Patent application title: R2R1/2 In Diagnosis and Therapy
Inventors:
Jeroen Marcel Maria Roger Aerssens (Nieuwrode, BE)
Ronald Karel Louisa Van Brempt (Rc Leiden, NL)
Pieter Johan Peeters (Dessel, BE)
Ronald Anthonius De Hoogt (Turnhoutseweg 30, BE)
Marcus Cornelis De Ruiter (Rc Leiden, NL)
Assignees:
JANSSEN PHARMACEUTICA NV
Academisch Ziekenhuis Leiden A/U Leiden Uni Medical Ctr
IPC8 Class: AG01N3368FI
USPC Class:
800 9
Class name: Multicellular living organisms and unmodified parts thereof and related processes nonhuman animal the nonhuman animal is a model for human disease
Publication date: 2014-03-06
Patent application number: 20140068794
Abstract:
The present invention stems from the finding that two genes designated
R2R1 and R2R2, play important roles in tissue development and
cancer biology. In particular, the inventors have discovered that these
two genes are expressed in pulmonary cells and are required for late
branching morphogenesis of pulmonary epithelium and endothelium and
support the development/maintenance of the refined three dimensional
architecture of the lung. These genes are essential in the squamous
differentiation program and development/maintenance of the progenitor
(Krt14 expressing) cell pool. Moreover, the inventors have identified
crucial roles for these genes in cancer biology, particularly processes
associated with the acquisition of an immortal and metastatic phenotype
(including cancer progression and metastasis) and pulmonary and cardiac
development. Accordingly, the invention provides compounds and methods
for use in the treatment of cardiac and pulmonary diseases and well as in
cancer.Claims:
1-12. (canceled)
13. An antibody or antigen binding fragment thereof capable of binding to an R2R1/2 protein or a fragment thereof.
14. The antibody of claim 13, wherein the R2R1/2 protein is encoded by a sequence selected from the group consisting of those designated SEQ ID NOS: 3, 6, 9 and 12, or a sequence at least 60% identical thereto.
15. A method of treating one or more diseases and/or conditions selected from the group consisting of: (a) cardiac diseases and/or conditions; (b) pulmonary diseases and/or conditions; and (c) cancer; said method comprising administering a therapeutically effective amount of: (i) a polynucleotide comprising an R2R1/2 gene, wherein the R2R1/2 gene is encoded by a sequence selected from the group consisting of those designated SEQ ID NOS: 1, 2, 4, 5, 7, 8, 10 and 11, or a sequence at least 60% identical thereto; (ii) a polypeptide comprising an R2R1/2 protein, wherein the R2R1/2 protein is encoded by a sequence selected from the group consisting of those designated SEQ ID NOS: 3, 6, 9 and 12 or a sequence at least 60% identical thereto; (iii) an antisense oligonucleotide capable of modulating the expression of an R2R1/2 gene; (iv) the antibody of claim 13; or (v) the antibody of claim 13, wherein the R2R1/2 protein is encoded by a sequence selected from the group consisting of those designated SEQ ID NOS: 3, 6, 9 and 12, or a sequence at least 60% identical thereto; to a subject in need thereof.
16. A method of diagnosing one or more diseases and/or conditions selected from the group consisting of: (a) a cardiac disease and/or condition; (d) a pulmonary disease and/or condition; (c) cancer; and (d) a susceptibility or predisposition to any of (a) to (c); said method comprising the step of detecting a level of R2R1 and/or R2R2 gene expression in a sample provided by a subject to be tested, wherein detection of an aberrant level of the R2R1 and/or R2R2 gene in said sample, is indicative of one or more of the diseases and/or conditions provided as (a)-(c) above and/or a susceptibility or predisposition thereto.
17. The method of claim 16, wherein the method comprises the use of an oligonucleotide probe and/or primer designed to hybridise under stringent conditions to all or part of a sequence selected from the group consisting of SEQ ID NOS: 1; 2; 4; 5; 7; 8; 10 and 11.
18. A method of identifying or obtaining agents which modulate the expression of an R2R1 and/or R2R2 genes, said method comprising the steps of contacting the R2R1 and/or R2R2 genes with a test agent and detecting any modulation of R2R1/2 gene expression.
19. A pharmaceutical composition comprising (i) an R2R1/2 gene provided by SEQ ID NOS: 1, 2, 4, 5, 7, 8, 10 or 11, or a sequence at least 60% identical thereto, in association with a pharmaceutically acceptable excipient, carrier or diluent; or (ii) an R2R1/2 protein provided by SEQ ID NOS: 3, 6, 9 and 12, or a sequence at least 60% identical thereto, in association with a pharmaceutically acceptable excipient, carrier or diluent.
20. An animal model for the study of a cardiac and/or pulmonary disease or condition, or cancer, wherein the animal model has been generated by modulation or disruption of an R2R1/2 gene.
Description:
FIELD OF THE INVENTION
[0001] The present invention provides novel genes that support the development/maintenance of the refined three-dimensional architecture of the lung as well as medicaments and compositions for treating pulmonary diseases and/or conditions.
BACKGROUND OF THE INVENTION
[0002] The expression of intermediate filament (Krt6), EDC (Epidermal Differentiation Complex) and SCC (Stratified epithelium-secreted protein gene complex) genes permits cellular survival in hostile environments. This expression profile leads to squamous differentiation and is a hallmark of skin epithelial cells. The inventors found that this transcriptional program is also vital for the refinement of pulmonary architecture (late branching morphogenesis): cells in distal airways and blood vessels need to assume a flat shape and acquire mechanical flexibility in an oxygen-rich environment. Proteins encoded by the intermediate filament group and associated EDC and SCC genes permit this kind of cell shape and flexibility. At the same time, the lung needs to maintain progenitor cells that are able to differentiate in cells capable of building such proteins. These progenitor or basal cells typically express the intermediate filament gene Krt14.
[0003] The lung is exposed to vast amounts of mechanical and oxidative stress and resembles the skin in this respect. The squamous differentiation program as mentioned above is the primary line of defence. Cells that line the distal airways and blood vessels of the lung need to withstand this stress and need to be replaced in case of cell death by a pool of progenitor cells.
Two human pathologies stand out by failure of this system.
[0004] 1. Bronchopulmonary dysplasia (BPD): Lungs of infants, which are born prematurely, are more susceptible to lung injury such as mechanical stress. Also, lungs of premature infants surviving lung injury often heal with significant scarring or bronchopulmonary dysplasia (BPD). These lungs have a limited or underdeveloped regenerative potential.
[0005] 2. Chronic obstructive pulmonary disease (COPD): Lungs of adult COPD patients seem to respond inappropriately to noxious stimuli such as smoking. Although they develop a barrier against these stimuli, their lack of regenerative capabilities in case of cell death leads to deformation of lung airways and blood vessels. Both BPD and COPD lead to significant morbidity and mortality.
SUMMARY OF THE INVENTION
[0006] The present invention stems from the finding that two genes play important roles in tissue development and cancer biology. In particular, the inventors have discovered that these two genes are expressed in pulmonary cells and are required for late branching morphogenesis of pulmonary epithelium and endothelium and support the development/maintenance of the refined three dimensional architecture of the lung. These genes are essential in the squamous differentiation program and development/maintenance of the progenitor (Krt14 expressing) cell pool. Moreover, the inventors have identified crucial roles for these genes in cancer biology, particularly processes associated with the acquisition of an immortal and metastatic phenotype (including cancer progression and metastasis) and cardiac development.
[0007] The inventors have ascertained the sequences of the murine and human forms of these genes. In view of their co-ordinated expression and function as regenerative genes for respiratory cells, the inventors have designated these genes R2R1 and R2R2. For simplicity, the bulk of this specification will use the term "R2R1/2" which is intended to represent the longer phrase "R2R1 and/or R2R2"
[0008] Accordingly, references to the R2R1/2 are to be understood as encompassing all mammalian forms of these genes including, for example, human and rodent (mouse, rabbit, guinea pig, rat etc) forms. Furthermore, in addition to encompassing the entire R2R1 and/or R2R2 gene sequences, it is to be understood that these designations also encompass fragments, portions, mutants, derivatives and/or homologous/orthologues of any of the genes described herein. In this regard, it should be understood that the term "R2R1" encompasses the murine sequence encoded by the cDNA sequences designated 2200001115Rik or RIKEN cDNA 2200001I15 as well as the seven human homologues designated FAM25A, FAM25B, FAM25C, FAM25D, FAM25E, FAM25G and FAM25HP.
[0009] In addition, where appropriate, the term "R2R1/2" encompasses the proteinaceous products of the R2R1 and/or R2R2 genes or fragments or portions thereof.
[0010] In particular, the term "R2R1/2" or indeed either of the terms "R2R1" or "R2R2", encompass the sequences given as SEQ ID NOS: 1-12 below or fragments, portions, analogues, variants or derivatives thereof.
[0011] The sequence of an exemplary transcript of the murine R2R1 gene is given below as SEQ ID NO: 1
TABLE-US-00001 SEQ ID NO: 1 acactgacacggaccgaaggagtggaaaaagctttacctgtcactgtct gctgccatacgATGCTGGGAGGCCTGGGGAAGCTGGCGGCCGAGGGCCT GGCCCACCGCACAGAGAAAGCCACTGGGGGAGCAGTTCACGCAGTGGAA GAGGTGGTGAGCGAGGTGGTGGGCCACGCCAAGGAGGTTGGAGAGAAGA CCATTAATGACGCCCTAAAGAAAGCCCAAGAATCAGGAGACAGGGTGGT GAAGGAGGTCACTGAGAAGGTCACCCACACCATCACTGATGCTGTTACC CATGCGGCAGAAGGCCTGGGAAGACTGGGACAGtgagcctgcctaccag catggctggcccttcctgaaggtcaataaagagtgtgaaacgtgaaaaa aaaaaaaaaaataacaaaaaaaaaaaaaaaaaa
[0012] The coding or translated part of this sequence is underlined and comprises some 267 nucleotides. This particular portion of SEQ ID NO: 1 has been designated SEQ ID NO: 2.
[0013] One of skill in this field will appreciate that the 267 translated nucleotides yield a protein comprising 89 amino acids and having the following sequence (designated SEQ ID NO: 3)
TABLE-US-00002 SEQ ID NO: 3 MLGGLGKLAAEGLAHRTEKATGGAVHAVEEVVSEVVGHAKEVGEKTIND ALKKAQESGDRVVKEVTEKVTHTITDAVTHAAEGLGRLGQ
[0014] In addition, the inventors have ascertained the complete sequence of an exemplary human transcript of the R2R1 gene and this is given as SEQ ID NO: 4 below:
TABLE-US-00003 SEQ ID NO: 4 actgtctgctgccacacgATGCTGGGAGGCCTGGGGAAGCTGGCTGCCG AAGGCCTGGCCCACCGCACCGAGAAGGCCACCGAGGGAGCCATTCATGC CGTGGAAGAAGTGGTGAAGGAGGTGGTGGGACACGCCAAGGAGACTGGA GAGAAAGCCATTGCTGAAGCCATAAAGAAAGCCCAAGAGTCAGGGGACA AAAAGATGAAGGAAATCACTGAGACAGTGACCAACACAGTCACAAATGC CATCACCCATGCAGCAGAGAGTCTGGACAAACTTGGACAGtgagtgcac ctgctaccacggcccttccccagtctcaataaaaagccatgacatgtg
[0015] The coding or translated part of this sequence is underlined and comprises some 267 nucleotides. This particular portion of SEQ ID NO: 4 has been designated SEQ ID NO: 5.
[0016] One of skill in this field will appreciate that the 267 translated nucleotides yield a protein comprising 89 amino acids and having the following sequence (designated SEQ ID NO: 6)
TABLE-US-00004 SEQ ID NO: 6 MLGGLGKLAAEGLAHRTEKATEGAIHAVEEVVKEVVGHAKETGEKAIAE AIKKAQESGDKKMKEITETVTNTVTNAITHAAESLDKLGQ
[0017] The sequence of an exemplary transcript of the murine R2R2 gene is given below as SEQ ID NO: 7
TABLE-US-00005 SEQ ID NO: 7 Gtgactggctgctgtctctagttgttgaggcctcttgggatctyggcgc tmacmccwtgctytagwgactccgatagctcccrmggctccagtgsasm cctcggkcggnggnagggaaaaggcacttgctggtagctctgctcaccc gcactgggacctggagctggaggactaagaagacagacggctgctgctt gccacagcctggaccATGGACCCCCATGAGATGGTTGTGAAGAATCCAT ATGCCCACATCAGCATTCCTCGGGCTCACCTGCGCTCTGACCTGGGGCA GCAGTTAGAGGAGGTTCCTTCTTCATCTTCCTCCTCTGAGACTCAGCCT CTGCCTGCAGGAACATGTATCCCAGAGCCAGTGGGCCTCTTACAAACTA CTGAAGCCCCTGGGCCCAAAGGTATCAAGGGCATCAAGGGTACTGCTCC TGAGCACGGCCAGCAGACCTGGCAGTCACCCTGCAATCCCTATAGCAGT GGGCAACGTCCATCGGGACTGACTTATGCTGGCCTGCCACCTGTAGGGC GTGGTGATGACATTGCCCACCACTGCTGCTGCTGCCCTTGCTGCTCCTG CTGCCACTGTCCTCGCTTCTGCCGTTGTCACAGCTGTTGTGTTATCTCC tagctgactattgaacctccagggctgtgcagcccaggttcctgctcaa tgccaaagtgttgctggacatcaggagcagccgttgtcatgagcatcag ccatttcctgccctgagcaggggagcctgtccaccagcgttcagctgta gccttctggaatagggttccagccactagccatgttggcaacaacaggg acacccttcacgtcctgcaagactttggcaataaagcaggatgagcgtt gctgnncctgntgaaaanaaamwaaawacwgccgttgtcacarcygttr tgttatctmmkagstgacwattgtaammtycagrgctgtrmagcccrgg kksckgctcaatgccaaagtgttgmtgsmcmtcrggrgsrgccaagctt tacgcggtacccgggattttttttgtacaaaaaggggccccctattagg
[0018] The coding or translated part of this sequence is underlined and comprises some 426 nucleotides. This particular portion of SEQ ID NO: 7 has been designated SEQ ID NO: 8.
[0019] One of skill in this field will appreciate that the 426 translated nucleotides yield a protein comprising 142 amino acids and having the following sequence (designated SEQ ID NO: 9)
TABLE-US-00006 SEQ ID NO: 9 MDPHEMVVKNPYAHISIPRAHLRSDLGQQLEEVPSSSSSSETQPLPAGT CIPEPVGLLQTTEAPGPKGIKGIKGTAPEHGQQTWQSPCNPYSSGQRPS GLTYAGLPPVGRGDDIAHHCCCCPCCSCCHCPRFCRCHSCCVIS
[0020] In addition, the inventors have ascertained the complete sequence of an exemplary human transcript of the R2R2 gene and this is given as SEQ ID NO: 10 below:
TABLE-US-00007 SEQ ID NO: 10 cttgaacccgggaggcagaggttgcagtgagccgagatcgcgcagctgc actccagcctgggcaacagagcaagactccatctcagaaaagaagcaga aagcctccaagagccaatggctctcaagcatcttggtctctgctaagaa gaggctcagaggcttagaagccctgcctcgccggggctttgaggtgtgt gagcaatggctggggactgcaggcccgggaatctgagggcctcacccca cttcctttccagagccgtgacctcaggctcacctcctgccctcctctca ggcaagctgcagatgccctttagggcccaggccatgccccggatgtgag gggctgagtcactggtttggcagtgcccctcagagcccaggcctgggct gccacccacctgaggacgagggctgggccagctgtcgtgctccagttgc tggggcctcttgggatcttgggaaccccatctctgagccccgccccATG GCCCCGCCCCTCCCAAGGAGGGAAAAGGCGGCTGCCAGTCGCTCAACTC AGGCACTGGGACCTAGAGCTCAGAAGACCGAGAGGACAGACTGCCGTGT TGCCACCACAGGCTGGACCATGGACCCCCAAGAGATGGTCGTCAAGAAC CCATATGCCCACATCAGCATCCCCCGGGCTCACCTGCGGCCTGACCTGG GGCAGCAGTTAGAGGTGGCTTCCACCTGTTCCTCATCCTCGGAGATGCA GCCCCTGCCAGTGGGGCCCTGTGCCCCAGAGCCAACCCACCTCTTGCAG CCGACCGAGGTCCCAGGGCCCAAGGGCGCCAAGGGTAACCAGGGGGCTG CCCCCATCCAGAACCAGCAGGCCTGGCAGCAGCCTGGCAACCCCTACAG CAGCAGTCAGCGCCAGGCCGGACTGACCTACGCTGGCCCTCCGCCCGCG GGGCGCGGGGATGACATCGCCCACCACTGCTGCTGCTGCCCCTGCTGCC ACTGCTGCCACTGCCCCCCCTTCTGCCGCTGCCACAGCTGCTGCTGCTG TGTCATCTCCtagcccagcccaccctgccagggccaggacccagacttc agcaaatgtggctcacacagtgccgggacatgccgggacatgcggggtg gctgttgtcatgggcgtctgccccttcacaccaggcactggggctcaga cccaccaggaaggtggccgttcagcccgagctcctgaaacggaatccca ggtcctggctggagagggacacccctgattaccttaaggcccaggcaat aaagcagggtgatcttc
[0021] The coding or translated part of this sequence is underlined and comprises some 552 nucleotides. This particular portion of SEQ ID NO: 10 has been designated SEQ ID NO: 11.
