Patent application title: Methods and compositions for detecting and treating retinal diseases
Inventors:
George Inana (Miami, FL, US)
Margaret Jean Mclaren (Miami, FL, US)
IPC8 Class: AA61K39395FI
USPC Class:
800 9
Class name: Multicellular living organisms and unmodified parts thereof and related processes nonhuman animal the nonhuman animal is a model for human disease
Publication date: 2012-09-20
Patent application number: 20120240245
Abstract:
The invention discloses multiple genes related to age-related macular
degeneration (AMD) and/or phagocytosis by RPE cells of the eye, and
methods and compositions for detecting and treating AMD and other retinal
degenerative conditions based on these phagocytosis-related and/or
AMD-related genes. Also provided are nonhuman transgenic animal models
useful for testing therapeutic compounds and treatment protocols for AMD,
and gene arrays including polymorphic variants of phagocytosis-related
and/or AMD-related genes, useful for genetic screening of nucleic acid
samples from subjects to obtain profiles of polymorphic variant sequences
in a plurality of genes associated with AMD. Several preferred
embodiments of the therapeutic compositions and animal models are based
on target genes MT1-MMP and casein kinase 1 epsilon (CK1ε),
phagocytosis-related genes found to be over-expressed in human donor eye
samples from patients having both wet and dry forms of AMD.Claims:
1. A method for treating a subject having a retinal or choroidal
degenerative disease or condition comprising administering to the eye of
said subject an antibody that specifically binds to a casein kinase 1
epsilon protein or peptide.
2. The method of claim 1, wherein said protein is a human casein kinase 1 epsilon encoded by the nucleic acid sequence set forth as SEQ ID NO:9, or a polymorphic variant thereof.
3. The method of claim 1, wherein said retinal or choroidal degenerative disease or condition is the wet or dry form of age-related macular degeneration (AMD).
4. The method of claim 1, wherein said antibody contacts a retinal cell type selected from a photoreceptor, an RPE cell, or a Muller cell, or a cell type of the choroid selected from an endothelial cell, a smooth muscle cell, a leukocyte, a macrophage, a melanocyte or a fibroblast.
5. The method of claim 1, wherein the antibody is administered by intraocular injection.
6. The method of claim 1, wherein the antibody is selected from the group consisting of a polyclonal antibody, a monoclonal antibody, a single chain antibody, an Fab fragment and an (Fab')2 fragment.
7. The method of claim 1, wherein the antibody is produced using a Fab library.
8. A method for treating a subject having a retinal or choroidal degenerative disease or condition comprising administering to the eye of said subject an inhibitory oligonucleotide that specifically downregulates the expression of casein kinase 1 epsilon, selected from the group consisting of a ribozyme, an antisense RNA, an interfering RNA (RNAi) molecule, a small inhibitory RNA (siRNA) molecule, and a triple helix forming molecule.
9. The method of claim 8, wherein the inhibitory oligonucleotide is a siRNA molecule.
10. The method of claim 9, wherein the siRNA molecule is a double-stranded RNA molecule consisting of the nucleotide sequences set forth as SEQ ID NOS:138 and 139.
11. A nonhuman transgenic animal comprising an isolated nucleic acid construct comprising a transgene driven by a promoter, wherein said construct causes over-expression, in at least one cell type of said animal, of a gene that is over-expressed in the eyes of human subjects with AMD, selected from the group consisting of membrane type matrix metalloproteinase 1 (MT1-MMP), casein kinase 1 epsilon (CK1.epsilon.), prostaglandin D2 synthase, and gene AMDP-3.
12. The nonhuman transgenic animal of claim 11, wherein said over-expression is conditionally controlled.
13. The nonhuman transgenic animal of claim 11, wherein said cell type is a retinal cell type selected from the group of consisting of a photoreceptor, an RPE cell, and a Muller cell, or a choroidal cell type selected from the group consisting of an endothelial cell, a smooth muscle cell, a leukocyte, a macrophage, a melanocyte, and a fibroblast.
14. The nonhuman transgenic animal of claim 12, wherein the transgene comprises a sequence that encodes MT1-MMP.
15. The nonhuman transgenic animal of claim 12, wherein the transgene comprises a sequence that encodes CK1 epsilon.
Description:
CROSS-REFERENCE TO RELATED APPLICATIONS
[0001] The present application is a continuation-in-part of co-pending U.S. patent application Ser. No. 11/924,346, filed Oct. 25, 2007, which is a continuation of U.S. patent application Ser. No. 10/773,446, filed Feb. 9, 2004, now U.S. Pat. No. 7,309,487 entitled "Methods and Compositions For Detecting and Treating Retinal Diseases," which are hereby incorporated by reference in their entireties.
BACKGROUND OF THE INVENTION
[0002] Age-related macular degeneration (AMD) is the number one cause of blindness for the elderly population over 60 years of age. It is a devastating disease that destroys central vision in the affected individuals, robbing them of their ability to perform activities necessary for everyday life such as reading and driving (Bressler et al., 1988; Evans, 2001; Gottlieb, 2002). In one study, the prevalence of AMD in persons 75 or older has been reported to be 7.8% (Klein et al., 1992).
[0003] AMD is a slow, progressive disease that involves cells of the outer retinal layers (including photoreceptors and the retinal pigment epithelial (RPE) cells that support the photoreceptors), as well as cells in the adjacent vascular layer of the eye known as the choroid. Macular degeneration is characterized by the breakdown of the macula, a small portion of the central retina (about 2 mm in diameter) responsible for high-acuity vision. Late-onset macular degeneration (i.e., AMD) is generally defined as either "dry" or "wet." The wet ("exudative") neovascular form of AMD affects approximately 10% of those with the disease, and is characterized by abnormal blood vessels growing from the choriocapillaris through the RPE, typically resulting in hemorrhage, exudation, scarring, and/or serous retinal detachment. Approximately 90% of patients with AMD have the non-neovascular dry form, characterized by atrophy of the RPE and loss of macular photoreceptors.
[0004] One of the clinical hallmarks of AMD is the presence of deposits of debris-like material, termed "drusen," that accumulate on Bruch's membrane, a multilayered composite of extracellular matrix components separating the RPE (the outermost layer of the retina) from the underlying choroid. Drusen can be observed by funduscopic eye examination. These deposits have been extensively characterized in microscopic studies of donor eyes from patients with AMD (Sarks, et al., 1988). The deposits observed in the living eye upon clinical examination are classified as either soft drusen or hard drusen, according to several criteria including relative size, abundance, and shape of the deposits (reviewed, for example, in Abdelsalam et al., 1999). Histochemical and immunocytochemical studies have shown that drusen contain a variety of lipids, polysaccharides, glycosaminoglycans and proteins (Abdelsalam et al., 1999; Hageman et al., 1999, 2001).
[0005] There is presently no cure for AMD. Several types of treatments are available, with laser photocoagulation of abnormal vessels in the wet form of the disease being the standard (Gottlieb, 2002; Algvere and Seregard, 2002). This treatment is limited by the fact that only well-delineated neovascular lesions can be treated in this way and that 50% of patients will suffer recurrence of the leakage from the vessels (Fine et al., 2000). Because of the energy of the laser required for this treatment, the photoreceptors in the treated area will also die, and the patient will also often suffer central blindness immediately after the treatment. New neovascular lesions will eventually develop, requiring repeated treatments.
[0006] Photodynamic therapy, which combines low energy laser activation with a photosensitive agent, has been a valuable addition to the laser treatment approach (Bressler, 2001). In this method, a photosensitive agent, i.e., verteporfin is used which has an affinity for abnormal new blood vessels. Selective targeting of these vessels can be activated by nonthermal laser to produce reactive oxygen species which can destroy the abnormal vessels. In a study group, only 33% of those receiving photodynamic therapy with verteporfin had substantial loss of vision, compared to 61% of those who did not receive verteporfin. The treatment, however, was only beneficial for patients with classic choroidal neovascular membranes. The full long-term benefit of this new treatment modality has yet to be established. Despite this advance, however, the treatment does not prevent the subsequent formation of new neovascular lesions.
[0007] Other available treatments for the wet form of AMD include submacular surgery and external-beam radiation therapy. Those under study include retinal translocation and inhibition of vascular endothelial growth factor (Algvere and Seregard, 2002). For prevention of progression to advanced AMD, treatment with antioxidants, including vitamins C and E, β-carotene, and zinc, was shown to be helpful, and prophylactic laser treatment is under study (Gottlieb, 2002).
[0008] Despite the above-described advances, it is recognized that current treatment for AMD is mostly palliative (Algvere and Seregard, 2002). None of the available treatments attacks the fundamental cause of the disease, which is unknown. The disease therefore can continue to progress following treatment, with re-development of neovascularization and destruction of the macula. Accordingly, there remains a compelling need to understand the molecular mechanism of this disease, so that therapeutic treatment or cure can be directed at its root cause.
