Patent application title: LYSM RECEPTOR-LIKE KINASES TO IMPROVE PLANT DEFENSE RESPONSE AGAINST FUNGAL PATHOGENS
Inventors:
Jinrong Wan (Columbia, MO, US)
Gary Stacey (Columbia, MO, US)
Minviluz Stacey (Columbia, MO, US)
Xuecheng Zhang (Columbia, MO, US)
Assignees:
THE CURATORS OF THE UNIVERSITY OF MISSOURI
IPC8 Class: AC12N1582FI
USPC Class:
800279
Class name: Multicellular living organisms and unmodified parts thereof and related processes method of introducing a polynucleotide molecule into or rearrangement of genetic material within a plant or plant part the polynucleotide confers pathogen or pest resistance
Publication date: 2014-03-27
Patent application number: 20140090106
Abstract:
Perception of chitin fragments (chitooligosaccharides) is an important
first step in plant defense response against fungal pathogen. LysM
receptor-like kinases (LysM RLKs) are instrumental in this perception
process. LysM RLKs also play a role in activating transcription of
chitin-responsive genes (CRGs) in plants. Mutations in the LysM kinase
receptor genes or the downstream CRGs may affect the fungal
susceptibility of a plant. Mutations in LysM RLKs or transgenes carrying
the same may be beneficial in imparting resistance against fungal
pathogens.Claims:
1. A transgenic plant comprising a transgene, said transgene having
substantial sequence similarity to a LysM receptor kinase family gene.
2. The transgenic plant according to claim 1, wherein the LysM receptor kinase family gene is selected from the group consisting of polynucleotides of SEQ ID Nos. 1-2 and 4-6.
3. The transgenic plant according to claim 1, wherein the LysM receptor kinase family gene has a coding sequence that is at least 95% identical to the coding sequence of the polynucleotide of SEQ ID No. 6.
4. The transgenic plant according to claim 1, wherein the LysM receptor kinase family gene is selected from the group consisting of polynucleotides of SEQ ID Nos. 54-95.
5. The transgenic plant according to claim 1, wherein the plant is soybean.
6. The transgenic plant according to claim 1, wherein the plant is Arabidopsis thaliana.
7. The transgenic plant according to claim 1 wherein the LysM receptor kinase family gene encodes a functional LysM receptor kinase.
8. The transgenic plant according to claim 1, further comprising at least one regulatory sequence operably linked to said LysM receptor kinase family gene or fragment, said regulatory sequence controlling the expression level of the LysM receptor kinase family gene or fragment.
9. A plant comprising a LysM receptor kinase family gene having at least one mutation, said mutated gene being derived from an endogenous LysM receptor kinase family gene.
10. The plant of claim 9, wherein the mutated LysM receptor kinase family gene encodes a LysM receptor kinase with a mutation selected from the group consisting of amino acid substitution, deletion and insertion.
11. The plant of claim 9, wherein the mutation of the LysM receptor kinase family gene alters the expression level of the encoded LysM receptor kinase in said plant.
12. A method for protecting a plant from fungal infection, comprising the step of introducing into said plant a transgene, said transgene having substantial sequence similarity to a LysM receptor kinase family gene or fragment thereof.
13. The method of claim 12, further comprising the step of expressing a LysM receptor kinase encoded by said transgene.
14. The method of claim 12, wherein the LysM receptor kinase family gene is selected from the group consisting of polynucleotides of SEQ ID Nos. 1-2 and 4-6.
15. The method of claim 12, wherein the LysM receptor kinase family gene has a coding sequence that is at least 95% identical to the coding sequence of the polynucleotide of SEQ ID No. 6.
16. The method of claim 12, wherein the LysM receptor kinase family gene is selected from the group consisting of polynucleotides of SEQ ID Nos. 54-95.
17. A method for preventing fungal infection, comprising the step of generating the plant of claim 9.
18. The method according to claim 17 wherein the plant is soybean.
19. The method according to claim 17 wherein the plant is Arabidopsis thaliana.
20. The method of claim 17, wherein the LysM receptor kinase family gene is selected from the group consisting of polynucleotides of SEQ ID Nos. 1-2, 4-6 and 54-95.
Description:
[0001] This application is a divisional application of pending U.S. patent
application Ser. No. 13/347,199 filed Jan. 10, 2012, which is a
divisional application of U.S. patent application Ser. No. 11/835,328
filed Aug. 7, 2007, which claims priority to U.S. provisional patent
application Ser. No. 60/836,084 filed on Aug. 7, 2006. All of the
aforementioned applications are incorporated herein by reference into
this application.
SEQUENCE LISTING
[0003] This application is accompanied by a sequence listing that accurately reproduces the sequences described herein.
BACKGROUND
[0004] This disclosure relates to the use of molecular genetic technology involving LysM receptor kinase family genes and the expression or nonexpression thereof to modulate plant defense responses, especially against fungal pathogens.
[0005] Fungal disease causes significant agricultural losses in the United States and other parts of the world. Control of these pathogens is particularly difficult, often requiring treatment of entire fields with biocidal compounds. Although effective, increasing concern about the environmental and economic costs of such treatments require the need for alternative control methods.
[0006] Phakopsora pachyrhizi is a fungus that causes a rust disease of soybean (Glycine max), also known as Asian Soybean Rust. The pathogen has spread from Asia to all other soybean production regions in the world, and is reported to have arrived in the United States in the fall of 2004. At present, there is no known durable resistance available in any soybean varieties. Uromyces appendiculatus is a fungus that causes rust on bean (Phaseolus vulgaris). Breeders are working to identify genes in bean that can be manipulated for rust resistance. United States soybean producers have anticipated the arrival of P. pachyrhizi, the fungus that causes soybean rust, since its reported occurrence in Brazil. The arrival of P. pachyrhizi in the U.S. in late 2004 ended the anticipation, and farmers must now respond to the potential annual occurrence of this new disease. Farmer concerns have been based on reports of losses ranging from 10 to 80% in other regions of the world when control measures were not successfully implemented.
[0007] As rust-inducing fungi, U. appendiculatus and P. pachyrhizi belong to the order Uredinales, within the class Basidiomycetes. U. appendiculatus produces five spore stages on a single host plant. P. pachyrhizi reproduces predominantly by uredospores on a single host plant. Uredospores are responsible for rapid spread of the fungus. P. pachyrhizi can infect dozens of legume species, in addition to soybean.
[0008] Uredospores of U. appendiculatus penetrate through foliar stomatal openings. P. pachyrhizi differs in that germinated uredospores penetrate directly through the leaf epidermal cell layer. Typically, a uredospore that lands on a leaf surface germinates to produce an infection pad (appressorium) that adheres to the surface. In both species, the appressorium produces a hyphal peg that penetrates the plant. After penetration, each fungus develops thread-like structures (hyphae) that grow inter-cellularly through leaf tissues. The hyphae enter host cells without killing them. There, they produce spherical structures (haustoria) that extract nutrients from the living leaf cells. Soon after infection each fungus forms uredia that produce additional spores.
[0009] Soybean producers are particularly concerned because no durable, natural resistance to rust has been discovered after testing more than 18,000 soybean varieties. The pathogen, P. pachyrhizi can potentially infect any cultivar produced. In anticipation of the arrival of the rust pathogen, a great deal of research has been conducted to identify effective fungicides, and emergency governmental clearance for application to soybean has been obtained. Traditional screening and breeding methods have identified no major resistance genes to the aforementioned pathogens, and particularly in the case of U. appendiculatus and P. pachyrhizi.
[0010] Fungicides will likely be the front-line of defense against these and other fungal pathogens for many years until new resistance genes or other forms of resistance are identified. Fungicides have not traditionally been used in most soybean production. Consequently, there is limited information concerning the costs of this disease management practice and its likely economic viability. Widespread use of fungicides may also raise environmental concerns. These concerns have led to variable estimates of the acreage in Missouri and other states that may be shifted from soybean to alternative crops.
[0011] To protect soybean farmers and to ensure that soybean production meets increasing market demand, it is imperative that alternatives to fungicides be developed as rapidly as possible. Biotechnology-based approaches for defense against plant diseases are more preferable due to the minimal use of chemicals and the relative ease of deployment.
[0012] Chitin is a polymer of N-acetyl-D-glucosamine, found in fungal cell walls, insect exoskeletons, and crustacean shells. It has been hypothesized that plant chitinases can degrade chitin in the fungal cell walls to directly affect the viability of the invading fungal pathogen and to release short fragments (chitooligosaccharides) that can act as a general elicitor of plant innate immunity pathways. See e.g., Shibuya et al., 2001 and Stacey et al., 1997. In support of the above hypothesis, purified chitooligosaccharides have been shown to induce various defense responses in plants or cultured cells, such as induction of defense related genes and synthesis of phytoalexin. Shibuya et al., 2001 and Ramonell et al., 2005.
[0013] More particularly, chitooligosaccharides have been shown to induce a large number of genes (including many defense-related genes), and mutations in selected chitooligosaccharide-responsive genes or chitin-responsive genes (CRGs) have been shown to increase the susceptibility of a plant to certain fungal pathogens. See Ramonell et al., 2002; and Ramonell et al., 2005. Taken together, these studies suggest that plants possess a specific system to recognize chitooligosaccharides which, in turn, activate defense genes. See generally, Day et al., 2001; Zhang et al., 2002; Wan et al., 2004; Kaku et al., 2006; and Libault et al., 2007.
[0014] Previous work has reported chitin recognition in rice and legumes. Stacey, G. and N. Shibuya, Plant and Soil 194: 161-169 (1997) The ability of Arabidopsis thaliana to recognize and respond to chitin has also been reported. A variety of genes have been shown to respond to chitin treatment. See e.g., Ramonell et al. Microarray analysis of chitin elicitation in Arabidopsis thaliana. Mol. Plant Pathol. 3 (1): 301-311 (2002) and Zhang et al., Characterization of Early, Chitin-Induced Gene Expression in Arabidopsis Mol. Plant-Microbe Int. 15: 963-970 (2002) and Wan et al., Activation of a potential mitogen-activated protein kinase pathway in Arabidopsis by chitin. Mol. Plant Pathol. 5(1): 125-135 (2004).
[0015] More specifically, chitin binding sites or proteins have been previously identified in membrane preparations of a variety of plant cells. Day et al., 2001; Ito et al., 1997; and Okada et al, 2002. More recently, a LysM domain-containing protein (CEBiP) has been shown to be involved in the binding and recognition of chitooligosaccharides in rice. Kaku et al., 2006. The LysM motif was originally identified in bacterial enzymes that degrade cell wall component peptidoglycan, which is structurally similar to chitin. Joris, 1992. Since CEBiP lacks a significant intracellular domain, it likely functions as part of a chitin receptor complex. Kaku et al., 2006. However, no such chitin receptor complexes have been identified.
SUMMARY
[0016] The present disclosure overcomes the problems outlined above and advances the art by providing methods to confer fungal resistance to plants. This disclosure addresses a new biotechnology-based approach to generate rust resistant soybean and to confer rust resistance upon soybean plants. The technology also involves the development and/or deployment of defense peptides against fungal pathogen, such as the Asian soybean rust fungus. The technology similarly applies to other pathogens in plants, such as the field bean (Phaseolus vulgaris) rust pathogen, Uromyces appendiculatus.
[0017] It is hereby disclosed a number of LysM-containing receptor like kinases ("LysM RLKs") in soybean, as well as in other legume or non-legume plants. The lysine motif (LysM) domain is an ancient and ubiquitous protein module that binds peptidoglycan and structurally related molecules. A genomic survey in a large number of species spanning all kingdoms reveals that the combination of LysM and receptor kinase domains is present exclusively in plants. Table 1 lists a number of genes encoding LysM containing proteins from both prokaryotes and eukaryotes, along with their accession numbers from GenBank or other databases.
TABLE-US-00001 TABLE 1 LysM family genes in prokaryotes and eukaryotes LysM motif kingdom domain species name sources accession number >AGRT52b Bacteria Proteobacteria alpha- Argobacterium_tumefaciens_C58 UniProt/TrEMBL Q8UEQ5 proteobacteria >ANASP1 Bacteria Cyanobacteria Nostoc_PCC UniProt/TrEMBL Q8YRU0 >BACAN1a Bacteria Firmicutes Bacillus_anthracis UniProt/TrEMBL Q81WS5 >BACAN2b Bacteria Firmicutes Bacillus_anthracis UniProt/TrEMBL Q81Y89 >BACAN7 Bacteria Firmicutes Bacillus_anthracis UniProt/TrEMBL Q81SZ3 >BORPE5 Bacteria Proteobacteria Beta- bordetella_pertussis UniProt/TrEMBL Q7W0R5 proteobacteria >BORPE6 Bacteria Proteobacteria Beta- bordetella_pertussis UniProt/TrEMBL Q7VY72 proteobacteria >BRAJA1 Bacteria Proteobacteria alpha- Bradyrhizobium_japonicum UniProt/TrEMBL Q89Y08 proteobacteria >BRAJA2 Bacteria Proteobacteria alpha- Bradyrhizobium_japonicum UniProt/TrEMBL Q89XF2 proteobacteria >BURPS1 Bacteria Proteobacteria Beta- Burkholderia_pasudomallei_1710b UniProt/TrEMBL Q63LR7 proteobacteria >BURPS4 Bacteria Proteobacteria Beta- Burkholderia_pasudomallei_1710b UniProt/TrEMBL Q63TI4 proteobacteria >BURPS6a Bacteria Proteobacteria Beta- Burkholderia_pasudomallei_1710b UniProt/TrEMBL Q63V96 proteobacteria >CHLAU2 Bacteria Chloroflexi Chloroflexus_aurantiacus UniProt/TrEMBL Q3E5J5 >ECOLI6 Bacteria Proteobacteria Gama- Escherichia_coli UniProt/TrEMBL P75954 proteobacteria >PELCD5a Bacteria Proteobacteria Delta- Pelobacter_carbinolicus_DSM UniProt/TrEMBL Q3A2X4 proteobacteria >RALSO3 Bacteria Proteobacteria Beta- Ralstonia_solanacearum UniProt/TrEMBL Q8Y0H0 proteobacteria >RALSO6 Bacteria Proteobacteria Beta- Ralstonia_solanacearum UniProt/TrEMBL Q8XZ88 proteobacteria >RHOPA4 Bacteria Proteobacteria Alpha- Rhodopseudomonas_palustris UniProt/TrEMBL Q379H8 proteobacteria >SALCH2 Bacteria Proteobacteria Gama- Salmonella_choleraesuis UniProt/TrEMBL Q5J4C2 proteobacteria >SALCH5 Bacteria Proteobacteria Gama- Salmonella_choleraesuis UniProt/TrEMBL Q57QE0 proteobacteria >STRCO4 Bacteria Firmicutes Actinobacteridae Streptomyces_coelicolor UniProt/TrEMBL Q9ACX5 >VIBCH4 Bacteria Proteobacteria Gama- Vibrio_cholerae UniProt/TrEMBL Q9KNA7 proteobacteria >VIBCH5a Bacteria Proteobacteria Gama- Vibrio_cholerae UniProt/TrEMBL Q9KV14 proteobacteria >WOLSU1a Bacteria Proteobacteria Epsilon- wolinella_succinogenes UniProt/TrEMBL Q7M7V0 proteobacteria >WOLSU1b Bacteria Proteobacteria Epsilon- wolinella_succinogenes UniProt/TrEMBL Q7M7V0 proteobacteria >CAEEL1 Eukaryota Metazoa Nematoda Caenorhabditis_elegans UniProt/TrEMBL P90882 >CAEEL6 Eukaryota Metazoa Nematoda Caenorhabditis_elegans UniProt/TrEMBL Q93715 >CHLRE1 Eukaryota Chlorophyta Chlamydomonas_reinhardtii UniProt/TrEMBL Q9M5B9 >DICDI2 Eukaryota Mycetozoa Dictyosteliida Dictyostelium_discoideum UniProt/TrEMBL Q54BF7 >DROME10 Eukaryota Metazoa Chordata Drosophila_melanoqaster UniProt/TrEMBL Q9V4P7 >DROME9 Eukaryota Metazoa Chordata Drosophila_melanoqaster UniProt/TrEMBL Q9VNA1 >HUMAN1 Eukaryota Metazoa Chordata Homo_sapiens UniProt/TrEMBL Q5TF95 >HUMAN7 Eukaryota Metazoa Chordata Homo_sapiens UniProt/TrEMBL Q7Z3D4 >MOUSE5 Eukaryota Metazoa Chordata Mus_musculus UniProt/TrEMBL Q99LE3 >MOUSE9 Eukaryota Metazoa Chordata Mus_musculus UniProt/TrEMBL Q6DFV7 >XENLA5 Eukaryota Metazoa Chordata Xenopus_laevis UniProt/TrEMBL Q5BJ38 >AtLYK1b Eukaryota Viridiplantae Streptophyta Arabidopsis_thaliana TAIR At2g21630 >AtLYK1c Eukaryota Viridiplantae Streptophyta Arabidopsis_thaliana TAIR At2g21630 >AtLYK2 Eukaryota Viridiplantae Streptophyta Arabidopsis_thaliana TAIR At3g01840 >AtLYK3 Eukaryota Viridiplantae Streptophyta Arabidopsis_thaliana TAIR At1g51940 >AtLYK4a Eukaryota Viridiplantae Streptophyta Arabidopsis_thaliana TAIR At2g23770 >AtLYK4b Eukaryota Viridiplantae Streptophyta Arabidopsis_thaliana TAIR At2g23770 >AtLYK4c Eukaryota Viridiplantae Streptophyta Arabidopsis_thaliana TAIR At2g23770 >AtLYK5a Eukaryota Viridiplantae Streptophyta Arabidopsis_thaliana TAIR At2g33580 >AtLYK5c Eukaryota Viridiplantae Streptophyta Arabidopsis_thaliana TAIR At2g33580 >AtLYP1b Eukaryota Viridiplantae Streptophyta Arabidopsis_thaliana TAIR At1g21880 >AtLYP2a Eukaryota Viridiplantae Streptophyta Arabidopsis_thaliana TAIR At1g77630 >AtLYP3a Eukaryota Viridiplantae Streptophyta Arabidopsis_thaliana TAIR At2g17120 >AtLYP3b Eukaryota Viridiplantae Streptophyta Arabidopsis_thaliana TAIR At2g17120 >AtLysMe1 Eukaryota Viridiplantae Streptophyta Arabidopsis_thaliana TAIR At3g52790 >AtLysMe2 Eukaryota Viridiplantae Streptophyta Arabidopsis_thaliana TAIR At4g25433 >AtLysMe3 Eukaryota Viridiplantae Streptophyta Arabidopsis_thaliana TAIR At5g62150 >AtLysMn1 Eukaryota Viridiplantae Streptophyta Arabidopsis_thaliana TAIR At1g55000 >AtLysMn2 Eukaryota Viridiplantae Streptophyta Arabidopsis_thaliana TAIR At5g08200 >AtLysMn3 Eukaryota Viridiplantae Streptophyta Arabidopsis_thaliana TAIR At5g23130 >GmLYK10b Eukaryota Viridiplantae Streptophyta Glycine_max this study GmW2080D08.12 >GmLYK10c Eukaryota Viridiplantae Streptophyta Glycine_max this study GmW2080D08.12 >GmLYK11 Eukaryota Viridiplantae Streptophyta Glycine_max this study GmW2042I24.15 >GmLYK2 Eukaryota Viridiplantae Streptophyta Glycine_max this study GmW2098N11.15 >GmLYK4b Eukaryota Viridiplantae Streptophyta Glycine_max this study GmW2095P01.22 >GmLYK4c Eukaryota Viridiplantae Streptophyta Glycine_max this study GmW2095P01.22 >GmLYK8a Eukaryota Viridiplantae Streptophyta Glycine_max this study GmW2098N11.2 >GmLYK8b Eukaryota Viridiplantae Streptophyta Glycine_max this study GmW2098N11.2 >GmLYK9b Eukaryota Viridiplantae Streptophyta Glycine_max this study GmW2069O12.22 >GmLYK9c Eukaryota Viridiplantae Streptophyta Glycine_max this study GmW2069O12.22 >GmLYP1b Eukaryota Viridiplantae Streptophyta Glycine_max TIGR >GmLYP2a Eukaryota Viridiplantae Streptophyta Glycine_max TIGR >GmLYP2b Eukaryota Viridiplantae Streptophyta Glycine_max TIGR >GmLYP3a Eukaryota Viridiplantae Streptophyta Glycine_max TIGR >GmLYP3b Eukaryota Viridiplantae Streptophyta Glycine_max TIGR >GmLysMe1 Eukaryota Viridiplantae Streptophyta Glycine_max TIGR >GmLysMe2 Eukaryota Viridiplantae Streptophyta Glycine_max TIGR >GmLysMe3 Eukaryota Viridiplantae Streptophyta Glycine_max TIGR >GmLysMe4 Eukaryota Viridiplantae Streptophyta Glycine_max TIGR >GmLysMn1 Eukaryota Viridiplantae Streptophyta Glycine_max TIGR >GmLysMn2 Eukaryota Viridiplantae Streptophyta Glycine_max TIGR >GmLysMn3 Eukaryota Viridiplantae Streptophyta Glycine_max TIGR >GmNFR1ab Eukaryota Viridiplantae Streptophyta Glycine_max this study >GmNFR1ac Eukaryota Viridiplantae Streptophyta Glycine_max this study >GmNFR5aa Eukaryota Viridiplantae Streptophyta Glycine_max this study >GmNFR5ab Eukaryota Viridiplantae Streptophyta Glycine_max this study >GmNFR5ac Eukaryota Viridiplantae Streptophyta Glycine_max this study >MtLYK10a Eukaryota Viridiplantae Streptophyta Medicago_truncatula Medicago truncatula AC148994_13 sequencing resources >MtLYK10b Eukaryota Viridiplantae Streptophyta Medicago_truncatula Medicago truncatula AC148994_13 sequencing resources >MtLYK10c Eukaryota Viridiplantae Streptophyta Medicago_truncatula Medicago truncatula AC148994_13 sequencing resources >MtLYK12b Eukaryota Viridiplantae Streptophyta Medicago_truncatula Medicago truncatula AC126779_3 sequencing resources >MtLYK12c Eukaryota Viridiplantae Streptophyta Medicago_truncatula Medicago truncatula AC126779_3 sequencing resources >MtLYK13a Eukaryota Viridiplantae Streptophyta Medicago_truncatula Medicago truncatula AC126779_4 sequencing resources >MtLYK13b Eukaryota Viridiplantae Streptophyta Medicago_truncatula Medicago truncatula AC126779_4 sequencing resources >MtLYK3b Eukaryota Viridiplantae Streptophyta Medicago_truncatula Gene Bank AY372402 >MtLYK3c Eukaryota Viridiplantae Streptophyta Medicago_truncatula Gene Bank AY372402 >MtLYK9a Eukaryota Viridiplantae Streptophyta Medicago_truncatula Medicago truncatula AC148241_11 sequencing resources >MtLYK9b Eukaryota Viridiplantae Streptophyta Medicago_truncatula Medicago truncatula AC148241_11 sequencing resources >MtLYK9c Eukaryota Viridiplantae Streptophyta Medicago_truncatula Medicago truncatula AC148241_11 sequencing resources >OsLYK2b Eukaryota Viridiplantae Streptophyta Oryza_sativa TIGR LOC_Os06g41980 >OsLYK2c Eukaryota Viridiplantae Streptophyta Oryza_sativa TIGR LOC_Os06g41980 >OsLYK3 Eukaryota Viridiplantae Streptophyta Oryza_sativa TIGR LOC_Os06g41960 >OsLYK4b Eukaryota Viridiplantae Streptophyta Oryza_sativa TIGR LOC_Os02g09960 >OsLYK4c Eukaryota Viridiplantae Streptophyta Oryza_sativa TIGR LOC_Os02g09960 >OsLYK5a Eukaryota Viridiplantae Streptophyta Oryza_sativa TIGR LOC_Os03g13080 >OsLYK5c Eukaryota Viridiplantae Streptophyta Oryza_sativa TIGR LOC_Os03g13080 >OsLYK6a Eukaryota Viridiplantae Streptophyta Oryza_sativa TIGR LOC_Os11g35330 >OsLYK6b Eukaryota Viridiplantae Streptophyta Oryza_sativa TIGR LOC_Os11g35330 >OsLYK6c Eukaryota Viridiplantae Streptophyta Oryza_sativa TIGR LOC_Os11g35330 >OsLYP1b Eukaryota Viridiplantae Streptophyta Oryza_sativa TIGR LOC_Os03g04110 >OsLYP2a Eukaryota Viridiplantae Streptophyta Oryza_sativa TIGR LOC_Os09g37600 >OsLYP2b Eukaryota Viridiplantae Streptophyta Oryza_sativa TIGR LOC_Os09g37600 >OsLYP3a Eukaryota Viridiplantae Streptophyta Oryza_sativa TIGR LOC_Os06g10660 >OsLYP3b Eukaryota Viridiplantae Streptophyta Oryza_sativa TIGR LOC_Os06g10660 >OsLYP5b Eukaryota Viridiplantae Streptophyta Oryza_sativa TIGR LOC_Os02g53000 >OsLYP6a Eukaryota Viridiplantae Streptophyta Oryza_sativa TIGR LOC_Os11g34570 >OsLYP6b Eukaryota Viridiplantae Streptophyta Oryza_sativa TIGR LOC_Os11g34570 >OsLysMe1 Eukaryota Viridiplantae Streptophyta Oryza_sativa TIGR LOC_Os01g57390 >osLysMe2 Eukaryota Viridiplantae Streptophyta Oryza_sativa TIGR LOC_Os01g57400 >OsLysMe3 Eukaryota Viridiplantae Streptophyta Oryza_sativa TIGR LOC_Os04g48380 >OsLysMn1 Eukaryota Viridiplantae Streptophyta Oryza_sativa TIGR LOC_Os03g49250 >OsLysMn2 Eukaryota Viridiplantae Streptophyta Oryza_sativa TIGR LOC_Os06g51360 >PtLysMe4 Eukaryota Viridiplantae Streptophyta Populus_trichocarpa DOE JGI EUGENE3.00051310 >PtLysMe8 Eukaryota Viridiplantae Streptophyta Populus_trichocarpa DOE JGI EUGENE3.00070396 >PtLysMe9 Eukaryota Viridiplantae Streptophyta Populus_trichocarpa DOE JGI EUGENE3.00110096 >PtLysMe11 Eukaryota Viridiplantae Streptophyta Populus_trichocarpa DOE
JGI EUGENE3.00070285
[0018] In comparison to the LysM proteins in other kingdoms, plant LYK proteins possess unique features: (1) the combination of LysM and kinase domains exists exclusively in the plant lineage; (2) plant LYK proteins have no more than three LysM motifs; (3) if more than two LysM motifs exist within a single plant LYK protein, they are always distinct from each other at the protein sequence level; and (4) the LysM domain sequences in plant LYK proteins are highly diversified due to different combinations of heterogenous LysM motifs. Based on the sequence phylogenies, LysM motifs (named LYKa, LYKb, and LYKc from the N to the C terminus) in plant LYK proteins largely fall into five clades (FIG. 1A). This distribution of LysM motifs was found in all six plant species studied (i.e. LysM motifs from dicots and rice are clustered together in each Glade, suggesting that the diversification event of plant LysM motifs predated the divergence of monocot and dicot plants).
[0019] The LysM motifs from non-kinase plant LysM proteins have also been investigated. These sequences have been retrieved using BLAST searches against genomic sequence databases of Arabidopsis, rice, and poplar and EST sequences of soybean. Based on their subcellular localization predictions and domain arrangements, non-kinase plant LysM proteins may be further categorized into three subgroups, including LysM-type receptor-like proteins (LYPs), extracellular LysM proteins (LysMe), and nonsecretory intracellular LysM proteins (LysMn; FIG. 1B). This grouping will be helpful in understanding the nature of each LysM protein and providing insightful clues to the biological functions.
[0020] FIG. 1B illustrates the general domain structure of different LysM containing proteins. As shown in FIG. 1B, LysM RLKs (also referred to as "LYK") typically possess one or more LysM domains, a transmembrane domain and a kinase domain. The LysM domain is known for its capability to bind chitin. The transmembrane domain may serve to anchor the LysM RLKs in the membrane of the cells, whereas the kinase domain extends into the cytoplasm where it may phosphorylate specific substrates in the cell. In one embodiment, it is conceivable that the transmembrane domain of an LYK may be replaced with a different transmembrane domain from another LYK, or from another transmembrane protein,
[0021] As shown in FIG. 1 and FIG. 2, LysM domains in plants are highly diversified and that at least six distinct types of LysM motifs exist in plant LysM kinase proteins, which are shown as Types I-V and VII in FIG. 1B. Five additional types of LysM motifs exist in non-kinase plant LysM proteins, designated as Types VI, VIII-XI as shown in FIG. 1B. See also Zhang et al., Plant Physiol. 144, 623-636 (2007), which is hereby expressly incorporated by reference. FIG. 2 shows sequence alignment of representative LysM domains. FIG. 2A shows an alignment of 93 LysM-containing proteins in plants. Shaded areas indicate conserved residues in FIG. 2. FIG. 2B is an alignment of LysM domains from the LYK Group I, which contains LysM motif Types II and IV. FIG. 2c is an alignment of LysM domains from the LYK Group II, which contains LysM motif Types I, II and V. FIG. 2D is an alignment of LysM domains from the LYK Group III, which contains LysM motif Type VII. FIG. 2E is an alignment of LysM domains from the LYP group, which typically contains LysM motif Types VI and VII, or VI and VIII. See also FIG. 1B. FIG. 2F is an alignment of LysM domains from the LysMe group, which typically contains LysM motif Types IX or X. FIG. 2G is an alignment of LysM domains from the LysMn group, which typically contains LysM motif Type XI.
[0022] As predicted by Pfam, LYP proteins have exactly two LysM motifs and LysMe and LysMn proteins have only one LysM motif. Sequence alignments show that, among the 11 types of LysM motifs, motif sequences of LysMn (the motif within LysMn proteins, LysM motif type XI), one group of LysMe (the motif within LysMe proteins, LysM motif type X), and one group of LYPb (the second motif from the N terminus within LYP proteins, LysM motif VII) are extremely conserved. In these motifs, the amino acid identities averaged across the alignments are 91% for LysMe (type X), 86% for LysMn (type XI), and 75% for LYP (type VII). LysMn motif sequences always start with a His and end with a Pro. Similarly, LYPb motif sequences always end with a Pro. LYKa motifs are seven to 10 residues shorter.
[0023] In one aspect of this disclosure, soybean plants may be made resistant to soybean rust where no durable resistance is currently available. Certain soybean strains may be susceptible to rust diseases because they lack a functional signaling pathway that can perceive the existence of chitin or pass such a signal into the plant cell. Other strains may lack certain functional chitin responsive proteins and thus are not capable of mounting successful defenses against the invaders. It is hereby disclosed a methodology whereby a transgene encoding a component of the chitin signaling pathway may be introduced into a plant, such as a soybean plant. Expression of the transgene may enhance the perception of chitin by the transgenic plant, augment subsequent signaling that leads to gene activation inside the cells, and/or increase the capability of effector proteins in fighting the fungal pathogens.
[0024] In another aspect, mutations may be identified, induced or introduced into a plant, such as soybean, in order to obtain a mutant plant that have enhanced chitin perception or response. Certain mutations in LysM RLKs may enhance recognition of chitin by plant cells, whereas some other mutations in LysM RLKs may abolish chitin recognition. Similar situations may apply to other protein involved in fungal defense, including but not limited to the CRGs. The polynucleotide sequences disclosed herein may aid identification, mapping and/or genetic analysis of such mutants.
[0025] In one embodiment, a transgenic plant may be prepared by identifying in a plant one or more genes that contain a LysM receptor kinase family gene with a level of expression that is regulated by treatment of the plant with chitin. Such a plant may then be transformed with one or more LysM receptor kinase genes. The resulting transgenic plant may be challenged with chitin, its derivatives, or a pathogen to confirm an enhanced plant defense response, if desired.
[0026] The same general techniques disclosed herein may be employed in plants other than soybean to help create strains that are resistant to fungal infection. Different plants may be used to effect the transformation, such as soybean, Arabidopsis thaliana, or others. For instance, genes identified in one species may be introduced into another species in order to obtain transgenic plants that have enhanced fungal defense.
[0027] In another embodiment, a gene which belongs to the LysM receptor kinase family may be knocked out. Alternatively, a LysM receptor kinase family gene may be overexpressed. The resultant mutant plants may exhibit improved plant defense responses when challenged with chitin or its derivatives, for example, by spraying the leaves with chitin, or when challenged with a chitin-expressing organism.
[0028] In another embodiment, the LysM receptor kinase family gene may be any genes having a coding sequence that has 70%, 80%, 90%, 95%, 99%, or 100% sequence identity to any one of the polynucleotides of SEQ ID Nos. 1-7 and 54-95. In another embodiment, the LysM receptor kinase family gene may be any genes having a coding sequence that has 70%, 80%, 90%, 95%, 99%, or 100% sequence identity to the polynucleotides of SEQ ID No. 6 (AtLyk4).
BRIEF DESCRIPTION OF THE DRAWINGS
[0029] FIG. 1 illustrates the eleven distinct types of LysM motifs in plants. FIG. 1(A) The evolutionary relationships of plant LysM motifs. FIG. 1(B) Subcellular localization and LysM domain structures of LysM proteins in plants.
[0030] FIGS. 2A-2G show a sequence comparison of various plant LysM sequences that were obtained by computerized searching. FIG. 2A shows an alignment of 93 LysM-containing proteins: MtLYK10 (SEQ ID No. 75), MtLYK11 (SEQ ID No. 76), MtLysMe2 (SEQ ID No. 150; HtLYK1 (SEQ ID No. 71); MtLysHe1 (SEQ ID No. 151); AtLYK1 (SEQ ID No. 3); GmLYK2 (SEQ ID No. J6); HtLYK3 (SEQ ID No. 72); HtLYK4 (SEQ ID No. 73); HtLYK2 (SEQ ID No. 152); LjNFR1a (SEQ ID No. 1); PtLysHn8 (SEQ ID No. 153); PtLYK1 (SEQ ID No. 85); GmLYK8 (SEQ ID No. 63); PtLYK3 (SEQ ID No. 87); AtLYK5 (SEQ ID No. 5); GmLYK6 (SEQ ID No. 61); GmLYK4 (SEQ ID No. 58); HtLYK12 (SEQ ID No. 77); PtLYK6 (SEQ ID No. 90); PtLYK9 (SEQ ID No. 93); AtLYK4 (SEQ ID No. 6); OsLYK1 (SEQ ID No. 79); PtLYK5 (SEQ ID No. 89); GnLYK10 (SEQ ID No. 65); PtLYK7 (SEQ ID No. 91); AtLYK2 (SEQ ID No. 4); OsLYK2 (SEQ ID No. 80); PtLYP1 (SEQ ID No. 154); PtLYK4 (SEQ ID No. 88); PtLYK10 (SEQ ID No. 94); GmLYK9 (SEQ ID No. 64); GnNFR5a (SEQ ID No. 59); LjNFR5 (SEQ ID No. 2); PsSYH10 (SEQ ID No. 155); HtLYK13 (SEQ ID No. 78); HtLYK9 (SEQ ID No. 74); PtLYK2 (SEQ ID No. 86); OsLYK3 (SEQ ID No. 81); AtLYK3 (SEQ ID No. 5); PtLYK8 (SEQ ID No. 92); GnLYK11 (SEQ ID No. 66); PtLYP2 (SEQ ID No. 156); PtLYP3 (SEQ ID No. 157); PtLYP6 (SEQ ID No. 158); AtLYP3 (SEQ ID No. 159); GmLYP1 (SEQ ID No. 160); OsLYP1 (SEQ ID No. 161); PtLYP7 (SEQ ID No. 162); PtLYP5 (SEQ ID No. 163); PtLYP4 (SEQ ID No. 164); GmLYP4 (SEQ ID No. 165); GmLYP2 (SEQ ID No. 166); AtLYP2 (SEQ ID No. 167); AtLYP2 (SEQ ID No. 168); GmLYP3 (SEQ ID No. 169); GmLysHe1 (SEQ ID No. 170); MtLysH16 (171); PtLysHe7 (SEQ ID No. 172); GmLysHe2 (SEQ ID No. 173); GmLysHe4 (SEQ ID No. 174); PtLysHe6 (SEQ ID No. 175); GmLysHe3 (SEQ ID No. 176); GmLysHe6 (SEQ ID No. 177); GmLysHe5 (SEQ ID No. 178); PtLysHe3 (SEQ ID No. 179); PtLysHe5 (SEQ ID No. 180); PtLysMe10 (SEQ ID No. 181); AtLysHe3 (SEQ ID No. 182); OsLysHe4 (SEQ ID No. 183); AtLysHe2 (SEQ ID No. 184); AtLysHe1 (SEQ ID No. 185); PtLysHn9 (SEQ ID No. 186); PtLysHn7 (SEQ ID No. 187); PtLysHe8 (SEQ ID No. 188); PtLysMe11 (SEQ ID No. 189); PtLysHe4 (SEQ ID No. 190); GmLysHn2 (SEQ ID No. 191); GmLysHn4 (SEQ ID No. 192); PtLysHn6 (SEQ ID No. 193); OsLysHn3 (SEQ ID No. 194); GmLysHn3 (SEQ ID No. 195); GmLysHn5 (SEQ ID No. 196); AtLysHn2 (SEQ ID No. 197); PtLysMn11 (SEQ ID No. 198); AtLysHn3 (SEQ ID No. 199); GmLysHn1 (SEQ ID No. 200); AtLysHn1 (SEQ ID No. 201); PtLysHn2 (SEQ ID No. 202); PtLysHn1 (SEQ ID No. 203); OsLysHn1 (SEQ ID No. 204); PtLysMn10 (SEQ ID No. 205); and PtLysHe9 (SEQ ID No. 206). FIG. 2B is an alignment of LysM domains from the following proteins: MtLYK3 (SEQ ID No. 72), MtLYK4 (SEQ ID No. 73), MtLYK2 (SEQ ID No. 152), GmNFR1a (SEQ ID No. 54; GmNFR1b (SEQ ID No. 55); LjNFR1a (SEQ ID No. 1); MtLYK1 (SEQ ID No. 71); AtLYK1 (SEQ ID No. 3); GmLYK3 (SEQ ID No. 57); MtLYK3 (SEQ ID No. 72); MtLYK4 (SEQ ID No. 73); MtLYK2 (SEQ ID No. 152); GmNFR1a (SEQ ID No. 54); GmNFR1b (SEQ ID No. 55); LjNFR1a (SEQ ID No. 1); MtLYK1 (SEQ ID No. 71); AtLYK1 (SEQ ID No. 3); and GmLYK3 (SEQ ID No. 57). FIG. 2c is an alignment of LysM domains from GmNFR5a (SEQ ID No. 59); LjNFR5 (SEQ ID No. 2); MtLYK9 (SEQ ID No. 74); PtLYK2 (SEQ ID No. 86); GmLYK10 (SEQ ID No. 65); PtLYK7 (SEQ ID No. 91); AtLYK5 (SEQ ID No. 7); GmLYK8 (SEQ ID No. 63); MtLYK10 (SEQ ID No. 75); MtLYK11 (SEQ ID No. 76); LjLYK4 (SEQ ID No. 69); MtLYK12 (SEQ ID No. 77); GmLYK4 (SEQ ID No. 58); PtLYK6 (SEQ ID No. 90); PtLYK9 (SEQ ID No. 93); AtLYK4 (SEQ ID No. 6); PtLYK10 (SEQ ID No. 94); PtLYK4 (SEQ ID No. 88); GmLYK9 (SEQ ID No. 62); MtLYK13 (SEQ ID No. 78); PsSYM10 (SEQ ID No. 155); PtLYK11 (SEQ ID No. 95); GmNFR5b (SEQ ID No. 60); GmNFR5a (SEQ ID No. 59); LjNFR5 (SEQ ID No. 2); MtLYK9 (SEQ ID No. 74); PtLYK2 (SEQ ID No. 86); GmLYK10 (SEQ ID No. 65); PtLYK7 (SEQ ID No. 91); AtLYK5 (SEQ ID No. 7); GmLYK8 (SEQ ID No. 63); MtLYK10 (SEQ ID No. 75); MtLYK11 (SEQ ID No. 76); LyLYK4 (SEQ ID No. 69); MtLYK12 (SEQ ID No. 77); GmLYK4 (SEQ ID No. 58); PtLYK6 (SEQ ID No. 90); PtLYK9 (SEQ ID No. 93); AtLYK4 (SEQ ID No. 6); PtLYK10 (SEQ ID No. 94); PtLYK4 (SEQ ID No. 88); GmLYK9 (SEQ ID No. 64); MtLYK13 (SEQ ID No. 78); PsSYM10 (SEQ ID No. 155); PtLYK11 (SEQ ID No. 95); and GmNFR5b (SEQ ID No. 60). FIG. 2D is an alignment of LysM domains from GmLYK6 (SEQ ID No. 61); PtLYK3 (SEQ ID No. 87); OsLYK1 (SEQ ID No. 79); PtLYK5 (SEQ ID No. 89); AtLYK2 (SEQ ID No. 4); GmLKY11 (SEQ ID No. 66); PtLYK8 (SEQ ID No. 92); AtLYK3 (SEQ ID No. 5); OsLYK2 (SEQ ID No. 80); PtLYK1 (SEQ ID No. 85); OsLYK3 (SEQ ID No. 81). FIG. 2E is an alignment of LysM domains from AtLyP1 (SEQ ID No. 167); AtLYP2 (SEQ ID No. 168); PtLYP5 (SEQ ID No. 163); PtLYP7 (SEQ ID No. 162); PtLYP4 (SEQ ID No. 164); GmLYP4 (SEQ ID No. 165); GmLYP2 (SEQ ID No. 166); OsLYP3 (SEQ ID No. 207); OsLYP5 (SEQ ID No. 208); OsLYP4 (SEQ ID No. 209); GmLYP3 (SEQ ID No. 169); OsLYP1 (SEQ ID No. 161); OsLYP6 (SEQ ID No. 210); OsLYP2 (SEQ ID No. 211); PtLYP2 (SEQ ID No. 156); PtLYP3 (SEQ ID No. 157); AtLYP3 (SEQ ID No. 159); GmLYP1 (SEQ ID No. 160; PtLYP6 (SEQ ID No. 158); and PtLYP1 (SEQ ID No. 154). FIG. 2F is an alignment of LysM domains from GMLYSMe1 (SEQ ID No. 170); PtLysMe7 (SEQ ID No. 172); AtLysMe1 (SEQ ID No. 185); AtLysMe3 (SEQ ID No. 182); GmLysMe4 (SEQ ID No. 174); GmLysMe2 (SEQ ID No. 173); GmLysMe3 (SEQ ID No. 176); GmLysMe5 (SEQ ID No. 178); GmLysMe6 (SEQ ID No. 177); PtLysMe6 (SEQ ID No. 175); PtLysMe3 (SEQ ID No. 179); PtLysMe5 (SEQ ID No. 180); PtLysMe10 (SEQ ID No. 181); OsLysMe4 (SEQ ID No. 183); AtLysMe2 (SEQ ID No. 184); PtLysMe11 (SEQ ID No. 189); PtLysMe8 (SEQ ID No. 188); PtLysMe4 (SEQ ID No. 190); OsLysMe2 (SEQ ID No. 212); OsLysMe1 (SEQ ID No. 213); PtLysMe9 (SEQ ID No. 206); OsLysMe3 (SEQ ID No. 214). FIG. 2G is an alignment of LysM domains from AtLYSMn1 (SEQ ID No. 201); AtLysMn1 (SEQ ID No. 201); GmLysMn1 (SEQ ID No. 200); PtLysMn2 (SEQ ID No. 202); PtLysMn1 (SEQ ID No. 203); OsLysMn1 (SEQ ID No. 204); GmLysMn2 (SEQ ID No. 191); GmLysMn4 (SEQ ID No. 192); PtLysMn6 (SEQ ID No. 193); OsLysMn3 (SEQ ID No. 194); GmLysMn3 (SEQ ID No. 195); GmLysMn5 (SEQ ID No. 196); AtLysMn2 (SEQ ID No. 197); PtLysMn11 (SEQ ID No. 198); AtLysMn3 (SEQ ID No. 199); OsLysMn2 (SEQ ID No. 215); PtLysMn10 (SEQ ID No. 205); PtLysMn7 (SEQ ID No. 187); PtLysMn9 (SEQ ID No. 186); and PtLysMn8 (SEQ ID No. 153).
[0031] FIG. 3 presents experimental results showing enhanced expression of the defense genes PR-1 and PR-2 in the LysM receptor kinase mutant L3. FIG. 3A shows enhanced expression of the defense gene PR-2 in the LysM receptor kinase mutant L3. FIG. 3B shows enhanced expression of the defense gene PR-1 in the LysM receptor kinase mutant L3.
[0032] FIG. 4A shows an improved defense response of the L3 mutant to infection by the necotrophic fungus Botrytis cinerea.
[0033] FIG. 4B shows an improved defense response of the L3 mutant to infection by Pseudomonas syringae.
[0034] FIG. 5 shows a model of the involvement of the LysM receptor kinases in plant defense.
[0035] FIG. 6 shows that the knockout of the AtLysM RLK1 gene blocks the induction of the selected chitooligosaccharide-responsive genes (CRGs). FIG. 6A shows that induction of all selected CRGs is almost completely blocked in another insertion mutant, corresponding to At3g21630, or AtLYK1 (designated hereafter as AtLysM RLK1). FIG. 6B illustrates the intron-exon structure of AtLysM RLK1 (Square boxes represent exons. Solid lines between them are introns). FIG. 6C illustrates the predicted domain structure of AtLysM RLK1 (S: signal peptide; LysM: LysM domain; TM: transmembrane domain; Ser/Thr Kinase: Serine/Threonine kinase domain)
[0036] FIG. 7 shows disruption of the AtLysM RLK1 gene expression by the T-DNA insertions. WT=Wild-type Col-0; Mu=AtLysM RLK1 mutant. Actin-2 was used as an internal control. FIG. 7A shows RT-PCR results showing that primers corresponding to the exon regions on the side of the 10th intron failed to detect mRNA expression in the AtLysM RLK1 mutant. FIG. 7B shows that a truncated transcript derived from the gene sequence before the intron can be detected in the mutant by RT-PCR.
[0037] FIG. 8 shows restoration of CRGs in the AtLysM RLK1 mutant by the ectopic expression of the AtLysM RLK1 gene. Actin-2 serves as an internal control.
[0038] FIG. 9 shows the tissue expression pattern of the AtLysM RLK1 gene. Actin-2 serves as an internal control.
[0039] FIG. 10 shows that AtLysM RLK1 is induced by chitooligosaccharides, but not by the flagellin-derived flg22 peptide.
[0040] FIG. 11 shows that the T-DNA insertions in the AtLysM RLK1 gene block the induction of virtually all CRGs. FIG. 11A is a microarray read-out showing that a total of 909 genes responded more than 1.5 fold (P<0.05) to chitooctaose elicitation in both the wild-type and mutant plants 30 minutes after the treatment with chitooctaose. FIG. 11B shows the number of genes that are up-regualted in the wild-type and the mutant out of the 909 genes tested. FIG. 11C shows the number of genes that are down-regualted in the wild-type and the mutant out of the 909 genes tested.
[0041] FIG. 12 shows the functional categorization by annotations of 909 CRGs-GO Biological Process.
[0042] FIG. 13 shows that the AtLysM RLK1 mutant is more susceptible to fungal pathogens than wild-type plants and that exogenously applied chitooligosaccharides enhances resistance in the wild-type plants, but not in the mutant. FIG. 13A shows photographs of the infected leaves six days after inoculation. FIG. 13B shows trypan blue staining of the infected leaves indicating that the AtLysM RLK1 mutant supported more hyphal growth and production of conidiophores earlier than the wild-type plants (FIG. 13B). Arrows indicate sites where conidiophores are forming. FIG. 13c shows photographs of the leaves of the plants three days after inoculation, showing that the mutant developed slightly bigger lesions than the wild-type plants, as measured by average diameter of the lesions. FIG. 13D shows lesion size of the mutant as compared to that of wild-type plants. FIG. 13E shows that the mutant plants produced more spores per lesion than the wild-type plants. FIG. 13F shows that defense against bacterial infection was not affected by the mutation.
[0043] FIG. 14 shows that the selected CRGs are still induced in the AtLysM RLK1 mutant by a fungal pathogen, but to a reduced level. FIG. 14A shows that chitin-responsive defense gene MPK3 was induced by the fungal pathogen A. brassicicola in the mutant, albeit to a lower level as compared to that in wild-type plants. FIG. 14B shows that chitin-responsive defense gene WRKY53 was induced by the fungal pathogen A. brassicicola in the mutant, albeit to a lower level as compared to that in wild-type plants.
[0044] FIG. 15 shows that mutations in the legume Nod signal receptor genes NFR1 and NFR5 do not affect the induction of the selected CRGs in Lotus japonicus.
[0045] FIG. 16 shows that the mutation in the AtLysM RLK1 gene does not affect other defense-related pathways. FIG. 16A shows that both the mutant and wild-type plants showed similar induction of PR-1 by SA as shown by quantitative RT-PCR. FIG. 16B shows that both the mutant and wild-type plants showed similar induction of PDF1.2 by MeJA or ACC as shown by quantitative RT-PCR. FIG. 16C shows that the mutant plants were fully responsive to another typical PAMP, the flagellin-derived peptide flg22.
[0046] FIG. 17 shows that the AtLysM RLK1 mutation does not block the induction of flagellin-responsive genes.
[0047] FIG. 18 shows similarity between the LysM receptor kinase-like genes in a variety of plants.
[0048] FIG. 19 shows the expression analysis of GmLysM receptor-like kinases in response to white-mold pathogen.
[0049] FIG. 20 shows the expression analysis of GmLysM receptor-like kinases in chitin-treated leaves.
[0050] FIG. 21 shows the results of tissue-specific expression analysis of LysM receptor-like kinases in soybean, M. truncatula, and rice.
DETAILED DESCRIPTION
[0051] The following detailed description is provided to aid those skilled in the art in practicing the present instrumentalities. Even so, this detailed description should not be construed to unduly limit the present invention as modifications and variations in the embodiments discussed herein can be made by those of ordinary skill in the art without departing from the spirit or scope of the present inventive discovery.
[0052] LysM domain-containing receptor-like kinases ("LysM RLKs," "LYK" or "LysM receptor kinase family proteins"), such as NFR1 and NFR5 in legumes, have been shown to be critical for the perception of modified chitooligosaccharides--Nod signals--in the legume-rhizobial symbiotic nodulation process. See Limpens et al., 2003; Madsen et al., 2003; and Radutoiu et al., 2003. Similar LysM receptor kinase family genes (or LysM RLK genes) are also present in non-leguminous plants. Zhang et al., 2007. For example, five LysM RLK genes have been identified in the model plant Arabidopsis thaliana. LysM domain-containing proteins are also found in animals but LysM RLKs appear to be unique to plants. Zhang et al., 2007.
[0053] For purpose of this disclosure, a "LysM receptor kinase family gene" (LysM RLK gene) is any gene that encodes a protein with at least a Lysine motif (LysM) domain, a kinase domain and a transmembrane domain between the LysM and the kinase domain as shown in FIG. 1. In one aspect, the kinase domain is a serine/threonine kinase domain. A gene may be shown to belong to different subfamilies of LysM genes by sequence comparison with a known LysM gene, as exemplified by the sequence alignment of different LysM containing proteins in FIG. 2. This classification may be decided by a predetermined level of sequence identity in one or more functional domains of a known LysM receptor kinase, such as 70%, 80%, 90%, 95%, 96%, 97%, 98% 99% or 100% sequence identity with respect to a functional domain or an entire coding sequence.
[0054] Two sequences may be said to have "substantial sequence similarity" when they have a degree of sequence identity that persons of ordinary skill in the art may expect them to provide similar functionality; or in some cases, a person of skills in the art may expect individual domains within the sequences to possess similar functionality due to the sequence similarity. As measured by computer algorithms that are designed to quantitate sequence identity or similarity, this may be at least 70% identity, or more preferably, this may be any value from 90% and up, such as 95%, 96%, 97%, 98% or 99% identity, with 100% identity being an exact match. When two sequences are of different length, the sequence identity refers to cumulative sequence identity between those segments of both sequences that when aligned generate the best possible matches of individual residues throughout the full length of the sequences. In general, higher sequence similarity between two sequences indicates that the two sequences are more likely to perform similar, if not identical function.
[0055] The term CRGs (or CRG) refers to genes that may be activated upon perception of chitin or its derivatives by plant cells. Examples of CRGs may include MPK3, WRKY22, WRKY29, WRKY33, WRKY53 and any other genes whose expression levels may be up- or down-regulated when the cells are exposed to chitin or its derivatives. In one embodiment, genetic engineering may be used to render certain CRGs constitutively expressed, or, more preferably, expression of CRGs may be placed under control of certain regulatory elements such that CRG proteins may be expressed before fungi have been detected by the plant.
[0056] Under certain circumstances, it may be desirable to generate mutations in the coding sequence of certain genes, such as the LYK genes or CRGs. One of skills in the art may recognize that certain sequence variations may not significantly affect the functionality of a DNA or RNA molecule, or a protein. For instance, one may align and compare the sequences of the LYK family genes, as shown in FIG. 2, to determine which residues may be less conservative than others. The term "conservative" is used to depict those nucleotide or amino acid residues that have not undergone significant changes over the course of evolution. The conservative residues are typically shown as matching residues in a multi-sequence alignment. Guided by such an alignment, one may substitue those residues that are less conservative without compromising the functionality of the genes, RNAs, or proteins. Such manipulations of polynucleotide or polypeptide molecules are within the scope of this disclosure.
[0057] A "functional" LysM receptor kinase refers to a protein encoded by one of the LysM receptor kinase family genes that is capable of performing the full function that is typically performed by other LysM receptor kinase family proteins.
[0058] "Secretion sequence" means a sequence that directs newly synthesized secretory or membrane proteins to and through membranes of the endoplasmic reticulum, or from the cytoplasm to the periplasm across the inner membrane of bacteria, or from the matrix of mitochondria into the inner space, or from the stroma of chloroplasts into the thylakoid. Fusion of such a sequence to a gene that is to be expressed in a heterologous host ensures secretion of the recombinant protein from the host cell.
[0059] A "recombinant polynucleotide" means a polynucleotide that is free of one or both of the nucleotide sequences which flank the polynucleotide in the naturally-occurring genome of the organism from which the polynucleotide is derived. The term includes, for example, a polynucleotide or fragment thereof that is incorporated into a vector or expression cassette; into an autonomously replicating plasmid or virus; into the genomic DNA of a prokaryote or eukaryote; or that exists as a separate molecule independent of other polynucleotides. It also includes a recombinant polynucleotide that is part of a hybrid polynucleotide, for example, one encoding a polypeptide sequence.
[0060] "PCR" means polymerase chain reaction.
[0061] As used herein "polynucleotide" and "oligonucleotide" are used interchangeably and refer to a polymeric (2 or more monomers) form of nucleotides of any length, either ribonucleotides or deoxyribonucleotides. Although nucleotides are usually joined by phosphodiester linkages, the term also includes polymeric nucleotides containing neutral amide backbone linkages composed of aminoethyl glycine units. This term refers only to the primary structure of the molecule. Thus, this term includes double- and single-stranded DNA and RNA. It also includes known types of modifications, for example, labels, methylation, "caps", substitution of one or more of the naturally occurring nucleotides with an analog, internucleotide modifications such as, for example, those with uncharged linkages (e.g., methyl phosphonates, phosphotriesters, phosphoamidates, carbamates, etc.), those containing pendant moieties, such as, for example, proteins (including for e.g., nucleases, toxins, antibodies, signal peptides, poly-L-lysine, etc.), those with intercalators (e.g., acridine, psoralen, etc.), those containing chelators (e.g., metals, radioactive metals, boron, oxidative metals, etc.), those containing alkylators, those with modified linkages (e.g., alpha anomeric nucleic acids, etc.), as well as unmodified forms of the polynucleotide. Polynucleotides include both sense and antisense strands.
[0062] "Sequence" means the linear order in which monomers occur in a polymer, for example, the order of amino acids in a polypeptide or the order of nucleotides in a polynucleotide.
[0063] "Peptide," "Protein" and "Polypeptide" are used interchangeably and mean a compound that consists of two or more amino acids that are linked by means of peptide bonds.
[0064] "Recombinant protein" means that the protein, whether comprising a native or mutant primary amino acid sequence, is obtained by expression of a gene carried by a recombinant DNA molecule in a cell other than the cell in which that gene and/or protein is naturally found. In other words, the gene is heterologous to the host in which it is expressed. It should be noted that any alteration of a gene, including the addition of a polynucleotide encoding an affinity purification moiety, makes that gene unnatural for the purposes of this definition, and thus that gene cannot be "naturally" found in any cell.
[0065] A "non-immunoglobulin peptide" means a peptide which is not an immunoglobulin, a recognized region of an immunoglobulin, or contains a region of an immunoglobulin. For example, a single chain variable region of an immunoglobulin would be excluded from this definition.
[0066] "Substantially pure" or "substantially purified" means that the substance is free from other contaminating proteins, nucleic acids, and other biologicals derived from the original source organism. Purity may be assayed by standard methods, and will ordinarily be at least about 40% pure, more ordinarily at least about 50% pure, generally at least about 60% pure, more generally at least about 70% pure, often at least about 75% pure, more often at least about 80% pure, typically at least about 85% pure, more typically at least about 90% pure, preferably at least about 95% pure, more preferably at least about 98% pure, and in even more preferred embodiments, at least 99% pure. The analysis may be weight or molar percentages, evaluated, e.g., by gel staining, spectrophotometry, or terminus labeling etc.
[0067] A "transgene" refers to a coding sequence that has been introduced into a host organism, which may be referred to as a transgenic organism, e.g., a transgenic animal, or a transgenic plant, when the transgene has been successfully introduced into said organism. Typically, a transgene is introduced into a host organism so that the coding sequence may be transcribed into RNA or, in most cases, be further expressed as a polypeptide. The introduction of a transgene into a host organism and subsequent expression of the transgene is generally known as a transgenic process. A transgenic plant may refer to a whole plant, or a tissue of a plant, such as a seed, that contains a transgene and has a potential to be grow into a plant.
[0068] A "mutation," as used herein, means a change in the nucleotide sequence of a polynucleotide molecule or a change in the amino acid sequence of a polypeptide. The molecule carrying such a change may be referred to as a mutant molecule, and the change is typically measured by comparing the sequence of the mutant molecule with that of the polynucleotide or polypeptide molecule from which the mutant molecule is derived. An organism with a mutation in one of its endogenous genes may be called a mutant.
[0069] A mutation in a gene may occur on either the coding region or the non-coding region of the gene. A mutated copy of a gene may be said to be "derived" from a endogenous copy of the same gene when one or more spontaneous or induced mutations occur on an endogenous gene, at which point the endogenous gene becomes a mutated gene because it is no longer the same as the original wild-type gene. The resultant organism may be called a mutant.
[0070] In one aspect, the polynucleotide sequences disclosed herein, including but not limited to LysM receptor kinase family genes and various CRGs, may be used to identify those mutants with mutations in one of the LysM receptor kinase family genes. For instance, a large number of mutants may be generated by either spontaneous or induced mutation. These mutants may be screened to identify those mutations that occur in one of the LysM receptor kinase family genes. Suitable methods for such a screening may include, for example, PCR, sequencing, or hybridization.
[0071] In another aspect, these polynucleotide sequences, including but not limited to LysM receptor kinase family genes and various CRGs, may also be used to design DNA construct for targeted insertion, deletion, or substitution of a specific LysM receptor kinase family gene. For instance, a DNA fragment may be first inserted into the coding region of a LysM receptor kinase family gene, the construct thus obtained may then be introduced into a host to create an insertional mutant by homologous recombination.
[0072] The traits of a plant may be modified. A modified plant may be coconsidered "fungal resistant" if the chance of a plant becoming infected by a specific fungal pathogen is at least 30% less than that of a wild-type plant from which the modified plant is derived.
[0073] A molecule is "endogenous" to an organism if the molecule exists or is encoded by a molecule that exists in the organism without requiring a transgenic process. For purpose of this disclosure, the terms "expression" and "express" refer to transcription of DNA into RNA, or translation of RNA into protein, or both.
[0074] Besides null mutation that renders a protein completely non-functional, a mutation may also render a protein dominant negative when it only abolishes partial function of a protein. The resultant partially functioning protein may act as a "dominant negative" protein when it continues to perform the remaining function. For example, a mutated protein may continue to bind a co-factor without activating that co-factor because the activation function has been lost. When such a mutated protein is expressed in a cell, it binds to many co-factors without activating them, thus rendering them unavailable for the wild-type protein. In this situation, the mutant protein is said to be acting in a dominant negative fashion.
[0075] The term "knocking out" or "knock out" means rendering a gene non-functional. "Knocking down" means lowering the expression levels of a gene or decrease the relative activity of the encoded protein. A gene can be knocked out or knocked down through deletion, insertion, substitution of a fragment or a residue in the coding region or in the regulatory regions of a gene.
[0076] Within the scope of the disclosed instrumentalities are recombinant oligonucleotides encoding peptides having antifungal activity. These recombinant oligonucleotides can be used to produce recombinant polynucleotides which are commonly used as cloning or expression vectors although other uses are possible. A cloning vector is a self-replicating DNA molecule that serves to transfer a DNA segment into a host cell. The three most common types of cloning vectors are bacterial plasmids, phages, and other viruses. An expression vector is a cloning vector designed so that a coding sequence inserted at a particular site will be transcribed and translated into a protein.
[0077] Both cloning and expression vectors may contain nucleotide sequences that allow the vectors to replicate in one or more suitable host cells. In cloning vectors, this sequence is generally one that enables the vector to replicate independently of the host cell chromosomes, and also includes either origins of replication or autonomously replicating sequences. Various bacterial and viral origins of replication are well known to those skilled in the art and include, but are not limited to the pBR322 plasmid origin, the 2μ plasmid origin, and the SV40, polyoma, adenovirus, VSV and BPV viral origins. An expression vector may contain an origin of replication so that it can be replicated independently from the host's chromosome. More preferably, an expression vector carrying the transgene of interest may have a means by which the DNA fragment containing the transgene may be integrated onto a chromosome of the host plant and thus may be replicated along with the host chromosomes.
[0078] The polynucleotide sequences of the present disclosure may be used to produce antifungal peptides by the use of recombinant expression vectors containing the polynucleotide sequence disclosed herein. For purpose of this disclosure, antifungal peptides may mean polypeptides or fragments thereof that may help prevent fungal infection of a plant. Examples of antifungal peptides may include but not limited to polypeptides encoded by the LysM receptor kinase genes or the CRGs. In one embodiment, these antifungal peptide may be expressed in vitro and be applied onto a plant or be injected into a plant to achieve the desired antifungal effects. Suitable expression vectors include chromosomal, non-chromosomal and synthetic DNA sequences, for example, SV 40 derivatives; bacterial plasmids; phage DNA; baculovirus; yeast plasmids; vectors derived from combinations of plasmids and phage DNA; and viral DNA such as vaccinia, adenovirus, fowl pox virus, and pseudorabies. In addition, any other vector that is replicable and viable in the host may be used. Suitable host for in vitro expression may include bacterial, yeast, plant or insect cells, among others.
[0079] The nucleotide sequence of interest may be inserted into the vector by a variety of methods. In the most common method the sequence is inserted into an appropriate restriction endonuclease site(s) using procedures commonly known to those skilled in the art and detailed in, for example, Sambrook et al., Molecular Cloning, A Laboratory Manual, 2nd ed., Cold Spring Harbor Press, (1989) and Ausubel et al., Short Protocols in Molecular Biology, 2nd ed., John Wiley & Sons (1992).
[0080] In an expression vector, the sequence of interest may be operably linked to a suitable regulatory elements, including but not limited to a promoter or a enhancer, that may be recognized by the host cell to direct mRNA synthesis. Promoters generally refer to untranslated sequences located upstream from the start codon of a structural gene that regulate the transcription and translation of nucleic acid sequences under their control. Promoters may be classified as either inducible or constitutive promoters. Inducible promoters are promoters that initiate increased levels of transcription from DNA under their control in response to some change in the environment, e.g. the presence or absence of a nutrient or a change in temperature. Constitutive promoters, in contrast, maintain a relatively constant level of transcription.
[0081] A nucleic acid sequence is operably linked when it is placed into a functional relationship with another nucleic acid sequence. For example, DNA for a presequence or secretory leader is operatively linked to DNA for a polypeptide if it is expressed as a preprotein which participates in the secretion of the polypeptide; a promoter is operably linked to a coding sequence if it affects the transcription of the sequence; or a ribosome binding site is operably linked to a coding sequence if it is positioned so as to facilitate translation. Generally, operably linked sequences are contiguous and, in the case of a secretory leader, contiguous and in reading phase. Linking is achieved by ligation at restriction enzyme sites. If suitable restriction sites are not available, then synthetic oligonucleotide adapters or linkers can be used as is known to those skilled in the art. Sambrook et al., Molecular Cloning, A Laboratory Manual, 2nd ed., Cold Spring Harbor Press, (1989) and Ausubel et al., Short Protocols in Molecular Biology, 2nd ed., John Wiley & Sons (1992).
[0082] Common promoters used in expression vectors include, but are not limited to, LTR or SV40 promoter, the E. coli lac or trp promoters, and the phage lambda PL promoter. Useful inducible plant promoters include heat-shock promoters (Ou-Lee et al. (1986) Proc. Natl. Acad. Sci. USA 83: 6815; Ainley et al. (1990) Plant Mol. Biol. 14: 949), a nitrate-inducible promoter derived from the spinach nitrite reductase gene (Back et al. (1991) Plant Mol. Biol. 17: 9), hormone-inducible promoters (Yamaguchi-Shinozaki et al. (1990) Plant Mol. Biol. 15: 905; Kares et al. (1990) Plant Mol. Biol. 15: 905), and light-inducible promoters associated with the small subunit of RuBP carboxylase and LHCP gene families (Kuhlemeier et al. (1989) Plant Cell 1: 471; Feinbaum et al. (1991) Mol. Gen. Genet. 226: 449; Weisshaar et al. (1991) EMBO J. 10: 1777; Lam and Chua (1990) Science 248: 471; Castresana et al. (1988) EMBO J. 7: 1929; Schulze-Lefert et al. (1989) EMBO J. 8: 651). Other promoters known to control the expression of genes in prokaryotic or eukaryotic cells can be used and are known to those skilled in the art. Expression vectors may also contain a ribosome binding site for translation initiation, and a transcription terminator. The vector may also contain sequences useful for the amplification of gene expression.
