-Pro or other bait proteins in vitro under physiological conditions may be completely cloned with 5' RACE procedures or other procedures known to the art.
Example 18
Production of Antiserum
[0104] To facilitate in vivo co-immunoprecipitation studies further outlined below, antisera recognizing SrMV HC-Pro or another protein involved in PTGS may be produced. Using a pET or pGEX expression plasmid construct as described above or another 6xhis construct known to the art, one may produce 6Xhis-tagged SrMV HC-Pro or other protein in E. coli, purify the fusion protein as described above, and utilize the protein as an antigen with rabbits or other animals to produce polyclonal antisera. Similarly, SrMV P1 and host proteins may also be utilized to produce antisera, depending upon the potential utility of the resultant serum. Other antisera production techniques may also be used to produce polyclonal or, where useful, monoclonal antibodies.
Example 19
Transgenic Plant Studies
[0105] Because of its specific mode of action, the SrMV P1/HC-Pro and other proteins identified as involved in PTGS using the above two-hybrid and in vitro methods may target one or more factors that are expressed and functional in silenced tissue. This may be confirmed in planta. Because the cDNA library in the above examples was constructed from a plant harboring a PTGS-silenced SrMV coat protein sequence, and the plant is resistant to SrMV, one cannot readily utilize mechanical transmission of SrMV as a source of SrMV HC-Pro or other viral proteins in such plants, and thus may resort to transgenic methodologies.
[0106] Plant transformation studies demonstrate the feasibility of this approach. To date, SrMV HC-Pro transformants and SrMV P1/HC-Pro transformants have been confirmed via Southern and Northern analyses. The plants were produced via particle gun transformation of embryogenic callus, as previously described (Ingelbrecht et al., 1999). Three different plant transformation vectors have been constructed: (1) pIVING1023 which contains the SrMV P1/HC-Pro polyprotein sequence, in which SrMV HC-Pro may be expected to be cleaved out by the SrMV P1 protease in planta, (2) pIVING1002 which contains only the SrMV P1 sequence, and (3) pIVING991-1 which contains only the SrMV HC-Pro sequence. In the present example, two selectable markers for sugarcane transformation have been used, the nptII gene in combination with geneticin, and the bar gene for selection with bialaphos. Bialaphos selection may be used on bar transformed cells, and geneticin selection may be used on bar transgenic cells to select for second transformation events of previously transformed tissue. Other transformation mechanisms and selection systems known to the art may also be used. Such sequentially transformed plants may serve as a source of material for in planta studies.
[0107] In a further example, the transgenic plant #16 that was used as a source for construction of the prey cDNA library above, was utilized to generate embryogenic callus. Plant #16 was previously transformed with the bar gene, and in this example was transformed again with the SrMV P1-HC-Pro construct using geneticin selection. Because SrMV P1 has been described as an enhancer of the PTGS-inhibition by SrMV HC-Pro, the capsid message levels may be higher in SrMV P1-HC-Pro transformed #16 plants as compared to SrMV HC-Pro transformed #16 plants. In addition to having a system of modulated PTGS-suppression, these transgenic plants also allow verification in planta of the interactions between the viral proteins and the identified prey proteins, utilizing SrMV HC-Pro specific antiserum in standard co-immunoprecipitation methodologies.
[0108] Sequence homology of the isolated host proteins with known proteins, if present, may be used to postulate their biological function and whether or not they are involved in PTGS in a manner similar to that employed with the genes identified in mutagenesis studies of Neurospora, C. Elegans and Arabadopsis.
[0109] One approach for determining functions of the isolated genes may be based on creating suppression phenotypes for these genes in sugarcane via genetic transformation. Efficient triggering of gene suppression can be achieved by simultaneous expression of sense and antisense constructs or constructs designed to produce dsRNA as demonstrated in tobacco, rice and Arabadopsis (Waterhouse et al., 1998; Smith et al., 2000; Chuang and Meyerowitz, 2000). See also FIG. 17 for depictions of the basic structure of DNA and resulting mRNA for such constructs. Such constructs may be created for each of the isolated host genes and introduced in plants that display PTGS of a capsid protein transgene, such as plant #16. The transformation methodologies described above can be followed to generate stably transformed plants. Alternatively, a protoplast-based system can be used in combination with electroporation or PEG-mediated transformation. Expression of the capsid protein will be analyzed and reversal of silencing may indicate that the PTGS mechanism in these transgenics is impaired and that the transformed host genes are involved in PTGS.
Example 20
In Planta Studies of HC-Pro and Rnase H-Like Protein
[0110] Embryonic sugarcane calli were bombarded with various constructs according to the above examples in order to provide transient expression to verify the role of HN-Pro and RNase H-like protein in PTGS. Double stranded mRNA relating to the various proteins was generated using constructs such as those show in FIG. 17 inserted into the vector of FIG. 18.