[0009] It is well recognized that genetic factors play an important role in the etiology of AMD. For example, it has been reported that people with a family history of AMD and siblings of AMD patients have a higher risk of developing AMD (Evans, 2001). Monozygotic twins have shown a higher concordance rate of clinical features of AMD compared to dizygotic twins (Klein et al., 1994). Another study found all monozygotic twins affected with AMD to be concordant for AMD while only 42% of dizygotic twins were concordant (Meyers et al., 1995). Accordingly, one major approach to understanding AMD etiology is to look for genes involved in AMD. For example, approaches such as linkage analysis in large families, allele sharing analysis among sib pairs, and association studies in populations have been used in attempts to identify genes associated with AMD (Guymer, 2001). Linkage to chromosomal region 1q was reported in a large AMD family (Klein et al., 1998). Results of an allele sharing analysis did not yield any new candidate genes (Weeks et al., 2000). An association of a mutation in hemicentin-1 has been reported in a familial form of age-related macular degeneration mapping to human chromosome 1q in a large family (Schultz et al., 2003).
[0010] Another genetic strategy for AMD is to test genes causing other forms of inherited macular degenerations as putative causative genes ("candidate genes") for AMD. Several macular diseases with a clearly hereditary pattern of inheritance (so-called "Mendelian macular degenerations") have been described that resemble AMD in phenotype. Examples of these diseases include Sorsby's fundus dystrophy, Stargardt's disease, Best disease, and Doyne's honeycomb retinal dystrophy (Guymer, 2001). Causative genes for these diseases have been analyzed as candidate genes for AMD. To date however, none of them has clearly demonstrated a causal relationship with AMD. For example, the ATP-binding cassette transporter gene (ABCR) was found to be the pathogenic gene for recessive Stargardt's disease (Hutchinson et al., 1997). ABCR was proposed as a candidate gene for AMD, and in one study, 16% of patients with AMD were initially shown to have mutations in this gene (Allikmets et al., 1997). This conclusion, however, has been challenged (Stone et al., 1998).
[0011] The most likely reason for the failure to find AMD genes through classical genetic approaches such as chromosomal mapping, genetic linkage analysis, and candidate gene analysis, is that AMD is a "multigene," or "complex" genetic disease. Complex genetic diseases are those diseases believed to be caused by changes in multiple genes. Such diseases characteristically demonstrate a complex pattern of inheritance (Heiba et al., 1994; Klein et al., 1994). In the case of AMD, a disease of old age, it is generally thought that the course of the disease is influenced not only by the combined effects of the above-described multiple genetic factors, but is further affected by certain environmental risk factors.
[0012] A second broad approach aimed at discovering causative genes in AMD has been hypothesis-based research aimed at elucidating the mechanism of the disease, with the goal of secondarily identifying the genes involved in the mechanism. Numerous hypotheses regarding the pathogenic mechanism of AMD have been proposed and tested, resulting in a voluminous literature on this subject.
[0013] Oxidative damage has been one major theme as a proposed mechanism for AMD (Winkler et al., 1999; Evans, 2001; Husain et al., 2002). The retina is known to have an extremely high consumption of oxygen, and the photoreceptors and RPE are in a very oxygen-rich environment. The RPE is situated immediately adjacent to the choriocapillaris, a rich capillary plexus coursing with highly oxygenated blood. The retina is a light-sensitive organ in which photoactivated events are constantly occurring during times of light exposure, resulting inter alia in the production of reactive oxygen species. In general support of the oxidative damage hypothesis, antioxidants tested in clinical studies have been reported to have a moderate beneficial effect of reducing progression to severe AMD (Hyman and Neborsky, 2002), although the results of several studies are conflicting (Flood et al., 2002). Smoking, which can reduce plasma levels of antioxidants, has been associated with increased risk of AMD (Mitchell et al., 2002). Adding support to the oxidative damage theory is a recent proteomic analysis of drusen, which demonstrated the presence in these deposits of several oxidation-modified products (Crabb et al., 2002).
[0014] It has been proposed that dysfunction in the RPE is central to the pathogenesis of AMD and can lead to drusen formation (Hogan, 1972). The earliest known sign of RPE dysfunction is accumulation of lipofuscin, which may lead to the characteristic thickening of Bruch's membrane, formation of drusen, and choroidal neovascularization observed in the wet form of AMD (Gass et al., 1985; Sarks et al., 1988; Green, 1999). Lipofuscin is composed of oxidized, polymeric molecules derived mostly from phagocytosed membranes of photoreceoptor outer segments (OS) (Katz, 1989; Kennedy et al., 1995). OS membranes are known to be rich in polyunsaturated fatty acids, which are an excellent substrate for peroxidation (Katz, 1989). It is believed that these molecules cannot be degraded and therefore begin to accumulate in the RPE cells as lipofuscin. At least one component of lipofuscin, i.e., the fluorophore A2E, a pyridinium bisretinoid, has been demonstrated to be toxic, causing membrane destabilization (De and Sakmar, 2002), and inhibition of cytochrome c oxidase and apoptosis in cultured porcine and human RPE cells (Shaban et al., 2002). Thus, A2E and lipofuscin accumulation in the RPE is thought to be directly related to dysfunction and demise of these cells with aging.
[0015] The processes of oxidative damage, lipofuscin accumulation, and drusen formation are not limited to AMD, but rather occur to some extent in all individuals with advancing age. Accordingly, a fundamental question that remains unanswered is why these processes are more advanced in some people than others, leading to AMD. Progress in developing new therapies targeting the root cause of AMD will require much greater knowledge of specific gene targets involved in the key cellular metabolic pathways in photoreceptors, RPE and choroidal cells that contribute to the observed pathology.
SUMMARY OF THE INVENTION
[0016] The invention provides novel methods and compositions for screening and treating retinal degenerative conditions, including age-related macular degeneration (AMD), as well as animal models useful for testing therapeutic compounds and methods. The invention is the product of a gene discovery strategy resulting in isolation of genes showing differential expression 1) in AMD-affected vs. normal eye tissues and 2) during the process of phagocytosis of outer segments (OS) by RPE cells. OS phagocytosis is a critical function of the RPE cells, involving a complex multi-step process, the byproducts of which contribute to generation of reactive oxygen species and lipofuscin accumulation in the RPE cells.
[0017] Using a novel expression cloning strategy termed CHANGE (for Comparative Hybridization ANalysis of Gene Expression) at least 200 AMD-related genes and at least 60 phagocytosis-related genes expressed in RPE cells were isolated. Five previously uncharacterized genes were identified by this strategy and demonstrated to be related to AMD and/or RPE phagocytosis. The nucleic acid sequences of cDNAs encoding the products of these genes are listed herein as SEQ ID NOS:1, 4, 5, 12, and 17.
[0018] A subset of six genes, termed "AMD/phagogenes," or "AMDP genes" are further described herein that fit the dual criteria of relatedness to AMD and to RPE phagocytosis. Four of these genes, i.e., prostaglandin D2 synthase (SEQ ID NO:2), casein kinase 1 epsilon (CK1ε) (SEQ ID NO:9), matrix metalloproteinase, membrane-type 1 (MT1-MMP) (SEQ ID NO:15), and unknown RPE-expressed cDNA AMDP-3 (SEQ ID NO:17) all demonstrate up-regulation in AMD. AMDP genes down-regulated in AMD include ferritin heavy polypeptide 1 (SEQ ID NO:10), and SWI/SNF related/OSA-1 nuclear protein (SEQ ID NO:16).
[0019] Other genes previously not known to be functionally related to RPE phagocytosis are disclosed herein, including unknown PHG-1 (SEQ ID NO:1), myelin basic protein (SEQ ID NO:3), unknown PHG-4 (SEQ ID NO:4), unknown PHG-5 (SEQ ID NO:5), peanut-like2/septin 4 (SEQ ID NO:6), coactosin-like 1 (SEQ ID NO:7), clusterin (SEQ ID NO:8), metargidin (SEQ ID NO:11), unknown PHG-13 (SEQ ID NO:12), retinaldehyde binding protein 1 (SEQ ID NO:13), and actin gamma 1 (SEQ ID NO:14).
[0020] An exemplary AMDP gene discovered by the above strategy is the membrane-type matrix metalloproteinase 1 (MT1-MMP) (SEQ ID NO:15). MT1-MMP is a gene encoding a protease involved in the remodeling of extracellular matrix, for example by specifically activating pro-gelatinase A. Gelatinase A is the major metalloproteinase responsible for specific cleavage of type IV collagen, the main structural component of basement membranes. MT1-MMP also shows activity against other extracellular matrix components.