[0083] Expression and cloning vectors can, and usually do, contain a selection gene or selection marker. Typically, this gene encodes a protein necessary for the survival or growth of the host cell transformed with the vector. Examples of suitable markers include dihydrofolate reductase (DHFR) or neomycin resistance for eukaryotic cells and tetracycline or ampicillin resistance for E. coli. Selection markers in plants include resistance to bleomycin, gentamycin, glyphosate, hygromycin, kanamycin, methotrexate, phleomycin, phosphinotricin, spectinomycin, streptomycin, sulfonamide and sulfonylureas. Maliga et al., Methods in Plant Molecular Biology, Cold Spring Harbor Press, 1995, p. 39.
[0084] In addition, expression vectors can also contain marker sequences operatively linked to a nucleotide sequence for a protein that encode an additional protein used as a marker. The result is a hybrid or fusion protein comprising two linked and different proteins. The marker protein can provide, for example, an immunological or enzymatic marker for the recombinant protein produced by the expression vector. Suitable markers include, but are not limited to, alkaline phosphatase (AP), myc, hemagglutinin (HA), β-glucuronidase (GUS), luciferase, and green fluorescent protein (GFP).
[0085] The polynucleotide sequences of the present disclosure may also be part of an expression cassette that at a minimum comprises, operably linked in the 5' to 3' direction, a regulatory sequence such as a promoter, a polynucleotide encoding a peptide of the present disclosure, and a transcriptional termination signal sequence functional in a host cell. The promoter can be of any of the types discussed herein, for example, a tissue specific promoter, a developmentally regulated promoter, an organelle specific promoter, a seed specific promoter, a plastid specific promoter, etc. The expression cassette can further comprise an operably linked targeting, transit, or secretion peptide coding region capable of directing transport of the protein produced. The expression cassette can also further comprise a nucleotide sequence encoding a selectable marker and/or a purification moiety.
[0086] More particularly, the present disclosure includes recombinant constructs comprising an isolated polynucleotide sequence encoding the antifungal peptides of the present disclosure. The constructs can include a vector, such as a plasmid or viral vector, into which the sequence has been inserted, either in the forward or reverse orientation. The recombinant construct can further comprise regulatory sequences, including, for example, a promoter operatively linked to the sequence. Large numbers of suitable vectors and promoters are known to those skilled in the art and are commercially available.
[0087] Different domains from different LysM RLKs may be combined to obtain chimeric proteins. Such a chimeric protein may possess a number of desirable properties that are not otherwise exhibited by one single protein that naturally exist in plants. Such desirable properties may include but are not limited to increased sensibility to chitin and its derivatives, increased kinase activity or enhanced kinase specificity.
[0088] A further embodiment of the present disclosure relates to transformed host cells containing constructs comprising the oligonucleotide sequences of the present disclosure. For instance, various combination of the LysM RLK genes, in wild-type or mutated forms, may be introduced as transgenes into a host plant, such as soybean. In a preferred embodiment, the host plants is susceptible to fungal infection and the expression of the LysM RLK transgenes may confer certain degree of fungal resistance to the host. In addition to the LysM RLK genes, the transgenes may include other genes that may play a role in fungal defense, such as any of the CRGs whose forced expression may enhance the host's capability to defend against fungal pathogens.
[0089] The host cell can be a higher eukaryotic cell, such as a mammalian or plant cell, or a lower eukaryotic cell such as a yeast cell, or the host can be a prokaryotic cell such as a bacterial cell. Introduction of the construct into the host cell can be accomplished by a variety of methods including calcium phosphate transfection, DEAE-dextran mediated transfection, Polybrene, protoplast fusion, liposomes, direct microinjection into the nuclei, scrape loading, and electroporation. In plants, a variety of different methods can be employed to introduce transformation/expression vectors into plant protoplasts, cells, callus tissue, leaf discs, meristems, etc., to generate transgenic plants. These methods include, for example, Agrobacterium-mediated transformation, particle gun delivery, microinjection, electroporation, polyethylene glycol-mediated protoplast transformation, liposome-mediated transformation, etc. (reviewed in Potrykus (1991) Annu. Rev. Plant Physiol. Plant Mol. Biol. 42: 205).
[0090] Peptides produced by expression of the polynucleotides of the present disclosure can be obtained by transforming a host cell by any of the previously described methods, growing the host cell under appropriate conditions, inducing expression of the polynucleotide and isolating the protein(s) of interest. If the protein in retained within the host cell, the protein can be obtained by lysis of the host cells, while if the protein is a secreted protein, it can be isolated from the culture medium. Several methods are available for purification of proteins and are known to those of ordinary skill in the art. These include precipitation by, for example, ammonium sulfate or ethanol precipitation, acid extraction, anion or cation exchange chromatography, phosphocellulose chromatography, hydrophobic interaction chromatography, affinity chromatography, hydroxylapatite chromatography, lectin chromatography, high performance liquid chromatography (HPLC), electrophoresis under native or denaturing conditions, isoelectric focusing, and immunoprecipitation.
[0091] Alternatively, peptides encoded by the polynucleotides of the present disclosure can be produced by chemical synthesis using either solid-phase peptide synthesis or by classical solution peptide synthesis also known as liquid-phase peptide synthesis. In oligomer-supported liquid phase synthesis, the growing product is attached to a large soluble polymeric group. The product from each step of the synthesis can then be separated from unreacted reactants based on the large difference in size between the relatively large polymer-attached product and the unreacted reactants. This permits reactions to take place in homogeneous solutions, and eliminates tedious purification steps associated with traditional liquid phase synthesis. Oligomer-supported liquid phase synthesis has also been adapted to automatic liquid phase synthesis of peptides.
[0092] For solid-phase peptide synthesis, the procedure entails the sequential assembly of the appropriate amino acids into a peptide of a desired sequence while the end of the growing peptide is linked to an insoluble support. Usually, the carboxyl terminus of the peptide is linked to a polymer from which it can be liberated upon treatment with a cleavage reagent. In a common method, an amino acid is bound to a resin particle, and the peptide generated in a stepwise manner by successive additions of protected amino acids to produce a chain of amino acids. Modifications of the technique described by Merrifield are commonly used (see, e.g., Merrifield, J. Am. Chem. Soc. 96: 2989-93, 1964). In an automated solid-phase method, peptides are synthesized by loading the carboxy-terminal amino acid onto an organic linker (e.g., PAM, 4-oxymethylphenylacetamidomethyl), which is covalently attached to an insoluble polystyrene resin cross-linked with divinyl benzene. The terminal amine may be protected by blocking with t-butyloxycarbonyl. Hydroxyl- and carboxyl-groups are commonly protected by blocking with O-benzyl groups. Synthesis is accomplished in an automated peptide synthesizer, a number of which are commercially available. Following synthesis, the product may be removed from the resin. The blocking groups are removed typically by using hydrofluoric acid or trifluoromethyl sulfonic acid according to established methods (e.g., Bergot and McCurdy, Applied Biosystems Bulletin, 1987). Following cleavage and purification, a yield of approximately 60 to 70% is typically produced. Purification of the product peptides is accomplished by, for example, crystallizing the peptide from an organic solvent such as methyl-butyl ether, then dissolving in distilled water, and using dialysis (if the molecular weight of the subject peptide is greater than about 500 daltons) or reverse high-pressure liquid chromatography (e.g., using a C18 column with 0.1% trifluoroacetic acid and acetonitrile as solvents) if the molecular weight of the peptide is less than 500 daltons. Purified peptide may be lyophilized and stored in a dry state until use. Analysis of the resulting peptides may be accomplished using the common methods of analytical high pressure liquid chromatography (HPLC) and electrospray mass spectrometry (ES-MS).
[0093] In general, transgenic plants comprising cells containing polynucleotides of the present disclosure can be produced by any of the foregoing methods; selecting plant cells that have been transformed on a selective medium; regenerating plant cells that have been transformed to produce differentiated plants; and selecting a transformed plant that expresses the protein(s) encoded by the polynucleotides of the present disclosure at a desired level.
[0094] Specific methods for transforming a wide variety of dicots and obtaining transgenic plants are well documented in the literature (Gasser and Fraley, Science 244:1293, 1989; Fisk and Dandekar, Scientia Horticulturae 55:5, 1993; Dandekar and Fisk, Plant transformation: agrobacterium-mediated gene transfer. Methods Mol. Biol. 2005; 286:35-46; Olhoft P M, Donovan C M, Somers D A, Soybean (Glycine max) transformation using mature cotyledonary node explants, Methods Mol. Biol. 2006; 343:385-96; Ko T S, Korban S S, Somers D A, Soybean (Glycine max) transformation using immature cotyledon explants, Methods Mol Biol. 2006; 343:397-405; and all references cited therein).
[0095] Successful transformation and plant regeneration have also been achieved in a variety of monocots. Specific examples are as follows: asparagus (Asparagus officinalis; Bytebier et al. (1987) Proc. Natl. Acad. Sci. USA 84: 5345); barley (Hordeum vulgarae; Wan and Lemaux (1994) Plant Physiol. 104: 37); maize (Zea mays; Rhodes et al. (1988) Science 240: 204; Gordon-Kamm et al. (1990) Plant Cell 2: 603; Fromm et al. (1990) Bio/Technology 8: 833; Koziel et al. (1993) Bio/Technology 11: 194); oats (Avena sativa; Somers et al. (1992) Bio/Technology 10: 1589); orchardgrass (Dactylis glomerata; Horn et al. (1988) Plant Cell Rep. 7: 469); rice (Oryza sativa, including indica and japonica varieties; Toriyama et al. (1988) Bio/Technology 6: 10; Zhang et al. (1988) Plant Cell Rep. 7: 379; Luo and Wu (1988) Plant Mol. Biol. Rep. 6: 165; Zhang and Wu (1988) Theor. Appl. Genet. 76: 835; Christou et al. (1991) Bio/Technology 9: 957); rye (Secale cereale; De la Pena et al. (1987) Nature 325: 274); sorghum (Sorghum bicolor; Cassas et al. (1993) Proc. Natl. Acad. Sci. USA 90: 11212); sugar cane (Saccharum spp.; Bower and Birch (1992) Plant J. 2: 409); tall fescue (Festuca arundinacea; Wang et al. (1992) Bio/Technology 10: 691); turfgrass (Agrostis palustris; Zhong et al. (1993) Plant Cell Rep. 13: 1); and wheat (Triticum aestivum; Vasil et al. (1992) Bio/Technology 10: 667; Weeks et al. (1993) Plant Physiol. 102: 1077; Becker et al. (1994) Plant J. 5: 299). All these references relate to transformation techniques in dicots or monocots and are hereby expressly incorporated into this disclosure by reference.
[0096] Various LysM RLK genes show tissue specific expression in plants. Tissue specific promoters or other regulatory elements may play a role in controlling these tissue specific expression patterns. DNA recombination utilizing these regulatory elements may be employed to manipulate the expression pattern and/or levels of the various LysM RLK genes, or other genes in general. For instance, expression construct containing a tissue specific promoter may be used to drive the expression of a LysM RLK which is not otherwise expressed in the particular tissue.
EXAMPLES
[0097] The following examples are intended to provide illustrations of the application of the present disclosure. The following examples are not intended to completely define or otherwise limit the scope of the invention.
Example 1
Plant LysM Domains are Highly Diversified
[0098] The sequences of NFR1 (SEQ ID No. 1) and NFR5 (SEQ ID No. 2) genes from Lotus japonicus, as reported by Radutoiu et al. (2003) were used to identify genes encoding LysM domain-containing proteins by searching public databases of Arabidopsis, rice, poplar, M. truncatula, and L. japonicus. Soybean LYK genes were identified by shotgun sequencing bacterial artificial chromosomes (BACs) with homologies to LysM-encoding ESTs. The resulting putative LYK protein sequences from all species were then searched against the Pfam server to verify LysM and kinase domains. Collectively, a total of 49 LYK genes were identified in the six plant genomes, namely, those of Arabidopsis, rice, poplar, M. truncatula, L. japonicus and soybean, as summarized in Table 2. The predicted amino acid sequences of these genes or fragments were obtained and compared by sequence alignment, with representative alignments shown in FIG. 2.
TABLE-US-00002 TABLE 2 LysM type receptor-like kinase genes from Arabidopsis, soybean, Lotus, Medicago, rice and poplar. Name (SEQ ID No.) Alias name Sources AtLYK1 (SEQ ID No.3) At3g21630 TAIR AtLYK2 (SEQ ID No.4) At3g01840 TAIR AtLYK3 (SEQ ID No.5) At1g51940 TAIR AtLYK4 (SEQ ID No.6) At2g23770 TAIR AtLYK5 (SEQ ID No.7) At2g33580 TAIR GmNFR1α (SEQ ID 54) GmW2098N11.16 this study GmNFR1β (SEQ ID 55) GmW2098N15.9 this study GmLYK2 (SEQ ID 56) GmW2098N11.15 this study GmLYK3 (SEQ ID 57) GmW2026N19.18 this study GmLYK4 (SEQ ID 58) GmW2095P01.22 this study GmNFR5α (SEQ ID 59) GmW2035N07.17 this study GmNFR5β (SEQ ID 60) GmW2095P01.23 this study GmLYK6 (SEQ ID 61) GmW2075N23 this study GmLYK7 (SEQ ID 62) GmW2035N07.16 this study GmLYK8 (SEQ ID 63) GmW2098N11.2 this study GmLYK9 (SEQ ID 64) GmW2069O12.22 this study GmLYK10 (SEQ ID 65) GmW2080D08.12 this study GmLYK11 (SEQ ID 66) GmW2042I24.15 this study LjNFR1 (SEQ ID 1) AJ575248 Gene Bank LjLYK2 (SEQ ID 67) TM0545.8 Kazusa LjLYK3 (SEQ ID 68) TM0545.9 Kazusa LjLYK4 (SEQ ID 69) TM0522.16 Kazusa LjNFR5 (SEQ ID 2) AJ575255 Gene Bank LjLYK6 (SEQ ID 70) TM0076a.10 Kazusa MtLYK1 (SEQ ID 71) CR936945.12 Medicago truncatula sequencing resources MtLYK3 (SEQ ID 72) AY372402 Gene Bank MtLYK4 (SEQ ID 73) AY372403 Gene Bank MtLYK9 (SEQ ID 74) AC148241_11 Medicago truncatula sequencing resources MtLYK10 (SEQ ID 75) AC148994_13 Medicago truncatula sequencing resources MtLYK11 (SEQ ID 76) AC148994_15 Medicago truncatula sequencing MtLYK12 (SEQ ID 77) AC126779_3 Medicago truncatula sequencing resources MtLYK13 (SEQ ID 78) AC126779_4 Medicago truncatula sequencing resources OsLYK1 (SEQ ID 79) LOC_Os01g36550 TIGR OsLYK2 (SEQ ID 80) LOC_Os06g41980 TIGR OsLYK3 (SEQ ID 81) LOC_Os06g41960 TIGR OsLYK4 (SEQ ID 82) LOC_Os02g09960 TIGR OsLYK5 (SEQ ID 83) LOC_Os03g13080 TIGR OsLYK6 (SEQ ID 84) LOC_Os11g35330 TIGR PtLYK1 (SEQ ID 85) FGENESH1_PG.C_LG_VIII001701 DOE JGI PtLYK2 (SEQ ID 86) FGENESH1_PG.C_LG_VII000997 DOE JGI PtLYK3 (SEQ ID 87) EUGENE3.00051645 DOE JGI PtLYK4 (SEQ ID 88) EUGENE3.00081504 DOE JGI PtLYK5 (SEQ ID 89) EUGENE3.00400189 DOE JGI PtLYK6 (SEQ ID 90) GRAIL3.0019013601 DOE JGI PtLYK7 (SEQ ID 91) GRAIL3.0017002501 DOE JGI PtLYK8 (SEQ ID 92) FGENESH1_PM.C_LG_I000490 DOE JGI PtLYK9 (SEQ ID 93) EUGENE3.00570233 DOE JGI PtLYK10 (SEQ ID 94) EUGENE3.00100714 DOE JGI PtLYK11 (SEQ ID 95) eugene3.00570235 DOE JGI
[0099] More specifically, plant LysM protein sequences were first searched using the key word LysM and BLASTp (1e-20) using the LysM domains of LjNFR1 (SEQ ID No. 1) and LjNFR5 (SEQ ID No. 2) against the following publicly available databases: Arabidopsis (Arabidopsis thaliana, database maintained by the Carnegie Institution of Washington Department of Plant Biology); rice (Oryza sativa, database maintained by the Institute for Genomic Research (TIGR)); poplar (Populus spp., database maintained by DOE's Joint Genome Institute); Medicago truncatula, database maintained by the lab of Nevin Young at the University of Minnesota); and Lotus japonicus (database maintained by the Kazusa DNA Research Institute in Japan. Domain structures of the resulting potential LysM proteins were analyzed with Pfam software and Inter-ProScan to identify LysM proteins. Soybean (Glycine max) LysM proteins were searched via tBLASTn (1e-5) using the same query sequences as above against two publicly available EST databases, one maintained by the Institute for Genomic Research, the other maintained by Monsanto.
[0100] Primers were designed based on the resulting soybean EST sequences to probe a six-dimensional BAC pool for LysM-containing BACs via a PCR-based approach. The probed LYK-containing BACs were verified and shotgun sequenced to either finished phase (phase 3) at the Arizona Genome Sequencing Center or prefinished phase (phase 2) at the Washington University Genome Sequencing Center. BAC sequences were annotated using the dicot species model and Arabidopsis matrix of FGENESH. Annotated proteins were similarly analyzed to screen for LYK proteins. Signal peptides and transmembrane domains were predicted with SignalP using both nearest-neighbor and hidden Markov model (HMM) algorithms and transmembrane HMM, respectively.
[0101] The GenBank accession numbers of soybean BACs are EF533702 for GMWb098N11; EF533695 for GMWb098N15; EF533696 for GMWb026N19; EF533701 for GMWb095P01; EF533697 for GMWb035N07; EF533699 for GMWb069O12; EF533700 for GMWb080D08; and EF533698 for GMWb042124. LysM protein sequences from species spanning all kingdoms were extracted from Pfam and searched for LysM motifs at an E-value cutoff of 0.1.
[0102] Sequence alignments were performed using ClustalX 1.83 (Thompson et al., 1997) with PHYLIP output format and edited in Jalview (Clamp et al., 2004). The average identities across the alignments for LysMe (type X), LysMn (type XI), and LYPb (type VII) were calculated based on the exported annotations in Jalview. An HMM profile calculated using hmmer (Eddy, 1998) for each alignment was used to realign (hmmalign) sequences at matching states (-m) to identify and remove indel regions.
[0103] Parsimony trees were generated using the program protpars of PHYLIP (Felsenstein, 2000), with maximum-likelihood branch lengths calculated using TREE-PUZZLE (Schmidt et al., 2002). Distance trees were calculated using the program Protdist and Fitch of the PHYLIP package. Maximum-likelihood trees were calculated using the program proml of the PHYLIP package. Bootstrap values were calculated using the program seqboot of the PHYLIP package. Trees were viewed and rooted using A Tree Viewer (Zmasek and Eddy, 2001). For calculation of nucleotide substitution rates, codon-aligned nucleic acid sequences were created using CodonAlign 2.0. All insertions and deletions were removed except that a gap of more than 30 nucleotides was preferably retained to demonstrate the lack of the p loop and the activation loop in the kinase domains of LjNFR5 orthologs (Limpens et al., 2003; Madsen et al., 2003; Arrighi et al., 2006). Nucleotide substitution levels were calculated using the program codeml of the PAML package (Yang, 1997) with a user-defined parsimony tree.
[0104] To build microsynteny maps, genomic sequences surrounding each LYK gene, about 0.5 to 0.9 Mb in length, were extracted from the above databases and from soybean BAC sequences, which are about 100 to 170 kb in length. The genomic sequences were annotated using dicot species model and Arabidopsis matrix of FGENESH for the five dicot plants and monocot species model and rice matrix for rice. The annotated protein sequences were compiled together into a peptide sequence database. Repetitive sequences were excluded from the databases. BLASTp was used to compare proteins against the database with an E-value cutoff of 1e-20 and a percent identity cutoff of 35% between species and 40% within same species and legumes. BLASTp results were then filtered once to remove retroelements. The microsynteny maps were finally drawn in Adobe Illustrator 10.0.
Example 2
Induction of Gene Expression in Arabidopsis Treated with Chitin
[0105] A total of five LysM receptor-like kinase genes were identified in Arabidopsis from the studies described in Example 1. The Genbank numbers of these five genes are AtLYK1 (GenBank accssion # At3g21630), AtLYK2 (GenBank accssion # At3g01840), AtLYK3 (GenBank accssion # At1g51940), AtLYK4 (GenBank accssion # At2g23770), AtLYK5 (GenBank accssion # At2g33580), and, which are designated as SEQ ID. Nos. 3-7, respectively. A DNA microarray experiment was performed by treating Arabidopsis plants with chitin. Leaves were treated by spraying with chitin (100 μM)+0.2% Tween-20. The Affymetrix 24K Arabidopsis genome chip was utilized according to the manufacturer's instructions for this test. The data obtained showed that transcription of 3 of the 5 Arabidopsis LysM RLK genes, At2g33580 (13-fold), At3g21630 (2-fold) and At2g23770 (2-fold), was significantly increased by treating the plants with chitin. The data implicate these genes in plant chitin response.
Example 3
LysM RLK Mutants in Arabidopsis
[0106] To test whether non-leguminous LysM RLKs may be involved in the perception of chitooligosaccharides and the subsequent induction of downstream genes that have been implicated in fungal defense, T-DNA insertion mutants were obtained for all five LysM RLK genes (i.e., At1g51940, At2g23770, At2g33580, At3g01840, and At3g21630) in Arabidopsis. The gene At3g21630 (SEQ ID. No. 3) is also termed AtLYK1 or AtLysM RLK1. Homozygous mutants were then treated with a purified chitooligosaccharide (chitooctaose) and the expression levels of known CRGs, such as MPK3 (At3g45640, SEQ ID. No. 8), WRKY22 (At4g01250, SEQ ID. No. 9), WRKY29 (At4g23550, SEQ ID. No. 10), WRKY33 (At2g38470, SEQ ID. No. 11), and WRKY53 (At4g23810, SEQ ID. No. 12), were measured.
[0107] More particularly, Arabidopsis seedlings were grown hydroponically as described by Ramonell et al., 2005. Fourteen-day old seedlings were treated with chitooctaose (Sigma, St. Louis, Mo., USA) at a concentration of 1 μM or with distilled water (as a control) for 30 minutes. To test flagellin-responsive genes, 14-day old seedlings were also treated with the flagellinderived flg22 peptide (dissolved in dimethyl sulfoxide, DMSO) at a final concentration of 10 μM or with an equivalent amount of DMSO (as a control) for 30 minutes. To test other defense pathways, Arabidopsis seedlings were also treated for 24 hours with 5 mM SA, 100 μM MeJA, and 0.5 mM ACC (all obtained from Sigma, St. Louis, Mo., USA) and dissolved in 0.1% ethanol. The control plants were similarly treated with an equivalent amount of ethanol. After treatment, the seedlings were collected and frozen in liquid nitrogen for RNA isolation.
[0108] Total RNA was isolated using the Trizol Reagent according to the manufacturer's instructions (Invitrogen, Carlsbad, Calif.). The isolated RNA was further purified using Qiagen RNeasy Mini Columns according to the manufacturer's instruction (Qiagen, Valencia, Calif., USA) and treated with Turbo® DNase (Ambion, Austin, Tex., USA). For semi-quantitative RT-PCR or quantitative 17 PCR, cDNA was synthesized using M-MLV reverse transcriptase according to the manufacturer's instructions (Promega, Madison, Wis., USA).
[0109] Semi-Quantitative RT-PCR.
[0110] The gene-specific primer pairs (forward and reverse) for detecting the following selected chitooligosaccharide-responsive genes (CRGs) are:
TABLE-US-00003 For MPK3 (At3g45640): (SEQ ID No. 13) 5'-CTCACGGAGGACAGTTCATAAG-3' and (SEQ ID No. 14) 5'-GAGATCAGATTCTGTCGGTGTG-3' For WRKY22 (At4g01250): (SEQ ID No. 15) 5'-GTAAGCTCATCAGCTACTACCAC-3' and (SEQ ID No. 16) 5'-ACCGCTAGATGATCCTCAACAG-3' for WRKY29 (At4g23550): (SEQ ID No. 17) 5'-ATGGACGAAGGAGACCTAGAAG-3' and (SEQ ID No. 18) 5'-CCGCTTGGTGCGTACTCGTTTC-3' For WRKY33 (At2g38470): (SEQ ID No. 19) 5'-CTCCGACCACAACTACAACTAC-3' and (SEQ ID No. 20) 5'-GGCTCTCTCACTGTCTTGCTTC-3' For WRKY53 (At4g23810): (SEQ ID No. 21) 5'-CCTACGAGAGATCTCTTCTTCTG-3' and (SEQ ID No. 22) 5'-AGATCGGAGAACTCTCCACGTG-3'
[0111] As an internal control, the following forward and reverse primers of actin-2 (At3g18780) were included in the same PCR reaction with each primer pair of the above genes:
TABLE-US-00004 (SEQ ID No. 23) 5'-GACTAAGAGAGAAAGTAAGAGATAATCCAG-3' and (SEQ ID No. 24) 5'-CAGCCTTTGATTTCAATTTGCATGTAAGAG-3'.
[0112] PCR reactions were conducted using Taq polymerase (Promega, Madison, Wis., USA) under the following conditions: 94° C., 3 minutes; 94° C., 30 seconds; 55° C., 30 seconds; 72° C., 1.5 minutes; 25 cycles; 72° C., 3 minutes. The corresponding CRG genes in Lotus japonicus were identified by blasting the cDNA sequences of the above Arabidopsis CRGs (and also actin-2) against the TIGR Lotus japonicus Gene Index. The closest hits were chosen and arbitrarily named after their Arabidopsis counterparts with the prefix Lj (standing for Lotus japonicus). The following primer pairs were designed to detect these genes:
TABLE-US-00005 For LjMPK3 (TC8079): (SEQ ID No. 25) 5'-CACCCTTGCGTAGAGAGTTTACTGATGTC-3', and (SEQ ID No. 26) 5'-GTTGACGAGGATATTGAGGAAGTTGTCTG-3'; For LjWRKY22 (AV423663): (SEQ ID No. 27) 5'-TCACCTTGCTGGTTCTGGTTCTGGTTCTG-3', and (SEQ ID No. 28) 5'-TCTGATAGGGGTGCAACCCCATCTTCTTC-3'; For LjWRKY33 (TC14849): (SEQ ID No. 29) 5'-AGTTGTGGTTCAGACCACCAGTGACATTG-3' and (SEQ ID No. 30) 5'-ACCCCATTGAGTTTCCAAACCCTGATGAG-3'; For LjWRKY53 (TC9074): (SEQ ID No. 31) 5'-CCCATCAAAAGAACCAACCACAACAAGAG-3' and (SEQ ID No. 32) 5'-ATCCGCACGCACTTGAACCATGTATTGTG-3'; For LjActin-2 (TC14247): (SEQ ID No. 33) 5'-AAGGTTCGTAAACGATGGCTGATGCTGAG-3' and (SEQ ID No. 34) 5'-ACCTTGATCTTCATGCTGCTAGGAGCAAG-3'.
[0113] LjActin-2 was used as an internal control.
[0114] Quantitative PCR.
[0115] To quantify gene expression using quantitative PCR, the forward and reverse primers of each gene were as follows:
TABLE-US-00006 For PR-1 (At2g14610, SEQ ID. No. 35): (SEQ ID No. 36) 5'-AACACGTGCAATGGAGTTTGTGGTCACT-3' and (SEQ ID No. 37) 5'-ACCATTGTTACACCTCACTTTGGCACAT-3'; For PDF1.2 (At5g44420, SEQ ID. No. 38): (SEQ ID No. 39) 5'-AGTGCATTAACCTTGAAGGAGCCAAACAT-3' and (SEQ ID No. 40) 5'-AACAGATACACTTGTGTGCTGGGAAGACA-3'; For MPK3 (At3g45640): (SEQ ID No. 41) 5'-TGGCCATTGATCTTGTTGACAGAATGTTGA-3' and (SEQ ID No. 42) 5'-TCGTGCAATTTAGCAAGGTACTGGTGATT-3'; for WRKY53 (At4g23810): (SEQ ID No. 43) 5'-TTTAGGCGCCAAATTCCCAAGGAGTTATT-3' and (SEQ ID No. 44) 5'-TCTGGACTTGTTTCGTTGCCCAACAGTTT-3'; For actin-2 (At3g18780): (SEQ ID No. 45) 5'-GGTATTCTTACCTTGAAGTATCCTATTG-3' and (SEQ ID No. 46) 5'-CTCATTGTAGAAAGTGTGATGCCAGATC-3'.
[0116] Actin-2 was used as an internal control to normalize gene expression across different samples. The reactions were conducted on a 7500 Real-Time PCR System (Applied Biosystems, Foster City, Calif., USA) using the SYBR®Green Master Mix (Applied Biosystems, Foster City, Calif., USA) with the following PCR conditions: 95° C., 10 minutes; 95° C., 15 seconds; 60° C., 1 minute; 40 cycles; followed by the dissociation curve analysis to verify single amplicon. The fold change in the target gene, normalized to actin-2 and relative to the gene expression in the control sample, was calculated as described in Ramonell et al., 2002.
[0117] The AtLysM RLK1 insertion mutant (096F09) used in the current work was generated in the context of the GABI-Kat program and provided by Bernd Weisshaar (MPI for Plant Breeding Research, Cologne, Germany) See Shibuya et al., 2001. The homozygous plants were identified by genotyping using the following gene-specific primers:
TABLE-US-00007 (SEQ ID No. 47) 5'- AGAATATATCCACGAGCACACGGTTCCAG-3' (forward), and (SEQ ID No. 48) 5'-GACGAAAAGAGAGTGGATAAAGCAACCAC-3' (reverse) together with the T-DNA left border primer: (SEQ ID No. 49) 5'-CCCATTTGGACGTGAATGTAGACAC-3'.
[0118] These two primers were also used to detect the expression of the AtLysM RLK1 gene via RT-PCR. The other primers used to detect the transcript 5' of the insertion site were as follows:
TABLE-US-00008 (SEQ ID No. 50) 5'-ATGAAGCTAAAGATTTCTCTAATCGCTC-3', and (SEQ ID No. 51) 5'-GAAATGCACCATTTGGATCTCTTCCAG-3'
[0119] The mutants of the other 4 Arabidopsis LysM RLK genes were obtained from the SALK Institute and Syngenta Incorporation through the Arabidopsis Basic Research Center (ABRC) or from the Martienssen lab at the Cold Spring Harbor Laboratory.
[0120] One insertion mutant designated as L3 with an insertion in gene At1g51940 exhibited an interesting phenotype. This mutant showed enhanced expression of some defense-related genes, such as PR-2 and PR-1, as shown in FIG. 3. The PR-2 gene (AT3G57260, SEQ ID. No. 52) encodes a β-1,3-glucanase, which is an enzyme that degrades the fungal cell wall component glucan to inhibit fungal infection. PR-1 (SEQ ID. No. 35) has also been shown to be involved in plant defense against pathogens, especially bacterial pathogens. This enhanced expression of defense genes suggests that the mutant may be resistant to fungal pathogens. The test of this mutant with a fungal pathogen called Botrytis cinerea demonstrated that the mutant is resistant to this fungal pathogen, as shown in FIG. 4A. B. cinerea is a necrotrophic fungus, and dead plants were assessed as those having no remaining green, only yellowish leaves. The L3 mutant demonstrated increased resistance relative to the wild-type plant.
[0121] In addition, the L3 mutant showed decreased susceptibility to the bacterial pathogen Pseudomonas syringae strain DC3000, as shown in FIG. 4B. The enhanced resistance is likely due to the knockout of the specific LysM receptor kinase gene. Analysis of expression of the At1g51940 gene in the L3 mutant failed to detect the mRNA. Therefore, the lack of At1g51940 gene expression correlates with elevated PR1 expression and enhanced disease resistance. This suggests that At1g51940 may act normally to repress the disease resistance response according to a model pathway of the involvement of the LYSM RK in plant defense, as shown in FIG. 5. Therefore, it is possible to make plants more resistant to fungal pathogens by either knocking out or knocking down the expression of this gene. Furthermore, dominant negative forms of this protein may also be made and employed to modulate plant fungal resistance.
[0122] Another insertion mutant, corresponding to At3g21630, or AtLYK1 (designated hereafter as AtLysM RLK1), almost completely blocked the induction of all the selected CRGs (FIG. 6A), suggesting a critical role of AtLysM RLK1 in the perception of chitooligosaccharides. Both the mutant (Mu) and wildtype (WT) plants were treated with purified chitooctaose or water (as a control) for 30 minutes and gene expression of the selected CRGs was detected using semi-quantitative RT-PCR. Actin-2 was used as an internal control. The amplification of both actin-2 and a CRG was conducted in the same tube.
[0123] The AtLysM RLK1 gene (SEQ ID. No. 3) is 2988 nucleotides (nts) long, with 11 introns (FIG. 6B) and a coding sequence of 1854 nts. Square boxes represent exons. Solid lines between them are introns. The start codon (ATG) and stop codon (TAG) are included in the first and last exon, respectively. The two T-DNA insertions (T-DNA1 and 2) inserted in the 10th intron in the AtLysM RLK1 mutant are indicated above the gene. LB: left border; RB: right border. The AtLysM RLK1 gene encodes a LysM RLK of 617 amino acids (SEQ ID No. 53), with an extracellular domain (containing 3 predicted LysM motifs), a transmembrane domain (TM), and an intracellular serine/threonine kinase domain. FIG. 6C illustrates the predicted domain structure of AtLysM RLK1. S: signal peptide; LysM: LysM domain; TM: transmembrane domain; Ser/Thr Kinase: Serine/Threonine kinase domain. AtLysM RLK1 has been shown to be phylogenetically related to the Nod signal receptor NFR1. Zhang et al., 2007.
[0124] Two T-DNA insertions were identified in the AtLysM RLK1 mutant, separated by 4 nts, in the 10th intron (FIG. 6B). RT-PCR analysis using primers corresponding to the exon regions on the side of the 10th intron failed to detect mRNA expression in the AtLysM RLK1 mutant; however, a truncated transcript derived from the gene sequence before the intron was detected by RT-PCR (FIGS. 7A and 7B), suggesting the T-DNA insertions in the intron blocked full-length transcription of the gene.
[0125] To confirm that the observed changes in CRGs expression were caused by the mutation in the AtLysM RLK1 gene, the mutant was complemented with the full-length AtLysM RLK1 cDNA driven by the constitutive Cauliflower Mosaic Virus (CMV) 35S promoter. More specifically, the full-length CDS (1854 nucleotides long) was obtained by RT-PCR and cloned in the Eco RV site of the pBluescript vector. The confirmed sequence was further cloned into the modified binary 16 vector pCAMBIA1200 that contains a 35S promoter-Multiple Cloning Sites (MCS)-poly A signal, downstream of the 35S promoter. The final construct was electroporated into Agrobacterium tumafaciens EHA105 according to the procedures described by Stacey and Shibuya, 1997. The resultant A. tumafaciens was then used to transform the homozygous AtLysM RLK1 mutant via floral dipping as described by Passarinho et al., 2002.
[0126] Multiple transgenic lines were obtained. The complemented plants were treated with chitooctaose or water (as a control) at a final concentration of 1 μM for 30 minutes. RT-PCR data show that the selected CRGs were induced to a level in the selected complemented plants (Com-1 and Com-2) similar to the level in the wild type (WT) plants (FIG. 8). Com-1 and Com-2 are two independent complemented lines and WT is wild-type Col-0 plants.
[0127] Thus, the complemented plants showed restored induction of those selected CRGs, confirming that it was the insertions in the AtLysM RLK1 gene that caused the observed change in gene expression. The complementation data also ruled out the possibility that a truncated protein translated from the observed truncated transcript may have affected the expression of the selected CRGs.
[0128] The expression pattern of the AtLysM RLK1 gene was also studied. RT-PCR data show that AtLysM RLK1 is expressed ubiquitously in the whole plant, in tissues such as root, rosette leaf, cauline leaf, stem, inflorescence, silique, flower bud, open flower, and pollen, with the lowest expression levels in pollen (FIG. 9). Interestingly, this gene was induced by chitooligosaccharides, but not by the flg22 peptide derived from flagellin, a PAMP (pathogen-associated molecular pattern) produced by pathogenic bacteria (FIG. 10) (Gomez-Gomez et al., 2000), suggesting a specific role of this gene in chitooligosaccharide signaling. More specifically, for experiment (A), fourteen-day-old, hydroponically grown seedlings were treated for 30 minutes with chitooctaose at a final concentration of 1 μM or with distilled water (as a control); for experiment (B), the seedlings were treated with flg22 (dissolved in DMSO) at a final concentration of 10 μM or with an equivalent amount of DMSO (as a control).
[0129] Gene expression profiles in the AtLysM RLK1 mutant in response to chitooctaose were studied using the Affymetrix Arabidopsis Whole Genome Array ATH1 (with ˜22000 genes), with wild-type plants as a control. Data analysis showed that a total of 909 genes responded more than 1.5 fold (P<0.05) to chitooctaose elicitation in both the wild-type and mutant plants 30 minutes after the treatment (FIG. 11A). A row represents a gene and each column represents a sample. WT-8mer=wild-type Col-0 treated with chitooctaose; WT-water=wild-type Col-0 treated with distilled water; Mu-water=the AtLysM RLK1 mutant treated with distilled water; Mu-8mer=the AtLysM RLK1 mutant treated with chitooctaose. The color bar below the cluster picture: the red color indicates the expression level of a gene is above the mean expression of the gene across all samples; the green color indicates expression lower than the mean. These genes can be separated into two groups: up- and down-regulated by chitooctaose, as represented by the two large clusters in FIG. 11A.
[0130] Out of the 909 genes tested, 890 showed a change in transcript levels in the wild-type plants in response to chitooctaose, with 663 up-regulated and 227 down-regulated (FIG. 11B, 11C and Table 3). Up-regulated genes: 1.5 fold, P<0.05. Down-regulated genes: 1.5 fold, P<0.05. By contrast, only 33 genes out of 909 were responsive in the mutant, with 16 up-regulated and 17 down-regulated (FIGS. 11B and 11C; Table 4). Among the 33 genes, 14 genes (3 up- and 11 down-regulated) were also similarly regulated in the wild-type plants (rows 1 to 15 in Table 4), leaving only 19 genes that appeared to be differentially regulated by chitooctaose in the mutant (rows 16-34 in Table 4). However, 13 of these genes showed a similar regulation trend (up- or down-regulation) in the wild-type plants to that in the mutant, although such a trend was not considered significant in the wild-type plants (rows 16 to 28 in Table 4).
TABLE-US-00009 TABLE 3 Genes that are Responsive to chitooctaose in Wild-type WT Mu Probe set Annotation Accession FC FC WT P Mu P 245613_at hypothetical protein At4g14450 72.79 -1.8 0.019018 0.836714 249197_at putative protein contains similarity to At5g42380 51.63 1.06 0.019359 0.866994 calmodulin; supported by full-length cDNA: Ceres: 99348. 258947_at hypothetical protein similar to calmodulin-like At3g01830 43.33 1.58 0.012148 0.103554 protein GB: CAB42906 [Arabidopsis thaliana]; Pfam HMM hit: EF hand; supported by full-length cDNA: Ceres: 7252. 260399_at putative lipoxygenase similar to lipoxygenase At1g72520 41.48 -1.11 0.009183 0.804844 GB: CAB56692 [Arabidopsis thaliana]; supported by cDNA: gi_15810254_gb_AY056166.1-- 257540_at hypothetical protein At3g21520 34.95 1.08 0.001687 0.921695 256526_at disease resistance protein, putative similar to disease At1g66090 33.68 -1.13 0.01628 0.597847 resistance protein RPP1-WsA [Arabidopsis thaliana] GI: 3860163; supported by full-length cDNA: Ceres: 93530. 250796_at putative protein similar to unknown protein At5g05300 31.56 1.1 0.004167 0.773037 (gb|AAF01528.1) 261474_at anionic peroxidase, putative similar to anionic At1g14540 30.96 -1.02 0.001414 0.955549 peroxidase GI: 170202 from [Nicotiana sylvestris] 245755_at hypothetical protein predicted by At1g35210 30.5 -1.11 0.01789 0.881393 genemark.hmm; supported by full-length cDNA: Ceres: 42217. 254231_at putative protein AR411-Arabidopsis thaliana (thale At4g23810 28.21 -1.25 0.018401 0.425568 cress), PID: g1669603; supported by cDNA: gi_13507100_gb_AF272748.1_AF272748 248322_at putative protein similar to unknown protein At5g52760 26.14 1.31 0.004693 0.152597 (emb|CAA71173.1) 249770_at unknown protein; supported by full-length cDNA: At5g24110 25.43 -1.06 0.035483 0.869658 Ceres: 6469. 247215_at Expressed protein; supported by full-length cDNA: At5g64905 24.84 1.24 0.04883 0.449026 Ceres: 3657. 265725_at putative alanine acetyl transferase At2g32030 23.73 1.08 0.027584 0.473742 266821_at putative ethylene response element binding protein At2g44840 23.69 -1.22 0.019899 0.271281 (EREBP); supported by full-length cDNA: Ceres: 6397. 248904_at Expressed protein; supported by full-length cDNA: At5g46295 23.69 -1.49 0.007953 0.428724 Ceres: 18973. 261648_at salt-tolerance zinc finger protein identical to salt- At1g27730 22.71 -1.09 0.016239 0.33423 tolerance zinc finger protein GB: CAA64820 GI: 1565227 from [Arabidopsis thaliana]; supported by cDNA: gi_14334649_gb_AY034998.1-- 262085_at hypothetical protein predicted by genemark.hmm At1g56060 22.37 1.36 0.001015 0.285112 261021_at hypothetical protein similar to reticuline oxidase-like At1g26380 22.2 2.37 0.004697 0.108353 protein GB: CAB45850 GI: 5262224 from [Arabidopsis thaliana]; supported by cDNA: gi_13430839_gb_AF360332.1_AF360332 263182_at Expressed protein; supported by full-length cDNA: At1g05575 19.34 -1.02 0.005652 0.804382 Ceres: 27081. 249417_at calcium-binding protein-like cbp1 calcium-binding At5g39670 18.84 -1.03 0.012777 0.874622 protein, Lotus japonicus, EMBL: LJA251808; supported by cDNA: gi_16648829_gb_AY058192.1-- 254120_at putative mitochondrial uncoupling protein At4g24570 18.47 -1.26 0.006894 0.121685 mitochondrial uncoupling protein, Arabidopsis thaliana (thale cress), PATX: E1316826; supported by full-length cDNA: Ceres: 119476. 264153_at disease resistance protein RPS4, putative similar to At1g65390 17.77 -1.01 0.006757 0.837017 disease resistance protein RPS4 GI: 5459305 from [Arabidopsis thaliana] 249264_s_at disease resistance protein-like At5g41740 17.16 1.01 0.016032 0.965149 246821_at calmodulin-binding-like protein calmodulin- At5g26920 16.58 -1.22 0.002033 0.305516 binding protein TCB60, Nicotiana tabacum, EMBL: U58971 265327_at unknown protein At2g18210 16.02 -1.08 0.007312 0.847704 252131_at BCS1 protein-like protein Homo sapiens h-bcs1 At3g50930 15.81 1.28 0.020727 0.19657 (BCS1) mRNA, nuclear gene encoding mitochondrial protein which is involved in the expression of functional mitochondrial ubiquinol- cytochrome c reductase complex probably via the control of expression of Riesk 245840_at hypothetical protein predicted by At1g58420 15.76 -1.16 0.001929 0.649675 genemark.hmm; supported by full-length cDNA: Ceres: 124269. 245041_at AR781, similar to yeast pheromone receptor At2g26530 15.71 -1.4 0.011514 0.105739 identical to GB: D88743, corrected a frameshift found in the original record (at 69530 bp), sequence submitted has been verified from 10 sequence electropherograms. The translation now starts from an upstream ATG. 248799_at ethylene responsive element binding factor 5 At5g47230 15.6 -1.36 0.005208 0.117072 (ATERF5) (sp|O80341); supported by cDNA: gi_14326511_gb_AF385709.1_AF385709 250149_at cinnamoyl CoA reductase-like protein cinnamoyl At5g14700 15.46 -1.13 0.022877 0.554222 CoA reductase, Populus tremuloides, EMBL: AF217958; supported by full-length cDNA: Ceres: 17229. 256306_at lipase, putative contains Pfam profile: PF01764: At1g30370 15.3 1.11 0.00866 0.751633 Lipase 246777_at RING-H2 zinc finger protein-like RING-H2 zinc At5g27420 14.67 -1.44 0.011803 0.037951 finger protein ATL6-Arabidopsis thaliana, EMBL: AF132016; supported by full-length cDNA: Ceres: 106078. 263783_at putative WRKY-type DNA binding protein; At2g46400 14.41 1.26 0.005589 0.126494 supported by cDNA: gi_15430276_gb_AY046275.1-- 245369_at Expressed protein; supported by full-length cDNA: At4g15975 14.34 1.03 0.00366 0.920236 Ceres: 124835. 251336_at putative protein hypothetical protein F4I18.26- At3g61190 14.31 1.07 0.007665 0.724447 Arabidopsis thaliana, PIR: T02471; supported by full- length cDNA: Ceres: 30454. 260046_at Expressed protein; supported by cDNA: At1g73800 13.55 1 0.010217 0.9771 gi_16648699_gb_AY058126.1-- 260068_at putative calmodulin-binding protein similar to At1g73805 13.35 1.16 0.007383 0.51944 calmodulin-binding protein GB: AAB37246 [Nicotiana tabacum] 266071_at unknown protein At2g18680 13.32 1.11 0.004216 0.713949 253643_at hypothetical protein; supported by full-length At4g29780 13.05 -1.11 0.002383 0.344174 cDNA: Ceres: 249769. 264213_at hypothetical protein contains similarity to lectin At1g65400 12.76 -1.17 0.021498 0.255697 polypeptide GI: 410436 from [Cucurbita maxima] 262382_at virus resistance protein, putative similar to virus At1g72920 12.68 -1.4 0.003189 0.073493 resistance protein GI: 558886 from [Nicotiana glutinosa] 247543_at DNA binding protein-like DNA binding protein At5g61600 12.31 -1.31 0.014862 0.112814 EREBP-4, Nicotiana tabacum, PIR: T02434; supported by full-length cDNA: Ceres: 92102. 256442_at hypothetical protein predicted by At3g10930 11.62 -1.07 0.025288 0.782271 genefinder; supported by full-length cDNA: Ceres: 12509. 253060_at putative protein predicted protein, Arabidopsis At4g37710 11.56 1.34 0.005721 0.412461 thaliana; supported by full-length cDNA: Ceres: 207350. 253915_at putative protein centrin, Marsilea vestita; supported At4g27280 11.5 -1.07 0.01185 0.333253 by full-length cDNA: Ceres: 13072. 249928_at CCR4-associated factor-like protein At5g22250 11.41 -1.17 0.009043 0.204172 245711_at putative c2h2 zinc finger transcription factor At5g04340 11.35 -1.11 0.016673 0.392119 261892_at transcription factor, putative similar to WRKY At1g80840 11.27 1.09 0.006954 0.498704 transcription factor GB: BAA87058 GI: 6472585 from [Nicotiana tabacum]; supported by full-length cDNA: Ceres: 6437. 261394_at wall-associated kinase 2, putative similar to wall- At1g79680 11.05 1.08 0.003074 0.819188 associated kinase 2 GI: 4826399 from [Arabidopsis thaliana] 251774_at nematode resistance protein-like protein Hs1pro-1 At3g55840 11.02 -1.37 0.007795 0.481526 nematode resistance gene, Beta procumbens, EMBL: BPU79733; supported by full-length cDNA: Ceres: 149697. 265723_at putative disease resistance protein At2g32140 10.99 1.18 0.016981 0.537825 255339_at hypothetical protein similar to A. thaliana At4g04480 10.83 1.32 0.012367 0.514455 hypothetical protein F1N20.130, GenBank accession number AL022140 251054_at receptor like protein kinase receptor like protein At5g01540 10.66 -1.01 0.003949 0.917593 kinase-Arabidopsis thaliana, EMBL: ATLECGENE; supported by cDNA: gi_13605542_gb_AF361597.1_AF361597 253827_at Expressed protein; supported by cDNA: At4g28085 10.37 -1.09 0.010797 0.530514 gi_15028040_gb_AY045877.1-- 255945_at putative protein At5g28610 10.06 1.21 0.010366 0.464865 249618_at putative protein predicted proteins, Arabidopsis At5g37490 9.99 -1.09 0.004719 0.814525 thalina 248934_at serine/threonine protein kinase-like protein At5g46080 9.94 -1.17 0.007605 0.645509 261037_at lipoxygenase identical to GB: CAB56692 from At1g17420 9.88 -1.1 0.002351 0.775492 (Arabidopsis thaliana) 267623_at unknown protein At2g39650 9.87 -1.12 0.006744 0.439834 259428_at MAP kinase, putative similar to MAP kinase 5 At1g01560 9.84 1.54 0.002458 0.135149 GI: 4239889 from [Zea mays] 246927_s_at nodulin-like protein nodulin, Glycine max, At5g25260 9.74 1.6 0.004623 0.163075 EMBL: AF065435 264758_at late embryogenesis abundant protein, putative At1g61340 9.73 1.17 0.016626 0.389155 similar to late embryogenesis abundant protein GI: 1350540 from [Picea glauca] 245329_at Expressed protein; supported by full-length cDNA: At4g14365 9.7 1.42 0.002486 0.029582 Ceres: 37809. 262072_at hypothetical protein predicted by At1g59590 9.54 -1.12 0.009452 0.599212 genemark.hmm; supported by full-length cDNA: Ceres: 99553. 255844_at putative protein kinase contains a protein kinase At2g33580 9.42 1.11 0.006611 0.553845 domain profile (PDOC00100) 253632_at senescence-associated protein homolog senescence- At4g30430 9.29 1.22 0.004184 0.351697 associated protein 5-Hemerocallis hybrid cultivar, PID: g3551954; supported by full-length cDNA: Ceres: 122632. 257511_at hypothetical protein At1g43000 9.29 -1.13 0.020984 0.846669 253999_at 1-aminocyclopropane-1-carboxylate synthase-like At4g26200 9.24 -1.56 0.004129 0.115048 protein ACC synthase, Malus domestica, U73816 265920_s_at unknown protein At2g15120 9.13 1.33 0.001682 0.33404 263800_at hypothetical protein predicted by genscan; supported At2g24600 8.97 1.02 0.014457 0.769925 by cDNA: gi_15810330_gb_AY056204.1-- 248164_at putative protein similar to unknown protein At5g54490 8.97 -1.17
0.008767 0.190232 (pir||T05752); supported by full-length cDNA: Ceres: 109272. 265597_at Expressed protein; supported by cDNA: At2g20145 8.96 -1 0.023429 0.965819 gi_13605516_gb_AF361584.1_AF361584 248327_at putative protein similar to unknown protein At5g52750 8.93 -1.04 0.017097 0.808905 (emb|CAA71173.1); supported by full-length cDNA: Ceres: 19542. 252908_at putative protein At4g39670 8.56 1.17 0.012661 0.466742 251400_at putative protein prib5, Ribes nigrum, At3g60420 8.53 1.64 0.029245 0.023237 EMBL: RNI7578; supported by full-length cDNA: Ceres: 31361. 261475_at anionic peroxidase, putative similar to anionic At1g14550 8.51 1.41 0.01103 0.395333 peroxidase GI: 170202 from [Nicotiana sylvestris] 256185_at dof zinc finger protein identical to dof zinc finger At1g51700 8.47 -1.09 0.001565 0.51485 protein [Arabidopsis thaliana] GI: 3608261; supported by cDNA: gi_3608260_dbj_AB017564.1_AB017564 250493_at putative protein various predicted proteins, At5g09800 8.28 -1.06 0.010787 0.857557 Arabidopsis thaliana 252679_at CCR4-associated factor 1-like protein At3g44260 8.27 -1.27 0.000484 0.065446 CAF1_MOUSE CCR4-ASSOCIATED FACTOR 1- Mus musculus, SWISSPROT: CAF1_MOUSE; supported by cDNA: gi_15292828_gb_AY050848.1-- 265797_at Expressed protein; supported by full-length cDNA: At2g35715 8.26 -1.27 0.005817 0.60028 Ceres: 9996. 248448_at putative protein contains similarity to ethylene At5g51190 8.25 -1.1 0.009635 0.575899 responsive element binding factor; supported by full- length cDNA: Ceres: 2347. 255884_at hypothetical protein predicted by At1g20310 8.15 -1.19 0.022852 0.204061 genemark.hmm; supported by full-length cDNA: Ceres: 8562. 261449_at putative ATPase similar to GB: AAF28353 from At1g21120 7.97 1.46 0.004955 0.182958 [Fragaria × ananassa] 265841_at putative glycogenin At2g35710 7.96 -1.27 0.011587 0.280483 251895_at class IV chitinase (CHIV) At3g54420 7.95 -1.09 0.003142 0.722712 263935_at unknown protein At2g35930 7.89 -1.06 0.006185 0.342267 255502_at contains similarity to a protein kinase domain (Pfam: At4g02410 7.89 -1.07 0.003365 0.691648 pkinase.hmm, score: 166.20) and to legume lectins beta domain (Pfam: lectin_legB.hmm, score: 139.32) 258787_at hypothetical protein predicted by genscan; supported At3g11840 7.84 -1.13 0.036463 0.37994 by full-length cDNA: Ceres: 100676. 266658_at Expressed protein; supported by full-length cDNA: At2g25735 7.71 -1.47 0.003942 0.026409 Ceres: 7152. 245250_at ethylene responsive element binding factor-like At4g17490 7.54 1.06 0.008242 0.704323 protein (AtERF6); supported by cDNA: gi_3298497_dbj_AB013301.1_AB013301 247487_at putative protein predicted protein, Arabidopsis At5g62150 7.39 1.01 0.005388 0.945249 thaliana 261470_at ethylene-responsive element binding factor, putative At1g28370 7.33 -1.17 0.005528 0.478834 similar to ethylene-responsive element binding factor GI: 8809573 from [Nicotiana sylvestris]; supported by full-length cDNA: Ceres: 27635. 262381_at virus resistance protein, putative similar to virus At1g72900 7.27 -1.19 0.006528 0.311035 resistance protein GI: 558886 from [Nicotiana glutinosa] 248123_at putative protein similar to unknown protein At5g54720 7.23 1.29 0.006214 0.245803 (gb|AAD32884.1) 263379_at putative CCCH-type zinc finger protein also an At2g40140 7.21 1.01 0.004911 0.848966 ankyrin-repeat protein 263584_at NAM (no apical meristem)-like protein similar to At2g17040 7.13 -1.29 0.006099 0.12014 petunia NAM (X92205) and A. thaliana sequences ATAF1 (X74755) and ATAF2 (X74756); probable DNA-binding protein; supported by cDNA: gi_13605646_gb_AF361804.1_AF361804 259566_at hypothetical protein At1g20520 7.04 -1.14 0.024145 0.734624 267028_at putative WRKY-type DNA binding protein At2g38470 7.02 -1.19 0.009642 0.27064 265008_at Mlo protein, putative similar to Mlo protein At1g61560 6.99 1.2 0.00298 0.470024 GI: 1877220 from [Hordeum vulgare]; supported by cDNA: gi_14091581_gb_AF369567.1_AF369567 247693_at putative protein leucine zipper-containing protein, At5g59730 6.97 1.01 0.004438 0.963142 Lycopersicon esculentum, PIR: S21495; supported by cDNA: gi_14334437_gb_AY034910.1-- 257748_at hypothetical protein predicted by genemark.hmm At3g18710 6.82 -1.16 0.009082 0.446456 258351_at hypothetical protein contains similarity to ion At3g17700 6.78 -1.02 0.004386 0.920019 channel protein from [Arabidopsis thaliana]; supported by cDNA: gi_8131897_gb_AF148541.1_AF148541 251745_at putative protein zinc finger transcription factor At3g55980 6.71 -1.36 0.001393 0.169064 (PEI1), Arabidopsis thaliana, EMBL: AF050463; supported by cDNA: gi_15810486_gb_AY056282.1-- 257536_at unknown protein At3g02800 6.46 1.24 0.011172 0.244827 246108_at putative protein retinal glutamic acid-rich protein, At5g28630 6.43 -1.14 0.017801 0.374617 bovine, PIR: A40437; supported by full-length cDNA: Ceres: 24151. 256046_at unknown protein At1g07135 6.42 -1.28 0.005287 0.339032 258436_at putative RING zinc finger protein similar to RING- At3g16720 6.39 -1.2 0.002525 0.290143 H2 zinc finger protein ATL6 GB: AAD33584 from [Arabidopsis thaliana]; supported by full-length cDNA: Ceres: 4581. 254255_at serine/threonine kinase-like protein At4g23220 6.39 1.58 0.01157 0.225448 serine/threonine kinase, Brassica oleracea; supported by cDNA: gi_14423417_gb_AF386946.1_AF386946 248686_at 33 kDa secretory protein-like; supported by cDNA: At5g48540 6.37 1.07 0.007302 0.530534 gi_15292980_gb_AY050924.1-- 248726_at RAS superfamily GTP-binding protein-like; At5g47960 6.34 -1 0.011505 0.996984 supported by cDNA: gi_12004622_gb_AF218121.1_AF218121 256633_at unknown protein At3g28340 6.32 -1.23 0.013314 0.277616 256183_at MAP kinase kinase 4 (ATMKK4) identical to MAP At1g51660 6.32 1.03 0.001255 0.842274 kinase kinase 4 [Arabidopsis thaliana]; supported by cDNA: gi_13265419_gb_AF324667.2_AF324667 247949_at cytochrome P450 At5g57220 6.31 -1.03 0.00798 0.740555 250098_at putative protein; supported by full-length cDNA: At5g17350 6.21 -1.09 0.005623 0.628534 Ceres: 1198. 255504_at drought-induced-19-like 1 similar to drought- At4g02200 6.14 1.1 0.002902 0.428306 induced-19, GenBank accession number X78584 similar to F2P16.10, GenBank accession number 2191179 identical to T10M13.20 253414_at putative protein At4g33050 6.08 -1.1 0.002073 0.284317 262731_at hypothetical protein similar to gb|AF098458 latex- At1g16420 6.07 1.18 0.016528 0.727417 abundant protein (LAR) from Hevea brasiliensis 247848_at resistance protein-like disease resistance protein At5g58120 6.07 -1.04 0.01295 0.876046 RPP1-WsA, Arabidopsis thaliana, EMBL: AF098962 254926_at ACC synthase (AtACS-6); supported by cDNA: At4g11280 6.04 -1.17 0.005123 0.161176 gi_16226285_gb_AF428292.1_AF428292 249719_at Expressed protein; supported by full-length cDNA: At5g35735 6.04 -1.08 0.005081 0.233393 Ceres: 32450. 247208_at nodulin-like; supported by full-length cDNA: At5g64870 6.04 1.22 0.001605 0.225756 Ceres: 142026. 257478_at hypothetical protein similar to putative At1g16130 5.96 -1.23 0.008918 0.562604 serine/threonine-specific protein kinase GI: 7270012 from [Arabidopsis thaliana] 246993_at Cys2/His2-type zinc finger protein 1 At5g67450 5.95 -1.06 0.005299 0.855156 (dbj|BAA85108.1) 252060_at putative protein other hypothetical proteins in At3g52430 5.94 1.2 0.005073 0.34486 Arabidopsis thaliana; supported by cDNA: gi_6457330_gb_AF188329.1_AF188329 267381_at unknown protein; supported by cDNA: At2g26190 5.9 -1.09 0.006528 0.587987 gi_16930468_gb_AF419588.1_AF419588 245038_at similar to latex allergen from Hevea brasiliensis; At2g26560 5.89 -1.06 0.019374 0.83179 supported by full-length cDNA: Ceres: 1999. 266800_at hypothetical protein predicted by genefinder At2g22880 5.86 -1.01 0.003336 0.993661 259211_at unknown protein identical to GB: AAD56318 At3g09020 5.82 1.08 0.00649 0.5476 (Arabidopsis thaliana) 253485_at Expressed protein; supported by full-length cDNA: At4g31800 5.82 -1.13 0.00494 0.428126 Ceres: 40692. 260211_at hypothetical protein similar to YGL010w-like At1g74440 5.77 1.06 0.003351 0.730279 protein GB: AAC32136 [Picea mariana] 256093_at predicted protein; supported by cDNA: At1g20823 5.74 -1.35 0.016068 0.107243 gi_15027984_gb_AY045849.1-- 267451_at putative AP2 domain transcription factor At2g33710 5.72 -1.17 0.015334 0.725714 260411_at hypothetical protein similar to GB: AAB61488 At1g69890 5.71 -1.29 0.011204 0.168552 [Arabidopsis thaliana]; supported by full-length cDNA: Ceres: 34864. 254592_at heat shock transcription factor-like protein heat At4g18880 5.7 -1.08 0.009829 0.552909 shock transcription factor, Zea mays, PIR2: S61448 264000_at putative mitochondrial dicarboxylate carrier protein; At2g22500 5.68 -1.18 0.004964 0.182153 supported by full-length cDNA: Ceres: 20723. 263475_at Expressed protein; supported by full-length cDNA: At2g31945 5.63 1 0.00655 0.971652 Ceres: 258917. 254408_at serine/threonine kinase-like protein serine/threonine At4g21390 5.63 1.2 0.003477 0.605633 kinase BRLK, Brassica oleracea, gb: Y12531 245209_at putative protein similarity to predicted protein, At5g12340 5.63 -1.23 0.004077 0.532051 Arabidopsis thaliana 259629_at disease resistance protein contains domains At1g56510 5.61 -1.13 0.009583 0.608416 associated with disease resistance genes in plants: TIR/NB-ARC/LRR 247655_at zinc finger protein Zat12; supported by full-length At5g59820 5.56 1.2 0.004335 0.099425 cDNA: Ceres: 40576. 266834_s_at putative protein phosphatase 2C At2g30020 5.52 -1.03 0.005778 0.730603 256181_at light repressible receptor protein kinase, putative At1g51820 5.51 -1.08 0.002365 0.605128 similar to light repressible receptor protein kinase GI: 1321686 from (Arabidopsis thaliana) 251705_at DNA-binding protein-like DNA-binding protein 4 At3g56400 5.5 -1.03 0.00667 0.83389 WRKY4-Nicotiana tabacum, EMBL: AF193771; supported by full-length cDNA: Ceres: 34847. 251097_at receptor like protein kinase receptor like protein At5g01560 5.48 -1.09 0.00945 0.858858 kinase-Arabidopsis thaliana, EMBL: ATLECGENE 248392_at integral membrane protein-like At5g52050 5.45 -1.21 0.005162 0.477438 254158_at putative protein dihydrofolate reductase- At4g24380 5.44 -1.17 0.013347 0.342417 Schizosaccharomyces pombe, PID: e1320950; supported by full-length cDNA: Ceres: 27155. 260406_at putative glutathione transferase similar to glutathione At1g69920 5.41 2.07 0.009635 0.082596 transferase GB: CAA09188 [Alopecurus myosuroides] 254241_at serine/threonine kinase-like protein serine/threonine At4g23190 5.37 1.09 0.001802 0.566443 kinase, Brassica oleracea