[0111] FIG. 19 shows that GUS under a Ubi promoter alone is expressed in embryonic callus. Introduction of a double stranded GUS mRNA reduces its expression by activating PTGS of GUS mRNA. However expression is restored by both introducing SrMV P1-HC-Pro into the callus and by inducing PTGS of the RNase H-like protein using a double stranded mRNA of that protein. Average results for three separate experiments of this nature are depicted in FIG. 20 and show that statistically significant results are consistently obtained. Overall, these experiments confirm that SrMV P1 and SrMV HC-Pro inhibits PTGS and that the RNase H-like protein is involved in effecting PTGS.
[0112] FIG. 21 illustrates the PTGS effect of the RNase H-like protein even more clearly. In these figures calli transiently transformed with GUS exhibit the protein. This protein production is severely disrupted by subsequent or simultaneous transformation with DNA that produces double stranded GUS mRNA, which triggers PTGS. PTGS is not turned off by the transformation of the GUS/double stranded GUS calli with RNase H. However, it is severely hampered by introduction of DNA that produces double stranded mRNA for RNase H-like protein, which induces PTGS of RNase H-like protein itself. These experiments demonstrate that the RNase-H like protein plays a role in PTGS. These results are summarized in Table 4.
TABLE-US-00004 TABLE 4 Transformation Constructs Callus Color 35S-int/GUS Blue 35S-int/GUS + 35S-int/dsGUS Not Blue 35S-int/GUS + 35S-int/dsGUS + Not Blue 35S-int/RNase H-like protein 35S-int/GUS + 35S-int/dsGUS + Blue 35S-int/ds RNase H-like protein
[0113] All references mentioned in any portion of the foregoing specification are incorporated by reference herein.
Sequence CWU
1
1312103DNAPotyvirus Sorghum Mosaic Virus 1gaattcgccc ttatttcagc catggcagga
gcatggaaca ctgtgactta caagtggagg 60ccgaatttgg acaacgcaag agatgttcga
aaagtgatgg aacattttgc agcaaagcac 120caagtttatg atgcaaagcg tgcagcagag
cataacagta gaatccttcg caggactttt 180gtacaagaaa ttgcaaaagc acctgaagag
aagacttcat acaaacctca ggtgtgggtc 240gaaaaacaag ataacaatcc aacgatccat
ctacactatg ttagattcaa aaacaaggag 300aaaaagatat tacccgagat cactccagga
tcggtcgcaa agttaacgcg acggatactt 360gagctcagca aaactacaaa gcttgaagtt
gaactcattg gtaagaaaag acgcaaatcg 420actaaactag caatcaaaag gcgcagaaac
agagagtatt tgcattgtga aactagacac 480gaaacaaaca aatttaagcg cgttgacatc
aacatagaac gacactggtt tccacttgtg 540aagaagattt caaagtgcta tagtcacata
tcaccaagaa tgtacaaaaa catgagcaaa 600ggcgacagtg ggttaacatt catccagaat
ggtgagttat ttataatccg aggaaaacga 660gatggcgtcc tacttaatag tatcaccaat
gaaactcgaa ttaatgaaat aacttatttt 720agcgatgctc aggcgaacga cttctggcga
ggttacacag atcatatggt cgaaaatagg 780ttaatttcta caactcatac agaacacata
cccacaataa atttagagaa gtgtggaaag 840aggatggcat tgttagagat attgtttcac
tccacattca aaattacgtg taagcactgt 900aacaatgacg atcttgaact atcggatgat
gagtttggag aaagactata taagaactta 960atcagaattg aagaaaagca aaaagaatat
ttagctgaag atcaaaagct taagcgaatg 1020atatcctttc tgaaggatag atgcaatcca
aaatttgagc atttaccatt attatggcag 1080gtcgctgaaa caattggaca ttacactgat