[0021] It has been demonstrated that MT1-MMP is a highly attractive therapeutic target for screening and treating AMD and other retinal conditions, based on the following findings: 1) MT1-MMP is upregulated in the RPE and photoreceptors in the eyes of patients with AMD, in a monkey model of AMD, and in the RCS rat, a model of retinal degeneration involving a defect in OS phagocytosis by the RPE; 2) MT1-MMP is directly involved in the mechanism of phagocytosis by RPE cells; 3) the progress of retinal degeneration in the RCS rat is significantly reduced by blocking activated MT1-MMP present in the subretinal space with an anti-MT1-MMP antibody; 4) a synonymous polymorphism of MT1-MMP (i.e., P259P) that could produce a splice variant of the mRNA resulting in a truncated protein, and a missense polymorphism of MT1-MMP (i.e., D273N) affecting the catalytic domain of the protein are found with higher frequency in the DNA of patients with AMD (54.8% vs. 31.6%) and familial maculopathies (68.2% vs. 31.6%).
[0022] Another exemplary AMDP gene discovered by the CHANGE strategy is casein kinase 1 epsilon (CK1ε) (SEQ ID NO:9). At the time of our initial discovery of this gene (in clone 57-29) by the CHANGE analysis, the function of this gene was unknown. It later became known from mapping studies that CK1ε is the gene associated with the tau mutantaton in Syrian hamsters and is a key regulator of circadian rhythmicity (Lowrey et al., 2000). Studies described herein have revealed that this gene is intimately involved in phagocytosis of photoreceptor OS by RPE cells and is over-expressed in several animal models of AMD. Importantly, cigarette smoking, the most significant environmental factor associated with AMD, is shown herein to cause over-expression of CK1ε in mice, with phase shifting of the peak time of expression and an apparent uncoupling of the cellular "clocks" that control diurnal gene expression in the retina and in the RPE/choroid of the eye. Such dysregulation in circadian rhythmicity between interacting cell types, i.e., photoreceptors and RPE cells within these two tissues, is likely to cause stress on the RPE cells, which are tasked with daily phagocytosis, already a metabolically demanding process in itself. Such stress over an extended period in the lifetime of the animal could lead to the accumulation of lipofuschin and drusen deposits characteristic of AMD.
[0023] Based on the foregoing discoveries, it is an object of the invention to provide a method for delaying or reversing a retinal or choroidal degenerative disease or condition in a subject. The method includes contacting a retinal or choridal cell of a subject having, or at risk of developing, a retinal or choroidal degenerative disease or condition with an agent that modulates the expression or activity of an AMDP-related or phagocytosis-related gene. The AMDP-related or phagocytosis-related gene can be human unknown PHG-1; prostaglandin D2 synthase; myelin basic protein; human unknown PHG-4; human unknown PHG-5; human peanut-like 2/septin 4; coactosin-like 1; clusterin; casein kinase 1 epsilon; ferritin heavy polypeptide 1; metargidin; human unknown PHG-13; retinaldehyde binding protein 1; actin gamma 1; matrix metalloproteinase, membrane-associated 1 (MT1-MMP); SWI/SNF related/OSA-1 nuclear protein; and human unknown AMDP-3. The foregoing AMDP-related or phagocytosis-related genes include, respectively, the nucleotide sequences identified herein as SEQ ID NOS:1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, and 17.
[0024] Preferred genes targeted for modulation of expression or activity are prostaglandin D2 synthase, CK1ε, MT1-MMP and unknown gene AMDP-3, shown herein to be up-regulated in AMD. In one particularly preferred embodiment, the agent is directed against a MT1-MMP nucleic acid or protein. In another particularly preferred embodiment, the agent is directed against a CK1ε nucleic acid or protein.
[0025] The retinal or choroidal degenerative disease or condition can be AMD. The method can be used to treat a subject suffering from AMD, or at risk of developing AMD.
[0026] The method can delay the retinal or choroidal degenerative disease or condition, or it can reverse the disease or condition.
[0027] The cell type to be contacted in the practice of the method can be a photoreceptor, an RPE cell or a Muller cell, or a cell type of the choroid, including an endothelial cell, a smooth muscle cell, a leukocyte, a macrophage, a melanocyte or a fibroblast.
[0028] In a preferred embodiment of the method, in which the AMDP-related or phagocytosis-related gene is MT1-MMP, the MT1-MMP may be located within the cell or in an extracellular matrix, such as an interphotoreceptor matrix.
[0029] In some embodiments of the method, the agent down-regulates expression of a nucleic acid or amino acid sequence of an AMDP-related or phagocytosis-related gene. In preferred embodiments, the targeted genes include MT1-MMP, CK1ε, prostaglandin D2 synthase and AMDP-3, which genes are shown herein to be over-expressed in AMD. The agent may be an oligonucleotide, for example a ribozyme, an antisense RNA, an interfering RNA (RNAi) molecule, a small inhibitory (siRNA) molecule or a triple helix forming molecule. The agent may also be an antibody that specifically binds to a MT1-MMP, CK1ε, prostaglandin D2 synthase or AMDP-3 protein or peptide. Preferably the antibody can neutralize at least one biological activity of the protein or peptide. For example, an antibody directed against MT1-MMP can neutralize activation of a progelatinase A, or degradation of an extracellular matrix component. An antibody directed against CK1ε can cause a change in circadian rhythmicity, for example, a change or "phase shift" in the timing of a biological function that follows a characteristic circadian rhythm, such as the process of shedding and phagocytosis of photoreceptor outer segments (OS) in the eye, in which phagocytosis of OS by the RPE cells normally peaks at the time of light onset in the morning.
[0030] In another embodiment, the agent that down-regulates expression of MT1-MMP, prostaglandin D2 synthase, CK1ε, or AMDP-3 can be a small molecule.
[0031] It is a further object of the invention to provide a method of determining risk of a subject of developing a retinal or choroidal degenerative disease or condition. The method includes screening a nucleic acid sequence of the subject for the presence of at least one polymorphism in at least one phagocytosis-related or AMDP-related gene, wherein the presence of a polymorphism indicates that the subject is at higher risk for developing a retinal degenerative disorder than a subject without the polymorphism. The phagocytosis-related genes can include, but are not limited to, unknown PHG-1, prostaglandin D2 synthase, myelin basic protein, unknown PHG-4, unknown PHG-5, peanut-like 2/septin 4, coactosin-like 1, clusterin, casein kinase 1 epsilon, ferritin heavy polypeptide 1, metargidin, unknown PHG-13, retinaldehyde binding protein 1, actin gamma 1, membrane type metalloprotinase 1 (MT1-MMP), SWI/SNF related/OSA-1 nuclear protein, and unknown AMDP-3. Nucleic acids encoding these phagocytosis-related gene products include, respectively, cDNA sequences listed herein as SEQ ID NOS:1-17.
[0032] The AMDP-related genes to be screened in the method can include, but are not limited to, prostaglandin D2 synthase, CK1ε, ferritin heavy polypeptide 1, SWI/SNF related/OSA-1 nuclear protein, and AMDP-3. Nucleic acids encoding these AMDP-related gene products include, respectively, cDNA sequences listed herein as SEQ ID NOS:2, 9, 10, 16 and 17.
[0033] The polymorphisms screened in the method can be within an intronic, exonic or promoter region of the gene of interest.
[0034] In a preferred embodiment of the screening method, the gene of interest is MT1-MMP. The polymorphism can be within a region of the human MT1-MMP gene that can be amplified by PCR using amplimer pairs having nucleic acid sequences selected from the following groups: SEQ ID NOS:18 and 19; 20 and 21; 22 and 23; 24 and 25; 26 and 27; 28 and 29; 30 and 31; 32 and 33; 34 and 35; 36 and 37; 38 and 39; 40 and 41; 42 and 43; 44 and 45; 46 and 47; 48 and 49; 50 and 51; 52 and 53; 54 and 55; 56 and 57; and 57 and 58.
[0035] In a particularly preferred embodiment of the method, the polymorphism is within a 285 bp fragment of exon 5 of the human MT1-MMP gene. Within this region, the polymorphisms can include a D273N missense polymorphism and a P259P synonymous polymorphism.
[0036] It is also an object of the invention to provide a method of treating a retinal or choroidal degenerative disease or condition in a subject. The method includes contacting a retinal or choroidal cell of the subject with a vector that includes a nucleic acid encoding an agent that down-regulates or inhibits expression of a phagocytosis-related or AMDP-related mRNA or protein. The agent included in the vector can be an antisense RNA, a ribozyme, or an interfering RNA (RNAi) molecule. In preferred embodiments, the phagocytosis-related or AMDP-related genes targeted for down-regulation are prostaglandin D2 synthase, MT1-MMP, and AMDP-3, comprising respectively the nucleic acid sequences shown herein as SEQ ID NOS:2, 15 and 17.
[0037] In another aspect, the invention provides a method of treating a retinal or choridal degenerative disease or condition using a vector to deliver a desired form of a phagocytosis-related or AMDP-related gene product to a subject in need thereof. The vector can include a nucleic acid encoding either a wild type or polymorphic variant of a phagocytosis-related or AMDP-related gene.