265674_at unknown protein; supported by full-length cDNA: At2g32190 5.3 1.24 0.013333 0.440285 Ceres: 40344. 264757_at receptor protein kinase (IRK1), putative similar to At1g61360 5.28 -1.05 0.002166 0.73136 receptor protein kinase (IRK1) GI: 836953 from [Ipomoea trifida] 248875_at disease resistance protein-like At5g46470 5.28 -1.01 0.004999 0.943089 247708_at putative protein COP1-interacting protein CIP8, At5g59550 5.28 -1.21 0.003861 0.156044 Arabidopsis thaliana, EMBL: AF162150; supported by cDNA: gi_15450686_gb_AY052711.1-- 260239_at putative receptor protein kinase similar to At1g74360 5.26 1.27 0.014165 0.212238 brassinosteroid insensitive 1 GB: AAC49810 (putative receptor protein kinase); contains Pfam profiles: PF00560 Leucine Rich Repeat (17 repeats), PF00069 Eukaryotic protein kinase domain; supported by cDNA: gi_158 255549_at predicted protein of unknown function At4g01950 5.23 -1.02 0.009729 0.893458 266992_at similar to Mlo proteins from H. vulgare; supported At2g39200 5.21 -1.12 0.008101 0.282992 by cDNA: gi_14091593_gb_AF369573.1_AF369573 261973_at hypothetical protein predicted by genemark.hmm At1g64610 5.19 -1.09 0.005786 0.674167 254242_at serine/threonine kinase-like protein serine/ At4g23200 5.19 1.03 0.007882 0.840853 threonine kinase, Brassica oleracea 260477_at Ser/Thr protein kinase isolog At1g11050 5.15 -1.34 0.029135 0.243484 265670_s_at unknown protein; supported by full-length cDNA: At2g32210 5.07 1.19 0.014682 0.138268 Ceres: 31665. 265199_s_at putative glucosyl transferase At2g36770 5.07 1.33 0.003771 0.194926 247493_at copine-like protein copine I, Homo sapiens, At5g61900 5.07 1.04 0.003077 0.714944 EMBL: HSU83246; supported by full-length cDNA: Ceres: 146738. 265737_at putative phosphatidic acid phosphatase; supported At2g01180 5.04 -1.05 0.00382 0.74519 by full-length cDNA: Ceres: 19163. 260243_at hypothetical protein similar to putative protein At1g63720 5.01 1.07 0.019639 0.772243 GB: CAA18164 [Arabidopsis thaliana]; supported by cDNA: gi_13878144_gb_AF370335.1_AF370335 252045_at putative protein arm repeat containing protein ARC1- At3g52450 5.01 1.25 0.012091 0.125673 Brassica napus, PID: g2558938 250153_at putative protein TMV response-related gene product, At5g15130 5 1.05 0.011689 0.809857 Nicotiana tabacum, EMBL: AB024510 247047_at putative protein contains similarity to unknown At5g66650 4.98 -1.01 0.006647 0.888192 protein (gb AAC17084.1); supported by cDNA: gi_14596230_gb_AY042903.1-- 261476_at hypothetical protein contains similarity to alpha- At1g14480 4.97 1.14 0.02789 0.562278 latroinsectotoxin precursor GI: 9537 from [Latrodectus tredecimguttatus] 247205_at unknown protein; supported by full-length cDNA: At5g64890 4.96 1.59 0.010532 0.389292 Ceres: 9242. 261450_s_at O-methyltransferase, putative similar to At1g21110 4.95 1.5 0.02203 0.137219 GB: AAF28353 from [Fragaria × ananassa] 252474_at putative protein several hypothetical proteins- At3g46620 4.94 -1.06 0.006633 0.705845 Arabidopsis thaliana 257840_at protein kinase, putative contains Pfam profile: At3g25250 4.93 1.19 0.013824 0.496857 PF00069 Eukaryotic protein kinase domain 248964_at cytochrome P450 At5g45340 4.93 -1.52 0.003815 0.013613 247071_at putative protein similar to unknown protein (emb At5g66640 4.92 -1.02 0.010559 0.987089 CAB16816.1) 246270_at putative protein At4g36500 4.92 -1.2 0.002823 0.230335 261033_at unknown protein; supported by full-length cDNA: At1g17380 4.84 -1.01 0.017643 0.96188 Ceres: 37370. 260261_at unknown protein At1g68450 4.78 -1.03 0.006946 0.882923 249485_at receptor protein kinase-like protein receptor-protein At5g39020 4.74 1.03 0.002268 0.823569 kinase-like protein, Arabidopsis thaliana, PIR: T45786 256487_at disease resistance gene, putative similar to downy At1g31540 4.73 1.14 0.01107 0.679977 mildew resistance protein RPP5 [Arabidopsis thaliana] GI: 6449046 249983_at putative protein S-receptor kinase PK3 precursor, At5g18470 4.69 1.03 0.006021 0.801393 maize, PIR: T02753; supported by full-length cDNA: Ceres: 154037. 258682_at putative ribosomal-protein S6 kinase (ATPK19) At3g08720 4.68 1.12 0.009464 0.260404 identical to putative ribosomal-protein S6 kinase (ATPK19) GB: D42061 [Arabidopsis thaliana] (FEBS Lett. 358 (2), 199-204 (1995)); supported by cDNA: gi_15292784_gb_AY050826.1-- 254487_at calcium-binding protein-like calcium-binding At4g20780 4.63 -1.43 0.015022 0.176224 protein, Solanum tuberosum, gb: L02830 265728_at hypothetical protein predicted by genscan At2g31990 4.62 -1.14 0.025876 0.616683 258792_at hypothetical protein predicted by At3g04640 4.62 -1.08 0.003809 0.521094 genefinder; supported by full-length cDNA: Ceres: 8992. 253535_at putaive DNA-binding protein DNA-binding protein At4g31550 4.62 -1.17 0.001616 0.072643 WRKY3-Petroselinum crispum, PIR2: S72445; supported by full-length cDNA: Ceres: 11953. 257751_at hypothetical protein predicted by At3g18690 4.6 -1.01 0.006195 0.939184 genemark.hmm; supported by full-length cDNA: Ceres: 104278. 261367_at protein kinase, putative similar to many predicted At1g53080 4.59 1.31 0.008723 0.439817 protein kinases 247240_at putative protein strong similarity to unknown protein At5g64660 4.57 -1.08 0.004191 0.392312 (emb|CAB89350.1) 261526_at protein kinase identical to protein kinase GI: 2852447 At1g14370 4.56 -1.08 0.004758 0.475696 from [Arabidopsis thaliana]; supported by cDNA: gi_2852446_dbj_D88206.1_D88206 254948_at putative protein various predicted proteins, At4g11000 4.55 1.03 0.020674 0.901116 Arabidopsis thaliana 245119_at unknown protein; supported by cDNA: At2g41640 4.54 -1.2 0.013528 0.323955 gi_16930450_gb_AF419579.1_AF419579 248319_at unknown protein At5g52710 4.5 -1.19 0.022646 0.498394 245765_at hypothetical protein similar to putative disease At1g33600 4.5 -1.01 0.00753 0.943437 resistance protein GB: AAC14512 GI: 2739389 from [Arabidopsis thaliana] 248821_at protein serine threonine kinase-like At5g47070 4.49 1.13 0.005807 0.220356 245272_at hypothetical protein; supported by cDNA: At4g17250 4.49 -1 0.016447 0.969266 gi_16323154_gb_AY057681.1-- 255595_at putative chitinase similar to peanut type II chitinase, At4g01700 4.48 1.09 0.009232 0.455046 GenBank accession number X82329, E.C. 3.2.1.14 249918_at putative protein predicted protein, Arabidopsis At5g19240 4.48 1.11 0.005605 0.490746 thaliana 263565_at unknown protein At2g15390 4.45 -1.28 0.011298 0.375612 261713_at protein kinase, putative identical to bHLH protein At1g32640 4.43 1.12 0.002007 0.392042 GB: CAA67885 GI: 1465368 from [Arabidopsis thaliana]; supported by cDNA: gi_14335047_gb_AY037203.1-- 262772_at puative calcium-transporting ATPase similar to At1g13210 4.4 -1.06 0.004192 0.641809 gb|AF038007 FIC1 gene from Homo sapiens and is a member of the PF|00122 E1-E2 ATPase family. ESTs gb|T45045 and gb|AA394473 come from this gene 258364_at unknown protein At3g14225 4.4 -1.49 0.013195 0.305266 257022_at zinc finger protein, putative similar to Cys2/His2- At3g19580 4.39 -1.04 0.01073 0.818188 type zinc finger protein 2 GB: BAA85107 from [Arabidopsis thaliana]; supported by cDNA: gi_15028256_gb_AY046043.1-- 252053_at syntaxin-like protein synt4; supported by full-length At3g52400 4.38 1.02 0.002866 0.837782 cDNA: Ceres: 37248. 250695_at lectin-like protein kinase At5g06740 4.38 -1.34 0.030543 0.436678 246293_at SigA binding protein; supported by cDNA: At3g56710 4.38 -1.01 0.005488 0.98387 gi_14596086_gb_AY042831.1-- 249032_at putative protein contains similarity to disease At5g44910 4.37 1.06 0.010921 0.589391 resistance protein 265189_at unknown protein; supported by cDNA: At1g23840 4.34 1.12 0.020118 0.585186 gi_14335017_gb_AY037188.1-- 265668_at putative alanine acetyl transferase; supported by At2g32020 4.31 1.45 0.006627 0.053107 full-length cDNA: Ceres: 21201. 264232_at putative protein kinase Pfam HMM hit: Eukaryotic At1g67470 4.3 -1.07 0.003961 0.651045 protein kinase domain; identical to GB: AAC18787 (Arabidopsis thaliana) 263948_at similar to harpin-induced protein hin1 from tobacco; At2g35980 4.28 1.34 0.007735 0.319605 supported by full-length cDNA: Ceres: 26418. 261748_at hypothetical protein predicted by At1g76070 4.27 -1.05 0.034903 0.781675 genemark.hmm; supported by full-length cDNA: Ceres: 39494. 252278_at NAC2-like protein NAC2-Arabidopsis thaliana, At3g49530 4.25 -1.01 0.001287 0.915747 EMBL: AF201456; supported by cDNA: gi_16604578_gb_AY059734.1-- 247137_at calcium-dependent protein kinase; supported by At5g66210 4.23 -1.01 0.004474 0.902625 full-length cDNA: Ceres: 18901. 255568_at putative DNA-binding protein; supported by At4g01250 4.21 -1.2 0.010495 0.218487 cDNA: gi_15028172_gb_AY045909.1-- 259479_at Expressed protein; supported by full-length cDNA: At1g19020 4.2 1.23 0.002707 0.175614 Ceres: 31015. 245247_at scarecrow-like 13 (SCL13); supported by cDNA: At4g17230 4.2 1.06 0.010533 0.625637 gi_16930432_gb_AF419570.1_AF419570 252470_at protein kinase 6-like protein protein kinase 6- At3g46930 4.19 1.13 0.012875 0.362838 Glycine max, PIR2: S29851 256050_at leucine zipper protein, putative similar to leucine At1g07000 4.16 1.04 0.018298 0.855313 zipper protein GI: 10177020 from [Arabidopsis thaliana] 261405_at unknown protein; supported by full-length cDNA: At1g18740 4.15 -1.11 0.00951 0.382476 Ceres: 40753. 267288_at similar to cold acclimation protein WCOR413 At2g23680 4.12 1.06 0.026303 0.758149 [Triticum aestivum] 252592_at mitogen-activated protein kinase 3; supported by At3g45640 4.12 -1.15 0.004807 0.119458 cDNA: gi_14423447_gb_AF386961.1_AF386961 247125_at putative protein contains similarity to unknown At5g66070 4.11 1 0.001239 0.979764 protein (gb|AAF18680.1) 265184_at unknown protein; supported by full-length cDNA: At1g23710 4.09 -1.18 0.014497 0.24269 Ceres: 36437. 247773_at putative protein At5g58630 4.09 -1.08 0.006176 0.825067 263478_at putative receptor-like protein kinase; supported by At2g31880 4.08 1.14 0.00624 0.158364 cDNA: gi_16648754_gb_AY058153.1-- 251910_at serine/threonine-specific kinase like protein At3g53810 4.05 -1.02 0.002869 0.843094 serine/threonine-specific kinase (EC 2.7.1.--) precursor-Arabidopsis thaliana, PIR: S68589 245662_at hypothetical protein predicted by genemark.hmm At1g28190 4.04 -1.23 0.0328 0.44426 259997_at unknown protein similar to N- At1g67880 4.03 1 0.005767 0.973619 acetylglucosaminyltransferase III GB: AAC53064 [Mus musculus] 252179_at putative protein UDP-glucose: (glucosyl) LPS At3g50760 4.03
-1.04 0.00304 0.802486 alpha1,3-glucosyltransferase WaaO, E. coli, EMBL: AF019746 252928_at putative protein more than 30 predicted proteins, At4g38940 4.01 1.07 0.000729 0.325455 Arabidopsis; supported by full-length cDNA: Ceres: 40069. 251832_at putative protein tomato leucine zipper-containing At3g55150 4.01 1.41 0.010257 0.134388 protein, Lycopersicon esculentum, PIR: S21495 266396_at unknown protein At2g38790 4 1.05 0.027395 0.850892 259400_at receptor-like protein kinase, putative similar to At1g17750 3.97 -1.02 0.042252 0.932069 receptor-like protein kinase INRPK1 GI: 1684913 from [Ipomoea nil] 255654_at Similar to receptor kinase At4g00970 3.97 -1.11 0.010838 0.737951 254587_at resistance protein RPP5-like downy mildew At4g19520 3.97 -1.05 0.00768 0.89806 resistance protein RPP5, Arabidopsis thaliana, PATX: G2109275 255753_at myb factor, putative similar to myb factor At1g18570 3.95 1.03 0.004424 0.830522 GI: 1946266 from [Oryza sativa]; supported by cDNA: gi_3941465_gb_AF062887.1_AF062887 246532_at putative protein beta-glucan-elicitor receptor- At5g15870 3.94 -1.02 0.015841 0.913394 Glycine max, EMBL: D78510 246631_at unknown protein; supported by full-length cDNA: At1g50740 3.93 1.04 0.006841 0.56351 Ceres: 34587. 252533_at putative protein predicted proteins, Arabidopsis At3g46110 3.9 1.02 0.017185 0.893955 thaliana 267384_at unknown protein highly similar to At2g44370 3.88 1.08 0.005016 0.736235 GP|2435515|AF024504 258650_at putative protein kinase similar to protein kinase At3g09830 3.88 1.11 0.012936 0.571912 (APK1A) GB: Q06548 [Arabidopsis thaliana]; contains Pfam profile: PF00069 Eukaryotic protein kinase domain 249339_at putative protein similar to unknown protein At5g41100 3.88 -1.05 0.004061 0.72083 (gb|AAB80666.1) 248794_at ethylene responsive element binding factor 2 At5g47220 3.87 -1.23 0.011156 0.098663 (ATERF2) (sp|O80338); supported by full-length cDNA: Ceres: 3012. 245457_s_at disease resistance RPP5 like protein At4g16960 3.86 1.18 0.010259 0.375561 248316_at putative protein similar to unknown protein At5g52670 3.84 -1.03 0.006334 0.875191 (emb|CAA71173.1) 253046_at cytochrome P450-like protein cytochrome P450, At4g37370 3.83 2.17 0.019261 0.016853 Glycyrrhiza echinata, AB001379; supported by full- length cDNA: Ceres: 253698. 262374_s_at flax rust resistance protein, putative similar to flax At1g72930 3.81 1.03 0.004406 0.567071 rust resistance protein GI: 4588066 from [Linumusitatissimum]; supported by full-length cDNA: Ceres: 2795. 258537_at putative disease resistance protein similar to disease At3g04210 3.81 1.09 0.005941 0.472196 resistance protein RPP1-WsC GB: AAC72979 [Arabidopsis thaliana]; supported by cDNA: gi_15982829_gb_AY057522.1-- 252648_at disease resistance protein homolog disease At3g44630 3.81 -1.23 0.007177 0.068548 resistance protein RPP1-WsB-Arabidopsis thaliana, EMBL: AF098963 247913_at unknown protein At5g57510 3.81 1.12 0.009476 0.608703 267411_at putative disease resistance protein At2g34930 3.8 -1.06 0.015151 0.825604 265440_at pEARLI 4 protein Same as GB: L43081; supported At2g20960 3.8 -1.08 0.001968 0.382136 by cDNA: gi_871781_gb_L43081.1_ATHPEARA 245252_at ethylene responsive element binding factor 1 At4g17500 3.8 -1.47 0.008058 0.087956 (frameshift !); supported by cDNA: gi_3434966_dbj_AB008103.1_AB008103 259033_at putative pectinacetylesterase similar to At3g09410 3.79 1.64 0.003796 0.052828 pectinacetylesterase precursor GB: CAA67728 [Vigna radiata] 246233_at putative protein At4g36550 3.79 -1.43 0.028755 0.228285 255599_at cyclic nucleotide gated channel (CNGC4) like At4g01010 3.78 -1.02 0.00606 0.91649 protein Arabidopsis thaliana cyclic nucleotide gated channel (CNGC4), PID: g4378659 262901_at hypothetical protein predicted by genemark.hmm At1g59910 3.77 -1.08 0.006294 0.540378 259952_at putative disease resistance protein similar to Cf-4 At1g71400 3.74 1.08 0.001393 0.408545 GB: CAA05268 from (Lycopersicon hirsutum) 246858_at receptor-like protein kinase-like receptor-like At5g25930 3.73 1.02 0.015786 0.964753 protein kinase 5, Arabidopsis thaliana, PIR: S27756 250435_at putative protein various predicted proteins, At5g10380 3.72 1.22 0.007856 0.106321 Arabidopsis thaliana 261650_at envelope Ca2+-ATPase identical to envelope Ca2+- At1g27770 3.71 1.05 0.00839 0.580228 ATPase GB: AAD10212 GI: 516118 from (Arabidopsis thaliana); supported by cDNA: gi_493621_dbj_D13983.1_ATHRCECAA 252906_at putative gamma-glutamyltransferase gamma- At4g39640 3.71 1.07 0.012355 0.562612 glutamyltransferase, Arabidopsis thaliana, PIR2: S58286 251636_at calcium-dependent protein kinase calcium- At3g57530 3.71 -1.26 0.016722 0.11982 dependent protein kinase-Fragaria × ananassa, EMBL: AF035944 247426_at putative protein contains similarity to calmodulin- At5g62570 3.67 1.02 0.018802 0.878551 binding protein 266685_at hypothetical protein At2g19710 3.66 -1 0.018487 0.952139 249903_at disease resistance protein-like At5g22690 3.65 -1.04 0.010635 0.754135 247925_at TCH4 protein (gb|AAA92363.1); supported by At5g57560 3.65 -1.28 0.003003 0.132214 cDNA: gi_14194112_gb_AF367262.1_AF367262 248611_at putative protein contains similarity to WRKY-type At5g49520 3.63 -1.45 0.010966 0.13904 DNA-binding protein 265221_s_at putative glutamate decarboxylase; supported by At2g02010 3.62 -1.12 0.01727 0.698419 cDNA: gi_13605709_gb_AF361836.1_AF361836 259792_at unknown protein; supported by cDNA: At1g29690 3.62 -1.05 0.013953 0.685925 gi_15809819_gb_AY054177.1-- 256576_at zinc finger protein (PMZ), putative identical to At3g28210 3.62 1.34 0.019514 0.107277 putative zinc finger protein (PMZ) GB: AAD37511 GI: 5006473 [Arabidopsis thaliana] 254784_at growth factor like protein antisense basic fibroblast At4g12720 3.62 1.06 0.012904 0.638871 growth factor GFG-Rattus norvegicus, PID: g1518635; supported by full-length cDNA: Ceres: 148575. 247177_at unknown protein; supported by cDNA: At5g65300 3.62 1.1 0.004863 0.387978 gi_13877834_gb_AF370180.1_AF370180 245226_at gene_id: K17E7.15~unknown protein At3g29970 3.6 1.76 0.01017 0.066452 256756_at ATPase II, putative similar to GB: AAD34706 from At3g25610 3.59 -1.01 0.009255 0.929097 [Homo sapiens] (Biochem. Biophys. Res. Commun. 257 (2), 333-339 (1999)) 253140_at RING-H2 finger protein RHA3b; supported by full- At4g35480 3.56 -1.04 0.013391 0.651703 length cDNA: Ceres: 31493. 250289_at putative protein; supported by full-length cDNA: At5g13190 3.56 1.18 0.000966 0.176346 Ceres: 5392. 247811_at leucine zipper-containing protein leucine zipper- At5g58430 3.56 -1.01 0.001344 0.933016 containing protein, Lycopersicon esculentum, PIR: S21495 261899_at cinnamoyl CoA reductase, putative similar to At1g80820 3.55 -1.11 0.01598 0.720481 cinnamoyl CoA reductase GB: AAF43141 GI: 7239228 from [Populus tremuloides]; supported by full-length cDNA: Ceres: 32255. 245866_s_at unknown protein At1g57990 3.55 -1.09 0.011056 0.501011 264867_at unknown protein At1g24150 3.53 -1 0.030643 0.978236 261193_at unknown protein; supported by cDNA: At1g32920 3.53 -1.12 0.009489 0.382199 gi_15450636_gb_AY052686.1-- 261339_at protein kinase, putative similar to many predicted At1g35710 3.51 1.32 0.013195 0.062019 protein kinases 267490_at putative receptor-like protein kinase At2g19130 3.5 1 0.015702 0.997521 259561_at hypothetical protein; supported by cDNA: At1g21250 3.49 1.52 0.005151 0.042781 gi_14532585_gb_AY039917.1-- 263228_at putative reticuline oxidase-like protein similar to At1g30700 3.48 1.07 0.007823 0.648304 GB: P30986 from [Eschscholzia californica] (berberine bridge-forming enzyme), ESTs gb|F19886, gb|Z30784 and gb|Z30785 come from this gene; supported by cDNA: gi_16930506_gb_AF419607.1_AF419607 255627_at Expressed protein; supported by full-length cDNA: At4g00955 3.48 1.08 0.009206 0.72176 Ceres: 93818. 254256_at serine/threonine kinase-like protein serine/threonine At4g23180 3.45 -1.2 0.002919 0.140829 kinase, Brassica oleracea; supported by cDNA: gi_13506744_gb_AF224705.1_AF224705 260135_at calmodulin-related protein similar to GB: P25070 At1g66400 3.44 -1.11 0.013883 0.371779 from [Arabidopsis thaliana], contains Pfam profile: PF00036 EF hand (4 copies); supported by full- length cDNA: Ceres: 95959. 260206_at putative protein kinase contains Pfam profile: At1g70740 3.43 -1.12 0.012329 0.420329 PF00069 Eukaryotic protein kinase domain 259887_at putative protein kinase similar to protein kinase At1g76360 3.42 1.1 0.008975 0.501823 (APK1A); contains Pfam profile: PF00069 Eukaryotic protein kinase domain 262383_at disease resistance protein, putative similar to disease At1g72940 3.41 1.18 0.011942 0.230832 resistance protein GI: 9758876 from [Arabidopsis thaliana] 256177_at protein kinase, putative contains Pfam profile: At1g51620 3.41 1.23 0.01444 0.359679 PF00069: Eukaryotic protein kinase domain 245777_at unknown protein contains similarity to At1g73540 3.41 -1.25 0.026487 0.341823 diphosphoinositol polyphosphate phosphohydrolase GI: 3978224 from [Homo sapiens] 249221_at serine/threonine protein kinase-like protein At5g42440 3.4 -1.02 0.005295 0.883947 245448_at disease resistance RPP5 like protein At4g16860 3.4 -1.15 0.027985 0.375642 254869_at protein kinase-like protein KI domain interacting At4g11890 3.37 2.12 0.007665 0.003284 kinase 1-Zea mays, PIR2: T02053 256755_at calmodulin, putative similar to GB: P07463 from At3g25600 3.37 -1.05 0.007284 0.663209 [Paramecium tetraurelia] (Cell 62 (1), 165-174 (1990)) 264107_s_at putative receptor-like protein kinase At2g13790 3.34 1.16 0.008131 0.293891 266017_at unknown protein; supported by cDNA: At2g18690 3.32 1.36 0.008527 0.108178 gi_14517479_gb_AY039575.1-- 263776_s_at putative cyclic nucleotide-regulated ion channel At2g46440 3.32 1.21 0.026465 0.278033 protein 245193_at F12A21.6 hypothetical protein At1g67810 3.32 1.17 0.00613 0.205789 256522_at unknown protein; supported by full-length cDNA: At1g66160 3.3 -1.22 0.004073 0.074994 Ceres: 35218. 248703_at dermal glycoprotein precursor, extracellular-like At5g48430 3.28 1.09 0.005001 0.574329 260434_at hypothetical protein predicted by genscan+ At1g68330 3.27 -1.14 0.006128 0.614427 252652_at putative chloroplast prephenate dehydratase similar At3g44720 3.23 1.08 0.004759 0.192206 to bacterial PheA gene products 260023_at unknown protein At1g30040 3.21 1.26 0.004354 0.301041 251640_at putative protein; supported by full-length cDNA: At3g57450 3.21 -1.03 0.002724 0.717428 Ceres: 12522.
264314_at unknown protein; supported by cDNA: At1g70420 3.18 1.24 0.00926 0.33473 gi_15010575_gb_AY045589.1-- 262549_at hypothetical protein similar to hypothetical protein At1g31290 3.18 1.36 0.017342 0.141779 GB: AAF24586 GI: 6692121 from [Arabidopsis thaliana] 261459_at O-methyltransferase, putative similar to At1g21100 3.18 1.37 0.006504 0.199125 GB: AAF28353 from [Fragaria × ananassa]; supported by cDNA: gi_15982843_gb_AY057529.1-- 249139_at Cys2/His2-type zinc finger protein 3 At5g43170 3.18 -1.11 0.014619 0.403291 (dbj|BAA85109.1); supported by full-length cDNA: Ceres: 9878. 248980_at putative protein similar to unknown protein At5g45090 3.18 -1.03 0.006572 0.837241 (pir||T04765) 264660_at putative glutamyl-tRNA reductase 2 precursor At1g09940 3.17 -1.02 0.009351 0.857849 similar to GB: P49294 and to A. thaliana HEMA2 (gb|U27118) 254014_at NPR1 like protein regulatory protein NPR1- At4g26120 3.17 1.03 0.021113 0.898299 Arabidopsis thaliana, PID: g1773295 252126_at putative disease resistance protein At3g50950 3.17 1.08 0.00517 0.256863 262228_at protein kinase, putative similar to protein kinase 1 At1g68690 3.16 1.18 0.018754 0.421396 GB: BAA94509 GI: 7573596 from [Populus nigra]; supported by cDNA: gi_14334805_gb_AY035076.1-- 259626_at bZIP transcription factor, putative contains Pfam At1g42990 3.15 1.08 0.006031 0.361959 profile: PF00170: bZIP transcription factor; supported by cDNA: gi_15028322_gb_AY045964.1-- 254063_at receptor kinase-like protein receptor-like protein At4g25390 3.15 -1.09 0.021274 0.509081 kinase, RLK3-Arabidopsis thaliana, PID: e1363211 259443_at chitinase, putative similar to chitinase GI: 1237025 At1g02360 3.14 1.33 0.010757 0.097826 from [Arachis hypogaea] 266615_s_at putative monooxygenase; supported by full-length At2g29720 3.13 -1 0.006073 0.993995 cDNA: Ceres: 34214. 251507_at putative protein CND41, chloroplast nucleoid DNA At3g59080 3.13 -1.26 0.019246 0.076416 binding protein-Nicotiana tabacum, EMBL: D26015; supported by cDNA: gi_15983375_gb_AF424562.1_AF424562 246870_at ferrochelatase-I At5g26030 3.12 -1.03 0.007971 0.563075 261063_at transcription factor scarecrow-like 14, putative At1g07520 3.09 1.05 0.0041 0.648222 similar to GB: AAD24412 from [Arabidopsis thaliana] (Plant J. 18 (1), 111-119 (1999)) 260296_at putative disease resistance protein similar to disease At1g63750 3.07 -1.24 0.035995 0.346342 resistance protein (RPP1-WsC) GB: AAC72979 [Arabidopsis thaliana] 248868_at putative protein similar to unknown protein At5g46780 3.07 1.08 0.012841 0.668687 (gb|AAC61815.1); supported by full-length cDNA: Ceres: 254442. 267069_at unknown protein At2g41010 3.06 -1 0.022932 0.942266 261143_at unknown protein At1g19770 3.06 -1.07 0.003012 0.469481 255116_at receptor protein kinase-like protein receptor protein At4g08850 3.06 1.13 0.013035 0.33618 kinase-like protein-Arabidopsis thaliana, PIR2: T05898 253284_at putative protein hydroxyproline-rich glycoprotein At4g34150 3.05 1.01 0.004615 0.829133 precursor, Nicotiana tabacum, PIR2: S06733; supported by cDNA: gi_15724315_gb_AF412098.1_AF412098 252903_at putative protein various predicted proteins, At4g39570 3.05 -1.05 0.005467 0.697229 Arabidopsis thaliana 254847_at putative phospholipase D-gamma phospholipase D- At4g11850 3.04 -1.01 0.014523 0.911568 gamma-Arabidopsis thaliana, PID: g2653885; supported by cDNA: gi_2653884_gb_AF027408.1_AF027408 251937_at putative protein predicted protein, Arabidopsis At3g53400 3.04 1.04 0.035806 0.858509 thaliana 256366_at protein kinase, putative contains Pfam profile: At1g66880 3.03 1.12 0.002701 0.411044 PF00069: Eukaryotic protein kinase domain 247393_at unknown protein At5g63130 3.03 -1.65 0.018398 0.063566 260556_at putative endochitinase At2g43620 3.02 1.32 0.003455 0.0287 259445_at dioxygenase, putative similar to dioxygenase At1g02400 3.01 1.16 0.012122 0.130623 GI: 1666096 from [Marah macrocarpus] 259298_at putative disease resistance protein similar to Cf-2 At3g05370 3.01 -1.08 0.040444 0.621247 disease resistance protein GB: AAC15780 from [Lycopersicon pimpinellifolium] 257644_at unknown protein; supported by full-length cDNA: At3g25780 3.01 1.19 0.022772 0.336306 Ceres: 3457. 253628_at xyloglucan endo-1,4-beta-D-glucanase-like protein At4g30280 3.01 1.29 0.005842 0.110446 xyloglucan endo-1,4-beta-D-glucanase (EC 3.2.1.--) XTR-3-Arabidopsis thaliana, PIR2: S71222; supported by full-length cDNA: Ceres: 142204. 249072_at putative protein similar to unknown protein At5g44060 3.01 1.08 0.007698 0.56653 (gb|AAD10670.1) 253257_at extra-large G-protein-like extra-large G-protein, At4g34390 3 -1.06 0.004333 0.352585 Arabidopsis thaliana, AF060942 253124_at putative protein unknown protein Arabidopsis At4g36030 3 -1.07 0.016993 0.706449 thaliana, PATX: E248475 250676_at harpin-induced protein-like; supported by cDNA: At5g06320 3 1.02 0.003772 0.798472 gi_9502175_gb_AF264699.1_AF264699 266037_at putative protein kinase contains a protein kinase At2g05940 2.99 1.03 0.011895 0.742301 domain profile (PDOC00100); supported by cDNA: gi_15810412_gb_AY056245.1-- 254314_at extensin-like protein hybrid proline-rich protein, At4g22470 2.98 -1.04 0.013677 0.797081 Zea mays, PIR2: JQ1663 252825_at small GTP-binding protein-like SR1 Nt-rab6, At4g39890 2.97 1.25 0.014269 0.471148 Nicotiana tabacum, L29273; supported by cDNA: gi_14423429_gb_AF386952.1_AF386952 260401_at unknown protein similar to hypothetical protein At1g69840 2.96 1.19 0.013016 0.197702 GB: CAA10289 [Cicer arietinum] 250821_at putative protein similar to unknown protein At5g05190 2.95 -1.11 0.008801 0.532383 (emb|CAB88044.1) 245265_at hypothetical protein; supported by cDNA: At4g14400 2.95 1.34 0.046774 0.092249 gi_15810232_gb_AY056155.1-- 264289_at hypothetical protein similar to hypothetical protein At1g61890 2.94 1.17 0.016735 0.217477 GI: 2894569 from [Arabidopsis thaliana]; supported by cDNA: gi_15028186_gb_AY045916.1-- 259410_at hypothetical protein predicted by genemark.hmm At1g13340 2.94 1.45 0.015002 0.097363 253958_at putative protein RING zinc finger protein, Gallus At4g26400 2.94 1.06 0.002619 0.621194 gallus 249078_at phytochelatin synthase (gb|AAD41794.1); At5g44070 2.94 -1.02 0.008033 0.806261 supported by cDNA: gi_14532653_gb_AY039951.1-- 267293_at hypothetical protein At2g23810 2.93 -1.06 0.004637 0.578539 259992_at putative heat shock transcription factor contains At1g67970 2.93 -1.01 0.006051 0.910383 Pfam profile: PF00447 HSF-type DNA-binding domain; N-terminal portion similar to heat shock transcription factor proteins: GB: CAA74397 [Arabidopsis thaliana], GB: S25478 [Lycopersicon esculentum] 252862_at putative L-ascorbate oxidase L-ascorbate oxidase, At4g39830 2.93 1.13 0.009756 0.383415 Cucumis sativus, PIR1: KSKVAO 249550_at protein kinase-like protein wall-associated kinase 4 At5g38210 2.93 -1.13 0.00676 0.38925 (wak4), Arabidopsis thaliana, EMBL: ATH9695 247279_at arabinogalactan-protein (gb|AAC77823.1); At5g64310 2.93 -1.01 0.00661 0.937671 supported by full-length cDNA: Ceres: 25423. 265450_at hypothetical protein predicted by genefinder At2g46620 2.92 -1.03 0.014924 0.733991 251479_at serine/threonine-specific kinase lecRK1 At3g59700 2.91 -1.08 0.008769 0.515335 precursor, lectin receptor-like 249418_at putative protein predicted protein, Arabidopsis At5g39780 2.91 1.1 0.015458 0.521455 thaliana 266247_at hypothetical protein predicted by genscan At2g27660 2.89 -1.11 0.009688 0.350456 249252_at putative protein contains similarity to unknown At5g42010 2.89 -1.05 0.014073 0.747236 protein (gb|AAF19687.1) 255291_at putative calcium dependent protein kinase At4g04700 2.88 -1.04 0.023496 0.890022 253747_at serine threonine-specific kinase like protein serine At4g29050 2.87 -1.09 0.011457 0.626019 threonine-specific kinase lecRK1-Arabidopsis thaliana, PIR2: S68589 250323_at putative protein hydroxyproline-rich glycoprotein, At5g12880 2.87 1.06 0.009216 0.469664 kidney bean, PIR: A29356 262801_at unknown protein; supported by full-length cDNA: At1g21010 2.86 1.08 0.017653 0.443505 Ceres: 17521. 251061_at putative protein hypothetical protein ARC1- At5g01830 2.86 1.18 0.015743 0.623238 Brassica napus, PIR: T08872 265132_at unknown protein; supported by cDNA: At1g23830 2.84 -1.07 0.017467 0.652241 gi_16604403_gb_AY058100.1-- 260439_at hypothetical protein predicted by At1g68340 2.84 -1.04 0.003917 0.840841 genscan+; supported by full-length cDNA: Ceres: 3385. 260227_at unknown protein similar to hypothetical proteins At1g74450 2.83 -1.16 0.009649 0.269848 GB: AAD39276 [Arabidopsis thaliana], GB: CAB53491 [Oryza sativa]; supported by full- length cDNA: Ceres: 108193. 261453_at O-methyltransferase, putative similar to At1g21130 2.82 -1.15 0.010888 0.513201 GB: AAF28353 from [Fragaria × ananassa]; supported by full-length cDNA: Ceres: 101583. 254432_at reticuline oxidase-like protein reticuline oxidase, At4g20830 2.82 1.19 0.046062 0.572211 Eschscholzia californica, PIR: A41533; supported by cDNA: gi_15983492_gb_AF424621.1_AF424621 253971_at fructose-bisphosphate aldolase-like protein At4g26530 2.82 -1.02 0.016712 0.805977 fructose-bisphosphate aldolase, Arabidopsis thaliana, PIR1: ADMU; supported by full-length cDNA: Ceres: 34690. 262165_at putative acyl-CoA: 1-acylglycerol-3-phosphate At1g75020 2.81 -1.13 0.010295 0.275107 acyltransferase similar to acyl-CoA: 1-acylglycerol- 3-phosphate acyltransferase GB: CAB09138 (Brassica napus); contains Pfam profile: PF01553 Acyltransferase; supported by full-length cDNA: Ceres: 115679. 258275_at unknown protein; supported by full-length cDNA: At3g15760 2.81 -1.09 0.002884 0.259472 Ceres: 8259. 255564_s_at hypothetical protein T15B16.8 At4g01750 2.81 1.28 0.004474 0.364426 253377_at putative protein NBS/LRR disease resistance protein At4g33300 2.81 1.03 0.008788 0.64371 (RFL1)-Arabidopsis thaliana, PID: g3309619 260220_at putative MYB family transcription factor contains At1g74650 2.8 -1.05 0.014801 0.787419 Pfam profile: PF00249 Myb-like DNA-binding domain 256583_at hypothetical protein At3g28850 2.8 1.08 0.009872 0.39554 252193_at R2R3-MYB transcription factor; supported by At3g50060 2.8 -1.67 0.007202 0.02821 cDNA: gi_15983427_gb_AF424588.1_AF424588 247509_at heat shock factor 6 At5g62020 2.8 1.11 0.004718 0.497285 246368_at light repressible receptor protein kinase, putative At1g51890 2.8 1.32 0.007014 0.17566 similar to light repressible receptor protein kinase
GI: 1321686 from [Arabidopsis thaliana] 259507_at unknown protein At1g43910 2.79 1.41 0.005884 0.156323 251769_at receptor kinase-like protein receptor kinase At3g55950 2.79 1.02 0.037029 0.858886 homolog CRINKLY4, maize, PIR: T04108 250335_at lysophospholipase-like protein lysophospholipase At5g11650 2.78 1.07 0.004853 0.539031 homolog LPL1, Oryza sativa, EMBL: AF039531; supported by full-length cDNA: Ceres: 15284. 248134_at putative protein contains similarity to integral At5g54860 2.78 1.09 0.010767 0.465367 membrane protein 246988_at putative protein strong similarity to unknown protein At5g67340 2.78 1.18 0.01807 0.609865 (pir||T00518) 247707_at scarecrow-like 11-like scarecrow-like 11, At5g59450 2.76 -1.06 0.028093 0.649019 Arabidopsis thaliana, EMBL: AF036307; supported by cDNA: gi_14334655_gb_AY035001.1-- 256497_at ORF1, putative similar to ORF1 GI: 457716 from At1g31580 2.75 1.39 0.004888 0.080053 (Arabidopsis thaliana); supported by cDNA: gi_16649160_gb_AY059950.1-- 264008_at unknown protein At2g21120 2.74 -1.01 0.003042 0.876414 264716_at matrix metalloproteinase, putative similar to matrix At1g70170 2.73 -1.02 0.005524 0.873758 metalloproteinase GI: 7159629 from [Cucumis sativus] 261445_at unknown protein; supported by cDNA: At1g28380 2.73 -1.06 0.02128 0.701956 gi_16604598_gb_AY059744.1-- 256968_at unknown protein At3g21070 2.73 -1.14 0.014315 0.494381 256763_at unknown protein At3g16860 2.73 -1.06 0.01099 0.724699 255605_at hypothetical protein At4g01090 2.73 -1.18 0.02941 0.263496 254652_at DNA binding-like protein SPF1 protein, sweet At4g18170 2.73 1.05 0.048645 0.839297 protein, PIR2: S51529 and WRKY protein family, Petroselinum crispum, MNOS: S72443, MNOS: S72444, MNOS: S72445 247532_at putative protein disease resistance protein kinase Pto, At5g61560 2.73 -1.03 0.020053 0.845513 Lycopersiocon esculentum, PIR: A49332 264106_at unknown protein At2g13780 2.71 1.2 0.013998 0.074305 265075_at hypothetical protein similar to embryo-abundant At1g55450 2.7 -1.08 0.016743 0.546091 protein GB: L47672 GI: 1350530 from [Picea glauca]; supported by cDNA: gi_14335021_gb_AY037190.1-- 256793_at unknown protein; supported by full-length cDNA: At3g22160 2.69 -1.09 0.013465 0.391312 Ceres: 8081. 258551_at hypothetical protein predicted by At3g06890 2.68 -1.02 0.016594 0.966946 genscan+; supported by full-length cDNA: Ceres: 262487. 255740_at wall-associated kinase, putative similar to wall- At1g25390 2.68 -1.15 0.012139 0.281008 associated kinase 1 GI: 3549626 from [Arabidopsis thaliana]; supported by cDNA: gi_15529241_gb_AY052245.1-- 246099_at blue copper binding protein; supported by full- At5g20230 2.67 1.7 0.008061 0.011289 length cDNA: Ceres: 7767. 264616_at unknown protein At2g17740 2.67 1 0.022917 0.884668 254042_at xyloglucan endo-1,4-beta-D-glucanase (XTR-6); At4g25810 2.66 1.07 0.002288 0.480301 supported by cDNA: gi_1244757_gb_U43488.1_ATU43488 246289_at putative protein predicted protein At2g41010- At3g56880 2.66 -1.02 0.010884 0.82618 Arabidopsis thaliana; EMBL: AC004261; supported by full-length cDNA: Ceres: 39584. 266792_at putative sucrose/H+ symporter At2g02860 2.65 1.05 0.005194 0.618209 265853_at putative RING zinc finger protein At2g42360 2.64 1.27 0.007875 0.101121 258786_at putative syntaxin contains Pfam profile: PF00804 At3g11820 2.64 1.16 0.005501 0.095192 syntaxin; supported by full-length cDNA: Ceres: 38899. 247940_at phosphatidylserine decarboxylase At5g57190 2.64 -1.08 0.02156 0.742477 257083_s_at non-race specific disease resistance protein, putative At3g20590 2.63 -1.1 0.022335 0.571353 contains non-consensus CT donor splice site at exon 1; potential pseudogene; similar to non-race specific disease resistance protein GB: AAB95208 [Arabidopsis thaliana] 264434_at hypothetical protein predicted by genscan; supported At1g10340 2.61 1.14 0.016538 0.421888 by cDNA: gi_13937239_gb_AF372975.1_AF372975 263804_at putative protein kinase contains a protein kinase At2g40270 2.61 1.02 0.002801 0.766513 domain profile (PDOC00100); supported by full- length cDNA: Ceres: 123911. 249896_at unknown protein; supported by cDNA: At5g22530 2.61 1.12 0.01975 0.439704 gi_14532613_gb_AY039931.1-- 249459_at peroxidase ATP24a At5g39580 2.61 -1.24 0.011537 0.098646 247740_at receptor-like protein kinase precursor-like receptor- At5g58940 2.61 1.11 0.013363 0.45095 like protein kinase precursor, Madagascar periwinkle, PIR: T10060 246931_at putative protein apoptosis-related protein PNAS-4, At5g25170 2.6 1.01 0.003003 0.89146 Homo sapiens, EMBL: AF229834; supported by full- length cDNA: Ceres: 263500. 265713_at putative integral membrane protein At2g03530 2.59 -1.18 0.010284 0.275593 263931_at unknown protein; supported by full-length cDNA: At2g36220 2.59 1.04 0.032332 0.640228 Ceres: 12251. 264834_at unknown protein similar to ESTs gb|AA605440 and At1g03730 2.58 1.02 0.006042 0.863016 gb|H37232; supported by full-length cDNA: Ceres: 30716. 259852_at disulfide bond formation protein, putative similar to At1g72280 2.58 1.24 0.014598 0.356183 GI: 6642925 from [Mus musculus] 252539_at putative protein At3g45730 2.58 1.3 0.009508 0.150314 252378_at receptor kinase-like protein protein kinase Xa21- At3g47570 2.58 1.12 0.02363 0.435178 Oryza sativa, PIR: A57676; supported by cDNA: gi_15810450_gb_AY056264.1-- 251684_at putative protein At3g56410 2.57 1.08 0.023561 0.592162 261719_at hypothetical protein similar to hypothetical protein At1g18380 2.56 1.36 0.016331 0.094821 GB: AAF25996 GI: 6714300 from [Arabidopsis thaliana] 254248_at serine/threonine kinase serine/threonine kinase, At4g23270 2.56 -1.04 0.006144 0.687459 Brassica oleracea 253204_at GTP binding protein beta subunit; supported by At4g34460 2.56 -1.01 0.007411 0.949586 cDNA: gi_15028006_gb_AY045860.1-- 249361_at protein kinase-like protein protein kinase ATN1, At5g40540 2.55 -1 0.004647 0.984046 Arabidopsis thaliana, PIR: S61766 248665_at Expressed protein; supported by full-length cDNA: At5g48655 2.55 1.02 0.009359 0.848153 Ceres: 12974. 253455_at putative protein At4g32020 2.54 -1.01 0.00825 0.888496 248978_at putative protein contains similarity to disease At5g45070 2.54 -1.05 0.030143 0.671649 resistance protein 248870_at putative protein similar to unknown protein At5g46710 2.54 1.03 0.004547 0.48662 (pir||T05076); supported by full-length cDNA: Ceres: 42747. 252170_at hypothetical protein; supported by cDNA: At3g50480 2.53 1.09 0.00647 0.431092 gi_13605735_gb_AF361849.1_AF361849 264636_at hypothetical protein predicted by At1g65490 2.52 1.04 0.019945 0.653146 genemark.hmm; supported by full-length cDNA: Ceres: 2118. 264400_at glucose-6-phosphate/phosphate-translocator At1g61800 2.51 -1.13 0.035853 0.380949 precursor, putative similar to glucose-6- phosphate/phosphate-translocator precursor GI: 2997591 from [Pisum sativum]; supported by cDNA: gi_14596172_gb_AY042874.1-- 245567_at germin precursor oxalate oxidase At4g14630 2.51 -1.12 0.010395 0.322763 264083_at ethylene reponse factor-like AP2 domain At2g31230 2.5 -1.09 0.007348 0.596007 transcription factor 261220_at ER lumen protein-retaining receptor similar to At1g19970 2.5 1.11 0.01247 0.321046 SP: O44017 from [Entamoeba histolytica] 259546_at unknown protein At1g35350 2.49 -1.02 0.009622 0.86289 266101_at unknown protein; supported by cDNA: At2g37940 2.47 1.05 0.006689 0.366431 gi_16604321_gb_AY058059.1-- 262384_at disease resistance protein, putative similar to disease At1g72950 2.47 -1.07 0.017043 0.63375 resistance protein GI: 9758876 from [Arabidopsis thaliana] 251423_at regulatory protein-like regulatory protein preg, At3g60550 2.47 1.04 0.005454 0.86361 Neurospora crassa, PIR: S52974 259312_at putative RING-H2 zinc finger protein ATL6 similar At3g05200 2.46 -1.11 0.020543 0.252889 to GB: AAD33584 from [Arabidopsis thaliana]; supported by cDNA: gi_4928402_gb_AF132016.1_AF132016 267624_at putative protein kinase At2g39660 2.45 -1.1 0.015362 0.313576 266230_at hypothetical protein predicted by genscan and At2g28830 2.45 1.03 0.03586 0.739871 genefinder; supported by cDNA: gi_14334729_gb_AY035038.1-- 260656_at hypothetical protein predicted by genemark.hmm At1g19380 2.45 1.12 0.012154 0.152235 253664_at NADPH-ferrihemoprotein reductase (ATR2) At4g30210 2.45 1.06 0.014893 0.463357 251259_at putative protein phosphoprotein phosphatase (EC At3g62260 2.45 1.09 0.008555 0.495561 3.1.3.16) 1A-alpha-Homo sapiens, PIR: S22423; supported by full-length cDNA: Ceres: 20050. 267357_at putative nematode-resistance protein; supported by At2g40000 2.44 1.16 0.031618 0.266241 full-length cDNA: Ceres: 35056. 254521_at putative protein similar to unknown protein At5g44810 2.44 -1.09 0.041638 0.416086 (gb|AAC79139.1) 263419_at putative protein kinase contains a protein kinase At2g17220 2.43 1.09 0.008341 0.27009 domain profile (PDOC00100); supported by full- length cDNA: Ceres: 13257. 253323_at putative protein protein phosphatase Wip1, Homo At4g33920 2.43 -1.13 0.025096 0.456412 sapiens, PID: g2218063; supported by full-length cDNA: Ceres: 40123. 258983_at putative aminotransferase similar to beta-alanine- At3g08860 2.42 1.14 0.006331 0.051755 pyruvate aminotransferase GB: BAA19549 [Rattus norvegicus], alanine-glyoxylate aminotransferase GB: Q64565 [Rattus norvegicus]; Pfam HMM hit: Aminotransferases class-III pyridoxal-phosphate 249583_at CALMODULIN-RELATED PROTEIN 2, TOUCH- At5g37770 2.42 -1.17 0.006305 0.208762 INDUCED (TCH2); supported by full-length cDNA: Ceres: 25475. 258046_at MAP kinase kinase 5 identical to GB: BAA28831 At3g21220 2.41 1.13 0.013633 0.416118 from [Arabidopsis thaliana]; supported by cDNA: gi_3219272_dbj_AB015316.1_AB015316 250990_at serine/threonine-specific protein kinase NAK; At5g02290 2.41 -1.12 0.012886 0.320809 supported by full-length cDNA: Ceres: 27477. 249423_at Expressed protein; supported by full-length cDNA: At5g39785 2.41 -1.13 0.026115 0.657212 Ceres: 118847. 248814_at putative protein similar to unknown protein At5g46910 2.4 -1.03 0.007949 0.787827 (pir||T06699) 254204_at putative protein CGI-58 protein-Homo At4g24160 2.38 -1.04 0.010591 0.590709 sapiens, PID: g4929585 252485_at disease resistance protein RPP13-like protein At3g46530 2.37 -1.05 0.012107 0.677972 disease resistance protein RPP8-Arabidopsis thaliana, EMBL: AF089710; supported by cDNA: gi_14334999_gb_AY037179.1-- 265620_at unknown protein At2g27310 2.35 -1.2 0.049206 0.345125 264756_at receptor protein kinase (IRK1), putative similar to At1g61370 2.35 -1.07 0.010494 0.634376 receptor protein kinase (IRK1) GI: 836953 from
[Ipomoea trifida] 266993_at nodulin-like protein; supported by cDNA: At2g39210 2.33 1.12 0.017636 0.371447 gi_16930478_gb_AF419593.1_AF419593 256735_at hypothetical protein predicted by genemark.hmm At3g29400 2.33 -1.12 0.006192 0.192654 256425_at disease resistance protein, putative similar to disease At1g33560 2.33 1.04 0.010206 0.488005 resistance protein RPP1-WsB GB: BAB01321 GI: 9279731 from (Arabidopsis thaliana) 250829_at disease resistance-like protein rpp8, Arabidopsis At5g04720 2.33 -1.08 0.013721 0.38143 thaliana, EMBL: AF089711; supported by cDNA: gi_15292720_gb_AY050794.1-- 248698_at receptor-like protein kinase; supported by cDNA: At5g48380 2.33 1.13 0.021685 0.326459 gi_13605826_gb_AF367312.1_AF367312 247594_at putative protein farnesylated protein GMFP5, At5g60800 2.33 1.37 0.027382 0.054007 Glycine max, EMBL: U64916 266166_at putative glucosyltransferase; supported by full- At2g28080 2.32 -1.05 0.018474 0.764619 length cDNA: Ceres: 13761. 262745_at lipase, putative contains Pfam profile: PF00657 At1g28600 2.32 -1.12 0.015545 0.267784 Lipase/Acylhydrolase with GDSL-like motif; supported by full-length cDNA: Ceres: 37307. 257407_at unknown protein At1g27100 2.32 -1.19 0.009875 0.068971 258282_at unknown protein At3g26910 2.31 1.14 0.003241 0.168885 252373_at disease resistance protein EDS1; supported by At3g48090 2.31 -1.07 0.011996 0.442811 cDNA: gi_15028150_gb_AY046025.1-- 250956_at putative protein At5g03210 2.31 -1.02 0.03092 0.993512 248851_s_at disease resistance protein-like; supported by cDNA: At5g46490 2.3 1.09 0.009234 0.638209 gi_16323098_gb_AY057653.1-- 254924_at MAP kinase (ATMPK5) possible internal deletion At4g11330 2.29 -1.14 0.010478 0.330061 at position 161, missing one A residue; reference GI: 457401; supported by cDNA: gi_457401_dbj_D21841.1_ATHATMPK5 250279_at ABA-responsive protein-like ABA-responsive At5g13200 2.29 -1.11 0.021853 0.295383 protein, Hordeum vulgare, EMBL: AF026538 263221_at UDP-galactose 4-epimerase-like protein similar to At1g30620 2.28 1.23 0.015145 0.343241 proteins from many bacterial species including [Bacillus subtilis] and [Methanobacterium thermoautotrophicum] 261718_at wall-associated kinase, putative similar to wall- At1g18390 2.28 1.07 0.008635 0.505026 associated kinase 2 GB: CAB42872 GI: 4826399 from [Arabidopsis thaliana] 250398_at putative protein predicted proteins, Arabidopsis At5g11000 2.28 1.16 0.012015 0.299864 thaliana; supported by full-length cDNA: Ceres: 263168. 256922_at hypothetical protein contains similarity to flavonol At3g19010 2.27 -1.04 0.029488 0.80973 synthase (FLS) GB: Q41452 from [Solanum tuberosum], contains Pfam profile: PF00671 Iron/Ascorbate oxidoreductase family; supported by full-length cDNA: Ceres: 41506. 267530_at putative receptor-like protein kinase At2g41890 2.26 -1.11 0.028311 0.389366 256627_at unknown protein; supported by cDNA: At3g19970 2.26 1.05 0.015238 0.715164 gi_14532501_gb_AY039875.1-- 255880_at hypothetical protein predicted by genscan+ At1g67060 2.26 -1.01 0.015412 0.926607 254660_at receptor serine/threonine kinase-like protein At4g18250 2.26 -1.06 0.024187 0.802259 receptor serine/threonine kinase PR5K, PATCHX: G1235680 264528_at hypothetical protein similar to Human XE169 At1g30810 2.25 1.03 0.00644 0.758062 protein (gi|3033385); similar to EST gb|T88128 257784_at geranylgeranylated protein, putative similar to At3g26970 2.25 1.17 0.00182 0.287058 ATGP4 GB: AAD00115 from [Arabidopsis thaliana] 255344_s_at putative receptor-like protein kinase At4g04540 2.25 1.01 0.022152 0.792265 255080_at arabinogalactan-protein homolog arabinogalactan- At4g09030 2.25 -1.04 0.036604 0.768432 protein-Arabidopsis thaliana, PID: g3883126; supported by cDNA: gi_10880496_gb_AF195891.1_AF195891 259325_at unknown protein At3g05320 2.24 -1.15 0.016669 0.38111 252851_at putative protein CLATHRIN COAT ASSEMBLY At4g40080 2.24 -1.08 0.014274 0.543837 PROTEIN AP180-Mus musculus, SWISSPROT: Q61548; supported by full-length cDNA: Ceres: 8970. 257071_at unknown protein; supported by cDNA: At3g28180 2.23 1.04 0.014928 0.697835 gi_15810494_gb_AY056286.1-- 253476_at S-receptor kinase-like protein serine/threonine- At4g32300 2.23 -1.04 0.009096 0.440827 specific protein kinase PK10 precursor, Oryza sativa, PIR2: S50767 254292_at putative protein At4g23030 2.22 1.13 0.007331 0.570308 249393_at disease resistance-like protein resistance gene Cf-4, At5g40170 2.22 -1.04 0.016127 0.647441 Lycopersicon hirsutum, EMBL: LHJ002235 249320_at disease resistance protein-like non-consensus TT At5g40910 2.22 1.13 0.049144 0.285783 donor splice site at exon 1 246327_at receptor-like serine/threonine kinase, putative similar At1g16670 2.22 1.02 0.008469 0.775987 to receptor-like serine/threonine kinase GI: 2465923 from [Arabidopsis thaliana]; supported by cDNA: gi_16649102_gb_AY059921.1-- 267537_at putative guanylate kinase; supported by cDNA: At2g41880 2.21 -1.02 0.012268 0.883883 gi_7861794_gb_AF204675.1_AF204675 251987_at CYTOCHROME P450 71B5; supported by cDNA: At3g53280 2.21 -1.27 0.010305 0.155225 gi_3164131_dbj_D78601.1_D78601 248981_at regulatory protein NPR1-like; transcription factor At5g45110 2.21 1.11 0.020899 0.556465 inhibitor I kappa B-like 265611_at unknown protein; supported by full-length cDNA: At2g25510 2.2 -1.04 0.010382 0.533642 Ceres: 10730. 259071_at unknown protein similar to hin1 GB: CAA68848 At3g11650 2.2 -1.02 0.008823 0.776008 [Nicotiana tabacum]; supported by cDNA: gi_9502173_gb_AF264698.1_AF264698 249029_at disease resistance protein-like At5g44870 2.2 1.01 0.033247 0.925701 265648_at putative beta-1,3-glucanase; supported by full- At2g27500 2.19 -1.07 0.015702 0.521836 length cDNA: Ceres: 1126. 252921_at putative protein DNA damage-inducible protein- At4g39030 2.19 1.57 0.026714 0.071711 Synechocystis sp., PIR2: S77364 266749_at putative protein kinase contains a protein kinase At2g47060 2.18 -1.05 0.013372 0.636302 domain profile (PDOC00100) 266231_at putative protein kinase At2g02220 2.18 -1.01 0.008538 0.969684 254878_at heat shock transcription factor-like protein heat At4g11660 2.18 1.15 0.015925 0.386918 shock transcription factor HSF29, Glycine max, PIR2: S59541 258764_at putative pectinesterase contains similarity to At3g10720 2.17 -1 0.01345 0.975443 pectinesterase GB: AAB57671 [Citrus sinensis] 266975_at hypothetical protein predicted by grail At2g39380 2.16 1.09 0.019477 0.60417 254921_at putative protein hypothetical protein F16G20.230- At4g11300 2.16 -1.01 0.016335 0.995903 Arabidopsis thaliana, PIR2: T05391; supported by full-length cDNA: Ceres: 17771. 259937_s_at putative ABC transporter contains Pfam profile: At1g71330 2.14 1.25 0.009288 0.060426 PF00005 ABC transporter 255524_at hypothetical protein similar to pectinesterase At4g02330 2.14 -1.08 0.006633 0.352183 250018_at putative protein similar to unknown protein At5g18150 2.14 -1.05 0.005427 0.652846 (emb|CAB87627.1) 249987_at putative protein predicted proteins, Arabidopsis At5g18490 2.14 1.03 0.016862 0.931777 thaliana; supported by full-length cDNA: Ceres: 32414. 265722_at putative chlorophyll a/b binding protein; supported At2g40100 2.13 1.35 0.029801 0.022089 by full-length cDNA: Ceres: 6454. 262540_at hypothetical protein predicted by genemark.hmm At1g34260 2.13 1.1 0.022736 0.377889 264767_at hypothetical protein similar to putative At1g61380 2.12 1.15 0.012495 0.215443 serine/threonine kinase GI: 4585880 from [Arabidopsis thaliana]; supported by full-length cDNA: Ceres: 13461. 251192_at alpha galactosyltransferase-like protein alpha At3g62720 2.12 -1.11 0.0119 0.381401 galactosyltransferase-Trigonella foenum-graecum, EMBL: TFO245478; supported by cDNA: gi_15983425_gb_AF424587.1_AF424587 249984_at putative protein rsc43, Dictyostelium discoideum, At5g18400 2.12 1.05 0.006326 0.639984 EMBL: AF011338; supported by full-length cDNA: Ceres: 6084. 249237_at putative protein similar to unknown protein At5g42050 2.12 1.01 0.019097 0.90689 (sp|P37707); supported by full-length cDNA: Ceres: 6903. 249021_at putative protein similar to unknown protein At5g44820 2.12 -1.03 0.014955 0.780757 (pir||T04881) 266452_at hypothetical protein predicted by genscan; supported At2g43320 2.11 1.01 0.010229 0.93189 by cDNA: gi_14517475_gb_AY039573.1-- 266168_at putative protease inhibitor; supported by full-length At2g38870 2.11 1.07 0.011954 0.24411 cDNA: Ceres: 11662. 257264_at hypothetical protein contains Pfam profile: PF01657 At3g22060 2.11 1.41 0.033325 0.224965 Domain of unknown function; supported by cDNA: gi_14334417_gb_AY034900.1-- 252133_at hypothetical protein hypothetical protein- At3g50900 2.11 -1.16 0.048974 0.506058 Arabidopsis thaliana chromosome 4 AP2 contig, PID: e353223; supported by full-length cDNA: Ceres: 10044. 248230_at putative protein similar to unknown protein At5g53830 2.11 -1.24 0.009004 0.419028 (gb|AAF34839.1); supported by cDNA: gi_13926341_gb_AF372918.1_AF372918 247571_at snap25a; supported by full-length cDNA: At5g61210 2.11 1.1 0.033279 0.210386 Ceres: 14562. 253147_at protein kinase-like protein serine/threonine- At4g35600 2.1 1.1 0.007538 0.342113 specific protein kinase APK1, Arabidopsis thaliana, PIR2: S28615 252976_s_at Phospholipase like protein Arabidopsis thaliana At4g38550 2.1 1.02 0.005166 0.820109 pEARLI 4 mRNA, PID: g871782 260975_at receptor-like serine/threonine kinase, putative At1g53430 2.09 -1.06 0.02462 0.698322 similar to receptor-like serine/threonine kinase GB: AAC50043 GI: 2465923 from [Arabidopsis thaliana] 256799_at unknown protein; supported by cDNA: At3g18560 2.09 1.14 0.021195 0.61081 gi_14190488_gb_AF380644.1_AF380644 246529_at serine/threonine-specific protein kinase-like protein At5g15730 2.09 1.07 0.010794 0.454295 serine/threonine-specific protein kinase NPK15- Nicotiana tabacum; supported by full-length cDNA: Ceres: 25636. 245731_at MAP kinase, putative similar to MAP kinase kinase At1g73500 2.09 -1.17 0.002926 0.06007 5 GI: 3219273 from [Arabidopsis thaliana]; supported by full-length cDNA: Ceres: 112118. 257785_at geranylgeranylated protein, putative similar to At3g26980 2.08 1.12 0.027564 0.130143 ATGP4 GB: AAD00115 from [Arabidopsis thaliana] 251248_at P-glycoprotein-like proetin P-glycoprotein-2- At3g62150 2.08 1.18 0.006349 0.219814 Arabidopsis thaliana, EMBL: Y10228 264841_at putative protein kinase similar to (Z71703), cdc2- At1g03740 2.07 1.03 0.009533 0.802311 like protein kinase; similar to ESTs gb|T20748, gb|T20464, and emb|Z17761; supported by cDNA: gi_14532735_gi_13430451 262360_at receptor protein kinase, putative similar to receptor At1g73080 2.07 1.07 0.049706 0.387954 protein kinase GI: 1389566 from [Arabidopsis