aatcaagcaa aacagatcct tgaagttaat 1140gaagcgctca taaaagtgaa cactctttct
gttgaagatg cagtcaaagc tagcgcatcg 1200ttgctagaga tttcaagatg gtacaagaat
aggaaagaat catcgaaaga aggtacactt 1260agtacattca ggaataaaat ttcacctaaa
agtactatta atacagcact gatgtgtgat 1320aatcagctcg atacaaatgg taacttccta
tggggaaaga gagaatatca tgccaagcga 1380ttctttacaa actattttga agctgttgat
ccaaaagaca cgtatgaaaa gcatgttact 1440cggttcaatc caaatggtca acgcaaactt
tcgattggaa aactagttat cccattagac 1500ttccagaaga ttcgtgaatc atttataggt
gttcaagttc aaaaacaagc aattagtaga 1560gcgtgcttaa gtaaaatcga aaataattac
atataccctt gctgttgtgt aactacagaa 1620tttggtcaac cggtttattc agagatcatt
ccaccaacta aaggtcatat tactattgga 1680aattcgaccg acccaaaaat tgtggatttg
cctaattccg acccaccaat gatgtacata 1740gcgaaagatg gttattgtta tttgaatata
tttttagctg ctctgataaa cgtcaatgaa 1800gattcagcaa aagattacac aaagtttttg
cgtgatgaac taattgaaag acttggaaag 1860tggccaaaac tcaaagacgt ggcgacagca
tgttatgcat tatcagtaat gtttccagaa 1920attaagaacg cggagcttcc acaaatacta
gtggaccacg aacataaaac catgcatgtg 1980atagattcgt acggatctct cagtgttggc
ttccacatac tcaaagcgaa cacaatagga 2040caattaatca aaatgcaata tgaatccatg
gaaagtgaaa tgagagagta tgtagtcggt 2100tag
21032694PRTPotyvirus Sorghum Mosaic
VirusMISC_FEATURE(694)..(694)"Xaa" may be any amino acid or not present
2Met Ala Gly Ala Trp Asn Thr Val Thr Tyr Lys Trp Arg Pro Asn Leu1
5 10 15Asp Asn Ala Arg Asp Val
Arg Lys Val Met Glu His Phe Ala Ala Lys 20 25
30His Gln Val Tyr Asp Ala Lys Arg Ala Ala Glu His Asn
Ser Arg Ile 35 40 45Leu Arg Arg
Thr Phe Val Gln Glu Ile Ala Lys Ala Pro Glu Glu Lys 50
55 60Thr Ser Tyr Lys Pro Gln Val Trp Val Glu Lys Gln
Asp Asn Asn Pro65 70 75
80Thr Ile His Leu His Tyr Val Arg Phe Lys Asn Lys Glu Lys Lys Ile
85 90 95Leu Pro Glu Ile Thr Pro
Gly Ser Val Ala Lys Leu Thr Arg Arg Ile 100
105 110Leu Glu Leu Ser Lys Thr Thr Lys Leu Glu Val Glu
Leu Ile Gly Lys 115 120 125Lys Arg
Arg Lys Ser Thr Lys Leu Ala Ile Lys Arg Arg Arg Asn Arg 130
135 140Glu Tyr Leu His Cys Glu Thr Arg His Glu Thr
Asn Lys Phe Lys Arg145 150 155
160Val Asp Ile Asn Ile Glu Arg His Trp Phe Pro Leu Val Lys Lys Ile
165 170 175Ser Lys Cys Tyr
Ser His Ile Ser Pro Arg Met Tyr Lys Asn Met Ser 180
185 190Lys Gly Asp Ser Gly Leu Thr Phe Ile Gln Asn
Gly Glu Leu Phe Ile 195 200 205Ile
Arg Gly Lys Arg Asp Gly Val Leu Leu Asn Ser Ile Thr Asn Glu 210
215 220Thr Arg Ile Asn Glu Ile Thr Tyr Phe Ser
Asp Ala Gln Ala Asn Asp225 230 235
240Phe Trp Arg Gly Tyr Thr Asp His Met Val Glu Asn Arg Leu Ile
Ser 245 250 255Thr Thr His
Thr Glu His Ile Pro Thr Ile Asn Leu Glu Lys Cys Gly 260
265 270Lys Arg Met Ala Leu Leu Glu Ile Leu Phe
His Ser Thr Phe Lys Ile 275 280
285Thr Cys Lys His Cys Asn Asn Asp Asp Leu Glu Leu Ser Asp Asp Glu 290
295 300Phe Gly Glu Arg Leu Tyr Lys Asn
Leu Ile Arg Ile Glu Glu Lys Gln305 310
315 320Lys Glu Tyr Leu Ala Glu Asp Gln Lys Leu Lys Arg
Met Ile Ser Phe 325 330
335Leu Lys Asp Arg Cys Asn Pro Lys Phe Glu His Leu Pro Leu Leu Trp
340 345 350Gln Val Ala Glu Thr Ile
Gly His Tyr Thr Asp Asn Gln Ala Lys Gln 355 360
365Ile Leu Glu Val Asn Glu Ala Leu Ile Lys Val Asn Thr Leu