[0038] Yet another embodiment of the invention is a composition for prevention or treatment of a retinal or choroidal degenerative disease or condition in a subject comprising an agent that blocks the expression or activity of a phagocytosis-related or AMDP-related gene. In some embodiments, the agent can be an antisense RNA, a ribozyme, an interfering RNA (RNAi) molecule or a small interfering RNA (siRNA) molecule. A particularly preferred siRNA molecule useful for knockdown of MT1-MMP is a double-stranded siRNA molecule consisting of the RNA sequences set forth in SEQ ID NOS:138 and 139.
[0039] The agent can also be an antibody or a small molecule.
[0040] Also within the invention are compositions for prevention or treatment of a retinal or choroidal degenerative disease or condition in a subject comprising a vector. In various embodiments, the vectors can include a nucleic acid encoding an agent that down-regulates or inhibits expression of a phagocytosis-related or AMDP-related mRNA or protein, or a nucleic acid that encodes a wild type or polymorphic variant of a phagocytosis-related or AMDP-related protein. In preferred embodiments, the phagocytosis-related or AMDP-related genes include MT1-MMP, CK1ε, prostaglandin D2 synthase and AMDP-3. In particularly preferred embodiments, the gene is MT1-MMP or CK1ε.
[0041] The invention further provides several embodiments of nonhuman transgenic animals useful, for example, as models of AMD and other retinal degenerative conditions. Preferably, the transgenic animal is a mammal, more preferably a rodent, and most preferably a mouse. In one embodiment, a transgenic animal includes an isolated nucleic acid construct that causes at least one cell type of the animal to over-express a phagocytosis-related or AMDP-related gene. The phagocytosis-related or AMDP-related gene is preferably MT1-MMP, CK1ε, prostaglandin D2 synthase, or AMDP-3. Some preferred versions of the transgenic animals are engineered to overexpress the phagocytosis-related or AMDP-related gene product in particular cell types, including retinal cell types selected from photoreceptors, RPE cells and Muller cells, and choroidal cell types including endothelial cells, smooth muscle cells, leukocytes, macrophages, melanocytes and fibroblasts. In some embodiments, the gene of interest is conditionally over-expressed.
[0042] Another preferred embodiment of an animal model of AMD/retinal degeneration is a nonhuman transgenic animal including an isolated nucleic acid construct that causes at least one cell type of the animal to express a polymorphic variant of a phagocytosis-related or AMDP-related nucleic acid and/or protein. In preferred embodiments, the nucleic acid and/or protein is MT1-MMP, CK1ε, prostaglandin D2 synthase, or AMDP-3. The polymorphic variant can be increased in incidence in a population of humans with AMD, compared to a normal control population.
[0043] Yet another embodiment is a nonhuman polytransgenic animal including at least a first isolated nucleic acid construct and at least a second isolated nucleic acid construct, the first construct causing at least one cell type of the animal to express a polymorphic variant of a first gene correlated with increased incidence of AMD, and the second nucleic acid construct causing at least one cell type of the animal to express a polymorphic variant of a second gene correlated with increased incidence of AMD, or having an association with RPE phagocytosis.
[0044] In preferred embodiments of the polytransgenic animals, the first gene is MT1-MMP and the second gene is selected from ABCR, apolipoprotein E, C--C chemokine receptor-2, cystatin C, hemicentin/FIBL-6, manganese superoxide dismutase, C--C chemokine ligand/monocyte chemoattractant protein 1, and paraoxonase.
[0045] In other preferred embodiments of the polytransgenic models, the first gene is MT1-MMP and the second gene is a phagocytosis-related or AMDP-related gene selected from human unknown PHG-1, prostaglandin D2 synthase, myelin basic protein, human unknown PHG-4, human unknown PHG-5, human peanut-like 2/septin 4, coactosin-like 1, clusterin, CK1ε, ferritin heavy polypeptide 1, metargidin, human unknown PHG-13, retinaldehyde binding protein 1, actin gamma 1, SWI/SNF related/OSA-1 nuclear protein, and human unknown AMDP-3.
[0046] Particularly preferred embodiments of the transgenic animals of the invention are mice, which provide the advantage of a relatively short life span, making them more suitable for study of age-related diseases than other longer-lived animal models such as monkeys.
[0047] In yet another aspect, the invention provides isolated nucleic acids encoding previously uncharacterized gene products shown herein to be phagocytosis-related and/or AMDP-related proteins. The nucleic acids encoding these proteins include nucleic acid sequences comprising SEQ ID NOS:1, 4, 5, 12, and 17.
[0048] The invention further provides a gene array including a plurality of isolated oligonucleotide sequences, said sequences being positioned within an intronic, exonic or promoter sequence of a native human AMD-related or phagocytosis-related gene. The genes represented by the oligonucleotide sequences in the array encode cDNAs comprising nucleic acid sequences shown herein as SEQ ID NOS:1-17 and SEQ ID NOS:62-69.
[0049] In preferred embodiments of the gene array, at least one gene is MT1-MMP and the oligonucleotide sequences include a P259P or a D273N polymorphic variant of the MT1-MMP coding sequence. These variants of MT1-MMP are shown herein to be increased in frequency in a population of patients with AMD and other macular degenerative conditions, relative to their frequency in a population of normal control subjects.
[0050] The gene array can further include at least one oligonucleotide sequence comprising at least one polymorphic variant of one or more AMD-related genes besides MT1-MMP. The polymorphic variant sequences can include: ABCR (D217N; G1961E), manganese superoxide dismutase (V47A), apolipoprotein E (C130, R176C and C130R, R176), cystatin C (A25T) and paraoxonase (Q192R, L54M).
[0051] The gene arrays of the invention are useful, for example, for screening DNA samples from subjects to determine the distribution of polymorphic variants of a plurality of AMD-related and/or phagocytosis-related genes in the subject's DNA. In keeping with the multi-gene (complex disease) etiology of AMD, it is contemplated that information pertaining to the distribution of combinations of particular polymorphic variants of AMD-related or phagocytosis-related genes in a subject's DNA can be used to predict the likelihood that the subject is at greater risk of developing a retinal disorder such as AMD than is a subject lacking said combination of particular polymorphic variants of AMD-related or phagocytosis-related genes.
[0052] The gene arrays of the invention, tailored to AMD and related disorders, can provide a convenient and relatively inexpensive means of testing polymorphic variants of a plurality of genes known to be related to AMD and related disorders.
[0053] These and other objects of the invention are set forth in more detail in the description and examples below, which are intended to illustrate the invention but not limit the scope thereof.
BRIEF DESCRIPTION OF THE DRAWINGS
[0054] The drawings form part of the present specification and are included to further demonstrate certain aspects of the present invention. The invention may be better understood by reference to one or more of the following drawings in combination with the detailed description of specific embodiments presented herein.
[0055] FIG. 1 is a photograph showing duplicate CHANGE array panels, each containing 96 genes (spots) hybridized with "+" and "-" probes (Probes 1 and 2), according to an embodiment of the invention. Up and down arrows indicate genes showing increased or decreased expression, respectively, upon hybridization with Probe 1 vs. Probe 2.
[0056] FIG. 2 (upper panel) shows a schematic drawing of a vital assay of rod outer segment (ROS) phagocytosis by cultured RPE cells. The lower panel shows black and white photographs of living BPEI-1 RPE cells undergoing phagocytosis after ROS feeding, according to an embodiment of the invention. When observed by fluorescence microscopy, lysosomes in the RPE cells appear red due to sulforhodamine (SR) staining and FITC-stained ROS appear green. During successive stages of phagocytosis, ROS are bound to the cell surfaces, then ingested by the RPE cells, first becoming phagosomes and then phagolysosomes (distinguishable by yellow-orange fluorescence) upon fusion with lysosomes.
[0057] FIG. 3 is a series of photographs showing different stages of ROS phagocytosis viewed in large scale cultures of living BPEI-1 RPE cells at the indicated times after feeding with FITC-ROS, according to an embodiment of the invention. The upper four panels show massive binding of ROS to the cell surfaces during the first 9-10 hours after feeding. The lower four panels show synchronous ROS ingestion and formation of phagolysosomes, starting approximately 11 hours after feeding with ROS.
[0058] FIG. 4 is a graph showing the mRNA expression profiles of 16 phagocytosis-related genes ("phagogenes") expressed by RPE cells, discovered by CHANGE, according to an embodiment of the invention. Expression levels of phagogenes fluctuate in RPE cells at selected times during the course of ROS phagocytosis in vitro. Identities of the phagogenes (PHG-1-16) are provided in Table 1, infra.
[0059] FIG. 5 is three photographs showing the grading system used to classify human donor eyes for AMD-related changes in the retina, according to an embodiment of the invention. Grades shown: 0-+1, minimal thickening of the Bruch's membrane; +2-+3, multiple small to mid size drusen, with thickened Bruch's membrane; +3-+4, large coalescing drusen.
[0060] FIG. 6 is a two Northern blots and a graph showing expression of MT1-MMP and actin mRNA during phagocytosis by cultured RPE cells at 4 and 13 hours after ROS feeding. Decreased expression at 4 hours and increased expression at 13 hours is seen, confirming results obtained by CHANGE. The amount of RNA present in each lane is estimated by actin hybridization, used to normalize the MT1-MMP hybridization signal.