thaliana] 249705_at serine/threonine protein kinase-like At5g35580 2.07 1.06 0.02776 0.584833 259876_at putative DnaJ protein similar to dnaJ-like protein At1g76700 2.06 1.04 0.040672 0.74219 GB: CAA72705 [Arabidopsis thaliana]; Pfam HMM hit: DnaJ, prokaryotic heat shock protein 266316_at unknown protein; supported by cDNA: At2g27080 2.05 1.1 0.008877 0.282687 gi_15450380_gb_AY052291.1-- 262183_at unknown protein At1g77900 2.05 1.11 0.020307 0.383917 260345_at receptor protein kinase, putative similar to receptor At1g69270 2.05 -1.16 0.02512 0.233028 protein kinase GI: 1389566 from (Arabidopsis thaliana); supported by cDNA: gi_4204848_gb_U55875.1_ATU55875 260635_at unknown protein At1g62420 2.04 -1.19 0.00958 0.160716 253780_at protein phosphatase 2C-like protein protein At4g28400 2.04 1.02 0.049077 0.837741 phosphatase 2C-fission yeast, PIR2: S54297; supported by cDNA: gi_16604584_gb_AY059737.1-- 251218_at CP12 protein precursor-like protein CP12 protein At3g62410 2.04 1.05 0.005991 0.528908 precursor, chloroplast-Pisum sativum, PIR: T06562; supported by full-length cDNA: Ceres: 2721. 245641_at Expressed protein; supported by full-length cDNA: At1g25370 2.04 -1.04 0.0479 0.829103 Ceres: 118770. 263915_at hypothetical protein predicted by genscan and At2g36430 2.03 -1.26 0.020455 0.076673 genefinder 254508_at putative protein gene F4P9.34 chromosome II BAC At4g20170 2.03 -1.03 0.032001 0.79323 F4P9, Arabidopsis thaliana 253292_at Expressed protein; supported by full-length cDNA: At4g33985 2.03 -1.02 0.00638 0.87138 Ceres: 9341. 265772_at putative protein kinase contains a protein kinase At2g48010 2.02 1.04 0.012695 0.802318 domain profile (PDOC00100); supported by cDNA: gi_14335115_gb_AY037237.1-- 265375_at unknown protein; supported by full-length cDNA: At2g06530 2.02 1.08 0.015552 0.48528 Ceres: 91878. 265208_at putative giberellin beta-hydroxylase contains At2g36690 2.01 -1.35 0.006868 0.034666 similarities to GA beta-20-hydroxylase from tobacco (GB: 3327245) and to ethylene forming enzyme from Picea glauca (GB: L42466) 264746_at unknown protein similar to putative DNA-binding At1g62300 2.01 1.04 0.035415 0.469191 protein GI: 7268215 from [Arabidopsis thaliana]; supported by cDNA: gi_12658409_gb_AF331712.1_AF331712 260312_at putative disease resistance protein similar to disease At1g63880 2.01 -1.17 0.018521 0.24588 resistance protein RPP1-WsC GB: AAC72979 [Arabidopsis thaliana] 258173_at putative protein kinase similar to serine/threonine At3g21630 2.01 1.07 0.010988 0.529339 protein kinase Pto GB: AAB47421 [Lycopersicon esculentum] (Plant Cell 9 (1), 61-73 (1997)) 247617_at receptor like protein kinase receptor like protein At5g60270 2 -1.35 0.002662 0.065021 kinase, Arabidopsis thaliana, PIR: T47484 259213_at putative receptor ser/thr protein kinase similar to At3g09010 1.99 1.14 0.025161 0.261501 receptor kinase GB: S70769 [Arabidopsis thaliana]; supported by full-length cDNA: Ceres: 124301. 258544_at disease resistance gene (RPM1) identical to disease At3g07040 1.99 -1.35 0.036667 0.073823 resistance gene (RPM1) GB: X87851 [Arabidopsis thaliana] 249777_at putative protein similar to unknown protein (gb At5g24210 1.99 -1.11 0.022053 0.175713 AAD29063.1) 249208_at allene oxide synthase (emb CAA73184.1); At5g42650 1.99 1.18 0.018657 0.090962 supported by cDNA: gi_6002956_gb_AF172727.1_AF172727 248014_at putative protein similar to unknown protein At5g56340 1.99 -1.05 0.045301 0.723728 (pir||T05064) 264223_s_at receptor kinase, putative similar to receptor kinase 1 At1g67520 1.98 1.1 0.021718 0.628018 GI: 9294449 from [Arabidopsis thaliana] 262082_s_at wall-associated kinase 2, putative similar to At1g56120 1.98 1.12 0.030141 0.183617 receptor-like serine/threonine kinase GB: AAC50043 GI: 2465923 from [Arabidopsis thaliana] 249835_s_at putative protein similar to unknown protein (gb At5g23510 1.98 -1.08 0.01545 0.400133 AAF01580.1) 245051_at putative WRKY-type DNA-binding protein; At2g23320 1.98 1.19 0.009782 0.0756 supported by cDNA: gi_13506742_gb_AF224704.1_AF224704 264351_at unknown protein Contains similarity to At1g03370 1.97 1.02 0.026436 0.91627 gb|AB011110 KIAA0538 protein from Homo sapiens brain and to phospholipid-binding domain C2 PF|00168. ESTs gb|AA585988 and gb|T04384 come from this gene 262926_s_at receptor kinase, putative similar to receptor kinase 1 At1g65790 1.97 -1.11 0.021308 0.451766 [Brassica rapa] GB: BAA23676 253326_at putative protein polygalacturonase(EC 3.2.1.15) At4g33440 1.97 -1.05 0.019179 0.7895 precursor - Erwinia carotovora, PID: g42330 246305_at putative protein protein At2g40060 - Arabidopsis At3g51890 1.97 1.2 0.016172 0.125232 thaliana, EMBL: AF002109; supported by full-length cDNA: Ceres: 93427. 245219_at viral resistance protein, putative similar to viral At1g59124 1.97 -1.03 0.010527 0.805754 resistance protein GI: 7110565 from [Arabidopsis thaliana] 267393_at similar to axi 1 protein from Nicotiana tabacum At2g44500 1.96 -1.24 0.025881 0.212221 259109_at putative serine threonine protein phosphatase type At3g05580 1.96 1.08 0.029112 0.485226 one similar to GB: AAC39461 252037_at putative calmodulin calmodulin - Tetrahymena At3g51920 1.96 1.1 0.015438 0.170162 pyriformis (SGC5), PIR1: MCTE; supported by cDNA: gi_14190470_gb_AF380635.1_AF380635 258176_at B regulatory subunit of PP2A, putative similar to B At3g21650 1.95 -1.17 0.03792 0.249846 regulatory subunit of PP2A GB: AAB58902 [Arabidopsis thaliana] 256169_at receptor protein kinase, putative contains Pfam At1g51800 1.95 1.2 0.02536 0.076434 profiles: PF00069: Eukaryotic protein kinase domain, multiple PF00560: Leucine Rich Repeat 260974_at receptor-like serine/threonine kinase, putative At1g53440 1.94 -1.02 0.009705 0.677274 similar to receptor-like serine/threonine kinase GB: AAC50043 GI: 2465923 from [Arabidopsis thaliana] 252310_at GTPase activating-like protein GTPase activating At3g49350 1.94 -1 0.026703 0.921316 protein gyp7, Yarrowia lipolytica, EMBL: YLGYP7 251790_at elicitor responsive/phloem-like protein FIERG2 At3g55470 1.94 1.05 0.012047 0.64204 protein, Oryza sativa, PIR: T04363 249480_s_at protein kinase - like protein receptor-like protein At5g38990 1.94 -1.2 0.019593 0.193368 kinase (EC 2.7.1.--) precursor, Madagascar periwinkle, PIR: T10060 249364_at putative protein predicted protein, Arabidopsis At5g40590 1.94 -1.06 0.023463 0.5486 thaliana 265385_at putative diacylglycerol kinase; supported by full- At2g20900 1.93 1.03 0.02312 0.814052 length cDNA: Ceres: 15863. 264580_at unknown protein EST gb|ATTS0295 comes from At1g05340 1.93 1.37 0.011774 0.077989 this gene; supported by full-length cDNA: Ceres: 20380. 258608_at unknown protein; supported by full-length cDNA: At3g03020 1.93 1.12 0.001615 0.228742 Ceres: 35949. 262868_at unknown protein At1g64980 1.92 1.03 0.010064 0.758096 260255_at putative protein kinase similar to p58 protein kinase At1g74330 1.92 -1.02 0.036098 0.867257 GB: AAB59449 (Homo sapiens); contains Pfam profile: PF00069 Eukaryotic protein kinase domain 257902_at receptor kinase, putative similar to receptor kinase At3g28450 1.92 1.01 0.015158 0.903391 GB: AAD02501 from [Arabidopsis thaliana] 254211_at phosphatase like protein phosphoprotein phosphatase At4g23570 1.92 1.08 0.021478 0.429462 (EC 3.1.3.16) PPT - rat 252009_at zinc finger - like protein zinc finger protein 216, At3g52800 1.92 1.01 0.025834 0.996803 Homo sapiens, EMBL: AF062072; supported by cDNA: gi_14596166_gb_AY042871.1-- 265460_at putative caltractin; supported by full-length cDNA: At2g46600 1.91 -1.28 0.011693 0.060574 Ceres: 7802. 262455_at Mlo protein, putative similar to Mlo protein At1g11310 1.91 -1 0.011858 0.991742 GB: Z83834 GI: 1877220 from [Hordeum vulgare]; supported by full-length cDNA: Ceres: 259664. 262119_s_at glutathione S-transferase, putative similar to At1g02930 1.91 1.13 0.011514 0.130465 glutathione S-transferase GI: 860955 from [Hyoscyamus muticus]; supported by cDNA: gi_15215607_gb_AY050332.1-- 257700_at unknown protein similar to unknown protein At3g12740 1.91 1.12 0.011076 0.18275 GB: AAD25612 from [Arabidopsis thaliana]; supported by full-length cDNA: Ceres: 37019. 253534_at cytochrome P450 monooxygenase; supported by At4g31500 1.91 1.1 0.008445 0.216603 full-length cDNA: Ceres: 13745. 248873_at disease resistance protein-like At5g46450 1.91 1.04 0.019311 0.616452 251071_at putative protein receptor protein kinases At5g01950 1.89 -1.02 0.019672 0.77446 250419_at RPP1 disease resistance protein - like disease At5g11250 1.89 -1.16 0.007043 0.23803 resistance protein RPP1-WsC, Arabidopsis thaliana, EMBL: AF098964 246018_at Expressed protein; supported by full-length cDNA: At5g10695 1.88 1.11 0.007166 0.390262 Ceres: 103171. 245151_at putative pectinesterase; supported by full-length At2g47550 1.88 1.02 0.027515 0.828056 cDNA: Ceres: 111254. 265499_at putative glucosyltransferase At2g15480 1.87 -1.11 0.043677 0.508495 263797_at putative WRKY-type DNA binding protein; At2g24570 1.87 1.08 0.018452 0.267205 supported by cDNA: gi_15991743_gb_AF425836.1_AF425836 263731_at metalloproteinase, putative similar to At1g59970 1.87 -1.03 0.019632 0.713464 metalloproteinase GI: 3128477 from [Arabidopsis thaliana] 252076_at LS1-like protein AT-LS1 product - Arabidopsis At3g51660 1.87 1.38 0.021217 0.088589 thaliana, EMBL: X58827; supported by full-length cDNA: Ceres: 107294. 258460_at unknown protein At3g17330 1.86 1.07 0.005904 0.601401 245254_at ATP-sulfurylase; supported by cDNA: At4g14680 1.86 -1.38 0.031698 0.050397 gi_459143_gb_U06275.1_ATU06275 266536_at hypothetical protein predicted by genefinder; At2g16900 1.85 -1.07 0.006846 0.487008 supported by cDNA: gi_14532491_gb_AY039870.1-- 265479_at hypothetical protein; supported by full-length At2g15760 1.85 -1.04 0.048838 0.747296 cDNA: Ceres: 5. 262873_at hypothetical protein predicted by genemark.hmm At1g64700 1.85 1.2 0.006355 0.233681 258207_at putative GTP pyrophosphokinase similar to GTP At3g14050 1.85 -1.03 0.021111 0.736124 PYROPHOSPHOKINASE GB: O87331 from [Corynebacterium glutamicum]; supported by cDNA: gi_7141305_gb_AF225703.1_AF225703 267335_s_at putative beta-1,3-glucanase At2g19440 1.84 -1.28 0.025894 0.194876 245218_s_at viral resistance protein, putative, 5 partial similar to At1g58842 1.84 -1.02 0.034348 0.880456
viral resistance protein GI: 7110565 from [Arabidopsis thaliana] 264082_at unknown protein; supported by full-length cDNA: At2g28570 1.83 1.16 0.032702 0.469306 Ceres: 36244. 260037_at putative DNA-binding protein (RAV2-like) identical At1g68840 1.83 -1.21 0.029408 0.263089 to residues 34-352 of RAV2 GB: BAA34251 (Arabidopsis thaliana); supported by full-length cDNA: Ceres: 19561. 258134_at rubisco expression protein, putative similar to At3g24530 1.83 1.06 0.014418 0.42968 GB: O22034 from [Cyanidium caldarium] (J. Plant Res. 110, 235-245 (1997)); supported by full-length cDNA: Ceres: 148454. 260314_at unknown protein similar to putative protein At1g63830 1.82 -1.01 0.00932 0.923314 GB: CAA20468 [Arabidopsis thaliana] 258956_at hypothetical protein predicted by At3g01440 1.82 -1.19 0.013406 0.260869 genscan+; supported by full-length cDNA: Ceres: 13653. 262649_at unknown protein contains similarity to xenotropic At1g14040 1.81 1.05 0.009938 0.587887 and polytropic retrovirus receptor GB: 4759334 257972_at putative protein kinase, ATN1 almost identical (1 At3g27560 1.81 -1.04 0.029115 0.724347 amino acid) to GB: S61766 from [Arabidopsis thaliana]; supported by cDNA: gi_16604327_gb_AY058062.1-- 250575_at putative protein At5g08240 1.81 1.08 0.022224 0.516011 259826_at arm repeat-containing protein, putative similar to At1g29340 1.8 1.01 0.02957 0.88125 GI: 2558938 from [Brassica napus] (Proc. Natl. Acad. Sci. U.S.A. 95 (1), 382-387 (1998)) 253364_at F-box protein family, AtFBX13 cotains similarity to At4g33160 1.8 1.01 0.015511 0.924255 fimbriata GI: 547307 from [Antirrhinum majus] 248895_at receptor protein kinase At5g46330 1.8 -1.03 0.014527 0.815519 263457_at unknown protein At2g22300 1.79 1.05 0.018298 0.6555 254553_at TMV resistance protein N-like TMV resistance At4g19530 1.79 -1.02 0.024648 0.800063 protein N, Nicotiana glutinosa, PIR2: A54810 254331_s_at cytochrome P450-like protein flavonoid 3,5- At4g22710 1.79 1.04 0.011357 0.507737 hydroxylase Hf1, Petunia x hybrida, PIR2: S38985 245838_at disease resistance protein, putative similar to disease At1g58410 1.79 1.04 0.045241 0.800057 resistance protein RPP8 GI: 8843900 from [Arabidopsis thaliana] 267392_at putative beta-glucosidase At2g44490 1.78 -1.01 0.01293 0.768374 264879_at cotton fiber expressed protein, putative similar to At1g61260 1.78 -1.08 0.020899 0.6028 cotton fiber expressed protein 1 GI: 3264828 from [Gossypium hirsutum] 251804_at beta-1,3-glucanase - like protein probable beta-1,3- At3g55430 1.78 -1.02 0.033278 0.91732 glucanase, Triticum aestivum, PIR: T06268; supported by full-length cDNA: Ceres: 8980. 249314_at receptor kinase-like protein At5g41180 1.78 1.15 0.034221 0.348074 245456_at disease resistance RPP5 like protein At4g16950 1.78 -1.02 0.014799 0.787739 267169_at putative oxidoreductase At2g37540 1.77 1.01 0.020016 0.912423 265079_at hypothetical protein contains similarity to zinc finger At1g55460 1.76 -1.02 0.015429 0.82749 protein rts2 GB: U16133 GI: 563244 from [Saccharomyces cerevisiae]; supported by cDNA: gi_13430439_gb_AF360132.1_AF360132 259230_at unknown protein; supported by cDNA: At3g07780 1.76 1.13 0.012523 0.041915 gi_15028084_gb_AY045899.1-- 250850_at putative protein; supported by cDNA: At5g04550 1.76 -1.14 0.015093 0.108003 gi_13605828_gb_AF367313.1_AF367313 261506_at choline kinase, putative similar to At1g71697 1.75 1.03 0.033638 0.818348 CHOLINE/ETHANOLAMINE KINASE GB: Q9Y259 from [Homo sapiens] 251028_at putative protein putative hydrolase At2g32150- At5g02230 1.75 -1.06 0.019924 0.548891 Arabidopsis thaliana, EMBL: AC006223; supported by full-length cDNA: Ceres: 36724. 258336_at putative ethylene-inducible protein similar to At3g16050 1.74 1.13 0.019656 0.136759 ethylene-inducible protein GB: M88254 from [Hevea brasiliensis]; supported by cDNA: gi_4103951_gb_AF029980.1_AF029980 253415_at putative protein peptidyl-prolyl cis-trans isomerase, At4g33060 1.74 1.15 0.022933 0.090678 Schizosaccharomyces pombe, gb: SPBC16H5 251643_at guanylate kinase-like protein guanylate kinase - At3g57550 1.74 1.12 0.014659 0.125227 Mus musculus, TREMBL: MMU53514_1; supported by cDNA: gi_7861797_gb_AF204676.1_AF204676 247384_at protein kinase; supported by cDNA: At5g63370 1.74 1.11 0.010852 0.400409 gi_16974578_gb_AY060555.1-- 265269_at hypothetical protein predicted by genscan At2g42950 1.72 1.09 0.017878 0.365888 262571_at hypothetical protein predicted by genscan+; At1g15430 1.72 1.12 0.022912 0.305054 supported by cDNA: gi_15293248_gb_AY051058.1-- 259466_at response regulator 5, putative similar to response At1g19050 1.72 -1.03 0.026729 0.64387 regulator 5 GI: 3953599 from [Arabidopsis thaliana]; supported by cDNA: gi_3953602_dbj_AB008490.1_AB008490 254723_at ammonium transport protein (AMT1); supported by At4g13510 1.72 1.08 0.011754 0.406577 cDNA: gi_14335079_gb_AY037219.1-- 253193_at putative protein SEC7 protein, Saccharomyces At4g35380 1.72 1.12 0.019937 0.504827 cerevisiae, PIR2: S49764 265461_at unknown protein similarity to ubiquitin family of At2g46500 1.71 1.18 0.019691 0.111325 proteins; supported by cDNA: gi_16930424_gb_AF419566.1_AF419566 253614_at putative protein heat shock protein 101 - Triticum At4g30350 1.71 -1.07 0.030479 0.677967 aestivum, PID: g4558484 247816_at similar to unknown protein (pir||S75584); supported At5g58260 1.71 -1.12 0.011236 0.191707 by full-length cDNA: Ceres: 3488. 262457_at hypothetical protein similar to hypothetical protein At1g11200 1.7 -1.03 0.017609 0.719233 GB: CAB36801 GI: 4455265 from [Arabidopsis thaliana]; supported by full-length cDNA: Ceres: 40975. 255512_at Expressed protein; supported by cDNA: At4g02195 1.69 1.06 0.014636 0.692316 gi_5059351_gb_AF154574.1_AF154574 251516_s_at putative protein hypothetical protein SPCC320.08 - At3g59310 1.69 -1.06 0.017534 0.387516 Schizosaccharomyces pombe, PIR: T41303 254103_at putative protein; supported by full-length cDNA: At4g25030 1.68 1.04 0.007754 0.433098 Ceres: 16463. 245757_at phosphate-induced (phi-1) protein, putative similar At1g35140 1.68 -1.51 0.004633 0.081204 to phi-1 GB: BAA33810 GI: 3759184 from [Nicotiana tabacum]; supported by full-length cDNA: Ceres: 118937. 253387_at P-Protein-like protein P-Protein precursor, Solanum At4g33010 1.66 1.04 0.019086 0.463263 tuberosum, gb: Z99770; supported by cDNA: gi_14596024_gb_AY042800.1-- 247272_at GTP cyclohydrolase II; 3,4-dihydroxy-2-butanone-4- At5g64300 1.66 1 0.011375 0.888475 phoshate synthase (emb|CAA03884.1) supported by cDNA: gi_940382_dbj_D45165.1_ATHGTPCII 261788_at unknown protein; supported by full-length cDNA: At1g15980 1.65 -1.05 0.013373 0.66035 Ceres: 122986. 249010_at unknown protein; supported by cDNA: At5g44580 1.65 1.05 0.008905 0.282782 gi_15027902_gb_AY045808.1-- 259074_at putative protein kinase contains Pfam profile: At3g02130 1.63 -1.03 0.007959 0.571341 Eukaryotic protein kinase domain 258394_at unknown protein; supported by full-length cDNA: At3g15530 1.63 1.07 0.004284 0.548019 Ceres: 15303. 258665_at thioredoxin-like protein similar to thioredoxin H- At3g08710 1.61 1.03 0.0083 0.731007 type GB: P29448 [Arabidopsis thaliana] 253317_at putative protein At4g33960 -1.83 -1.77 0.010341 0.022397 260126_at putative hydroxymethyltransferase similar to serine At1g36370 -1.93 -1.86 0.005701 0.006964 hydroxymethyltransferage GB: P50433 from [Solanum tuberosum]; supported by full-length cDNA: Ceres: 122515. 246926_at putative protein At5g25240 -2.09 -2.21 0.019603 0.017979 258217_at unknown protein contains Pfam profile PF00398 At3g17990 -2.21 -2.27 0.009887 0.0037 Ribosomal RNA adenine dimethylases 258218_at methyltransferase, putative similar to At3g18000 -2.21 -2.29 0.00667 0.009294 methyltransferase GB: AAC01738 from [Amycolatopsis mediterranei] 254343_at PRH26 protein; supported by full-length cDNA: At4g21990 -2.22 -1.83 0.012838 0.031291 Ceres: 36866. 265121_at similar to flavin-containing monooxygenase At1g62560 -2.37 -1.87 0.020126 0.00922 (sp|P36366); similar to ESTs gb|R30018, gb|H36886, gb|N37822, and gb|T88100 similar to flavin- containing monooxygenase GB: AAA21178 GI: 349534 from [Oryctolagus cuniculus]; supported by cDNA: gi_13877746_gb_AF37013 251039_at putative protein hypothetical protein T6H20.90 - At5g02020 -3.73 -1.91 0.001899 0.021967 Arabidopsis thaliana, EMBL: AL096859; supported by cDNA: gi_16648747_gb_AY058150.1-- 259015_at unknown protein similar to hypothetical protein At3g07350 -3.79 -1.81 0.001762 0.010373 GB: AAC17612 [Arabidopsis thaliana]; supported by full-length cDNA: Ceres: 251012. 248676_at putative protein similar to unknown protein At5g48850 -5.55 -4.23 0.003428 0.003335 (gb|AAC72543.1) 249752_at putative protein similar to unknown protein (emb At5g24660 -5.87 -2.27 0.002654 0.005949 CAB62461.1); supported by full-length cDNA: Ceres: 268701. 246935_at leucine-rich repeats containing protein grr1 - At5g25350 -1.64 -1.09 0.01077 0.205242 Glycine max. EMBL: AF019910 261957_at methionine/cystathionine gamma lyase, putative At1g64660 -1.66 1.1 0.017509 0.258584 similar to methionine gamma-lyase GB: CAA04124.1 GI: 2330885 from [Trichomonas vaginalis]; supported by cDNA: gi_15450931_gb_AY054546.1-- 263284_at unknown protein At2g36100 -1.68 1.21 0.009385 0.046069 263064_at putative bZIP transcription factor contains a bZIP At2g18160 -1.68 -1.04 0.003705 0.553739 transcription factor basic domain signature (PDOC00036); supported by cDNA: gi_14335073_gb_AY037216.1-- 265102_at putative peroxidase similar to cationic peroxidase At1g30870 -1.69 1.01 0.008534 0.760053 (gi|1232069); similar to EST gb|AI100412; supported by full-length cDNA: Ceres: 123968. 259773_at auxin-induced protein, putative similar to At1g29500 -1.69 1.03 0.017479 0.683467 SP: P33083 from [Glycine max] 258181_at nitrate transporter identical to nitrate transporter At3g21670 -1.7 -1.68 0.013713 0.025046 GB: CAB38706 [Arabidopsis thaliana]; supported by full-length cDNA: Ceres: 111089. 252220_at putative protein hypothetical protein - Arabidopsis At3g49940 -1.7 -1.08 0.010242 0.277047 thaliana, EMBL: CAB38293; supported by full-length cDNA: Ceres: 17840. 251524_at 3-isopropylmalate dehydratase-like protein (small At3g58990 -1.71 -1.3 0.015617 0.116619 subunit) 3-isopropylmalate dehydratase, small subunit - Thermotoga maritima, PIR: A72363 258008_at putative late embryogenesis abundant protein similar At3g19430 -1.72 1.26 0.007578 0.089324 to GB: AAB01570 from [Picea glauca] 263227_at Expressed protein; supported by cDNA: At1g30750 -1.73 1.1 0.00931 0.284316 gi_15292976_gb_AY050922.1-- 263118_at putative 3-methylcrotonyl-CoA carboxylase ESTs At1g03090 -1.73 -1.22 0.009453 0.064506 gb|H35836, gb|AA651295 and gb|AA721862 come from this gene; supported by cDNA: gi_533706_gb_U12536.1_ATU12536
248252_at putative protein similar to unknown protein At5g53250 -1.73 1.16 0.005785 0.142465 (emb|CAB71094.1) 256598_at cytochrome P450 homolog, putative similar to At3g30180 -1.74 1.01 0.019165 0.939678 cytochrome P450 homolog GB: U54770 GI: 1421740 from [Lycopersicon esculentum]; supported by full- length cDNA: Ceres: 11278. 256062_at unknown protein; supported by full-length cDNA: At1g07090 -1.75 1.03 0.007971 0.599694 Ceres: 28780. 263490_at F-box protein ORE9, AtFBL7 identical to F-box At2g42620 -1.76 1.14 0.014384 0.078788 containing protein ORE9 GI: 15420162 from [Arabidopsis thaliana] 247477_at putative protein 21K protein precursor, Medicago At5g62340 -1.76 1.03 0.019734 0.869872 sativa, PIR: T09390 262399_at unknown protein; supported by full-length cDNA: At1g49500 -1.77 -1.05 0.018923 0.609256 Ceres: 33047. 259856_at unknown protein; supported by full-length cDNA: At1g68440 -1.77 -1.41 0.017035 0.048737 Ceres: 34166. 253510_at hypothetical protein At4g31730 -1.77 1.29 0.034578 0.053073 251017_at protein phosphatase - like protein protein At5g02760 -1.77 -1.04 0.007317 0.502031 phosphatase 2C homolog, Mesembryanthemum crystallinum, EMBL: AF097667 248279_at putative protein similar to unknown protein At5g52910 -1.77 -1.28 0.016383 0.116964 (pir||T13959) 266191_at putative peroxidase At2g39040 -1.78 1.26 0.012061 0.114533 253217_at actin depolymerizing factor - like protein actin At4g34970 -1.78 1.31 0.011164 0.086098 depolymerizing factor1, Arabidopsis thaliana, PID: G1408471 262717_s_at putative cytochrome P450 similar to gb|AF069494 At1g16410 -1.79 -1.32 0.016905 0.014389 cytochrome P450 from Sinapis alba and is a member of the PF|00067 Cytochrome P450 family. EST gb|F14190 comes from this gene; supported by cDNA: gi_15208670_gb_AY035021.2-- 262517_at putative glutathione transferase Second of three At1g17180 -1.79 1.02 0.02066 0.906386 repeated putative glutathione transferases. 72% identical to glutathione transferase [Arabidopsis thaliana] (gi|4006934). Location of ests 191A10T7 (gb|R90188) and 171N13T7 (gb|R65532) 256926_at hypothetical protein predicted by genemark.hmm At3g22540 -1.79 1.21 0.036851 0.289954 256252_at glucosyl transferase, putative similar to zeatin O- At3g11340 -1.79 1.59 0.019921 0.01385 xylosyltransferase SP: P56725 [Phaseolus vulgaris (Kidney bean) (French bean)] 261226_at expansin S2 precursor, putative similar to At1g20190 -1.8 -1.05 0.012655 0.608442 GB: U30460 from [Cucumis sativus]; supported by full-length cDNA: Ceres: 11011. 251144_at anthranilate N-benzoyltransferase-like protein At5g01210 -1.8 1.11 0.008524 0.106988 anthranilate N-benzoyltransferase, clove pink, PIR: T10717; supported by cDNA: gi_15912268_gb_AY056412.1-- 265645_at unknown protein At2g27370 -1.81 1.1 0.025976 0.504975 249923_at conglutin gamma-like protein conglutin gamma At5g19120 -1.81 -1.05 0.007868 0.519593 precursor, Lupinus angustifolius, PIR: S21426; supported by cDNA: gi_15010797_gb_AY045700.1-- 247914_at xyloglucan endotransglycosylase At5g57540 -1.81 -1.03 0.026871 0.814009 265048_at jasmonate inducible protein, putative similar to At1g52050 -1.82 1.15 0.022645 0.325821 jasmonate inducible protein GI: 9279642 from [Arabidopsis thaliana] 252970_at small auxin up RNA (SAUR-AC1); supported by At4g38850 -1.82 1.19 0.007042 0.093178 full-length cDNA: Ceres: 14973. 249862_at PGPD14 protein; supported by full-length cDNA: At5g22920 -1.82 -1.2 0.013335 0.024948 Ceres: 41666. 266820_at putative AP2 domain transcription factor pFAM At2g44940 -1.84 -1.27 0.027529 0.279728 domain (PF00847) supported by full-length cDNA: Ceres: 31044. 258038_at unknown protein; supported by full-length cDNA: At3g21260 -1.84 -1.25 0.024483 0.131574 Ceres: 260109. 252250_at putative protein predicted protein, Arabidopsis At3g49790 -1.85 -1.2 0.011715 0.131586 thaliana 247337_at putative protein similar to unknown protein At5g63660 -1.85 1 0.021642 0.941607 (pir||S51637) 260167_at hypothetical protein predicted by genscan+ At1g71970 -1.86 -1.06 0.02425 0.744455 257162_s_at ammonium transporter, putative similar to At3g24290 -1.86 -1.03 0.01777 0.655292 GB: AAD54638 from [Arabidopsis thaliana] (Plant Cell (1999) 11 (5), 937-948) 246275_at putative protein; supported by full-length cDNA: At4g36540 -1.86 1.06 0.011645 0.640095 Ceres: 123997. 245586_at hypothetical protein At4g14980 -1.86 1.16 0.037731 0.349881 245136_at putative auxin-regulated protein At2g45210 -1.86 1.1 0.020869 0.28948 262850_at signal response protein (GAI) identical to GAI At1g14920 -1.87 -1.05 0.012647 0.589803 GB: CAA75492 GI: 2569938 [Arabidopsis thaliana] (Genes Dev. In press); supported by cDNA: gi_16648833_gb_AY058194.1-- 258080_at unknown protein; supported by full-length cDNA: At3g25930 -1.87 1.11 0.025451 0.601329 Ceres: 2767. 253255_at putative auxin-regulated protein auxin-induced At4g34760 -1.87 -1.2 0.007683 0.154545 protein X15, Glycine max, PIR2: JQ1097; supported by full-length cDNA: Ceres: 10510. 246996_at putative protein similar to unknown protein At5g67420 -1.87 -1.17 0.029109 0.184764 (emb|CAB62102.1); supported by full-length cDNA: Ceres: 40250. 265511_at putative glycine-rich protein; supported by cDNA: At2g05540 -1.88 -1.36 0.004653 0.037214 gi_15215617_gb_AY050337.1-- 264957_at F-box protein family, AtFBL5 contains similarity to At1g77000 -1.88 -1.06 0.021505 0.577258 F-box protein FBL2 GI: 6063090 from [Homo sapiens]; supported by full-length cDNA: Ceres: 3549. 264467_at unknown protein similar to EST At1g10140 -1.88 1.26 0.012415 0.023258 gb|AA598098; supported by full-length cDNA: Ceres: 23916. 256828_at unknown protein At3g22970 -1.88 1.18 0.017776 0.232803 248178_at root cap protein 2-like protein At5g54370 -1.88 1.29 0.00898 0.087446 262396_at unknown protein; supported by full-length cDNA: At1g49470 -1.89 -1.17 0.02115 0.065156 Ceres: 95546. 259976_at hypothetical protein; supported by full-length At1g76560 -1.89 -1.22 0.011496 0.142412 cDNA: Ceres: 147838. 252834_at putative protein RING-H2 zinc finger protein ATL6- At4g40070 -1.89 1.24 0.030751 0.235116 Arabidopsis thaliana, PID: g4928403; supported by cDNA: gi_16930492_gb_AF419600.1_AF419600 250860_at amino acid transport-like protein amino acid At5g04770 -1.89 -1.19 0.02117 0.317647 transport protein AAT1, Arabidopsis thaliana, PIR: S51171; supported by full-length cDNA: Ceres: 158156. 265049_at jasmonate inducible protein, putative similar to At1g52060 -1.9 1.31 0.010222 0.06857 jasmonate inducible protein GI: 9279642 from [Arabidopsis thaliana] 265050_at jasmonate inducible protein, putative similar to At1g52070 -1.91 1.32 0.019022 0.277021 jasmonate inducible protein GI: 9279641 from [Arabidopsis thaliana] 252991_at protein kinase like protein protein kinase 6 (EC At4g38470 -1.91 -1.43 0.022882 0.053733 2.7.1.--)-soybean, PIR2: S29851 250157_at prx10 peroxidase-like protein prx10 peroxidase, At5g15180 -1.91 1.05 0.009746 0.667075 Spinacia oleracea, EMBL: SOY16776 267457_at putative proline-rich protein At2g33790 -1.92 1.55 0.02688 0.053166 266882_at unknown protein; supported by full-length cDNA: At2g44670 -1.92 -1.35 0.00935 0.103065 Ceres: 40641. 263208_at zinc finger protein 5, ZFP5 possible transcription At1g10480 -1.93 1.06 0.009367 0.572822 factor with C2H2 zinc finger; supported by full- length cDNA: Ceres: 23664. 253722_at putative protein zinc finger transcription factor- At4g29190 -1.93 1.08 0.004592 0.184244 Arabidopsis thaliana, PID: g2961542; supported by full-length cDNA: Ceres: 16432. 251356_at putative protein hypothetical proteins-Arabidopsis At3g61060 -1.93 -1.13 0.007848 0.181064 thaliana; supported by cDNA: gi_14334587_gb_AY034967.1-- 245176_at unknown protein similar to GP|2104534|AF001308 At2g47440 -1.93 -1.64 0.031285 0.016259 (T10M13.11) 262170_at hypothetical protein predicted by At1g74940 -1.94 1.09 0.007492 0.34681 genemark.hmm; supported by full-length cDNA: Ceres: 24864. 260900_s_at branched-chain alpha keto-acid dehydrogenase, At1g21400 -1.94 -1.33 0.003536 0.091842 putative similar to branched-chain alpha keto-acid dehydrogenase GB: AAC69851 GI: 3822223 from [Arabidopsis thaliana] 260058_at unknown protein; supported by cDNA: At1g78100 -1.94 1.22 0.026338 0.07061 gi_15450975_gb_AY054568.1-- 259854_at RING-H2 zinc finger protein ATL3, putative similar At1g72200 -1.94 1.07 0.006318 0.364739 to GI: 4928397 from [Arabidopsis thaliana] (Plant Mol. Biol. 40 (4), 579-590 (1999)) 258145_at integral membrane protein, putative similar to At3g18200 -1.94 1.08 0.032913 0.537375 MtN21 (nodulation-induced gene) GB: CAA75575 [Medicago truncatula] 253763_at xyloglucan endotransglycosylase-like protein At4g28850 -1.94 -1.07 0.010403 0.604484 xyloglucan endotransglycosylase 1, Fagus sylvatica, PID: e1354157 249008_at putative protein contains similarity to DNA-3- At5g44680 -1.94 1.04 0.019733 0.431852 methyladenine glycosylase I; supported by full-length cDNA: Ceres: 29551. 261711_at unknown protein similar to hypothetical protein At1g32700 -1.95 -1.07 0.022252 0.357576 GB: AAF25968 GI: 6714272 from [Arabidopsis thaliana]; supported by full-length cDNA: Ceres: 206224. 260887_at ascorbate oxidase promoter-binding protein, putative At1g29160 -1.95 -1.05 0.008773 0.547417 similar to ascorbate oxidase promoter-binding protein GB: D45066 GI: 853689 from [Cucurbita maxima] 254718_at putative protein disease resistance response protein At4g13580 -1.95 1.13 0.008166 0.148335 206-d, Pisum sativum, U11716 253103_at putative auxin-induced protein high similarity to At4g36110 -1.95 1.24 0.007143 0.125412 auxin-induced protein 15A, soybean, PIR2: JQ1096; supported by cDNA: gi_13194817_gb_AF349524.1_AF349524 245987_at NAM-like protein hypothetical protein SENU5, At5g13180 -1.95 -1 0.030979 0.994604 senescence up-regulated-Lycopersicon esculentum, EMBL: Z75524; supported by cDNA: gi_14326559_gb_AF385734.1_AF385734 264130_at hypothetical protein predicted by genemark.hmm At1g79160 -1.96 -1.01 0.007603 0.925019 257076_at unknown protein At3g19680 -1.96 -1.32 0.00642 0.015347 248564_at putative protein contains similarity to AT-hook At5g49700 -1.96 1.11 0.017313 0.355178 DNA-binding protein 246228_at peroxidase like protein At4g36430 -1.96 1.3 0.014156 0.048404 245090_at putative integral membrane protein nodulin At2g40900 -1.96 1.07 0.023157 0.443533 265031_at serine/threonine protein kinase, putative similar to At1g61590 -1.97 1.11 0.0313 0.357745 serine/threonine protein kinase GI: 1066501 from [Arabidopsis thaliana]
263981_at unknown protein; supported by full-length cDNA: At2g42870 -1.97 -1.13 0.013425 0.350209 Ceres: 102453. 252570_at isovaleryl-CoA-dehydrogenase precursor (IVD); At3g45300 -1.97 -1.07 0.015973 0.428779 supported by full-length cDNA: Ceres: 33674. 248432_at putative protein similar to unknown protein At5g51390 -1.97 -1.14 0.006297 0.177565 (gb|AAB68039.1); supported by full-length cDNA: Ceres: 1076. 267628_at unknown protein similar to GP|2262147|AC002330 At2g42280 -1.98 1.01 0.023113 0.929253 266941_at peroxidase (ATP22a) identical to GB: Y08781 At2g18980 -1.98 -1.16 0.014024 0.253565 266838_at similar to jasmonate-inducible proteins from At2g25980 -1.98 1.03 0.013791 0.844383 Brassica napus 263318_at Expressed protein; supported by full-length cDNA: At2g24762 -1.98 1.02 0.019425 0.816736 Ceres: 19631. 260081_at unknown protein At1g78170 -1.98 1.1 0.010912 0.443013 257654_at DnaJ protein, putative contains Pfam profile: At3g13310 -1.98 1.19 0.018497 0.11787 PF00226 DnaJ domain; supported by full-length cDNA: Ceres: 31309. 257294_at non-phototropic hypocotyl protein, putative similar At3g15570 -1.98 -1.4 0.017594 0.029437 to GB: AAF05914 from [Arabidopsis thaliana]; supported by full-length cDNA: Ceres: 118259. 254606_at nodulin-26-like protein major intrinsic protein, At4g19030 -1.98 1.03 0.008673 0.680712 Oryza sativa, PIR2: S52003; supported by full-length cDNA: Ceres: 109513. 264014_at putative auxin-regulated protein At2g21210 -1.99 -1.17 0.002926 0.036113 260770_at RING-H2 finger protein RHA3a, putative similar to At1g49200 -1.99 1.24 0.024855 0.128926 RING-H2 finger protein RHA3a GI: 3790573 from [Arabidopsis thaliana]; supported by cDNA: gi_14517431_gb_AY039551.1-- 260693_at peptide transporter PTR2-B, putative similar to At1g32450 -1.99 -1 0.029791 0.955651 SP: P46032 from [Arabidopsis thaliana] 257448_s_at putative protein various predicted proteins At3g45800 -1.99 -1.12 0.018199 0.206911 Arabidopsis thaliana 259328_at putative lectin contains Pfam profile: PF01419 At3g16440 -2 1.24 0.010314 0.191237 jacalin-like lectin domain; similar to jasmonate inducible protein GB: Y11483 (Brassica napus), myrosinase binding protein GB: BAA84545 (Arabidopsis thaliana); supported by cDNA: gi_6694742_gb_AF214573.1_AF2145 263151_at hypothetical protein predicted by At1g54120 -2.01 -1.32 0.026676 0.072957 genemark.hmm; supported by full-length cDNA: Ceres: 94743. 262427_s_at thioglucosidase, putative similar to thioglucosidase At1g47600 -2.01 1.2 0.008411 0.161898 GI: 871992 from [Arabidopsis thaliana] 261822_at unknown protein; supported by full-length cDNA: At1g11380 -2.01 -1.42 0.035581 0.129626 Ceres: 113571. 265245_at unknown protein At2g43060 -2.03 -1.03 0.010559 0.598558 258511_at unknown protein; supported by full-length cDNA: At3g06590 -2.03 -1.06 0.005721 0.306173 Ceres: 9391. 251072_at putative protein wound-inducible protein wun1 At5g01740 -2.03 -1.26 0.023991 0.03577 protein-Solanum tuberosum, PIR: JQ0398; supported by full-length cDNA: Ceres: 248967. 267178_at unknown protein; supported by full-length cDNA: At2g37750 -2.04 1.3 0.011938 0.023415 Ceres: 28529. 262236_at hypothetical protein similar to hypothetical protein At1g48330 -2.04 -1.17 0.028943 0.116441 GI: 9294146 from [Arabidopsis thaliana] 250717_at putative protein similar to unknown protein At5g06200 -2.04 1.1 0.014766 0.475991 (gb|AAF00668.1) 263265_at hypothetical protein predicted by genscan and At2g38820 -2.05 -1.04 0.028413 0.735506 genefinder; supported by cDNA: gi_16649128_gb_AY059934.1-- 263150_at heat-shock protein, putative similar to heat-shock At1g54050 -2.05 1.3 0.019374 0.275131 protein GI: 472939 from [Helianthus annuus]; supported by full-length cDNA: Ceres: 97415. 254820_s_at pEARLI 1-like protein Arabidopsis thaliana At4g12510 -2.05 1.07 0.00646 0.52646 pEARLI 1 mRNA, completecds, PID: g871780 251174_at putative protein latex protein allergen Hev b 7- At3g63200 -2.05 -1.09 0.011679 0.253402 Hevea brasiliensis, EMBL: AF113546; supported by cDNA: gi_15912226_gb_AY056391.1-- 250469_at pollen allergen-like protein SAH7 protein, At5g10130 -2.05 -1.05 0.021086 0.612309 Arabidopsis thaliana, EMBL: ATH133639 249606_at putative protein DNA-binding protein CCA1, At5g37260 -2.05 1.11 0.011138 0.453754 Arabidopsis thaliana, PIR: T02684 252368_at cytochrome P450-like protein cytochrome P450 At3g48520 -2.06 1.18 0.017948 0.40172 CYP94A1-Vicia sativa, PIR2: T08014 245277_at glucosyltransferase like protein; supported by At4g15550 -2.07 -1.21 0.004725 0.325423 cDNA: gi_2149126_gb_U81293.1_ATU81293 260266_at putative B-box zinc finger protein contains Pfam At1g68520 -2.08 -1.06 0.023551 0.259093 profile: PF00643 B-box zinc finger; supported by full-length cDNA: Ceres: 108109. 260741_at hypothetical protein contains Pfam profile: PF00117 At1g15045 -2.09 1.12 0.016477 0.263068 Glutamine amidotransferase class-I 257858_at hypothetical protein predicted by At3g12920 -2.1 1 0.019142 0.98788 genefinder; supported by full-length cDNA: Ceres: 924. 266372_at putative two-component response regulator 3 At2g41310 -2.11 -1.24 0.013281 0.195422 protein identical to GB: AB010917, contains a response regulator receiver domain; supported by cDNA: gi_3273199_dbj_AB010917.1_AB010917 266072_at putative trehalose-6-phosphate synthase At2g18700 -2.11 -1.08 0.005616 0.502618 255858_at zinc finger protein (ZFP6) identical to zinc finger At1g67030 -2.11 1.3 0.017196 0.066902 protein GI: 790683 from [Arabidopsis thaliana]; supported by cDNA: gi_15215716_gb_AY050387.1-- 247954_at beta-galactosidase (emb|CAB64740.1); supported At5g56870 -2.12 -1.36 0.01322 0.072012 by cDNA: gi_15451017_gb_AY054589.1-- 252036_at putative protein; supported by full-length cDNA: At3g52070 -2.13 -1.21 0.011681 0.137087 Ceres: 118329. 258497_at putative flowering-time gene CONSTANS (COL2) At3g02380 -2.14 -1.47 0.018979 0.03124 identical to putative flowering-time gene CONSTANS (COL2) GB: AAB67879 GI: 1507699 (Arabidopsis thaliana); supported by full-length cDNA: Ceres: 949. 253829_at Medicago nodulin N21-like protein MtN21 gene, At4g28040 -2.14 -1.2 0.006927 0.185535 Medicago truncatula, Y15293; supported by cDNA: gi_13899060_gb_AF370525.1_AF370525 248801_at homeobox-leucine zipper protein-like; supported by At5g47370 -2.14 1.06 0.008347 0.456353 cDNA: gi_15450446_gb_AY052324.1-- 247921_at CONSTANS-like B-box zinc finger protein-like; At5g57660 -2.14 -1.13 0.003117 0.120344 supported by full-length cDNA: Ceres: 6639. 257615_at unknown protein At3g26510 -2.16 -1.13 0.004838 0.273425 266140_at nodulin-like protein; supported by cDNA: At2g28120 -2.17 -1.3 0.008244 0.055633 gi_16209713_gb_AY057618.1-- 257643_at AP2 domain transcription factor contains Pfam At3g25730 -2.17 -1.32 0.038334 0.152701 profile: PF00847 AP2 domain; similar to RAV1 (DNA-binding protein) GB: BAA34250 [Arabidopsis thaliana] (Nucleic Acids Res. 27 (2), 470-478 (1999)); supported by full-length cDNA: Ceres: 39877. 248528_at putative protein similar to unknown protein At5g50760 -2.18 1.01 0.008566 0.918675 (emb|CAB86483.1) 264788_at putative DnaJ protein; supported by full-length At2g17880 -2.19 -1.25 0.011101 0.265694 cDNA: Ceres: 22711. 253957_at putative protein; supported by cDNA: At4g26320 -2.19 -1.16 0.006508 0.414219 gi_10880502_gb_AF195894.1_AF195894 247199_at DNA binding protein TGA1a homolog; supported At5g65210 -2.19 1.09 0.010073 0.256741 by full-length cDNA: Ceres: 31032. 247170_at putative protein contains similarity to lectin-like At5g65530 -2.19 1.2 0.020752 0.326964 protein kinase 250099_at unknown protein; supported by cDNA: At5g17300 -2.2 1.13 0.018091 0.37707 gi_14190364_gb_AF378860.1_AF378860 247474_at putative protein predicted proteins, Arabidopsis At5g62280 -2.21 -1.01 0.016382 0.887761 thaliana 261768_at gibberellin 3 beta-hydroxylase, putative similar to At1g15550 -2.22 1.01 0.037882 0.920313 gibberellin 3 beta-hydroxylase GI: 3982753 from [Arabidopsis thaliana]; supported by cDNA: gi_1945343_gb_L37126.1_ATHGA4A 259264_at putative aldose 1-epimerase shows similarity to At3g01260 -2.22 1.22 0.023126 0.227322 aldose epimerases 253812_at putative wound induced protein wound-induced At4g28240 -2.25 -1.09 0.00548 0.137021 protein-tomato (fragment), PIR2: S19773; supported by full-length cDNA: Ceres: 20161. 246917_at serine-rich protein; supported by full-length cDNA: At5g25280 -2.25 1.24 0.008947 0.039883 Ceres: 99323. 261265_at hypothetical protein predicted by At1g26800 -2.26 1.2 0.022793 0.218643 genscan+; supported by full-length cDNA: Ceres: 250127. 246229_at pectinesterase like protein At4g37160 -2.26 1.07 0.01383 0.725863 250012_x_at auxin-induced protein-like At5g18060 -2.27 1.11 0.022574 0.446969 259751_at putative transcription factor similar to myb-related At1g71030 -2.29 -1.31 0.004307 0.051811 transcription factor 24 GB: S71287; supported by full- length cDNA: Ceres: 31592. 246932_at ethylene-responsive element-like protein ethylene- At5g25190 -2.31 -1.28 0.015246 0.042175 responsive element binding protein homolog, Stylosanthes hamata, EMBL: U91857; supported by cDNA: gi_15010715_gb_AY045659.1-- 264463_at unknown protein similar to ESTs gb|T20511, At1g10150 -2.32 -1.04 0.007428 0.701582 gb|T45308, gb|H36493, and gb|AA651176; supported by full-length cDNA: Ceres: 2558. 252178_at putative protein various predicted proteins At3g50750 -2.33 1.17 0.016542 0.191688 247149_at unknown protein; supported by full-length cDNA: At5g65660 -2.33 -1.03 0.011232 0.82378 Ceres: 25419. 260855_at phosphatidylinositol-4-phosphate 5-kinase, putative At1g21920 -2.34 1.02 0.010684 0.812177 similar to phosphatidylinositol-4-phosphate 5-kinase GB: CAB53377 GI: 5777366 from [Arabidopsis thaliana]; supported by full-length cDNA: Ceres: 37462. 256743_at Expressed protein; supported by full-length cDNA: At3g29370 -2.34 1.09 0.007379 0.34665 Ceres: 22461. 249065_at putative protein similar to unknown protein (gb At5g44260 -2.34 -1.05 0.002162 0.688129 AAD10689.1); supported by cDNA: gi_14334449_gb_AY034916.1-- 264524_at tat-binding protein, putative Highly Similar to At1g10070 -2.35 -1.06 0.006931 0.464612 branched-chain amino acid aminotransferase; Location of EST gb|T44177 and gb|AA395381; supported by cDNA: gi_15293208_gb_AY051038.1-- 264521_at unknown protein Location of EST gb|T41885 and At1g10020 -2.37 -1.37 0.002442 0.03961 gb|AA395021 258091_at hypothetical protein predicted by genmark; supported At3g14560 -2.37 1.27 0.014798 0.126209
by full-length cDNA: Ceres: 19279. 261480_at phytochrome kinase substrate 1, putative similar to At1g14280 -2.39 1.06 0.004881 0.498893 phytochrome kinase substrate 1 GI: 5020168 from [Arabidopsis thaliana]; supported by full-length cDNA: Ceres: 97569. 252040_at putative protein hypothetical protein F10M6.70- At3g52060 -2.39 -1.02 0.011127 0.860596 Arabidopsis thaliana, PIR3: T05399; supported by cDNA: gi_15293266_gb_AY051067.1-- 246001_at putative protein predicted protein, Arabidopsis At5g20790 -2.39 -1.63 0.007949 0.0182 thaliana; supported by full-length cDNA: Ceres: 267031. 258809_at NAM-like protein (no apical meristem) similar to At3g04070 -2.4 -1.24 0.010367 0.050869 NAM GB: CAA63101 [Petunia × hybrida] 258362_at unknown protein At3g14280 -2.41 -1.2 0.007995 0.150833 249467_at NAM/CUC2-like protein CUC2, Arabidopsis At5g39610 -2.41 -1.48 0.007479 0.036136 thaliana, EMBL: ATAB2560; supported by full- length cDNA: Ceres: 113779. 251665_at responce reactor 4; supported by cDNA: At3g57040 -2.42 -1.1 0.008685 0.45678 gi_3273201_dbj_AB010918.1_AB010918 263382_at putative anthranilate N- At2g40230 -2.45 -1.06 0.018135 0.712794 hydroxycinnamoyl/benzoyltransferase; supported by full-length cDNA: Ceres: 105546. 246071_at ids4-like protein ids-4 protein-Hordeum vulgare, At5g20150 -2.47 -1.14 0.002464 0.214703 PIR: T05905; supported by full-length cDNA: Ceres: 32843. 247585_at putative protein predicted proteins, Arabidopsis At5g60680 -2.5 -1.09 0.002909 0.183267 thaliana; supported by full-length cDNA: Ceres: 16638. 264210_at putative myb-related transcription factor Similar to At1g22640 -2.51 1.26 0.00562 0.068761 myb-related transcription factor (THM27) gb|X95296 from Solanum lycopersicum. ESTs gb|T42000, gb|T04118, gb|AA598042, gb|AA394757 and gb|AA598046 come from this gene; supported by cDNA: gi_3941409_gb_AF 252965_at putative auxin-induced protein auxin-induced At4g38860 -2.53 -1.12 0.006173 0.26078 protein 10A, Glycine max., PIR2: JQ1099 253814_at putative protein; supported by full-length cDNA: At4g28290 -2.55 -1.46 0.006294 0.044367 Ceres: 10077. 246523_at CONSTANS-like 1 At5g15850 -2.55 1.01 0.013312 0.900077 263325_at putative RING zinc finger protein; supported by At2g04240 -2.56 1.18 0.004531 0.13068 cDNA: gi_13265496_gb_AF324691.2_AF324691 265342_at hypothetical protein predicted by genscan; supported At2g18300 -2.58 1.1 0.014912 0.315678 by cDNA: gi_15724317_gb_AF412099.1_AF412099 253872_at putative protein Arabidopsis thaliana nap At4g27410 -2.58 1.21 0.004591 0.057522 gene, PID: e1234813; supported by full-length cDNA: Ceres: 38344. 264783_at putative calcium-dependent protein kinase (U90439) At1g08650 -2.6 -1.61 0.010263 0.046318 similar to ESTs gb|T46119, gb|H76837, and gb|H36948; supported by cDNA: gi_6318612_gb_AF162660.1_AF162660 266363_at hypothetical protein predicted by genscan and At2g41250 -2.64 -1.63 0.002674 0.015646 genefinder 260070_at putative helix-loop-helix DNA-binding protein At1g73830 -2.64 -1.19 0.010198 0.127406 contains Pfam profile: PF00010 Helix-loop-helix DNA-binding domain 250844_at putative protein; supported by full-length cDNA: At5g04470 -2.64 1.22 0.009233 0.425631 Ceres: 13812. 256589_at cytochrome P450, putative contains Pfam profile: At3g28740 -2.66 -1.69 0.010428 0.037959 PF00067 cytochrome P450; supported by cDNA: gi_15292830_gb_AY050849.1-- 265067_at hypothetical protein predicted by At1g03850 -2.7 1.37 0.004124 0.138732 genefinder; supported by full-length cDNA: Ceres: 271253. 256914_at hypothetical protein At3g23880 -2.72 1.01 0.02091 0.915885 251169_at putative protein putative protein At2g25690- At3g63210 -2.73 1.13 0.005839 0.38507 Arabidopsis thaliana, EMBL: AC006053; supported by full-length cDNA: Ceres: 40080. 255934_at cytochrome P450, putative similar to cytochrome At1g12740 -2.74 -1.11 0.010048 0.828407 P450 GI: 4176420 from [Arabidopsis thaliana] 266150_s_at hypothetical protein At2g12290 -2.77 1.08 0.009686 0.668542 259502_at unknown protein; supported by cDNA: At1g15670 -2.77 -1.23 0.002226 0.019701 gi_15146331_gb_AY049307.1-- 263283_at hypothetical protein predicted by genscan and At2g36090 -2.79 1.26 0.004 0.034558 genefinder 253125_at DnaJ-like protein DnaJ-like protein, Phaseolus At4g36040 -2.83 1.3 0.002096 0.029069 vulgaris, PATX: G1684851 248208_at unknown protein At5g53980 -2.83 -1.12 0.002744 0.19142 264021_at putative auxin-regulated protein; supported by full- At2g21200 -2.85 1.14 0.01116 0.255677 length cDNA: Ceres: 7141. 249755_at unknown protein; supported by full-length cDNA: At5g24580 -2.87 -1.02 0.01127 0.906115 Ceres: 6393. 255284_at 5-adenylylsulfate reductase; supported by full- At4g04610 -2.9 -1.53 0.007861 0.08308 length cDNA: Ceres: 40330. 253207_at putative protein small auxin up-regulated RNA, At4g34770 -2.9 -1.24 0.004842 0.041275 Malus domestica, gb: Z93766 252118_at putative protein various predicted proteins, At3g51400 -2.9 -1.18 0.019242 0.328518 Arabidopsis thaliana; supported by full-length cDNA: Ceres: 14797. 247540_at ethylene responsive element binding factor-like At5g61590 -2.99 1.03 0.003616 0.728822 ethylene responsive element binding factor 5, Arabidopsis thaliana, SWISSPROT: ERF5_ARATH; supported by full- length cDNA: Ceres: 19893. 264379_at hypothetical protein predicted by grail At2g25200 -3 -1.3 0.023213 0.145635 263688_at unknown protein Location of EST 228A16T7A, At1g26920 -3.05 -1.22 0.00728 0.304232 gb|N65686; supported by full-length cDNA: Ceres: 24946. 246522_at bZIP DNA-binding protein-like putative bZIP At5g15830 -3.09 -1.46 0.010759 0.157233 DNA-binding protein-Capsicum chinense, EMBL: AF127797 258059_at NAM-like protein (No Apical Meristem) similar to At3g29035 -3.25 1.32 0.004473 0.140279 GB: CAA63101 from [Petunia × hybrida] (Cell 85 (2), 159-170 (1996)) 259982_at putative RING zinc finger protein contains Pfam At1g76410 -3.31 -1.01 0.011262 0.945494 profile: PF00097 Zinc finger, C3HC4 type (RING finger); supported by full-length cDNA: Ceres: 27464. 262986_at unknown protein similar to hypothetical protein At1g23390 -3.44 -1.22 0.008132 0.177105 GB: AAF27090 GI: 6730669 from (Arabidopsis thaliana); supported by full-length cDNA: Ceres: 101865. 260287_at unknown protein contains two Kelch motifs; At1g80440 -3.57 -1.18 0.008035 0.221837 supported by full-length cDNA: Ceres: 32885. 247754_at putative protein At5g59080 -3.77 -1.55 0.004279 0.021198 267238_at unknown protein; supported by full-length cDNA: At2g44130 -3.86 1.18 0.003036 0.152629 Ceres: 6950. 266156_at hypothetical protein predicted by genscan At2g28110 -3.99 1.1 0.004914 0.561885 266322_at putative auxin-regulated protein At2g46690 -4 -1.09 0.003394 0.327782 258367_at putative protein kinase similar to protein kinase At3g14370 -4.02 -1.07 0.005242 0.532182 homolog GB: AAC78477 from [Arabidopsis thaliana]; supported by full-length cDNA: Ceres: 96699. 253155_at putative protein predicted protein, Arabidopsis At4g35720 -4.2 -1.28 0.00697 0.318015 thaliana 265573_at putative zinc-finger protein similar to zinc-finger At2g28200 -4.25 -1.33 0.00752 0.040354 protein GB: AAC98446 247696_at MYB27 protein-like MYB27 protein, Arabidopsis At5g59780 -4.32 1.37 0.003111 0.017905 thaliana, PIR: T46166; supported by cDNA: gi_3941479_gb_AF062894.1_AF062894 250937_at putative protein various predicted proteins, At5g03230 -4.33 1.47 0.010367 0.036447 Arabidopsis thaliana; supported by cDNA: gi_13878024_gb_AF370275.1_AF370275 251443_at putative protein unknown protein At2g44130- At3g59940 -4.72 1.12 0.001814 0.186012 Arabidopsis thaliana, EMBL: AC004005; supported by full-length cDNA: Ceres: 8014. 261177_at hypothetical protein predicted by genemark.hmm At1g04770 -5.29 -1.41 0.001922 0.031579 249454_at expressed protein predicted protein, Synechocystis At5g39520 -5.74 -1.29 0.003609 0.135283 sp., PIR: S77152; supported by full-length cDNA: Ceres: 5331. 265387_at unknown protein; supported by full-length cDNA: At2g20670 -6.17 -1.57 0.002564 0.056602 Ceres: 34827. 254265_s_at serine threonine kinase-like protein KI domain At4g23140 2.94 1.76 0.151164 0.015078 interacting kinase 1 (KIK1), Zea mays; supported by cDNA: gi_13506746_gb_AF224706.1_AF224706 263539_at putative tyrosine aminotransferase; supported by At2g24850 2.01 2.2 0.066727 0.012756 full-length cDNA: Ceres: 14570. 265837_at unknown protein At2g14560 1.91 2.1 0.278938 0.013727 263402_at hypothetical protein similar to hypothetical protein At2g04050 1.65 1.87 0.126971 0.028665 GB: AAC27412 256766_at Expressed protein; supported by cDNA: At3g22231 1.62 1.87 0.168568 0.005967 gi_14335055_gb_AY037207.1-- 263061_at putative AAA-type ATPase At2g18190 1.48 1.94 0.324983 0.049719 267024_s_at putative aquaporin (plasma membrane intrinsic At2g34390 1.46 1.99 0.056378 0.04848 protein) 245035_at unknown protein similar to At2g26400 1.39 1.78 0.302025 0.044811 GP|2244827|gnl|PID|e326818|Z97336 252746_at sucrose synthase-like protein SUCROSE At3g43190 1.35 2.67 0.11372 0.012763 SYNTHASE (SUCROSE-UDP GLUCOSYLTRANSFERASE), Arabidopsis thalina, SWISSPROT: SUS1_ARATH; supported by cDNA: gi_14334569_gb_AY034958.1-- 263401_at hypothetical protein similar to hypothetical protein At2g04070 1.22 2.22 0.734988 0.005027 GB: AAC27412 245306_at Expressed protein; supported by full-length cDNA: At4g14690 1.2 2.16 0.22188 0.009285 Ceres: 95834. 258277_at putative cytochrome P450 similar to cytochrome At3g26830 1.04 2.61 0.754479 0.013858 P450 71B2 GB: O65788 [Arabidopsis thaliana] 252882_at Expressed protein; supported by full-length cDNA: At4g39675 -1.17 1.94 0.106839 0.012488 Ceres: 14423. 261913_at flavin-containing monooxygenase FMO3, putative At1g65860 -1.63 -1.85 0.058793 0.008577 similar to flavin-containing monooxygenase FMO3 GI: 349533 from [Oryctolagus cuniculus] 249727_at putative protein similar to unknown protein At5g35490 -1.2 -2.09 0.190584 0.008082 (gb|AAB61527.1) 254474_at putative protein predicted proteins, Arabidopsis At4g20390 -1.06 -1.71 0.572517 0.017439 thaliana; supported by full-length cDNA: Ceres: 248721. 260856_at TINY-like protein similar to TINY GB: CAA64359 At1g21910 1.17 -2.2 0.268116 0.009082 GI: 1246403 from [Arabidopsis thaliana]; supported by full-length cDNA: Ceres: 19721. 249215_at dihydroflavonol 4-reductase At5g42800 1.61 -1.77 0.401965 0.048093 254283_s_at anthocyanidin synthase-like protein putative At4g22870 2.09 -1.98 0.161214 0.018458
leucoanthocyanidin dioxygenase, Arabidopsis thaliana, PID: g1575699
TABLE-US-00010 TABLE 4 Genes that are Responsive to chitooctaose in the AtLysM RLK1 mutant WT Mu Probe set Annotation Accession FC FC WT P Mu P 253046_at cytochrome P450 - like protein cytochrome P450, At4g37370 3.83 2.17 0.019261 0.016853 Glycyrrhiza echinata, AB001379; supported by full- length cDNA: Ceres: 253698. 254869_at protein kinase - like protein KI domain interacting At4g11890 3.37 2.12 0.007665 0.003284 kinase 1 -Zea mays, PIR2: T02053 246099_at blue copper binding protein; supported by full- At5g20230 2.67 1.7 0.008061 0.011289 length cDNA: Ceres: 7767. 253317_at putative protein At4g33960 -1.83 -1.77 0.010341 0.022397 260126_at putative hydroxymethyltransferase similar to serine At1g36370 -1.93 -1.86 0.005701 0.006964 hydroxymethyltransferage GB: P50433 from [Solanum tuberosum]; supported by full-length cDNA: Ceres: 122515. 246926_at putative protein At5g25240 -2.09 -2.21 0.019603 0.017979 258217_at unknown protein contains Pfam profile PF00398 At3g17990 -2.21 -2.27 0.009887 0.0037 Ribosomal RNA adenine dimethylases 258218_at methyltransferase, putative similar to At3g18000 -2.21 -2.29 0.00667 0.009294 methyltransferase GB: AAC01738 from [Amycolatopsis mediterranei] 254343_at PRH26 protein; supported by full-length cDNA: At4g21990 -2.22 -1.83 0.012838 0.031291 Ceres: 36866. 265121_at similar to flavin-containing monooxygenase At1g62560 -2.37 -1.87 0.020126 0.00922 (sp|P36366); similar to ESTs gb|R30018, gb|H36886, gb|N37822, and gb|T88100 similar to flavin- containing monooxygenase GB: AAA21178 GI:349534 from [Oryctolagus cuniculus]; supported by cDNA: gi_13877746_gb_AF37013 251039_at putative protein hypothetical protein At5g02020 -3.73 -1.91 0.001899 0.021967 T6H20.90 - Arabidopsis thaliana, EMBL: AL096859; supported by cDNA: gi_16648747_gb_AY058150.1-- 259015_at unknown protein similar to hypothetical protein At3g07350 -3.79 -1.81 0.001762 0.010373 GB: AAC17612 [Arabidopsis thaliana]; supported by full-length cDNA: Ceres: 251012. 248676_at putative protein similar to unknown protein At5g48850 -5.55 -4.23 0.003428 0.003335 (gb|AAC72543.1) 249752_at putative protein similar to unknown protein (emb At5g24660 -5.87 -2.27 0.002654 0.005949 CAB62461.1); supported by full-length cDNA: Ceres: 268701. 254265_s_at serine threonine kinase - like protein KI domain At4g23140 2.94 1.76 0.151164 0.015078 interacting kinase 1 (KIK1), Zea mays; supported by cDNA: gi_13506746_gb_AF224706.1_AF224706 263539_at putative tyrosine aminotransferase; supported by At2g24850 2.01 2.2 0.066727 0.012756 full-length cDNA: Ceres: 14570. 265837_at unknown protein At2g14560 1.91 2.1 0.278938 0.013727 263402_at hypothetical protein similar to hypothetical protein At2g04050 1.65 1.87 0.126971 0.028665 GB: AAC27412 256766_at Expressed protein; supported by cDNA: At3g22231 1.62 1.87 0.168568 0.005967 gi_14335055_gb_AY037207.1-- 263061_at putative AAA-type ATPase At2g18190 1.48 1.94 0.324983 0.049719 267024_s_at putative aquaporin (plasma membrane intrinsic At2g34390 1.46 1.99 0.056378 0.04848 protein) 245035_at unknown protein similar to At2g26400 1.39 1.78 0.302025 0.044811 GP|2244827|gnl|PID|e326818|Z97336 252746_at sucrose synthase -like protein SUCROSE At3g43190 1.35 2.67 0.11372 0.012763 SYNTHASE (SUCROSE-UDP GLUCOSYLTRANSFERASE), Arabidopsis thalina, SWISSPROT: SUS1_ARATH; supported by cDNA: gi_14334569_gb_AY034958.1-- 263401_at hypothetical protein similar to hypothetical protein At2g04070 1.22 2.22 0.734988 0.005027 GB: AAC27412 245306_at Expressed protein; supported by full-length cDNA: At4g14690 1.2 2.16 0.22188 0.009285 Ceres: 95834. 261913_at flavin-containing monooxygenase FMO3, putative At1g65860 -1.63 -1.85 0.058793 0.008577 similar to flavin-containing monooxygenase FMO3 GI:349533 from [Oryctolagus cuniculus] 249727_at putative protein similar to unknown protein At5g35490 -1.2 -2.09 0.190584 0.008082 (gb|AAB61527.1) 258277_at putative cytochrome P450 similar to cytochrome At3g26830 1.04 2.61 0.754479 0.013858 P450 71B2 GB: O65788 [Arabidopsis thaliana] 252882_at Expressed protein; supported by full-length cDNA: At4g39675 -1.17 1.94 0.106839 0.012488 Ceres: 14423. 254474_at putative protein predicted proteins, Arabidopsis At4g20390 -1.06 -1.71 0.572517 0.017439 thaliana; supported by full-length cDNA: Ceres: 248721. 260856_at TINY-like protein similar to TINY GB: CAA64359 At1g21910 1.17 -2.2 0.268116 0.009082 GI:1246403 from [Arabidopsis thaliana]; supported by full-length cDNA: Ceres: 19721. 249215_at dihydroflavonol 4-reductase At5g42800 1.61 -1.77 0.401965 0.048093 254283_s_at anthocyanidin synthase - like protein putative At4g22870 2.09 -1.98 0.161214 0.018458 leucoanthocyanidin dioxygenase, Arabidopsis thaliana, PID: g1575699
[0131] The similar regulation patterns for this small number of genes in both the mutant and wild-type plants may be due to some redundant function provided by one of the other four LysM RLKs in the mutant. Eventually, only 6 genes appeared to behave differentially in both the mutant and wild-type (last 6 rows in Table 4). The cause of such a discrepancy is not clear. Since these few genes were only weakly to moderately regulated (-1.7 to 2.6 fold), experimental variation is a possible cause.
[0132] To determine the functional classification of the 909 genes described above, information of these genes were input into the TAIR web-based GO annotation software. The output shows that the CRGs disclosed here include many defense-related genes (such as genes encoding pathogenesis-related proteins and disease resistance proteins) and signal transduction-related genes (such as various kinases and transcription factors) (FIG. 12), suggesting a potential relationship between gene induction and plant defense mediated by chitooligosaccharides.
[0133] Since the mutation in the AtLysM RLK1 gene blocked the regulation of almost all CRGs (˜98%) by the chitooligosaccharide, AtLysM RLK1 is very likely the chitin receptor (or part of the receptor complex) that is responsible for perceiving the chitooligosaccharide elicitor and initiating cellular signaling leading to downstream gene expression. This notion is also indirectly supported by its structural features and the findings that LysM RLKs NFR1 and NFR5 in the legume Lotus japonicus serve as the receptors for the lipo-chitin Nod signal. See Limpens et al., 2003; Madsen et al., 2003; Radutoiu et al., 2003.
[0134] Because many receptor kinases form heterodimers, it has been suggested that the legume NFR1 and NFR5 may function as a heterodimer complex. See e.g., Goring et al., 2004. It is likely that AtLysRLK1 may require a partner protein, either another LysM RLK or a protein similar to the rice CEBiP. Kaku et al., 2006. However, since mutations in the other four AtLysM RLK genes had no obvious effect on the expression of selected CRGs, it seems unlikely that the products of these four genes are essential for the receptor function. There are three CEBiP-like proteins in Arabidopsis, which are encoded by At1g21880, At1g77630 and At2g17120, respectively.
[0135] If chitooligosaccharide recognition is an integral part of the response pathway by which plants defend against fungal pathogens, mutations in the AtLysM RLK1 gene should affect plant resistance to fungal pathogens. To test this hypothesis, three week-old mutant and wild-type plants were inoculated with the biotrophic powdery mildew fungal pathogen Erysiphe cichoracearum. Ten days later, the mutant plants appeared to support more fungal growth than the wild-type plants.
[0136] The susceptibility appeared to be less than that observed in NahG plants, which express salicylate hydrolase, preventing the accumulation of salicylic acid, and are therefore very susceptible to the fungal pathogen (FIG. 13A). Trypan blue staining of the infected leaves also showed the AtLysM RLK1 mutant supported more hyphal growth and production of conidiophores earlier than the wild-type plants (FIG. 13B). Arrows indicate sites where conidiophores are forming. All photographs were taken six days after inoculation. Bar=0.1 mm 4-week-old plants were also inoculated with the necrotrophic fungus Alternaria brassicicola. Three days after inoculation, the mutant developed slightly bigger lesions than the wild-type plants, as measured by average diameter of the lesions (FIGS. 13C and 13D). In agreement with this, the mutant plants also produced more spores per lesion than the wild-type plants (FIG. 13E).
[0137] To test the specificity of AtLysM RLK1 in fungal disease resistance, the response of the mutant to the bacterial pathogen Pseudomonas syringae pv. Tomato DC3000 was also examined. After infiltration with the pathogen, both the mutant and wild-type plants supported a similar bacterial growth three days after inoculation (FIG. 13F), indicating that defense against bacterial infection was not affected by the mutation. WT=wild-type Col-0; Mu=the AtLysM RLK1 mutant. CSC=crab shell chitin, 8mer=chitooctaose, and water=distilled water. Empty columns=WT Col-0 and solid black columns=the AtLysM RLK1 mutant. Asterisks indicate statistically significant differences between the mutant and wild-type plants (P<0.05). Error bars=standard error. Each experiment was repeated at least twice with similar results.
[0138] The mutation in the AtLysM RLK1 gene led to only moderate susceptibility to fungal pathogens, suggesting that AtLysM RLK1 plays a role in mediating basal or general resistance to fungal pathogens through the recognition of the chitooligosaccharide PAMP derived from fungal cell walls. This result is not surprising because it is well documented that fungal pathogens produce multiple elicitors that induce plant innate immunity. See e.g., Chisholm et al., 2006. Thus, blocking the chitin response pathway would not be expected to completely block all defense responses mounted by the plant against the fungal pathogen.