Ser Val 370 375 380Glu Asp Ala Val Lys
Ala Ser Ala Ser Leu Leu Glu Ile Ser Arg Trp385 390
395 400Tyr Lys Asn Arg Lys Glu Ser Ser Lys Glu
Gly Thr Leu Ser Thr Phe 405 410
415Arg Asn Lys Ile Ser Pro Lys Ser Thr Ile Asn Thr Ala Leu Met Cys
420 425 430Asp Asn Gln Leu Asp
Thr Asn Gly Asn Phe Leu Trp Gly Lys Arg Glu 435
440 445Tyr His Ala Lys Arg Phe Phe Thr Asn Tyr Phe Glu
Ala Val Asp Pro 450 455 460Lys Asp Thr
Tyr Glu Lys His Val Thr Arg Phe Asn Pro Asn Gly Gln465
470 475 480Arg Lys Leu Ser Ile Gly Lys
Leu Val Ile Pro Leu Asp Phe Gln Lys 485
490 495Ile Arg Glu Ser Phe Ile Gly Val Gln Val Gln Lys
Gln Ala Ile Ser 500 505 510Arg
Ala Cys Leu Ser Lys Ile Glu Asn Asn Tyr Ile Tyr Pro Cys Cys 515
520 525Cys Val Thr Thr Glu Phe Gly Gln Pro
Val Tyr Ser Glu Ile Ile Pro 530 535
540Pro Thr Lys Gly His Ile Thr Ile Gly Asn Ser Thr Asp Pro Lys Ile545
550 555 560Val Asp Leu Pro
Asn Ser Asp Pro Pro Met Met Tyr Ile Ala Lys Asp 565
570 575Gly Tyr Cys Tyr Leu Asn Ile Phe Leu Ala
Ala Leu Ile Asn Val Asn 580 585
590Glu Asp Ser Ala Lys Asp Tyr Thr Lys Phe Leu Arg Asp Glu Leu Ile
595 600 605Glu Arg Leu Gly Lys Trp Pro
Lys Leu Lys Asp Val Ala Thr Ala Cys 610 615
620Tyr Ala Leu Ser Val Met Phe Pro Glu Ile Lys Asn Ala Glu Leu
Pro625 630 635 640Gln Ile
Leu Val Asp His Glu His Lys Thr Met His Val Ile Asp Ser
645 650 655Tyr Gly Ser Leu Ser Val Gly
Phe His Ile Leu Lys Ala Asn Thr Ile 660 665
670Gly Gln Leu Ile Lys Met Gln Tyr Glu Ser Met Glu Ser Glu
Met Arg 675 680 685Glu Tyr Val Val
Gly Xaa 6903937DNASaccharum hybrid cultivar CP72-1210 3ggccattatg
gccggggaga acactgtatg aagaacatgg ccgcttctta tagtcaagac 60gcgcagctcg
atctcgtact tcctgacgct cccgtcgatg ctagcgcgtc tcgttctgaa 120cattcatctc
agcttgctag ctctaactgg agatctgtga ttcaaaactc cccacctgat 180ctcctatgcg
gatgcggtag accggcaatt aggcgcacgg cagagactgc gaagaacaat 240ggccgcatct
ttcgcacgtg tccggcgtgc aaaatatgga tttggcagga tctgctggac 300agctatgtga
atgctttgat aagctactgt cgtgatgcct ccattgattc ccttcagtca 360gagcttgaat
ctagccgttt attaatttcc gagaagcagg cacagatttc acgtttggag 420aaacaattgg
agacgctgca gccacttatc tcaaaatata ctgaacaatc tcgcagcatt 480gctcaggcct
ccattccatc atcacttttt ttcttggaag cctgcagtct ccgacatcag 540cggcggaggg
tgaatgaaaa tcaaggaggg gctaatttca aaagaagcta ctggagcgat 600taaagcttcg
gttaaaaata agcaagaatc taacacccag agcacaaatt ccatgagctg 660gctttttttt
gggacaccct ttcatttttc atcaaaaaag ggggggcacc cccagtttcc 720tccaaaaggc
tccccctgtc cgacatcata ggtgatgtga ttacccaaaa acaggttgtc 780ccgcttgctg
actcgatgcc aaatttggat tcaatgctgc tcctgttgtt ttaacaatca 840atcattttga
ctaaaagcat tccccttaaa attgttgtta aatttattgt caaacttatt 900accgcaaagt
ccgttggcag gtaatccccc ccttttt
9374200PRTSaccharum hybrid cultivar CP72-1210 4Gly His Tyr Gly Arg Gly
Glu His Cys Met Lys Asn Met Ala Ala Ser1 5
10 15Tyr Ser Gln Asp Ala Gln Leu Asp Leu Val Leu Pro
Asp Ala Pro Val 20 25 30Asp
Ala Ser Ala Ser Arg Ser Glu His Ser Ser Gln Leu Ala Ser Ser 35
40 45Asn Trp Arg Ser Val Ile Gln Asn Ser
Pro Pro Asp Leu Leu Cys Gly 50 55
60Cys Gly Arg Pro Ala Ile Arg Arg Thr Ala Glu Thr Ala Lys Asn Asn65
70 75 80Gly Arg Ile Phe Arg
Thr Cys Pro Ala Cys Lys Ile Trp Ile Trp Gln 85