[0022] One of skill in this field will appreciate that the 552 translated nucleotides yield a protein comprising 184 amino acids and having the following sequence (designated SEQ ID NO: 12)
TABLE-US-00008 SEQ ID NO: 12 MAPPLPRREKAAASRSTQALGPRAQKTERTDCRVATTGWTMDPQEMVVK NPYAHISIPRAHLRPDLGQQLEVASTCSSSSEMQPLPVGPCAPEPTHLL QPTEVPGPKGAKGNQGAAPIQNQQAWQQPGNPYSSSQRQAGLTYAGPPP AGRGDDIAHHCCCCPCCHCCHCPPFCRCHSCCCCVIS
[0023] As such, the present invention relates to the genes encoded by the sequences designated as SEQ ID NOS: 1, 2, 4, 5, 7, 8, 10 and 11, as well as any fragments, portions, mutants, variants, derivatives and/or homologues/orthologues thereof. Typically, fragments, portions, mutants, variants, derivatives and/or homologues/orthologues are functional or active--that is, they retain the function of the wild type R2R1/2 genes.
[0024] The term "mutants" may encompass naturally occurring mutants or those artificially created by the introduction of one or more nucleic acid additions, deletions, substitutions or inversions.
[0025] One of skill in this field will readily understand that genes homologous to the human and murine R2R1/2 genes detailed above may be found in a number of different species, including, for example, other mammalian species. Homologous genes may exhibit as little as approximately 20 or 30% sequence homology or identity however, in other cases, homologous genes may exhibit at least 40, 50, 60, 65 70, 75, 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99% homology to the various nucleotide sequences given above. As such, homologous genes from other species are to be included within the scope of this invention.
[0026] Using the various nucleic acid and amino acid sequences described herein, one of skill in the art could readily identify related sequences in other species, such as other mammals etc. For example, nucleic acid obtained from a particular species may be probed using the probes described herein, for homologous or closely related sequences.
[0027] In addition, it should be understood that the present invention also relates to the products of the genes encompassed by this invention and in particular the peptides encoded by SEQ ID NOS: 3, 6, 9 and 12. Furthermore, fragments, portions, analogues, variants, derivatives of any of these or homologous and/or identical proteins are also within the scope of this invention. Typically, fragments, portions, derivatives, variants and/or homologues are functional or active--that is they retain the function of the wild type R2R1/2 protein.
[0028] In addition, proteins, polypeptides/peptides homologous/identical to any of the proteins encoded by SEQ ID NOS: 3, 6, 9 and 12 are also within the scope of this invention. Protein or polypeptide/peptide sequences which are considered as homologous or identical to any of the sequences described herein may exhibit as little as 20% or 30% sequence identity/homology. However, homologous/identical sequences may be at least 40, 50, 60, 65, 70, 75, 80, 85, 90, 95, 96, 97, 98 or 99% homologous or identical. Insofar as the invention relates to fragments of any of the protein or polypeptide/peptide sequences described herein, it should be understood that a fragment may comprise anywhere between about 10 and n-1 amino acids (where "n" is the number of amino acids in the complete sequence). For example, a fragment may comprise about 10, 15, 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120 or about 125 amino acids (the maximum number of amino acids being determined by the number of amino acids in the complete sequence)--such fragments/portions may be referred to as peptide fragments. In one embodiment, the peptide fragments may be antigenic and/or immunogenic--that is to say, they retain the ability to bind antibodies exhibiting specificity, affinity and/or selectivity for the native (complete) antigen--such as those encoded by SEQ ID NOs: 3, 6, 9 and 12.
[0029] One of skill in this field will readily understand that for the various nucleic acid sequences and polypeptides described herein, natural variations due to, for example, polymorphisms, may exist between R2R1/2 genes and proteins isolated from any given species. In addition, it is well known in the art, that the degeneracy of the genetic code permits substitution of one or more bases in a codon without alteration to the primary amino acid sequence. As such, genetic degeneracy may be exploited in order to yield variant nucleic acid sequences which encode peptide or protein sequences substantially identical to the antigen sequences described herein. Indeed, variant sequences provided by this invention may manifest as proteins and/or genes that exhibit one or more amino acid/nucleic acid substitutions, additions, deletions and/or inversions relative to a reference sequence (for example any of the sequences described above).
[0030] As such, it is to be understood that all such variants, especially those that are functional or display the desired activity, are to be included within the scope of this invention.
[0031] In other embodiment, the invention relates to derivatives of any of the R2R1/2 sequences described herein. The term "derivatives" may encompass R2R1/2 gene or peptide sequences which, relative to those described herein, comprise one or more amino acid substitutions, deletions, additions and/or inversions.
[0032] Additionally, or alternatively, analogues of the various peptides described herein may be produced by introducing one or more conservative amino acid substitutions into the primary sequence. One of skill in this field will understand that the term "conservative substitution" is intended to embrace the act of replacing one or more amino acids of a protein or peptide with an alternate amino acid with similar properties and which does not substantially alter the physcio-chemical properties and/or structure or function of the native (or wild type) protein. Analogues of this type are also encompassed with the scope of this invention.
[0033] As is well known in the art, the degeneracy of the genetic code permits substitution of one or more bases in a codon without changing the primary amino acid sequence. Consequently, although the sequences described in this application are known to encode the R2R1/2 proteins described herein, the degeneracy of the code may be exploited to yield variant nucleic acid sequences which encode the same primary amino acid sequences.
[0034] As stated, the present inventors have discovered that the R2R1/2 genes (and their protein products) are involved in pulmonary and cardiac morphogenesis events (such as cell signalling/transition etc.) and support the development of the three dimensional architecture of the lung and heart. As such, a first aspect of this invention provides the R2R1/2 genes and/or R2R1/2 proteins, for use as medicaments or for use in treating diseases affecting cell/tissue development/structure, differentiation, proliferation and/or morphogenesis. For example, the R2R1/2 genes and/or R2R1/2 proteins may be for use in treating, for example pulmonary and/or cardiac diseases and/or conditions as well as cancer--particularly cancers affecting tissues of the pulmonary or cardiac system.
[0035] In certain embodiments, the invention may provide the R2R1/2 genes and/or R2R1/2 proteins for use in modulating cell transition events such as, for example, mesenchymal to epithelial transition (MET) events and events involved in the reverse process, epithelial to mesenchymal transition (EMT).
[0036] In a second aspect, the present invention provides the R2R1/2 genes and/or R2R1/2 proteins for use in treating pulmonary diseases and/or conditions or their use in the manufacture of a medicament for the treatment of pulmonary diseases and/or conditions.
[0037] In a further aspect, the invention provides the R2R1 gene and/or protein for use in treating cardiac diseases. In one embodiment, the R2R1 gene and/or protein may be used to treat diseases affecting the development/structure, differentiation, proliferation and/or morphogenesis of cardiac cells/tissues. In one embodiment, the invention may provide the R2R1 gene and/or protein for use in treating diseases and/or conditions affecting the development and/or formation of the ventricular septum.
[0038] In a further aspect, the invention provides the R2R1 gene and/or protein for use in treating cancer. In one embodiment, the R2R1 gene and/or protein may be used to treat cancers affecting a variety of tissues including, for example, pulmonary and/or cardiac tissue. More generally, the invention may extend to the treatment of any cancer involving aberrant/defective MET/EMT processes. Examples of at least some of the cancers which may be treated using the genes and/or proteins of the invention are detailed below.
[0039] It is reiterated that the terms "R2R1", "R2R2" and "R2R1/2" encompass not only the complete gene/peptide sequences as described above, but also fragments, analogues, homologues, orthologues, variants and derivatives thereof.
[0040] The terms "pulmonary disease" or "pulmonary condition" may include pathologies of the lung such as those affecting lung development or the 3D architecture of pulmonary tissues. For example, "pulmonary diseases" may include diseases and/or conditions affecting the epithelial cells lining the airways of the lung, the endothelial cells of the pulmonary vessel network and/or the differentiation and/or growth of these cells. As such, pulmonary diseases may include diseases affecting pulmonary morphogenesis pathways and events. Pulmonary diseases and/or conditions affecting the differentiation of certain pulmonary cell types (for example the squamous epithelial cells) or the generation and/or maintenance of progenitor (basal) cell populations may also be treated with the compounds, medicaments and methods described herein. By way of specific example, diseases such as Bronchopulmonary Dysplasia (BPD) and/or Chronic obstructive pulmonary disease (COPD) may be treated using the compounds described herein. In other embodiments, the terms "pulmonary disease" or "pulmonary condition" may include cell proliferation or neoplastic disorders such as cancer, including, for example, non-small cell lung cancer (NSCLC) and/or small cell lung cancer (SCLC).
[0041] The terms "cardiac disease" or "cardiac condition" may include pathologies of the heart such as those affecting cardiac development or the 3D architecture of cardic tissues. Cardiac diseases may include diseases affecting cardiac morphogenesis pathways and events, in particular mesenchymal to epithelial and epithelial to mesenchymal transition. By way of specific example, diseases such as atrial and ventricular septal defects, atrioventricular canal defects, malformation of cardiac (atrioventricular) valves and coronary arteries may be treated using the compounds described herein.
[0042] It should also be understood that since the R2R1/2 genes/proteins described herein have been shown to play a role in morphogenesis events (such as cell signalling etc.) and support the development of the three dimensional architecture of complex tissues such as the lung and/or heart, they may find application in regenerative medicine. By way of example, where stem cells (for example adult, embryionic or reprogrammed somatic cells (such as iPS cells) are used to repair or reconstruct, for example, damaged or diseased tissue, the proteins and/or genes provided by this invention may be used to facilitate tissue development.
[0043] The R2R1/2 protein and genes play an important role in the morphogenesis events of the lung and heart as gatekeepers of epithelial to mesenchymal and mesenchymal to epithelial transition, Therefore, they may find application in cancer biology and cancer treatment. By way of example, diseases such as transformation of localized cancer to cancer metastasis may be treated using compounds described herein.
[0044] In addition to providing various uses and medicaments involving the R2R1/2 genes and/or proteins described herein, the present invention also provides methods of treating subjects suffering from any of the diseases and/or conditions described herein, including any of the cardiac/pulmonary diseases and/or conditions outlined above. As such a third aspect of this invention provides a method of treating a cardiac/pulmonary disease and/or condition comprising the steps of administering to a subject in need thereof, a therapeutically effective amount of the R2R1 and/or R2R2 genes and/or R2R1 and/or R2R2 proteins described herein.
[0045] Where a cardiac or pulmonary disease or condition results from, or is associated with, a lack of or defective, R2R1/2 gene and/or protein expression/function, the use of a functional R2R1/2 gene or protein as described herein, may provide a means of restoring normal (or wild type) R2R1/2 gene/protein function so as to treat and/or alleviating the symptoms of the disease or condition.
[0046] It will be appreciated that the uses, medicaments and methods of treatment described herein may require the generation of recombinant R2R1/2 genes/proteins and as such, the present invention further contemplates methods of generating and/or expressing recombinant R2R1/2 genes and/or proteins. One of skill in this field will appreciate that PCR techniques may be exploited to selectively obtain R2R1/2 gene sequences from a variety of sources including, for example, lung tissue. These sequences may be ligated to various expression or regulatory control sequences such as, for example, promoter, operator, inducer, enhancer and/or silencer elements, ribosome binding sites and/or terminator sequences. Suitable regulatory or expression control sequences may be selected by those skilled in this field for any given host. In a further embodiment, the PCR derived R2R1/2 gene sequences may be introduced into a vector (such as a plasmid or expression cassette). In one embodiment, the vector may further comprise a nucleotide sequence of a tag or label to assist in protein purification procedures.
[0047] A host cell may be transformed with the vector and maintained under conditions suitable to induce expression of the R2R1/2 gene sequence and production of recombinant R2R1/2. Vectors into which R2R1/2 gene sequences (or fragments thereof) have been cloned, may be introduced or transfected into cells using a variety of techniques--such techniques may otherwise be referred to as transfection protocols. Transfection protocols utilise conditions which render cell membranes permeable to compounds such as nucleic acids. By way of example, it may be possible to facilitate the transfection of vectors, including expression vectors, into cells using electroporation, heat shock, chemical compounds such, for example, calcium phosphate, stronitium phosphate, microinjection techniques and/or gene guns.
[0048] Techniques used to purify recombinant proteins generated in this way are known and, where the recombinant protein is tagged or labelled, these may include the use of, for example, affinity chromatography techniques.
[0049] In view of the above, the fourth and fifth aspects of this invention provide an expression vector comprising an R2R1/2 gene sequence and a host cell transformed therewith, respectively.
[0050] In addition to providing the R2R1/2 genes and/or proteins as a means of treating various diseases and/or conditions (for example cardiac and/or pulmonary diseases and/or conditions), the present invention also provides compounds capable of modulating the expression of the R2R1/2 genes and which may be useful in the treatment of conditions that result from, or are associated with R2R1/2 gene/protein over-expression. Such compounds may be oligonucleotides, preferably antisense oligonucleotides which may take the form of, for example DNA and/or RNA. In one embodiment, the oligonucleotides are RNA molecules known to those skilled in this field as small/short interfering and/or silencing RNA and which will be referred to hereinafter as siRNA. Such siRNA oligonucleotides may take the form of native RNA duplexes or duplexes that have been modified in some way (for example by chemical modification) to be nuclease resistant. Additionally, or alternatively, the siRNA oligonucleotides may take the form of short hairpin RNA (shRNA) expression or plasmid constructs that correspond to, or comprise, the siRNAs described herein.
[0051] The oligonucleotides provided by this invention may be designed to modulate the expression the R2R1/2 genes. By analysing native or wild-type R2R1/2 sequences and with the aid of algorithms such as BIOPREDsi, one of skill in the art could easily determine or computationally predict nucleic acid sequences that have an optimal knockdown effect for these genes (see for example: http://www.biopredsi.org/start.html). Accordingly, the skilled man may generate and test an array or library of different oligonucleotides to determine whether or not they are capable of modulating the expression of the R2R1/2 genes.
[0052] In view of the above, the antisense oligonulcoeotides and/or siRNA molecules described herein may be used (i) treat any of the diseases and/or conditions described herein--in particular (ii) to treat pulmonary diseases and/or disorders, (iii) to treat cardiac diseases and/or conditions and (iv) to treat cancer. Furthermore, the antisense oligonulcoeotides and/or siRNA molecules described herein may be used in the manufacture of medicaments for treating the diseases outlined as (i)-(iv) above or in methods of treating subjects suffereing from such diseases and/or disorders.
[0053] In addition, antibodies (or antigen binding fragments thereof) capable of binding to the R2R1/2 proteins may be useful in the treatment of the diseases and/or conditions described herein, including, for example, cardiac and/or pulmonary diseases and/or conditions. Antibodies which block or neutralise the function of the R2R1/2 proteins may be particularly useful where a disease and/or condition results from over expression of an R2R1/2 protein. The techniques used to generate monoclonal antibodies (mAbs) are well known and can easily be exploited to generate mAbs specific for either of the R2R1 and/or R2R2 proteins or fragments thereof. Similalry, the processes used to generate polyclonal antibodies are also well established and may be used to generate antibodies specific for either of the R2R1 and/or R2R2 proteins or fragments thereof.
[0054] Other compounds useful in the treatment of diseases and/or conditions described herein (for example cardiac and/or pulmonary conditions and/or disorders or cancer) may include for example, proteins, peptides, amino acids, carbohydrates and other small organic molecules.