[0139] This notion is supported by the observation that chitin-responsive defense genes (e.g., MPK3 and WRKY53) were still induced by the fungal pathogen A. brassicicola in the mutant, albeit to a lower level compared with that in wild-type plants (FIG. 14). The gene induction by the fungal pathogen is monitored by quantitative RT-PCR at different time points after inoculation. Each data point is the average of the relative gene expression (fold change, normalized to actin-2 and relative to the time 0 sample) from three replicates. Error bar=standard error. WT=wild-type Col-0; Mu=the AtLysM RLK1 mutant. This low level expression of some defense genes in the mutant may explain why the mutant was only moderately susceptible to the fungal pathogens as compared to the wild-type plants.
[0140] Pretreatment of rice plants by chitooligosaccharide has been shown to enhance fungal resistance in rice. Tanabe et al., 2006. It is shown here that pretreatment of wild-type plants with chitooligosaccharides reduced disease symptoms upon inoculation with the fungal pathogen A. brassicicola, as evidenced by reduced lesion size and spore production (FIGS. 13C, 13D and 13E). Pretreatment also inhibited the growth of the bacterial pathogen P. syringae pv. tomato DC3000 (FIG. 13F), reflecting a general induction of plant innate immunity. In contrast, similar pretreatment of the AtLysM RLK1 mutant plants did not enhance resistance. These data further support the critical role for AtLysM RLK1 in mediating the perception of chitooligosaccharides by plants.
[0141] Chitin is present in the cell walls of all true fungi, but not in plants. Fungal pathogens with defects in chitin synthesis are significantly less virulent on susceptible hosts, including both plants and animals. See Bulawa et al., 1995; and Soulie et al., 2006. As disclosed herein, AtLysM RLK1 is likely a receptor for chitin PAMP of fungal pathogen. AtLysM RLK1 is only the third pattern recognition receptor (PRR) identified in plants. The other two PRRs are both leucine-rich repeat receptor-like kinases (LRR RLKs). Nurnberger et al., 2006. Therefore, this finding adds a new class of proteins to the family of plant PRRs.
[0142] LysM RLK NFR1 and NFR5 Nod signal receptors specifically recognize a lipochitin molecule with a back-bone of 4-5 units of N-acetyl-D-glucosamine. For review, see Stacey et al. 2006. This specificity is supported by the findings that mutations in either of the NFR1 and NFR5 genes in L. japonicus did not block the induction of the selected CRGs in this plant (FIG. 15). In more details, both the wild type (Gifu) and the Nod signal receptor mutants nfr1-1 and nfr5-1 were treated with chitooctaose for 30 minutes at a concentration of 1 μM or with water (as a control). The selected CRGS were detected using semi-quantitative RT-PCR. LjActin-2 was used as an internal control.
[0143] However, previous results suggested that both the legume NFR genes and AtLysM RLK1 are under negative selection, implying the functional conservation between legume NFR and AtLysM RLK1 genes. Zhang et al., 2007. Perhaps, the discrepancy in substrate specificity lies in the difference in their extracellular LysM domains, since legume NFR proteins have two LysM motifs while AtLysM RLK1 has three. Zhang et al., 2007.
[0144] To determine whether the AtLysM RLK1 mutation affects other defense-related pathways, such as the salicylic acid (SA) and jasmonic acid/ethylene (JA/ETH) responsive pathways, the AtLysM RLK1 mutant and wild-type plants were treated with SA, methyl jasmonic acid (MeJA), and 1-aminocyclopropane-1-carboxylic acid (ACC), respectively, and expression of the pathway hallmark genes, PR-1 (the SA pathway) and PDF1.2 (the JA/ETH pathway) was examined.
[0145] Quantitative RT-PCR data demonstrated that both the mutant and wild-type plants showed similar induction of PR-1 by SA and of PDF1.2 by MeJA or ACC, indicating that the mutation did not affect these defense pathways (FIG. 16). In addition, the mutant plants were fully responsive to another typical PAMP, the flagellin-derived peptide flg22 (FIG. 16). Each data point is the average of the relative gene expression (fold change, normalized to actin-2 and relative to the control sample) from three replicates. Error bar=standard error. No statistically significant differences are found between the mutant and wild type in the induction of the above genes.
[0146] Interestingly, as shown by FIG. 17, the AtLysM RLK1 mutation does not block the induction of flagellin-responsive genes. Collectively, the data indicate that AtLysM RLK1 is the primary, specific receptor for recognition of the chitooligosaccharide PAMP derived from fungal pathogen cell walls. This recognition is a crucial step in the elicitation of protective innate immunity responses in the plant.
Example 4
Forced Expression of Certain LysM RLK Genes Enhanced Fungal Resistance in Plants
[0147] Over-expression of one LysM RLK gene, At2g33580, under a strong cauliflower mosaic virus (CMV) 35S promoter in transgenic plants, resulted in enhanced disease resistance. Therefore, this gene may function in a positive way to elevate disease resistance, similar to the mechanism of the At3g21630 gene, as shown in FIG. 5. Together with the data in Example 3 showing that forced expression of AtLysM RLK1 gene can restore induction of CRGs in a AtLysM RLK1 insertion mutant, these data confirm that the expression of specific LysM RLK genes in transgenic plants may confer enhanced disease resistance.
Example 5
Induction of Gene Expression by Chitin or its Derivatives
[0148] Based on the genome-wide gene expression studies using microarrays, quantitative reverse transcriptase-polymerase chain reaction (qRT-PCR) was conducted to identify over 120 different transcription factors in Arabidopsis thaliana whose expression is enhanced by chitin treatment. Many of these transcription factors are known to be involved in fungal pathogen response. This again shows the correlation of chitin response to disease resistance. Each of these transcription factors is a potential target for genetic manipulation in order to enhance disease resistance. The results of this study are described in greater details in Libault et al., 2007, which is hereby expressly incorporated by reference.
[0149] Some of the genes disclosed herein, including the LysM RLK genes, the CRGs such as the transcription factors described above, may play a positive role in plant fungal defense. Such genes may be called positive regulators. Transgenic plants may be generated wherein these positive regulators are expressed at an elevated level to increase the expression of transcription factors or other downstream genes. Some of the genes, on the other hand, may inhibit a plant's fungal defense capability. Such genes may be called negative regulators (also called "negative regulatory genes"). Deletion mutants of such genes that play a negative role in fungal defense may be created to obtain plants with enhanced fungal defense capability. Alternatively, dominant negative mutants of the negative regulatory genes may be introduced into a wild-type plant to inhibit the function of negative factors. Host plants may include any plants that may be susceptible to fungal infection, such as Arabidopsis thaliana, soybean, and others.
Example 6
LysM Containing Proteins in Soybean
[0150] Utilizing the gene sequences from Arabidopsis, as described above, a total of 13 LysM RLK genes were identified in the soybean genome by searching the dbEST sequence database maintained by the Institute of Genomic Research. These 13 soybean LysM RLKs are: GmNFR1α (also referred to as GmNFR1a in this disclosure), GmNFR1β (also referred to as GmNFR1b in this disclosure), GmLYK2, GmLYK3, GmLYK4 (previously known as GmLysM17), GmNFR5α (also referred to as GmNFR5a in this disclosure), GmNFR5β (also referred to as GmNFR1b in this disclosure), GmLYK6 (previously known as GmLysM14), GmLYK7, GmLYK8 (previously known as GmLysM4), GmLYK9 (previously known as GmLysM16), GmLYK10, GmLYK11, and are designated as SEQ ID Nos. 54-66, respectively.
[0151] In addition, an additional fifteen soybean genes were identified that appear to have a LysM domain, but no associated kinase domain. PCR primers were developed for several of these genes, and their location on the soybean bacterial artificial chromosome (BAC)-based physical map was determined by probing pools of individual BAC clones. In this way, BAC clones encoding the various LysM domain proteins were identified. At this time, twelve BACs have been sequenced and the genomic sequences, including the regulatory regions, have been obtained for several LysM RLK genes.
[0152] Sequence comparisons between the soybean BAC sequences and other plant species showed examples where gene order (microsynteny) was well conserved. For example, FIG. 18 shows microsynteny between the soybean GmNFR5 and LysM17 (GmLYK4) gene-encoding regions and corresponding regions in poplar (Pt), Lotus japonicus (Lj), and Medicago trancatula (Mt) Arabidopsis thaliana (At) and rice (Os). These regions of microsynteny may be expanded by use of these methods to other plant species. Thus, knowledge of the genomic location in soybean can allow for the identification of the likely functional orthologue in other plant species, or vice versa.
[0153] In some cases, mapping the gene to the physical map also gave a genetic map location, due to genetic markers associated with the BAC clones. Table 5 shows examples of such regions that are in proximity to LysM RLK encoding regions. The locations of these genes were correlated with known quantitative trait loci (QTLs) associated with fungal resistance, i.e., Sclerotina white mold, Asian soybean rust and sudden death syndrome. In each case, a close correlation existed between the location of the LysM RLK and a known QTL for disease resistance. Thus, mapping of the LysM RLK may aid in the localization of disease resistance QTLs in soybean. The sequence of the LysM RLK gene can also be used to define better genetic markers for fine mapping of the associated QTLs. For instance, soybean mutants may be generated and selected for fungal resistant phenotypes. The close genetic link between certain QTLs and some LysM RLK genes may allow one to use the LysM RLK gene sequences to trace segregation of the QTLs. For example, molecular markers in the form of PCR primers, oligonucleotide probes, single nucleotide polymorphisms, restriction fragment polymorphisms, among others, derived from the DNA sequence of the LYK genes could be very useful in following a specific QTL in a breeding process.
TABLE-US-00011 TABLE 5 Associations of GmLysM-RLKs with known fungal resistance QTLs Genetic Genetic Linkage position Soybean LysM-RLKs marker Group (cM) Sclerotinia sclerotiorum QTL Satt172 D1b 100.89 Sclerotinia sclerotiorum QTL Satt459 D1b 118.62 K411_1 D1b 119.34 GmLysM4, GmLysM26, GmNFR1a A343_2 D1b 120.97 Sclerotinia sclerotiorum QTL Satt143 L 30.19 GmLysM23 Sat_388 L 30.86 Sclerotinia sclerotiorum QTL Satt481 L 54.57 GmLysM25 Sat_402 C2 103.33 Asian soybean rust QTL Satt460 C2 117.77 Fusarium solani f. sp glycines (SDS) Satt307 C2 121.27 QTL
[0154] More particularly, all or a fragment of the polynucleotides of the instant disclosure may be used as probes for genetically and physically mapping the genes of which they are a part, and can be further used as markers for traits linked to those genes. Such information may be useful in plant breeding in order to develop lines with desired phenotypes. For example, the instant nucleic acid fragments may be used as restriction fragment length polymorphism (RFLP) markers. Southern blots of restriction-digested plant genomic DNA may be probed with the nucleic acid fragments of the instant disclosure. The resulting banding patterns may then be subjected to genetic analyses using computer programs such as MapMaker (Lander et al. (1987) Genomics 1:174-181) in order to construct a genetic map. In addition, the nucleic acid fragments of the instant disclosure may be used to probe Southern blots containing restriction endonuclease-treated genomic DNAs of a set of individuals representing parent and progeny of a defined genetic cross. Segregation of the DNA polymorphisms is noted and used to calculate the position of the instant nucleic acid sequence in the genetic map previously obtained using this population (Botstein et al. (1980) Am. J. Hum. Genet. 32:314-331).
[0155] The production and use of plant gene-derived probes for use in genetic mapping is described in Bernatzky and Tanksley (1986) Plant Mol. Biol. Reporter 4:37-41. Numerous publications describe genetic mapping of specific cDNA clones using the methodology outlined above or variations thereof. For example, F2 intercross populations, backcross populations, randomly mated populations, near isogenic lines, and other sets of individuals may be used for mapping. Such methodologies are well known to those skilled in the art.
[0156] Nucleic acid probes derived from the instant nucleic acid sequences may also be used for physical mapping (i.e., placement of sequences on physical maps; see Hoheisel et al. In: Non mammalian Genomic Analysis: A Practical Guide, Academic press 1996, pp. 319-346, and references cited therein). In another embodiment, nucleic acid probes derived from the instant nucleic acid sequences may be used in direct fluorescence in sits hybridization (FISH) mapping (Trask (1991) Trends Genet. 7:149-154). Although current methods of FISH mapping favor use of large clones ranging from a few Kb to several hundred Kb; see Laan et al. (1995) Genome Res. 5:13-20), improvements in sensitivity may allow performance of FISH mapping using shorter probes.
[0157] A variety of nucleic acid amplification-based methods of genetic and physical mapping may be carried out using the nucleic acid sequences of the instant disclosure. Examples include allele-specific amplification (Kazazian (1989) J. Lab. Clin. Med. 11:95-96), polymorphism of PCR-amplified fragments (CAPS; Sheffield et al. (1993) Genomics 16:325-332), allele-specific ligation (Landegren et al. (1988) Science 241:1077-1080), nucleotide extension reactions (Sokolov (1990) Nucleic Acid Res. 18:3671), Radiation Hybrid Mapping (Walter et al. (1997) Nat. Genet. 7:22-28) and Happy Mapping (Dear and Cook (1989) Nucleic Acid Res. 17:6795-6807). For these methods, the sequence of a nucleic acid fragment is used to design and produce primer pairs for use in the amplification reaction or in primer extension reactions. The design of such primers is well known to those skilled in the art. In methods employing PCR-based genetic mapping, it may be necessary to identify DNA sequence differences between the parents of the mapping cross in the region corresponding to the instant nucleic acid sequence. This, however, is generally not necessary for mapping methods.
[0158] Soybean genotypes used for mapping of soybean QTLs for white mold resistance included Corsoy 79 and Dassel. As shown in FIG. 19, treatment of leaves of these plants, as well as Williams 82 as a control, resulted in strong induction of three of the LysM RLK genes out of a total of six such genes tested. These genes are, therefore, excellent targets for genetic manipulation using the methods demonstrated in Arabidopsis to create soybean plants with decreased disease susceptibility. Finally, as was the case with Arabidopsis, some of the soybean LysM RLK genes are induced upon treatment of soybean with chitin, as confirmed by the results shown in FIG. 20. Leaves were treated by spraying with chitin (100 μM)+0.2% Tween-20.
Example 7
Tissue Specific Expression of LysM-Containing Proteins in Soybean and Other Plants
[0159] Other experiments examined the expression of the various LysM domain-containing genes under various conditions. For the six plant species in this study, tissue expression levels of LYK genes have only been reported for M. truncatula (Limpens et al., 2003; Arrighi et al., 2006) and Arabidopsis. Therefore, LYK gene expression was measured using quantitative reverse transcription (RT)-PCR in different tissues of soybean, M. truncatula, and rice plants.
[0160] More particularly, Soybean, M. truncatula, and rice plants were grown in the greenhouse at 28° C. to 30° C. with a 16-h light/8-h dark cycle. Roots and vegetative tissues were sampled about 3 weeks after planting and flowers were sampled about 3 months after planting. Total RNAs were extracted using Trizol (Invitrogen) followed by Turbo DNase (Ambion) treatment to remove genomic DNA contamination. First-strand cDNAs were synthesized using Moloney murine leukemia virus reverse transcriptase (Promega). Quantitative RT-PCR was performed using a 7500 real-time PCR system (Applied Biosystems) following standard procedures. The primer sequences are listed in Table 6. q-RT-PCR of GmSubi2, MtActin2, OsEF1α are used to normalize the expression data of all of the other genes in the respective species.
TABLE-US-00012 TABLE 6 Plant LYK primers for qRT-PCR (5' to 3' from left to right) SEQ SEQ Genes Forward ID No. Reverse ID No. GmSubi2 AGCTATTCGCAGTTCCCAAAT 96 CAGAGACGAACCTTGAGGAGA 97 GmNFR1a AAGAACATCCGTGGAAAGGTT 98 AATGTTCCCACAAGACGAGTG 99 GmNFR1b TGACATATGCCAATCTCACCA 100 GTGACATTAACCGTGGCATTT 101 GmLYK2 GATCCACAACAACGTCCAAAT 102 ATGGAAGCAATATCCCAATCC 103 GmLYK3 TAACGGTGACGTTGATGTTCA 104 GTTGTCGAGGTTGATTTCTCG 105 GmLYK4 AGATGTGCTTGTCCCACAAAG 106 CAGAATCACCCCAGTTTACCA 107 GmNFR5a ACCGCTCTTTTGCCAATATCT 108 AACGGGGTTTAAATCCATCAC 109 GmNFR5b CATGGCCAGAACTTTTACCAA 110 GTTGTCATGGCTTTCCTACCA 111 GmLYK9 TGATCTCCTACGTCGTCCAAC 112 GCGTCAATGATGGACTGTTCT 113 GmLYK10 CCTCTCTCTCCAACCTCACCT 114 CTGATCCTGGGAGAGGAACTC 115 GmLYK11 TTCGGTTCCTGGTGAGTCTTA 116 TCATGGGGTACATGAGCTTTC 117 MtActin2 TGGCATCACTCAGTACCTTTCAACAG 118 ACCCAAAGCATCAAATAATAAGTCAACC 119 MtLYK1 CATGAGCATTCAGTGCCTGT 120 TGCAGAATCAGTAAGCCTGGT 121 MtLYK3 TGCTAAGGGTTCAGCTGTTGGTA 122 AAATGCCCTAGAAGTTTGTGGAAG 123 MtLYK4 CGCAAGATGGATGTGTATGC 124 CATGGCTCTCGAACTCGTTT 125 MtLYK9 CACTCATATTCTTTTCTGCCACCCA 126 TGCAATGGATTGAGGACTGGTGT 127 MtLYK10 GGAAATGGAGAAATGGCAAA 128 CGCCTTGACCAAGAAACCTA 129 MtLYK11 GGCATTGATGGGTCAGAACT 130 TGCAAAGAGGATCACACTGC 131 MtLYK12 CTCTTCTTCTTCTTCTTCGTCAGCA 132 GGTATGCTTGGCATGTTTGAGTTT 133 MtLYK13 GGTTGTTCTCGGAATCTTCG 134 ATGCATGTATTGCAGACCGA 135 OsEF1α TTTCACTCTTGGTGTGAAGCAGAT 136 GACTTCCTTCACGATTTCATCGTAA 137 OsLYK1 ATGGCGATATGGGTGACATT 138 TCCACATGGAAGGTGAATGA 139 OsLYK2 GTTCTTGCGTCTGGTGCTCT 140 CTCCTTATCCGGAGCCAAC 141 OsLYK3 ATGGAGGAGGTGTTCGTCAC 142 CCGAGGACCATAGAAGCTGA 143 OsLYK4 CATGGTCACCTACCTCGTCA 144 TATGATGGAGCTCTCGGTGA 145 OsLYK5 GTTCATCGACAAACCGATCA 146 TAATACGAGCTGCCGAGCTT 147 OsLYK6 GTGACGAGGAGAATGGAGGA 148 CTCGATCAGCTTCACCATCA 149
[0161] The data agree well with previous results on MtLYK expression levels (Limpens et al., 2003; Arrighi et al., 2006). It was also found that plant LYK expression was generally tissue specific and that most plant LYK genes were expressed predominantly in the root in soybean (FIG. 21A), M. truncatula (FIG. 21B), rice (FIG. 21C), L. japonicus (Madsen et al., 2003; Radutoiu et al., 2003), and Arabidopsis, although a few genes were expressed in stems and leaves. Expression levels of each LYK gene were displayed in artificial scales relative to particular housekeeping genes. Data were collected from three biological replicates. Error bars represent SDs.
[0162] As predicted from their orthologous relationships (Zhang et al, 2007), GmNFR1, GmNFR1, MtLYK3, and LjNFR1 (Radutoiu et al., 2003) showed similar patterns of root-specific expression. Similarly, GmNFR5, GmNFR5, MtLYK13, OsLYK5, and LjNFR5 (Madsen et al., 2003) showed root-specific expression. These results are reasonable, from a biological perspective, because these receptors need to efficiently contact Nod factors secreted by soilborne symbiotic bacteria.
[0163] Additionally, the following sets of orthologous genes also exhibit similar expression patterns: GmLYK4 and MtLYK12; GmLYK10 and AtLYK2; GmLYK8 (data not shown); and AtLYK5. Interestingly, several duplicated genes displayed different expression patterns. For example, GmLYK2 and MtLYK11 expression is dramatically different from duplicated partners, GmNFR1 and MtLYK10, respectively. MtLYK9, paralogous to MtLYK13, is expressed differently from the latter. These data suggest the functional diversification of LYK genes after duplications.
[0164] In light of the detailed description of the invention and the examples presented above, it can be appreciated that the several aspects of the invention are achieved.
[0165] It is to be understood that the present invention has been described in detail by way of illustration and example in order to acquaint others skilled in the art with the invention, its principles, and its practical application. Particular formulations and processes of the present invention are not limited to the descriptions of the specific embodiments presented, but rather the descriptions and examples should be viewed in terms of the claims that follow and their equivalents. While some of the examples and descriptions above include some conclusions about the way the invention may function, the inventors do not intend to be bound by those conclusions and functions, but put them forth only as possible explanations.
[0166] Moreover, while most of the examples provided use Arabidopsis or soybean as the host plant, it is to be understood that the transgenic and plant breeding procedures described herein are broadly applicable to other plant species as well. These other plants may include but are not limited to: Rice, Wheat, Barley, poplar, M. truncatula, L. japonicus and many other crops, vegetables, and trees. Although transformation and breeding procedures differ from one plant species to another, it is within the skills of an ordinary artisan to modify the teaching of this disclosure for use in other plants. The methodology for conferring fungal resistance upon a plant or for selecting for a fungal resistant plant may be applicable to a broad spectrum of fungi, such as Fusarium, Powdery mildew, and the variety of fungi that cause soybean rust, among others.
[0167] It is to be further understood that the specific embodiments of the present invention as set forth are not intended as being exhaustive or limiting of the invention, and that many alternatives, modifications, and variations will be apparent to those of ordinary skill in the art in light of the foregoing examples and detailed description. Accordingly, this invention is intended to embrace all such alternatives, modifications, and variations that fall within the spirit and scope of the following claims.
REFERENCES
[0168] Full citations of references that are not fully cited in the text are listed below. All references, including those that are not listed below but are fully cited in the text, are hereby incorporated by reference to the same extent as though fully disclosed herein:
[0169] 1. Arrighi J, Bane A, Ben Amor B, Bersoult A, Soriano L, Mirabella R, Carvalho-Niebel F, Journet E, Gherardi M, Huguet T, et al (2006) The Medicago truncatula lysine motif-receptor-like kinase gene family includes NFP and new nodule-expressed genes. Plant Physiol 142: 265-279.
[0170] 2. Bulawa, C. E., D. W. Miller, L. K. Henry, J. M. Becker, Proc. Natl. Acad. Sci. U.S.A. 92, 10570-10574 (1995).
[0171] 3. Chisholm, S. T., G. Coaker, B. Day, B. J. Staskawicz, Cell 124, 803-814 (2006).
[0172] 4. Clamp M, Cuff J, Searle S M, Barton G J (2004) The Jalview Java alignment editor. Bioinformatics 20: 426-427.
[0173] 5. Day, R. B. et al., Plant Physiol. 126, 1162-1173 (2001).
[0174] 6. Eddy S R (1998) Profile hidden Markov models. Bioinformatics 14: 755-763.
[0175] 7. Felsenstein J (2000) PHYLIP (Phylogeny Inference Package), Ed 3.6. University of Washington, Seattle.
[0176] 8. Gomez-Gomez, L., T. Boller, Mol. Cell 5, 1003-1011 (2000).
[0177] 9. D. R. Goring, J. C. Walker, Science 303, 1474-1475 (2004).
[0178] 10. Ito, Y., H. Kaku, N. Shibuya, Plant J. 12, 347-356 (1997).
[0179] 11. Joris, B., FEMS Microbiol. Lett. 70, 257-264 (1992).
[0180] 12. Kaku, H. et al., Proc. Natl. Acad. Sci. U.S.A. 103, 11086-11091 (2006).
[0181] 13. Limpens et al., Science 302, 630-633 (2003).
[0182] 14. Libault, M., J. Wan, T. Czechowski, M. Udvardi, G. Stacey, Mol. Plant-Microbe Interact. (in press) (2007).
[0183] 15. Libault M, Wan J, Joshi T, Zhang X, Czechowski T, Xu D, Udvardi M, Stacey G (2006) The regulation of Arabidopsis transcription factor and ubiquitin-ligase genes by chitin allows the identification of a G-box motif highly represented in their promoter sequences. Plant Physiol. (submitted).
[0184] 16. Libault, M., J. Wan, T. Czechowski, M. Udvardi, and G. Stacey, Identification of 118 Arabidopsis Transcription Factor and 30 Ubiquitin-Ligase Genes Responding to Chitin, a Plant-Defense Elicitor. Molecular Plant-Microbe Interactions, Vol. 20, No. 8, 2007, pp. 900-911.
[0185] 17. Madsen, E B; Madsen, L H; Radutoiu, S; Olbryt, M; Rakwalska, M; Szczyglowski, K; Sato, S; Kaneko, T; Tabata, S; Sandal, N; Stougaard, J. 2003. A receptor kinase gene of the LysM type is involved in legume perception of rhizobial signals. Nature 425 (6958): 637-640.
[0186] 18. Nurnberger, T., B. Kemmerling, Trends Plant Sci. 11, 519-522 (2006).
[0187] 19. Okada, M. and M. Matsumura, Y. Ito, N. Shibuya, Plant Cell Physiol. 43, 505-512 (2002).
[0188] 20. Passarinho, P., S. C. de Vries, In: The Arabidopsis Book, American Society of Plant Biologists (2002).
[0189] 21. Radutoiu, S; Madsen, L H; Madsen, E B; Felle, H H; Umehara, Y; Gronlund, M; Sato, S; Nakamura, Y; Tabata, S; Sandal, N; Stougaard, J. 2003. Plant recognition of symbiotic bacteria requires two LysM receptor-like kinases. Nature 425 (6958): 585-592.
[0190] 22. Ramonell, K., B. Zhang, R. Ewing, Y. Chen, D. Xu, G. Stacey, and S. Somerville. 2002 Microarray analysis of chitin elicitation in Arabidopsis thaliana. Mol. Plant Pathol. 3 (1): 301-311.
[0191] 23. Ramonell K, Berrocal-Lobo M, Koh S, Wan J, Edwards H, Stacey G and Somerville S. 2005. Loss-of-function mutations in four chitin responsive genes show increased susceptibility to the powdery mildew pathogen, Erysiphe cichoracearum. Plant Physiol. 138: 1027-1036
[0192] 24. Schmidt H A, Strimmer K, Vingron M, von Haeseler A (2002) TREE-PUZZLE: maximum likelihood phylogenetic analysis using quartets and parallel computing. Bioinformatics 18: 502-504.
[0193] 25. Shibuya, N., E. Minami, Physiol. Mol. Plant Pathol. 59, 223-233 (2001).
[0194] 26. Soulie, M. C. et al., Cell Microbiol. 8, 1310-1321 (2006).
[0195] 27. Stacey, G. and N. Shibuya. 1997. Chitin recognition in rice and legumes. Plant and Soil 194: 161-169.
[0196] 28. G. Stacey, M. Libault, L. Brechenmacher, J. Wan, G. D. May, Curr. Opin. Plant. Biol. 9, 110-121 (2006).
[0197] 29. Tanabe, S., et al., Biosci. Biotechnol. Biochem. 70, 1599-1605 (2006).
[0198] 30. Thompson J D, Gibson T J, Plewniak F, Jeanmougin F, Higgins D G (1997) The Clustal-X windows interface-flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res 25: 4876-4882
[0199] 31. Wan, Jinrong, Shuqun Zhang, and Gary Stacey. 2004. Activation of a potential mitogen-activated protein kinase pathway in Arabidopsis by chitin. Mol. Plant Pathol. 5(1): 125-135.
[0200] 32. Yang Z (1997) PAML: a program package for phylogenetic analysis by maximum likelihood. Comput Appl Biosci 13: 555-556.
[0201] 33. Zhang, B., K. Ramonell, S. Somerville, and G. Stacey. 2002. Characterization of Early, Chitin-Induced Gene Expression in Arabidopsis Mol. Plant-Microbe Int. 15: 963-970.
[0202] 34. Zhang, X.-C. et al., Plant Physiol. 144, 623-636 (2007).
[0203] 35. Zmasek C M, Eddy S R (2001) ATV: display and manipulation of annotated phylogenetic trees. Bioinformatics 17: 383-384.
Sequence CWU
1
1
21512205DNALotus japonicus 1aagtgtgaca ttagtttcaa gagaaaaata aatgatcaaa
acctggtaga gagtcctaga 60aattcaatgt tctgatttct ttcattcatc tctgctgcca
ttttgatttg cacaatgaag 120ctaaaaactg gtctactttt gtttttcatt cttttgctgg
ggcatgtttg tttccatgtg 180gaatcaaact gtctgaaggg gtgtgatcta gctttagctt
cctattatat cttgcctggt 240gttttcatct tacaaaacat aacaaccttt atgcaatcag
agattgtctc aagtaatgat 300gccataacca gctacaacaa agacaaaatt ctcaatgata
tcaacatcca atcctttcaa 360agactcaaca ttccatttcc atgtgactgt attggtggtg
agtttctagg gcatgtattt 420gagtactcag cttcaaaagg agacacttat gaaactattg
ccaacctcta ctatgcaaat 480ttgacaacag ttgatctttt gaaaaggttc aacagctatg
atccaaaaaa catacctgtt 540aatgccaagg ttaatgtcac tgttaattgt tcttgtggga
acagccaggt ttcaaaagat 600tatggcttgt ttattaccta tcccattagg cctggggata
cactgcagga tattgcaaac 660cagagtagtc ttgatgcagg gttgatacag agtttcaacc
caagtgtcaa tttcagcaaa 720gatagtggga tagctttcat tcctggaaga tataaaaatg
gagtctatgt tcccttgtac 780cacagaaccg caggtctagc tagtggtgca gctgttggta
tatctattgc aggaaccttc 840gtgcttctgt tactagcatt ttgtatgtat gttagatacc
agaagaagga agaagagaaa 900gctaaattgc caacagatat ttctatggcc ctttcaacac
aagatgcctc tagtagtgca 960gaatatgaaa cttctggatc cagtgggcca gggactgcta
gtgctacagg tcttactagc 1020attatggtgg cgaaatcaat ggagttctca tatcaggaac
tagcgaaggc tacaaataac 1080tttagcttgg ataataaaat tggtcaaggt ggatttggag
ctgtctatta tgcagaattg 1140agaggcaaga aaacagcaat taagaagatg gatgtacaag
catcaacaga atttctttgt 1200gagttgaagg tcttaacaca tgttcaccac ttgaatctgg
tgcgcttgat tggatactgc 1260gttgagggat ctctattcct tgtttatgaa catattgaca
atggaaactt aggccaatat 1320ttgcatggtt caggtaaaga accattgcca tggtctagcc
gagtacaaat agctctagat 1380gcagcaagag gccttgaata cattcatgag cacactgtgc
ctgtgtatat ccatcgcgat 1440gtgaaatctg caaacatatt gatagataag aacttgcgtg
gaaaggttgc agattttggc 1500ttgaccaagc ttattgaagt tgggaactcc acactacaaa
ctcgtctggt gggaacattt 1560ggatacatgc ccccagaata tgctcaatat ggtgatattt
ctccaaaaat agatgtatat 1620gcatttggag ttgttctttt tgaacttatt tctgcaaaga
atgctgttct gaagacaggt 1680gaattagttg ctgaatcaaa gggccttgta gctttgtttg
aagaagcact taataagagt 1740gatccttgtg atgctcttcg caaactggtg gatcctaggc
ttggagaaaa ctatccaatt 1800gattctgttc tcaagattgc acaactaggg agagcttgta
caagagataa tccactgcta 1860agaccaagta tgagatcttt agttgttgct cttatgaccc
tttcatcact tactgaggat 1920tgtgatgatg aatcttccta cgaaagtcaa actctcataa
atttactgtc tgtgagataa 1980aggttctcca tgcaaatgca tgtttgttat atatatcttg
tagtacaact aagcagacaa 2040aaagttttgt actttgaatg taaatcgagt cagggtgttt
acattttatt actccaatgt 2100ttaattgcca aaaccatcaa aaagtcctag gccagacttc
ctgtaattat atttagcaaa 2160gttgcagatt ctaagttcag tttttttaaa aaaaaaaaaa
aaaaa 220522292DNALotus japonicus 2ttattgatat
actaaaccac aggatatttt attgacaatg tgaatgttcc atattttcaa 60caatgctgat
tccctctgat aaagaacaag ttccttttct ctttccctgt taactatcat 120ttgttcccca
cttcacaaac atggctgtct tctttcttac ctctggctct ctgagtcttt 180ttcttgcact
cacgttgctt ttcactaaca tcgccgctcg atcagaaaag attagcggcc 240cagacttttc
atgccctgtt gactcacctc cttcttgtga aacatatgtg acatacacag 300ctcagtctcc
aaatcttctg agcctgacaa acatatctga tatatttgat atcagtcctt 360tgtccattgc
aagagccagt aacatagatg cagggaagga caagctggtt ccaggccaag 420tcttactggt
acctgtaact tgcggttgcg ccggaaacca ctcttctgcc aatacctcct 480accaaatcca
gctaggtgat agctacgact ttgttgcaac cactttatat gagaacctta 540caaattggaa
tatagtacaa gcttcaaacc caggggtaaa tccatatttg ttgccagagc 600gcgtcaaagt
agtattccct ttattctgca ggtgcccttc aaagaaccag ttgaacaaag 660ggattcagta
tctgattact tatgtgtgga agcccaatga caatgtttcc cttgtgagtg 720ccaagtttgg
tgcatcccca gcggacatat tgactgaaaa ccgctacggt caagacttca 780ctgctgcaac
caaccttcca attttgatcc cagtgacaca gttgccagag cttactcaac 840cttcttcaaa
tggaaggaaa agcagcattc atcttctggt tatacttggt attaccctgg 900gatgcacgtt
gctaactgca gttttaaccg ggaccctcgt atatgtatac tgccgcagaa 960agaaggctct
gaataggact gcttcatcag ctgagactgc tgataaacta ctttctggag 1020tttcaggcta
tgtaagcaag ccaaacgtgt atgaaatcga cgagataatg gaagctacga 1080aggatttcag
cgatgagtgc aaggttgggg aatcagtgta caaggccaac atagaaggtc 1140gggttgtagc
ggtaaagaaa atcaaggaag gtggtgccaa tgaggaactg aaaattctgc 1200agaaggtaaa
tcatggaaat ctggtgaaac taatgggtgt ctcctcaggc tatgatggaa 1260actgtttctt
ggtttatgaa tatgctgaaa atgggtctct tgctgagtgg ctgttctcca 1320agtcttcagg
aaccccaaac tcccttacat ggtctcaaag gataagcata gcagtggatg 1380ttgctgtggg
tctgcaatac atgcatgaac atacctatcc aagaataata cacagggaca 1440tcacaacaag
taatatcctt ctcgactcga acttcaaggc caagatagcg aatttcgcca 1500tggccagaac
ttcgaccaac cccatgatgc caaaaatcga tgtcttcgct ttcggggtgc 1560ttctgataga
gttgctcacc ggaaggaaag ccatgacaac caaggagaac ggcgaggtgg 1620ttatgctgtg
gaaggatatg tgggagatct ttgacataga agagaataga gaggagagga 1680tcagaaaatg
gatggatcct aatttagaga gcttttatca tatagataat gctctcagct 1740tggcatcctt
agcagtgaat tgcacagctg ataagtcttt gtctcgaccc tccatggctg 1800aaattgttct
tagcctctcc tttctcactc aacaatcatc taaccccaca ttagagagat 1860ccttgacttc
ttctgggtta gatgtagaag atgatgctca tattgtgact tccattactg 1920cacgttaagc
aagggaaggt aattcagttt ctcatcaaat tgatcaagat gcactttgtt 1980tgcgtggtta
ctattacatt tttaactagc tatttgctta tttctctgta tttatttgtc 2040agacactgga
attgaatatc atatgatgga ggagttgtct gttaatacat gtgctaataa 2100caaattcagg
caagatagtt aattgcattt gaaatacata tttctgctca gagatggtga 2160acatccatgc
tccgaagctc atattaagtg tggtagctat tttcttttca tctttttggg 2220gtgaatgcgt
gttcatgtaa ctcgtaaggt gttatatatt acagaagtcg tatacgtcgt 2280tccaaaaaaa
aa
229232063DNAArabidopsis thaliana 3aaacaaaaaa aacagagacc cttcctctgg
ggaatctaag gaagcttcaa aatgaagcta 60aagatttctc taatcgctcc gattcttctt
ctcttctcat tcttcttcgc cgtggaatct 120aagtgcagga ctagctgtcc tttagctcta
gcttcgtact atctcgagaa cggaacaaca 180ctctccgtca tcaaccaaaa cctcaattct
tcaatcgcgc cttacgatca aatcaatttc 240gatccaatcc tcaggtacaa cagtaacatt
aaagacaaag atagaatcca gatgggctct 300agggttcttg tacctttccc ttgcgaatgt
caacctggtg atttcctagg gcacaatttc 360agctacagtg ttcgacagga agatacttac
gaaagagtcg cgattagtaa ttacgcgaat 420ctcacgacga tggagtcgtt acaggcgagg
aatccttttc cggcgactaa catacctctc 480tctgcgacgc ttaatgtatt ggtgaattgt
tcttgtggtg atgagagtgt ttcgaaagat 540tttggtttgt ttgttacgta tccgcttcgt
cctgaagaca gtctcagttc tattgcgaga 600tcttccggtg tatcggcgga tattctgcag
agatataatc ccggtgttaa ttttaactcc 660gggaatggaa tcgtttatgt gcctggaaga
gatccaaatg gtgcatttcc accattcaaa 720tcaagtaaac aagatggtgt tggtgctgga
gttattgctg gtatagttat aggagtgatt 780gtggctttgt tgttgatctt gtttatcgta
tattatgctt accggaagaa taagtcgaag 840ggtgattcgt tttcttcttc tattccgttg
tctactaagg ctgatcatgc ttcttctact 900agtctccaaa gtggaggttt gggtggtgcc
ggagtgtctc ctggcattgc tgccataagc 960gtggacaaat ctgttgagtt ttcgttggag
gaactagcaa aggctactga taatttcaat 1020ttgtctttta agattgggca aggtggtttt
ggggctgttt actatgcaga gctgagagga 1080gaaaaagctg cgattaagaa gatggacatg
gaggcatcga aacagttctt ggcggaacta 1140aaagtcttaa cgcgtgtaca tcatgtcaac
ctggttcgcc tgattggata ttgtgttgag 1200ggatcacttt tcttggtgta tgaatatgtt
gagaatggta accttggaca acatttacat 1260gggtcaggac gagaaccatt accgtggact
aagagagtgc agattgcact agactcagct 1320agaggtttag aatatatcca cgagcacacg
gttccagttt atgtccatag ggacattaaa 1380tctgccaata ttttgataga ccagaaattc
cgagcaaagg tagcagattt cgggttaaca 1440aaactgacag aagttggagg ttcagcaact
cggggtgcaa tgggtacatt tggttacatg 1500gcaccagaga ctgtttatgg agaagtgtct
gcaaaagtag atgtatatgc atttggagtt 1560gtcctttacg aattgatttc tgcgaaaggt
gcggttgtca aaatgacaga agccgttggt 1620gaatttagag gccttgttgg tgtgttcgaa
gaatcattca aggaaaccga caaagaagaa 1680gcactacgca agattataga cccgaggctc
ggtgatagtt acccgtttga ttcggtatac 1740aagatggcgg aattagggaa agcatgtaca
caagagaatg cgcagctacg tccgagtatg 1800agatacattg tggttgcttt atccactctc
ttttcgtcta ccggaaattg ggatgttgga 1860aacttccaaa acgaagattt agtcagtctt
atgtccggcc ggtagactcg ttttccggtt 1920tgctgttgtt atatagaaat gattgttttt
tggtatgctc acgtatattt tgtctgtcta 1980tacgaacttt acatgtcaca actcacaagt
tgacttttat atgattttct ataacgctct 2040atatgtcaat tattcaacgt ctc
206341979DNAArabidopsis thaliana
4tctagaagtc agacatggct gtttcagtta gtaagcagta catgacaagt cttgtagtga
60tcttgctctt catctcctta tcatctctct ctccaacttc aacttcacac agctgtgacc
120ctgtcgaaga agaagaagaa gcctcctcct tcggttacgt ttgccactca aacctccaaa
180agtgccacac tttcgccatt ctcagagcca aacctccctt ctattccctc tcggacctga
240gtcgccatct tggtctcgac gcagacgacg agtatgttcc gaaaggtcag ttactcttga
300tcccaataga gtgcaggtgt aatggaagca tctacgaggc cagtctcatt aaaaattgcg
360ttaaaggaga cacctttcgc tctgtctctc agtctctgca aggcttgacc acctgcctat
420ctatcagaga gaagaatcca catatttccg aagacaagct tggcgacaac attaaactgc
480gtttggcgat caggtgctct tgtccacaag aaggcgtttc aaacgctagt tttctcgtta
540cgtatccggt gggcgtccgc gacagcgttt caagcctggc ggttaggttt aacacaacag
600aggatgccat tgtctctgca aacaacaaat ctggtgtggt tccactcaag cctgctctca
660tccctcttga tcacaaacca gagaaacaag gatcccggaa aagaaatcca tccaagaaga
720aacggtcaaa gatgaagctc atgatcgctg tgagcagcgc gattgctgga gtttgcggtc
780ttgtcactct catggtgttt ggttacttac actggaagaa agagacgcag attcaaacgc
840aaacgcaaaa gtggattagc aacaaagacc ccgagacgcg gcagctgagt ctgagcatcc
900gaaccacgag tgacaagaaa atctcgtttg aagggtcgca ggacggttcc atcttggatt
960ctcacaacac cgtcggcacc acaacgcccc gaaaacccgt tctggagatt tacgcgtttg
1020aagagttaga gaaggccacg gagaatttca gttcaagcaa tcacatcaaa ggttcggttt
1080atttcggttc gctcaaaggc aaagacttgg ctataaagca agtcaatgca gatgaaatga
1140aaagattcga tttcgggctt ctcaatgatc agtcacacta ctataaccac aatgtgatta
1200gggttcttgg aacatgtttt agagaaatcg atcaagattc ttatctggtt ttcgagtatg
1260caaggaatgg gtccctgtgg gattggattc agaacaaatt ggccatcaag aatcaattca
1320tcgagtcttg ttactgtttc ttggcatgga aacagaggat caagatatgt cacgatgtcg
1380ccatcgcgtt aaagtatatg catcggatca actacgtcca cggcaacatc aagagcagaa
1440acatcttctt gaacgaagat ctcagaggta aagtcggaaa ctttgggatg tcaaagtgcg
1500taaccaatga attagccaca gaagagaacc ttatagagag ttcactgtct ccagcttctg
1560acatatttgc ttacgggata atcgtaatgg aggttttgtc tggacaaacc ccagatatgt
1620tgcttggatt gcaagaagta gagacaacat cccttgggac acaagaaact tttgtttccg
1680aatggagtag attaagaagg cttctcgggg acaaggaaaa gctgagggaa gtgatggaca
1740gcacattggg agagagctat tcggttgatt ctgcgtttga aatcgcaagt attgcaagag
1800attgtactgc agaagaagcc gagtcaaggc cgagcgcggt tgagattgca gagagggttt
1860caagattggt ggatgacgat gaagatgaag aagatgaggc agtaatagat agagagagta
1920ccttaatttc agagagttca tacaagcctt tggtaaagaa gagtagtata atagattaa
197952321DNAArabidopsis thaliana 5gctgatttca tctcttctct acttcttcct
cttctccatt aaccaaatca accaataccc 60atttctcaat tcaccatatc ttcatcaaat
caagattctt tcttccttaa tccctaaaaa 120aaaccttctt tcaattctga ttttcactta
ggtttttctc aatttctact tcacacaatg 180aatcttacct tctacatctt cttcttaagt
cttttaccaa gtttctcatc ttcaaagcca 240atgaattgct ctgacactac tcgtctctgc
agttccttcc ttgctttcaa accaaaccaa 300aaccaatctt tctctgtaat ccaaagcatg
ttcgatgtat taccacaaga cataaccgcc 360gacatctccg gaggttactt ctttattaaa
aagaactgtt cttgtcttac tacgactcat 420caatacacaa caaacacaac attcacaatc
agacaaaacg ttggatacgt ttacaatgtt 480accgtctctg cttactctgg tctcgcgttt
ccaccgaata caactagagc agctcgcgct 540ggtgcagtgg tttctgttca gcttctttgt
ggttgctcta gtggtctctg gaactatctt 600atgagctacg ttgctatggc tggagatagt
gtacaatctc tctcgagcag atttggtgtt 660agtatggatc gaattgagga tgttaatggg
atcttgaatc ttgataatat cacagctggt 720gatctccttt atatcccact tgattccgta
cctggagagc cttatgagac aagtaagata 780aatccaccag ctccttctcc tgcgcctgca
tcatcattag ctaatggcaa tatctcagat 840gaccaggtga atcacactgc aaagagtggt
agtcatgtgc cttatatatg gattgttggt 900ggtcttggag ttgtgcttgc acttcttgtt
ctgtgcatac ttgtatgtat ctgtttgaga 960tcatctagtt gcagttccag tgaggaagat
ggtaatggac acaacttcca aatccttaga 1020aaatccggtt tcttttgcgg ttctggtcgg
tataactgct gtagatcagg ggattttaga 1080caaaccaatg gtgaaactca agttgttgct
attcctaaag ctcttggtga tggaatgttt 1140gagatagaga agcctatggt gtttacttat
gaagaaattc gtgcggctac ggatgagttt 1200tcggattcta atcttcttgg tcatggaaac
tatggttctg tatactttgg tctacttaga 1260gaacaggaag ttgctgtcaa aaggatgact
gctacaaaaa ctaaagagtt tgcagcagag 1320atgaaagtgc tttgcaaagt tcatcattcc
aatctggtag aattgattgg ttacgctgca 1380accgttgacg agcttttcgt agtttatgaa
tatgtccgaa agggaatgct gaaaagccat 1440ttgcatgatc ctcagagcaa aggaaatacc
ccactttctt ggataatgag gaatcagatt 1500gcacttgacg cagcaagagg cttggaatat
attcatgagc atactaaaac tcattatgtt 1560caccgagata tcaagacaag caatattttg
ctcgacgagg cattcagggc taagatatcg 1620gattttggac ttgcaaaact cgttgagaaa
actggagagg gagaaatatc agtaactaaa 1680gtagttggta catatggtta tcttgcacca
gaatatctaa gcgacggtct tgccacctcg 1740aaaagtgata tttacgcctt tggtgttgtt
cttttcgaaa ttatatccgg aagagaagct 1800gttataagaa cggaggcgat tggaactaag
aatccagaaa gacgtccatt ggcatctatt 1860atgttagcag tgctaaagaa ctcaccagac
tctatgaaca tgtcgagttt gaaggagttt 1920gttgacccca acatgatgga tttatacccg
catgactgtt tgttcaagat tgctacgttg 1980gcaaagcaat gtgtggatga tgatccgatt
ctaagaccaa acatgaagca agtggttata 2040tcgctctcgc agatactcct gtcttccatt
gaatgggaag ccactcttgc tggaaacagc 2100caagtcttta gtggtctagt ccaaggaaga
taaaaacaat aaaaacttag aaccctttct 2160ttattttcaa tttatgaatg tcgccattgc
ctagaagaag cttagtttgt ctaaaccaaa 2220agatcgttcg cagtctaact aagtagatag
ttacagagac aaatctctat atgattattc 2280ttttggagaa tcaaatgaag agatgtatga
gtttatatgc t 232161839DNAArabidopsis thaliana
6atgatctcgt tttcatttca tctcctcgtc ttcatcctcc taagtctctc ttccttcgca
60acagcacaac agccttatgt cggaatatca acaactgatt gctctgtttc cgacaacaca
120acctctgttt tcggctactc ttgtaatggt ctcaacaaaa cttgccaagc ttacgtcatt
180ttccgatcta ctccttcttt ctccaccgtc acctcaatct cttccctctt ctctgtcgac
240ccatctctcg tctcctccct caatgacgct tcaccttcaa cttctttccc ttctggtcaa
300caagtcatta tccctttaac ttgttcttgc accggtgacg attcacaatc caatattaca
360tacacaatcc aaccaaacga ttcgtatttc gccattgcta acgacactct tcaggggctt
420tcgacatgtc aagctttggc aaaacagaac aatgtttctt ctcaatcttt gtttcctggt
480atgaggatcg ttgttccgat ccgttgtgcg tgtcccacgg ctaaacagat caacgaagat
540ggcgtgaagt atctgatgag ttatactgtg gtttttgaag acactatcgc gatcatcagc
600gatagattcg gggttgagac gagcaagact ttaaaagcta acgaaatgtc ttttgagaat
660tctgaagttt tccctttcac aacaatcctg atccctttgg tgaatcctcc tgcaaatact
720aactctctca ttcctcctcc tcctcctcct cctccacaat cggtttctcc tcctccattg
780tcgccggatg gtaggaaatc gaagaagaaa acatgggtct atgctcttgc cggagtacta
840ggaggagcat tggttttgag tgtgatcggt gcagccatat tttgcctgtc taagaagaaa
900acgaaaacgc aaacgcaaga ggaaacggga aacctcgata gcttcatggg caagaaacca
960ccgatgtctg atcaagaatt cgatccgcta gatggtttgt caggtatggt ggtagaatcc
1020ttaaaagttt acaaatttca cgagctgcaa tcagctacaa gcgatttcac atcgtctagc
1080tcgattggag gatcagggta catcggaaaa atcaacggcg atggcgcaat gatcaagaaa
1140atcgaaggaa acgcatcgga ggaagtcaac ttattgtcta aactaaacca tcttaacatc
1200atccgtctct ctggattctg tttccatgaa ggagattggt acttagtcta cgaacacgct
1260tcaaacggaa gcttaagtga atggatccac acgacgaaat cgttactaag cttgacacaa
1320aaactacaaa tcgctttgga tatagcaaca gggctaaact atctacacaa cttcgctgat
1380cctccatacg ttcacagaga tttgaacagc aacaatgtgt tcctcgacct cgagtttcga
1440gcaaagattg gtagtttagg ttcagcaaga tcaacgactg aagatttcgt gttgaccaag
1500cacgtggaag gaacaagagg gtacttagct ccggagtatt tggaacatgg tttagtttca
1560actaagcttg atgtctatgc ttttggagtt gtcttgttgg agattgttac tggtaaagaa
1620gcttctgaat tgaagaaaga gatcgatgaa ggtaaagcta ttgatgagat tttgattcat
1680ggaagattgt tacctgaagg attaacaagt tttgttgaga gattagtggt ggattgttta
1740aagaaagatc atttgaatcg tccatcgatg gatgaaaatg ttatgtcttt gtctaagatc
1800ttggcggcta cgcagaactg ggaagaatcg tcgtactaa
183972239DNAArabidopsis thaliana 7gatcagaaca ccacaaaaca tggctgcgtg
tacactccac gcgctctcag tcaccttatt 60cctccttctc ttctttgccg tgtcaccggc
gaaagctcag caaccgtacg tcaacaacca 120ccagctcgcc tgcgaggtcc gtgtctacga
caacataacc aacggattca catgtaacgg 180cccaccttct tgccgctcat acctcacttt
ctggtctcaa ccaccgtaca acacggcgga 240ctcaatcgcc aaactcctca acgtctccgc
cgcagagatc caatcaatca acaacctccc 300cacagccacc accagaatcc caacccgtga
attagtcgtg atcccagcta actgctcctg 360ctcctcctcc agcggaggat tttaccaaca
caacgccact tacaatctct ccggtaacag 420aggagatgaa acctatttct cagtggctaa
cgatacttac caagctttat ccacgtgtca 480agccatgatg tcacaaaacc gttacggcga
gagacaacta acccccggct taaacctcct 540tgttcctctc cgatgtgctt gtcccaccgc
caaacaaacc accgccggat ttaaatatct 600cctgacttac ttagtcgcca tgggagatag
tatctccggc atcgccgaga tgttcaacag 660cacatccgcc gccataaccg aaggtaacga
gcttacatca gacaatatct tcttcttcac 720accggttcta gttcctctca caactgaacc
taccaaaatc gttatatctc cgtcgcctcc 780tcctccaccc gttgttgcta cgccgcctca
aacgccagtt gatcctccgg gatcttcttc 840ttctcacaaa tggatctaca tcggaattgg
aatcggagct ggtttgcttc tcttactctc 900aatcttagct ctctgcttct acaaacgaag
gtctaagaag aagtcattac cgtcgtcgtt 960gccggaggag aacaagctct ttgattcatc
aaccaaacaa tctattccca caacaacaac 1020gactcaatgg tcaatagatt tatccaattc
atcagaagct ttcggtttaa aatccgccat 1080agaatctcta acactataca gattcaacga
tcttcaatca gctacttcga atttcagcga 1140cgaaaacaga atcaaaggct ctgtttatcg
cgcgacaatc aacggcgacg atgccgctgt 1200gaaagtgatc aaaggagatg tttcttcctc
tgagatcaat cttttgaaga agctaaatca 1260ttctaatatc atccgtctct caggtttctg
tatccgtgaa ggaacatcgt acctcgtctt 1320cgagtattca gagaatggat cgatcagtga
ttggcttcac tcgtcgggca agaagagttt 1380gacatggaaa cagagagttg aaatagcaag
ggatgtggca gaggcgttag attatctcca 1440taactatata actccacctc atattcataa
gaacttggaa tcaacaaaca tacttctgga 1500ttctaatttc agagccaaga ttgcgaattt
cggtgttgcg aggattcttg atgaaggtga 1560tcttgatctt cagttaacaa gacatgttga
aggaacacaa ggctacttag ctccagagta 1620tgtggagaat ggagtcatta cttcgaaact
agacgtgttt gcttttggag ttgcggttct 1680tgagcttctt tcggggagag aagcagtaac
gatacataag aagaaggaag gagaagaaga 1740agtggagatg ttgtgtaaag tgataaacag
tgtgcttgga ggagagaatg tgagagagaa 1800gttaaaagag tttatggatc catctctagg
gaatgagtat ccgttggagc tggcttacac 1860catggctcag cttgctaaga gctgtgtcgc
aactgatctt aactcgcgtc catctgtcac 1920tcaggttcta accacgctct caatgatcgt
ctcctcctcc atcgattggg agccttctga 1980tgaccttctt cgttccggct ctcttggcaa
ctagttcagt tttttttttc tattggtagt 2040caataacaag tatttagact ttagataagg
accatacggc taaaaagttg agatctaata 2100agtaccccac tgtgaaacat agtaataaga
acatatcata gagaaacttt atttagattt 2160tgacatgttg cttcagtagt cttttgtatt
tcagaaaaaa ataatcattt tcttccaatg 2220agaagtagaa tcttgagga
223981159DNAArabidopsis thaliana
8atgaacaccg gcggtggcca atacacggat tttccggcgg tggaaactca cggaggacag
60ttcataagtt acgatatctt cggtagttta ttcgagatca catctaagta tcgtcctccg
120attataccaa ttggtcgtgg agcttatgga atcgtttgct ctgtgttgga tacggagacg
180aacgagctag tagcgatgaa gaagatagct aatgcttttg ataatcatat ggatgctaag
240cgtacgcttc gtgagatcaa gcttcttcgt catcttgatc atgaaaacat tatagctata
300agagatgttg ttccaccacc actaagaaga cagttcagtg atgtttatat ctctactgaa
360ttaatggata ctgatcttca tcaaatcatc agatctaacc agagtttatc agaagaacac
420tgtcagtact tcttgtacca gctacttcga ggactgaagt atatccactc agctaacatt
480attcataggg atttaaagcc gagcaatctt ctgttgaacg cgaattgcga tttaaagatt
540tgtgatttcg gtcttgctag acctacttca gagaatgatt ttatgactga gtatgttgtt
600acgagatggt atagagcacc tgagcttctg ttgaactctt cagattacac agctgctatt
660gatgtttggt ctgttggttg tatctttatg gagcttatga atagaaagcc tttgttccct
720ggtaaagacc atgttcatca aatgcgctta ttgacagagt tgcttggcac accgacagaa
780tctgatctcg gttttactca caatgaggat gcgaaaagat acatccggca acttcccaac
840ttcccacgtc agcccttagc taaacttttc tctcatgtta acccaatggc cattgatctt
900gttgacagaa tgttgacgtt tgaccccaac agaagaatca ctgttgaaca agctctgaat
960caccagtacc ttgctaaatt gcacgacccg aatgatgagc caatctgtca aaagccattc
1020tcttttgagt tcgaacaaca gcctctggat gaggaacaga taaaagagat gatctaccaa
1080gaagccatag cactcaatcc aacatacggt tagaagtgca gcagccccgt gaatgcctgg
1140tattacccaa taaccatcc
115991219DNAArabidopsis thaliana 9tcagcgctct cttttgtctc ctctgctttt
ttctcttctc tcctcagaga ttcgaagctt 60tttgtctccc ctgagtaacc aaattcaatg
gccgacgatt gggatctcca cgccgtagtc 120agaggctgct cagccgtaag ctcatcagct
actaccaccg tatattcccc cggcgtttca 180tctcacacaa accctatatt caccgtcgga
cgacaaagta atgccgtctc cttcggagag 240attcgagatc tctacacacc gttcacacaa
gaatctgtcg tctcttcgtt ttcttgtata 300aactacccag aagaacctag aaagccacag
aaccagaaac gtcctctttc tctctctgct 360tcttccggta gcgtcactag caaacccagt
ggctccaata cctctagatc taaaagaaga 420aagatacagc ataagaaagt gtgccatgta
gcagcagaag ctttaaactc cgatgtctgg 480gcatggcgaa agtacggaca gaaacccatc
aaaggttcac catatccaag aggatactac 540agatgtagta catcaaaagg ttgtttagcc
cgtaaacaag tggagcgaaa tagatccgac 600ccgaagatgt ttatcgtcac ttacacggcg
gagcataatc atccagctcc gacacaccgt 660aattctctcg ccggaagcac acgtcagaaa
ccatccgatc aacagacgag taaatctccg 720acgaccacta ttgctactta ttcatcgtct
ccggtgactt cagccgacga atttgttttg 780cctgttgagg atcatctagc ggtgggagat