90 95Asp Leu Leu Asp Ser Tyr Val Asn Ala Leu Ile
Ser Tyr Cys Arg Asp 100 105
110Ala Ser Ile Asp Ser Leu Gln Ser Glu Leu Glu Ser Ser Arg Leu Leu
115 120 125Ile Ser Glu Lys Gln Ala Gln
Ile Ser Arg Leu Glu Lys Gln Leu Glu 130 135
140Thr Leu Gln Pro Leu Ile Ser Lys Tyr Thr Glu Gln Ser Arg Ser
Ile145 150 155 160Ala Gln
Ala Ser Ile Pro Ser Ser Leu Phe Phe Leu Glu Ala Cys Ser
165 170 175Leu Arg His Gln Arg Arg Arg
Val Asn Glu Asn Gln Gly Gly Ala Asn 180 185
190 Phe Lys Arg Ser Tyr Trp Ser Asp 195
2005449DNASaccharum hybrid cultivar CP72-1210 5gaattcggcc attatggccg
gggcatgtgc agtgaatgcg ccaaggtcct gaggtaccaa 60accactcggt gccccatctg
caggcagcct gttgagcgtc tcctcgagat caaagtgagc 120aacaaatctg aagagcagca
gcagacgccc caatcgccgc cgctcccagc cccagctctg 180cagcaggaag aggtgtagcc
gtgattaaag tcagttctga gacattatat ggaactagtt 240tgcggccttc aggcctttcc
ctaaaggttt gttctctcat ctgagcaacg gggaatgtaa 300ccggtacttt acctttagcc
tatgtaagct tctggcatcg catggctttg ccgacctctg 360ctgtacctgc ttatctggag
gtcggagacc aagatgccaa ggaaagtgtg taccgtatat 420taaaaaaaaa aaaaaaaaaa
aaaaaaaaa 4496149PRTSaccharum hybrid
cultivar CP72-1210MISC_FEATURE(66)..(141)"Xaa" may be any amino acid or
not present 6Glu Phe Gly His Tyr Gly Arg Gly Met Cys Ser Glu Cys Ala Lys
Val1 5 10 15Leu Arg Tyr
Gln Thr Thr Arg Cys Pro Ile Cys Arg Gln Pro Val Glu 20
25 30Arg Leu Leu Glu Ile Lys Val Ser Asn Lys
Ser Glu Glu Gln Gln Gln 35 40
45Thr Pro Gln Ser Pro Pro Leu Pro Ala Pro Ala Leu Gln Gln Glu Glu 50
55 60Val Xaa Pro Xaa Leu Lys Ser Val Leu
Arg His Tyr Met Glu Leu Val65 70 75
80Cys Gly Leu Gln Ala Phe Pro Xaa Arg Phe Val Leu Ser Ser
Glu Gln 85 90 95Arg Gly
Met Xaa Pro Val Leu Tyr Leu Xaa Pro Met Xaa Ala Ser Gly 100
105 110Ile Ala Trp Leu Cys Arg Pro Leu Leu
Tyr Leu Leu Ile Trp Arg Ser 115 120
125Glu Thr Lys Met Pro Arg Lys Val Cys Thr Val Tyr Xaa Lys Lys Lys
130 135 140Lys Lys Lys Lys
Lys1457872DNASaccharum hybrid cultivar CP72-1210 7gaattcggcc attatggccg
ggaccaggaa ctccaggaat cagatgacaa ctcggggtat 60agagctcctg aagtgaccat
gtccggtcag tattctcaaa agagtgatgt ttacagcttt 120ggtgtcgtca tgcttgagct
actgactgga cagaaagcat ttgacagctc tcgggcaagg 180tcccagcaat cactagtccg
gtgggcttca ccgcagctgc acgacatcga ctcgctagat 240cagatggttg atccaacctt
agaggggctg taccatgcga aatcactctc tcggttcgca 300gacgcaatcg ctctctgtgt
ccagcctgaa ccagaattca ggccaccaat gtcggaggtc 360gtccagtcac tggtccgtct
tgtgcagcga gcaagcatgg ggacagcact aagcagcgag 420tggaattctt gccagttcga
tgaatctggt gatcacacgc tctaggggaa aatgatgtgt 480atttcctaga gagtctgatg
aggaactata gaaggctcac aagtcataga aacttgcagc 540ttggcattgt tgtgagttgt
gacggtgtga catgtgccag tgtcaggtga aatgtgactt 600ttacctatgc cattttactg
agagtctgct gcaacctgaa gtaggggtga aaagaaagtt 660ccttctttaa aaatatatat
ggttcattcg gacgtgtata tgaatatctt ttgaagacaa 720tcaactttct gatttcgtct
ctgatcgctg tccaaaaatt atcagggaag atgtagcact 780agtcctgcca cagaattagt
catctgtata tcctcagaaa tccgaaccat atccaggaaa 840catcaacaga ggacacgtcc
acatattcga ac 8728290PRTSaccharum hybrid
cultivar CP72-1210MISC_FEATURE(155)..(248)"Xaa" may be any amino acid or
not present 8Glu Phe Gly His Tyr Gly Arg Asp Gln Glu Leu Gln Glu Ser Asp