[0055] In addition to the above, isolated R2R1/2 nucleotide and/or protein sequences may be used as the basis for the design of probes and/or primers for use in ex vivo and/or in situ detection and expression studies. Typical detection studies include, for example, polymerase chain reaction (PCR), hybridisation studies, sequencing protocols and immunological and/or Southern/Northern blotting detection techniques.
[0056] In principle any polynucleotide (or oligonucleotides) or polypeptide fragment designed from the sequences described above may be used in said detection and/or expression studies.
[0057] Typically, polynucleotide fragments for use as probes and/or primers, will comprise 10-30 nucleotides (although other lengths may be useful for certain applications) and exhibit a degree of specificity for a particular sequence and will not bind unrelated sequences. Similarly, polypeptide fragments to be used as probes may also be relatively short and typically may comprise 5-20 amino acids (although other slightly shorter or longer lengths may be useful for some applications).
[0058] It will be readily understood that careful selection of the primer/probe sequence and the use of stringent (preferably highly stringent) hybridisation conditions will minimise any non-selective binding.
[0059] Accordingly, oligonucleotides probe and/or primer sequences having at least 50%, at least 75%, at least 90% or at least 95% complementarity as well as those having exact (i.e. 100%) complementarity to all or part of the nucleotide sequences described herein are to be considered as encompassed within this invention.
[0060] Hybridisation between a probe/primer and a nucleic acid sequence (such as any described herein) may be effected at a temperature of about 40° C.-75° C. in 2-6×SSC (i.e. 2-6×NaCl 17.5 g/l and sodium citrate (SC) at 8.8 g/l) buffered saline containing 0.1% sodium dodecyl sulphate (SDS). Of course, depending on the degree of similarity between the probe/primer and the sequence, buffers with a reduced SSC concentration (i.e. 1×SSC containing 0.1% SDS, 0.5×SSC containing 0.1% SDS and 0.1×SSC containing 0.1% SDS).
[0061] Polypeptide probes having at least 30%, 50%, 70%, 75%, 80%, 85%, 90% or 95% identity as well as those having exact (i.e. 100% identity) to all or part of the amino acid sequences disclosed herein, are to be considered as within the scope of this invention.
[0062] As such, a further aspect of this invention provides oligonucleotides probes and/or primers designed to hybridise to all or part of a sequence selected from the group consisting of SEQ ID NOS: 1; 2; 4; 5; 7; 8; 10 and 11. In addition, a further aspect provides polypeptide probes designed to bind to all or part of a sequence selected from the group consisting of SEQ ID NOS: 3; 6; 9 and 12
[0063] In another aspect the present invention provides a method of diagnosing a pulmonary disease or condition and/or susceptibility thereto, wherein the method comprises determining if the R2R1 and/or R2R2 genes in a subject are aberrantly expressed.
[0064] Subjects diagnosed as suffering from, for example cancer and/or a cardiac and/or pulmonary disease and/or condition may exhibit aberrant (i.e. increased or decreased) R2R1 and/or R2R2 gene/protein expression. The term "aberrant expression" should be understood to encompass levels of gene expression that are either increased and/or decreased relative to the expression observed in sample derived from a healthy subject or a subject who is not suffering from the diseases and/or condition (namely a cardiac or pulmonary disease and/or condition or cancer).
[0065] The term "sample" should be understood as including samples of bodily fluids such as whole blood, plasma, serum, saliva, sweat and/or semen. In other instances "samples" such as tissue biopsies and/or scrapings may be used. In particular lung tissue biopsies and/or scrapings may be used. In addition, a sample may comprise a tissue or gland secretion and washing protocols may be used to obtain sample of fluid secreted into, for example, the lung. Suitable washing protocols may include broncho-alveolar lavage procedures. One of the skill in this field will appreciate that the samples described above may yield quantities of R2R1/2 nucleic acid (i.e. DNA or RNA) and/or R2R1/2 proteins, peptides (or fragments thereof). Furthermore, these methods may comprise the first step of providing a sample from a subject suspected of suffering from a pulmonary disease and/or condition or who may be at risk of developing a pulmonary disease and/or condition.
[0066] An increase in the level of R2R1 and/or R2R2 gene/protein expression may be associated with any of the diseases and/or conditions described above or susceptibility thereto. For example an increase in R2R1/2 gene/protein expression may be indicative of excessive cellular proliferation and/or differentiation--and may be associated with, for example, a neoplastic condition such as cancer (i.e, lung cancer). Decreases in R2R1 and/or R2R2 gene/protein expression may be indicative of pathological conditions characterised by poor or impaired cardiac/lung development. Such conditions may result in tissue injury (due to mechanical stresses acting within the heart/lung), scarring, loss of structural integrity of the heart/lung and deformed or damaged lung airways or cardiac structure.
[0067] One of skill in the art will be familiar with the techniques that may be used to identify levels of proteins and/or genes, such as the R2R1/2 genes and/or proteins, in samples such as those listed above.
[0068] Such techniques may include, for example, polymerase chain reaction (PCR) based techniques such as real-time PCR (otherwise known as quantitative PCR). In the present case, real time-PCR may used to determine the level of expression of the genes encoding the R2R1 and/or R2R2 proteins. Typically, and in order to quantify the level of expression of a particular nucleic acid sequence, reverse transcriptase PCR may be used to reverse transcribe the relevant mRNA to complementary DNA (cDNA). Preferably, the reverse transcriptase protocol may use primers designed to specifically amplify an mRNA sequence of interest. Thereafter, PCR may be used to amplify the cDNA generated by reverse transcription.
[0069] Typically, the cDNA is amplified using primers designed to specifically hybridise with a certain sequence and the nucleotides used for PCR may be labelled with fluorescent or radiolabelled compounds.
[0070] One of skill in the art will be familiar with the technique of using labelled nucleotides to allow quantification of the amount of DNA produced during a PCR. Briefly, and by way of example, the amount of labelled amplified nucleic acid may be determined by monitoring the amount of incorporated labelled nucleotide during the cycling of the PCR.
[0071] Further information regarding the PCR based techniques described herein may be found in, for example, PCR Primer: A Laboratory Manual, Second Edition Edited by Carl W. Dieffenbach & Gabriela S. Dveksler: Cold Spring Harbour Laboratory Press and Molecular Cloning: A Laboratory Manual by Joseph Sambrook & David Russell: Cold Spring Harbour Laboratory Press.
[0072] Other techniques that may be used to determine the level of R2R1 and/or R2R2 gene expression in a sample include, for example, Northern and/or Southern Blot techniques. A Northern blot may be used to determine the amount of a particular mRNA present in a sample and as such, could be used to determine the amount of R2R1 and/or R2R2 gene expression. Briefly, mRNA may be extracted from, for example, a cell based or cell free system modified to include expressible R2R1 and/or R2R2 genes using techniques known to the skilled artisan, and subjected to electrophoresis. A nucleic acid probe, designed to hybridise (i.e. complementary to) an mRNA sequence of interest--in this case the mRNA encoding the R2R1 and/or R2R2 proteins, may then be used to detect and quantify the amount of a particular mRNA present in a sample.
[0073] Additionally, or alternatively, a level of R2R1 and/or R2R2 gene expression may be identified by way of microarray analysis. Such a method would involve the use of a DNA microarray that comprises nucleic acid derived from the R2R1 and/or R2R2 genes. To identify the level of R2R1 and/or R2R2 gene expression, one of skill in the art may extract the nucleic acid, preferably the mRNA from a system (either cell based or cell free) subjected to the method described in the first aspect of this invention, and subject it to an amplification protocol such as, reverse transcriptase PCR to generate cDNA. Preferably, primers specific for a certain mRNA sequence--in this case the sequences encoding the R2R1 and/or R2R2 genes may be used.
[0074] The amplified R2R1 and/or R2R2 cDNA may be subjected to a further amplification step, optionally in the presence of labelled nucleotides (as described above). Thereafter, the optionally labelled amplified cDNA may be contacted with the microarray under conditions that permit binding with the DNA of the microarray. In this way, it may be possible to identify a level of R2R1 and/or R2R2 gene expression.
[0075] Further information regarding the above described techniques may be found in, for example, PCR Primer: A Laboratory Manual, Second Edition Edited by Carl W. Dieffenbach & Gabriela S. Dveksler: Cold Spring Harbour Laboratory Press and Molecular Cloning. A Laboratory Manual by Joseph Sambrook & David Russell: Cold Spring Harbour Laboratory Press.
[0076] In order to determine the level of R2R1/2 proteins in a sample, immunological techniques exploiting agents capable of binding R2R1/2 proteins, may be used.
[0077] In one embodiment, the diagnostic methods described above may comprise the step of contacting a substrate (or portion thereof) with a sample to be tested, under conditions which permit the association, interaction, binding and/or immobilization of any R2R1/2 protein present in the sample, to said substrate.
[0078] Suitable substrates may include, for example, glass, nitrocellulose, paper, agarose and/or plastics. A substrate such as, for example, a plastic material, may take the form of a microtitre plate.
[0079] Alternatively, the substrate to be contacted with the sample to be tested may comprise an agent capable of binding R2R1/2 protein(s). Preferably, the agent capable of binding the R2R1/2 proteins is/are bound to the substrate (or at least a portion thereof). Suitable binding agents may include, for example, antibodies such as monoclonal or polyclonal antibodies and/or other types of peptide or small molecule capable of binding to the R2R1/2 proteins. It is to be understood that this definition applies to all types of binding agent mentioned herein. As such, the substrate (or a portion thereof) may be contacted with the sample to be tested under conditions that permit binding or interaction between the agents capable of binding the R2R1/2 protein and any R2R1/2 protein present in the sample.
[0080] Any R2R1/2 protein bound to the substrate or agents capable of binding the R2R1/2 protein(s), may be detected with the use of a further agent capable of binding the R2R1/2 protein(s) (referred to hereinafter as the "primary binding agent"). Additionally, or alternatively, the primary binding agents may have affinity for, or bind to, R2R1/R2R2 protein: substrate complexes or complexes comprising the R2R1/2 proteins and the abovementioned agents capable of binding the R2R1/2 proteins.
[0081] The primary binding agents may be conjugated to moieties which permit them to be detected (referred to hereinafter as "detectable moieties"). For example, the primary agents may be conjugated to an enzyme capable of reporting a level via a colorimetric chemiluminescent reaction. Such conjugated enzymes may include but are not limited to Horse Radish Peroxidase (HRP) and Alkaline Phosphatase (AlkP). Additionally, or alternatively, the primary binding agents may be conjugated to a fluorescent molecule such as, for example a fluorophore, such as FITC, rhodamine or Texas Red. Other types of molecule that may be conjugated to binding agents include radiolabelled moieties.
[0082] Alternatively, any R2R1/2 protein bound to the substrate or agents capable of binding the R2R1/2 proteins, may be detected by means of a yet further binding agent (referred to hereinafter as "secondary binding agents") having affinity for the primary binding agents. Preferably, the secondary binding agents are conjugated to detectable moieties.
[0083] The amount of primary binding agent (or secondary binding agent bound thereto) bound to R2R1/2 protein(s), may represent the level of R2R1/2 protein(s) present in the sample tested.
[0084] In one embodiment, the methods for identifying a level of R2R1/2 protein, may take the form of "dip-stick" test, wherein a substrate (or portion thereof) is contacted with a sample to be tested under conditions which permit the binding of any R2R1/2 protein(s) present in the sample to the substrate or a binding agent bound or immobilised thereto.
[0085] In a further embodiment, the methods may take the form of an immunological assay such as, for example, an enzyme-linked immunosorbent assay (ELISA). An ELISA may take the form of a "capture" ELISA wherein, a sample to be tested is contacted with a substrate, and any R2R1/2 protein(s) present in the sample is/are "captured" or bound by a binding agent (capable of binding the R2R1/2 proteins) bound or immobilized to the substrate. Alternatively, the sample may be contacted with the substrate under conditions that permit "direct" binding between any R2R1/2 protein(s) present in the sample and the substrate.
[0086] Each of the ELISA methods described above may comprise a "direct" protein detection step or an "indirect" identification step. ELISAs involving such steps may be known as "direct" ELISAs or "indirect" ELISAs.
[0087] A "direct" ELISA may involve contacting the sample to be tested with a substrate under conditions that permit the binding of any R2R1/2 protein(s) present in the sample to the substrate and/or a binding agent bound thereto. After an optional blocking step, bound R2R1/2 protein(s) may be detected by way of an agent capable of binding the R2R1/2 proteins (i.e. a primary binding agent). Preferably, the primary binding agents are conjugated to a detectable moiety.
[0088] An "indirect" ELISA may comprise the further step of, after contacting the R2R1/2 protein(s) with a primary binding agent, using a further binding agent (secondary binding agent) with affinity or specificity for the primary binding agent. Preferably, the secondary binding agent may be conjugated to a detectable moiety.
[0089] Other immunological techniques which may be used to identify a level of R2R1/2 protein in a sample include, for example, immunohistochemistry wherein binding agents, such as antibodies capable of binding the R2R1/2 protein(s), are contacted with a sample, preferably a tissue sample, under conditions which permit binding between any R2R1/2 protein(s) present in the sample and the R2R1/2 protein binding agent. Typically, prior to contacting the sample with the binding agent, the sample is treated with, for example a detergent such as Triton X100. Such a technique may be referred to as "direct" immunohistochemical staining.
[0090] Alternatively, the sample to be tested may be subjected to an indirect immunohistochemical staining protocol wherein, after the sample has been contacted with a R2R1/2 protein binding agent, a further binding agent (a secondary binding agent) which is specific for, has affinity for, or is capable of binding the R2R1/2 protein binding agent, is used to detect R2R1/2 protein/binding agent complexes.
[0091] The skilled man will understand that in both direct and indirect immunohistochemical techniques, the binding agent or secondary binding agent may be conjugated to a detectable moiety. Preferably, the binding agent or secondary binding agent is conjugated to a moiety capable of reporting a level of bound binding agent or secondary binding agent, via a colorimetric chemiluminescent reaction.
[0092] In order to identify the levels of R2R1/2 protein(s) present in the sample, one may compare the results of an immunohistochemical stain with the results of an immunohistochemical stain conducted on a reference sample. By way of example, a sample that reveals more or less bound R2R1/2 protein binding agent (or secondary binding agent) than in a reference sample, may have been provided by a subject with a particular disease and/or condition.
[0093] Other techniques that exploit the use of agents capable of binding the R2R1/2 proteins include, for example, techniques such as Western blot or dot blot. A Western blot may involve subjecting a sample to electrophoresis so as to separate or resolve the components, for example the proteinaceous components, of the sample. The resolved components may then be transferred to a substrate, such as nitrocellulose. In order to identify any R2R1/2 protein(s) present in the sample, the substrate may be contacted with a binding agent capable of binding R2R1/2 protein(s) under conditions which permit binding between any R2R1/2 protein(s) present in the sample and the agents capable of binding R2R1/2 protein(s).
[0094] Advantageously, the agents capable of binding R2R1/2 protein(s) may be conjugated to a detectable moiety.
[0095] Alternatively, the substrate may be contacted with a further binding agent having affinity for the binding agent(s) capable of binding R2R1/2 protein(s). Advantageously, the further binding agent may be conjugated to a detectable moiety.
[0096] In the case of a dot blot, the sample or a portion thereof, may be contacted with a substrate such that any R2R1/2 protein(s) present in the sample is bound to or immobilised on the substrate. Identification of any bound or immobilised R2R1/2 protein(s) may be conducted as described above.
[0097] In any of the abovementioned techniques, the amount of primary or secondary binding agent detected is representative of, or proportional to, the amount of R2R1/2 protein present in the sample. Furthermore, the results obtained from any or all of the diagnostic methods described herein may be compared with the results obtained from reference or control samples derived from healthy subjects known not to be suffering from, or susceptible to, a particular disease or disorder (such as, for example a cardiac and/or pulmonary disease or disorder and/or cancer).
[0098] A further aspect of this invention provides a method of identifying or obtaining agents which modulate the expression of the R2R1 and/or R2R2 genes, said method comprising the steps of contacting the R2R1 and/or R2R2 genes with a test agent and detecting any modulation of R2R1/2 gene expression.
[0099] One of skill in this field will appreciate that a method such as that described in this 9th aspect of the invention may be conducted in systems such as, for example, cell based or cell free systems, modified to include the R2R1 and/or R2R2 genes. By way of example, cells may be transfected with nucleic acid comprising either the R2R1 or R2R2 gene. In one embodiment, the nucleic acid may take the form of a vector (for example a plasmid or expression cassette as described above).