cttgacggag aagaagatct gttatctttg 840tcggatacgg tggttagcga tgatttcttc
gatgggttag aggaattcgc agccggagat 900agcttttccg ggaactcggc tccggcgagt
tttgatctct cttgggttgt gaacagtgcc 960gccactacca ccggaggaat atgattagat
tacgacggct tagaatactc ttattaggac 1020agatttatag gattaaggaa ttattctcgg
agcatatgta aaaataggat aaaagaaaat 1080gttctttgtt actttttttc gggttttctt
cctattgttt ctaaacatct tagaaaaaat 1140ttaattgtat attccttaag ctcgatacat
cttgtttaac tcgagtacga tttacttatg 1200gagttatggt ctttgtctc
121910927DNAArabidopsis thaliana
10ggcttatatg gacgaaggag acctagaagc aatagtcaga ggctactccg gctccggaga
60cgcgttttcc ggcgaaagtt ccggtacgtt ttcaccttcg ttttgcctac cgatggagac
120gtctagtttc tacgaaccgg agatggagac aagtggctta gatgagctcg gtgaacttta
180caaacccttt taccctttct ccacacaaac gatcctcaca agctcggtct ctctccctga
240agattcaaaa cctttccgag atgacaagaa acaacgatca catggttgtc ttttatccaa
300cggatcaaga gctgatcata tccgaatttc agaatccaaa tcaaagaaaa gcaagaagaa
360tcaacagaag agagttgttg agcaagtgaa agaagagaat ctgttgtcgg acgcatgggc
420gtggcgtaaa tacgggcaga aacccatcaa aggatctcca tacccaagga gttattacag
480atgcagtagc tcaaaagggt gtttggcaag aaaacaagtc gaaagaaatc ctcaaaaccc
540ggagaaattc accataacat acactaatga gcacaatcat gaactaccaa cccggagaaa
600ctcattagcc ggttcgactc gagcaaaaac ttcccaaccc aaaccaacct taaccaaaaa
660atccgaaaaa gaagttgttt cttcccctac aagtaatcct atgatcccat ccgctgatga
720atcttctgtt gcggttcaag aaatgagcgt tgcggaaacg agtacgcacc aagcggctgg
780agcaatcgag ggccgccgct tgagtaacgg tttaccatcg gatttgatgt ccgggagcgg
840aacttttcca agttttaccg gtgacttcga tgaactattg aatagccaag agttcttcag
900tgggtattta tggaattact agagagc
927111804DNAArabidopsis thaliana 11aaacttccat ttttcgtatg gctgcttctt
ttcttacaat ggacaatagc agaaccagac 60aaaacatgaa tggttctgct aattggtcac
aacaatccgg aagaacatct acttcctctt 120tggaagatct tgagatacca aagttcagat
cttttgctcc ttcttcaatc tctatctctc 180cttctcttgt ctctccttcc acttgtttca
gtccctctct ttttctcgat tcccctgctt 240ttgtctcctc ctctgctaac gttctagctt
ctccaaccac aggagcttta atcacaaacg 300taactaacca gaaaggtata aatgaaggag
acaagagcaa caacaacaac tttaacttat 360tcgatttctc attccacaca caatcatcag
gagtttctgc tccgaccaca actacaacta 420caactacaac tacaacaaca acaaacagtt
ctatctttca atctcaggaa caacagaaga 480agaaccagtc agaacaatgg agccaaaccg
agactcgtcc aaacaatcaa gctgtatctt 540acaatggaag agagcaaagg aaaggagagg
atggttacaa ttggagaaag tacggacaaa 600aacaggtgaa aggaagtgag aatcctcgga
gttactataa gtgtactttc cctaattgtc 660caacgaagaa gaaagtggag agatctttgg
aaggtcagat cacagagatt gtgtataaag 720gaagccacaa ccatcctaaa cctcagtcta
ctagaagatc ttcttcgtct tcttcgactt 780ttcattcagc tgtgtacaat gccagtttgg
atcataatcg tcaagcttct tctgatcagc 840ctaattccaa taatagcttt catcagtctg
attcctttgg gatgcaacaa gaggataata 900ctacttctga ttctgttggt gacgatgagt
tcgaacaagg ctcatcgatt gtcagcagag 960acgaagaaga ttgtgggagt gaacctgaag
caaagagatg gaaaggggac aatgaaacaa 1020atggtgggaa tggtggtgga agcaagacag
tgagagagcc gagaatcgta gtgcagacaa 1080cgagtgatat tgacattctt gacgacggtt
acagatggag aaaatacggc cagaaagtcg 1140ttaagggaaa cccaaatcca agaagctact
acaagtgcac aaccatcggt tgtccagtga 1200ggaaacatgt ggagagagca tcacacgaca
tgagagcagt aatcacaacc tacgaaggga 1260aacacaacca cgatgttcct gcagctcgtg
gtagcggtta cgccacaaac agagcaccac 1320aggattcgtc ttcagtcccg attagaccag
ctgctattgc tggtcactcc aactacacta 1380cttcttctca agcaccatat acacttcaga
tgctgcacaa caacaacact aataccgggc 1440cttttggtta cgccatgaac aacaataaca
acaacagcaa ccttcaaacg caacaaaact 1500ttgttggtgg tggattctct agagcaaagg
aagaaccaaa cgaggagacc tcatttttcg 1560attcgtttat gccctgaaga aaaaaggaac
cagttgttcc ttttttatgt tccttttgta 1620catttctgcc accaaaggat tttactactt
actagttatc ctgcaggata gtaagtcaga 1680cttctatagt ccatagagaa attttttcat
ttgttttttc acacgcctgt aatatgttta 1740atgtttgtac tttgtaccat agaactagaa
cacggaataa aaccaatcaa ttttcagttt 1800cttc
1804121514DNAArabidopsis thaliana
12aatcctataa cactctcatt ctcatcatat cattcttcaa tctatataac ccattcttaa
60ttatactcaa cacacattat atttttctga tcatatcatt ctttcagtcc atctatataa
120ccaattcttg atttatactt aaaacacaca ttatacatct ttctcatcat agtttgtatc
180aatttcctag agtaaactac ctaaaggaaa aaaaaaatct attttgggaa tcatatacta
240aaaatggaag gaagagatat gttaagttgg gagcaaaaga cattgctaag cgagcttatc
300aatggatttg atgcggccaa aaagcttcag gcacgactta gagaagctcc gtcgccgtcg
360tcatcatttt catcaccggc gacggctgtt gctgagacta acgagattct ggtgaagcag
420atagtttctt cctacgagag atctcttctt ctgctaaact ggtcatcctc accgagcgta
480caacttattc cgacgccggt tactgtagtc ccggtggcaa atcccggcag tgttccagaa
540tctccggcat cgataaacgg aagtccgaga agtgaagagt ttgccgatgg aggaggttct
600agcgagagtc atcatcgcca agattacatt ttcaattcaa agaaaagaaa gatgttacca
660aagtggtcag aaaaagtgag aataagccca gagagaggct tagaaggacc tcaagatgat
720gtctttagct ggagaaaata tggtcaaaaa gacattttag gcgccaaatt cccaaggagt
780tattacagat gcacacatcg tagcacacaa aactgttggg caacgaaaca agtccagaga
840tcagacgggg atgctacggt tttcgaagtg acgtacagag gaacacacac ttgttcgcag
900gcgatcacaa gaacaccacc attagcctcg ccggagaagc gacaagacac cagagtcaaa
960ccagccatta cccaaaagcc aaaggatatt ctcgagagtc ttaaatccaa cttaaccgtt
1020cgaaccgatg ggcttgatga tggtaaagac gttttctcgt tccctgatac gccgccgttt
1080tacaattacg gaactatcaa cggcgagttc ggccacgtgg agagttctcc gatcttcgac
1140gttgttgact ggttcaatcc aacggtcgag attgacacaa ctttccccgc gtttttacac
1200gagtcgattt attattaatt aaaatttgta acagagaaat agatagtaac tagtaagtaa
1260tgatcagcga gagttaaaac ataaaagtac ttagagtaat ctaacgatgc ataataagga
1320atgttcaaca ggacttgaac atgatttcaa tactaagaga gatttatcta gctactggta
1380gtagccgcag acttcttgtt gtaagcttcc acttcctttt tgtatgcttc catcagtttt
1440gcggcctttc cgttgtaaac ttgcttctct tcttcactca attcctgcaa agacgtattc
1500gcacagatag atta
15141322DNAArtificialPCR primer 13ctcacggagg acagttcata ag
221422DNAArtificialPCR primer 14gagatcagat
tctgtcggtg tg
221523DNAArtificialPCR primer 15gtaagctcat cagctactac cac
231622DNAArtificialPCR primer 16accgctagat
gatcctcaac ag
221722DNAArtificialPCR primer 17atggacgaag gagacctaga ag
221822DNAArtificialPCR Primer 18ccgcttggtg
cgtactcgtt tc
221922DNAArtificialPCR primer 19ctccgaccac aactacaact ac
222022DNAArtificialPCR primer 20ggctctctca
ctgtcttgct tc
222123DNAArtificialPCR primer 21cctacgagag atctcttctt ctg
232222DNAArtificialPCR primer 22agatcggaga
actctccacg tg
222330DNAArtificialPCR primer 23gactaagaga gaaagtaaga gataatccag
302430DNAArtificialPCR primer 24cagcctttga
tttcaatttg catgtaagag
302529DNAArtificialPCR primer 1 25cacccttgcg tagagagttt actgatgtc
292629DNAArtificialPCR primer 26gttgacgagg
atattgagga agttgtctg
292729DNAArtificialPCR primer 27tcaccttgct ggttctggtt ctggttctg
292829DNAArtificialPCR primer 28tctgataggg
gtgcaacccc atcttcttc
292929DNAArtificialPCR primer 29agttgtggtt cagaccacca gtgacattg
293029DNAArtificialPCR primer 30accccattga
gtttccaaac cctgatgag
293129DNAArtificialPCR primer 31cccatcaaaa gaaccaacca caacaagag
293229DNAArtificialPCR primer 32atccgcacgc
acttgaacca tgtattgtg
293329DNAArtificialPCR primer 33aaggttcgta aacgatggct gatgctgag
293429DNAArtificialPCR primer 34accttgatct
tcatgctgct aggagcaag
2935760DNAArabidopsis thaliana 35ataacacaac aataaccatt atcaacttag
aaaaatgaat tttactggct attctcgatt 60tttaatcgtc tttgtagctc ttgtaggtgc
tcttgttctt ccctcgaaag ctcaagatag 120cccacaagat tatctaaggg ttcacaacca
ggcacgagga gcggtaggcg taggtcccat 180gcagtgggac gagagggttg cagcctatgc
tcggagctac gcagaacaac taagaggcaa 240ctgcagactc atacactctg gtgggcctta
cggggaaaac ttagcctggg gtagcggtga 300cttgtctggc gtctccgccg tgaacatgtg
ggttagcgag aaggctaact acaactacgc 360tgcgaacacg tgcaatggag tttgtggtca
ctacactcaa gttgtttgga gaaagtcagt 420gagactcgga tgtgccaaag tgaggtgtaa
caatggtgga accataatca gttgcaacta 480tgatcctcgt gggaattatg tgaacgagaa
gccatactaa tgaagtaatg atgtgatcat 540gcatacacac gtacataaag gacgtgtata
tgtatcagta tttcaataag gagcatcata 600tgcaggatgt atcaatattt atcaaataat
acaaataaga gctgagatta cgagaatcta 660tttaaattaa aagttacata cttaattatt
atagttatat atgtaaaata tgtggccttt 720ttaaaagtta cataattaat tattatagtt
aatgtctttc 7603628DNAArtificialPCR primer
36aacacgtgca atggagtttg tggtcact
283728DNAArtificialPCR primer 2 37accattgtta cacctcactt tggcacat
2838438DNAArabidopsis thaliana 38acacaacaca
tacatctata cattgaaaac aaaatagtaa taatcatcat ggctaagttt 60gcttccatca
tcacccttat cttcgctgct cttgttctct ttgctgcttt cgacgcaccg 120gcaatggtgg
aagcacagaa gttgtgcgag aagccaagtg ggacatggtc aggggtttgc 180ggaaacagta
atgcatgcaa gaatcagtgc attaaccttg aaggagccaa acatggatca 240tgcaactatg
tcttcccagc acacaagtgt atctgttacg tcccatgtta aatctaccaa 300taatctttgg
tgctaaatcg tgtgtatttt acataaaata agtctctgtc actctatgag 360taactttatg
acatgcatat ttctgtttta atgtttattt tcccgttgtt gttacaataa 420tataaaaata
atttatgt
4383929DNAArtificialPCR primer 39agtgcattaa ccttgaagga gccaaacat
294029DNAArtificialPCR primer 40aacagataca
cttgtgtgct gggaagaca
294130DNAArtificialPCR primer 41tggccattga tcttgttgac agaatgttga
304229DNAArtificialPCR primer 42tcgtgcaatt
tagcaaggta ctggtgatt
294329DNAArtificialPCR primer 43tttaggcgcc aaattcccaa ggagttatt
294429DNAArtificialPCR primer 44tctggacttg
tttcgttgcc caacagttt
294528DNAArtificialPCR primer 45ggtattctta ccttgaagta tcctattg
284628DNAArtificialPCR primer 46ctcattgtag
aaagtgtgat gccagatc
284729DNAArtificialPCR primer 47agaatatatc cacgagcaca cggttccag
294829DNAArtificialPCR primer 48gacgaaaaga
gagtggataa agcaaccac
294925DNAArtificialPCR primer 49cccatttgga cgtgaatgta gacac
255028DNAArtificialPCR primer 50atgaagctaa
agatttctct aatcgctc
285127DNAArtificialPCR primer 51gaaatgcacc atttggatct cttccag
27521249DNAArabidopsis thaliana 52atatcatttt
tcacagaatc atagaaaaat caagaaaatg tctgaatcaa ggagcttagc 60ctcaccacca
atgttgatga ttcttctcag ccttgtaata gcttccttct tcaaccacac 120agctggacaa
atcggagtat gctacgggat gctaggcgat accttgccaa gtccatcgga 180cgttgtggct
ctttacaaac aacaaaacat ccagcgaatg cggctctacg gccctgaccc 240aggcgctctt
gccgctctcc gtggctctga catcgagctc atcctcgacg ttcccagttc 300agatcttgaa
cgtctcgcct ccagtcaaac ggaggccgac aagtgggttc aagaaaacgt 360tcagagctac
agagatggtg tcagattccg gtacatcaac gttggaaatg aggtgaaacc 420ctcagttggg
gggtttctct tacaagcaat gcagaacatc gagaacgcgg tttctggagc 480agggcttgaa
gtcaaggtct caacagctat agccactgac accaccactg atacgtctcc 540tccgtctcaa
ggaaggttca gggatgagta taagagcttt ctcgaaccag tgataggttt 600cttggcaagc
aagcaatctc ccttgctcgt gaatctctac ccttacttca gctacatggg 660agacacggcc
aacatccatc tagactacgc tctgttcacc gcccagtcca ctgttgataa 720cgatccaggg
tactcatacc aaaacctatt cgacgcaaat ctcgactcgg tttatgcagc 780attggagaaa
tcagggggcg gatcgttgga aatcgtggtg tcggagaccg gttggcccac 840agagggagca
gtcgggacga gtgtggaaaa cgcaaagact tatgttaaca atttgataca 900acatgtgaag
aatggatcac cgagaaggcc agggaaagct atagagactt atatattcgc 960tatgttcgat
gagaataaga aggaaccaac gtatgagaag ttttggggac tgtttcatcc 1020agatcgacag
tctaagtatg aagttaattt caactaatcc ttagagactt gtgggctttt 1080tatgtaagcg
tatttaaaaa ttgggaactt gttgtagtaa taaggaataa ttaatgcgct 1140ttcagcgtgt
agtatgttgt tatttttaag gttataaatg agctgcaagc ataaataagg 1200aaaaaaaata
gcatgggcct ataggcccaa taataaaaca agcttgctt
124953617PRTArabidopsis thaliana 53Met Lys Leu Lys Ile Ser Leu Ile Ala
Pro Ile Leu Leu Leu Phe Ser 1 5 10
15 Phe Phe Phe Ala Val Glu Ser Lys Cys Arg Thr Ser Cys Pro
Leu Ala 20 25 30
Leu Ala Ser Tyr Tyr Leu Glu Asn Gly Thr Thr Leu Ser Val Ile Asn
35 40 45 Gln Asn Leu Asn
Ser Ser Ile Ala Pro Tyr Asp Gln Ile Asn Phe Asp 50
55 60 Pro Ile Leu Arg Tyr Asn Ser Asn
Ile Lys Asp Lys Asp Arg Ile Gln 65 70
75 80 Met Gly Ser Arg Val Leu Val Pro Phe Pro Cys Glu
Cys Gln Pro Gly 85 90
95 Asp Phe Leu Gly His Asn Phe Ser Tyr Ser Val Arg Gln Glu Asp Thr
100 105 110 Tyr Glu Arg
Val Ala Ile Ser Asn Tyr Ala Asn Leu Thr Thr Met Glu 115
120 125 Ser Leu Gln Ala Arg Asn Pro Phe
Pro Ala Thr Asn Ile Pro Leu Ser 130 135
140 Ala Thr Leu Asn Val Leu Val Asn Cys Ser Cys Gly Asp
Glu Ser Val 145 150 155
160 Ser Lys Asp Phe Gly Leu Phe Val Thr Tyr Pro Leu Arg Pro Glu Asp
165 170 175 Ser Leu Ser Ser
Ile Ala Arg Ser Ser Gly Val Ser Ala Asp Ile Leu 180
185 190 Gln Arg Tyr Asn Pro Gly Val Asn Phe
Asn Ser Gly Asn Gly Ile Val 195 200
205 Tyr Val Pro Gly Arg Asp Pro Asn Gly Ala Phe Pro Pro Phe
Lys Ser 210 215 220
Ser Lys Gln Asp Gly Val Gly Ala Gly Val Ile Ala Gly Ile Val Ile 225
230 235 240 Gly Val Ile Val Ala
Leu Leu Leu Ile Leu Phe Ile Val Tyr Tyr Ala 245
250 255 Tyr Arg Lys Asn Lys Ser Lys Gly Asp Ser
Phe Ser Ser Ser Ile Pro 260 265
270 Leu Ser Thr Lys Ala Asp His Ala Ser Ser Thr Ser Leu Gln Ser
Gly 275 280 285 Gly
Leu Gly Gly Ala Gly Val Ser Pro Gly Ile Ala Ala Ile Ser Val 290
295 300 Asp Lys Ser Val Glu Phe
Ser Leu Glu Glu Leu Ala Lys Ala Thr Asp 305 310
315 320 Asn Phe Asn Leu Ser Phe Lys Ile Gly Gln Gly
Gly Phe Gly Ala Val 325 330
335 Tyr Tyr Ala Glu Leu Arg Gly Glu Lys Ala Ala Ile Lys Lys Met Asp
340 345 350 Met Glu
Ala Ser Lys Gln Phe Leu Ala Glu Leu Lys Val Leu Thr Arg 355
360 365 Val His His Val Asn Leu Val
Arg Leu Ile Gly Tyr Cys Val Glu Gly 370 375
380 Ser Leu Phe Leu Val Tyr Glu Tyr Val Glu Asn Gly
Asn Leu Gly Gln 385 390 395
400 His Leu His Gly Ser Gly Arg Glu Pro Leu Pro Trp Thr Lys Arg Val
405 410 415 Gln Ile Ala
Leu Asp Ser Ala Arg Gly Leu Glu Tyr Ile His Glu His 420
425 430 Thr Val Pro Val Tyr Val His Arg
Asp Ile Lys Ser Ala Asn Ile Leu 435 440
445 Ile Asp Gln Lys Phe Arg Ala Lys Val Ala Asp Phe Gly
Leu Thr Lys 450 455 460
Leu Thr Glu Val Gly Gly Ser Ala Thr Arg Gly Ala Met Gly Thr Phe 465
470 475 480 Gly Tyr Met Ala
Pro Glu Thr Val Tyr Gly Glu Val Ser Ala Lys Val 485
490 495 Asp Val Tyr Ala Phe Gly Val Val Leu
Tyr Glu Leu Ile Ser Ala Lys 500 505
510 Gly Ala Val Val Lys Met Thr Glu Ala Val Gly Glu Phe Arg
Gly Leu 515 520 525
Val Gly Val Phe Glu Glu Ser Phe Lys Glu Thr Asp Lys Glu Glu Ala 530
535 540 Leu Arg Lys Ile Ile
Asp Pro Arg Leu Gly Asp Ser Tyr Pro Phe Asp 545 550
555 560 Ser Val Tyr Lys Met Ala Glu Leu Gly Lys
Ala Cys Thr Gln Glu Asn 565 570
575 Ala Gln Leu Arg Pro Ser Met Arg Tyr Ile Val Val Ala Leu Ser
Thr 580 585 590 Leu
Phe Ser Ser Thr Gly Asn Trp Asp Val Gly Asn Phe Gln Asn Glu 595
600 605 Asp Leu Val Ser Leu Met
Ser Gly Arg 610 615 541872DNAGlycine max
54atggaactca aaaaagggtt acttgtgttc tttttgctgc tggagtgtgt ttgttacaat
60gtggaatcca agtgtgtgaa gggatgtgat gtagctttcg cttcctacta tgtcagtccg
120gatttaagct tagaaaatat agcgcggttg atggaatcaa gcattgaagt tataatcagc
180ttcaatgaag acaatatatc gaatggttat ccgctatcct tttacagact caatattcca
240ttcccctgtg actgtattgg tggtgagttt ctggggcatg tgtttgagta ctcagcttct
300gcaggtgaca cctatgattc gattgcgaaa gtgacatacg ccaatctcac caccgttgag
360cttttgcgga ggttcaatgg ctatgatcaa aatggtatac ctgcaaatgc cagggttaat
420gtcacggtca attgttcttg tgggaacagc caggtttcaa aagattatgg gatgtttatt
480acctatccac tcaggcctgg gaataatttg catgatattg ccaatgaggc tcgtcttgat
540gcacagttgc tgcagcgtta caatcctggt gtcaatttca gcaaagagag tgggactgtt
600ttcattccag gaagagatca acatggagac tatgttccct tgtacccgag aaaaacaggt
660cttgctaggg gtgctgcagt tggaatatct atagcaggaa tatgcagtct tctattatta
720gtaatttgct tatatggcaa gtacttccag aagaaggaag gagagaaaac taaattgcca
780acagaaaatt ctatggcgtt ttcaactcaa gatggtacgg tctctggaag tgcagaatat
840gaaacttcag gatccagtgg gactgctagt gctactggcc tcacaggcat tatggtggca
900aaatcaatgg agttctcata tcaagaacta gccaaggcta caaataactt tagcttggag
960aataaaattg gtcaaggtgg atttggagct gtctattatg cagaactgag aggcgagaaa
1020actgcaatta agaagatgga tgtgcaagca tcgacagaat ttctttgcga gttgaaggtc
1080ttaactcacg ttcatcactt taatctggta cagcatcctt caaacaaccc caagcatgtg
1140cgcttgattg gatattgtgt tgagggatcc cttttccttg tatatgaata tattgacaat
1200ggaaacttag gccaatattt gcatggtaca gggaaagatc ctttgccatg gtctggtcga
1260gtgcaaattg cgctagattc agcaagaggc cttgaatata ttcacgagca cactgtgcct
1320gtgtatatcc atcgtgatgt aaaatcagca aatatattaa tagacaagaa catccgtgga
1380aaggttgcag attttggctt gaccaaactt attgaagttg gaggctccac acttcacact
1440cgtcttgtgg gaacatttgg atacatgcca ccagaatatg ctcaatatgg tgacatttct
1500ccaaaagtag atgtatatgc ttttggagtg gttctttatg aacttatttc tgcaaagaat
1560gctgttctaa agacaggtga atctgttgct gaatcaaagg gccttgtagc tttgtttgaa
1620gaagcactta atcagagtaa tccttcagaa agtattcgca aactggtgga tcctaggctt
1680ggcgaaaact atccaattga ttcagttctc aagattgctc aacttgggag agcttgtaca
1740agagataacc cactactacg cccgagtatg aggtctatag ttgttgctct catgacactt
1800tcatcaccta ctgaggattg cgacacttcc tacgaaaatc agactctcat aaatctactg
1860tctgtgagat ga
1872551860DNAGlycine max 55atggaactca aaaaatggtt actgttcttt ttgttgctgg
agtatgtttg ttgcaatgcg 60gagtctaagt gtgtgaaggg atgtgatgta gctttagctt
catactatgt tagtccaggg 120tatttactct tggaaaatat aacgcgcttg atggaatcaa
ttgttctgtc caattctgat 180gttataatct acaacaaaga caaaatattc aatgaaaatg
tgctagcatt ttccagactc 240aatattccat tcccctgtgg ctgtatcgat ggtgagtttc
tggggcatgt gtttgagtac 300tcagcttctg caggtgacac ctatgattcg attgcgaaag
tgacatatgc caatctcacc 360actgttgagc ttttgcggag gttcaacagt tatgatcaaa
atggtatacc tgcaaatgcc 420acggttaatg tcacggtcaa ttgttcttgt gggaacagcc
aggtttcaaa agattatggg 480ctgtttatta cctatctact caggcctggg aataatttgc
atgatattgc caacgaggct 540cgccttgatg cacagttgct acagagttac aatcctggtg
tcaatttcag caaagagagt 600ggggatattg ttttcattcc aggaaaagat caacatggag
attatgttcc cttgtaccct 660agaaaaacag caggtcttgc tacgagtgct tcagttggaa
taccaatagc aggaatatgc 720gttcttctat tagtaatttg catatatgtc aagtacttcc
agaagaagga aggagagaaa 780gctaaattgg caacagaaaa ttctatggcg ttttcaactc
aagatgtctc tggaagtgca 840gaatatgaaa cttcaggatc cagtgggact gctagtacta
gtgcaactgg ccttacaggc 900attatggtgg caaaatcaat ggagttctca tatcaagaac
tagccaaggc tacaaataac 960ttcagcttgg agaataaaat tggtcaaggt ggatttggaa
ttgtctatta tgcagaactg 1020agaggcgaga aaactgcaat caagaagatg gacgtgcaag
catcaacaga atttctttgc 1080gagttgaagg tcttaactca tgttcatcac ttgaatctgg
tgcgcttgat tggatattgt 1140gttgaggggt ctcttttcct tgtatatgaa tatattgaca
atggaaactt aggccaatat 1200ttgcatggta cagggaaaga tcctttccta tggtctagcc
gagtgcaaat tgcactagat 1260tcagcaagag gccttgaata tattcacgag cacactgtgc
cagtgtatat ccatcgtgat 1320gtaaaatctg caaatatatt aatagacaaa aacttccgtg
gaaaggttgc agattttggt 1380ttgaccaaac ttattgaagt tggaggttcc acacttcaaa
ctcgtcttgt gggaacattt 1440ggatacatgc caccagaata tgctcaatat ggtgacattt
ctccaaaagt agatgtatat 1500gcttttggag ttgttcttta tgaacttatt tctgcaaaga
atgctgtcct aaagacagta 1560gaatctgttg ctgaatcaaa gggccttgta gctttgtttg
aagaagcact taatcagagt 1620aatccttcag aaagtattcg caaactggtg gatcctaggc
ttggagaaaa ctatccaatc 1680gattcagttc tcaagattgc tcaacttggg agagcttgta
caagagataa cccactacta 1740cgcccgagta tgaggtctat agttgttgct ctcttgacac
tttcatcacc tactgaggat 1800tgctatgatg acacttccta cgaaaatcag actctcataa
atctactgtc tgtgagatga 1860561803DNAGlycine max 56atggaacaca gtttcagatt
accagttttc ttcttgttat gtgcctctat agcgttcagt 60gcagaatcca agtgtagcag
gggttgtgat ttagctctag cttcctacta tctatcacaa 120ggtgacttga catatgtatc
aaagcttatg gaatctgagg ttgtttcaaa acctgaagat 180attctcagct acaacactga
caccataaca aacaaagacc tgttgcctgc ctctatcaga 240gtgaacgttc cattcccttg
tgactgcatt gatgaagagt ttcttggcca tacttttcaa 300tacaacctta caacaggaga
cacttatttg tccattgcca ctcagaacta ctctaatttg 360accactgctg agtggttgcg
gagcttcaac agatatttac cagctaatat tcctgatagt 420gggactctta atgtcaccat
taactgttcc tgtgggaata gtgaagtttc caaggattat 480ggattgttca tcacgtaccc
tcttagacct gaggattctt tgcagtcgat tgccaacgag 540actggcgttg atcgtgactt
gctggttaag tacaacccgg gtgtaaattt tagccaaggg 600agtggtctgg tttatattcc
aggaaaaggt cttgcgggtg gtgttattgc tggaatatct 660attggagtag taacaggact
tctgctattg gcattttgtg tgtatgttac atattaccga 720agaaagaagg tatggaagaa
ggatttgctc tcagaagaat ccaggaagaa ctctgctaga 780gttaagaatg tccctctttc
agatgaagcc tctggtgatt cggctgcaga aggtggtact 840aacaccattg gcattagggt
gaacaaatca gcagagtttt catatgagga actagccaat 900gccacaaata acttcagttt
ggctaataaa attggtcaag gtggttttgg ggtagtctat 960tatgcagagc tgaatggaga
gaaagctgca ataaaaaaga tggacataca agcaacaaga 1020gaatttcttg cggaattgaa
agtgttgaca catgttcatc acttgaacct ggtgcgcttg 1080attggatatt gtgttgaggg
ctcccttttt cttgtctatg agtacattga gaatggcaac 1140ttaggacaac atctacgtaa
atcaggtttc aatcctttgc catggtctac ccgagttcaa 1200attgctctgg attcagccag
aggtcttcaa tacattcatg agcatacggt acctgtatat 1260atccatcgtg acataaagtc
ggaaaacatt ttaatagaca aaaacttcgg tgcaaaggtt 1320gcagactttg gattaaccaa
gttgattgat gttggaagtt catcacttcc cactgttaat 1380atgaagggca catttggtta
catgccacca gaatatgcat atggcaatgt ttctcccaaa 1440atagatgtct atgcttttgg
agttgttctt tatgaactaa tttctggtaa agaagcattg 1500agcagaggtg gtgtctctgg
tgctgaacta aagggccttt ttgatgaagt atttgatcag 1560caagatacca cagaaggtct
taaaaaactg gtggatccta ggcttggaga taactaccca 1620attgattcag tttgcaagat
ggcacaactt gctagagcat gcacagagag cgatccacaa 1680caacgtccaa atatgagttc
tgttgtggtt actctcacag cacttacttc aactactgag 1740gattgggata ttgcttccat
cattgaaaat ccaactcttg caaatctaat gtctggtaaa 1800taa
1803571740DNAGlycine max
57atgaccacac acccaactac aaaatcaaaa ccaccccatg ttttcttcct tctcttaatt
60cagttattga tttcgattac aagagtcaaa ggtagttgcg ttactggctg caaccttgct
120ttagcttcct attacttagg gaacggcaca aacctcactt acattagcaa cctcttcgga
180aggccaacat cggagattct aaaatacaac ccaagtgtta agaacccaaa cgtgattctt
240agccaaacac ggatcaacgt gcccttttcg tgtgactgct tgaacggtgc gtttttgggg
300cacactttct cgtacgcgat ccaacacgga aacacgtaca agatagttgc cgaggttgat
360ttctcgaacc tcacgacgga ggattgggtt ggcagggtca acagttaccc gccgaatcag
420atacccgaca acgtgaacat caacgtcacc gttaactgtt cctgcggaaa ccgacacgtg
480tccaaggatt atgggctgtt catgacgtac ccgcttcggg tcggtgacag cttacagcgg
540gttgcggccg aagccggcgt gccggcggag ctgctgctga ggtacaaccc cacggcggat
600ttcggtgctg ggaatggact cgtgtttgtg ccggctaaag ttgaagacca gtgcacattg
660cacctctgga tgaccctagg gccagccctg gttgtcttct ggaaatctgg tgggatgatt
720tgtaagggtt gggggcactc cttgcaccct tatcctatta tgtatttcca ttttcattta
780ctatacacaa atggtccaag cacaaagact caacatatga aaaatggaaa ctttccacca
840atgcagttga gatcaggaat atctagtgga gccattgctg gcatagctgt tggaggagct
900gttggggtct taattttggc acttcttcta tatgttggac tacgcagaag aaggaaagtg
960gctgaggtat cccttctccc agtcccagga gcatctgaag atcagtgtag tccactccaa
1020ctccatcatg gtattggttg tggaagcagc ttggacaagg catctgaatc ttctgttgta
1080gcctctccaa ggttgacagg gattacagtg gacaagtcag tggagttccc atatgaggag
1140ctagacaagg ctactgatgg ctttagtgcg gctaatataa ttggtcgagg tggctttgga
1200tctgtttact atgctgagct ccggaatgag gtgaggttaa taggatattg tgttgagggc
1260tccttatttt tggtttatga gtacattgag aacggcaatt taagtcaaca tttgcgaggc
1320tcagggaggg atccgttaac atgggcagct cgagttcaaa ttgcccttga tgcagcaaga
1380ggattggaat acatccatga gcacactgtt cctgtctaca tccatagaga tattaaatca
1440gcaaacattt tgatagacaa aaactttcgt gcaaagtttg aagaagttct tggtctgtca
1500gatccaaaag tagatcttcg tcaacttatt gaccctacac ttggtgacaa ctaccctctt
1560gactcggtat ttaaggtgtc tcagcttgcc aaagcatgta cacatgagaa ccctcaactt
1620aggccaagca tgagatcaat tgtagttgcc ttaatgacac tttcatctgc aactgaggat
1680tgggatgttg gctccttcta tgaaaaccaa gctttggtgc acctaatgtc tggaaggtag
1740581902DNAGlycine max 58atggatctct ttcccttcat ccccatcatc attttcacac
tattgatcca caacttctct 60ctgattctgg ggcagcaacc ttacattggt ttaggcacag
tagcgtgccc aagaaggggt 120aacaaaaatt ctatccgtgg ttacacttgc aatggtgcaa
accatagctg ccaaagttac 180ctcaccttca gatctcaacc catctacaac tctgtcaaga
caatatcaac tttgttgggt 240tctgacccat cccagcttgc taaaataaac tcagtttcca
tgaatgacac ctttgaaaca 300aacaagttgg ttattgttcc ggtcaactgt tcctgtgcag
gtgagtatta tcaaacaaac 360acatcctatg agttccataa ttcagaaact tacttcttga
ttgccaacaa tacttttgag 420ggcctcacaa catgccaagc tttggagaac caaaaccaca
accctgcaaa catataccct 480ggtagaaggc ttttagtgcc tcttagatgt gcttgtccca
caaagaatca aactgagaaa 540ggcatcaggt acctcctaag ttacttggta aactggggtg
attctgtttc attcattagt 600gagaaatttg gtgtcaactt tatgaccact cttgaagcta
atacacttac tctcacccaa 660gccacgatct atccctttac cacaattcta gttccccttc
atgacaagcc ctcaagttct 720caaactgttt cgccaactcg gcgcactcca ccaccctctc
ctccctcttc tgatcatagc 780tcaaacaaaa catgggtgta tgtagttgtt ggggttgttg
tgggagctat tgccttaata 840tcggttctct gtgctgtcat tttcttcaca cgctatcgca
aaaatagaaa gaaagatgac 900tcagtggtag tagggtccaa gagttttgag gcaattgagg
aaaaaccaga agtgaaagtg 960aatgaaaaat tgtcagagat catatctggc atagctcagt
ctttcaaagt gtataatttt 1020gaggaactac agcgtgcaac agataacttt agtcctagca
gctggatcaa agggtctgtt 1080tatcgcggtg tgattaacgg tgatttggct gcaattaaaa
ggatagaagg agatgtgtca 1140aaagagatag agatactgaa caaaatcaac cattccaatg
ttatacgcct ttccggggtt 1200agcttccacg aggggggttg gtaccttgtt tatgagtatg
ccgctaatgg ggacttgagt 1260gaatggatct acttccacaa cgtgaatggg aaatttctga
gttggacgca gagaatgcag 1320attgcattgg atgtggccac aggacttgac tatcttcaca
gtttcacttc tcctcctcat 1380atccacaagg atataaacag cagtaacatt cttctggatg
gtgatttcag gggaaaggtc 1440acgaatttaa gccttgctag gtgtttggaa ggaggggacg
atcaacttcc cgcgacgagg 1500cacattgttg ggacaagagg ctacatggct ccagagtatt
tggaaaatgg tcttgtgtct 1560acaaagcttg atgtgtatgc atttggggta cttatgctgg
aaatggtcac tggaaaagag 1620gttgctgcta ttttaactga agatgagaca aaattgtcac
atgttttaag tggcatactt 1680ggtgaggaaa gtggcaagga gatgttgaag gagtttgtgg
atccctcttt gggagaaaat 1740tgtccattgg aacttgctat gtttgtgatt gaaatgattg
ataattgcat aaagacagat 1800ccggcaagtc gccctagtgt gcatgagatt gtgcaatcta
tgtcaagaac cctgaagtct 1860tcactgagtt gggaaaggtc aatgaatgtc ccacgaaatt
aa 1902591797DNAGlycine max 59atggctgtct tctttccctt
tcttcctctc cactctcaga ttctttgtct tgtgatcatg 60ttgttttcca ctaatattgt
agctcaatca caacaggaca atagaacaaa cttttcatgc 120ccttctgatt caccgccttc
atgtgaaacc tatgtaacat acattgctca gtctccaaat 180tttttgagtc taaccaacat
atccaatata tttgacacaa gccctttatc cattgcaaga 240gccagtaact tagagcctat
ggatgacaag ctagtcaaag accaagtctt actcgtacca 300gtaacctgtg gttgcactgg
aaaccgctct tttgccaata tctcctatga gatcaaccaa 360ggtgatagct tctactttgt
tgcaaccact tcatacgaga atctcacgaa ttggcgtgca 420gtgatggatt taaaccccgt
tctaagtcca aataagttgc caataggaat ccaagtagta 480tttcctttat tctgcaagtg
cccttcaaag aaccagttgg acaaagagat aaagtacctg 540attacatacg tgtggaagcc
cggtgacaat gtttcccttg taagtgacaa gtttggtgca 600tcaccagagg acataatgag
tgaaaacaac tatggtcaga actttactgc tgcaaacaac 660cttccagttc tgatcccagt
gacacgcttg ccagttcttg ctcgatctcc ttcggacgga 720agaaaaggcg gaattcgtct
tccggttata attggtatta gcttgggatg cacgctactg 780gttctggttt tagcagtgtt
actggtgtat gtatattgtc tgaaaatgaa gactttgaat 840aggagtgctt catcggctga
aactgcagat aaactacttt ctggagtttc aggctatgta 900agtaagccta ccatgtatga
aactgatgcg atcatggaag ctacaatgaa cctcagtgag 960cagtgcaaga ttggggaatc
agtgtacaag gcaaacatag agggtaaggt tttggcagta 1020aaaagattca aggaagatgt
cacggaagag ctgaaaattc tgcagaaggt gaatcatggg 1080aatctggtga aactaatggg
tgtctcatca gacaatgatg gaaactgttt tgtggtttat 1140gaatacgctg aaaatgggtc
tcttgatgag tggctattct ccaagtcttg ttcagacaca 1200tcaaactcaa gggcatccct
tacatggtgt cagaggataa gcatggcagt ggatgttgcg 1260atgggtttgc agtacatgca
tgaacatgct tatccaagaa tagtccacag ggacatcaca 1320agcagtaata tccttcttga
ctcgaacttt aaggccaaga tagcaaattt ctccatggcc 1380agaactttta ccaaccccat
gatgccaaag atagatgtct ttgcatttgg ggtggttctg 1440attgagttgc ttaccggaag
gaaagccatg acaaccaagg aaaatggtga ggtggtcatg 1500ctgtggaagg acatttggaa
gatctttgat caagaagaga atagagagga gaggctcaaa 1560aaatggatgg atcctaagtt
agagagttat tatcctatag attacgctct cagcttggcc 1620tccttggcgg tgaattgtac
tgcagataag tctttgtcca gaccaaccat tgcagaaatt 1680gtccttagcc tctcccttct
cactcaacca tctcccgcaa cattggagag atccttgact 1740tcttctggat tggatgtaga
agctactcaa attgtcactt ccatagcagc tcgttga 1797601800DNAGlycine max
60atggctgtct tcttttcctt tcttccgctc cgttctcaga ttctttgtct tgtacttatg
60ttgtttttca ctaatattgt agctcaatca caacagacca atgaaacaaa cttttcatgc
120ccttctgatt caccaccgcc ttcatgtgaa acctatgtaa catacattgc tcagtctcca
180aattttttga gtctaaccag catatccaat atatttgaca caagtccttt atccattgca
240agagcaagta acttagagcc tgaagacgac aagctgatcg cagaccaagt cttactgata
300ccagtaacct gtggttgcac tggaaaccgt tctttcgcca atatctccta tgagatcaac
360ccaggtgata gcttctactt tgttgcaacc acttcatacg agaatctcac gaattggcgt
420gtagtgatgg atttaaaccc cagtctaagt ccaaatacgt tgccaatagg aatccaagta
480gtatttcctt tattctgcaa gtgtccttca aagaaccagt tggacaaagg gataaagtac
540ctgattacat acgtgtggca gcccagtgac aatgtttccc ttgtaagtga aaagtttggt
600gcatcaccag aggacatatt gagtgaaaac aactatggtc agaactttac tgctgcaaac
660aaccttccag ttctgatccc agtgacacgc ttgcctgttc ttgctcaatc tccttcagat
720gtaagaaaag gcggaattcg tcttccagtt ataattggta ttagcttggg atgcacgcta
780ctggtcgtgg ttttagcagt attactggtg tatgtatact gtctgaaaat taagagtttg
840aataggagtg cttcatcagc tgaaactgca gataaactac tttctggagt ttcaggctat
900gtaagtaagc ctaccatgta tgaaactgat gcgatcatgg aagctaccat gaacctcagt
960gagcagtgca agattgggga atcagtgtac aaggcaaaca tagagggtaa ggttttggca
1020gtaaaaagat tcaaggaaaa tgtcacagag gagttgaaaa ttctgcagaa ggtgaatcat
1080ggaaatctgg tgaaattaat gggtgtctcg tcagacaatg atggaaattg ttttgtggtt
1140tatgaatatg ctcaaaatgg atctcttgat gagtggctat tctacaagtc ttgttcagac
1200acatcagact caagggcctc ccttacatgg tgtcagagga taagcatagc agtggatgtt
1260gcaatgggtt tgcagtacat gcatgaacat gcatatccaa gaatagtcca cagggacatc
1320gcaagcagca atatccttct tgactcaaac ttcaaggcca agatagcaaa tttctccatg
1380gccagaactt ttaccaaccc cacgatgcca aagatagatg tctttgcatt tggggtggtt
1440ctgatagagt tgcttactgg taggaaagcc atgacaacca aggaaaatgg tgaggtagtt
1500atgctgtgga aggacatttg gaagatcttt gatcaagaag agaatagaga ggagaggctc
1560aaaaaatgga tggatcctaa gttagagagt tattatccta tagattatgc tctcagcttg
1620gcctccttgg cagtgaattg tactgcagat aagtctttgt ccagatcaac cattgcagaa
1680attgtcctta gcctctccct tctcactcaa ccatctcccg tgacattgga gagatccttg
1740acttcttctg gattagatgt agaagctact caaattgtca cttccatagc agctcgttga
1800611794DNAGlycine max 61atggggcctt caccttcctt cctttacact tgcaatggct
tcaacaaaac ctgcatgtcc 60ttcctcatct tcaaatccaa acctcctttc aactccataa
ctacaatctc caacctcaca 120tcatcaaacc cagaagagct tgcaagaatc aatgatgtca
ctgtgctcaa agtgtttcca 180actgggaaag aggtgattgt ccctttgaac tgttcttgtt
taactaggga atattaccaa 240gctgaaacca aatatgtatt ggggcaatcc ccaacatatt
tcacggtggc aaatgatact 300tttgagggtt tgaccacttg tgataccctc atgcgtgcta
attcatatgg tgaacttgat 360ttgcttcctg gcatggaact gcacgtgcca ctcagatgtg
cttgtcctac atggcatcag 420atcacaaatg gcaccaagta tttgctgaca tactcagtga
actggggtga tagcattaaa 480aatattgctg caaggttcaa tgtagcagca ggtaatgtgg
tcgatgccaa tggtttttcc 540acacaaacac aaactatttt tccattcaca actgtactaa
ttcctttgcc aagtgaacct 600gtgagttcaa tggccataat tgtcaatggt ccaccagctg
tgtcacctct tcctgtttgc 660agctcagaaa agtgcaactc aaggaggaaa ctttacattg
ttattgccac cactgggggt 720tccatgctgg ttctttgtgt tgtcttattt gggggttttc
tgtgcagaaa gagatcagca 780aggttcatta agagaggtga gcaaagtgag aaagcaaaga
agttgtcttc ggaagatatc 840cgcggcaaga tagccatcat tgaacatcac tccaaggtgt
acaagtttga ggaaatagag 900gaggccactg aaaactttgg ttcaaagaat agaattaaag
gttctgtgtt tcgtggagta 960ttcggtaaag agaaaaacat attggctgtc aagaaaatga
gaggagatgc atccatggaa 1020gtgaatttgc tggaaaggat caatcatttc aacctgataa
aactgcaagg ttactgtgaa 1080aatgatggtt tcccctatct tgtttatgag tttatggaaa
atggctcttt gagagaatgg 1140ctaagcagaa acaggtcgaa ggagcaccag agtttggcat
ggaggatcct gattgctctg 1200gatgttgcca atggtcttca gtatcttcac aacttcacag
aaccttgcta tgtgcacagg 1260aacataaaca gtggaaacat tctactgaac agagatctaa
gggccaagat agcaaacttt 1320gctcttgttg aagaatcaga aagtaaaata acttctggtt
gtgccgcatc acatgtcgtg 1380aaatcgaggg gttatacggc tccggagtat ctggaggcag
ggatggtcac taccaaaatg 1440gacgtttttg ccttcggagt ggtgctgttg gaattgatca
cgggtaaaga ttctgttacc 1500ctacacgatg gaagagaagt gatgcttcat gcaatcatag
taaatctcat tggtaaagag 1560aatttagaag agaaggtgag tttgttcatt gatccttgcc
tcactgtcac tggaaacagt 1620gagatagtat gtgctcccca gctagttaaa cttggtctgg
catgcttgat ccaagaacca 1680gcagagagac caaccatggt agaggtagtc tctagcttat
tgaaaatata tacaagttat 1740atggagcaaa taataccacc tagcatcagc aacagtccta
gcatggagag gtga 1794621905DNAGlycine max 62atgcatctct ttcccttcat
ccccataatc attttcacac tattgatcca caacttctct 60ctgattctgg ggcagcaacc
ttatattggt ttaggcacag tagcgtgtcc aagaaggggt 120aacaaaaatt ctatccgagg
ttacacttgc aatggtgcaa accatagctg ccaaagttac 180ctcaccttca gatctcaacc
catttacaac tctgtcaaga caatatcaac tttgttgggt 240tctgacccat cccagcttgc
taaaataaac tcagtttcca tgaatgacac ctttgagaca 300aacaagttgg tgattgttcc
ggtcaactgt tcctgttcag gtgagtatta tcaaacaaac 360acatcctatg tgttccagaa
ttcagaaact tacttgttga ttgctaacaa cacttttgag 420ggcctcacaa catgtcaagc
tttggagaac caaaaccaca accctgcaaa catatacccc 480ggtagaagac ttttagtgcc
tctcagatgt gcttgtccca caaagaacca aaccaagaaa 540ggcatcaggt acctcttgag
ttacttggtg aactggggtg attctgtttc attcattagt 600gagaaatttg gtgtcaactt
tatgtctact cttgaagcta atacccttac tctcacacaa 660gccatgatct atccctttac
gacaatttta gttccccttc atgacaagcc ctcaagttct 720caaaccgttt cgccaactca
acgcattagt ccaccaccct cacctccctc ttctgatcat 780agctcaaaca aaacatgggt
gtatgtagtt gttggggttg ttgtgggagc tattgcctta 840acatcggttc tctgtgctgt
cattttcttc aaacgctatc gcaaaaatag aaacaaagat 900gactcattgg tggcagtgcc
taagagtttt gaggcaattg aggaaaaacc tcaagtgaaa 960gtgaatgaaa aattgtcaga
gaacatatct ggcatagctc agtctttcaa agtgtataac 1020tttgaggaac tacagcgtgc
aacagataac tttagtccta gcagctggat caaagggtct 1080gtttatcgcg gtgtgattaa
tggtgatttg gctgcaatta aaaagataga aggagatgtg 1140tcaaaagaga tagagatact
gaacaaaatc aaccatacca acgttatacg cctttctgga 1200gttagcttcc atgagggtcg
ttggtacctt gtttatgtgt atgctactaa tggggacttg 1260agtgaatgga tctacttcaa
caacgtggac gggaagtttt tgagttggac tcaaagaatg 1320caaattgcat tggatgtggc
aacaggactt gactatcttc acagtttcac ttctcctcct 1380cacatccaca aggatattaa
cagcagtaac attcttctgg atggtgattt caggggaaag 1440gtcgcgaatt taagccttgc
taggtgtttg gaaggagggg atgatcaatt tcccacgacg 1500aggcacattg ttgggacaag
aggctacatg gctccagagt atttggaaaa tggtcttgtg 1560tccacaaagc ttgacgtata
tgcatttggg gtactgatgc tggaaatggt cactggaaaa 1620gaggtcgctg ctattttaac
tgaagatgag acaaaattgt cacatgtttt aagtggcata 1680cctggtgaga gaagtggcaa
ggagtggttg aaggagtttg tggatccctc tttgggagag 1740aattgtccat tggaacttgc
tatgtttgtg attgaaatga ttgatgattg cataaagaca 1800gatccagcaa gtcgccctag
tgtgcatgag attgtgcaat ctctttcaag aacagtgaac 1860tcttcactga gttgggaaag
gtcaatgaat gtcccacgaa attaa 1905631992DNAGlycine max
63atggccacaa tccagtgcgc cacaaccacc ctaatactgt tactcctcct cctcctcatc
60ataccaagat ccaattccca acaagagtac gtaaacaaca agcaactaga ctgcaacaac
120gaatacaact ccacaaaggg taacctctgc aacagtctcc cgtcatgcac ctcctacctc
180accttcaagt cctccccgcc ggaatacacc acccccgccg ccatctcctt cctcctcaac
240tccactccgg ccctcatcgc cgccgccaac aacatcaccg acgtccagac cctcccggcc
300gacaccctcg tcaccgtccc cgtcaactgc tcctgctccg gcccctacta ccagcacaac
360gcctcctaca ccatcaaagt ccaaggcgaa acctacttct ccatcgccaa caacacctac
420caggccctca ccacgtgtca ggccctcgag cttcaaaaca ccgtcggcat gcgcgacctc
480ctcaagggtc agaacctcca cgtgcctctc aggtgcgcgt gtcccacgca gaagcagcgc
540gaggcagggt tcaagtactt gctcacttac ttggtttctc aaggagaatc ggtttccgcc
600attggagata tcttcggtgt cgacgaacag agcattctcg acgccaacga gctttctact
660tcttccgtta ttttctactt cacgccgatt tcggttcctt tgaaaacaga gccaccggtt
720acaataccga gagcagcgat tcccccggag gattccccgt cgccgccgct gccgcctgct
780ccggcggggg acggagattc cgactcttcc aagaaatggg tcattgtcgg aatcgtggtt
840ggggttgttg tgttgcttat tttaggtgct gctctgtttt acctctgttt ctatcggcgg
900cggcggcggg tggaacaccc tccgccgccg ccttcagcga aggctttctc gggctctacc
960accacgaagg cgactattcc gacgacgcag tcgtggtctc tttcctcgga aggggttcgt
1020tacgcgattg aatcgttgag tgtgtacaag ttcgaggagt tgcagaaggc tacggggttc
1080ttcggcgaag agaacaagat caagggttct gtttataggg cttctttcaa gggtgattat
1140gccgcggtga agattctcaa aggtgatgtg tcgggtgaga ttaaccttct caggaggatt
1200aaccacttca acattataag gttgtcgggt ttctgtgtct acaaaggtga cacctatctt
1260gtgtacgagt tcgcggagaa tgattctctt gaggattggc ttcacagtgg tagcaagaag
1320tacgagaatt ctacatctct gagttgggtg cagagggttc atattgctca tgatgttgct
1380gacgcactta actaccttca taactatact agtcctcctc acgtacacaa gaatttgaag
1440agtgggaacg tgcttttgga tggcaatttt agggctaaag tttcgaattt gggattggca
1500agagctgtgg aggatcacgg ggatgatggg gggtttcaat tgacaaggca tgtggtgggg
1560actcatgggt acatggcacc tgagtacatt gagaatggtt tgattactcc aaagatggat
1620gtttttgcgt ttggggttgt gcttttggag cttctttcgg ggagagaagc tgttgttggt
1680ggagaccaga atggatctgg ggagaagatg ctgtcggcaa ctgtgaatca cgtgcttgaa
1740ggagagaatg ttagagagaa acttagaggt ttcatggatc caaatttgag agatgaatat
1800cccttggaac tggcttattc catggccgaa cttgccaaac tctgtgttgc tcgtgacctc
1860aatgcaaggc cacagatttc tgaagctttc atgattttgt ctaagattca atcctccacg
1920ttggattggg atccgtctga cgagcttgaa cggtctagat ctgttggcca aatctctgat
1980agcagcagat ag
1992641854DNAGlycine max 64atgcatcagt tcttctctgc tgtcttcctc ttcctcctcc
ttccactttg cagcaacgct 60caaacagcac gacaagcgaa caacacgggc ttcacgtgca
acttcacacg cacgtgcacg 120tcctacgcct tctaccgggc caccgccccc aacttcaccg
acctggcctc catcggcgac 180ctcttctcgg tgagccgcct catgatatca accccatcca
acatctcctc ttcttccctc 240aacacccctt tactcccaaa tacacccctc ttcgtcccct
taacctgctc ctgcaacccc 300gtcaacgcct ccttcggctc cctctcctac gccaacatct
cctacaccat caaccccggc 360gacaccttct tcttagtctc caccatcaag ttccagaacc
tcaccacttt cccctccgtt 420gaagtcgtca accccaccct cctcgccacc aacctctcca
tcggccagga caccatcttc 480cccatcttct gcaaatgccc ccccaacagc caaggaacaa
attacatgat ctcctacgtc 540gtccaacccg aggataacat gtcgtcaata gcttcaactt
tcggtgctga agaacagtcc 600atcattgacg ccaacggcgg tgagacaact ctacatgact
atgacaccat tttcgtgccc 660gtggcgcggc tgccggctct gtcgcagccc gcggtggttc
ctcccgcgcc gccgccagta 720attgggagca ataatgatga taggacaggg actgttagag
ggttgggggt tggattgggg 780attgtggggt tgttgttgat tttggtgagt ggggtttggg
tgtacagaga ggttgttgtg 840atgaagggag tggtgagaga tgatgaggag aagaatgtgt
atttgggagg gaaagcggag 900ggtaagaatt tggatgtcaa gttgatggct aatgtgtctg
attgtttgga taagtatagg 960gtttttggga ttgatgagct ggtggaggcc actgatgggt
ttgatcagag ttgcttgatt 1020caggggtcgg tgtacaaagg cgaaattgat gggcatgtct
ttgccatcaa gaagatgaag 1080tggaatgcct atgaggaact caagatcttg cagaaggtaa
accatgggaa tctagtgaag 1140ttggagggat tttgcataga cccagaagaa gcaaattgct
atctagtcta tgaatatgta 1200gagaatggat ctctatactc atggctgcat gagggcaaaa
aagaaaagtt gagctggaaa 1260ataaggctaa gaatagccat tgacattgcc aatggtctcc
aatacatcca cgagcacaca 1320aggccacggg ttgtgcacaa agacatcaag agcagcaaca
ttctcttgga ctccaacatg 1380agagccaaaa ttgccaattt tgggcttgca aagtctggaa
tgaatgccat taccatgcac 1440attgtgggaa cacagggcta tattgcccct gaatatcttg
ctgatggtgt ggtctctaca 1500aagatggatg tgttcgcatt tggggttgtg ttgcttgaat
taatttctgg gaaggaagtc 1560attaacgagg agggaaatct cttgtgggca agtgccatca
aaacctttga agtggacaat 1620gagcaagaga agacaaggag gctgaaggaa tggttggaca
aggatatttt aagggagaca 1680ttctcaatgg aaagtttgat gggtgctctc accgttgcaa
ttgcttgctt gcatagagac 1740ccgtctaaga ggcccagcat aatggatatc gtctatgcac
tgagtaaaag tgaagatatg 1800ggctttgata tttccgatga tgggattggt tccccacggg
tgatagctag gtga 1854652055DNAGlycine max 65atgatcattg catttcacat
gaaggctttg gtctttttcc tttggctgtt tgtcccctca 60ttgggcaaag acttgctaag
ctgtgagaca acttcacctg atgcctctgg ctaccattgc 120attgaaaatg tgtcacagaa
ccaatgtgag acatttgcac tctttttaac aaactcttac 180tactcctctc tctccaacct
cacctcttat ttgggactaa acaagtttgt catagcacaa 240gcaaatggtt tctctgcaga
cactgagttc ctctcccagg atcagcctct actggtacct 300attcattgta aatgcatagg
tggcttctct caggctgagt taaccaaaac taccgtcaaa 360ggagagagct tctatggcat
tgctcaatca ctagagggat tgacaacttg caaggctatt 420agggacaaca accctggtgt
gtcaccttgg aatcttgatg acaaagtgag attggttgtt 480ccattgagat gttcctgccc
attttcatct caagttagac cacaaccaaa gcttctgctt 540tcttatccag taagtgaagg
tgatactatt tccaatttgg cctcaaagtt caatattact 600aaagaagcta ttgtttatgc
taataacata tcatcacaag gccttaggac tagaagtagt 660cttgcaccct ttacctctat
tctcattcca cttaatggta agcctatcat tggtcctttg 720gttaagccca aggaacccga
ttctggcaat caaacaacca gcatcccagt gacaagtcca 780cacaagaaat caccaatgtg
gaagactgaa ttgtgcattg gcctagctgg ggttgcactt 840ggagtctgca ttgcttttgc
tgccgccttt ttcttcatca gattgaaaca caagaaggag 900gaagagaatt catgcaaaga
aggggacttg gagctgcaat atctgaacca aagtgtgaga 960accacctcaa ctagtgacaa
gaaagtttca tttgaagggt ctcaggatgc tcttgatgtg 1020aaaatcgtgg acgccctgcc
gcggaagttg ttgctggata catacaccat tgaagatgtg 1080agaaaagcaa ctgaagattt
cagctcaagc aatcacattg aagggtcagt gtatcacggt 1140cgtctgaacg ggaagaacat
ggcaatcaaa gggacaaagg cagaagtagt gtcaaagata 1200gatcttggcc ttttccatga
tgcacttcac catcatccca acatactcag gcttcttgga 1260actagcatgt tagaggggga
gcagcaagaa gagtcatttt tagtttttga gtatgccaaa 1320aatggttcat tgaaagattg
gctccatggt ggattagcca tcaagaacca attcattgct 1380tcctgctact gtttcctgac
ttggagccaa aggctcagga tctgccttga tgtggccggt 1440gccttgcagt atatgcacca
tgttatgaat ccaagctatg tgcatagaaa tgtaaagagc 1500aggaacatct ttttggatga
agaatttggt gccaagatag ggaattttgg tatggcaggt 1560tgtgttgaga atgacactga
ggaccctcaa ttctattcca ccaaccctgc ctcttggagt 1620cttggttatt tggcacctga
atatgtgcac caaggtgtaa tttccccaag tgttgatatc 1680tttgcttatg gggtggtttt
gttggaagtt ttgtccggtc aaacacctat aagcaggccc 1740aatgagaagg gagaaggaag
catttggctt acagataaaa tcaggtccat tttggtgtca 1800gaaaatgtga atgaactaag
ggattggata gacagtgcat tgggggagaa ctattcattt 1860gatgcagctg tgacacttgc
caacattgca agagcttgtg tggaggaaga ttcctctttg 1920agaccaagtg caagggaaat
tgttgagaag ctatcaagat tggtggagga attaccacaa 1980ggggaaaatg acatgttaat
gtgtgaaagc tctagcaaac ctctggtgaa ggcagtggaa 2040aacaatgttg aataa
2055662340DNAGlycine max
66atgaactgca ccgacacaag tcgtgtttgc acttctttct tggcctttaa gcctcatcaa
60aaccaaacct tggctgtgat acaaagcatg tttgatgtgt tgcccggtga gatcaccgtt
120gaaggcaatg gctgggatta tatattcatc aggaaaaact gttcttgtgc tgctggtatg
180aagaagtatg tgtctaacac cacactcact gtgaaatcca atggaggatt tgagcatgat
240ttggtgatgg aagcctatga caggcttgct ctcttgccca acaccacgac gcgttgggca
300agagaaggcg gtatcatatc tctgagcttg ttctgtagtt gctctagtgg actgtggaac
360tatctgatga gctatgtcat cagagatggg gacagtgttg aatcactagc aagcaggttt
420ggggttagta tggatagcat tgagacagtg aatggcattg acaatcctac tgttggttct
480ctcgtttata tacctttgaa ttcggttcct ggtgagtctt accacttgat gaatgatact
540cctccagctc caaccccttc gccgtctgtt aataattttt cagctgacca agtcaaccag
600aaggctcatg taccccatga atggatcata ggaggtttag gggttggtct tgctctgata
660atattaacca ttattgtgtg tgtggcccta agatcaccca attgtttggt tgaagccgga
720aataatgcaa aagattcttc aggaaagatc tctaataagt tctatgtttt tggaaatcca
780agtttgtttt gtggatgtgt caaacctgtg gaccagaagc aaactgatgg tgaatccagc
840agtcaccaaa ttaccggtac caaaacatca actctaatac ctgacatgtt ggacatggat
900aagcctgtag ttttttcata tgaagaaatt ttttcttcaa ctgatggttt ctctgattca
960aatctacttg ggcacagaac atatggctct gtttactatg gtctcctcgg tgaccaggaa
1020gttgctatta aaagaatgac atctactaaa actaaagaat ttatgtcaga ggtaaaagtt
1080ctgtgcaagg ttcatcatgc taatctggta gaattgatcg gctatgcggt tagccatgat
1140gagtttttcc ttatttatga atttgctcag aaaggttcac tcagcagcca tttgcatgat
1200cctcaaagta agggtcattc acctctttct tggatcacaa gggtccagat tgcacttgat
1260gctgctaggg gccttgaata catacacgag catacaaaaa ctcgttacgt ccatcaagat
1320atcaagacaa gcaacatttt acttgatgct tcctttagag ccaagatatc agattttggg
1380ttagcaaaac ttgtaggtaa aacaaatgag ggagaaaccg cagcaaccaa agttgtaaat
1440gcatatggat atcttgctcc agaatacttg agcaatggcc ttgcaacgac caaaagtgat
1500gtctatgcat ttggtgttgt tctttttgag attatatcag ggaaggaagc cattattcaa
1560acacaaggcc ctgaaaaacg atcattggca tctattatgt tggcagttct taggaactca
1620cctgataccg tgagcatgtc aagcacgaga aacctcgttg atcctattat gatggatatg
1680tatccccatg attgtgtata taagatggcc atgctggcaa agcaatgtgt ggatcaggat
1740cctgtattac gccctgatat gaaacaagtt gtgatttccc tctcacaaac cctgttgtct
1800tctgttgagt gggaagccac tcttgctggg aacagccagt tgaaactggt gaaacattgg
1860tacaaggcta aggcttactt gacaaggtgg atcaaaagat cttctggaat aaaggacggc
1920ccaattatca atcaagagaa cgtccccttt ttgccaagga atggcaacag attcttcctc
1980cagaatgtgg agacaatcat agacaatgtg agaggggtac ttctccttca tcctttgata
2040caccacgcgg ctaagaacaa tgattcaagg aaagcttttt gggttttgac ggagtccctg
2100agaggaaccg aacttgcagt tgcaggtgga ggagacacca cagcggggaa tggaactccg
2160ttcacaagct tctgcccggg aacttggatt tccaccaatg aacttcccat gttttctccc
2220acagtcaccc gccacggagg aaaggataac tttagagacg gaaactctga atgcgaccag
2280tgtcatatgg gctatcctat ggcccattta gcgcactcac aacccatggg ctcatgctag
2340671809DNALotus japonicus 67atggaaccaa aattaacgtt ttcactttcg
ttcttgttaa cgttgttgtc cccatttgca 60gaatcaaagt gcatcaaagg ttgtgattta
gctctagctt catactacca atggagtggt 120tcaaacctca cctatatctc aaagatcatg
gagtctcaga ttctttcaaa accacaagac 180attgtcacct acaacaaagg caaaagaaac
tttggtgttt tctccacaag ggttaacgtt 240ccatttcctt gtgattgcat caatggtgag
tttttgggtc acacgtttga gtaccagctt 300cagccagagg agacatacac aacagttgca
agtgaaacgt tttccaattt gactgttgat 360gtgtggatgc aagggtttaa catttatcca
ccgaccaata ttcctgattt tgctgtgttg 420aatgttactg ttaattgttc ttgtgggaat
agtgaggttt ctaaggatta tgggttgttt 480atcacgtatc cgcttagaat tgaggatagt
ttgcagtcca ttgcggaaga gatgaagctt 540gaggctgagt tgctgcagag gtacaatcct
ggtgtgaatt tcagccaagg aagtggcctg 600gttttcattc ctggaaaagg cttttcaggc
ggggttattg ctggaatatc tgttggagta 660ctagttgggc ttttgttagt tgcattttgt
gtgtatacta aacatctgca aaagaaaaaa 720gcattggaga aaaaattgat cttggatgat
tccacagtta actctgctca agttagtaat 780gattctggtg gcattatgat ggacaaatca
cgcgagttct catataaaga actagccgat 840gccacaaata actttagcgt ggctaataga
attggtgaag gtggttttgg aacagtttac 900tatgcagatc tgagtggcga gacaatacca
gaaggcaata atgcgaaaca atcaagagcc 960actgacaata aaactgccat caagaagatg
aacatgctcg catcgagaga atttcttgct 1020gaagtgaaag tcttagctaa tgttcatcac
ttgaacctgg tacggttgat tggatactgc 1080attgagggtt ctctcttcct tgtatatgaa
tacattgaca atggaaactt aaaacaaagt 1140ttgcatgatt tagaaagaga gcctttgcca
tggtctacta gggtgcaaat tgctttggat 1200tcagccagag ctcttgaata cattcacgag
catacagtgc atgtgtatat tcatcgtgac 1260ataaagtcgg aaaatatttt actagacaat
agcttccatg caaaggttgc agattttgga 1320ttgtccaagc tggttcaagt tggaaattcg
attggttctt ctgttaatat gatgaagggt 1380acatttggtt acatgccacc agaatatgca
cgtggtgttg tctctccttc tcccaaaata 1440gatgtttatg cctttggagt tgttctttat
gaacttattt ctgctaaaga agctgtaatc 1500agggatggtg ctcaatctaa gggcctcgta
gcattgtttg atgaagtttt gggtaatcag 1560ctagatccta gggaaagtct tgtaagtttg
gtggatccta ggcttcaaga taactactca 1620attgactcag tttgcaagat ggcacaactt
gccaaagtgt gcacagagcg tgatccaacg 1680ggacgtccaa gtatgagatc tgttatggtt
gctctaatga cacttagttc tacaactcaa 1740agttgggaca ttgcatcctt ctatgaaaat
ccagctcttg taaatcgaat gtcgggaaga 1800ttggagtag
1809681794DNALotus japonicus
68atggaatcta gggttctttc agaacctgaa gatattatca actacaacca agaccatgtt
60aggaacccag atgtattaca ggttcacacc agggttaacg ttccattccc ttgtgattgt
120atcaatggcg agtttttagg tcatatattt cggcatgagt ttcatgaagg ggacacctat
180ccatccgttg caggcacggt cttttccaat ttgaccaccg atgcgtggtt gcaaagcacg
240aacatttatg gaccaacctc gattcctgtt ttagcgaagg ttgatgtcac ggttaactgt
300tcctgtgggg acatcaaggt ttccaaggat tatgggttgt tcatcacata cccacttaga
360gctgaggata ctttggagtc cattgcagaa gaggcgaagc ttcaaccgca cttgctgcag
420aggtacaacc cgggtgtgga tttcagccgc ggaaatggtc tggtttttat tccggggaag
480gatgaaaatg gagtttatgt tcccttgcac attagaaaag caggtctaga tagggtagtt
540gcaggagtat ccataggagg tacatgtggg cttctgttat ttgcactttg tatatatatg
600agatactttc ggaagaagga aggagaagag gctaaatttc caccaaaaga atctatggag
660ccttcaattc aagatgatag taaaattcat cctgctgcca atggaagtgc tggctttaaa
720tatattatga tggatagatc atcagagttt tcatatgaag aacttgccaa tgccacaaat
780gacttcaatt tggctaataa aattggtcaa ggtggtttcg gagaggtcta ctatgctgag
840ctgagaggcg agaaagttgc tatcaagaag atgaagatcc aagcatcacg ggaatttctt
900gctgaattga aagtcttaac aagtgttcat cacctgaact tggtacgctt gattggatat
960tgcgttgaga gatctctctt tcttgtctac gaatacatgg acaatggaaa tttaagccaa
1020catttacgcg agtcagagag agaactgatg acatggtcta ccaggttgca aattgcactg
1080gatgtagcga gaggccttga atacattcat gactatacag tgcctgtgta tatccaccgg
1140gacattaaac cagataatat tttattaaat aaaaacttca atgccaaggt tgcagatttt
1200gggctaacca agctgactga catagaaagt tctgcaatta acactgatca tatggcaggc
1260acatttggtt acatgccgcc agaaaatgca cttggacgtg tttctcgcaa gatagatgtt
1320tatgcatttg gagttgttct ttatgaatta atttctgcca aagaagctgt ggtcgagata
1380aaggagtcat ctacagagct caaaagcctt gaaattaaga ctgatgaacc tagtgttgag
1440tttaagagcc ttgtagcttt gtttgatgaa gttattgacc atgaggggaa tcccattgaa
1500ggtcttagaa aactggtgga tccgaggctt ggagagaact actcaattga ttccattcgt
1560gagatggctc aacttgccaa agcttgcaca gaccgagatc caaaacaacg tccaccaatg
1620agatctgttg tggttgttct aatggcttta aattctgcta ctgatgatag gatgagtcat
1680gctgaggggg aaaatctgat tatggaaggc attcagaaaa attcagggaa ttgttattgt
1740ggagagacag gctctcgaaa tcctgagagg agagataggc tctcaaaaac ctga
1794691902DNALotus japonicus 69atgcattgct ttcccttcac ccttatcttc
ctttccaccc tcttcatctt cttcaccttc 60cctatcactc tagctcagca accttacatt
ggtttagcca ccaatgcgtg ccctcgccgg 120ggaaactcaa attccatccg cggctacact
tgcaacggcg gcagtgcaaa tcacacttgc 180caagcttacc tcaccttcag aactcaaccc
atttacaact ctgtctacac aatttccact 240ctgttatcct ctgacgcacg ccacctagct
gaaacaatgt cagtctcaca aaacaccacc 300ttcgaaacaa acaagcttgt aatcgttcct
gttcaatgtt cttgtgctgg tgagtattac 360caggcaaaca catcatatgc atttcagaac
acagacaccc ccttttcgat tgctaacaac 420acttttgaag gcctcacaac atgtcaagct
ttgatgcatg agaatcacaa ccctggtcat 480ctataccttg gtagagaact cacagttcct
cttagatgtg cttgtcccac taagaatcaa 540acagagaaag gaatcaagta cctcttgagt
tatcttgtga attggggaga ttctatttca 600gtcatcagtg agaaatttgg tgtcagctgt
aataacactc ttgaagctaa ttccctttct 660ctcacaaaag ccaaaattta tccctttacc
acacttttag taccccttca tgataagccc 720tccaattcac aaaccatttt gcaatctcaa
ccatcttcaa cctcatcttc accaccaccc 780tcttcttcca cacatcaaag ctcaaacaaa