Asp1 5 10 15Asn Ser Gly
Tyr Arg Ala Pro Glu Val Thr Met Ser Gly Gln Tyr Ser 20
25 30Gln Lys Ser Asp Val Tyr Ser Phe Gly Val
Val Met Leu Glu Leu Leu 35 40
45Thr Gly Gln Lys Ala Phe Asp Ser Ser Arg Ala Arg Ser Gln Gln Ser 50
55 60Leu Val Arg Trp Ala Ser Pro Gln Leu
His Asp Ile Asp Ser Leu Asp65 70 75
80Gln Met Val Asp Pro Thr Leu Glu Gly Leu Tyr His Ala Lys
Ser Leu 85 90 95Ser Arg
Phe Ala Asp Ala Ile Ala Leu Cys Val Gln Pro Glu Pro Glu 100
105 110Phe Arg Pro Pro Met Ser Glu Val Val
Gln Ser Leu Val Arg Leu Val 115 120
125Gln Arg Ala Ser Met Gly Thr Ala Leu Ser Ser Glu Trp Asn Ser Cys
130 135 140Gln Phe Asp Glu Ser Gly Asp
His Thr Leu Xaa Gly Lys Met Met Cys145 150
155 160Ile Ser Xaa Arg Val Xaa Xaa Gly Thr Ile Glu Gly
Ser Gln Val Ile 165 170
175Glu Thr Cys Ser Leu Ala Leu Leu Xaa Val Val Thr Val Xaa His Val
180 185 190Pro Val Ser Gly Glu Met
Xaa Leu Leu Pro Met Pro Phe Tyr Xaa Glu 195 200
205Ser Ala Ala Thr Xaa Ser Arg Gly Glu Lys Lys Val Pro Ser
Leu Lys 210 215 220Ile Tyr Met Val His
Ser Asp Val Tyr Met Asn Ile Phe Xaa Arg Gln225 230
235 240Ser Thr Phe Xaa Phe Arg Leu Xaa Ser Leu
Ser Lys Asn Tyr Gln Gly 245 250
255Arg Cys Ser Thr Ser Pro Ala Thr Glu Leu Val Ile Cys Ile Ser Ser
260 265 270Glu Ile Arg Thr Ile
Ser Arg Lys His Gln Gln Arg Thr Arg Pro His 275
280 285Ile Arg 29091141DNASaccharum hybrid cultivar
CP72-1210 9gaattcggcc attatggccg gggaccgcag ttcccccgac cacaccgttc
cgccgcgcac 60agaggccagc cccgcgccag gagtaagttt gttcttttta acaatatgtc
gagggaggag 120aatgtttaca tggccaagct ggctgagcag gccgaaaggt atgaggagat
ggttgagtat 180atggagaagg tggctaagac tgtagatgtt gaagagctca ctgtggagga
gcgtaacctc 240ctgtctgtcg catacaagaa tgtgattggg gctcgccgtg cttcatggcg
cattgtctct 300tccattgaac agaaggagga gtcccgtaag aacgaagagc atgtgaacct
tatcaaggaa 360taccgcggga agattgaggc tgaactgagc aacatctgtg atggcatcct
gaaactgctt 420gactcccacc tagtgccttc ctctactgct gctgaatcaa aggtcttcta
cctcaagatg 480aagggtgact atcacaggta tcttgcggaa tttaagactg gtgctgagag
gaaggaatct 540gctgagagca caatggtagc ctacaaggct gctcaggaca ttgctctggc
tgagctggca 600cctacacatc cgataaggct tgggcttgct cttaacttct cagtgttcta
ttatgagatt 660ctgaactccc cagacaaagc ttgcaacctt gcaaagcagg cgtttgatga
agctatctct 720gagttagaca cccttgggga ggagtcatac aaagatagca ctctgatcat
gcagctcctg 780agggacaact tgaccctttg gacctctgac ctcacggagg atggtgctga
tgagggcaaa 840gaagcctcaa aaggtgatgc tggcgaggga cagtaatctt cggagagggc
atgttgttcc 900agcctggttt tagatgctct atgctgtcga agctgtgccg tgccattatt
gtagcagatt 960tcctctcccc ctcacttcat ttgcctcata ttagtaggct ggtagtggtc
gaattagttc 1020ccattgcttt gtgttgcagc tagttggcac taggtccgtg tggactggta
ttgttcccct 1080ggatttgaca agcatgtcct gtggtcgctc tagcgtttta ttgagctttg
aagcctcgat 1140t
114110380PRTSaccharum hybrid cultivar
CP72-1210MISC_FEATURE(292)..(377)"Xaa" may be any amino acid or not
present 10Glu Phe Gly His Tyr Gly Arg Gly Pro Gln Phe Pro Arg Pro His
Arg1 5 10 15Ser Ala Ala
His Arg Gly Gln Pro Arg Ala Arg Ser Lys Phe Val Leu 20
25 30Phe Asn Asn Met Ser Arg Glu Glu Asn Val
Tyr Met Ala Lys Leu Ala 35 40
45Glu Gln Ala Glu Arg Tyr Glu Glu Met Val Glu Tyr Met Glu Lys Val 50