[0061] FIG. 7 is a graph showing a fluctuating (diurnal) pattern of expression of MT1-MMP mRNA in the normal rat retina, according to an embodiment of the invention. The highest level of MT1-MMP expression occurs at 6 AM, approximately 1-2 hours before the time of maximal shedding and phagocytosis of the photoreceptor (OS) in vivo.
[0062] FIG. 8 is eight photomicrographs (phase contrast and fluorescence) showing immunofluorescent staining of normal rat retina fixed at various times of day and immunostained with an anti-MT1-MMP antibody, according to an embodiment of the invention. Diurnal variation is seen in the immunofluorescence level of MT1-MMP protein present in the OS and RPE, with the highest level of signal observed at 6 AM, less at 10 AM, and no signal at 10 PM, consistent with the diurnal pattern of MT1-MMP mRNA expression levels shown in FIG. 7.
[0063] FIG. 9 is a fluorescence micrograph of a section of human retina stained with anti-MT1-MMP antibody, showing localization of the MT1-MMP protein in the OS of rod and cone photoreceptors and in phagosomes within the RPE cells, according to an embodiment of the invention.
[0064] FIG. 10 (A-C) is three fluorescence micrographs showing the effect of anti-MT1-MMP antibody on ROS phagocytosis by RPE cells in culture, according to an embodiment of the invention. Ingestion of the fed ROS (fluorescence) is evident in the cytoplasm in control cells not incubated with antibody (B) and in cells incubated with an unrelated (X-arrestin) antibody (C), whereas ROS binding and phagocytosis does not occur in cells incubated with anti-MT1-MMP antibody prior to feeding with ROS (A).
[0065] FIG. 11 (A-D) is four micrographs of H&E stained paraffin sections of normal rat retina showing the effect of subretinal injection of anti-MT1-MMP antibody on the structure of the outer retina. Pronounced lengthening and abnormal orientation of the OS, consistent with inhibited OS phagocytosis, is observed in the anti-MT1-MMP antibody injected left eye, O.S. (A, B). In contrast, retinal architecture is normal in the uninjected right eye (O.D.) of the same animal (C). Subretinal injection of an unrelated (X-arrestin) antibody has no effect (D).
[0066] FIG. 12 shows Northern blot analysis of MT1-MMP mRNA expression levels in the RPE/choroid and retina of a subject affected with AMD (A) compared to a normal control subject (N). A 5.5-fold increase in the level of MT1-MMP mRNA is seen in the affected retina, with a 1.2-fold increase in the RPE/choroid of this subject. The Northern blot hybridization signals are normalized with respect to the amount of RNA present in each lane using actin hybridization as a reference.
[0067] FIG. 13 is a graph showing a positive correlation of level of expression of MT1-MMP mRNA with increasing severity of AMD-related pathology (grade 0-+4 changes) in retinas of subjects affected with AMD.
[0068] FIG. 14 shows the nucleic acid sequence of a 285 bp PCR product including exon 5 of human MT1-MMP. The positions of codons 259 and 273 are underlined. Bases showing changes in polymorphisms P259P and D273N found in AMD and macular degeneration patients are indicated in boldface.
[0069] FIG. 15 is two micrographs showing a delay in inherited retinal degeneration in an RCS rat injected subretinally on postnatal day 7 with an anti-MT1-MMP antibody and fixed at 30 days of age. The delay in retinal degeneration is evidenced by the greater number of photoreceptor nuclei (approximately double) remaining in the outer nuclear layer of the retina of the injected eye (A), compared to a comparable mid-central region in the uninjected control eye of the same animal (B).
[0070] FIG. 16 is a graph showing Northern blot analysis of CK1ε mRNA expression levels in samples of retina from human subjects affected with AMD of various grades of pathology and normal control retinas.
[0071] FIG. 17A-B is two graphs showing results of expression studies using real-time quantitative PCR analysis to determine levels of CK1ε expression in human eye (HE) retina (A) and RPE/choroid eyecups (EC) samples (B), according to grade of AMD pathology observed in the samples.
[0072] FIG. 18A-B is two graphs showing expression of CK1ε in the retinas (A) and RPE/choroid eyecups (B) of mice subjected to cigarette smoke for up to 6 weeks.
[0073] FIG. 19A-B is two graphs showing expression of CK1ε in the retinas (A) and RPE/choroid eyecups (B) of mice from 1-14 days following laser photocoagulation treatment to induce formation of choroidal neovascular (CNV) membranes.
[0074] FIG. 20A-B is two graphs showing diurnal expression of CK1ε at 3 hour intervals during the 24 hour Light:Dark cycle, in the retinas (A) and RPE/choroid eyecups (B) of normal control mice and mice exposed to cigarette smoke for 3 weeks.
[0075] FIG. 21 is a schematic diagram showing a pTRE-TBI-mCk1ε vector used in the construction of an inducible CK1ε over-expression transgenic mouse model, according to an embodiment of the invention.
[0076] FIG. 22A-F is a series of six photomicrographs showing histological features of AMD-like pathology observed in an inducible MT1-MMP over-expression transgenic mouse model, according to an embodiment of the invention. See specification for details.
[0077] FIG. 23A-C is a series of fluorescence photomicrographs showing the development of CNV lesions in the mouse laser photocoagulation model of CNV formation, in control mice at a few hours, and at 14 days after laser burn (A, B), and at 14 days after laser burn+treatment with an anti-MT1-MMP or anti-VEGF antibody.
DETAILED DESCRIPTION OF THE INVENTION
[0078] Based on the foregoing discoveries, the invention provides novel genes related to AMD and/or phagocytosis by RPE cells, methods and compositions for detecting and treating AMD and other retinal degenerative conditions, and animal models based on phagocytosis-related and/or AMDP-related genes useful, inter alia, for testing therapeutic compounds and treatment protocols for AMD. The below described preferred embodiments illustrate adaptations of these compositions and methods. Nonetheless, from the description of these embodiments, other aspects of the invention can be made and/or practiced based on the description provided below.
Biological Methods
[0079] Methods involving conventional molecular biology techniques are described herein. Such techniques are generally known in the art and are described in detail in methodology treatises such as Molecular Cloning: A Laboratory Manual, 2nd ed., vol. 1-3, ed. Sambrook et al., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1989; and Current Protocols in Molecular Biology, ed. Ausubel et al., Greene Publishing and Wiley-Interscience, New York, 1992 (with periodic updates). Various techniques using polymerase chain reaction (PCR) are described, for example, in Innis et al., PCR Protocols: A Guide to Methods and Applications, Academic Press: San Diego, 1990. Methods for chemical synthesis of nucleic acids are discussed, for example, in Beaucage and Carruthers, Tetra. Letts. 22:1859-1862, 1981, and Matteucci et al., J. Am. Chem. Soc. 103:3185, 1981. Chemical synthesis of nucleic acids can be performed, for example, on commercial automated oligonucleotide synthesizers. Immunological methods (for example, preparation of antigen-specific antibodies, immunoprecipitation, and immunoblotting) are described, for example, in Current Protocols in Immunology, ed. Coligan et al., John Wiley & Sons, New York, 1991; and Methods of Immunological Analysis, ed. Masseyeff et al., John Wiley & Sons, New York, 1992. Conventional methods of gene transfer and gene therapy can also be adapted for use in the present invention. See, for example, Gene Therapy: Principles and Applications, ed. T. Blackenstein, Springer Verlag, 1999; Gene Therapy Protocols (Methods in Molecular Medicine), ed. P. D. Robbins, Humana Press, 1997; and Retro-vectors for Human Gene Therapy, ed. C. P. Hodgson, Springer Verlag, 1996.
Phagocytosis-Related Genes Isolated by CHANGE
[0080] Studies leading to the invention were performed to identify genes involved in OS phagocytosis by RPE cells that, when perturbed, could result in stress and dysfunction in the RPE. Such stresses could lead to one or more undesirable changes associated with macular, retinal or choroidal diseases, such as enhanced lipofuscin accumulation, drusen formation, or formation of neovascular membranes. The gene discoveries described herein were based on the premise that dysfunction in phagocytosis by the RPE is a key factor leading to such AMD-related changes. RPE cells perform the crucial function of sustaining the homeostasis of the photoreceptors. This demanding task includes inter alia a daily process of phagocytosis and digestion of OS membranes which are renewed and shed daily from the tips of the OS of the photoreceptors (Young and Bok, 1969). As further described below, the phagocytic process includes the steps of binding, ingestion and digestion of OS membranes. Under normal circumstances, RPE cells are non-dividing cells. Thus, throughout the lifetime of an individual, the daily process of OS phagocytosis represents not only an enormous metabolic load on these cells, but also contributes to the accumulation within these cells of undigested material, particularly lipofuscin, a complex amalgam of cellular waste products including toxic photoreceptor-derived materials such as A2E.