acgtggttgt atgtagttgt tggaggagtt 840tttgccttaa ttgttctcac tgctgtcatt
ttctgcatac actatcacaa gggtaggaag 900aaagatgatt ctttgagtca attgactgtg
tctgagagtt ttgaaaacca acaattaggg 960aaggaaatga agaaaggaga tggaaaattg
tcagagttca tacatggcat agctcaatct 1020ttcaaagtgt acagctttga ggaaatacag
cgtgcaacaa ataatttgaa ttcaagcagt 1080ttgataaaag gttctgttta tcgcggtgtt
atgaatggtg atttggttgc aattaagaaa 1140acagaaggag atgtatcaaa agagatacaa
attctgaaca aggtgaacca ttctaatgtt 1200atacgtctct ctggggttag cttcaacgag
ggtcaatggt accttgttta tgagtatgct 1260gctaatggac ccttgagtga gtggattttc
tttgggaagt ttctgagttg gactcagaga 1320atacagattg cactggatgt ggccatagga
cttgactatc ttcatagttt cacttctcct 1380cctcacatcc acaaggatct gaagagtagt
aatattcttc tggacagtga tttccgagca 1440aagattgcaa atttaagcct tgcaaggtct
gtgaaaggag tggatgagga tgatcaattt 1500ctcgcgacaa ggaatattgt tgggacaaga
ggttacatgg ctccagagta tttggaaaat 1560gggcttgtat ccacaaagct tgatgtgtat
gcatttgggg tactgatgct ggaaatcctc 1620acaggaaaag aggttgctgc tattttagca
gaagataata ataagaattt gtcaggcgtt 1680ttaagtgctg tacttggtga ggaaaggctg
aaggagttta tggatccctc tcttcaatca 1740aattatccat ttgaacttgc tatgtttgtg
tttgaaatta tagttggttg catagagaaa 1800gatccagcta gtcgcccttc catgcaggag
attgtaccaa ctctttcaag aaccatgaac 1860tcttcattga gttgggaaat gtcagtgaac
atctcaggat ag 1902703861DNALotus japonicus
70atggcttctc taactcatcc tctatgtgtt ctccttactc taatggctgc agcttcattt
60gcaagtgtgt tttcccttga agtttcatcc aaaacaactt acatggaacc ttttaactgc
120tctacaaaga tcagaacatg caattccttg ctctaccaca taagcatcgg tctcaaggtc
180gaagaaatag cccgctttta ctcggttaat ctctcccgaa tcaagccaat aactcgcggt
240accaagcaag attaccttgt ctcagtgcct tgtacttgca gaaacaccaa tggccttaat
300ggatatttct atcatacatc ctacaaggtt aaggttaatg acagttttgt ggatattcag
360aacctgttct atagtggaca agcttggcct gtgaatgaag atttggtggt tccaaatgag
420actatgacaa tacatattcc ttgtgggtgt tcagaaagtg gctctcaaat tgtcgtcaca
480tacacagttc aaaggaatga tacaccatta tcaattgctc ttttgctaaa tgctacggtt
540gaaggcatgg tgagtgtgaa ctcagttatg gctccgaatc ccacattcat agatgttggt
600tgggtgttat atgttcccaa ggagttgaat ccaatttccc atggaaaaga aaataaacac
660aagctggaga aaattattgg catcttagcg ggtgtgatat tactttcaat tattaccttg
720atcattctta tcgtcaggag aaatagatcc tatgaaacct gcaaagatga tccacgcgct
780atctcaaaaa gatcaatcgg caaaagaact agttccttaa tgaaccgcga ctttcacaaa
840gaatacatgg aagatgcgac atcatttgac tcagaaagac cagtaattta tactcttgag
900gagattgaac aagctactaa tgacttcgat gaaactcgaa ggattggagt cggtggatat
960ggaacagtgt attttggagt gttaggggag aaggtatgga ttatggaaat cccacctaca
1020ttctttaatg gtcctcccca tgctagcatt tctttttcat gctgcaacaa agcattgaag
1080aataatagct attttgattt tcaggaggtt gctataaaga agatgaaatc taacaaatcc
1140aaagaattct atgcagaact caaggccttg tgtaagatcc atcacattaa cattgtggag
1200ttattaggat atgccagcgg agatgaccac ctttacttgg tgtatgagta tgtgcccaat
1260ggatctctca gtgaacatct tcatgatccc ttactgaaag tctgcaacgt tggtcctacc
1320gtttgtttgt taatggagga ggtttacgtg gatggtcact tagctgatgt ggaagttgaa
1380ttagagagag aaagagacca gcaactgatt cagccttctt cctcgcctct agggactcgt
1440tcgttcttta tctcttcctt ttcgctaaat ctcaagcttc gaaacatctt cctcacggtg
1500ctctgttttt taaggaggaa gagggtagaa gatgaccgtt gcaggtctca cgtgcaactc
1560acgtgtttcc ctctctcttc agtccttctt tctctctcca attggacatc cacaaatcta
1620aattgtggta ctggaacagg tcaccagcct ctttcttggt gtgctaggat tcaaattgca
1680ctggattcag caaaaggtat tgaatacata catgattaca caaaagcaca gtatgtgcac
1740cgcgatataa agactagtaa tattcttctt gatgagaagc tcagagcaaa ggtagcagat
1800tttgggcttg caaagctagt agaacgaacc aatgatgaag aattcatagc aacaaggctt
1860gttggaacac caggctatct tccaccagaa tctctaaagg agcttcaagt gacagtaaaa
1920actgatgtat ttgcatttgg ggtggttatg ttagagttga taacagggaa acgtgcacta
1980tttcgtgaca accaagaagc caacaatatg agatcacttg ttgcagttgt taaccaaata
2040tttcaagaag ataaccctga gactgcttta gaagttaccg tggatgggaa tctacaacgt
2100agctatccta tggaagatgt ctacaatatg gcagaactat cacactggtg cttgcgcgaa
2160aatccggtgg acaggcctga aatgagtgag atcgttgtga aattgtcaaa gattataatg
2220tcctcgatag agtgggaagc atcacttggc ggagacagcc aagtcttcag cgggtttaag
2280caatatctga agcaaagtcc cacactgacc aaaacaatgg gcaagcaggt tttaggttca
2340cgcaaaggga gcataaagct cacaaagacg gtctttgaag ttttgcttga gtgtaagcat
2400gggaagagca gtcctcgtcc tcttccttct ccctattctt tcaacaccgt gacgctcgtt
2460gatctgattg gaaattggga tccagctcca gcagcaatgg caggggcagc ttctgctctg
2520ttcctcctcg acatcaaagg ccgcgtcctc atctggcgcg actaccgcgg cgacgtcacc
2580gccctcgacg ccgaacgctt cttcaccaag ctcatcgaaa aacaggctga tgcgcagtct
2640caagatccgg ttgtccacga taacggtatc agctacatgt tcgttcagca cagcaatgtt
2700tacctcatga tagccaccag acaaaactgc aatgccgcta gcctcctctt cttcctccat
2760cgagtagtcg acgtttttaa gcattatttt gaagaattgg aagaggaatc acttagggat
2820aactttgtcg ttgtgtatga actgcttgat gaaattatgg actttggcta ccctcagtac
2880actgaggcga agattcttag tgagtttatc aagaccgatg cttatagaat ggaagttacg
2940cagagacctc ccatggctgt gactaatgcc gtgtcttggc gcagtgaagg gataagctac
3000aagaaaaatg aggtagctgt taggaaaagt gaagaatgcg tactggtttt cttggatgtg
3060gtggagagtg tgaacatact tgtcaatagc aatggacaaa taattagatc tgatgttgtt
3120ggggctctca agatgagaac ttatttgagt ggtatgcctg agtgtaaact tggcttaaat
3180gatagagtat tattagaggc gcaaggtaga acaaccaaag gaaaggcgat agacttggaa
3240gacatcaaat ttcaccagta cggttgcttg actttcttgt ctttactaac tgatcagtta
3300gagtgtgtcc gtttggctcg atttgaaaat gatcgaacaa tttccttcat accacctgat
3360ggagcatttg atttgatgac atacaggctc agtacacagg ttaagccatt aatttgggtg
3420gaagcaactg ttgaaaagca ttctaaaagt cggattgaga taatggtaaa agctagaagt
3480caatttaagg aacgtagcac tgccacaaat gttgagattg agttgcctgt gcctgttgat
3540gcaatgaatc caaatgttcg aacttcaatg ggatctgcag catatgcacc tgaaaaagat
3600gcattaatct ggaaaattcg atccttccct ggaggcaagg agtatatgtt aagagcagag
3660tttcgtcttc ccagtataac agctgaggaa gcaacccctg agagaaaagc tcctatacgt
3720gtgaaatttg aaatacctta ttttactgtt tctggaatac aggtaagata tttgaagatt
3780attgagaaaa gtgggtatca ggctcttccg tgggtgagat acataacaat ggctggagag
3840tatgaactaa ggcttattta a
3861711773DNAMedicago truncatula 71atgaaaccaa tcaaattcat attgtcactt
ttacttatgt tgctggcctc ctctagtgca 60gaatcaaaat gtagcaaaac ttgtgactta
gctttagcat cctattatat atgggaaggt 120acaaacttga catatatttc aaacatcatg
caatccaatg ttgtttcaaa acctctagat 180attttcagct acaatactga tactctacca
aacctagata tgctccgttt ttcttcaaga 240ctgaatgttc cattcccttg tgattgcatt
aatgacgagt ttcttggtca cacgtttttg 300tacgagtttc acccaagaga gacctatgct
tccattgctg agttgacttt tagcaatttg 360accaataaag agtggatgga gaaggtcaat
gttcctgatt ctgtgaaggt taatgttact 420gttaactgtt cctgtggaga caaaatggtt
tccaaggatt atggtttgtt cataacatat 480cctcttagtt ctgaggacac tttggagtcc
attgcgaagc atacaaaagt taagcctgag 540ttgctacaga agtacacccc tggtgtgaat
ttcagcaaag gaagtgggct ggtttttatt 600ccagggaaag ataaaaatgg agtttatgtt
cccttgcccc acggaaaagc aggtcattta 660gctaggagtt tagctactgc tgtcggagga
acatgcacgg ttctgttatt agcaattagt 720atatatgcta tatactttcg aaataagaat
gcaaaggagt ctaaattgcc atcaaaatat 780attgtggtgg acaaatcacc taaattttca
tacgaagaac tagccaatgc tactgataaa 840ttcagcttgg ctaacaaaat tggtcaaggt
ggttttggtg aagtctacta tggagagccg 900agaggcaaga aaactgcaat aaaaaagatg
aagatgcaag caacaaggga atttcttgct 960gaactgaaaa tcttaacaag agttcaccac
tgcaacctgg tacacttgat tggatattgt 1020gttgagggat ctctattcct tgtttatgaa
tacatcgaca atggaaactt aagccaaaat 1080ctacatgatt cagagagagg accgatgacg
tggtctacca ggatgcaaat tgctctggat 1140gtagcaagag gccttgaata cattcatgag
cattcagtgc ctgtatacat ccatcgcgac 1200atcaaatcag ataatatttt attaaatgaa
aactttactg ggaagattgc agattttgga 1260ttaaccaggc ttactgattc tgcaaattca
acagataaca ctcttcatgt ggctggcaca 1320tttggttaca tgccacctga aaatgtatat
ggacgtattt ctcgcaagat agatgtatat 1380gcttttggag ttgttcttta tgaactaatt
tctgctaaac cagctgtgat caagatcgat 1440aaaaccgagt tcgagagcga gattaggacc
aatgaatcta ttgatgaata caagagcctt 1500gtagctttgt ttgatgaagt tattgatcaa
aagggagatc ctattgaagg tctaagaaat 1560ctggtggatc caaggctaga agataattat
tcaattgatt ccattagcaa gatggcaaag 1620cttgccaggg cttgcttaaa ccgagatccg
aaacgtcgtc caacaatgag agctgttgtg 1680gtttctctaa tgacacttaa ttctacaatt
gatgatggaa gtaggtcagc cagtgcagca 1740ttgagtactg ttatggagca tgactcgaaa
tga 1773721863DNAMedicago truncatula
72atgaatctca aaaatggatt actattgttc attctgtttc tggattgtgt ttttttcaaa
60gttgaatcca aatgtgtaaa agggtgtgat gtagctttag cttcctacta tattatacca
120tcaattcaac tcagaaatat atcaaacttt atgcaatcaa agattgttct taccaattcc
180tttgatgtta taatgagcta caatagagac gtagtattcg ataaatctgg tcttatttcc
240tatactagaa tcaacgttcc gttcccatgt gaatgtattg gaggtgaatt tctaggacat
300gtgtttgaat atacaacaaa agaaggagac gattatgatt taattgcaaa tacttattac
360gcaagtttga caactgttga gttattgaaa aagttcaaca gctatgatcc aaatcatata
420cctgttaagg ctaagattaa tgtcactgta atttgttcat gtgggaatag ccagatttca
480aaagattatg gcttgtttgt tacctatcca ctcaggtctg atgatactct tgcgaaaatt
540gcgaccaaag ctggtcttga tgaagggttg atacaaaatt tcaatcaaga tgccaatttc
600agcataggaa gtgggatagt gttcattcca ggaagagatc aaaatggaca tttttttcct
660ttgtattcta gaacaggtat tgctaagggt tcagctgttg gtatagctat ggcaggaata
720tttggacttc tattatttgt tatctatata tatgccaaat acttccaaaa gaaggaagaa
780gagaaaacta aacttccaca aacttctagg gcattttcaa ctcaagatgc ctcaggtagt
840gcagaatatg aaacttcagg atccagtggg catgctactg gtagtgctgc cggccttaca
900ggcattatgg tggcaaagtc gacagagttt acgtatcaag aattagccaa ggcgacaaat
960aatttcagct tggataataa aattggtcaa ggtggatttg gagctgtcta ttatgcagaa
1020cttagaggcg agaaaacagc aattaagaag atggatgtac aagcatcgtc cgaatttctc
1080tgtgagttga aggtcttaac acatgttcat cacttgaatc tggtgcggtt gattggatat
1140tgcgttgaag ggtcactttt cctcgtatat gaacatattg acaatggaaa cttgggtcaa
1200tatttacatg gtataggtac agaaccatta ccatggtcta gtagagtgca gattgctcta
1260gattcagcca gaggcctaga atacattcat gaacacactg tgcctgttta tatccatcgc
1320gacgtaaaat cagcaaatat attgatagac aaaaatttgc gtggaaaggt tgctgatttt
1380ggcttgacca aacttattga agttggaaac tcgacacttc acactcgtct tgtgggaaca
1440tttggataca tgccaccaga atatgctcaa tatggcgatg tttctccaaa aatagatgta
1500tatgcttttg gcgttgttct ttatgaactt attactgcaa agaatgctgt cctgaagaca
1560ggtgaatctg ttgcagaatc aaagggtctt gtacaattgt ttgaagaagc acttcatcga
1620atggatcctt tagaaggtct tcgaaaattg gtggatccta ggcttaaaga aaactatccc
1680attgattctg ttctcaagat ggctcaactt gggagagcat gtacgagaga caatccgcta
1740ctacgcccaa gcatgagatc tatagttgtt gctcttatga cactttcatc accaactgaa
1800gattgtgatg atgactcttc atatgaaaat caatctctca taaatctgtt gtcaactaga
1860tga
1863731875DNAMedicago truncatula 73atgaatctca aaaatggatt actattgttc
attctgtttc tggattgtgt ttttttcaaa 60gttgaaacca aatgtgtaaa agggtgtgat
gtagctttag cttcctacta tattatgcca 120tcaattcaac tcataaatgt atcaaacttt
atacaatcaa agattgttct taccaattcc 180tttgatgtta taatgagcta caatagagtc
gtagtattcg ataaatctgg tcttatttcc 240tatactagaa tcaacgttcc gttcccatgt
gaatgtattg gaggtgaatt tctaggacat 300gtgtttgaat atacaacaaa agaaggagac
gattatgatt taattgcaaa tacttattat 360gcaagtttga caactgttga gttattgaaa
aagttcaaca gctatgatcc aaatcatata 420cctgttaagg ctaagattaa tgtcactgta
atttgttcgt gtgggaatag ccagatttca 480aaagattttg gcttgtttgt tacctatcca
ctcaggtctg atgatactct tgcgaaaatt 540gcgaccaaag ctgatcttga tgaagggttg
ttacaaaatt tcaatcaaga tgccaatttc 600agcaaaggaa gtgggatagt gttcattcca
ggaagagatg aaaatggagt ttatgttccc 660ttgccctcta gaaaagcagg tcacttagct
agaagtttag ttgctgctgg aatatgtatt 720cgaggagtat gcatggttct gctattagca
atttgtatat atgttagata ctttcgcaag 780aagaatggag aagagtctaa attgccacca
gaagattcta tgtcaccttc aactaaagat 840ggtgataaag atagctatag tgatactaga
tccaaatata tattggtgga caaatcacca 900aagttctcat acaaagtact agccaatgct
acagagaact tcagtttggc taaaaaaatt 960ggtcaaggtg gttttggtga ggtctactat
ggagtgctgg gaggcaagaa agttgcaatc 1020aagaagatga agacgcaagc aacaagagaa
tttctttctg aactgaaagt cttaacaagt 1080gttcgtcact taaacctggt acacttgatt
ggatattgtg ttgagggatt tctatttctt 1140gtgtatgaat acatggaaaa tggaaactta
agccaacatc tacataattc agagaaagaa 1200ctgatgacct tgtctagaag gatgaaaatt
gctctggatg tagcaagagg ccttgagtat 1260attcatgatc attcggtgcc tgtatatatc
catcgtgaca taaaatcaga taatatttta 1320ttaaataaaa acttcaatgg gaagattgca
gattttggat taaccaagct gactaatatc 1380gcaaattcaa cagataacac taatcacatg
gcaggcacat ttggttacat gccgcccgaa 1440aatgcatacg ggcgtatttc tcgcaagatg
gatgtgtatg cttttggagt tgttctttat 1500gagctaattt ctgctaaagc agccgtgatc
atgatcgata aaaacgagtt cgagagccat 1560gagattaaga ccaatgaatc tactgatgag
tacaagagcc ttgtagcttt gtttgatgaa 1620gttatggatc aaaagggaga tcctatagaa
ggtctaagaa agctggtgga tccaaggcta 1680ggagataact attcaattga ttccattagc
aagatggcaa agcttgccaa ggcttgcata 1740aaccgagatc cgaaacaacg tccaaaaatg
agagatgttg tggtttctct tatgaaacta 1800atttctacta ttgatgatga aagtaggaca
gatagcgcag agttgagtct tgatgtggag 1860catgattcaa attga
1875741773DNAMedicago truncatula
74atggtttctt ccttttttca tacactcata ttcttttctg ccacccatat cctacttcaa
60ttgccacaag ccaatggaaa aaacttctca tgtactttga attcctctcc ttcatgtgac
120acatatgttg catactttgc taactctcca aactttttaa ccctcacagc catatctgat
180atatttgaca ccagtcctca atccattgca agagcaagca acataaaaga tgaaaatatg
240aacctcattc atggccaact tttgcttata cctataactt gtggttgcaa tggaaatgga
300aactactctt ttgccaatat ctcacactta atcaaagaaa gtgaaagtta ctattatctt
360tcgaccattt cgtatcagaa tctcactaat tggcagacag tggaagattc aaaccctaat
420ctgaatccat atttgttgaa aataggcacc aaaataaaca tccctttgtt ttgtaggtgc
480ccttcaaact attttgcaaa agggatagag tatctcatta cttatgtttg gcagcctaat
540gataatctta cacttgtagc ttccaagctt ggtgcatcac caaaggacat aatcacagca
600aatacaaaca actttggtca aaacttcact gttgcaatca accttccagt ttttattcct
660gtgaaaaact taccagctct ttctcaatca tactattctt caagtgaaag aaagagaatc
720aatcattttt ctattataat ttccattggt atatgtctag gatgcactat tctgatttca
780ctattattat tactttttta tgtttattgt ttaaggaaga gaaaggcttg tgagaataag
840tgtgtgcctt ctgtggagat aacagataag ttaatttccg aagtttcaaa ttatgtaagt
900aagccaacag tgtatgaagt tggtatgatt atgaaagcta ccatgaacct taatgaaatg
960tgtaagatag ggaaatcagt gtacaaagct aaaatagatg gtttggtttt ggctgtgaaa
1020aatgtaaagg gacatatcac agtcacagaa gagttgatga ttttacaaaa ggtaaatcat
1080gcaaatctag tgaaactagt aggtgtctct tcaggatatg atggaaatca ctttcttgta
1140tatgaatatg ctgaaaatgg atctctttat aactggttat tatctgagtt ttgcactctt
1200agttggagtc agaggttaag catagcagtt gatattgcaa tagggctgca atacttgcat
1260gaacacacac aaccatgtat agtccatagg aacatcaaat caagtaacat tcttcttgac
1320tcaaaattta aggccaagat agcaaatttt tctgttgcaa gaactacaaa gaatcctatg
1380ataacaaaag ttgatgtttt aggttatgga atggttctga tggagctaat aacaggaaag
1440aagtttttgt cgtatagcga acatagcgag gtgaatatgt tgtggaaaga ttttaagtgt
1500gtgtttgata cagaacaaaa gagagaagaa attgttagaa gatggatgga tcctaagtta
1560gggaggtttt ataatgttgt tgaagctctt agtttattca ctttggcagt gaattgcata
1620gaagaacaac ctttgttaag accaactatg ggagaagttg ttcttagtct ttctcttctc
1680actcaaccct ctcctacttt gttagaggtg tcttggactt atggattaga tgtagaggtt
1740gctgaaatgg ttactcccat catagctcgt tga
1773752313DNAMedicago truncatula 75atgaatacct tggatcttaa actacacctc
aaaattgaag tgcaactcat caaactcatg 60aagtttcttt atctttacat tctcttatgt
atgttacctt actcaatcaa ctgccaacag 120atattactta acaccactgt cactgattgc
tcaggcaccc cttcagcacc aaaaggatac 180ctatgcaata gtcctcaaaa ctcatgcaat
tcattcttaa ctttcaggtc aaaaccttct 240tatgacaacc ctacaagtat tgcttacctt
cttggctcag aagcatcgac catagcctca 300atcaacaaca tctcaagaaa tgaaaaactt
ccaaccaata aaacaatcat tgtccctatc 360ttgtgctcat gttctggaaa tatctaccaa
cataacactc cttacactgt ccaaaaaggt 420gatacatact ttcacttggt aaatgaaact
taccaaagcc tcacaacatg tcaagctttg 480aagggtcaga attattatgc ttctgaaaac
attgcaattg gtgccgaggt aacagttcca 540gtactttgtg cttgtcctac aacaacacag
atggcgaaag ggatcacttc cttgctggtt 600tacatagtga actatggtga aactgttaag
tctataggag aggcttatgg tgttgatgaa 660caaagcatat tagaggcaaa tgagttgcaa
ccaagtgaaa acagaagtgt gattctcttt 720gcgttgacac cgatattact tcctcttaga
ggcaagagct gcaaagagga tcctgatagt 780ttctactgta cttgttctca gggaaggcta
gcagatggaa gttgtaatga atcccatggt 840caaaagtttc ctgccaagtt ggttgctgca
ttaggtgttg gtattggtgc aggctttctg 900gtattgtttc tgcttagtta caggttatat
caatacataa agaaaaagag agcgagtatt 960cgtaaggaaa agttattcag gcaaaacggt
ggttacttgt tacaagagaa gttatcatca 1020tatggaaatg gagaaatggc aaagcttttt
acagcagagg agctccaaag agcaacagat 1080gactacaacc aaagtaggtt tcttggtcaa
ggcggctatg gcacagtgta caaaggaatg 1140ctaccagatg gaaccatagt cgcagttaaa
aagtcaaaac accttgacag aaaccaaata 1200gaaacgtttg tcaacgaagt agtcatctta
tctcagatca accacaggaa cattgtcaaa 1260cttttaggtt gttgtcttga gacagaaact
ccattacttg tctatgaata cattcacagc 1320ggaactcttt cccaacatat ccatgggaaa
gaccgtgatt catccctttc atgggaaagt 1380cgccttagaa ttgcatgtga ggttgccgga
gcagtgacat atatgcattt ctcagcttct 1440attccaatct tccatagaga cataaaacca
agtaacatac tcctagacaa taactatagt 1500gccaaagtgt ctgattttgg aacatctaga
tcaatcccat tagataagac tcatttaacc 1560acagcagtag gaggtacttt tgggtacatg
gatcctgaat attttcagtc aagtcaattt 1620acagataaga gtgatgtata tagtttcggc
gtggtgcttg ttgagcttat aaccggtaga 1680aagccaataa cattcaatga tgaagatgag
ggtcagaata tgacggcaca tttcatttca 1740gtgatgaagg aaaaccaact tcctcaaatt
ttagataacg cattggttaa tgaagcaagg 1800aaagatgaca ttctagctat tgcaaatctt
gcaatgagat gtttgagact taatggtaag 1860aaaagaccaa caatgaaaga ggtttcaatg
gaattagaag cactaagaaa ggaaatatct 1920accaacataa caccccctac actgtcaaaa
aaaggtgatt cctattatca cttggtaaat 1980ggtacttacc aaagaaagcc tcacgccatg
tcaggctacg aagggtcaga attactatgc 2040ttctgtgaaa acatttcaat tgatgttgag
ctcacatttg atcaaggaga cagagggttg 2100ggactcctta agttcttggt cagcagttgt
ggaagcagga tagagatatc tagaatcacc 2160attgatagag attcctgggt aattcttatc
aagattcaag attgtgtatc agagaactct 2220ggcttggaac ttttgttcca gccatttgag
tgtccgtttt cactatgtta tccccgtgaa 2280gcagtttctc acccacatgt gtcaaacaat
tag 2313762040DNAMedicago truncatula
76atggaacttt acaactacat tcttcacttc acagttctgt tcctatgcat gttttctcag
60ttattcaact gccagcaagt atacctcaat aacactgtct ttgactgcac taacccctct
120actgtaccaa aaggatacct ctgcaatggc cttaaaaaat catgtacttc ctttttagtt
180ttcaagtcaa aacctctcta tgataaccct acaaaaattg cctaccttct tcgctctgaa
240gcatcagcca tagcctcaat caataagatc cctttaaatg aaaagattcc ttcaaataaa
300tcaatcattg tcccagtctt ttgttcatgt gatggaaata tctaccagca tagtacatct
360tactctgtaa agcagaatga tacttactat gagttggtaa aagaaactta ccaaggcctt
420acaacctgcc aggcattgat gggtcagaac tactatgctc ctgtctcaat tcaacttgat
480gctgagctca cagttccaat actatgtgct tgtcccacag caaacctgac ggcaaaaggg
540gttacctcct tactcgttca catggtgaac tatggtgaaa cggttaagtc gataggagaa
600gcttatggtg ttgatgaaca tagtatgcga gaggccaatg agttgtcagg gctgcaaagt
660gcaaacagca gtgtgatcct ctttgcatcg acgcctatac tagttcctct gagacgcaag
720aattgtaaag agaactcaga tagattctat tgcaaatgtt ccgaggcact acatggagat
780gaaagctcca aggggatcta ctgtgacgaa tctcctagac gaaaagttcc tgcaaaattg
840gttgctgcct caggtatggg aattggtaca gtcttactgt gtttgtttct tttgagttgc
900aaattatatc aacacataaa gaaaaggaga gcaagtactc ataaggaaaa gttattcagg
960caaaacggtg gttacttgtt acaagagaag ttatcatctt atggaaatgg agaaatggca
1020aagcttttta cggcagaaga gctgcaaaga gcaacagata actacaaccg gagtagattt
1080cttggtcaag gtggctatgg cacggtgtac aaaggaatgt taccagatgg aaccatagtt
1140gcagttaaaa agtcaaaaga actagaaagg aaccagatag agacttttgt caatgaagtg
1200gtcatcttgt cacagatcaa ccacaggaat attgtcaaac tcttaggctg ttgtcttgaa
1260acagaaactc cattactagt ttatgaattt attcccaacg gaactctttc ccaacatata
1320catatgaaag accaagagtc ctccctttca tgggaaaacc gcctaagaat tgcatgtgaa
1380gttgctggag cagtggcata tatgcatttt tcggcttcta ttccaatctt ccacagagac
1440ataaaaccca caaacatact actagacagt aactttagtg ccaaggtatc tgattttggg
1500acatcaagat caataccact agataagact cacctaacca catttgtagg tggaacttac
1560gggtacatag atcctgaata tttccagtcc aatcaattca ctaataagag tgatgtatac
1620agtttcgggg tcgtgcttgt agagctcata accagtagaa agcctatttc attctatgat
1680gaagatgacg gtcagaattt gattgcacat ttcatttcag tgatgaagga gaaccaagtt
1740tctcaaatta tagatgctag actgcaaaag gaagcaggga aagacaccat tcttgccatt
1800tcaagtcttg caaggagatg cttgagactt aatcataaga aaagaccaac aatgaaagaa
1860gtttcagcag aattagaaac actaagaaag gcacaaagtt cttttgaaat caaccatgat
1920catgattctt catcaagtga tgaagaatca tttgggcacg gaattaatga atcaacagac
1980caagagtcca aagaggaaag caatttattt tccttacaaa tagaatccgc gtccttctga
2040771914DNAMedicago truncatula 77atgcatctct ttcccttcat cccttttttc
attttcaatc tcttcttctt cttcttcgtc 60agcaacatct ccatcatcct aggacaacaa
ccttatattg gtttagcaac aaatgcatgt 120ccacgaaaag gcgattcaaa atcaatccga
ggttacacat gtaacggcaa aactcaaaca 180tgccaagcat acctcacctt cagaactcaa
ccaatttact cctcagtttc aacaatatca 240tcattactag gctcaaatcc atctcaactc
gccgaaataa actccgtttc tttaaacgaa 300acattcgaaa caaacaaaat ggtaattgtt
cctgtcaatt gttcttgttc tggtaactat 360tatcaagcaa atacatccta tgtttttcaa
aacacagaca cttattttat agttgctaac 420aacacttttg aaggactttc aacatgtcaa
gctttgatgc atgaaaatca taatcctggt 480gatgtatatc ctggtagaaa actacttgtt
cctcttagat gtgcttgtcc tacaaagaat 540caaactcaga aaaacataaa gtatcttttg
agttatttgg ttgattgggg tgattctgtt 600tcattcatta gtgataaatt tggtgttaac
tttagaacca ctcttgaagc taatacactt 660tctttaacac aatctacaat ttatcccttt
acaacacttc ttgttcctct ttttgataag 720ccctcaagtt ctcaaattca aacacatcat
tctccttcat catccccacc tttatcttct 780tcttcttcga ccgacaaaaa atcgaaaaaa
acttgggttt atgttgttgt tggagtactt 840ggaggagttg taattgtagc tttaatatta
tttttatatg ctttcatttc cttcaaaaag 900ggtaaaaaga aaaatgattt tttggtgagt
gtttctgaaa gtactatttt tgaggaaaaa 960gagaaaccaa tgaagaaaga agatgaaaaa
ttgtctgaga ttatatgtgg tatagctcaa 1020tctttcaaag tgtatgattt tgaggaaatt
aaggttgcaa ctgataattt tagtccaagt 1080tgtcgggtta aaggaactgt ttatcgcggc
cttattaaag gcgatttggc tgcgattaag 1140aagacagaag gagatgtttc aaaagagata
caaattctaa acaaagttaa ccattccaat 1200gttattcgtc tttccggtgt tagcttcaat
caaggacatt ggtaccttgt ttatgagtat 1260gctgctaatg gagcattaag tgattggtta
ttttctaaca aaaaaatgga tgatggaaat 1320attctgagtt ggattcggag aataaagatt
gcattagatg tggcaatagg agttgaatat 1380cttcatagtt tcacttctcc tccacatatt
cataaggatc taaagtgtag taacatactt 1440cttgatagtg attttaaagc aaaggttgca
aatttaaggc atgtaaggtg tgtggaagaa 1500gttgaaaatg atgaagaatt tgttgctaca
agacatattg ttgggacaag aggttacatg 1560gctcctgagt atttggaaaa tggtcttgtt
tctacaaagc ttgatgtgta tgcatttggt 1620attttgatgt tggaaattat tacaggaaaa
gaggttggtt ttatgatatc aaaagataat 1680gagaatttgt tggatgtttt gagtggaata
cttggtgaga aaagtggtga tgagaagttg 1740aaggagttta tggatccttc attgcaagga
aattatccat ttgaacttgc tatgtttgtg 1800attgaaatta ttcagaattg tttaaacaag
gatccaggaa atagacctgc tatggatgag 1860attgtaccag ttttgtcaag aacattgaat
tcttcattga gttgggaaat gtaa 1914781788DNAMedicago truncatula
78atgtctgcct tctttcttcc ttctagttct catgctcttt ttcttgtcct catgttgttt
60tttctcacta acatctcagc tcaaccacta tatataagtg aaacaaactt tacatgccct
120gtggattctc ctccttcatg tgaaacctat gtagcataca gagctcagtc tccaaatttt
180ttgagcctat ctaacatatc tgatattttt aatttgagtc ctttacgcat tgcaaaagcc
240agtaacatag aagccgagga taagaagctg attccagatc aactgttact agtacctgta
300acttgtggtt gcactaaaaa tcactctttc gcgaatatca cctactcaat caagcaaggt
360gacaacttct tcatactttc aatcacttca taccagaatc tcaccaatta tcttgaattt
420aaaaatttca accccaatct aagtccaact ctattgccac tagacaccaa agtttcagtc
480cctttattct gcaagtgccc ttcaaagaat caattgaaca aaggaataaa gtatcttatt
540acttatgtgt ggcaggataa tgacaatgtt acccttgtta gttcaaagtt tggtgcatca
600caagttgaaa tgttagctga aaacaatcat aacttcactg cttcaaccaa ccgttcagtt
660ttgatccctg tgacaagttt accaaaactt gatcaaccat cttcaaatgg aagaaaaagc
720agcagtcaaa atctggctct tataattggt atcagcctag gaagtgcttt tttcatttta
780gttttaacac tatcacttgt gtacgtatat tgtctcaaaa tgaagagatt gaatagaagt
840acttcatcgt ccgagactgc agataagtta ctttcaggtg tttcaggtta tgtaagcaag
900ccaacaatgt atgaaattga tgcaatcatg gaaggtacaa cgaatctgag tgacaattgt
960aagattggtg aatcagttta caaggctaat atagatggta gagttttagc agtgaaaaaa
1020atcaaaaaag atgcttcaga ggagctgaaa attctgcaga aggtaaacca tggaaatctt
1080gtgaaactaa tgggtgtgtc ttctgacaat gatggaaact gttttctggt ctatgagtat
1140gctgaaaatg gatcacttga agagtggttg ttctcggaat cttcgaaaac ttcaaactcg
1200gtggtttcgc ttacatggtc tcagagaata accattgcca tggatgttgc aatcggtctg
1260caatacatgc atgaacatac ttatccacga ataatccaca gagacatcac aacaagtaat
1320atccttcttg gctcaaattt taaggccaag atagcgaatt tcgggatggc tagaacttca
1380accaactcaa tgatgccgaa aattgatgtt ttcgcttttg gggtggttct aatagagttg
1440ttaactggca agaaagctat gacaactaaa gaaaatggtg aggttgttat tctgtggaag
1500gatttttgga agatttttga tcttgaaggg aatagagaag agaggttaag aaaatggatg
1560gatcctaagt tagagagttt ttatcctata gataatgctc taagtttggc ttctttggca
1620gtaaattgca ctgctgataa atcattgtct agaccaacca ttgcagaaat tgttctttgt
1680ctttctcttc tcaaccaacc atcatctgaa ccaatgttag aaagatcctt gacatctggt
1740ttagatgctg aagctactca tgttgttact tctgtaatag ctcgttaa
1788791653DNAOryza sativa 79atgtccaagc atggtttcct gttcttcctc ctcttctttg
tgcaacacct tcacaccatc 60atctctttgc ttgattcacc tggaggaagc ttagcatcac
ttccagaatg gcagcccatg 120ccatgcaagt ctgtatctgt caacccctta tgcagctcat
acctctatgt caccccagaa 180gggcgcaatt tgtccaaagt agcctctgat ttcagtggga
atgcatctct tttccagcgg 240atcacacggc tctctggttc agaggacttg ctggtgaatg
tgccgtgtgt gtgcgaagcg 300atcaatgcca ccatgactgg tctcttccat gacactaact
acagggtgaa ggatggcgat 360atgggtgaca ttatcaacag caagactttc agtgggctcg
cgttgaatgt tggtgatggt 420cagatcctgc acaaagaaga gaagttgatc attcaccttc
catgtggatg ctcctcgaca 480gcgccagagg gagtgttgtc ttatgcagtt caggatggag
ataccttggg caacatcgca 540agcttgttca gatcgagttg gaaagatatc ttggatttga
acccaagagt cgcaaatcct 600gatttcataa agccaggatg gatcttgttt atcccgatgg
gagttgctgg tccttctaac 660aagaaaattg atccattcca aacagagagg cctgtgattt
tcagcttgag agcaattgaa 720gatgctacat ctaattttga tgagaagagg aagattggtg
agggaggata tggaagtgtc 780tacctcggtt tcataggaac acacgaaata gcagtcaaga
agatgaaagc aagcaaatca 840aaggagttct ttgctgagtt gaaagtgctg tgcaaaatac
atcacataaa tgtggttgaa 900ttgattggtt atgctgctgg ggatgatcat ctttacctcg
tttatgagta tgtccagaat 960ggatcgctta gtgaacatct ccatgatcct ttgctgaaag
gccaccaacc tctatcatgg 1020actgcaagaa cacagatagc aatggactct gcacgtggta
ttgaatacat ccatgatcat 1080acaaagacct gctatgtgca ccgtgacatc aaaaccagca
atattctgct agacaatggt 1140ctacgagcta aagttgcaga tttcggcttg gtcaagctcg
ttcagcgtag tgatgaagat 1200gaatgtctgg caactcgttt ggttggaacg ccaggctacc
ttccaccaga atcagttctt 1260gagcttcaca tgactaccaa atccgatgtg tatgcattcg
gagtagttct tgcagagctt 1320attactggtc tccgtgcact tgtacgggac aacaaggaag
ctaacaagac aaaatctcta 1380atctcaatca tgaggaaagc attcaaacca gaagatctgg
agagttccct ggagacaatt 1440gtagatccct acctgaagga caattacccc atagaagaag
tttgcaagtt ggcaaacatt 1500tcgatgtggt gcctcagtga ggatccactg caccgccctg
aaatgaggga ggttatgccg 1560attttggctc aaattcatat ggcttccata gagtgggaag
catcactcgg aggcgatggt 1620gaagtcttca gtggcgtttc caatggtaga tga
1653802004DNAOryza sativa 80atgtcgatgc tgccgcgctt
cctcctcctc gtcgtgctgc tcgcggtgcc gatgacggcg 60gcgcagcagc agtacgaggc
caacgcgcag ggcgactgct acaccgacaa cggcagctcc 120gtcctcggct acacctgcgg
caccgccgcc tccccgccgc cgccgccatg cacggcctac 180ctcaccttcc gctccgcccc
accgagctac gcctccccga tcaccgtctc ctacctcctc 240aacgccagcg tccccgccgt
cgccgccgcc aactccgtcc cggtctcgcc tccggtcgcc 300cgcgacggcc tcctcctcgt
ccccgtcccc tgcgcctgca ccgccgccgg gtactaccag 360cacgacgccg gctacgtcat
ccagttcgac gacgagacct acttcgtcat ggcgaacgac 420acgtaccagg ggctcacgac
gtgccaggcg ctcatggcgc agaacccggc gcacgacagc 480ctcgacctct acccgggaat
cagactcacc gtgccgctgc gctgcgcgtg cccgtcgccg 540gcgcaggccg ccgccggcgt
gaggtaccta gtgacgtacc tcctcggctg ggacgacgac 600tcgtccaccg tcgccgaccg
cttcggcgcc gactaccagg ccgttctctt cgccaacaac 660ctcaccgacg actccacggt
gtatcccttc accacgatgc ttgtcccgct caagcatcgg 720cccaagcccg acgtgacggt
cttgccggag ccggggcctc cttctccggc tccggctccg 780gcagtgtcag ctccgccgcc
gccggcggtg ccttccagcg aatcgggcag tgggaggtgg 840aagaaatcct ttcgcggtag
gtgtatcggc attggagttg gtgttgggtt cgccgttctt 900gcgtctggtg ctctgcttgc
tctgttcttg ctacggcgtc ggtggcggtg gcgtggcaac 960ggcgagcttc acgatgtgcc
gttggctccg gataaggagg gagcgaaggc gacgccgccg 1020ccgtggatgc tgccgacgac
ggtggcggac gtcgacgtgc gcgatgccgt gggatcgatg 1080gccgtgtacg agtacggcga
gctggagcgg gtgacggccg ggttcgcgga ggagcggcgc 1140atcggggact cgtcggtcta
ccgcgcggtg atcaacggcg acgtcgcggc ggcggtgaag 1200cgcgtcgccg gcgacgtggg
cgccgaggtg agcgtcctgg gacgcgtcag ccactcgtgc 1260ctcgtccgcc tgttcggcct
ctgcgtgcac cgcggcgaca cgtacctggt gttcgagctc 1320gccgagaacg gcgcgctcag
cgactggatc cgcggcgaca acggcggccg cgccctctcg 1380tggaggcagc ggatgcaggc
ggcgctcgac gtggccgacg ggctcaacta cctccacaac 1440tacaccaggc cgccctacgt
gcacaagaac ctcaagagca gcaacgtcct cctcgacgcc 1500gacttccgcg ccaaggtctc
caacttcggc ctggcgcgca ccgtcgccgg cgccggcggg 1560cagatgacgt cgcgcgtcgt
gggtacgcag gggtacatgg cgccggagta cctggagcac 1620gggctgatcg ggcctcacct
cgacgtgttc gccttcggtg tcgtcctgct cgagctcctg 1680tccgggaagg aggcagcgcc
tgcgcgagac ggcggcgagg gtggcgatgg cgaagcattg 1740gcgttgctgc tgtgggagga
agcggagggg cagctggtgg tcgacggcga cgatgacgac 1800gcgcggggca aggtggcggc
gttcatggac tcccggttgc gtggcgacta cccgtcggag 1860gtggctctcg cgatggctgc
gctggcgctg cggtgcgtcg cgcgggagcc ccgcgcgcgg 1920ccgtcgatgg tggaggtgtt
cctctcgctc tcggcgttgc acggcacaac gttggattgg 1980gccccccatg caaccctgag
ttga 2004811893DNAOryza sativa
81atgccgcccg gcgcgcgagc aagacgcatc gccgcactcg acctgatctt cctcgtgttt
60ctccggctcg ccagcgcgta tcaacgacca tccgacgcgc atatcaactg catcgacagc
120ttctgtctcg gcggctacac ctgcagcgaa accaccgcca ccacgtcctg caccgcctac
180ctcaccttcc gctccgaccc cccgctctcc gtcgcctacc tcctcaacgc gacgccctcc
240gccgtcgccg ccgccaactc tgtccccctc gccgtctccc ccgtcgacgg cacgcagctc
300ctcctcgtcc ctgtcccctg ctcctgcaac cgggccaccg gctactacca acacaacacc
360acctacgcca ttcaggaact tgataccttc ttcctcatcg ccaataacac gttccagggg
420ctcacgacgt accaaagcat catcgccaat aaccctgcaa gcgaagctat gtctccagtg
480atcaacggcc ccctcgccgt gccgctccgc tgcgcgtgcc cctctgcgac tacgggacga
540ataaacaatc tgctgacgta cgtggtccag gagggggaca acgtgaccag catcgcgcgc
600agattcaact ctacgcacgg cgacgtgctt gccgccaaca cgttgttggt cccgctcgtc
660cacccgccgc attcgcgagt ggtgctcgcg aatactacta ttacttccac gactccgccg
720gaatctcaga aattttacgt gtcgagtccc tgcagcaatg gcttgctggc tggcttgggt
780atcggcgtcg gatgcggcgt ctctgcgtgg gctgccgttc ttgctgtgtt cttgctatgg
840cgtcgaaggc ggcggcgccc cgtcggcgac agctcaggca tggccaggga gacccccctg
900gtagcggctg tgcgtggcgc ggtggagact ctggctgcct acagctacgc ggacatcgag
960acggcgacgg cggggttcgc ggaggagcgg agagtggccg ccggatcgtc ggtgtaccgc
1020gcggtgatca acggcgaggc tttcgcggtg aaacgcgtgg ccgccggcgg cgacgacgtg
1080cgcggtgagg tcgacgtcct cggccgcgtc aaccactccg gcctcgtccg gctccggggc
1140ctgtgcgcga acggagacga cacctacctg gtgctcgagt tcgccgagaa cggcgcgctc
1200agcgagtggc ttcaccccgg cagcgcagcc gcgtgtctcc gccgtgtcct cggctggaag
1260cagcgcgttc tggtggcgct cgacgtcgcg ggtgggctca actacctgca ccacttcacc
1320aaccctccct acgtgcacaa gaatctcaac agcggcaacg tcctcctcga cgcgaacctc
1380cgcgcgaagg tctcgagcct cgggttcgcg cgcgccgtgg ccgtcgccgt cgcagccgga
1440gacgacagca tcgccctgat gacacaccac gtcgtgggca cccacggtta cctggcgccg
1500gagtacctgg agcacggcct gatcagtccc aagctcgacg tgttcagctt cggtgtcatc
1560cagctcgagc tcttgtccgg gaagacggcg gcgtttgtca ccgacgatga cgggcagaac
1620atgctgctgt ggcaggcggc ggacgggctc gtcgacggcg atggcgcatg gttcaagctg
1680agggcgttca tggaccctca gctgcaaggc cactacccga tcggtgtcgc gtccgcggtg
1740gccgcgctgg ccgttcggtg cgtggcgcgg gagccacggg cgcggccttc catggaggag
1800gtgttcgtca cgctctcagc ggtgtacaac ctcacggttg attgggatcc tcagaattac
1860agcgcatcag cttctatggt cctcggtagg tag
1893822070DNAOryza sativa 82atggcggaaa ctccacgccg aatccgccgc ggcggcgccg
ggctctcgct cgtcgtcgtc 60gccctcctcc tcgccgtcgc ggcgagggcg cagcaggagt
acgaggcgaa ccagcagaac 120gcctgctacg ccaccaacgc gagctccacc ctcggctaca
cctgcaacgc cacctccgcc 180tccgccgccg ccgcggcgcc ctgcgacgcg tacctggtgt
tccgctcctc cccgcccctc 240tacgcctccg ctgtctccat ctcctacctc ctcaacgtcg
ccgccgccgc cgtcgccgac 300tccaacgccg tcgacccggt cgcccccgtc gccgccgacc
gcctcgtgct cgcgcccgtg 360ccgtgcggct gctccccggg cggctactac cagcacaacg
cctcccacac catccgcgac 420accggcgtcg agacctactt catcatcgcc aacctcacct
accagggcct ctcgacgtgc 480caggctctca tcgcgcagaa tcccctccac gacagccgcg
gcctcgtcgc cggcgacaac 540ctcaccgtcc cgctccgctg cgcctgcccc tcgccgccgc
aggccgccgc gggcgtcaag 600cacatggtca cctacctcgt cacctggggc gacaccgtct
ccgccatcgc cgcgcggttc 660cgcgtcgacg cgcaggaggt gctcgacgcc aacacgctca
ccgagagctc catcatatac 720cccttcacca cgctgctggt cccgctcaag aacgcgccca
cgccggacat gctcgcgccg 780ccggcccagg cgccgccgcc ccccgcgccc gcgccgccgc
gggcgcagcc gccgcctggt 840gggtcgggca gcgggaaggg agtcgccgtt ggggtaggcg
tcggttgcgg cgttcttgcg 900ttggcgggtg tctttggctt gctgttcttc tgtctccggc
ggcgacgcgg cgttggggaa 960gaaagtgttc gtccggggaa ggtggtcggc gacgtgtcct
cttcggcgga gtacggcgcc 1020ctggcatcgg ggaagcagac gacgacggca acctcgatgt
cgtcgctgtc cgcggcgagg 1080tcgctgatgg cgagcgaagt gcgcgaggcg ttggagtcgc
taacggtgta caagtactcg 1140gagctggaga aggccacggc ggggttctcg gaggagcgga
gggtgcccgg caccgccgtg 1200taccgcggcg tgttcaacgg cgatgcggcg gcggtgaagc
gtgtttccgg cgacgtgagc 1260ggcgaggtcg gcatcctgaa gcgcgtgaac cactgcagcc
tgatccgcct ctccggcctc 1320tgcgtccacc gcggcgacac ctacctcgtc ttcgagtacg
ccgagaacgg cgcgctcagc 1380gactggctcc acggcggcga cgccgccacc ggcgtcctgg
ggtggaggca gcgcgtgcag 1440gtggcgttcg acgtcgccga cgggctcaac tacctccacc
actacaccaa cccgccatgt 1500gtgcacaaga acatcaagag cagcaacatc ctcctcgacg
ccgacctcca cggcaagatg 1560tccagcttcg ggctagcccg cgcgctcccc gcgggcgacg
gcgccgccgc cgccgccgcg 1620cagctgacgc gccacgtcgt gggcacccag ggctacctct
ccccggagta cctcgagcac 1680ggcctcatca cgcccaagct cgacgtgttc gccttcggcg
tcgtcctcct cgagctcctc 1740tccggcaagg aggcggcgtc ctccggcgac ggcgagaacg
gcgaggcgct gctgctgtgg 1800gagtccgcgg cggaggcgct cgtcgacggc ggcggcgagg
acgcggggag caacgtgcgc 1860gcgttcatgg acccgcggct tggcggcgac tacccgctcg
acctggccat ggccgtggcg 1920tcgctggcgg cgcggtgcgt ggcgaggcag cccgccgcgc
ggccggccat ggacgaggtg 1980ttcgtctcgc tcgcggcggt gtacggctcc acggtggact
ggaacccctc ggatcacggc 2040aactccgggt cgtcgctgat tgggaggtag
2070831875DNAOryza sativa 83atggaacaca agggtttgtg
catcctcgcc gtcgtcatcg ccttccagct cgccggcggg 60gaggccgtca ccgatgccac
tgcccgggca cgtcgcttcg cctgtaacgt gtcggcgccg 120tgcgacacgt tcgtcgtgta
ccggacgcag tcgccggggt tcctcgacct cggcaacatc 180tcggacctgt tcggcgtgag
ccgggcgctg atcgccagcg ccaacaagct gaccaccgag 240gacggggtgc tcctgccggg
gcagccgctg ctcgtgccgg tcaagtgcgg ctgcacgggc 300gcgcgctcct tcgccaacgt
cacgtacccc atccggcctc gcgacacctt cttcgggctc 360gccgtcaccg cgttcgagaa
cctcaccgac ttcgtcctcg tcgaggagct caacccggcg 420gcggaggcga ccaggctgga
gccgtggcag gaggtcgtcg tgccgctctt ctgccggtgc 480ccgacgcggg aggagctcag
cgccgggtca cggctcctcg tcacctacgt gtggcagccc 540ggggacgacg tgtccgtggt
gagcgcgctg atgaacgcct ccgctgccaa catcgccgcg 600tcgaacggcg tcgcgggcaa
ctccaccttc gcgacggggc agcccgtgct gatcccggtg 660tcgcagccgc cgcgttttcc
tccgctgacc tacggtgcca tcgccgccga tcccggagcg 720ggcaagcacc gccacggcat
catcgtggcg acgagcatcg cggggtcttt cgtcgcgtgc 780gccgtgctgt gcacggcgat
cttggcgtac cggaggtacc gcaagaaggc gccggtgcca 840aagcacgtca gcccgaagct
ttcttggacc aagagcctga acagattcga cagcaatagc 900tccattgctc gcatgatcaa
tggaggggac aagctgctca ccagcgtgtc gcagttcatc 960gacaaaccga tcatctttag
agaggaggaa atcatggaag cgacgatgaa cttggacgaa 1020cagtgcaagc tcggcagctc
gtattaccgc gcgaaccttg aaagggaggt gttcgcggtg 1080aagccggcga aaggcaacgt
tgccggggag ctgaggatga tgcagatggt gaaccacgcc 1140aacctgacca agctggccgg
catatccatc ggcgcggacg gcgactacgc cttcctcgtg 1200tacgagttcg ccgagaaggg
ctcgcttgac aagtggctgt accagaagcc gccgtgctcg 1260cagccgtcgt cgagctccgt
ggcaactctg tcgtgggacc agaggctggg catcgcgctg 1320gacgtcgcga acggcttgct
ctacctgcac gagcacacgc agccgagcat ggtgcacggc 1380gacgtccgtg cccggaacat
cctcctcacc gcgggcttca gggcgaagct gtccaacttc 1440tccctggcca agccggccgc
catggtcgac gcggcggcga cgagcagcga cgtgttcgcg 1500ttcgggctgc tcctcctcga
gctcctctcc gggaggaggg cggtggaggc gcgcgtcggg 1560gtggagatcg gcatgctgcg
gacggagatc cgcaccgtgc tggacgccgg cggggacaag 1620agggcggcga agctgaggaa
gtggatggac ccgaccctcg gcggtgagta cggcgtggac 1680gcggcgctca gcttggccgg
catggcgagg gcgtgcaccg aggaggacgc ggcgcggcgg 1740cccaagatgg ccgagatcgc
gttcagcctc tcggtgctcg gacagccgct gtccgtctcc 1800gacgcgttcg agaggctatg
gcagcccagc tcggaggaca gcatcgggat tgggaacgag 1860gtggcagcta gatag
1875842055DNAOryza sativa
84atggcgcaat cgcgttcaca cgccatggcc gcgttcgcct tcgtcgtcgt cttcctcctc
60tccggcgcgc cgggagcccg gtcgcagcag ccgtacggct cgcaggtggc cgactgcccc
120aacaagcaca acgacaccgg cctcctgggc tacttctgca gcagcggcgg cggcggcggc
180ggcgcgccgt cgtcgtcgtc gtcgtcgtgc cagacctacc tcaccttcca cgccacgccg
240cgctaccccg acctcgccgc catcgcgtcg ctcctcggcg ccgacgcgtc cagcctcgcc
300gccgcgaact ccgcggcgtt gcccaccgcg gcgctcgcgc cgggcgccaa ggtgctcgtc
360ccggcgacct gctcctgcac gggcgccgcc tactaccagc ggaacgcgac gtacgtggcg
420gtcgccggcg acacgctgct ggtgatcgcg aacgacacgt tccagggcct gtccacgtgc
480caggcggtgc aggagcaggc cctcggcgac gcgccggcga ggtcgctcct ggcggggcag
540cgcgtcaccg tgccgctccg gtgcgcgtgc ccgagcgccg cccaggccgc cgccggcgtg
600aggtacctgg tgacgtacct ggtcgacgag ttcgacgagg tgggcgccat cgctgcgagg
660ttcggcgtcg acgccgggaa catctcggcg gccaacgaga tggccattac tgacaccata
720taccctttca caacgctgct catccccgtc aagtccaagc ccgacgtgtc gcagctccgg
780tcgccgccgc cgccgcctcc tccgccgccg gcggcgcctg ctccgaccac gaaccgcaag
840aaccacaccg gagtctacgt cggcatcggt gcggccgccg tggctgttct cgccgtggtc
900accgccgttg tggctgccct tgctgtgagg gcgaggaggc agcgacggcg agccaccgcc
960gctgtcgccg ccgccggagg caagggcggt aaaggcaacg acaaggcgtc gccggcgttc
1020accggcggcg aggtgtccgt gtcgatcagc gaggcgttct cgggcctctc cgacatcaag
1080tcctccctga aggtgttcac ctacgcggag ctcgcggcgg cgaccgacgg cttcagcccg
1140gaccgccgcg tcggcgggtc ggtgtaccgc gccgtgttca acggcgacgc ggccgccgtg
1200gaggtcgtgg accgggacgt gtcggcggag gtggagatca tgaggaagat caaccacctc
1260aaccttgtca ggctcatcgg cctctgccac caccgcggcc ggtggtacct cgtctcggag
1320tacgccgagc acggcacgct ccgcgaccgc ctcctcgccg gcggcggcgc gccgccgctg
1380agctggtcgc agcgcgtgca ggtggccctc gacgtcgccg aggggctccg ctacctgcac
1440gggtacacgc ggccgccgta cgtgcacatg gacgtcagca gcgacagcgt cctcctcgcc
1500ggcggcgccg acctccgcgg caagctccgc aacttcggcg gcgccagggt catccgcggc
1560ggcggcggcg aggcgttcac gatgacgagc aacatcgccg ggacgcgcgg gtacacggcg
1620ccggagtacc tggagcacgg cgtcgtgtcg cccaaggccg acgtgtactc cctcggcgtc
1680gtgctcctgg agctcgtcac cggcaagggc gtcgacgagc tggaggccga cggcgccggc
1740gacccgttcg ccggcatgaa cgcgctggcc ggagacctcg acggcggcag cgaagacgac
1800gcggcggtga cgaggagaat ggaggagttc ttggacccgg cgatggcggc gaccgggagc
1860agctgcccgc gcgaggccgt ggcgatgatg gtgaagctga tcgagaggtg cgtccgccgc
1920gacgcggcgg cgcggccggg aatgggggag gtggcgcagc atctgctgat gttgcacggc
1980gtctccggcg acggctggca cagctcgctg gagcactacc ggagctccgg cggcgacggc
2040ggcgaacagc catag
2055851680DNAPopulus sp. 85atgacttccc acaatttcct tcttggtctc ctgatcactt
taatggcaac cctagtagct 60ttaggttttg cctctgatga ctccaccaat tcaccaaata
tgtacccatt tacttgctct 120gattctgaag acatcaaaat ttgtaatgcc tcactgtacc
ataccaacta cgacagctta 180cagaaagaac agcttgcttc catttatggt gtcagcccag
ctcagatcat ttctatttcc 240tatgcaagcc gacaagacta tcttgtaacg gtaccttgct
cgtgcaaaaa tatcaatggc 300actgttggat acttctatga tgcaatccac aacgtaagtc
aaggagaact gttttctaat 360gtttcagctc agatttttaa tgggcaagct tggtgggttg
aagatgaagc gtggttgttt 420aatccaagaa ataatttctc catgcatctc ttgtgtgggt
gtacaaaaag caaatcccaa 480attgtggtaa catatactgt tcagcagcat gacacattat
cagatatttc aaccaggcta 540tcatctacag taggtggcat acagagcatg aacataaatc
tgattaagaa cccaagttcc 600ataaatgtgg attgggtgct gtttgttccc atggacagta
aaccggcctc tggaaaagga 660tcaggaaggg gacacaaatg gataataatc gtcggcatac
tatcttctct gacagcactt 720tcaatcatca cattggtaat cctccttctt tggagaaaag
gatgccgaca gaacagcgag 780gaggacttga aacctgtacc caaaagcatg agcacgaaca
gagcattttc cttgcagagt 840ctgtacaagg gaagtaacga agatgggaca gcttttgaat
cagaaaaacc agtcgtcttt 900agtctagaag agattgaaga ggctacaaaa agctttgaca
aaacgaagaa aatcggagag 960ggtggatatg gatgtgtgta ccatggatta ttgcgaggac
aggaggttgc agtaaagaag 1020atgagatcaa acaaatccca tgagtttttt gcagagctca
aggtcttatg taagatccat 1080cacataaatg tggtggaact tctgggttat gccggtggag
atgatcactt ctacttgatt 1140tatgagtatg ttcgaaatgg gtcacttagt gatcaccttc
acgatccatt gctaaaaggt 1200caccagccac tctcttggac agcaagagca cagattgccc
ttgatgctgc aaagggaatt 1260gaatacatcc atgaccacac caaagcacgc tatgtgcaca
gagatataaa gacaagtaac 1320attctactcg atgagggact gcgagcaaag gttgcagatt
ttggattggc aaagcttgtt 1380gaacgaacga gtgatgaaga tttaatagca acacgactgg
tctacaaaat ttttgaagat 1440aaagatccag agactgcttt agcagatgtc atcgacaaaa
atctccgcaa cagctacccc 1500atggaggatg tctacaagat ggcagaaatt gctgactggt
gtttgagcga agaagcaatt 1560aatagaccag agatgcgaga agtagttcaa aaattatcta
tgatcgtagt gtcctcagta 1620gagtgggagg cttcactggg agggagcagc atatttagtg
gggtttttaa tggaagatga 1680861797DNAPopulus sp. 86atggcaattt ctttgctctc
ttcctttttc actcaagctc tatttttctt ggttcttgta 60tttttctcca cctatgtcac
agcacaagct ccacccggca caaatttttc atgccctgtt 120gattcaccga cctcatgtcc
tacttatatt agttaccttg cccagccacc agattttttg 180gacctcggaa agatttctca
tctatttggg atcagtcgta cgctaatagc aagtgccagt 240aacctggtgt ctgaggacac
cccattattt ccaaatcaac tcttgcttgt acctatacgt 300tgtggttgca ctggtagcca
atcttttgtc aatatcacct accaaatcca gcaaggtgat 360agcatctact cggtttcaac
tatttcattt gagaacctca cccgttggca agaggtggaa 420gctttgaacc gcagtctgac
tccaaccctc ttgcatgctg gtgacgaggt tatatttcct 480ctgttctgta agtgcccttc
aaggactcat ttggagaatg ggattgaaca tctcattact 540tatgtgtggc aacctggtga
tgatctcaag aaagtagctg ctatgcttaa tgcatctgaa 600cgtaatattg tgattgaaaa
caactatgat aactttaatg ctgcagtcta taacccgata 660gtgatccctg tgtccaagtt
accagttctc tctcaaccat acctcactcc tgaaagaaga 720ggatctaagc atctttggat
tgtcattgtt gctgcaagca tagcaagcac tttttttacc 780tgtcctttgg ttgcttttct
gatccacaag cgttgttcat ataaggcaac caaagctttg 840gatcgtactg gttcctgttt
ggagaccagc gatctgcttc caggggtttt aggttgtctg 900gacaagtcaa tcatttatga
ggttaaagca attatggagg gaaccatgga tctacacgaa 960cactacaaga taggaggatc
agtatatagg gccaacatca atggctgtgt cttagcagtg 1020aagaaaacca aggatgatgt
cacggaagag ctaaagattt tgcagaaagt cagtcatgca 1080aatctggtga aactaatggg
gatgtcatct gaatctgata gagaaggcaa tcgcttcttg 1140gtctatgaat atgcagaaaa
cgggtcactg gacaagtggt tgcatcccaa gtctgaatct 1200tcctcaagct ccgtgggttt
cctcacttgg aagcaaagga tgcaagtagc actagatgtg 1260gccaacggcc tgcaatactt
gcatgaacat actcaaccaa gaaccgttca caaggatatt 1320cgaacgagta atattcttct
cgattccaca tttcgggcaa agatagcaaa tttctcaatg 1380gctagagctg ccacagactc
gatgatgcct aaagatgatg tatttgattt tggagttgtt 1440ttgttagagt tgctttctgg
aaagaaggcc atggtaacca aagaaaaagg cgagattgtt 1500ctgttgtgca gggaaataaa
agatgtcttg gaaatggaag agaaaagaga ggagaggtta 1560agaaagtgga tggatcccaa
cttggagaga ttttatccca ttgatagtgc catgagcttg 1620gcaaccttgg ctaggttgtg
cacactggaa aagtcttcag aaaggccaag catggcagag 1680attgtcttta acctcacagt
tctcacccag tcatctccag agacattaga aaggtggact 1740tctgaggtgg aaacggaaga
ttttactcgg ctcgtcagcc ctgtcacagc tcgttga 1797871965DNAPopulus sp.
87atggactttc ttctactgta tctgtatgtg gtgctgctgc tctcaccagc actggtacaa
60gggcaacaaa cctatgtagc caaccaccaa ctggactgct acaacaatgc cttcaatgaa
120acgaccaagg gatttctgtg caacggagtt caatcatcat gccaatccta cctcaccttc
180cgatctatgc ctccctacaa ttctcctgtc cttattgcct atctcttagg cgtcccacaa
240tccgccactc gcatcgcctc catcaataac ctctcctctg acactgccac tattcccacc
300aacacccagg tcgtggttcc agtcaactgc tcctgttatg cccgtcaata ttaccagcac
360aattccacct atcagctcaa ggacaaatcc gaaacctact tcagcgtggc caacaacacc
420taccaaggcc tcaccacatg ccagtccttg atgtctcaga atccctacgg cgatcggaat
480ctgtcactcg gtctcactct ccaaataccc ctgaggtgtg cttgcccaac ttccaaccag
540aatgcttcag ggatcaacca cttgctcacc tacatggtca cttggggcga ttccatctct
600tccattgccc agctgtttgg cgttgacaag cagagggtac ttgatgcaaa caagctgtct
660tcctccaata tcattttccc cttcactccg attctggttc ctctgcccac tgagcccacc
720aaaattgaac agccatcggc agcccctccg cctgccgcac catctccgca gactcccaat
780gtttccgttg ggggctcttc cgatcacaaa gccttgtatg tgggtgttgg gataggagct
840gctttcctca ttcttttatt tgctgcgttt ggatttctgt tttggcaccg caaatctcgt
900aagcaacaga agcctgtctc cacttcagaa cccgaaacgc tgccatcagt ctctactgat
960ttcactgtac tcccagtctc caacaacaaa tcttggtctc tctcttctca tgacgcccga
1020tatgctattg agtccttgac tgtctacaaa tacgaggact tacaagtggc caccgggtac
1080ttcgcccaag ctaatctgat caagggctcg gtttataggg gatctttcaa gggtgacaca
1140gccgcagtca aggttgtgaa aggagatgtc tccagcgaga tcaacatttt gaagatgatc
1200aatcactcca acgtcatcag gctctctggt ttctgcttac atgagggcaa cacttacctt
1260gtttatgagt acgccgacaa tggctctctc actgattggc ttcactctaa caacatatac
1320cgaattcttg cttggaagca gagagttcgg attgcctacg atgtggctga tgccctcaat
1380taccttcaca actacaccaa cccgtcctat atccataaga acttgaagac cagcaacatt
1440cttttggatg ccaacttgag agccaaggtt gctaatttcg gcttggcaag aacactggaa
1500aatggccaag atggtggact gcaactgaca agacatgtgg taggcactca aggttatttg
1560gcccccgagt acatcgagaa tggagttatc accccgaaat tagatgtttt tgctttcggg
1620gttgtgatgt tggagctctt atctgggaag gaagcagctg ctacagctat tgacaagatt
1680gcaggagacg acttgctctc tgtaatgata atgcgtgtgc ttgaggggga caatgtgaga
1740gagaaactct ctgccttcct ggacccttgc ctaagagacg agtacccctt ggatctagct
1800ttctcaatgg cccaactggc taaaagctgt gtcgagcatg atctcaatac acgaccttca
1860atgcctcaag ttttcatgat gttgtccaag atcctctcgt cttcgttgga ctgggatcca
1920tccgatgagc tcaatcgatc taggtctata gacagtggca ggtag
1965881575DNAPopulus sp. 88atgttcttgc aacaccatca tattagcact agcatttcct
ctgcaaacat cacatatacc 60atcgaggcag gcaacacttt ctacattgtc tcgactaaat
acttccaaaa ccttacgacc 120taccagtctg ttgagctttt caaccctaca cttatccccg
aactactcga cataggagta 180gaggtgatct ttccaatatt ttgcaagtgt cctcatcaaa
cccagttgca aaacaaggtg 240aattatctgg tatcttatgt gtttcagcct tctgataact
tatcttcagt tgcttcaaca 300tttggagttg aaacacaatc tattgttgat gttaatggca
ataacatcca gccttatgat 360accatattcg ttccagtcaa tcaacttcca caactggcac
aacctacagt agttgttcct 420tctggggcgc cgccccctga gaagacagag aggaaaggtg
tgattatagg attagcagtt 480gggctaggaa ttgctgggct tttactggtc ttggtaagtg
gggtttggtt ttatagagag 540ggtgtgttga agaagagaag agatgttgag aaagttgagg
agaagcgtag gatgcagttg 600aatgggggaa gtaaagggtt gaaggatata gaagtgagtt
tgatggcaga tgtttcagat 660tgcttggata agtacagggt ctttaagatt gatgaactga
aagaagctac taacgggttc 720agtgaaaatt gcttgattga aggatctgtg tttaaggggt
ccataaatgg agagacctat 780gccatcaaga agatgaagtg gaatgcctgt gaggagctca
agatattgca gaaggtaaac 840catggcaact tggtgaagct agagggcttt tgcatagacc
ccgaggatgc aaattgctat 900ctggtctatg agttcgtaga cagtggctct ctacattcgt
ggttgcatcg caacgagaaa 960gaaaaactaa gctggaaaac aaggttacgc gttgcaattg
atgttgcaaa tggtctccaa 1020tacatccatg agcacactag gccaagggtt gtgcacaaag
acattaaaag cagcaatatc 1080ctcttggact ccagcatgag agccaaaatt gccaactttg
gactagcaaa aacaggctgc 1140aacgccataa caatgcacat tgttggcact caaggctaca
ttgcgcctga atatttagct 1200gatggtgtgg tgtcaacgag aatggatgtt ttctcttttg
gtgtggtttt gcttgagcta 1260atctcaggaa gagaagcaat tgatgaagaa ggcaaggttt
tgtgggcaga agctattgga 1320gttctggaag gaaatgttga agagaggagg aaggtaaaga
gattgacagc atggatggac 1380aaggttcttt tagaggagtc atgctcaatg gagagtgtaa
tgaatacaat ggctgttgca 1440attgcttgct tgcatagaga tccatcaaag aggcctagca
tggtggatat tgtctatgcc 1500ttgtgcaaga gcgatgattt gttttttgac atctcagaag
acggattgtc aaaccctcag 1560gtaatggcaa gataa
1575891830DNAPopulus sp. 89atggcaaact tctttctcag
tttgctcact ctcagttttc tcacttcata tgctaatgct 60caacaaaact actcgaagga
ttcagcacta gactgcaacg ccaatgatga tgcaggacca 120tcctctgcat ttctttacac
ctacaatggc caggaccaat cttgccaggc ctttctgatc 180ttcaaatctc aaccttcttt
taactcggtg ccttcaatct cagcccttac gtcggcaaac 240caagaagagc tggcaagaat
caacaatgtt acaaggctat cagagttccc taccaacaat 300gaggtgattg ttcctgtaaa
ttgtttttgt tttggccaat attaccaggc caacactaca 360attcaggtca cgactactcg
tggaacctac tatgtcatag cgaatgaaac ttacgaggga 420ttatctactt gtgctgcgct
taagcatttg aatatacatg gtgaatatga tttgttgcct 480ggtgaggaac tgcaagtacc
acttcgatgt gcttgcccta caacgaatca aatgataaga 540ggaacaaagt atctagtgac
ttaccctctt agctcagatg ataacattcc tgatattgct 600gacagattca aagtaagcac
caaggatata cttgatgcaa atggtatgga agaaaatcca 660actctttatc ctgatacaac
aattctcata cctctgccta ctcaacctac aagttcacaa 720acaataattc acagcaaccc
aaacatttct cctccatcag ctttgagtcc tagaaataga 780ggatcaaaga aaaaacatta
tgagtctgct gggcttgctg cagcttgctc tttgctagtc 840atcagtatca ttacagctgt
cgtttttctc tcttgcaaaa agacaagaga aaaggtttct 900gggagaggta gagaaagaaa
acaggcagtg ccagaagaca ttcgtgtcga aattgcaagc 960tatgagcaag ttttaaaagt
cttcaaattc gaagaggtaa ggaaagctac tgaaaatctc 1020agttcagaaa gtagaataaa
tggctctgtg tatcgtggag agtttggtgg ggagatctta 1080gctgtcaaga agatgagtag
agatgtaaca aaggaagtga acattttgaa gaggatcaac 1140cacttcaatt tgattaagct
tgaaggtgta tgcgaaaatc gtggctgttt ctaccttgtt 1200ttggagtata tggaaaatgg
gtcccttaga gagtggctgt cctgtaaaaa gttcgaagaa 1260actggaaatt gggcacagag
aattcagatt gctctggatg ttgctaatgg actttactat 1320cttcacagct tcaccgaacc
tgcctatgtg cacaaggaca tcaaaagcag caatgttctg 1380ttaaacggca atctaagggc
caagattgca aatttcagtc ttgcgagagc agcgacaagc 1440gctgccatga caaaacatgt
tgtgggctct ataggttaca tggcgcctga gtacgtacgg 1500gaaggacaag tgacccccaa
gattgatgtt tatgcttttg gagtcatttt gctggaactg 1560atcacaggga aagatgctgt
tttcacacag gatggcaggg aagccctcct ttcaacggaa 1620atattttcta tcatggagaa
taaaaaccct gaagttgagc tggatttctt tgtcgatcct 1680gctctaaaag gaagttgtgg
aacaaacttc gcattatgct tggctaaagt aagcgtagcc 1740tgcttgatga aagaaccagc
aaggagacct agtatggagg aagtggtatc agttctgttg 1800aaaattcagg caaatgtaca
gaaatcataa 1830901913DNAPopulus sp.