55 60Ala Lys Thr Val Asp Val Glu Glu Leu
Thr Val Glu Glu Arg Asn Leu65 70 75
80Leu Ser Val Ala Tyr Lys Asn Val Ile Gly Ala Arg Arg Ala
Ser Trp 85 90 95Arg Ile
Val Ser Ser Ile Glu Gln Lys Glu Glu Ser Arg Lys Asn Glu 100
105 110Glu His Val Asn Leu Ile Lys Glu Tyr
Arg Gly Lys Ile Glu Ala Glu 115 120
125Leu Ser Asn Ile Cys Asp Gly Ile Leu Lys Leu Leu Asp Ser His Leu
130 135 140Val Pro Ser Ser Thr Ala Ala
Glu Ser Lys Val Phe Tyr Leu Lys Met145 150
155 160Lys Gly Asp Tyr His Arg Tyr Leu Ala Glu Phe Lys
Thr Gly Ala Glu 165 170
175Arg Lys Glu Ser Ala Glu Ser Thr Met Val Ala Tyr Lys Ala Ala Gln
180 185 190Asp Ile Ala Leu Ala Glu
Leu Ala Pro Thr His Pro Ile Arg Leu Gly 195 200
205Leu Ala Leu Asn Phe Ser Val Phe Tyr Tyr Glu Ile Leu Asn
Ser Pro 210 215 220Asp Lys Ala Cys Asn
Leu Ala Lys Gln Ala Phe Asp Glu Ala Ile Ser225 230
235 240Glu Leu Asp Thr Leu Gly Glu Glu Ser Tyr
Lys Asp Ser Thr Leu Ile 245 250
255Met Gln Leu Leu Arg Asp Asn Leu Thr Leu Trp Thr Ser Asp Leu Thr
260 265 270Glu Asp Gly Ala Asp
Glu Gly Lys Glu Ala Ser Lys Gly Asp Ala Gly 275
280 285Glu Gly Gln Xaa Ser Ser Glu Arg Ala Cys Cys Ser
Ser Leu Val Leu 290 295 300Asp Ala Leu
Cys Cys Arg Ser Cys Ala Val Pro Leu Leu Xaa Gln Ile305
310 315 320Ser Ser Pro Pro His Phe Ile
Cys Leu Ile Leu Val Gly Trp Xaa Trp 325
330 335Ser Asn Xaa Phe Pro Leu Leu Cys Val Ala Ala Ser
Trp His Xaa Val 340 345 350Arg
Val Asp Trp Tyr Cys Ser Pro Gly Phe Asp Lys His Val Leu Trp 355
360 365Ser Leu Xaa Arg Phe Ile Glu Leu Xaa
Ser Leu Asp 370 375 380
11829DNASaccharum hybrid cultivar CP72-1210 11gaattcggcc attatggccg
ggctgttcaa gctggaccgt tatatggtat gggacaccat 60ggatcttcca ccacaattgc
ttatggcggt gcatacttgc catattcttc ctcaactgga 120caatcgagca ataatcatca
agagcatgga tttcctgagc ggccagggca gcctgagtgt 180caatatttta tgaggactgg
aggttgcaaa tttggaacta tgtgtaaata taaccatcct 240cgagattgga gcactcctaa
gtccaactac atgttcagtc atctctgcct tccacttcgt 300ccgggtgctc agccttgtgc
gtactatgca caaaatggat attgcagata tggagttgca 360tgcaaatatg atcacccaat
gggtacacta ggctacagtt catctgcttt acccctatct 420gacatgccaa ttgctcccta
ccctatcggc ttctctgttg ccacgttggc tccatcttca 480tcttccccag aatatatttc
aaccaaagat ccatcaatca accaagtagc atcaccagtg 540cagcacccga acatgttgga
acaatcttgc caaaaggggt ttcccttcgg atccattatg 600cgaactcaac ttctacaagt
gtcggcagtt caagcctggg gggcgctgat tttctgactg 660ggggatgatc cttaacacaa
atttctatac ttgaacagtt tgaagccttc aaggaataaa 720aactggggcc ttgaaaaacc
gggaggggtt cttcccaaat aaaactgtgg tcaacactca 780tcctgaattg gtttcctatt
caaacggaag aggtttagga gtcacattg 82912276PRTSaccharum
hybrid cultivar CP72-1210MISC_FEATURE(219)..(262)"Xaa" may be any amino
acid or not present 12Glu Phe Gly His Tyr Gly Arg Ala Val Gln Ala Gly Pro
Leu Tyr Gly1 5 10 15Met
Gly His His Gly Ser Ser Thr Thr Ile Ala Tyr Gly Gly Ala Tyr 20
25 30Leu Pro Tyr Ser Ser Ser Thr Gly
Gln Ser Ser Asn Asn His Gln Glu 35 40
45His Gly Phe Pro Glu Arg Pro Gly Gln Pro Glu Cys Gln Tyr Phe Met
50 55 60Arg Thr Gly Gly Cys Lys Phe Gly
Thr Met Cys Lys Tyr Asn His Pro65 70 75
80Arg Asp Trp Ser Thr Pro Lys Ser Asn Tyr Met Phe Ser
His Leu Cys 85 90 95Leu