[0100] In one embodiment, the results obtained from the methods described above may be compared to those obtained from a control method in which the R2R1 and/or R2R2 genes have not been contacted with a test agent. In this way, it may be possible to determine whether or not said agent is capable of modulating the expression of the R2R1 and/or R2R2 genes. Where the level of R2R1 and/or R2R2 gene expression is less or greater than the level of expression detected in the control method, the test agent may be useful as a modulator of R2R1 and/or R2R2 gene expression. Where the level of expression is the same as that observed in the control methods, the test agent is most likely not capable of modulating the expression of the R2R1 and/or R2R2 genes.
[0101] Suitable test agents may take the form of nucleic acids, for example the antisense oligonucleotides described above, proteins, peptides, amino acids, antibodies (and fragments thereof), carbohydrates and other small organic molecules.
[0102] In further aspect, the present invention provides pharmaceutical compositions comprising the R2R1/2 genes/proteins described above, antisense oligonucleotides (DNA or RNA) as described herein and/or any of the agents identified by the methods provided by the 9th aspect of this invention and which are capable of modulating the expression or function of the R2R1 and/or R2R2 genes/proteins, in association with a pharmaceutically acceptable excipient, carrier or diluent. Such compositions may find application in, for example, the treatment of the various diseases and/or conditions described herein including the cardiac and/or pulmonary diseases and/or cancers described above.
[0103] Preferably, the pharmaceutical compositions provided by this invention are formulated as sterile pharmaceutical compositions. Suitable excipients, carriers or diluents may include, for example, water, saline, phosphate buffered saline, dextrose, glycerol, ethanol, ion exchangers, alumina, aluminium stearate, lecithin, serum proteins, such as serum albumin, buffer substances such as phosphates, glycine, sorbic acid, potassium sorbate, partial glyceride mixtures of saturated vegetable fatty acids, water salts or electrolytes, such as protamine sulphate, disodium hydrogen phosphate, potassium hydrogen phosphate, sodium chloride, zinc salts, colloidal silica, magnesium trisilicate, polyvinyl pyrrolidone, cellulose-based substances, polyethylene glycon, sodium carboxymethylcellulose, polyacrylates, waxes, polyethylene-polypropylene-block polymers, polyethylene glycol and wool fat and the like, or combinations thereof.
[0104] Said pharmaceutical formulation may be formulated, for example, in a form suitable for oral, parenteral or topical administration. In one embodiment, the pharmaceutical composition may be formulated such that it can be inhaled. Compositions that are to be administered by inhalation may take the form of fine powders or solutions which can be aerosolised and inhaled as droplets. One of skill in this field will be familiar with devices that may be used to deliver compositions directly to the lung by, for example, inhalation. The droplet or particle size of the composition can be altered such that the drug can access different regions of the lung. For example, once inhaled, small particles or droplets may penetrate deep into the lung tissue and in some cases may reach the alveoli.
[0105] Pharmaceutical compositions formulated for topical administration may be presented as an ointment, solution or a suspension in an aqueous or non-aqueous liquid, or as an oil-in-water liquid emulsion.
[0106] Compounds capable of modulating the R2R1 and/or R2R2 genes, such as, for example, those identified by the methods described herein or the R2R1/2 gene/protein fragments, antisense oligonucleotides, and antibodies described herein, may find further application as modulators of cell differentiation. As stated, the inventors have determined that the R2R1 and R2R2 genes and their products are involved in the pathways that modulate the differentiation of pulmonary epithelial cells, particularly the squamous epithelial cells generated from the pulmonary epithelial progenitor or basal cell population (i.e. Krt14+ve cells).
[0107] Compounds that modulate cell (for example, pulmonary epithelial cell) differentiation may be particularly useful in the treatment of disorders such as BPD which yield damaged and scarred lungs with a reduced or underdeveloped regenerative potential. As such, by administering or using a compound which modulates cell differentiation, it may be possible to improve or restore the regenerative potential of the lung. One of skill in this field will readily understand that compounds capable of enhancing or promoting R2R1 and/or R2R2 gene expression or which restore the function of these genes may be particularly useful.
[0108] In a further aspect, the present invention provides an animal model for studying tissue development and/or cell transition events as well as certain diseases and/or conditions (including cardiac and/or pulmonary diseases and/or conditions and cancer). In one embodiment, the animal model may be generated by manipulating or modulating (i.e. up or down regulating) the expression of the R2R1/2 genes/proteins. Additionally or alternatively, an animal model may be generated by disrupting R2R1/2 gene expression. As described above, the disclosure of a number of human and murine R2R1/2 gene/protein sequences herein ensure that the skilled man could readily manipulate/modulate these sequences in situ to generate animal models. For example, "knock-out" animals may be created, that is, the expression of R2R1/2 gene sequences is reduced or substantially eliminated. Such models are useful for testing the effects of drugs potentially useful in the treatment of the diseases and/or disorders described herein and/or for determining the function or role of a particular gene. Alternatively, animal models in which the expression of the R2R1/2 genes/proteins are upregulated, may also be created. In this way, the efficacy and function of drugs aimed at suppressing upregulated R2R1/2 gene/protein expression may be tested. It is also possible to investigate the effect of upregulated R2R1/2 gene protein expression in these animals.
[0109] In other embodiments, substitutions, additions, deletions and/or inversions may be introduced into the R2R1/2 gene sequences in order to effect changes in the activity of the proteins and to help elucidate the function of certain domains, amino acids and the like. Additionally, or alternatively, knockout animals, such as those described above, may be transformed with any of the gene sequences described herein. This is particularly useful where the effect of a variant or mutated R2R1/2 gene or efficacy of a drug or compound capable of suppressing the expression or function of said variant or mutated sequence, is to be investigated.
BRIEF DESCRIPTION OF THE DRAWINGS
[0110] The present invention will now be described in detail with reference to the following Figures which show:
[0111] FIG. 1. Comparison of endothelial and epithelial cells from Vegf+/+ at E16.5. Volcanoplot showing on the y-axis the significance of each gene when testing for the effect of cellular origin (based on LIMMA). The x-axis shows the fold change when comparing the 2 tissues. Ker+ cells express archetypical epithelial genes, while il+ cells transcribe representative endothelial genes. Endothelial Cldn5 and epithelial Foxa1 are highlighted because both were also present as top genes in the Y-axis (PC2) of the unsupervised spectral map analysis (see FIG. 3).
[0112] FIG. 2. Schematic representation of the laser capture microdissection proces. Embryonic thoraces (A) were cut in 8 mm sections and placed on slides (B). Double immunohistochemical staining on these tissue sections (C) with anti-`pan` keratin and GS-IB4 isolectin separated pulmonary airway cells with epithelial (D) distinctiveness from the surrounding cells with endothelial (E) characteristics. Specific cell groups isolated by laser capture microdissection are shown in F and G. (L=lung, H=heart).
[0113] FIG. 3. Spectral map analysis with first principal component (PC1 on X-axis) and second principal component (PC2 on Y-axis). Time effect or embryonic age was uncovered in PC1 (accountable for 35% of the variation in the data set). Gene probes of genes expressed later in embryonic life as Sftpc were located further on the X-axis. Difference in cellular origin (il+ versus ker+ cells) was found in PC2. Gene probes of epithelial genes (`EPI`) were located further on the Y-axis in contrast to gene probes of endothelial gene (`ENDO`) families. The different sample groups were separated in the spectral map along PC1 (embryonic age) and PC2 (cellular origin). Ker+ samples gathered on the epithelial side of the Y-axis, and il+ samples grouped on the endothelial side. The samples also distributed along the X-axis according to their embryonic age. Samples of later embryonic days were located further on the X-axis. The panel below shows 3 expression profiles: (1) showing the difference between epithelial and endothelial cells for Foxal expression, (2) showing the effect of embryonic age for Afp expression and (3) showing the genotype dependent profile over embryonic age for Him expression.
[0114] FIG. 4, Comparison of epithelial cells from Vegf+/+ and Vegf120/120 genotypes. A. Volcanoplot showing on the y-axis the significance of each gene for testing whether the expression profile over embryonic age differs between the Vegf+/+ and Vegf120/120 genotypes (based on LIMMA). The x-axis shows the fold change in induction when comparing the 2 genotypes, Vegf+/+ versus Vegf120/120 at E16.5. Upregulation of Krt6a, EDC and SCC genes in wild type ker+ cells is highlighted. B. Airway epithelial cells in E16.5 Vegf+/+ lungs defined by anti-cytokeratin 4, 5, 6, 8, 10, 13, and 18 staining. Distal airway cells develop a flat or squamous cell morphology (yellow arrow) in contrast to their proximal neighbours (white arrow) C. Allocation of differential expression allong chromosome 3. This highlights the downregulation of the genes in the EDC of Vegf120/120 as compared to wildtype.
[0115] FIG. 5A. IF=Intermediate Filament Keratins (red) group anchoring to the Desmosome (orange) containing Dsc1. Pkp1 (purple) links Intermediate Keratin Filaments to Cadherin proteins of the Adherens type junction (yellow). Pkp1 also regulates the protein content of the Desmosome. EDC and SCC cluster genes interact with Intermediate Filament Keratins. The star symbol highlights the upregulated genes and gene clusters in the Intermediate Filament network. Upregulation of Eps811 (coding for an actin-capping protein) coordinates Intermediate Filament with Actin remodelling.
[0116] B. Volcanoplot showing on the y-axis the significance of each gene for testing whether the expression profile over embryonic age differs between endothelial cells of Vegf+/+ and Vegf120/120 genotypes (based on LIMMA). The x-axis shows the fold change in induction when comparing the 2 genotypes, Vegf+/+ versus Vegf120/120 at E16.5. The Krt, Dsc1, Pkp1, EDC and SCC cluster genes are highlighted. The Krt genes upregulated in wild type Vegf+/+ il+ cells differ from the Krt genes expressed in the equivalent wild type ker+ cells. Krt14 and Krt1 are trademark basal cell genes.
[0117] C. Immunofluorescent image (40× magnification) of GS-IB4 isolectin staining cells (il+ cells) in wild type E16.5 embryonic lungs. Il+ cells follow the same architectural pattern of the ker+ cells of the distal airways.
[0118] FIG. 6. Volcanoplot based on LIMMA analysis of the group of anti-cytokeratin 4-5-6-8-10-13-18 staining cells. (Linear Models for Microarray Data: differences between Vegf120/120 knockout and wild type littermates in expression profiles over embryonic age were tested through a two-way interaction of Vegf genotype and time). The RIKEN cDNA 2200001I15 gene and RIKEN cDNA 2310002J15 gene are highlighted.
[0119] FIG. 7. Volcanoplot based on LIMMA analysis of the group of GS-IB4 binding cells. (Linear Models for Microarray Data: differences between Vegf120/120 knockout and wild type littermates in expression profiles over embryonic age were tested through a two-way interaction of Vegf genotype and time). The RIKEN cDNA 2200001I15 gene and RIKEN cDNA 2310002J15 gene are highlighted.
[0120] FIG. 8: cDNA sequence alignments of the human hR4RA transcript (=human R2R1) versus mouse R4Ra transcript (=mouse R2R1). It should be noted that the largest contig constructed from sequenced RIKEN cDNA 2200001I15 gene clones is referred to as `mouse R4Ra transcript (=mouse R2R1)`.
[0121] FIG. 9: Protein sequence alignments of the translated human hR4RA (=human R2R1) transcript versus translated mouse R4Ra (=mouse R2R1) transcript.
[0122] FIG. 10: cDNA sequence alignments of the human hR4RD transcript (=human R2R2) versus mouse R4Rd transcript (=mouse R2R2). It should be noted that the largest contig constructed from sequenced RIKEN cDNA 2310002J15 gene clones is referred to as `mouse R4Rd transcript (=mouse R2R2)`.
[0123] FIG. 11: Protein sequence alignments of the translated human hR4RD (=human R2R2) transcript versus translated mouse R4Rd (=mouse R2R2) transcript.
[0124] FIG. 12: VEGF-A (VEGF164 dependent upregulation of R2R1 in the developing ventricular septum of the mouse embryo. Red=wild type, blue=vEGF120/120 knockout mouse, lacking the VEGF164 isoform.
[0125] FIG. 13: Human R2R2 homologue (=C9orf196) expression over time (24-72 h) in human adult primary lung epithelial cells.
[0126] FIG. 14: siRNA knockdown of VEGF165 leads to knockdown of the genes responsible for the basal cell program and squamous differentiation program. siRNA knockdown in human Primary Bronchial Epithelial Cells (PBEC's) reliably models the effect of gene knockdown of genes of interest. (a) RTqPCR: siRNA mediated knockdown of VEGFA and VEGF165 expression leads to knockdown of KRT14 (basal cell marker) expression. KRT14 relative expression (PGK1 normalized), VEGFA and VEGF165 are plotted versus siRNA. Included are two negative controls (called nr 10 and 11). siRNA directed against KRT14 expression is included as a positive control. siRNA VEGFA is directed against all VEGFA isoforms.
[0127] FIG. 15 A-J: Global gene expression changes brought about by siRNA mediated knockdown of VEGFA in PBEC's. Changes in global gene expression match exactly the changes observed in the Vegf120/120 knockout mouse, Graphs (a)-(j) depict the significance of changes in gene expression after administration during 24 h of an siRNA directed against VEGF. Graphs (a)-(j) are created by automated pathway analysis. All pathways are involved in keratinocyte differentiation: basal cell regeneration and squamous differentiation.
[0128] FIG. 16 A-C: siRNA's against the human R2R1 and R2R2 homologues. The human R2R1 homologue(s) comprise the FAM25 family encompasing the seven human paralogues designated FAM25A, FAM25B, FAM25C, FAM25D, FAM25E, FAM25G and FAM25HP. These are the human homologues of mouse the murine sequence encoded by the cDNA sequences designated 2200001I15Rik or RIKEN cDNA 2200001I15. The similarity of the FAM25 paralogues precludes specific RTqPCR of the different paralogues. The FAM25 paralogues are also absent on Affymetrix Human Gene expression arrays such as HT HG-U133 or HT HG-U219. We have selected the RTqPCR primer-probe set Hs04194072_m1 (Applied Biosystems) for the measurement of differential gene expression of the FAM25 family. Applied Biosystems states that this primer-probe set doesn't differentiate between the different paralogues. Three siRNA's designed against the FAM25 family were selected on their ability to downregulate Hs04194072_m1 expression in PBEC's. We have numbered the siRNA's nr 18, 20 and 22 respectively. Graph (a) illustrates the downregulation of FAM25 (Hs04194072_m1) expression (PGK1 normalized) after 24 h administration of the respective siRNA's in PBEC's. The human R2R2 homologue: Two siRNA's (numbered 15 and 17) designed against C9orf169 (at concentrations of 5 and 20 nM respectively) were selected on the basis of their ability to downregulate C9orf169 expression in PBEC's. C9orf169 expression was evaluated by microarray analysis and RTqPCR. Probes for C9orf169 are present on the Affymetrix HT HG-U219 Human Gene Expression Array. Graph (b) illustrates the downregulation (assessed by microarray analysis) of C9orf169 expression after 24 h administration of the respective siRNA's in PBEC's. The administration of siRNA's directed against the FAM25 family or VEGFA has no effect on C9orf169 expression. siRNA's directed against R2R1 (human FAM25 family) and R2R2 (human C9orf169) downregulate KRT14 expression and is the response similar to the one observed after administration of siRNA's directed against VEGFA and VEGF165 siRNA mediated knockdown of the FAM25 family and C9orf169 leads to downregulation of KRT14 gene expression (basal cell marker). This response is most obvious after administration of siRNA's directed against C9orf169. Graph (c) illustrates the downregulation KRT14 expression (PGK1 normalized) after 24 h administration of the respective siRNA's in PBEC's. Included are also the siRNA's directed against VEGFA and KRT14 for reason of comparison.
[0129] FIG. 17: is a schematic representation of the effects of the R2R homologues. Expression R2R homologues leads to simultaneous modulation of HIF1A signaling (conferring oxygen tolerance) (Box 5), AND modulation of a specific (PERP) anti-apoptotic pathway (Box 4). This will permit the (re)generation of strong epithelial cells (with a major defense barrier against stress), without the installment of unlimited growth potential. In other words, the modulation of a specific anti-apoptotic pathway is not a `permit` for general tolerance to apoptosis. General tolerance to apoptosis would lead to the dangerous situation of immortalization of the cell: the cell develops into a cancer cell. The pathway was constructed on the basis of microarray analysis of siRNA mediated knockdown of the human R2R1 homologue (FAM25 family) and human R2R2 homologue (C9orf169).