90atgagcttcc gttctgttgt ttctcctttt gctctcttca ttctttattg ctgttctttg
60attcaagctc agcagcctta cgtggggaaa ggtacaacaa aatgttcaaa cacagaaaat
120tccgctctgg ggtattcttg caatggcctg aacaagagtt gccaagctta tctgatcttc
180agatcccaac ctccttacag tactgttgcc tccatatcta ctcttttggg ttcagaccca
240tctcagctct ctcaaataaa ttcagtttct gagactacat catttccaac aaaccagttg
300gtgctggttc ctgtcaactg ctcatgttca ggcgattatt ttcaggcaaa cgcatcttat
360attgttcaat caggtaacac tcctttcttg attgctaata acacttatca aggcctctca
420acctgtcaag ctatccggaa tgaaaaaggt acacgaactg tcaatatatt tgctggtgaa
480acactcactg ttcctcttag atgtgcctgt cctacaaaga accagagtga tttaggtatc
540aggtatctgt taagttactt agtcacatgg ggcgatacag tttcaattgc tggtgtacga
600tttggtgcag atatcgggag agctcttgaa gctaatgaaa tcagtgagaa aaatcccacg
660atttacccct tcacaacact cctaattcct ttaaaaaacc caccaacaag ttcacaaact
720gtagtgccac cgccacctcc agcttcacct tcaccttcac ctccatcacc gtctccaaac
780tccgacaaaa gcgcaaacaa aacatggatt tatgttttcg ttggggctgt tggaggaatt
840gttcttacac tagtcattgg aaccattatt ttcttcatgc tcttccgaaa aagtaagaag
900caaccgggtc caattatcgt gtcacaaagc tttgaggcac atgagaaacc actcaacagg
960aagttggatg aagaacctca ggatttgtta gagagtgttt atagcatagc tcaatccatc
1020aaagtctaca actatgaaga tctgaaagct gcaacagata acttcagtcc cagtttttgg
1080atcaaagggt ctgtttttcg tggcctaatc aatggtgatt tcgctgccat taagaagatg
1140aacggagacg tgtccaagga gatagattta ttgaataaga tcaaccactc taatctaatt
1200cgcctctccg gtgtttgttt caatgatggg cattggtacc tggtttacga gtatgctgcc
1260aatggacctt tgagtgattg gatttatgtt agcagtaacg aaggaaagtt tttgaaatgg
1320acacaaagga tacagattgc tacggatgta gctacagggc ttaactatct gcatagtttt
1380actaactatc ctcatgtcca caaggatata aagagcagca acatacttct tgataaagat
1440ttaagggcta agattgcgaa cttttccctg gcaaggtcaa cagatgggcc ggaaggtgaa
1500tttgcattga caaggcacat cgttgggact aaaggttaca tggctcccga gtacttggaa
1560aatggtatta tctgtacaaa acttgatgtc tatgcatttg gaattctcac actggagata
1620atgactggga aagaagttgc tgctttatac agagaggaaa acagggagct atctgatgtt
1680ttaaatggtg ttctttctga ggaaggtggg ctggaggaga gtctgagtca acttattgat
1740ccttcaatgc aagggaatta tccttcagga ctcgccgttt taatggtcag attgattgat
1800agttgcttga acaaaaatcc agcaggtcgc ccagccatgg atgaaatcgt gcagtctctc
1860tcaggaattt tgattacttc tctggcctgg gaattgtcaa acaacacatc tag
1913912043DNAPopulus sp. 91atgggcatgg ttccacttat cagtaagctc tactttggag
ctttcgcctt tatattaatc 60tgtttacttg tctctgcact tggacagaac ttgttgagct
gccagacaac atctccagat 120gcttctggct accattgtaa cagtaacgga ttgcaggatc
agtgcaagac atttgctatt 180cttcacacta gctcatactt ctcatctctt tccaacttaa
gcttttattt gggcttagat 240cggtttgtga tcgcggcaac aaacggcttt tctgccaaca
ctgagtttct gccgaaagat 300cagcctttgc tgataccgat tgactgcaaa tgcaatggtg
gcttctttca ggctcttgtg 360acgaaaacta ccatcaaagg ggagagtttt tacagcattt
ctaagtcact ggagggacta 420acaacctgca aagccattcg agagaagaac ccaggtatct
caccggagaa tctaaatggt 480aaagttcagt tacaggtacc cttgagatgt gcttgtccat
cctctactga agtcattcta 540gcaactagac tattgctttc ttatccggta agtgctggcg
acacaatctc gaaccttgct 600attaagttca atactactcc agaagctatt acatccgcaa
ataacagatc attgacaacc 660tttaaaccca caagccttgt acctcttaca tctcttttga
taccactagg tggcaagcct 720accctcggtc cccttgcaaa acctaatgaa cccaatttgc
atattcctgc aagtagcctt 780ccagtgatca atccacataa gaaaaggtcc aagatgtgga
ggattggtgt ttatattgct 840gttactggag ctgtagttgg agtaagcatt gctattgcag
cagctttctt ggtgatccaa 900ttgaagaaga agaagcaagt tttaagcaag gaggcagata
cggagcttca gcagcttagt 960ttaagtgtaa gaactacaag tgacaagaaa gtctcattcg
atgactcaca aaatcatttt 1020gatagtcaga tcactgacac cacacccggt aaggtgtttg
tggagactta cactgtagag 1080gagctcaaaa gagcaaccga ggacttcaat tcaagcaatc
aaatcgaagg ttctgtgtat 1140catggtcgtc tcaatgggaa gaacttggca ataaagcgcg
tgcaaccaga aactatttca 1200aaagttgagc ttgggctttt tcaagatgca actcaccatc
atccaaacat aatcagagtg 1260gtgggaacat gtttgagtga aggtcctgat tcatttttgg
tttttgagta cgctaaaaat 1320ggatctttga aggattggct tcatggggga ctggccatga
agaaccaatt cattgcctcc 1380tgctactgtt tcctaacgtg gaatcagagg ctgaagatat
gtcttgatgt agcagtggcc 1440ctacaatata tgcaccatat catgcaccct agctatgttc
atagaaacat caagagccgg 1500aacatcttcc tcgatgaaga attcaatgcg aagataggaa
atttcggcat ggcagggtgt 1560gttgaagatg ataccaagga gccagacttt aattcaacca
accctgcctc ctggagcctt 1620ggatacttgg ctcctgaggc tcatcaaggt gtagtttcct
ctagcacaga tatattttct 1680tttggggtgg ttctgatgga ggttctatct ggacaaacac
cgataacaag gcctaacgac 1740aacggagaag ggagcatttg gctgtcgaag aaaatcaagt
ccattttgct atcagaaaat 1800gcggatgagc tgagagaatg gattgacagt gcaatggggg
agaattactc atttgatgag 1860gctgccactt tggccaatat tgcacgagct tgcacagaag
aagacccttc tttgagacca 1920acttctggtg aaatcgttga aaagctgttg agattggtgg
aagaatccac agaaggagag 1980cagatactaa tctgtgaaag ctcttgcaaa cctctagtca
agtcaactgc aacaagtgtt 2040taa
2043921902DNAPopulus sp. 92atgaactgta cggacactac
acgactgtgc acatcgtttt tggcctttaa accccaggag 60aacctaactc tagcaatgat
tcaaagcatg tttgatgtgt taccccaaga tgtgaccaga 120gaaggcaatg gccatggcta
catattcatc aagaagaact gctcttgttt gtccaaggac 180aaagtctatg tgactaactc
tacatataca gtgaaattca gtgggggtta tgtttatgac 240atcgtgatca atgcttatga
cgggcttgct ttcttgccca acacaacaag acaagcaaag 300gttggtgctg ttgtgtcatt
gaggttgttc tgtgggtgtt caagtgggtt gtggaattat 360ttggtgagtt atgtgatgaa
ggaaggagat actgttcagt ccttgtctag tcgatttggt 420gttagtatgg ataatattga
gacagtgaat gggattcaga atcctgataa tgttacagct 480ggtgcgctgt attatattcc
tctgaattca gttcccgggg aaccttatcc cttggagaat 540gataatcctc ctgctcctgt
tcctgcacct ccggatgaca ttttctcagc aaatattcca 600acaattcaca aggctcgtgt
accatatgga tggatcatag ggggtcttgg gatcggtctt 660gcactgattg tattatgtat
aattatttgt gtgtctctta agtcctcaag ttgcttatct 720gaatctcgag gaagtcacgc
caagcctcct gatggcaaga tttctcaaaa gtttcatatt 780cttcgcaagc aaagcttttg
ttgtacttct agaaggtcca tctgctgcaa atctgtagac 840tggaagcaaa caaatgggga
gtctagcagc caccagatta ctatacctaa aggtctagca 900actgatgtat ttgatgagaa
gcctgtggtt tttacatacg aagaaattct ttttgcaact 960gatgaatttt tggattctag
tcttcttggt catggaacat atggttctgt gtattatggt 1020caccttcatg accaggaagt
tgctattaag agaatgactg ctacaaaaac taaagaattt 1080atggctgaga tgaaaatcct
gtgcaaggtc catcatacaa atctggtaga attgattggc 1140tacgcagcta gtgatgcaga
gctcttcgtg atctatgaat atgcacagaa gggttcactt 1200agaagtcatt tacatgatcc
ccaaaataag ggtcatacac cactttcatg gatcatgcgg 1260gtccagattg cacttgatgc
agctagaggc ctggagtaca tccatgagca cactaaaaca 1320cactatgtcc accgggatat
caagacaagc aacatcttac ttgatggttc cttcagggcc 1380aagatttcag attttggatt
ggcaaaactg gttggcaaaa caggcgaggg agaagctaca 1440gcaacaaaag ttgttggtac
atatggttat ctagcaccag aatacttgag tgatggcctt 1500gccacaacca agagtgatgt
ttatgcattt ggtgtcgttc tttttgagat catatctggg 1560aaggaagcca taataagaac
tgaaggtgcg gttacaaaaa atcctgaaag acgttcactg 1620gcatctacta tgctagcagc
tcttaggaac acacctgact ccatgagcat gtcaagcttg 1680aaagacctca ttgatcctaa
tatgatggat ttatatcccc atgattgtgt attcaagttg 1740gccatgctag caaagcaatg
tgtggatgag gatcccatcc tacggcctga catgaagcaa 1800gttgtgattt ccctgtcaca
aattgtttta tcctccattg agtgggaagc aactcttgct 1860gggaacagcc aagttttcag
tggtcttgtc caaggaaggt ag 1902931953DNAPopulus sp.
93atgaacccgg gtccaggatt tcatgcgttg caagttttag agattatatc aggtttagga
60ggttttctcg ggtttacttg ttctttgatt caagctcagc aaccatatgt agggaaagct
120acgacaaatt gttccaacac ggaaaattct gctcttgggt attcttgcaa cgccctaaac
180aagagctgcc aagcttatct catcttcaga tcccagcctc cttacaacac tgttgcctcc
240atatctaccc ttttgggatc tgacccatct cagctctctg aagtaaattc agtttctgag
300accacatcat tcccatcaaa ccagttggtg atagttccag taaactgctc atgttcaggc
360gagtactctc aggcaaacgc atcttatatt gttcaaccga acgatactct tttcttgatt
420gctaataaca cctatcaagg cctctcaacc tgtcaagctc tccagaatca aaaaactacg
480cgaactgacg atatacttag cggtgaaaca ctcactgttc ctcttagatg tgcctgtccc
540acaaagaacc agagtgattt aggtataagg tatctcttaa gttacttagt cacaccggga
600gatgacgttc cagctattag tgaacaattt ggcgcagcta ctgggagaac tctcgaggct
660aatggactcc ctgagcaaaa tcccaccatt tttcccttca caacactcct aattccttta
720caaagcacgc caacaagttc tcaaactgta gtgccaccac cacctccagc ttcatcttca
780ccaccatcac catctccaaa ccctgaaaaa agctcaaaga aaacatggct ttatgttgtg
840gttggggttg ttggaggaat tgctcttaca atagtcattg gaaccattat tttcttcatg
900ttgtcccgta aaagtaagaa gcaacccggt ccagttatcg aatcgcaaag ctttgaggca
960catgagaaac cactcaacaa gaagttggat gaagaatctc aggagttttt cgagagtata
1020tccgctatag ctcaatccat caaagtctac aagtttgaag atctgaaagc tgcaactgat
1080aacttcagtc ctagctgttg gatcaaaggg tctgtttatc gtggcctaat caatggtgac
1140tttgctgcca ttaagaagat gaatggagac gtgtctaaag agatagaatt gttgaataag
1200atcaaccatt ctaatctaat tcgcctctcc ggtgtttgtt tcaatgatgg tcattggtac
1260ctggtttacg agtatgctgc tagtggacaa ttgagtgatt ggatttatga tagaagcaat
1320gaagggaagt ttttaaattg gacaaaaaga atacagattg cttccgatgt cgccacggga
1380cttaattatc tacatagttt cactaactat ccgcatgtcc acaaggatat aaagagcagc
1440aacattcttc ttgacagtga tttaagggct aaaattgcaa acttttccct ggcaaggtca
1500acaggtgacc aggatgatga atttgttttg acaaggcaca ttgttgggac aaaaggttac
1560atggctcctg agtacttgga aaatggagtt gtctcctcaa agcttgatgt ctatgcattt
1620gggattctca ctctggagat aattaccggg aaagaagttg ctgctttaca cagtgaggaa
1680agcaggaact tatcagatgt tttaaatggt gctctatctg aggtagatgg gcaggaggag
1740agtttgaagc aactcattga tccttcactg catgagaact atccttcagg acttgctgtt
1800ttagtggtca gattgattga tagttgctta aacaaaaacc caggagatcg cccgaccatg
1860gatgaaatcg tgcagtctct ctcaagaatt ttgactactt cgctggcctg ggaattgtct
1920agcaatgtat ctggctacca tatctctagt tag
1953941878DNAPopulus sp. 94atgacatcca aatcccatct agtcttctct ctcctctttt
ttatctatta cagtactatc 60ctccaccatt tgcaagccca accaagcacc caaggattca
cctgcacagc caatcagagt 120tcctttccat gccaaaccta tgccttctac cgagctacag
ctcctaactt ccttgacctt 180tcctcaatcg gtgacctttt ctcggttagc cgccttatga
tatcaaaacc aagtaacatc 240tcctctccag cctctcctct catccccaat caacccttgt
ttgtcccttt atcatgttct 300tgcaacacca tgaatggcac tagtatctcc tttgcaaaca
tcacatatac cataaagcca 360aatgacactt tctaccttgt ctcaactgaa tacttcggaa
accttactac ctaccagtct 420gttcaacttg ttaaccctac gcttatcccc acactactcc
aaataggagt agaagtgatc 480tttccaatat tttgcaagtg tcctaatcaa actcaattgc
aaaacaaggt gaattatctg 540gtatcttatg tgtttcagcc ttctgataac ttatcatcag
ttgcttcaac atttggagtc 600gaaacacaat ctattgtgga tgctaatggc aataacatac
agccttttga taccatattc 660ataccagtaa atcaacttcc acaactggca caacctacgg
tttttccttc tttggcgcct 720tctgggaaga ctcagaggaa aggtttgatc ataggattag
cagttggact aggaattgcc 780gggcttttgt tggtcttggt aagcggggtt tgttttttta
gagacggtgt attgaagaag 840agacgggatt ttgagagaga tgatcaggag aagcagagga
tgcagttcaa tggaggaagg 900aaagggttga aggatataga agtgaatttg atggcagatg
tttcagattg cttggataag 960tacagggtct ttaagattga tgaactgaaa gaagctactg
atgagttcgg tgagaattgc 1020ttgattgaag gatctgtgtt taaagggtcc ataaatggag
agacctatgc catcaagaag 1080atgaagtgga atgcttgtga ggagctcaag atattgcaga
aggtaaacca tggcaacttg 1140gtgaagctag aaggcttttg catagaccct gaggatgcaa
attgctatct ggtctacgag 1200tacatcgaca atggctctct gcattcgtgg ttgcatggta
acgagaaaga aaaactaagc 1260tggaaaacaa ggttacgcat tgcaattgac gttgcaaatg
gtctccaata catccatgag 1320cacactaggc caagggttgt acacaaagac attagaagca
gcaacattct tctagactcg 1380agcatgagag ccaaaattgc caactttgga ctagcaaaat
caggctacaa tgccataaca 1440atgcacattg tcggcaccca aggctacatt gcacctgaat
atttagctga cggtgtggtg 1500tcaacaagaa tggatgtttt ctcatttggt gtggttttgc
ttgagctaat ctcaggcaaa 1560gaagcaattg acgaagaagg caaggttttg tgggcagaag
ctagtggaat tttgcaggga 1620aatgttgaag agaggaaggt gaagagattg acaccatgga
tggataaggt tctattagag 1680cagtcatgct taatggagag tgtaatgaat gcaatggttg
ttgcaattgc ttgcttgcat 1740agagatccat caaagaggcc cagcatggtg gatattgttt
atgccttgtg taagaccgat 1800gatctgtttt ttgacatctc agaagacgta ttgtcagacc
ctcaggcatc tttagcctat 1860gggcgtactc ctttttaa
1878951728DNAPopulus sp. 95atggcaatat ctttgctccc
tttcttttcc acacaagctc ttcttctctt gatcctttta 60ttttcctcca catatgtcac
ctcacaagca ccacctgcct caaacttttc atgtcctgtt 120gattcaccta ccacatgctc
tacatatttc acttaccttg cccagccacc aaatttcttg 180gacctcggaa acatttctga
tctatttggg gtcagtcgta aggaaatagc aactgcaagc 240aacctggagt ctgaggacac
cccattattt ccaaatcaac ttttgcttgt acctaaacct 300tgtggttgca caggtaaccg
gtcttttgcc aacataactt accaaatcca gcaaggtgaa 360agcttctact tggtttcgac
tacttcattt gagaacctca cccgttggca agaggtggaa 420gctttgaacc ccagtctgac
tccaaccctc ttgcatgctg gtgacaaggt tatatttcct 480ttgttctgta agtgcccttc
aaatactcat ttggagagcg gaattgctta tctcattact 540tatgtgtggc aacctagtga
tgatctcacg aaagttgctg ctaaacttaa tgcctctgaa 600cgtaacattg tgattgaaaa
caactatgtg aactttactg ctgcagtcta cctcccggta 660ttgatccctg tgtcccagtt
gccagttctc tctcaaacta aagcttttga tcgcactgga 720tcttgtttgg agaccggcga
tctcattcaa acaaaggaac ttacaaaact ggagagtttt 780gaggcaaaga tcacaccaga
tgagctgctt ccaggggttt caggatatct gagcaagccg 840atcatttatg aggttaaaga
gatcatggag ggcacgatgg atctaaatga acattataag 900atacgaggat cagtatatag
ggccaaaatc aatggccggg tcttggcagt gaagaaaacc 960aaggatgatg tcacagaaga
actaaagatt ttgcagaaag tcagtcatgc aaatctggtt 1020aaactaatgg ggatgtcatc
tggatttgac agagaaggaa atcgcttctt ggtctatgaa 1080tatgcagaaa atgggtcgct
ggaaaaatgg ttgcatccca cctctgaatc ttcctcaagc 1140tctgcgggtt tcctcacttg
gagtcaaagg ttacatgtag cactagatgt ggccaatggc 1200ctgcaataca tgcatgaaca
cactcaacca agcatcgttc acaaggatat tcgaacaact 1260aatattcttc tcgattccac
atttcgggcg aaaatagcaa atttctcaat ggctagacct 1320gccacagact cattgatgcc
aaaagttgat gtatttgatt atggagttgt tttgttacaa 1380ttgctttctg gaaagaaggc
catggtaacc aaagaaaatg gcgagattgt tctgctgtgc 1440aaggaaatca aagccgtctt
ggaaattgaa gagaaaagag aagagagtct aagaaagtgg 1500atggatccct gcttggagag
gttttatccc attgatagtg ctctgagctt ggcaaccttg 1560gcaaggttgt gcactctgga
ggagtcttca gaaaggccaa gcatggcaga aattgtcttc 1620aacctcacag ttctcactca
gtcacctcct gagacatttg aaagatggac atctgggatg 1680gaaacagaag attttactcg
actcatcagc cctgtcacag ctcgttga 17289621DNAArtificialPCR
primer 96agctattcgc agttcccaaa t
219721DNAArtificialPCR primer 97cagagacgaa ccttgaggag a
219821DNAArtificialPCR primer
98aagaacatcc gtggaaaggt t
219921DNAArtificialPCR primer 99aatgttccca caagacgagt g
2110021DNAArtificialPCR primer 100tgacatatgc
caatctcacc a
2110121DNAArtificialPCR primer 101gtgacattaa ccgtggcatt t
2110221DNAArtificialPCR primer
102gatccacaac aacgtccaaa t
2110321DNAArtificialPCR primer 103atggaagcaa tatcccaatc c
2110421DNAArtificialPCR primer
104taacggtgac gttgatgttc a
2110521DNAArtificialPCR primer 105gttgtcgagg ttgatttctc g
2110621DNAArtificialPCR primer
106agatgtgctt gtcccacaaa g
2110721DNAArtificialPCR primer 107cagaatcacc ccagtttacc a
2110821DNAArtificialPcR primer
108accgctcttt tgccaatatc t
2110921DNAArtificialPCR primer 109aacggggttt aaatccatca c
2111021DNAArtificialPCR primer 1
110catggccaga acttttacca a
2111121DNAArtificialPCR primer 111gttgtcatgg ctttcctacc a
2111221DNAArtificialPCR primer 1
112tgatctccta cgtcgtccaa c
2111321DNAArtificialPCR primer 113gcgtcaatga tggactgttc t
2111421DNAArtificialPCR primer
114cctctctctc caacctcacc t
2111521DNAArtificialPCR primer 115ctgatcctgg gagaggaact c
2111621DNAArtificialPCR primer 1
116ttcggttcct ggtgagtctt a
2111721DNAArtificialPCR primer 117tcatggggta catgagcttt c
2111826DNAArtificialPCR primer
118tggcatcact cagtaccttt caacag
2611928DNAArtificialPCR primer 119acccaaagca tcaaataata agtcaacc
2812020DNAArtificialPCR primer
120catgagcatt cagtgcctgt
2012121DNAArtificialPCR primer 121tgcagaatca gtaagcctgg t
2112223DNAArtificialPCR primer
122tgctaagggt tcagctgttg gta
2312324DNAArtificialPCR primer 123aaatgcccta gaagtttgtg gaag
2412420DNAArtificialPCR primer
124cgcaagatgg atgtgtatgc
2012520DNAArtificialPCR primer 125catggctctc gaactcgttt
2012625DNAArtificialPCR primer
126cactcatatt cttttctgcc accca
2512723DNAArtificialPCR primer 127tgcaatggat tgaggactgg tgt
2312820DNAArtificialPCR primer
128ggaaatggag aaatggcaaa
2012920DNAArtificialPcR primer 129cgccttgacc aagaaaccta
2013020DNAArtificialPCR primer
130ggcattgatg ggtcagaact
2013120DNAArtificialPcR primer 131tgcaaagagg atcacactgc
2013225DNAArtificialPCR primer
132ctcttcttct tcttcttcgt cagca
2513324DNAArtificialPCR primer 133ggtatgcttg gcatgtttga gttt
2413420DNAArtificialPCR primer
134ggttgttctc ggaatcttcg
2013520DNAArtificialPCR primer 135atgcatgtat tgcagaccga
2013624DNAArtificialPCR primer
136tttcactctt ggtgtgaagc agat
2413725DNAArtificialPcR primer 137gacttccttc acgatttcat cgtaa
2513820DNAArtificialPCR primer
138atggcgatat gggtgacatt
2013920DNAArtificialPCR primer 139tccacatgga aggtgaatga
2014020DNAArtificialPCR primer
140gttcttgcgt ctggtgctct
2014119DNAArtificialPCR primer 141ctccttatcc ggagccaac
1914220DNAArtificialPCR primer
142atggaggagg tgttcgtcac
2014320DNAArtificialPCR primer 143ccgaggacca tagaagctga
2014420DNAArtificialPcR primer
144catggtcacc tacctcgtca
2014520DNAArtificialPCR primer 145tatgatggag ctctcggtga
2014620DNAArtificialPCR primer
146gttcatcgac aaaccgatca
2014720DNAArtificialPCR primer 147taatacgagc tgccgagctt
2014820DNAArtificialPCR primer
148gtgacgagga gaatggagga
2014920DNAArtificialPCR primer 149ctcgatcagc ttcaccatca
2015029PRTMedicago truncatula 150Thr Tyr
Pro Ile Arg Ser Glu Asp Thr Leu Glu Ser Ile Ala Lys Gly 1 5
10 15 Ala Glu Ile Glu Ala Glu Leu
Ile Gln Arg Tyr Asn Pro 20 25
15130PRTMedicago truncatula 151Ile Thr Tyr Pro Leu Arg Pro Glu Asp Ser
Leu Glu Leu Ile Ser Asn 1 5 10
15 Lys Tyr Glu Ile Asp Ala Glu Leu Ile Gln Lys Tyr Asn Pro
20 25 30 152107PRTMedicago
truncatula 152Tyr Ser Ala Lys Glu Gly Asp Thr Tyr Asp Leu Ile Ala Asn Ser
Tyr 1 5 10 15 Tyr
Ala Ser Leu Thr Thr Val Glu Leu Leu Lys Lys Phe Asn Ser Tyr
20 25 30 Asp Gln Asp His Ile
Pro Ala Lys Ala Lys Val Asn Val Thr Val Asn 35
40 45 Cys Ser Cys Gly Asn Ser Gln Ile Ser
Lys Asp Tyr Gly Leu Phe Ile 50 55
60 Thr Tyr Pro Leu Arg Thr Asp Asp Thr Leu Gln Lys Ile
Ala Asn Gln 65 70 75
80 Ser Asn Leu Asp Glu Gly Leu Ile Gln Ser Tyr Asn Ser Gly Val Asn
85 90 95 Phe Ser Asn Gly
Ser Gly Ile Val Phe Ile Pro 100 105
15346PRTPopulus sp. 153Tyr Thr Val Gln Glu Asn Asp Thr Leu Thr Gly Ile
Ala Glu Leu Leu 1 5 10
15 Ser Ala Glu Leu Thr Gly Ile Glu Asn Leu Asn Glu Arg Phe Thr Arg
20 25 30 Asn Pro Asn
Leu Ile Asp Val Gly Trp Val Leu Phe Val Pro 35
40 45 154112PRTPopulus sp. 154Ile Ser Ser Ala Asn
Ile Thr Tyr Thr Ile Glu Ala Gly Asn Thr Phe 1 5
10 15 Tyr Ile Val Ser Thr Glu Tyr Phe Gln Asn
Leu Thr Thr Tyr Gln Ser 20 25
30 Val Glu Leu Phe Asn Pro Thr Leu Ile Pro Glu Leu Ile Asp Ile
Gly 35 40 45 Val
Glu Val Pro Ile Phe Cys Lys Cys Pro Asn Gln Thr Gln Leu Gln 50
55 60 Asn Lys Val Asn Tyr Leu
Val Ser Tyr Val Phe Gln Pro Ser Asp Asn 65 70
75 80 Leu Ser Ser Val Ala Ser Thr Phe Gly Val Glu
Thr Gln Ser Ile Val 85 90
95 Asp Val Asn Gly Asn Asn Ile Gln Pro Tyr Asp Thr Ile Phe Val Pro
100 105 110
155116PRTPisum sativum 155Arg Tyr Phe Ala Asn Phe Thr Tyr Thr Ile Lys Leu
Gly Asp Asn Tyr 1 5 10
15 Phe Ile Val Ser Thr Thr Ser Tyr Gln Asn Leu Thr Asn Tyr Val Glu
20 25 30 Met Glu Asn
Phe Asn Pro Asn Leu Ser Pro Asn Leu Ile Pro Pro Glu 35
40 45 Ile Lys Val Val Val Pro Leu Phe
Cys Lys Cys Pro Ser Lys Asn Gln 50 55
60 Leu Ser Lys Gly Ile Lys His Leu Ile Thr Tyr Val Met
Gln Ala Asn 65 70 75
80 Asp Asn Val Thr Arg Val Ser Ser Lys Phe Gly Ala Ser Gln Val Asp
85 90 95 Met Phe Thr Glu
Asn Asn Gln Asn Phe Thr Ala Ser Thr Asn Val Pro 100
105 110 Ile Leu Ile Pro 115
156107PRTPopulus sp. 156Tyr Thr Val Gln Lys Asp Asp Gly Leu Phe His Ile
Ala Ala Glu Val 1 5 10
15 Phe Ser Gly Leu Val Thr Tyr Gln Glu Ile Ala Ala Val Asn Asn Ile
20 25 30 Ser Asp Val
Asn Leu Ile Lys Val Gly Gln Lys Leu Leu Ile Pro Leu 35
40 45 Phe Cys Asn Cys Asp Asp Val Asp
Gly Val Lys Val Val His Tyr Gly 50 55
60 His Val Val Glu Ala Gly Ser Ser Leu Glu Leu Ile Ala
Gln Glu Tyr 65 70 75
80 Gly Thr Ser Thr Asp Thr Leu Val Lys Leu Asn Gly Val Asn Asp Ser
85 90 95 Ser Leu Leu Ala
Gly Gln Val Leu Asp Val Pro 100 105
157110PRTPopulus sp. 157Tyr Thr Val Gln Lys Asp Asp Gly Leu Tyr His Ile
Ala Ala Glu Val 1 5 10
15 Phe Ser Gly Leu Val Thr Tyr Gln Glu Ile Ala Ala Val Asn Asn Asn
20 25 30 Val Thr Asp
Val Asn Leu Ile Glu Val Gly Gln Glu Leu Met Ile Pro 35
40 45 Leu Pro Cys Ser Cys Asp Asp Val
Asp Gly Val Lys Val Val His Tyr 50 55
60 Gly His Val Val Glu Ala Gly Ser Ser Leu Glu Phe Ile
Ala Gln Glu 65 70 75
80 Tyr Gly Thr Ser Arg Asn Thr Ile Met Lys Leu Asn Gly Ile Ala Asn
85 90 95 Gly Ser Ser Leu
Leu Ala Gly Gln Leu Ser Leu Leu Val Ser 100
105 110 158108PRTPopulus sp. 158Tyr Thr Val Gln Pro Asp
Asp Gly Leu Tyr Tyr Ile Ala Asn Asn Val 1 5
10 15 Phe Met Gly Leu Leu Ala His Gln Arg Ile Gln
Gln Val Asn Arg Ile 20 25
30 Glu Asn Pro Asn Val Ile Tyr Val Gly Gln Glu Leu Trp Ile Pro
Leu 35 40 45 Pro
Cys Ser Cys Glu Glu Val Glu Gly Glu Arg Val Val His Tyr Ala 50
55 60 His Leu Val Glu Glu Gly
Ser Thr Val Glu Glu Ile Ala Glu Lys Phe 65 70
75 80 Gly Thr Thr Asn Asp Thr Leu Tyr Arg Leu Asn
Gly Ile Thr Asn Asn 85 90
95 Ser Gln Leu Ile Ala Ala Thr Ala Phe Asp Val Pro 100
105 159107PRTArabidopsis thaliana 159Thr Ile
Lys Lys Asp Asp Ile Leu Ser Phe Val Ala Thr Glu Ile Phe 1 5
10 15 Gly Gly Leu Val Thr Tyr Glu
Lys Ile Ser Glu Val Asn Lys Ile Pro 20 25
30 Asp Pro Asn Lys Ile Glu Ile Gly Gln Lys Phe Trp
Ile Pro Leu Pro 35 40 45
Cys Ser Cys Asp Lys Leu Asn Gly Glu Asp Val Val His Tyr Ala His
50 55 60 Val Val Lys
Leu Gly Ser Ser Leu Gly Glu Ile Ala Ala Gln Phe Gly 65
70 75 80 Thr Asp Asn Thr Thr Leu Ala
Gln Leu Asn Gly Ile Ile Gly Asp Ser 85
90 95 Gln Leu Leu Ala Asp Lys Pro Leu Asp Val Pro
100 105 160110PRTGlycine max 160Pro Leu
Tyr Arg Ile Lys Lys Gly Asp Thr Leu Tyr Tyr Ile Ala Thr 1 5
10 15 Thr Thr Phe Ala Gly Leu Met
Lys Trp Pro Gln Ile Gln Val Ala Asn 20 25
30 Asn Ile Ala Asn Ala Asn Asn Ile Thr Thr Gly Asp
Met Leu Tyr Ile 35 40 45
Pro Leu Pro Cys Ser Cys Asp Glu Val Gly Gly Lys Ser Val Val His
50 55 60 Tyr Ala His
Leu Val Ala Pro Gln Ser Thr Val Glu Gly Ile Ala Glu 65
70 75 80 Glu Phe Gly Tyr Tyr Gln Gln
Ile Leu Leu Asn Leu Asn Gly Ile Ser 85
90 95 Asp Pro Lys Asn Leu Gln Ala Gly Gln Ile Leu
Asp Val Pro 100 105 110
161108PRTOryza sativa 161Tyr Val Val Gln Pro Gln Asp Gly Leu Asp Ala Ile
Ala Arg Asn Val 1 5 10
15 Phe Asn Ala Phe Val Thr Tyr Gln Glu Ile Ala Ala Ala Asn Asn Ile
20 25 30 Pro Asp Pro
Asn Lys Ile Asn Val Ser Gln Thr Leu Trp Ile Pro Leu 35
40 45 Pro Cys Ser Cys Asp Lys Glu Glu
Gly Ser Asn Val Met His Leu Ala 50 55
60 Tyr Ser Val Gly Lys Gly Glu Asn Thr Ser Ala Ile Ala
Ala Lys Tyr 65 70 75
80 Gly Val Thr Glu Ser Thr Leu Leu Thr Arg Asn Lys Ile Asp Asp Pro
85 90 95 Thr Lys Leu Gln
Met Gly Gln Ile Leu Asp Val Pro 100 105
162109PRTPopulus sp. 162Tyr Lys Thr Arg Pro Ser Asp Thr Leu Ser Thr
Ile Ala Asp Ser Ile 1 5 10
15 Tyr Ala Gly Leu Val Ser Ala Asp Gln Ile Lys Glu Ala Asn Ser Ile
20 25 30 Asp Asp
Pro Ser Val Leu Asp Val Gly Gln Ser Leu Val Val Pro Leu 35
40 45 Pro Cys Thr Cys Phe Asn Gly
Thr Asp Asn Ser Leu Pro Ala Ile Tyr 50 55
60 Leu Ser Tyr Val Val Lys Glu Val Asp Thr Leu Ala
Ala Ile Ala Ala 65 70 75
80 Arg Tyr Ala Thr Thr Leu Thr Asp Ile Met Asn Val Asn Ala Met Gly
85 90 95 Ser Val Ala
Ile Met Ala Gly Asp Ile Leu Ala Val Pro 100
105 163110PRTPopulus sp. 163His Tyr Lys Thr Arg Pro Ser
Asp Thr Leu Ser Thr Ile Ala Asp Ser 1 5
10 15 Ile Tyr Ala Gly Leu Val Ser Ala Asp Gln Ile
Lys Glu Ala Asn Ser 20 25
30 Ile Asp Asp Pro Ser Val Ile Asp Val Gly Gln Ser Leu Val Val
Pro 35 40 45 Leu
Pro Cys Thr Cys Phe Asn Gly Thr Asp Asn Ser Leu Pro Ala Ile 50
55 60 Tyr Leu Ser Tyr Val Val
Lys Glu Val Asp Thr Leu Ala Ala Ile Ala 65 70
75 80 Ala Arg Tyr Ala Thr Thr Leu Thr Asp Ile His
Asn Val Asn Ala Met 85 90
95 Gly Ser Val Ala Ile Met Ala Gly Asp Ile Leu Ala Val Pro
100 105 110 164110PRTPopulus sp.
164His Tyr Lys Thr Arg Pro Ser Asp Thr Leu Ser Ser Ile Ala Asp Ser 1
5 10 15 Ile Tyr Ala Gly
Leu Val Ser Ala Asp Gln Ile Lys Glu Ala Asn Ser 20
25 30 Ile Asp Asp Pro Ser Val Ile Asp Val
Gly Gln Ser Leu Val Val Pro 35 40
45 Leu Pro Cys Thr Cys Phe Asn Gly Thr Asp Asn Ser Leu Pro
Ala Ile 50 55 60
Tyr Leu Ser Tyr Val Val Lys Glu Val Asp Thr Leu Ala Ala Ile Ala 65
70 75 80 Ala Arg Tyr Glu Thr
Thr Leu Thr Asp Ile Met Asn Val Asn Ala Met 85
90 95 Gly Ser Ala Ala Ile Lys Ala Gly Asp Ile
Leu Ala Val Pro 100 105 110
16579PRTGlycine max 165Ser Asp Pro Ser Val Ile Asp Val Gly Gln Asn Leu
Val Val Pro Leu 1 5 10
15 Pro Cys Thr Cys Phe Asn Gly Ser Asp Asn Ser Leu Pro Ala Ile Tyr
20 25 30 Leu Ser Tyr
Val Val Arg Pro Val Asp Thr Leu Ala Ala Ile Ala Ala 35
40 45 Arg Tyr Phe Thr Thr Leu Thr Asp
Leu Met Asn Val Asn Ala Met Gly 50 55
60 Ser Tyr Ala Ile Asn Asp Gly Asp Ile Leu Ala Val Pro
Ile Pro 65 70 75
166100PRTGlycine max 166His Tyr Arg Thr Arg Pro Ser Asp Thr Leu Ser Ser
Ile Ala Asn Ser 1 5 10
15 Ile Tyr Gly Gly Leu Val Ser Pro Asp Gln Leu Arg Glu Ala Asn Ser
20 25 30 Ile Gly Asp
Asp Pro Ser Val Ile Asp Val Gly Leu Asn Leu Val Val 35
40 45 Pro Leu Pro Cys Thr Cys Phe Asn
Glu Ser Asp Asn Ser Leu Pro Ser 50 55
60 Ile Tyr Leu Ser Tyr Val Val Gln Pro Ile Asp Thr Leu
Ala Ala Ile 65 70 75
80 Ala Ala Arg Tyr Phe Thr Thr Phe Thr Asp Ile His Asn Val Asn Asp
85 90 95 Met Gly Thr Thr
100 167100PRTArabidopsis thaliana 167Gly Ser Ile Ala Asp Ser
Val Tyr Gly Gly Leu Val Ser Ala Glu Gln 1 5
10 15 Ile Gln Glu Ala Asn Ser Val Asn Asp Pro Ser
Leu Ile Asp Val Gly 20 25
30 Thr Ser Leu Val Ile Pro Leu Pro Cys Ala Cys Phe Asn Gly Thr
Asp 35 40 45 Asn
Ser Leu Pro Ala Val Tyr Leu Ser Tyr Val Val Lys Glu Ile Asp 50
55 60 Thr Leu Val Gly Ile Ala
Arg Arg Tyr Ser Thr Thr Ile Thr Asp Leu 65 70
75 80 Met Asn Val Asn Ala Met Gly Ala Pro Asp Val
Ser Ser Gly Asp Ile 85 90
95 Leu Ala Val Pro 100 168108PRTArabidopsis thaliana
168Lys Thr Arg Thr Ser Asp Thr Leu Gly Ser Ile Ala Asp Ser Val Tyr 1
5 10 15 Gly Gly Leu Val
Ser Pro Glu Gln Ile Gln Val Ala Asn Ser Glu Thr 20
25 30 Asp Leu Ser Val Leu Asp Val Gly Thr
Lys Leu Val Ile Pro Leu Pro 35 40
45 Cys Ala Cys Phe Asn Gly Thr Asp Glu Ser Leu Pro Ala Leu
Tyr Leu 50 55 60
Ser Tyr Val Val Arg Gly Ile Asp Thr Met Ala Gly Ile Ala Lys Arg 65
70 75 80 Phe Ser Thr Ser Val
Thr Asp Leu Thr Asn Val Asn Ala Met Gly Ala 85
90 95 Pro Asp Ile Asn Pro Gly Asp Ile Leu Ala
Val Pro 100 105 169109PRTGlycine
maxmisc_feature(76)..(76)Xaa can be any naturally occurring amino acid
169Ile Tyr Thr Val His Ala Ala Asp Thr Leu Ala Ser Ile Ser Glu Gly 1
5 10 15 Tyr Gly Gly Leu
Val Ser Ala Glu Gln Ile Lys Ile Val Asn Ala Ile 20
25 30 Asn Ala Thr Asn Pro Leu Thr Tyr Arg
Gly Thr Leu Val Ile Pro Leu 35 40
45 Pro Cys Thr Cys Phe Asp Asn Val Asn Asn Gly Gly Asn Ala
Ile Tyr 50 55 60
Met Ser Tyr Val Val Gln Arg Arg Glu Ser Leu Xaa Ser Ile Ala Thr 65
70 75 80 Lys Phe Gly Thr Thr
Val Ser Asp Leu Glu Thr Val Asn Gly Phe Gly 85
90 95 Glu Ala Thr Val Asp Pro Gly Asp Ile Leu
Ser Ile Pro 100 105
17045PRTGlycine max 170Ile Tyr Val Val Arg Glu Gly Glu Thr Leu Gln Thr
Ile Ser Glu Lys 1 5 10
15 Cys Gly Asp Pro Tyr Ile Val Glu Glu Asn Phe His Ile His Asp Phe
20 25 30 Asp Asp Val
Phe Pro Gly Leu Val Ile Lys Ile Asn Pro 35 40
45 17144PRTMedicago truncatula 171Tyr Val Val Arg Glu Gly
Glu Thr Leu His Thr Ile Ser Glu Lys Cys 1 5
10 15 Gly Asp Pro Phe Ile Val Glu Glu Asn Phe His
Ile His Asp Phe Asp 20 25
30 Asp Val Phe Pro Gly Leu Val Ile Lys Ile Asn Pro 35
40 17244PRTPopulus sp. 172Tyr Val Val Arg
Glu Gly Glu Thr Leu Asn Thr Ile Ser Glu Lys Cys 1 5
10 15 Gly Asp Pro Tyr Ile Val Glu Glu Asn
Phe His Ile His Asp Phe Asp 20 25
30 Asp Val Phe Pro Gly Leu Val Ile Lys Ile Thr Pro
35 40 17345PRTGlycine max 173Ile Tyr
Val Val Gly Glu Gly Glu Thr Leu Phe Thr Ile Ser Asp Lys 1 5
10 15 Cys Asn Asp Pro Phe Ile Val
Glu Arg Asn Phe His Ile His Asp Phe 20 25
30 Asp Asp Val Phe Pro Gly Leu Val Ile Lys Ile Thr
Pro 35 40 45 17445PRTGlycine
max 174Ile Tyr Val Val Gly Glu Gly Glu Thr Leu His Thr Ile Ser Asp Lys 1
5 10 15 Cys Gly Asp
Pro Phe Ile Val Glu Arg Asn Phe His Ile His Asp Phe 20
25 30 Asp Asp Val Phe Pro Gly Leu Val
Ile Lys Ile Thr Pro 35 40 45
17544PRTPopulus sp. 175Tyr Val Val Gly Glu Gly Glu Thr Leu His Thr Ile
Ser Asp Lys Cys 1 5 10
15 Gly Asp Pro Phe Ile Val Glu Gln Asn Phe His Ile His Asp Phe Asp
20 25 30 Asp Val Phe
Pro Gly Leu Val Ile Lys Ile Thr Pro 35 40
17643PRTGlycine max 176Ile Tyr Val Val Gly Glu Gly Glu Thr Leu
His Thr Ile Ser Asp Lys 1 5 10
15 Cys Gly Asp Pro Phe Ile Val Glu Lys Asn Phe His Ile His Asp
Phe 20 25 30 Asp
Asp Val Phe Pro Gly Leu Val Leu Lys Ile 35 40
17745PRTGlycine max 177Ile Tyr Val Val Gly Glu Gly Glu Thr Leu
His Thr Ile Ser Asp Lys 1 5 10
15 Cys Gly Asp Pro Phe Ile Val Glu Lys Asn Phe His Ile His Asp
Phe 20 25 30 Asp
Asp Val Phe Pro Gly Leu Val Ile Lys Ile Thr Pro 35
40 45 17832PRTGlycine max 178Ile Tyr Val Val Gly Glu
Gly Glu Thr Leu His Thr Ile Ser Asp Lys 1 5
10 15 Cys Gly Asp Pro Phe Ile Val Glu Asn Asn Phe
His Ile His Asp Phe 20 25
30 17944PRTPopulus sp. 179Tyr Val Val Lys Glu Gly Glu Thr Leu
His Thr Ile Ser Asp Lys Cys 1 5 10
15 Gly Asp Pro Phe Ile Val Glu Glu Asn Phe His Ile His Asp
Phe Asp 20 25 30
Asp Val Tyr Pro Gly Leu Val Ile Lys Ile Thr Pro 35
40 18044PRTPopulus sp. 180Tyr Val Val Lys Glu Gly Glu
Thr Leu His Thr Ile Ser Asp Lys Cys 1 5
10 15 Asp Asp Pro Phe Ile Val Glu Glu Asn Phe His
Ile His Asp Phe Asp 20 25
30 Asp Val Phe Pro Gly Leu Val Ile Lys Ile Thr Pro 35
40 18143PRTPopulus sp. 181Ile Tyr Val Val
Gly Glu Gly Glu Thr Leu Asn Thr Ile Ser Glu Lys 1 5
10 15 Cys Asp Asp Pro Phe Ile Val Glu Gln
Asn Phe His Ile His Asp Phe 20 25
30 Asp Asp Val Tyr Pro Gly Leu Val Ile Lys Ile 35
40 18243PRTArabidopsis thaliana 182Ile Val
Gly Glu Gly Glu Thr Leu His Thr Ile Gly Asp Lys Cys Gly 1 5
10 15 Asp Pro Phe Ile Val Glu Arg
Asn Phe His Ile His Asp Phe Asp Asp 20 25
30 Val Phe Pro Gly Leu Val Leu Lys Ile Ala Pro
35 40 18346PRTOryza sativa 183Glu Ile
Tyr Val Val Glu Glu Gly Glu Thr Leu His Ser Ile Ser Asp 1 5
10 15 Arg Cys Gly Asp Pro Tyr Ile
Leu Glu Gln Asn Phe His Val His Asp 20 25
30 Phe Asp Asp Val Phe Pro Gly Leu Val Ile Lys Ile
Thr Pro 35 40 45
18443PRTArabidopsis thaliana 184Tyr Val Val Gly Glu Gly Asp Thr Leu His
Ser Ile Ser Glu Lys Cys 1 5 10
15 Gly Asp Pro Phe Ile Val Glu Arg Asn Phe His Ile His Asp Phe
Asp 20 25 30 Asp
Val Phe Pro Gly Leu Leu Ile Lys Leu His 35 40
18543PRTArabidopsis thaliana 185Glu Val Lys Glu Gly Glu Thr Leu
Gln Thr Ile Ser Glu Lys Cys Gly 1 5 10
15 Asp Pro Tyr Ile Val Glu Gly Asn Phe His Ile His Asp
His Asp Asp 20 25 30
Leu Phe Pro Gly Leu Leu Ile Arg Ile Thr Pro 35
40 18645PRTPopulus sp. 186His Val Val Lys Glu Gly Glu Thr
Leu Thr Ser Ile Ser Lys Gln Tyr 1 5 10
15 Gly Val Ser Ile Tyr Ser Val Ala Ala Ala Asn Lys Asn
Ile Leu Asp 20 25 30
Val Asp Leu Val Phe Glu Gly Gln Ile Leu Asn Ile Phe 35
40 45 18749PRTPopulus sp. 187His Asp Val Val
Arg Gly Asp Thr Leu Ser Ala Ile Ala Lys Lys Phe 1 5
10 15 Tyr Gly Asp Ala Asn Lys Tyr Pro Val
Ile Phe Glu Ala Asn Lys Phe 20 25
30 Met Leu Ser His Phe Asp Lys Ile Tyr Pro Gly Gln Lys Leu
Arg Ile 35 40 45
Phe 18843PRTPopulus sp. 188Val Gly Ala Ala Ser Gly Asp Thr Cys Phe Thr
Ile Ala Gln Ser Phe 1 5 10
15 Asn Leu Thr Ala Ala Ser Phe Asp Ala Ile Asn Phe Asn Leu Asn Cys
20 25 30 Thr Ala
Leu Phe Val Gly Gln Trp Leu Cys Val 35 40
18946PRTPopulus sp. 189Val Val Gly Val Ala Ser Gly Asp Thr Cys Phe
Thr Ile Ala Gln Ser 1 5 10
15 Phe Asn Leu Thr Ala Ala Ser Phe Asp Ala Ile Asn Phe Asn Ile Ser
20 25 30 Cys Asn
Ala Leu Phe Val Gly Gln Trp Leu Cys Val Ala Gly 35
40 45 19043PRTPopulus sp. 190His Gly Val Val Thr
Gly Asp Thr Cys Ile Ala Val Glu Lys Gln Phe 1 5
10 15 Asp Leu Thr Ala Asn Asp Phe Leu Ala Ile
Asn Phe Asn Leu Asp Cys 20 25
30 Asp Lys Leu Phe Val Gly Gln Trp Leu Cys Val 35
40 19145PRTGlycine max 191Glu His Gln Val Ser
Lys Leu Asp Thr Leu Ala Gly Val Ala Ile Lys 1 5
10 15 Tyr Gly Val Glu Val Ala Asp Ile Lys Arg
Met Asn Gly Leu Ala Thr 20 25
30 Asp Leu Gln Met Phe Ala Leu Lys Thr Leu Lys Ile Phe
35 40 45 19246PRTGlycine max 192His
Gln Val Ser Lys Leu Asp Thr Leu Ala Gly Val Ala Ile Lys Tyr 1
5 10 15 Gly Val Glu Val Ala Asp
Ile Lys Arg Met Asn Gly Leu Ala Thr Asp 20
25 30 Leu Gln Met Phe Ala Leu Lys Thr Leu Lys
Ile Pro Leu Pro 35 40 45
19345PRTPopulus sp. 193Glu His Gln Val Ser Lys Arg Asp Thr Leu Ala Gly
Val Ala Ile Lys 1 5 10
15 Tyr Gly Val Glu Val Ala Asp Val Lys Arg Leu Asn Gly Leu Ser Thr
20 25 30 Asp Leu Gln
Met Phe Ala Leu Lys Thr Leu Leu Ile Pro 35 40
45 19444PRTOryza sativa 194His Arg Val Gly Lys Leu Asp Thr
Leu Ala Gly Ile Ala Ile Lys Tyr 1 5 10
15 Gly Val Glu Val Ala Asp Ile Lys Arg Leu Asn Gly Leu
Ser Thr Asp 20 25 30
Leu Gln Met Phe Ala His Lys Thr Leu Arg Ile Pro 35
40 19547PRTGlycine max 195Asp His Arg Val Ser Lys
Phe Asp Thr Leu Ala Gly Val Ala Ile Lys 1 5
10 15 Tyr Gly Val Glu Val Ala Asp Ile Arg Lys Met
Asn Asn Leu Val Thr 20 25
30 Asp His Gln Met Phe Ala Leu Lys Thr Leu His Ile Phe Leu Pro
35 40 45 19650PRTGlycine
max 196Gly Tyr Ile Glu His His Val Ser Lys Phe Asp Thr Leu Ala Gly Val 1
5 10 15 Ala Ile Lys
Tyr Gly Val Glu Val Ala Asp Ile Arg Lys Met Asn Ser 20
25 30 Leu Val Thr Asp His Gln Met Phe
Ala Leu Lys Thr Leu His Ile Phe 35 40
45 Leu Pro 50 19744PRTArabidopsis thaliana 197His
Arg Ile Ser Lys Phe Asp Thr Leu Ala Gly Val Ala Ile Lys Tyr 1
5 10 15 Gly Val Glu Val Ala Asp
Val Lys Lys Met Asn Asn Leu Val Thr Asp 20
25 30 Leu Gln Met Phe Ala Leu Lys Ser Leu Gln
Ile Phe 35 40 19850PRTPopulus
sp. 198Gly Phe Ile Glu His Pro Val Ser Lys Leu Asp Thr Leu Ala Gly Val 1
5 10 15 Ala Ile Lys
Tyr Gly Val Glu Val Ala Asp Ile Lys Lys Met Asn Gly 20
25 30 Leu Val Thr Asp Leu Gln Met Phe
Ala Leu Lys Ser Leu Gln Ile Phe 35 40
45 Leu Pro 50 19945PRTArabidopsis thaliana 199Glu
His Arg Val Ser Lys Phe Asp Thr Leu Ala Gly Ile Ala Ile Lys 1
5 10 15 Tyr Gly Val Glu Val Ala
Asp Ile Thr Lys Leu Asn Gly Leu Val Thr 20
25 30 Asp Leu Gln Met Phe Ala Leu Glu Ser Leu
Arg Ile Phe 35 40 45
20044PRTGlycine maxmisc_feature(41)..(41)Xaa can be any naturally
occurring amino acid 200Ser His His Ile Thr Arg Gly Asp Thr Val Ala Ser
Leu Ala Val Lys 1 5 10
15 Tyr Ser Val Gln Val Met Asp Ile Lys Arg Leu Asn Asn His His Ser
20 25 30 Asp His Gly
Ile Tyr Ser Arg Glu Xaa Ile Asp Phe 35 40
20144PRTArabidopsis thaliana 201His Arg Ile Cys Arg Gly Asp Ser
Val Thr Ser Leu Ala Val Lys Tyr 1 5 10
15 Ala Val Gln Val His Asp Ile Lys Arg Leu Asn Asn His
His Ser Asp 20 25 30
His Gly Ile Tyr Ser Arg Asp Arg Leu Leu Ile Phe 35
40 20245PRTPopulus sp. 202Ser His Lys Ile Val Lys
Gly Asp Ser Val Ala Ser Leu Ala Val Lys 1 5
10 15 Tyr Ser Val Gln Val His Asp Ile Lys Arg Ile
Asn Asn His Thr Ser 20 25
30 Asp His Gly Ile Asn Ser Arg Glu Arg Leu Leu Ile Phe
35 40 45 20345PRTPopulus sp. 203Ser
His Lys Ile Ala Arg Glu Asp Ser Val Thr Ser Leu Ala Lys Lys 1
5 10 15 Tyr Ser Val Gln Val Arg
Asp Ile Lys Leu Leu Asn Asn His Thr Ser 20
25 30 Asp Asn Gly Ile Tyr Ser Met Glu Arg Leu
Leu Ile Phe 35 40 45
20444PRTOryza sativa 204His Thr Val Arg Arg Gly Asp Thr Val Phe Gly Ile
Ala Leu Lys Tyr 1 5 10
15 Ser Ile Gln Val Thr Asp Ile Lys Arg Phe Asn Asn His His Ser Asp
20 25 30 His Gly Ile
Tyr Leu Arg Glu Arg Leu Leu Ile Phe 35 40
20546PRTPopulus sp. 205Ile Lys Ala Arg Lys Arg Asp Thr Leu Ile
Ser Val Ala Asn Arg Tyr 1 5 10
15 Gly Val Ser Ala Ser Asn Leu Ala Asp Trp Asn Asp Leu Lys Ser
Ser 20 25 30 Ala
Thr Leu His Ala Gly Gln Ser Leu Val Ala Tyr Leu Phe 35
40 45 20645PRTPopulus sp. 206Tyr Tyr Val Trp
Gln Gly Ser Asn Leu Thr Tyr Ile Ser Thr Ile Phe 1 5
10 15 Asn Gln Ser Ile Thr Glu Ile Ile Arg
Tyr Asn Phe Lys Val Pro Asn 20 25
30 Gln Asp Ser Ile Arg Ser Asp Thr Arg Leu Asn Val Phe
35 40 45 207112PRTOryza sativa
207Tyr Ser Ala Arg Pro Ala Asp Thr Leu Ala Ser Val Ala Asp Val Val 1
5 10 15 Phe Ala Gly Lys
Ala Ser Ala Asp Gln Ile Arg Arg Ala Asn Gly Leu 20
25 30 Ser Ala Glu Asp Pro Asp Ala Pro Leu
Asp Ala Gly Ala Thr Leu Val 35 40
45 Val Pro Leu Pro Cys Ala Cys Phe Asn Ser Thr Asp Asn Asn
Leu Pro 50 55 60
Ala Val Tyr Leu Ser Tyr Val Val Arg Val Gly Asp Thr Val Gln Ser 65
70 75 80 Ile Ala Ala Thr His
Ala Thr Thr Val Thr Asp Ile Ser Asn Val Asn 85
90 95 Ala Met Gly Ser Pro Ile Val Ala Pro Gly
Asp Ile Leu Ala Ile Pro 100 105
110 208112PRTOryza sativa 208Tyr Ala Ala Arg Pro Ala Asp Thr
Leu Ala Thr Val Ala Asp Gly Val 1 5 10
15 Phe Ala Gly Leu Ala Phe Ala Asp Gln Ile Arg Asn Ala
Asn Ala Val 20 25 30
Ala Ser Ala Asp Pro Asp Ala Pro Leu Asp Pro Gly Gln Lys Leu Val
35 40 45 Val Pro Leu Pro
Cys Val Cys Phe Asn Ser Ser Asp Asn Asn Leu Pro 50
55 60 Ala Val Tyr Leu Ser Tyr Val Val
Gln Val Gly Asp Thr Val Pro Ala 65 70
75 80 Ile Ala Ala Ser Tyr Glu Thr Thr Val Thr Asp Val
Met Asn Val Asn 85 90
95 Ala Met Gly Ser Pro Ile Ala Ala Pro Gly Asp Ile Leu Ala Ile Pro
100 105 110
209112PRTOryza sativa 209Tyr Val Ala Arg Pro Gly Asp Thr Leu Ala Ser Val
Ala Ser Ser Val 1 5 10
15 Tyr Gly Gly Leu Thr Thr Pro Asp Trp Ile Ser Asp Ser Asn Gly Ile
20 25 30 Leu Gly Ala
Lys Pro Asp Ala Ala Val Asp Ala Gly Thr Thr Leu Phe 35
40 45 Val Pro Leu His Cys Ala Cys Phe
Gly Gly Val Asp Asn Gly Leu Pro 50 55
60 Ala Val Tyr Leu Thr Tyr Val Ala Gly Lys Gly Asp Thr
Val Ala Ala 65 70 75
80 Val Ala Gln Arg Tyr Arg Thr Thr Ala Thr Asp Leu Met Ser Val Asn
85 90 95 Asp Met Ala Thr
Pro Glu Leu Ala Ala Gly Asp Ile Ile Val Val Pro 100
105 110 210107PRTOryza sativa 210His Ile
Ile Gln Ile Asn Asp Asn Met Ser Tyr Ile Ala Ala Gln Phe 1 5
10 15 Asp Asp Phe Val Thr Tyr Gln
Glu Ile Ala Ala Ala Ser Asn Ile Ser 20 25
30 Asn Pro Asp Phe Leu Glu Val Gly Gln Glu Leu Trp
Ile Pro Leu Pro 35 40 45
Cys Ser Cys Asp Gln Val Glu Gly Asn Asn Val Thr His Phe Ala Tyr
50 55 60 Lys Val Arg
Ala Ala Asp Asn Val Ser Lys Ile Ala Ala Arg Phe Gly 65
70 75 80 Val Lys Glu Ser Thr Leu Leu
Lys Ile Asn Gly Ile Thr Asp Pro Lys 85
90 95 Asn Leu Thr Gln Gly Gln Ile Leu Asp Val Pro
100 105 211108PRTOryza sativa 211Tyr
Arg Ile Arg Ala Gly Asp Thr Leu Asp Ala Ile Ala Arg Gly Val 1
5 10 15 Phe Ala Gly Leu Val Thr
Tyr Gln Asp Ile Ala Ala Ala Asn Asn Val 20
25 30 Ser Asp Pro Asn Lys Ile Ala Val Gly Gln
Glu Leu Trp Ile Pro Val 35 40
45 Pro Cys Ser Cys Asp Pro Val Ala Gly Gln Pro Val Val His
Tyr Thr 50 55 60
Tyr Val Val Pro Pro Gly Ala Ser Val Ala Ala Ile Ala Gln Asp Phe 65
70 75 80 Ala Thr Thr Glu Ala
Thr Val Leu Ala Leu Asn Arg Met Pro Asp Ala 85
90 95 Lys Ser Leu Leu Ala Gly Gln Val Leu Asp
Val Pro 100 105 21238PRTOryza
sativa 212His Gly Val Gln Ala Ser Glu Thr Cys Phe Ser Val Ser Gln Ser Ala
1 5 10 15 Gly Leu
Thr Gln Asp Gln Phe Leu Ala Phe Asn Pro Asn Ile Asn Cys 20
25 30 Ala Lys Val Phe Val Gly
35 21336PRTOryza sativa 213His Gly Val Glu Ala Gly Glu
Thr Cys Asp Ser Ile Ala Arg Arg Phe 1 5
10 15 His Ala Gly Leu Gly Arg Ala Pro Phe Phe Arg
Leu Val Ser Leu Asn 20 25
30 Pro Asn Ile Asn 35 21444PRTOryza sativa 214Tyr
Ala Ala Arg Pro Ala Asp Thr Leu Ala Thr Val Ala Asp Gly Val 1
5 10 15 Phe Ala Gly Leu Ala Phe
Ala Asp Gln Ile Arg Asn Ala Asn Pro Asp 20
25 30 Ala Pro Leu Asp Pro Gly Gln Lys Leu Val
Val Pro 35 40 21542PRTOryza
sativa 215His Gln Val Ser Arg Met Asp Thr Leu Pro Gly Leu Ala Ile Lys Tyr
1 5 10 15 Gly Val
Glu Ile Ser Asp Ile Lys Arg Ala Asn Ser Leu Met Thr Asp 20
25 30 Ser Gln Met Phe Ala His Lys
Met Leu Leu 35 40
User Contributions:
Comment about this patent or add new information about this topic:
| People who visited this patent also read: | |
| Patent application number | Title |
|---|---|
| 20160189672 | System for Accurately and Precisely Representing Image Color Information |
| 20160189671 | ORGANIC LIGHT EMITTING DISPLAY DEVICE |
| 20160189670 | ORGANIC LIGHT EMITTING DISPLAY DEVICE |
| 20160189669 | DISPLAY PANEL |
| 20160189668 | MULTI EMBEDDED TIMING CONTROLLER, DISPLAY PANEL, AND COMPUTER SYSTEM HAVING THE SAME |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
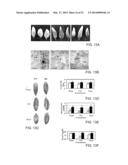 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |
| New patent applications in this class: | |
| Date | Title |
|---|---|
| 2018-01-25 | Methods for plant transformation using spectinomycin selection |
| 2017-08-17 | Insecticidal polypeptides having broad spectrum activity and uses thereof |
| 2017-08-17 | Process for the prevention and suppression of bacterial diseases in plants |
| 2017-08-17 | Insecticidal polypeptides having broad spectrum activity and uses thereof |
| 2016-12-29 | Novel insecticidal proteins and methods of use |
| New patent applications from these inventors: | |
| Date | Title |
|---|---|
| 2014-02-06 | Metabolic engineering of plants for increased homogentisate and tocochromanol production |
| 2013-03-28 | Protozoan glycosidases and related methods |
| 2012-08-02 | Soybean transcription factors and other genes and methods of their use |
| 2012-05-03 | Lysm receptor-like kinases to improve plant defense response against fungal pathogens |
| Top Inventors for class "Multicellular living organisms and unmodified parts thereof and related processes" | |
| Rank | Inventor's name |
|---|---|
| 1 | Gregory J. Holland |
| 2 | William H. Eby |
| 3 | Richard G. Stelpflug |
| 4 | Laron L. Peters |
| 5 | Justin T. Mason |