Pro Leu Arg Pro Gly Ala Gln Pro Cys Ala Tyr Tyr Ala Gln Asn
100 105 110Gly Tyr Cys Arg Tyr Gly Val
Ala Cys Lys Tyr Asp His Pro Met Gly 115 120
125Thr Leu Gly Tyr Ser Ser Ser Ala Leu Pro Leu Ser Asp Met Pro
Ile 130 135 140Ala Pro Tyr Pro Ile Gly
Phe Ser Val Ala Thr Leu Ala Pro Ser Ser145 150
155 160Ser Ser Pro Glu Tyr Ile Ser Thr Lys Asp Pro
Ser Ile Asn Gln Val 165 170
175Ala Ser Pro Val Gln His Pro Asn Met Leu Glu Gln Ser Cys Gln Lys
180 185 190Gly Phe Pro Phe Gly Ser
Ile Met Arg Thr Gln Leu Leu Gln Val Ser 195 200
205Ala Val Gln Ala Trp Gly Ala Leu Ile Phe Xaa Leu Gly Asp
Asp Pro 210 215 220Xaa His Lys Phe Leu
Tyr Leu Asn Ser Leu Lys Pro Ser Arg Asn Lys225 230
235 240Asn Trp Gly Leu Glu Lys Pro Gly Gly Val
Leu Pro Lys Xaa Asn Cys 245 250
255Gly Gln His Ser Ser Xaa Ile Gly Phe Leu Phe Lys Arg Lys Arg Phe
260 265 270Arg Ser His Ile
275139PRTSaccharum hybrid cultivar CP72-1210 13Val Ile Gln Asn Ser Pro
Pro Asp Leu1 5
User Contributions:
Comment about this patent or add new information about this topic:
 |  |
 |  |
 |  |
 |  |
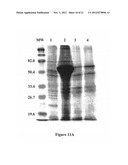 | 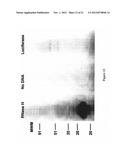 |
 | 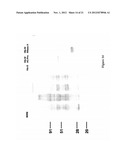 |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
| Similar patent applications: | |
| Date | Title |
|---|---|
| 2013-03-07 | Transgenic aloe plants for production of proteins and related methods |
| 2011-07-21 | Defensin polynucleotides and methods of use |
| 2013-03-07 | Screening of drug for attention deficit hyperactive disorder by using git1 knock-out mice as a novel adhd mouse model |
| 2013-01-31 | Autonomous lux reporter system and methods of use |
| 2013-01-31 | Plant transcriptional regulators of abiotic stress ii |
| New patent applications in this class: | |
| Date | Title |
|---|---|
| 2018-01-25 | Methods for plant transformation using spectinomycin selection |
| 2017-08-17 | Insecticidal polypeptides having broad spectrum activity and uses thereof |
| 2017-08-17 | Process for the prevention and suppression of bacterial diseases in plants |
| 2017-08-17 | Insecticidal polypeptides having broad spectrum activity and uses thereof |
| 2016-12-29 | Novel insecticidal proteins and methods of use |
| New patent applications from these inventors: | |
| Date | Title |
|---|---|
| 2017-05-18 | Methods, compositions, and systems for culturing and characterizing fastidious plant microbes |
| 2015-09-17 | Compositions, organisms, systems, and methods for expressing a gene product in plants |
| 2014-09-18 | Compositions, organisms, systems, and methods for altering cold, drought, and salt tolerance in plants |
| 2014-07-24 | Compositions, organisms, systems, and methods for expressing a gene product in plants |
| 2014-04-24 | Pathogen resistant citrus compositions, organisms, systems, and methods |
| Top Inventors for class "Multicellular living organisms and unmodified parts thereof and related processes" | |
| Rank | Inventor's name |
|---|---|
| 1 | Gregory J. Holland |
| 2 | William H. Eby |
| 3 | Richard G. Stelpflug |
| 4 | Laron L. Peters |
| 5 | Justin T. Mason |