[0130] FIG. 18: The R2R homologues are essential for `Keratinocyte regeneration and differentiation` (Box 1) and `Apoptosis modulation` (Box 2) in the VEGFA-VEGF165 pathway. The effects on gene expression by the R2R homologues on `Keratinocyte regeneration and differentiation`(Box 1) are apparent in FIGS. 18A and 18B. The effects on gene expression by the R2R homologues on `Apoptosis modulation`(Box 2) are apparent in FIG. 18C. They are the final end-effects of Box 3, 4 and 5.
[0131] FIG. 19 A-B: The effect on cellular respiration (see Box 3 in FIG. 17) is highly specific: siRNA mediated knockdown of the R2R homologues will down-regulate oxidative phosphorylation. Expression of the R2R homologues will upregulate oxidative phosphorylation. We can observe this effect at it's most profound in the expression of mitochondrial ATP5A1-ATP synthase (H+ transporting) (FIG. 19A). siRNA's directed at the R2R homologues will downregulate ATP5A1 expression. FIG. 19 B illustrates the expression of ATP5A1 among global changes in gene expression of the oxidative phosphorylation pathway.
[0132] FIG. 20: The effects of the R2R homologues on P53-P63 signaling (see Box 4 in FIG. 17). PERP (TP53 apoptosis effector) is downregulated by siRNA mediated knockdown of the human R2R homologues. `PERP is a p63-Regulated Gene Essential for Epithelial Integrity. p63 is a master regulator of stratified epithelial development that is both necessary and sufficient for specifying this multifaceted program. Perp, a tetraspan membrane protein originally identified as an apoptosis-associated target of the p53 tumor suppressor, is the first direct target of p63 clearly involved in mediating this developmental program in vivo`: this was demonstrated by Rebecca A. Ihrie et al. in 2005 (Cell, Vol. 120, 843-856, Mar. 25, 2005) (italics quoting author in the abstract of the paper). The graph demonstrates the downregulation of PERP expression after siRNA mediated knockdown of the human R2R homologues.
[0133] FIG. 21: An overview of the expression of PERP among global changes in gene expression of the P53 pathway after administration of FAM25 nr 20 (directed at human homologue R2R1)
[0134] FIG. 22: An overview of the expression of PERP among global changes in gene expression of the P53 pathway after administration of FAM25 nr 18 (directed at human homologue R2R1)
[0135] FIG. 23: An overview of the expression of PERP among global changes in gene expression of the P53 pathway after administration of C9orf169 nr 15 (directed at human homologue R2R2)
[0136] FIG. 24: An overview of the expression of PERP among global changes in gene expression of the P53 pathway after administration of C9orf169 nr 17 (directed at human homologue R2R2)
[0137] FIG. 25 A-C: The expression of HIF1A (SEE Box 5 IN FIG. 17) is tightly regulated by the expression of the human R2R homologues. siRNA mediated knockdown of the human R2R homologues will downregulate HIF1A expression (FIG. 25A). The downstream HIF1A effects on cellular respiration (oxidative phosphorylation) were described in 3 (see FIG. 17 Cellular respiration). The R2R homologues drive `PERP type p53-p63` and HIF1A expression. As such `tough` epithelial cells, equipped with the HIF1A armory, could develop unlimited growth potential under the influence of VEGFA/VEGF165. However, we observe at the same time that the epithelial cells are precluded from entering an immortal state. Expression of the R2R homologues will act as a brake for the expression of genes that could immortalize the cell. This is very obvious in the case of BCL2A1 (FIG. 25B), an anti-apoptosis gene whose expression confers resistance to therapy in cancer cells. siRNA mediated knockdown of the R2R homologues leads to upregulation of BCL2A1 expression. The R2R homologues act as a brake for BCL2A1 expression. R2R homologues will also promote the expression of genes that permit cells to enter apoptosis if needed. This is demonstrated by siRNA mediated knockdown of the R2R homologues: the knockdown will cause a downregulation MAP2K4 (FIG. 25c) a tumor suppressor in lung adenocarcinoma.
DETAILED DESCRIPTION
[0138] Table 1. RIKEN cDNA 2200001I15 gene and RIKEN cDNA 2310002J15 gene among the top list of genes in SAM analysis of differential gene expression on E16.5 in wild type and Vegf120/120 knockout anti-cytokeratin 4-5-6-8-10-13-18 staining cells.
[0139] Table 2. RIKEN cDNA 2200001I15 gene and RIKEN cDNA 2310002J15 gene among the top list of genes in SAM analysis of differential gene expression on E16.5 in wild type and Vegf120/120 knockout GS-IB4 staining cells.
[0140] Table 3. Distribution of embryos according to age and Vegf genotype status. wt/wt=homozygous wild type (Vegf+/+), 120/wt=heterozygous Vegf120/+, and 120/120=homozygous Vegf120/120 knockout. (NG=not genotyped because of poor embryologic morphology).
TABLE-US-00009 TABLE 1 Probe Set ID Variance UniGene ID Alignments Gene Title Gene Symbol 1448745_s_at 44.327 Mm.1121 chr3: 92166199-9216906 loricrin Lor 1451613_at 25.931 Mm.208047 chr3: 93405151-9341897 hornerin Hrnr 1449986_at 21.731 Mm.160339 chr16: 88647705-886486 RIKEN cDNA 2310034C09 gene 2310034C09Rik 1440186_s_at 19.595 Mm.44242 chr5: 36523188-3652342 RIKEN cDNA 2310020A21 gene 2310020A21Rik 1421575_at 18.532 Mm.358728 chr16: 88666693-886676 RIKEN cDNA 2310057N15 gene 2310057N15Rik 1453218_at 16.908 Mm.292458 chr3: 92764649-9276632 RIKEN cDNA 1110014K05 gene 1110014K05Rik 1459897_a_at 16.677 Mm.250717 chr7: 30460230-3046489 suprabasin Sbsn 1420676_at 16.402 Mm.41969 chr3: 92731934-9273371 small proline rich-like 3 Sprrl3 1420358_at 13.819 Mm.353193 chr16: 88647705-887093 keratin associated protein 13 Krtap13 1437019_at 13.44 Mm.27156 chr14: 33180971-331844 RIKEN cDNA 2200001I15 gene 2200001I15Rik 1456248_at 12.776 Mm.46390 chr3: 93078529-9307882 RIKEN cDNA 2310002A05 gene /// 2310002A05Rik /// LOC630971 1434227_at 12.373 Mm.268167 chr7: 30496664-3049985 keratinocyte differentiation associa Krtdap 1420677_x_at 12.312 Mm.41969 chr3: 92731934-9273371 small proline rich-like 3 Sprrl3 1439630_x_at 12.212 Mm.250717 chr7: 30463567-3046489 suprabasin Sbsn 1420350_at 11.873 Mm.279773 chr3: 92754015-9275571 small proline rich-like 2 Sprrl2 1428781_at 11.64 Mm.30138 chr7: 30485165-3048982 RIKEN cDNA 1110014F24 gene 1110014F24Rik 1452732_at 11.02 Mm.183043 chr6: 86593812-8659533 RIKEN cDNA 2300003P22 gene 2300003P22Rik 1419394_s_at 10.266 Mm.21567 chr3: 90754997-9075596 S100 calcium binding protein A8 (c S100a8 1453092_at 10.127 Mm.35806 chr3: 93099607-9310107 RIKEN cDNA 2300002G24 gene 2300002G24Rik 1435111_at 9.396 Mm.32861 chr7: 24063581-2406386 RIKEN cDNA 2310011E23 gene 2310011E23Rik 1419709_at 9.22 Mm.136573 chr16: 36369769-363746 stefin A3 Stfa3 1435761_at 9.202 Mm.383370 chr16: 36196367-362046 stefin A3 Stfa3 1422784_at 7.244 Mm.302399 chr16: 101517949-10152 keratin complex 2, basic, gene 6a Krt2-6a 1435760_at 7.165 Mm.300592 chr18: 42299151-422997 cystatin A Csta 1448756_at 6.71 Mm.2128 chr3: 90778558-9078122 S100 calcium binding protein A9 (c S100a9 1447669_s_at 6.627 Mm.215394 chr13: 13619774-136200 guanine nucleotide binding protein Gng4 1425336_x_at 5.61 Mm.422886 chr17: 33606481-336107 histocompatibility 2, K1, K region H2-K1 1437145_s_at 5.455 Mm.46431 chr2: 25060827-2506109 RIKEN cDNA 2310002J15 gene 2310002J15Rik 1420741_x_at 5.247 Mm.291782 chr3: 92862612-9286430 RIKEN cDNA 2310069N01 gene 2310089N01Rik 1442339_at 4.062 Mm.187847 chr16: 36076009-360811 stefin A2 like 1 Stfa2l1 1427492_at 3.963 Mm.34964 chrX: 108755370-108762 premature ovarian failure 1B Pof1b 1418722_at 3.693 Mm.236225 chr9: 110265612-110268 neutrophilic granule protein Ngp 1419409_at 3.69 Mm.291769 chr3: 92741051-9274165 small proline rich-like 5 Sprrl5 1436936_s_at 3.42 Mm.274770 chrX: 99684922-9968596 inactive X specific transcripts Xist 1427262_at 3.408 Mm.274770 chrX: 99663093-9968593 inactive X specific transcripts Xist 1430567_at 3.377 Mm.35369 chr18: 44114683-441478 serine peptidase inhibitor, Kazal ty Spink5 1422667_at 3.336 Mm.38498 chr11: 99947848-999520 keratin complex 1, acidic, gene 15 Krt1-15 1448881_at 3.243 Mm.26730 chr8: 112464257-112468 haptoglobin HP 1423547_at 2.71 Mm.45436 chr10: 116681442-11668 lysozyme Lyzs 1449586_at 1.939 Mm.4494 chr1: 137687809-137735 plakophilin 1 Pkp1 1449133_at 1.932 Mm.331191 chr3: 92569339-9257128 small proline-rich protein 1A Sprr1a 1449106_at 1.901 Mm.200916 chr11: 54746194-547537 glutathione peroxidase 3 Gpx3 1452543_a_at 1.862 Mm.2258 chr19: 9150684-9154982 secretoglobin, family 1A, member Scgb1a1 1450633_at 1.859 Mm.21075 chr13: 3837003-3837919 calmodulin 4 Calm4 1459898_at 1.826 Mm.250717 chr7: 30460230-3046489 suprabasin Sbsn 1429565_s_at 1.815 Mm.292457 chr3: 93103210-9310446 late cornified envelope 5A Lce5a 1429540_at 1.727 Mm.34382 chr7: 25076381-2507848 cornifelin Cnfn 1453801_at 1.597 Mm.180200 chr3: 94427502-9443259 thioesterase superfamily member 5 Them5 1422672_at 1.568 Mm.140151 chr3: 92522208-9252419 small proline-rich protein 1B Sprr1b indicates data missing or illegible when filed
TABLE-US-00010 TABLE 2 Probe Set ID Variance UniGene ID Gene Title Gene Symbol 1449586_at 32.877 Mm.4494 plakophilin 1 Pkp1 1423935_x_at 30.173 Mm.6974 keratin complex 1, acidic, gene 14 Krt1-14 1460347_at 28.662 Mm.6974 keratin complex 1, acidic, gene 14 Krt1-14 1438856_x_at 23.207 Mm.268618 serine (or cysteine) peptidase inhibitor, clade B, member 5 Serpinb5 1422667_at 21.017 Mm.38498 keratin complex 1, acidic, gene 15 Krt1-15 1422481_at 20.962 Mm.183137 keratin complex 2, basic, gene 1 Krt2-1 1424096_at 20.14 Mm.383993 keratin complex 2, basic, gene 5 Krt2-5 1451613_at 20.135 Mm.208047 hornerin Hrnr 1453218_at 18.792 Mm.292458 RIKEN cDNA 1110014K05 gene 1110014K05Rik 1441941_x_at 18.5 Mm.268618 serine (or cysteine) peptidase inhibitor, clade B, member 5 Serpinb5 1456203_at 17.757 -- RIKEN cDNA 1110020A10 gene 1110020A10Rik 1434227_at 16.95 -- keratinocyte differentiation associated protein Krtdap 1429067_at 16.396 -- calpain, small subunit 2 Capns2 1440186_s_at 16.186 Mm.378865 Transcribed locus -- 1419709_at 15.648 Mm.136573 stefin A3 Stfa3 1422939_at 14.807 Mm.337362 serine (or cysteine) peptidase inhibitor, clade B (ovalbumin), member 3B Serpinb3b 1422308_a_at 14.219 Mm.20973 lectin, galactose binding, soluble 7 Lgals7 1437019_at 13.876 Mm.27156 RIKEN cDNA 2200001I15 gene 2200001I15Rik 1450633_at 13.842 Mm.21075 calmodulin 4 Calm4 1421117_at 12.773 Mm.336625 dystonin Dst 1421752_a_at 12.718 Mm.268618 serine (or cysteine) peptidase inhibitor, clade B, member 5 Serpinb5 1418799_a_at 12.316 Mm.1225 procollagen, type XVII, alpha 1 Col17a1 1453801_at 11.671 Mm.180200 thioesterase superfamily member 5 Them5 1422940_x_at 11.333 Mm.337362 serine (or cysteine) peptidase inhibitor, clade B (ovalbumin), member 3B Serpinb3b 1436392_s_at 11.112 Mm.3629 transcription factor AP-2, gamma Tcfap2c 1452166_a_at 10.901 Mm.22662 keratin complex 1, acidic, gene 10 Krt1-10 1449938_at 10.491 Mm.1001 placental protein 11 related Pp11r 1421040_a_at 10.27 Mm.371562 glutathione S-transferase, alpha 2 (Yc2) Gsta2 1437232_at 10.198 Mm.107214 bactericidal/permeability-increasing protein-like 2 Bpil2 1424623_at 9.782 Mm.268618 serine (or cysteine) peptidase inhibitor, clade B, member 5 Serpinb5 1418748_at 9.085 Mm.20940 caspase 14 Casp14 1418158_at 9.029 Mm.20894 transformation related protein 63 Trp63 1430551_s_at 8.917 Mm.195937 lipase-like, ab-hydrolase domain containing 3 LipI3 1430550_at 8.679 Mm.195937 lipase-like, ab-hydrolase domain containing 3 LipI3 1435761_at 8.652 Mm.136573 stefin A3 Stfa3 1448397_at 8.545 Mm.25652 gap junction membrane channel protein beta 6 Gjb6 1459898_at 8.407 Mm.250717 suprabasin MGI: 2446326 1422672_at 8.373 Mm.140151 small proline-rich protein 1B Sprr1b 1459897_a_at 8.371 Mm.250717 suprabasin MGI: 2446326 1428781_at 8.218 Mm.30138 RIKEN cDNA 1110014F24 gene 1110014F24Rik 1419492_s_at 8.128 Mm.5341 defensin beta 1 Defb1 1439183_at 8.102 Mm.218784 N-acylsphingosine amidohydrolase (alkaline ceramidase) 3 Asah3 1419491_at 8.028 Mm.5341 defensin beta 1 Defb1 1427263_at 7.99 -- inactive X specific transcripts Xist 1416930_at 7.471 Mm.878 lymphocyte antigen 6 complex, locus D Ly6d 1435760_at 7.251 Mm.300592 cystatin A /// similar to Stefin homolog MGI: 3524930 /// LOC547252 1439630_x_at 7.031 Mm.250717 suprabasin MGI: 2446326 1435639_at 6.509 -- RIKEN cDNA 2610528A11 gene 2610528A11Rik 1424976_at 6.23 Mm.120274 ras homolog gene family, member V Rhov 1455519_at 6.163 Mm.383274 desmoglein 1 beta Dsg1b 1455715_at 5.932 Mm.373656 PREDICTED: Mus musculus RIKEN cDNA 2700099C18 gene -- (2700099C18Rik), mRNA 1434534_at 5.693 -- -- -- 1426048_s_at 5.603 Mm.85544 transcription factor AP-2, alpha Tcfap2a 1449500_at 5.535 Mm.66015 serine (or cysteine) peptidase inhibitor, clade B, member 7 Serpinb7 1430582_at 5.508 Mm.133101 SNF2 histone linker PHD RING helicase Shprh 1435670_at 5.464 Mm.137021 transcription factor AP-2 beta Tcfap2b 1422588_at 5.427 Mm.358617 keratin complex 2, basic, gene 6b Krt2-6b 1445187_at 5.408 Mm.329504 RIKEN cDNA 9430070O13 gene /// gene model 979, (NCBI) 9430070O13Rik /// Gm979 1421996_at 5.28 Mm.85544 transcription factor AP-2, alpha Tcfap2a 1423323_at 5.278 Mm.154045 tumor-associated calcium signal transducer 2 Tacstd2 1452228_at 5.242 Mm.257819 RIKEN cDNA 4930451A13 gene 4930451A13Rik 1449959_x_at 5.214 Mm.23784 small proline rich-like 9 Sprrl9 1419731_at 5.211 Mm.14098 cytochrome P450, family 2, subfamily b, polypeptide 19 Cyp2b19 1442279_at 5.14 Mm.312133 Enhancer of polycomb homolog 1 (Drosophila) (Epc1), transcript variant 1, Epc1 mRNA 1455408_at 5.008 Mm.24880 RIKEN cDNA 4732472I07 gene 4732472I07Rik 1416271_at 4.925 Mm.28209 PERP, TP53 apoptosis effector Perp 1418722_at 4.888 Mm.236225 neutrophilic granule protein Ngp 1437351_at 4.809 Mm.224814 CXXC finger 4 Cxxc4 1440162_x_at 4.662 Mm.208144 hypothetical protein A630043P06 A630043P06 1426641_at 4.647 Mm.266679 tribbles homolog 2 (Drosophila) Trib2 1442349_at 4.643 Mm.259334 RIKEN cDNA C630028N24 gene C630028N24Rik 1425624_at 4.593 Mm.209005 EPM2A (laforin) interacting protein 1 Epm2aip1 1460038_at 4.577 Mm.297371 POU domain, class 3, transcription factor 1 Pou3f1 1440523_at 4.562 Mm.98096 retinal short chain dehydrogenase reductase 2 MGI: 2668443 1431211_s_at 4.438 Mm.180200 thioesterase superfamily member 5 Them5 1447329_at 4.351 -- -- -- 1443687_x_at 4.344 -- -- -- 1420988_at 4.306 Mm.311585 polymerase (DNA directed), eta (RAD 30 related) Polh 1456248_at 4.189 Mm.46390 RIKEN cDNA 2310002A05 gene 2310002A05Rik 1430000_at 4.157 -- RIKEN cDNA B230117O15 gene B230117O15Rik 1446490_at 4.154 Mm.29966 Polypyrimidine tract binding protein 2, mRNA (cDNA clone MGC: 11671 Ptbp2 IMAGE: 3709255) 1441909_s_at 4.1 Mm.225253 RIKEN cDNA 9530066K23 gene 9530066K23Rik 1427747_a_at 4.035 Mm.9537 lipocalin 2 Lcn2 1437145_s_at 3.994 Mm.46431 RIKEN cDNA 2310002J15 gene 2310002J15Rik 1419463_at 3.985 Mm.20897 chloride channel calcium activated 2 Clca2 1441440_at 3.976 Mm.277366 autophagy-related 4C (yeast) Atg4c 1437705_at 3.854 -- -- -- 1418028_at 3.846 Mm.19987 dopachrome tautomerase Dct 1442786_s_at 3.804 Mm.270469 DNA segment, Chr 5, Brigham & Women's Genetics 0860 expressed D5Bwg0860e