[0081] Accordingly, in one aspect, the invention provides nucleic acid and protein sequences of genes previously unknown to be functionally related to the process of phagocytosis by RPE cells. Prior to the invention, there had not been a systematic search for genes involved in the mechanism of OS phagocytosis by RPE cells, herein also designated "phagocytosis-related genes," or "phagogenes," abbreviated to "PHG." Consistent with the knowledge that AMD is a complex, multi-gene disease, and that RPE phagocytosis is a multi-step cellular process necessarily involving many different gene products, the inventors sought to identify phagocytosis-related genes based on the realization that subtle changes, such as polymorphisms, in the DNA sequences of one or more phagocytosis-related genes, or a polymorphism in a phagocytosis-related gene in combination with a polymorphism in another gene, are likely to cooperate to produce the phenotype observed in AMD.
[0082] To obtain genes of interest by differential expression, as further described in the examples below, a custom expression profiling strategy was developed, termed CHANGE (for Comparative Hybridization ANalysis of Gene Expression). The CHANGE array included approximately 10,000 genes expressed in the RPE, arrayed in panels each comprising 96 cDNAs. (See FIG. 1.) To obtain phagogenes, the CHANGE array of RPE-expressed genes was screened with pairs of "+/- OS" hybridization probes made from total RNA expressed in a phagocytic RPE cell line during OS phagocytosis in vitro (+ OS probe) and in control cells without feeding of OS (- OS probe). Genes in the array were selected for further analysis based upon a showing of altered (i.e., increased or decreased) expression during OS phagocytosis, evidenced by a changed hybridization signal upon hybridization with the + OS vs. - OS probes, as indicated by arrows in FIG. 1. Of the approximately 10,000 genes screened, about 60 putative phagocytosis-related genes were identified on the basis of altered gene expression detected by CHANGE. Of these, 16 genes demonstrating very pronounced change in hybridization intensity upon phagocytic challenge (i.e., screening with +/- OS probes) were randomly selected for further study and confirmation of their functional relationship to RPE phagocytosis. Table 1 provides a listing of the above-described phagogenes with subsequently confirmed association with OS phagocytosis by RPE cells. These genes are further described in Example 2, infra. See also FIG. 4 showing mRNA expression profiles of these genes during phagocytosis of OS by RPE cells in vitro.
TABLE-US-00001 TABLE 1 Human Phagocytosis-related Genes Isolated by CHANGE NUCLEIC AMINOACID CLONE ACID SEQ SEQ ID NAME NUMBER ID NO. NO(S) IDENTITY PHG-1 6-29 1 71-79 Unknown PHG-2 33-25 2 80 Prostaglandin D2 synthase PHG-3 33-74 3 81 Myelin basic protein PHG-4 43-16 4 82-84 Unknown PHG-5 45-88 5 85 Unknown PHG-6 53-7 6 86 Peanut-like 2/septin 4 PHG-7 55-26 7 87 Coactosin- like 1 PHG-8 55-28 8 88 Clusterin PHG-9 57-29 9 89 Casein kinase 1 epsilon PHG-10 57-29 9 89 Casein kinase 1 epsilon (duplicate) PHG-11 73-51 10 90 Ferritin heavy polypeptide 1 PHG-12 74-39 11 91 Metargidin PHG-13 78-70a 12 92-98 Unknown PHG-14 78-70c 13 99 Retinaldehyde binding protein 1 PHG-15 80-31 14 100 Actin gamma 1 PHG-16 91-40 15 101 Matrix metallo- proteinase, membrane- associated 1 (MT1-MMP)
AMDP-Related Genes Isolated by CHANGE
[0083] In another aspect, the invention provides nucleic acid and protein sequences of genes previously unknown to be associated with AMD. To obtain AMD-related genes, the CHANGE array of 10,000 RPE-expressed genes was iteratively screened, as described above, using other pairs of "+/-" probes. The +/- probes used to identify AMD-related genes were made from total RNA extracted from the RPE/choroid of AMD-affected and unaffected human donor eyes, and from age-matched normal and affected eyes from a monkey model of AMD. Genes in the array were selected for further analysis based upon a showing of differential (i.e., increased or decreased) expression in AMD relative to aged normal control eyes. Based on the criterion of altered gene expression detected by CHANGE, approximately 200 AMD-related genes were identified.
[0084] To identify AMD-related phagogenes ("AMDP genes"), the data from the above-described two CHANGE screenings were compared, to identify a subset of RPE genes differentially expressed both in OS phagocytosis by RPE cells and in AMD. As described above, the phagocytosis CHANGE screening yielded approximately 60 phagogenes and the putative AMD-related genes numbered approximately 200. Initial comparison of the two databases yielded a subset of 6 genes showing changed expression in both phagocytosis and AMD (Table 2). These genes are herein designated "AMD-related phagogenes" or "AMD/phagogenes," abbreviated to "AMDP."
TABLE-US-00002 TABLE 2 AMD-Related Phagogenes ("AMDP" Genes) Isolated by Iterative CHANGE Analysis NUCLEIC AMINOACID CLONE ACID SEQ SEQ ID NAME NUMBER ID NO. NO(S) IDENTITY AMDP-1 33-25 2 80 Prostaglandin D2 synthase AMDP-2 37-14 16 102 SWI/SNF related/ OSA-1 nuclear protein AMDP-3 47-94 17 103-121 Unknown AMDP-4 57-29 9 89 Casein kinase 1 epsilon AMDP-5 73-51 10 90 Ferritin heavy polypeptide 1 AMDP-6 91-40 15 101 Matrix metallo- proteinase, membrane associated 1 (MT1-MMP)
[0085] Of the above listed genes, the CHANGE hybridization analysis indicated that mRNAs for genes AMDP-1,3,4, and 6 were expressed at higher levels in AMD eyes than in controls, whereas the expression levels of genes AMDP-2 and AMDP-5 were lower in AMD eyes than in controls. AMDP genes are further described in Example 3, infra.
Nucleic Acids Encoding Phagocytosis-Related and/or AMDP-Related Gene Products and Polymorphic Variants Thereof
[0086] As described above, the invention provides nucleic acid and amino acid sequences relating to genes discovered by a differential cloning strategy (CHANGE) to exhibit altered expression during RPE phagocytosis and/or in AMD. In one aspect, the invention provides novel purified nucleic acids (polynucleotides) isolated by this strategy. Previously unknown nucleic acids of the invention include nucleic acid sequences identified herein as PHG-1 (SEQ ID NO:1); PHG-4 (SEQ ID NO. 4); PHG-5 (SEQ ID NO: 5); PHG-13 (SEQ ID NO:12); and AMDP-3 (SEQ ID NO:17). These nucleic acids encode, respectively, polypeptides having the amino acid sequences identified herein as SEQ ID NOS:71-79; 82-84; 85; 92-98; and 103-121.
[0087] The invention also encompasses use of characterized nucleic acids and polypeptides previously unknown to be related to RPE phagocytosis and/or AMD. The relationship of the previously characterized genes to phagocytosis and AMD was discovered on the basis of changed expression during RPE phagocytosis and/or in AMD patients. Nucleic acids of the latter group include prostaglandin D2 synthase (SEQ ID NO:2), myelin basic protein (SEQ ID NO:3), peanut-like 2/septin 4 (SEQ ID NO:6); coactosin-like 1 (SEQ ID NO:7); clusterin (SEQ ID NO:8); casein kinase 1 epsilon (SEQ ID NO:9); ferritin heavy polypeptide 1 (SEQ ID NO:10); metargidin (SEQ ID NO:11); retinaldehyde binding protein 1 (SEQ ID NO:13); actin gamma 1 (SEQ ID NO:14); matrix metalloproteinase, membrane associated 1 (SEQ ID NO: 15); and SWI/SNF related/OSA-1 nuclear protein (SEQ ID NO:16).
[0088] Nucleic acid molecules of the present invention can be in the form of RNA or in the form of DNA (for example, cDNA, genomic DNA, and synthetic DNA). Preferred nucleic acid molecules of the invention are the respective native polynucleotides, including the nucleotide sequences shown herein as SEQ ID NOS:1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16 and 17.
[0089] The coding sequences which encode native phagocytosis-related and/or AMDP-related genes may be identical to the those of nucleotide sequences shown in SEQ ID NOS:1-17. They may also be different coding sequences which, as a result of the redundancy or degeneracy of the genetic code, encode the same polypeptides as the polynucleotides of SEQ ID NOS:1-17. Other nucleic acid molecules within the invention are variants of SEQ ID NOS:1-17 such as those that encode fragments, analogs and derivatives of the phagocytosis-related and AMDP-related genes described herein. Such variants may be, for example, naturally occurring allelic variants of native phagocytosis-related and AMDP-related genes, homologs of native phagocytosis-related and/or AMDP-related genes, splice variants, or non-naturally occurring variants of phagocytosis-related and/or AMDP-related genes. These variants have a nucleotide sequence that differs from the corresponding native SEQ ID NOS:1-17 in one or more bases. For example, the nucleotide sequence of such variants can feature a deletion, addition, or substitution of one or more nucleotides of native phagocytosis-related and/or AMDP-related genes.