TABLE-US-00011 TABLE 3 E7733 E12.5 A wt/wt B wt/120 C 120/120 D wt/120 E wt/120 F wt/wt G wt/wt H 120/120 E7734 E12.5 A 120/120 B wt/120 C wt/120 D wt/120 E wt/120 F wt/120 G wt/120 H 120/120 I wt/wt K 120/120 E7232 E14.5 A 120/wt B 120/wt C 120/120 D wt/wt E 120/wt F 120/wt G wt/wt H 120/120 I 120/wt K wt/wt L wt/wt E7494 E14.5 A 120/120 B wt/wt C 120/wt D 120/wt E 120/120 F 120/wt G 120/wt H 120/wt I wt/wt E7119 E16.5 A 120/wt B 120/wt C 120/120 D 120/wt E 120/wt F 120/wt G 120/120 H 120/wt I wt/wt J 120/wt K 120/wt L 120/wt E7477 E16.5 A 120/120 B wt/wt C ? D 120/+ E wt/wt F wt/wt G ? H 120/wt I 120/wt K 120/wt L 120/120 E7478 E16.5 A 120/120 B wt/wt C 120/wt D 120/wt E 120/wt F 120/120 G wt/wt H 120/wt I 120/wt K wt/wt L 120/120 M wt/wt
Introduction
[0141] The R2R genes, transcripts and respective proteins were discovered in a VEGF mouse knockout model. The Vegf120/120 knockout mouse is unable to produce the Vegf164 and Vegf188 isoforms (Vegf164 is the homologue of human VEGF165). The lungs of Vegf120/120 knockout mice are hypoplastic at birth and peripheral airway and vascular differentiation becomes severely impaired in Vegf120/120 knockout embryonic lungs. A genomics approach led to the discovery that Vegf164 in the mouse and VEGF165 in man drive a very specific gene expression program. This program consists of two components. The first component is the (re)generation of basal cells of the airway epithelium: basal cells are the source of at least the cellular population of the proximal airways. The basal cells also generate strong intercellullar connections by building hemidesmosome and focal adhesion connections. The basal cells strengthen their intracellular architecture by the intermediate filament proteins KRT14 and KRT5. These two proteins are glued to the (hemi)desmosome.
[0142] The second component is a differentiation program: it's a `fortification program` (`squamous differentiation`) of the cells lining the airways. These cells need to be tough at birth, as they will be exposed to mechanical stress and high levels of oxygen. The fortification is made possible by a family of proteins that strengthen cellular architecture. This family of proteins consists of the intermediate filament group and the proteins that strengthen the intermediate filaments (SPRR proteins, LOR, HRNR, etc). These two programs can be summarized under the headings `keratinocyte differentiation`, `epidermal cell differentiation`, `intermediate filament reorganization`, `cornified envelope`, `keratinocyte differentiation`. `cytoskeleton remodeling keratin filaments`.
[0143] While it is tempting to use the VEGF165 protein for the regeneration of damaged lungs in humans, VEGFA and VEGF165 are important regulators of a vast diversity of processes in the organism. Hence, the administration of VEGFA or VEGF165 would result in too many side effects.
[0144] Two novel genes (R2R1 and R2R2) were discovered in the gene expression program that's driven by Vegf164 in the mouse and VEGF165 in man. These novel genes, their transcripts and translated proteins are not related to the genes and their downstream products of the basal and squamous gene expression program.
[0145] Experiments were undertaken to demonstrate that these genes are important modulators of the basal and squamous differentiation program. One of the most important findings of these experiments is that these genes are important modulators of HIF1α and PERP expression in the cell. In our experiments, the R2R genes are positive regulators and interference with this mechanism opens therapeutic possibilities in cancer therapy.
Material and Methods
[0146] Mouse Embryos and Tissue Processing.
[0147] All animal experiments were approved by the Animal Ethics Committee of Leiden University Medical Center and performed according to the Guide for the Care and Use of Laboratory Animals published by the NIH. Heterozygous Vegf+/120 mice were crossed to obtain Vegf120/120 embryos and Vegf+/+ wild type littermates. The morning of the vaginal plug was defined embryonic day (E) 0.5. Pregnant females were sacrificed by cervical dislocation. E12.5, 14.5 and 16.5 embryos were isolated in sterile PBS. The embryonic thoraces were carefully dissected in Rnase-free conditions, placed in tissue freezing medium (TBS, Triangle Biomedical Sciences, Durham N.C.), frozen and stored at -80° C. The distribution of embryos according to age and maternal origin is represented in Table S1.
[0148] Cryostat sections (8 μm) were cut and attached to SuperFrost Plus microscope slides (Menzel Gmbh & Co KG, Braunschweig Germany). Sectioning and further immunohistochemical processing of embryonic thoraces of different ages was performed at random.
Immunohistochemistry and Laser Capture Microdissection.
[0149] Three tissue sections from each embryonic thorax were selected at the level of the biventricular view of the heart. These were immunohistochemically processed in one batch. The cryostat sections were fixed by placing the slides in cold acetone (4° C.) during 2 minutes after removal from the -80° C. freezer. All further immunohistochemical steps were performed at 4° C., and all buffers and antibody solutions kept at 4° C. Rnase free PBS or D-PBS buffer was prepared by diluting RNAsecure (25×, AM7006, Ambion TX) to 1× in the desired buffer. All antibody solutions were prepared in PBS whereas the isolectin GS-IB4 conjugate was diluted in D-PBS. Superase.In (AM2696, Ambion, Austin, Tex.) was added to each antibody solution in a final concentration of 1U/μl. The slides were air-dried and the tissue sections circumscribed with a hydrophobic barrier pen. After placing the slides on a cold metal block (4° C.), 30 ml of PBS was applied to each tissue section and drained off. Subsequently, 30 ml of mouse anti-keratin pan (4, 5, 6, 8, 10, 13, 18) monoclonal antibody (MAB1636, Chemicon) in a concentration of 10 μg/100 μl was dripped on the specimen. The antibody solution was drained off after 2 minutes and the tissue section gently rinsed with 250 ml of PBS. Thirty μl of Alexa-fluor-488 chicken anti-mouse IgG (H+L) conjugate (A21200, Invitrogen CA) in a concentration of 10 μg/100 μl was then applied for 2 minutes, followed again by a gentle wash with 250 μl of PBS. Finally, a third cycle of 2 minutes staining with 30 μl of isolectin GS-IB4 Alexa Fluor 594 conjugate (I21413, Invitrogen, CA) in a concentration of 10 μg/100 μl completed the staining procedure. The tissue sections were dehydrated at room temperature: 75% EtOH (30 sec), 95% EtOH (30 sec), 100% EtOH (30 sec), 100% EtOH (120 sec), xylene (180 sec). Laser capture microdissection was performed on a Veritas Microdissection Instrument (Arcturus Bioscience Inc., Mountain View, Calif.) immediately after the dehydration steps. We dissected 3×300 to 400 cells (as triplicate samples) from intrapulmonary airways or blood vessels in the embryonic lungs of three tissue sections at the level of the biventricular view of the heart. Cells staining for mouse anti-keratin pan monoclonal antibody/chicken anti-mouse IgG Alexa-fluor-488 conjugate were identified as green fluorescent cells (blue filter). These green fluorescent cells were defined as airway epithelial cells (ker+ cells) and were randomly dissected, irrespective of their proximal or distal airway morphology. Cells staining for isolectin GS-IB4 Alexa-fluor-594 conjugate were identified as red fluorescent cells (green filter). These cells were defined as mesenchymal cells with endothelial features (il+ cells). Staining of cells for both markers was not observed on the three embryonic time points (E 12.5, 14.5, 16.5). In fact, the green and red fluorescent cells could be observed as a positive/negative image from each other. The microdissected ker+ or il+ cells were collected in a Gene Amp tube (Applied Biosystems, Foster City Calif.) filled with 75 μl of RNeasy lysis buffer (RLT; Qiagen, Hilden, Germany) containing 0.14 M β-mercaptoethanol and 200 ng polyinosinic acid (Sigma).
RNA Isolation, Amplification, Labeling, and Microarray Hybridization.
[0150] Laser-captured samples were incubated at 42° C. for 20 minutes and then chilled on ice. Samples were stored at -80° C. until further processing. After thawing, an equal volume of 70% ethanol was added to each sample and then transferred to RNeasy MinElute Spin Columns (Qiagen). RNA was cleaned up according to the manufacturer's instructions, eluted in 14 μl of RNase-free water, and adjusted to 4 μl by vacuum drying. Two rounds of linear mRNA amplification were needed to generate sufficient quantities of cRNA. Two-cycle cDNA synthesis and synthesis of biotin-labeled cRNA was performed according to the GeneChip Eukaryotic Sample and Array Processing Manual (Affymetrix, Santa Clara, Calif.). As "spike-in" controls, the GeneChip Poly-A RNA control kit (Affymetrix) was used. MEGAscript T7 kit (Ambion, Austin, Tex.) was used for in vitro transcription of the second cDNA strand in the first round of amplification, yielding 112 to 457 ng of aRNA. The second round of amplification, starting from 100 ng of first round aRNA, yielded 11 to 86 μg of cRNA using the GeneChip in vitro transcription (IVT) labelling kit. Labeled RNA was hybridized to mouse genome MG-430--2.0 GeneChip arrays (Affymetrix). Hybridization was performed using 12.5 μg of biotin-labeled RNA at 45° C. for 16 hours under continuous rotation. Arrays were stained in Affymetrix Fluidics stations using streptavidin-phycoerythrin (SAPE), followed by staining with anti-streptavidin antibody and a second SAPE staining. Subsequently, arrays were scanned with an Agilent Laserscanner (Affymetrix).
Statistical Analysis.
[0151] The Affymetrix probe level data were summarized using FARMS (Factor Analysis for Robust Microarray Summarization)1. Raw intensities were log2 transformed to get data normally distributed. First, an unsupervised multivariate projection method, Spectral Map Analysis2, was applied to reduce the complexity of highly dimensional data (n genes vs. p samples). Spectral Map Analysis provides an unbiased identification of the predominant clusters of genes and subjects that are present in the data set. Second, tests for differential gene expression between the two cellular origins (ker+ versus il+ cells) was performed in LIMMA (Linear Models for Microarray Data)3, as this method uses information across genes making the analyses stable even for experiments with small number of arrays3. Third, differences between Vegf120/120 knockout and wild type littermates in expression profiles over embryonic age were tested through a two-way interaction of Vegf genotype and time, again using LIMMA3. The data of E12.5 and E14.5 were pooled, as we were only interested in the contrasting time profile of E16.5 versus E14.5 and E12.5. This test was performed on the ker+ and il+ samples separately, because these two tissues originated from the same embryos. Models like LIMMA assume that all the samples have been randomly and independently collected. Correction for this dependency would have required too complex models if the ker+ and il+ would have been analysed concurrently. Genomic variation from the single interaction of tissue type (ker+ versus il+ samples) was also uncovered by LIMMA analysis. Allocation of differential expression along the whole genome was performed using MACT (Microarray Chromosome Analysis Tool).
Results/Discussion
[0152] At birth, O2 and CO2 need to be exchanged in the lung across a large interface of distal airways and blood vessels. Embryonic lung development in the mouse undergoes a striking shift at E (=post conceptual day) 16.51. At this time, the intertwined airway and vascular trees sprout by multiplying and refining their distal branches. The distal airways or respiratory tubules multiply by subdivision into thin walled sacculi before birth. These sacculi eventually develop into postnatal alveoli2. Thin walled airways require flat cells to facilitate gas transport. The phenotypic differentiation to flat airway cells, which occurs around E16.5, is therefore a crucial phase in embryonic lung development. Epithelial cells covering the airways originate from branching foregut endoderm. From E16.5, epithelial cells in the distal airway start to flatten out whereas proximal cell preserve their columnar shape. The most distal of these cells will line the sacculi and alveoli and develop a flat or even squamous morphology by E18.5. The capillaries are lined with flat endothelial cells and represent the distal blood vessels of the vascular tree. Endothelial cells covering pulmonary blood vessels derive from mesodermal mesenchyme. Their growth needs to closely match the growth of their epithelial counterparts in order to provide the large alveolar-capillary interface over which gas exchange initiates at birth. Reciprocal crosstalk between endodermally derived airway epithelium and surrounding mesodermal mesenchyme initiates at early lung morphogenesis3,4. Starting at E9.5, Fgf10 produced by mesenchymal cells in surrounding mesoderm is the most important cue for branching endoderm. Close interaction with at least Shh, Bmp, TGF-β, and Wnt signaling factors modulates this early branching mechanism. However, the molecular machinery underpinning the later cellular phenotypical changes and epithelial-endothelial crosstalk at E16.5 is less understood.
[0153] In order to gain further insight into late lung differentiation after E12.5, we developed a RNA-friendly immunohistochemical staining protocol for laser capture microdissection of epithelial cells in the developing airway. We reasoned that downstream gene expression profiling of RNA isolated from airway cells sharing a common epithelial antigen at different embryonic ages, should highlight their transcriptional changes over time. This program should at least reflect epithelial features, preferably of the pulmonary airway type. The same assumption was tested on pulmonary cells marked for an endothelial marker universally expressed at different embryonic ages. Additionally, a mouse knockout model with late abnormal pulmonary branching morphogenesis was incorporated in this approach. We chose the Vegf120/120 model, as peripheral airway and vascular differentiation5,6,7 becomes severely impaired in these Vegf120/120 knockout embryonic lungs. Wild type epithelial and endothelial cells were expected to express a set of airway and vascular differentiation genes, lacking in their Vegf120/120 knockout counterparts. Vegf120/120 mice lack VEGF-A isoforms 164 and 188, but still express isoform 120. VEGF-A isoforms 164 and 188 (VEGF164 and VEGF188) are more tightly bound to the extracellular matrix than the more soluble VEGF120 variant, and concentrate locally around distal airways. The standard view states that pulmonary epithelial cells secrete these VEGF-A isoforms, whereby VEGF164 and VEGF188 encourage local growth of pulmonary endothelial cells through stimulation of receptor tyrosine kinases Flk1 (VEGF receptor-2) and Flt1 (VEGF receptor-1). Confined expansion of endothelial cells refines the pulmonary vascular tree and allows a matching growth of epithelial cells8,9. This epithelial-endothelial crosstalk permits gas exchange at birth by the formation of a close bond between the alveoli of the distal airways and the capillaries of the lung vasculature. However, this type of interaction cannot explain the presence of VEGF-A in mesenchymal cells surrounding the distal airway epithelial cells.
[0154] Immunohistochemical staining was performed on frozen tissue sections cut from embryonic thoraces isolated at E12.5, E14.5 and E16.5 (FIG. 2). The genomic distribution of the embryos is shown in Table S1. We selected antibodies exhibiting sufficient bandwidth for binding epithelial or endothelial antigens in the embryonic time frame of our study. Anti-cytokeratin (directed against cytokeratin 4, 5, 6, 8, 10, 13, and 18) was chosen for labelling of epithelial cells (ker+ cells) lining the airways, as primitive and differentiated epithelial cells globally express intermediate filaments of different keratins. Endothelial cells in the same tissue section were stained with isolectin GS-IB4 (Griffonia simplicifolia) Alexa-fluor-594 conjugate, which binds early10 and late endothelial cells11,12 in the mouse (il+ cells). Staining of cells for both immunohistochemical markers was not observed on the three embryonic time points. Three hundred to four hundred ker+ and il+ cells were selectively isolated by laser capture microdissection. Two rounds of linear mRNA amplification yielded sufficient cRNA for hybridization to Affymetrix Mouse 430--2.0 Genechips.
[0155] First, we examined if downstream gene expression profiling was reflecting differentiation with respect to embryonic age and epithelial versus endothelial origin. Exploratory, unsupervised analysis of gene expression data revealed that gene expression changes during embryonic development accounted for the largest variation (35%) in the data set. This variation was graphically well represented in the first principal component (X-axis or PC1) of a spectral map13 (FIG. 3). Genes displaying the strongest expression changes over the three embryonic stages lie at the extremes of the X-axis. One of these genes, surfactant associated protein C (Sftpc), is known to have an important physiological upregulation during embryogenesis, and sufficient amounts of its protein product are necessary for normal breathing at birth14,15. The second principal component (Y-axis or PC2), explaining another 17% of the variation in the dataset, could be assigned to gene expression differences in cellular origin. Some of the most extreme probe sets on the PC2 axis represent genes that are known to be highly characteristic for either pulmonary epithelium or endothelium. Among the eight most extreme probe sets, we identified CD 93 antigen (CD93)16 and claudin 5 (Cldn5)17 as illustrative endothelial genes, and on the opposite side of the Y-axis, forkhead box A1 (Foxa1)18 and keratin 8 (Krt8)19 as expressive epithelial genes. Superposition of the different samples on the spectral map showed their distribution along the first two principal components. The ker+ and il+ groups were clearly separated according to their cellular origin along PC2 (FIG. 3). Ker+ samples grouped towards epithelial type genes and il+ samples assembled towards endothelial type genes. Both cellular origins gathered together on PC1 at the same embryonic ages. This indicates that the overall developmental gene expression changes are similar for epithelial (ker+) and endothelial (il+) cells. As a whole, the samples cluster together with respect to cellular origin (ker+ versus il+ cells) and embryonic age when applying an unsupervised data-driven analysis. The spectral map analysis underscores that selective laser capture microdissection revealed a good resolution of gene expression profiles. An independent supervised univariate (gene-by-gene) analysis of the effect of tissue origin of the samples (ker+ versus il+) further confirmed that ker+ and il+ cells corresponded at the genomic level with epithelial or endothelial cells, respectively (FIG. 1).
[0156] Next, the transcriptional profile associated with abnormal branching morphogenesis in the Vegf120/120 knockout phenotype was charted in il+ and ker+ cells. For every gene, we tested whether its expression profile over embryonic age differed significantly between Vegf120/120 knockout and wild type (Vegf+/+) littermates. This difference in age dependant expression profile between Vegf+/+ and Vegf120/120 unfolded a genomic roadmap in three directions. There were several genes identified with only in the Vegf+/+ genotype a clear upregulation over embryonic age, e.g. Hmr (FIG. 2). The Vegf120/120 genotype showed an impairment in that age dependant induction.
[0157] First, the cause of the structural deficit in the airways of Vegf120/120 knockout lungs became apparent. Wild type ker+ cells highly expressed a surplus of 44 epithelial-specific genes on E16.5 compared to their Vegf120/120 knockout counterparts. A group of genes of the Epidermal Differentiation Complex (EDC) dominated the expression profile (FIG. 4). S100a8 and S100a9 figured among this EDC subset and are known to be VEGF-A responsive20 chemoattractants. Other elements of the EDC such as small proline rich region (Spry) genes and late constituents of the epidermal cornified envelope were also present. Cytoskeletal keratin Krt2-6 was co-expressed with this EDC subset in wild type ker+ cells on E16.5. Serine-cysteine proteinase inhibitors and the three genes of the SCC (stratified epithelium-secreted protein gene) complex completed the cohort of upregulated genes (FIG. 4).
[0158] Study of the interaction between embryonic age and genotype in wild type il+ cells uncovered again a profound upregulation of a limited set of genes on E16.5. This response led towards adoption of an epithelial-specific transformation program in wild type il+ cells on E16.5 compared to Vegf120/120 knockout cells. As in wild type ker+ cells, the EDC cluster, the SCC cluster, the cysteine proteinase inhibitors and exclusive keratin genes were clearly upregulated over embryonic age. Riken1110020A10, corresponding to the Dsc1 gene, was highly upregulated in wild type il+ cells on E16.5. Furthermore, Pkp1 (plakophilin 1) displayed an identical transcriptional profile on E16.5 (FIG. 5). A highly logical pattern appeared to drive the clustered upregulation of these genes. Keratins are intermediate filament proteins lending structural strength to the cell, most typically in epithelial cells21. Proteins encoded by the EDC and SCC cluster, and serine-cysteine proteinase inhibitors fortify this keratin network. Dsc1 (desmocollin 1) codes for one of the proteins shaping the desmosomes22,23. Intermediate keratin filaments are linked to intercellular desmosomes that form the cell junction together with gap and adherens junctions. The protein encoded by Pkp1 is most of all a positive regulator of desmosomal protein content24,25 and is a constituent of the desmosomal complex itself. Pkp1 also links intermediate keratin filaments to the cadherin proteins of the adherens junctions. These results identify receptor tyrosine kinase stimulation by VEGF-A isoforms 164 and 188 as a master switch in the assembly of the desmosomal/intermediate filament machinery in the lung. This mechanism adds another building block to the cytoskeletal and intercellular architecture on top of the Wnt/β-catenin-dependent adherens junction (E-cadherin). In fact, the upregulation of Eps811 on E16.5 in wild type il+ cells revealed even direct interference with actin, a key structural protein unrelated to the intermediate filament system. The coordinated and clustered expression of these cytoskeletal and desmosomal genes permits shaping of flat or squamous cellular arrangements in distal airways.
[0159] Second, aside from activation of genes coding for specific structural proteins, an intriguing finding was the upregulation of Mapkapk3 in wild type il+ cells on E16.5. Mapkapk3 integrates ERK and p38 signaling26 pathways in stress and mitogen responses such as VEGF-A stimulation of the endothelial cell. Cdkn2b (p15.sup.ink4b or Ink4b), part of the Ink4b-ARF-Ink4a tumor suppressor locus, was simultaneously upregulated. Substantial evidence points to suppression of this locus by associated polycomb group (Pcg) repressor complexes. Derepression of the locus occurs upon dissociation of Peg complexes by activation or overexpression of Mapkapk327. This brake pedal in the cell cycle allows differentiation during proliferative stimuli28 and was demonstrated here for the first time in vivo. In effect, cell cycle arrest permitting epithelial transformation of il+ cells is VEGF164- and VEGF188-dependent in the lung. Intriguingly, upregulation of Krt5, Krt14 and Tcfap2c was strikingly similar to the expression fingerprint of the basal cell progenitor in airway epithelium29. The basal cell phenotype only appears at birth in the pulmonary airway epithelium and typically binds isolectin. On the other hand, Krt1, EDC and SCC cluster gene expression is a squamous differentiation program. The protein ANp63 and TAap63 drive the keratin progenitor and squamous differentiation program respectively. The Trp63 gene coding for these two proteins was upregulated in the wild type il+ cells. As mentioned, staining of cells for both anti-cytokeratin (anti-4, 5, 6, 8, 10, 13, and 18) and isolectin GS-IB4 was not observed at the time points studied. The lack of Krt1 and Krt14 binding by the anti-cytokeratin antibody, allowed therefore the identification of the specific epithelial transformation program of wild type il+ cells at E16.5. It seems unlikely that ker+ epithelial cells generate this epithelial transcriptional program. This would require losing the binding capacity for the anti-cytokeratin antibody in order to escape ker+ labeling. At the same time, the ker+ cells would have to acquire exclusive isolectin GS-IB4 staining. As a result, we propose that pulmonary il+ cells harbor a reservoir of cells growing into epithelial maturity at E16.5. In other words, pulmonary mesenchymal il+ cells encompass cells with endothelial and epithelial competence.
[0160] Third, the gene represented by Affymetrix probe 1437019_at (RIKEN cDNA 2200001I15 gene) and the gene represented by Affymietrix probe 1437145_s_at (RIKEN cDNA 2310002J15 gene) were both upregulated on E16.5 in wild type anti-cytokeratin 4-5-6-8-10-13-18 staining epithelial cells and wild type GS-IB4 binding cells. We found these two genes (lacking biological annotation) to be tightly co-expressed with the squamous and basal cell transcriptional program. They play an important role in production and regeneration of differentiated airway cells. (FIG. 6, FIG. 7, Table 1, Table 2). Starting from the longest contig constructed from sequenced clones, we discovered the human homologue of the RIKEN cDNA 2200001I15 gene transcript (FIG. 8) and RIKEN cDNA 2310002J15 gene transcript (FIG. 10). Furthermore, protein sequence alignments confirmed the existence of a human homologous protein for the respective translated protein of the RIKEN cDNA 2200001I15 gene (FIG. 9) and RIKEN cDNA 2310002J15 gene (FIG. 11). We suggest the name R2R1 for the mammalian homologues of the RIKEN cDNA 2200001I15 gene, transcript and protein, and R2R2 for RIKEN cDNA 2310002J15 gene, transcript and protein as these genes and their protein products are responsible for regenerative function in the respiratory system.
[0161] Fourth, we found that VEGF-A is expressed in ker+ and il+ cells, independent of wild type or Vegf120/120 knockout status. Moreover, the gene coding for VEGF receptor 1 (Flk-1 or Kdr) was abundantly expressed in il+ cells, but also increased significantly as early as E14.5 in ker+ cells. This pattern of VEGF-A and VEGF receptor expression challenges the classic viewpoint of mesenchyme passively waiting for a VEGF-A stimulus from pulmonary epithelium. It coincides essentially with recent work demonstrating the need of endogenous VEGF-A expression and autocrine signaling in survival of endothelial cells30,31. From the genomic footprint of wild type versus Vegf120/120 knockout il+ cells, it became also apparent that the il+ cells are the ones responsible for sending epithelial transformation stimuli. Upregulation of Fgfbp1, Lgals7, Lgals3 and Il18 in wild type il+ cells on E16.5 represented these stimuli. The upregulation of the gene encoding fibroblast growth factor-binding protein 1 (Fgfbp1) indicates that primordial FGF-controlled lung budding is also at work in the final stages of lung differentiation. Fgfbp1 acts by concentrating FGF2 and is a fine modulator of growth and differentiation of epithelial tissues in response to FGF stimuli from mesenchyme. The FGF receptor 2 gene (Fgfr2) was in this respect richly expressed in the ker+ and il+ compartment.
[0162] In summary, selective laser capture microdissection of cells sharing a specific marker at different embryonic ages was performed. This allowed differentiation of gene expression profiles with respect to embryonic age, cellular origin and Vegf genotype. The transcriptional program revealed by this approach highlighted the importance of the intermediate filament and desmosomal network in the refinement of pulmonary architecture. This mechanism adds another building block to the cytoskeletal and intercellular architecture on top of the Wnt/β-catenin-dependent adherens junction (E-cadherin). Intermediate filaments provide the necessary strength to cells that will be exposed to vast amounts of mechanical and oxidant stress, Not surprisingly, pulmonary cells adopt the same defensive genomic program as skin cells, which are exposed to the same challenges. In parallel with cytoskeletal sophistication, the basal cell progenitor program is acquired by i/+ cells late in embryonic life, and is VEGF 164-188-dependent,
[0163] We have demonstrated that downregulation of FAM25 family and downregulation of C9orf169 gene expression lowers KRT14 expression. Further, we know that expression of the FAM25 family (the human R2R1 homologue) and C9orf169 (the human R2R2 homologue) is upregulated by VEGFA/VEGF165. How do these genes act in the VEGFA/VEGF165 pathway? siRNA mediated knockdown of the R2R homologues can be used to uncover the specific role of the R2R homologues in the pathway, Indeed the R2R homologues' role is to modulate the VEGFA/VEGF165 response in the cell. The basal expression of VEGFA and the VEGF165 isoform is high in the pulmonary epithelium. Epithelial functions and structures are far more complex than their endothelial counterparts. Therefore, the typical VEGFA/VEGF165 effects of `grow and multiply` in the endothelium need to be more refined in the endothelium. How is this realized? Expression R2R homologues leads to simultaneous modulation of HIF1A signaling (conferring oxygen tolerance), AND modulation of a specific (PERP) anti-apoptotic pathway. This will permit the (re)generation of strong epithelial cells (with a major defense barrier against stress), without the installment of unlimited growth potential. In other words, the modulation of a specific anti-apoptotic pathway is not a `permit` for general tolerance to apoptosis. General tolerance to apoptosis would lead to the dangerous situation of immortalization of the cell: the cell develops into a cancer cell.
[0164] This study sheds new light on certain human lung diseases. In preterm neonates, late embryonic pulmonary development is disrupted when the infant is prematurely delivered. Insufficient levels of surfactant protein in pulmonary alveoli lead to severe respiratory distress in a large group of premature infants. Instillation of surfactant in the neonatal lung has aided the prevention and treatment of respiratory failure in premature infants. However, high oxygen levels and tensile stress-forces of mechanical ventilation still lead to chronic lung damage or bronchopulmonary dysplasia (BPD). Accelerating squamous differentiation in distal airways of premature infants may prevent this debilitating condition. The intermediate filament network and synchronized replenishment of the basal cell pool may also be of paramount importance in the search for a cure of certain adult lung diseases. Adoption of a squamous phenotype with expression of some EDC cluster genes characterizes squamous metaplasia in the airways of adults with chronic obstructive pulmonary disease (COPD). This defense mechanism against noxious stimuli is however accompanied with loss of regenerative basal cells32. In contrast, the embryo succeeds in developing a squamous differentiation program while simultaneously building a basal cell reservoir by a well-defined roadmap. This roadmap serves as a guide in pharmacological intervention in the intermediate filament or basal cell transcriptional machinery. It also points to transformed il+ cells as a source for pulmonary epithelial cells.
Functional Characteristics of R2R1 and R2R2
[0165] The VEGF-A (mouse isoform VEGF164=isoform VEGF165) dependent expression of the intermediate filament group of genes has been confirmed in the mouse embryo. Expression of these intermediate filament genes leads to differentiation of pulmonary epithelial cells and development of the basal cell program in pulmonary mesenchymal cells.
[0166] Furthermore, VEGF-A (mouse isoform VEGF164=isoform VEGF165) dependent expression of intermediate filament genes has also been confirmed in adult human primary epithelial cells. Stimulation of these cells by VEGF-A isoform VEGF165 leads to upregulation of intermediate filament gene expression. Intermediate filament gene expression in these cells is downregulated by isoform VEGF165 specific siRNA's. In summary, the expression of intermediate filament genes serves as a paradigm of airway differentiation and regeneration. R2R1 and R2R2 play a specific role in this expression program.