[0090] In some applications, variant nucleic acid molecules encode polypeptides that substantially maintain a phagocytosis-related and/or AMDP-related functional activity. For other applications, variant nucleic acid molecules encode polypeptides that lack or feature a significant reduction in a phagocytosis-related and/or AMDP-related gene functional activity. Where it is desired to retain a functional activity of a native phagocytosis-related and/or AMDP-related gene, preferred variant nucleic acids feature silent or conservative nucleotide changes.
[0091] In other applications, variant phagocytosis-related and/or AMDP-related polypeptides displaying substantial changes in one or more functional activities of native phagocytosis-related and/or AMDP-related genes can be generated by making nucleotide substitutions that cause less than conservative changes in the encoded polypeptide. Examples of such nucleotide substitutions are those that cause changes in (a) the structure of the polypeptide backbone; (b) the charge or hydrophobicity of the polypeptide; or (c) the bulk of an amino acid side chain. Nucleotide substitutions generally expected to produce the greatest changes in protein properties are those that cause non-conservative changes in codons. Examples of codon changes that are likely to cause major changes in protein structure are those that cause substitution of (a) a hydrophilic residue, for example, serine or threonine, by a hydrophobic residue, for example, leucine, isoleucine, phenylalanine, valine or alanine; (b) a cysteine or proline by any other residue; (c) a residue having an electropositive side chain, for example, lysine, arginine, or histidine, by an electronegative residue, for example, glutamine or asparagine; or (d) a residue having a bulky side chain, for example, phenylalanine, by one not having a side chain, for example, glycine.
[0092] Naturally occurring allelic variants of native phagocytosis-related and/or AMDP-related genes within the invention are nucleic acids that have at least 75% (for example, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, and 99%) sequence identity with native phagocytosis-related and/or AMDP-related genes, and encode polypeptides having at least one functional activity in common with native phagocytosis-related and/or AMDP-related genes. Homologs of native phagocytosis-related and/or AMDP-related genes within the invention are nucleic acids isolated from non-human species that have at least 75% (for example, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, and 99%) sequence identity with native phagocytosis-related and/or AMDP-related genes, and encode polypeptides having at least one functional activity in common with native phagocytosis-related and/or AMDP-related genes.
[0093] Naturally occurring allelic variants of phagocytosis-related and/or AMDP-related genes and homologs of phagocytosis-related and/or AMDP-related genes can be isolated by screening for a native functional activity of a phagocytosis-related and/or AMDP-related gene (for example, activation of progelatinase A, in the case MT1-MMP) using techniques known in the art. The nucleotide sequence of such homologs and allelic variants can be determined by conventional DNA sequencing methods. Alternatively, public or non-proprietary nucleic acid databases can be searched to identify other nucleic acid molecules having a high percent (for example, 70, 80, 90%, 95% or more) sequence identity to a native phagocytosis-related and/or AMDP-related gene.
[0094] Non-naturally occurring variants of phagocytosis-related and/or AMDP-related genes are nucleic acids that do not occur in nature (for example, are made by the hand of man), have at least 75% (for example, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, and 99%) sequence identity with native phagocytosis-related and/or AMDP-related genes and encode polypeptides having at least one functional activity in common with native phagocytosis-related and/or AMDP-related genes. Examples of non-naturally occurring phagocytosis-related and/or AMDP-related nucleic acids are those that encode a fragment of a phagocytosis-related and/or AMDP-related protein, those that hybridize to a native phagocytosis-related and/or AMDP-related gene, or a complement of a native phagocytosis-related and/or AMDP-related genes under stringent conditions, those that share at least 65% sequence identity with a native phagocytosis-related and/or AMDP-related gene, or a complement of a native phagocytosis-related and/or AMDP-related gene, and those that encode a phagocytosis-related and/or AMDP-related gene fusion protein.
[0095] Nucleic acids encoding fragments of phagocytosis-related and/or AMDP-related genes within the invention are those that encode, for example, 2, 5, 10, 25, 50, 100, 150, 200, 250, 300, or more amino acid residues of the respective phagocytosis-related and/or AMDP-related proteins. Shorter oligonucleotides (for example, those of 6, 12, 20, 30, 50, 100, 125, 150 or 200 bases in length) that encode or hybridize with nucleic acids that encode fragments of phagocytosis-related and/or AMDP-related genes can be used as probes, primers, or antisense molecules. Longer polynucleotides (for example, those of 300, 400, 500, 600, 700, 800, 900, 1000, 1100, 1200, 1300, 1400, 1500, 1600, 1700, 1800, 1900, 2000, 2100, 2200, 2300, 2400, 2500, 2600, 2700, 2800, 2900, 3000 or more bases, such as 4000, 5000, 6000, 7000, 8000, and 9000 bases) that encode or hybridize with nucleic acids that encode fragments of phagocytosis-related and/or AMDP-related genes can be used in place of native phagocytosis-related and/or AMDP-related genes in applications where it is desired to modulate a functional activity of native phagocytosis-related and/or AMDP-related gene. Nucleic acids encoding fragments of phagocytosis-related and/or AMDP-related genes can be made by enzymatic digestion (for example, using a restriction enzyme) or chemical degradation of full length sequences of phagocytosis-related and/or AMDP-related genes, or variants thereof.
[0096] Nucleic acids that hybridize under stringent conditions to the nucleic acid of SEQ ID NOS:1, 4, 5, 12 and 17 or the complement of SEQ ID NOS:1, 4, 5, 12 and 17 are also within the invention. For example, such nucleic acids can be those that hybridize to SEQ ID NOS:1, 4, 5, 12 and 17 or the complement of SEQ ID NOS:1, 4, 5, 12 and 17 under low stringency conditions, moderate stringency conditions, or high stringency conditions. Preferred such nucleic acids are those having a nucleotide sequence that is the complement of all or a portion of SEQ ID NOS:1, 4, 5, 12 or 17. Other variants of SEQ ID NOS:1, 4, 5, 12 and 17 within the invention are polynucleotides that share at least 65% (for example, 65, 70, 75, 80, 85, 90, 91, 92, 93, 94, 95, 96, 97, 98, and 99%) sequence identity to SEQ ID NOS:1, 4, 5, 12 and 17 or the complement of SEQ ID NOS:1, 4, 5, 12 and 17. Nucleic acids that hybridize under stringent conditions or share at least 65% sequence identity with SEQ ID NOS:1, 4, 5, 12 and 17 or the complement of SEQ ID NOS:1, 4, 5, 12 and 17 can be obtained by techniques known in the art.
[0097] Nucleic acid molecules encoding fusion proteins of phagocytosis-related and/or AMDP-related genes, for example those encoded by nucleic acids described herein as SEQ ID NOS:1-17, are also within the invention. Such nucleic acids can be made by preparing a construct (for example, an expression vector) that expresses a phagocytosis-related and/or AMDP-related fusion protein when introduced into a suitable host. For example, such a construct can be made by ligating a first polynucleotide encoding a phagocytosis-related and/or AMDP-related protein, for example MT1-MMP, fused in frame with a second polynucleotide encoding another protein such that expression of the construct in a suitable expression system yields a fusion protein.
[0098] The invention encompasses labeled nucleic acid probes capable of hybridizing to a nucleic acid encoding a phagocytosis-related and/or AMDP-related polypeptide, as described above. The nucleic acid molecules of the invention allow those skilled in the art to construct nucleotide probes for use in the detection of nucleic acid sequences of the invention in biological materials. The probe may be used in hybridization to detect a phagocytosis-related and/or AMDP-related gene. The technique generally involves contacting and incubating nucleic acids (for example mRNA molecules) obtained from a sample from a patient or other cellular source with a probe of the present invention under conditions favorable for the specific annealing of the probes to complementary sequences in the nucleic acids. After incubation, the non-annealed nucleic acids are removed, and the presence of nucleic acids that have hybridized to the probe, if any, are detected.
[0099] The detection of nucleic acid molecules of the invention may involve the amplification of specific gene sequences using an amplification method (for example PCR), followed by the analysis of the amplified molecules using techniques known to those skilled in the art. Suitable primers can be routinely designed by one of skill in the art. For example, primers may be designed using commercially available software, such as OLIGO 4.06 Primer Analysis software (National Biosciences, Plymouth Minn.) or another appropriate program, to be about 22 to 30 nucleotides in length, to have a GC content of about 50% or more, and to anneal to the template at temperatures of about 60° C. to 72° C.