R2R1
[0167] The expression of R2R1 is VEGF-A (mouse isoform VEGF164=human isoform VEGF165) dependent in the mouse. Mesenchymal cells of the lung (GS-IB4 positive staining cells) acquire a basal epithelial cell gene expression program. The expression of R2R1 is essential for mesenchymal to epithelial transition (MET).
[0168] The role of R2R1 in MET has now been confirmed in embryonic tissue other than the lung: completion of the ventricular septum of the heart is accomplished by MET. In the mouse, R2R1 is highly expressed in the developing ventricular septum and is VEGF-A (mouse isoform VEGF164=human isoform VEGF165) dependent. Contrary, R2R1 expression is not found in the developing right ventricular outflow tract: development of this structure is known to be MET independent (see FIG. 12)
[0169] R2R1 is a candidate gene in mouse and human for mesenchymal to epithelial transition: the expression of this gene transduces the VEGF-A (mouse isoform VEGF164=human isoform VEGF165) effect on MET. The reverse process of MET is EMT (epithelial to mesenchymal transition). EMT is an essential part of cancer progression and metastasis. R2R1 may therefore play an important role in cancer biology and therapy.
[0170] Furthermore, in silico analysis revealed that the protein product of R2R1 likely interacts with the ribosome. Aside from the scientific impact, the interaction of the R2R1 protein structure with the ribosome has implications for drug development in the field of cancer biology and MET.
R2R2
[0171] R2R2 has been found to be very highly expressed in adult human primary lung epithelial cells. In vitro, the expression of R2R1 has been found to be upregulated over time and to be VEGF-A (isoform VEGF165) dependent. The R2R2 protein product is important for normal differentiation and maintenance of the adult pulmonary epithelium (see FIG. 13).
Sequence CWU
1
1
121425DNAMus musculus 1acactgacac ggaccgaagg agtggaaaaa gctttacctg
tcactgtctg ctgccatacg 60atgctgggag gcctggggaa gctggcggcc gagggcctgg
cccaccgcac agagaaagcc 120actgggggag cagttcacgc agtggaagag gtggtgagcg
aggtggtggg ccacgccaag 180gaggttggag agaagaccat taatgacgcc ctaaagaaag
cccaagaatc aggagacagg 240gtggtgaagg aggtcactga gaaggtcacc cacaccatca
ctgatgctgt tacccatgcg 300gcagaaggcc tgggaagact gggacagtga gcctgcctac
cagcatggct ggcccttcct 360gaaggtcaat aaagagtgtg aaacgtgaaa aaaaaaaaaa
aaataacaaa aaaaaaaaaa 420aaaaa
4252267DNAMus musculus 2atgctgggag gcctggggaa
gctggcggcc gagggcctgg cccaccgcac agagaaagcc 60actgggggag cagttcacgc
agtggaagag gtggtgagcg aggtggtggg ccacgccaag 120gaggttggag agaagaccat
taatgacgcc ctaaagaaag cccaagaatc aggagacagg 180gtggtgaagg aggtcactga
gaaggtcacc cacaccatca ctgatgctgt tacccatgcg 240gcagaaggcc tgggaagact
gggacag 267389PRTMus musculus 3Met
Leu Gly Gly Leu Gly Lys Leu Ala Ala Glu Gly Leu Ala His Arg 1
5 10 15 Thr Glu Lys Ala Thr Gly
Gly Ala Val His Ala Val Glu Glu Val Val 20
25 30 Ser Glu Val Val Gly His Ala Lys Glu Val
Gly Glu Lys Thr Ile Asn 35 40
45 Asp Ala Leu Lys Lys Ala Gln Glu Ser Gly Asp Arg Val Val
Lys Glu 50 55 60
Val Thr Glu Lys Val Thr His Thr Ile Thr Asp Ala Val Thr His Ala 65
70 75 80 Ala Glu Gly Leu Gly
Arg Leu Gly Gln 85 4342DNAHomo sapiens
4actgtctgct gccacacgat gctgggaggc ctggggaagc tggctgccga aggcctggcc
60caccgcaccg agaaggccac cgagggagcc attcatgccg tggaagaagt ggtgaaggag
120gtggtgggac acgccaagga gactggagag aaagccattg ctgaagccat aaagaaagcc
180caagagtcag gggacaaaaa gatgaaggaa atcactgaga cagtgaccaa cacagtcaca
240aatgccatca cccatgcagc agagagtctg gacaaacttg gacagtgagt gcacctgcta
300ccacggccct tccccagtct caataaaaag ccatgacatg tg
3425267DNAHomo sapiens 5atgctgggag gcctggggaa gctggctgcc gaaggcctgg
cccaccgcac cgagaaggcc 60accgagggag ccattcatgc cgtggaagaa gtggtgaagg
aggtggtggg acacgccaag 120gagactggag agaaagccat tgctgaagcc ataaagaaag
cccaagagtc aggggacaaa 180aagatgaagg aaatcactga gacagtgacc aacacagtca
caaatgccat cacccatgca 240gcagagagtc tggacaaact tggacag
267689PRTHomo sapiens 6Met Leu Gly Gly Leu Gly Lys
Leu Ala Ala Glu Gly Leu Ala His Arg 1 5
10 15 Thr Glu Lys Ala Thr Glu Gly Ala Ile His Ala
Val Glu Glu Val Val 20 25
30 Lys Glu Val Val Gly His Ala Lys Glu Thr Gly Glu Lys Ala Ile
Ala 35 40 45 Glu
Ala Ile Lys Lys Ala Gln Glu Ser Gly Asp Lys Lys Met Lys Glu 50
55 60 Ile Thr Glu Thr Val Thr
Asn Thr Val Thr Asn Ala Ile Thr His Ala 65 70
75 80 Ala Glu Ser Leu Asp Lys Leu Gly Gln
85 71078DNAMus musculusmisc_feature(109)..(109)n
is a, c, g, or t 7gtgactggct gctgtctcta gttgttgagg cctcttggga tctyggcgct
macmccwtgc 60tytagwgact ccgatagctc ccrmggctcc agtgsasmcc tcggkcggng
gnagggaaaa 120ggcacttgct ggtagctctg ctcacccgca ctgggacctg gagctggagg
actaagaaga 180cagacggctg ctgcttgcca cagcctggac catggacccc catgagatgg
ttgtgaagaa 240tccatatgcc cacatcagca ttcctcgggc tcacctgcgc tctgacctgg
ggcagcagtt 300agaggaggtt ccttcttcat cttcctcctc tgagactcag cctctgcctg
caggaacatg 360tatcccagag ccagtgggcc tcttacaaac tactgaagcc cctgggccca
aaggtatcaa 420gggcatcaag ggtactgctc ctgagcacgg ccagcagacc tggcagtcac
cctgcaatcc 480ctatagcagt gggcaacgtc catcgggact gacttatgct ggcctgccac
ctgtagggcg 540tggtgatgac attgcccacc actgctgctg ctgcccttgc tgctcctgct
gccactgtcc 600tcgcttctgc cgttgtcaca gctgttgtgt tatctcctag ctgactattg
aacctccagg 660gctgtgcagc ccaggttcct gctcaatgcc aaagtgttgc tggacatcag
gagcagccgt 720tgtcatgagc atcagccatt tcctgccctg agcaggggag cctgtccacc
agcgttcagc 780tgtagccttc tggaataggg ttccagccac tagccatgtt ggcaacaaca
gggacaccct 840tcacgtcctg caagactttg gcaataaagc aggatgagcg ttgctgnncc
tgntgaaaan 900aaamwaaawa cwgccgttgt cacarcygtt rtgttatctm mkagstgacw
attgtaammt 960ycagrgctgt rmagcccrgg kksckgctca atgccaaagt gttgmtgsmc
mtcrggrgsr 1020gccaagcttt acgcggtacc cgggattttt tttgtacaaa aaggggcccc
ctattagg 10788426DNAMus musculus 8atggaccccc atgagatggt tgtgaagaat
ccatatgccc acatcagcat tcctcgggct 60cacctgcgct ctgacctggg gcagcagtta
gaggaggttc cttcttcatc ttcctcctct 120gagactcagc ctctgcctgc aggaacatgt
atcccagagc cagtgggcct cttacaaact 180actgaagccc ctgggcccaa aggtatcaag
ggcatcaagg gtactgctcc tgagcacggc 240cagcagacct ggcagtcacc ctgcaatccc
tatagcagtg ggcaacgtcc atcgggactg 300acttatgctg gcctgccacc tgtagggcgt
ggtgatgaca ttgcccacca ctgctgctgc 360tgcccttgct gctcctgctg ccactgtcct
cgcttctgcc gttgtcacag ctgttgtgtt 420atctcc
4269142PRTMus musculus 9Met Asp Pro His
Glu Met Val Val Lys Asn Pro Tyr Ala His Ile Ser 1 5
10 15 Ile Pro Arg Ala His Leu Arg Ser Asp
Leu Gly Gln Gln Leu Glu Glu 20 25
30 Val Pro Ser Ser Ser Ser Ser Ser Glu Thr Gln Pro Leu Pro
Ala Gly 35 40 45
Thr Cys Ile Pro Glu Pro Val Gly Leu Leu Gln Thr Thr Glu Ala Pro 50
55 60 Gly Pro Lys Gly Ile
Lys Gly Ile Lys Gly Thr Ala Pro Glu His Gly 65 70
75 80 Gln Gln Thr Trp Gln Ser Pro Cys Asn Pro
Tyr Ser Ser Gly Gln Arg 85 90
95 Pro Ser Gly Leu Thr Tyr Ala Gly Leu Pro Pro Val Gly Arg Gly
Asp 100 105 110 Asp
Ile Ala His His Cys Cys Cys Cys Pro Cys Cys Ser Cys Cys His 115
120 125 Cys Pro Arg Phe Cys
Arg Cys His Ser Cys Cys Val Ile Ser 130 135
140 101291DNAHomo sapiens 10cttgaacccg ggaggcagag
gttgcagtga gccgagatcg cgcagctgca ctccagcctg 60ggcaacagag caagactcca
tctcagaaaa gaagcagaaa gcctccaaga gccaatggct 120ctcaagcatc ttggtctctg
ctaagaagag gctcagaggc ttagaagccc tgcctcgccg 180gggctttgag gtgtgtgagc
aatggctggg gactgcaggc ccgggaatct gagggcctca 240ccccacttcc tttccagagc
cgtgacctca ggctcacctc ctgccctcct ctcaggcaag 300ctgcagatgc cctttagggc
ccaggccatg ccccggatgt gaggggctga gtcactggtt 360tggcagtgcc cctcagagcc
caggcctggg ctgccaccca cctgaggacg agggctgggc 420cagctgtcgt gctccagttg
ctggggcctc ttgggatctt gggaacccca tctctgagcc 480ccgccccatg gccccgcccc
tcccaaggag ggaaaaggcg gctgccagtc gctcaactca 540ggcactggga cctagagctc
agaagaccga gaggacagac tgccgtgttg ccaccacagg 600ctggaccatg gacccccaag
agatggtcgt caagaaccca tatgcccaca tcagcatccc 660ccgggctcac ctgcggcctg
acctggggca gcagttagag gtggcttcca cctgttcctc 720atcctcggag atgcagcccc
tgccagtggg gccctgtgcc ccagagccaa cccacctctt 780gcagccgacc gaggtcccag
ggcccaaggg cgccaagggt aaccaggggg ctgcccccat 840ccagaaccag caggcctggc
agcagcctgg caacccctac agcagcagtc agcgccaggc 900cggactgacc tacgctggcc
ctccgcccgc ggggcgcggg gatgacatcg cccaccactg 960ctgctgctgc ccctgctgcc
actgctgcca ctgccccccc ttctgccgct gccacagctg 1020ctgctgctgt gtcatctcct
agcccagccc accctgccag ggccaggacc cagacttcag 1080caaatgtggc tcacacagtg
ccgggacatg ccgggacatg cggggtggct gttgtcatgg 1140gcgtctgccc cttcacacca
ggcactgggg ctcagaccca ccaggaaggt ggccgttcag 1200cccgagctcc tgaaacggaa
tcccaggtcc tggctggaga gggacacccc tgattacctt 1260aaggcccagg caataaagca
gggtgatctt c 129111552DNAHomo sapiens
11atggccccgc ccctcccaag gagggaaaag gcggctgcca gtcgctcaac tcaggcactg
60ggacctagag ctcagaagac cgagaggaca gactgccgtg ttgccaccac aggctggacc
120atggaccccc aagagatggt cgtcaagaac ccatatgccc acatcagcat cccccgggct
180cacctgcggc ctgacctggg gcagcagtta gaggtggctt ccacctgttc ctcatcctcg
240gagatgcagc ccctgccagt ggggccctgt gccccagagc caacccacct cttgcagccg
300accgaggtcc cagggcccaa gggcgccaag ggtaaccagg gggctgcccc catccagaac
360cagcaggcct ggcagcagcc tggcaacccc tacagcagca gtcagcgcca ggccggactg
420acctacgctg gccctccgcc cgcggggcgc ggggatgaca tcgcccacca ctgctgctgc
480tgcccctgct gccactgctg ccactgcccc cccttctgcc gctgccacag ctgctgctgc
540tgtgtcatct cc
55212184PRTHomo sapiens 12Met Ala Pro Pro Leu Pro Arg Arg Glu Lys Ala Ala
Ala Ser Arg Ser 1 5 10
15 Thr Gln Ala Leu Gly Pro Arg Ala Gln Lys Thr Glu Arg Thr Asp Cys
20 25 30 Arg Val Ala
Thr Thr Gly Trp Thr Met Asp Pro Gln Glu Met Val Val 35
40 45 Lys Asn Pro Tyr Ala His Ile Ser
Ile Pro Arg Ala His Leu Arg Pro 50 55
60 Asp Leu Gly Gln Gln Leu Glu Val Ala Ser Thr Cys Ser
Ser Ser Ser 65 70 75
80 Glu Met Gln Pro Leu Pro Val Gly Pro Cys Ala Pro Glu Pro Thr His
85 90 95 Leu Leu Gln Pro
Thr Glu Val Pro Gly Pro Lys Gly Ala Lys Gly Asn 100
105 110 Gln Gly Ala Ala Pro Ile Gln Asn Gln
Gln Ala Trp Gln Gln Pro Gly 115 120
125 Asn Pro Tyr Ser Ser Ser Gln Arg Gln Ala Gly Leu Thr Tyr
Ala Gly 130 135 140
Pro Pro Pro Ala Gly Arg Gly Asp Asp Ile Ala His His Cys Cys Cys 145
150 155 160 Cys Pro Cys Cys His
Cys Cys His Cys Pro Pro Phe Cys Arg Cys His 165
170 175 Ser Cys Cys Cys Cys Val Ile Ser
180
User Contributions:
Comment about this patent or add new information about this topic:
 |  |
 |  |
 |  |
 |  |
 | 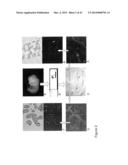 |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |
| Similar patent applications: | |
| Date | Title |
|---|---|
| 2013-06-20 | Oral insulin therapy |
| 2014-07-10 | Transgenic plants with enhanced agronomic traits |
| 2009-10-01 | Method for in vitro mass culture of aloe vera |
| 2011-10-20 | Method for diagnostic marker development |
| 2014-07-10 | Acyl-coa: diacylglycerol acyltransferase 1-like gene (ptdgat1) and uses thereof |
| New patent applications in this class: | |
| Date | Title |
|---|---|
| 2016-12-29 | Transgenic non-human mammal that expresses human mmp2 |
| 2016-07-07 | Humanized mice expressing the pyrin domain only protein 2 |
| 2016-07-07 | Method for creating endometriotic cells and endometriosis model animal |
| 2016-06-23 | Genetically modified rat models for pharmacokinetics |
| 2016-06-23 | Genetically modified rat models for pain |
| New patent applications from these inventors: | |
| Date | Title |
|---|---|
| 2017-07-13 | R2r1/2 in diagnosis and therapy |
| Top Inventors for class "Multicellular living organisms and unmodified parts thereof and related processes" | |
| Rank | Inventor's name |
|---|---|
| 1 | Gregory J. Holland |
| 2 | William H. Eby |
| 3 | Richard G. Stelpflug |
| 4 | Laron L. Peters |
| 5 | Justin T. Mason |