[0100] Hybridization and amplification techniques described herein may be used to assay qualitative and quantitative aspects of phagocytosis-related and/or AMDP-related gene expression. For example, RNA may be isolated from a cell type or tissue known to express a phagocytosis-related and/or AMDP-related gene, for example genes having SEQ ID NOS:1-17, and tested utilizing the hybridization (for example, standard Northern analyses) or PCR techniques referred to herein. The techniques may be used, for example, to detect differences in transcript size that may be due to normal or abnormal alternative splicing. The techniques may be used to detect quantitative differences between levels of full length and/or alternatively spliced transcripts detected in normal individuals relative to those individuals exhibiting symptoms of a disease. The primers and probes may be used in the above-described methods in situ, i.e., directly on tissue sections (fixed and/or frozen) of patient tissue obtained from biopsies, resections or eyebank eyes. Particular uses of the probes and primers of the invention are further described in the examples below.
Genetic Screening of Phagocytosis-Related and/or AMD-Related Nucleic Acids
[0101] In another aspect, the invention provides a method for determining the risk of a subject of developing a retinal or choroidal disease or degenerative condition. As used herein, a "retinal or choroidal disease or degenerative condition" includes but is not limited to any condition of the retina or choroid of the eye which results in injury or death of photoreceptors, RPE cells or other cell types of the retina, or injury, death or abnormal proliferation of choroidal cell types including but not limited to endothelial cells, melanocytes, smooth muscle cells, fibroblasts, lymphocytes, neutrophils, eosinophils, megokaryocytes, monocytes, macrophages and mast cells.
[0102] Degenerative conditions affecting the retina and/or choroid include age-related and other maculopathies, including but not limited to age-related macular degeneration (AMD), hereditary and early onset forms of macular degeneration ("familial AMD") such as Stargardt's disease/fundus flavimaculatus, Best disease/vitelliform dystrophy, congenital diffuse drusen/Doyne's honeycomb dystrophy, pattern dystrophies, Sorsby's macular dystrophy, juxtafoveal telangiectasia, choroidal atrophy, dominant drusen, crystalline drusen, annular macular dystrophy, occult choroidal neovascular membrane, choroideremia, idiopathic bulls-eye maculopathies, gyrate atrophy and the various forms of hereditary retinitis pigmentosa conditions. Other diseases or degenerative conditions of the retina and choroid include toxic maculopathies, for example, drug-induced maculopathies such as plaquenil toxicity, retinal disorders including retinal detachment, photic retinopathies, retinopathies induced by surgery, toxic retinopathies, retinopathy of prematurity, viral retinopathies such as CMV or HIV retinopathy related to AIDS, uveitis, ischemic retinopathies due to venous or arterial occlusion or other vascular disorders, retinopathies due to trauma or penetrating lesions of the eye, peripheral vitreoretinopathy, and cancers affecting the eye such as retinoblastoma and choriodal melanoma.
[0103] The method for determining risk involves screening a nucleic acid of a subject for the presence of polymorphisms in AMD-related or phagocytosis-related genes, wherein the presence of a polymorphism indicates that the subject is at higher risk for developing a retinal or choroidal disease or degenerative disorder than a control subject without the polymorphism. As used herein, a "normal" or "wild type" nucleotide is a base located at a particular position in a subject's DNA that is known to be the predominant base at that position in the general population. A "polymorphism," "polymorphic variant," or "polymorphic base or nucleotide," is a naturally occurring base change that occurs at lower frequency in the general population than the base representing the "wild type." A "polymorphism" as used herein can include a base change recognized as a "mutation."
[0104] A phagocytosis-related and/or AMDP-related nucleic acid of the invention, either alone or in combination with one or more other nucleic acids, may be used in hybridization, amplification and screening assays of biological samples to detect abnormalities, including point mutations, insertions, deletions, and chromosomal rearrangements. Genetic screening methods are well known in the art of molecular medicine. For example, using genomic DNA, direct sequencing, single stranded conformational polymorphism analyses, heteroduplex analysis, denaturing gradient gel electrophoresis, chemical mismatch cleavage, and oligonucleotide hybridization (including hybridization to oligonucleotides in a gene array) may be utilized. In general, a genomic DNA sample is obtained from a subject, for example from the subject's peripheral blood, or from a biological sample prepared from donated tissue such as an eyebank eye. The DNA is used for amplification of specific gene sequences, for example a particular exonic, intronic or promoter sequence of interest. To detect the presence of polymorphisms in a subject's DNA, single strand conformation polymorphism (SSCP) analysis, heteroduplex analysis, and automated versions thereof can be used, followed by DNA sequence analysis to determine the particular base change(s). These methods are also useful for confirming reported polymorphisms, for example those available in the Human Genome Single Nucleotide Polymorphism (SNP) database.
[0105] The invention provides methods for screening a subject for polymorphic variants of genes related to RPE phagocytosis and/or AMD. In one preferred method, pairs of sense and antisense primers (amplimers) are designed based on the nucleic acid sequence of a gene of interest and are used to amplify one or more exons, introns or promoter sequences within the gene. One preferred group of genes useful for screening for mutations and polymorphisms in patients with AMD and other macular diseases includes previously unknown genes shown herein to be correlated with phagocytosis and/or AMD, the cDNA sequences of which are identified herein as SEQ ID NOS:1, 4, 5, 12, and 17. Other preferred genes, also disclosed herein to be related to phagocytosis and/or AMD, have nucleic acid (cDNA) sequences described herein as SEQ ID NOS:2, 3, 6, 7, 8, 9, 10, 11, 13, 14, 15, and 16. (See Tables 1 and 2, supra.) As shown herein, an exemplary gene related to AMD and phagocytosis is MT1-MMP (SEQ ID NO:15). Any amplimers suitable for amplifying an exonic, intronic or promoter sequence of a phagocytosis-related and/or AMDP-related genes disclosed herein can be designed by those of skill in the art of molecular biology and used to screen DNA samples for mutations and/or polymorphisms. As an example, specific amplimer pairs, suitable for amplification of Exons 1-10, introns 1-9 and promoter regions of the human MT1-MMP gene are disclosed in Table 3 below.
[0106] The nucleic acids of the invention can also be used for screening of multiple genes in an array. Oligonucleotides or longer fragments derived from any of the nucleic acid molecules of the invention may be used as targets in a gene array such as a microarray. The gene targets in the array can include, for example, nucleic acids derived from any combination of phagocytosis-related and/or AMDP-related genes disclosed herein (i.e., SEQ ID NOS: 1-17) and any previously described nucleic acids, for example those previously associated with RPE phagocytosis and/or AMD, including but not limited to those derived from sequences identified herein as SEQ ID NOS:62-69. The oligonucleotide sequences included in the array can be derived from sequences positioned within an intronic, exonic or promoter sequence of the native human gene of interest. Preferably the arrays include oligonucleotide sequences encompassing all known polymorphic variants of the genes of interest. Particularly preferred custom arrays, suitable for example for screening the DNA of patients with eye diseases such as AMD, include all known polymorphic variants of genes shown to exhibit particular polymorphic variants with increased incidence in populations of patients with AMD and related disorders, relative to control populations of normal subjects. For a listing of genes with previously reported polymorphisms or mutations correlated with AMD, see Table 3, infra. Accordingly, genes suitable for inclusion in a custom array of the invention useful for AMD screening, and the relevant polymorphic variants thereof showing increased incidence in AMD (in parentheses) can include, but are not limited to: MT1-MMP (P259P; D273N); ABCR (D217N; G1961E); manganese superoxide dismutase (V47A); apolipoprotein E (C130, R176C and C130R, R176); cystatin C (A25T) and paraoxonase (Q192R, L54M).
[0107] The gene arrays of the invention can be used, for example, to simultaneously monitor the expression levels of large numbers of genes, and to identify genetic variants, mutations, and polymorphisms in a plurality of genes. The information derived from the analysis of the hybridization of patient DNA samples to the array can be used, for example, to determine gene function, to understand the genetic basis of a disorder, to diagnose or predict the likelihood of developing a disorder, or to develop and monitor the activities of therapeutic agents. The preparation, use, and analysis of gene arrays, including microarrays are well known to persons skilled in the art. (See, for example, Brennan, T. M. et al. (1995) U.S. Pat. No. 5,474,796; Schena, et al. (1996) Proc. Natl. Acad. Sci. 93:10614-10619; Baldeschweiler et al. (1995), PCT Application WO95/251116; Shalon, D. et al. (I 995) PCT application WO95/35505; Heller, R. A. et al. (1997) Proc. Natl. Acad. Sci. 94:2150-2155; and Heller, M. J. et al. (1997) U.S. Pat. No. 5,605,662 and Cronin, M. et al. (2003) U.S. Pat. No. 6,632,605.
Agents That Modulate Expression or Activity of Phagocytosis-Related and AMDP-Related Gene Products
[0108] In another aspect, the invention provides agents that modulate expression levels of mRNA or protein of phagocytosis-related and/or AMDP-related genes. Preferred genes/proteins to be targeted for down-regulation are those showing increased expression in AMD and related disorders, including, as demonstrated herein, prostaglandin D2 synthase, PD2S (respective nucleic acid and amino acid sequences: SEQ ID NOS:2 and 80), MT1-MMP (SEQ ID NOS:15 and 101), casein kinase 1 epsilon (CK1ε) (SEQ ID NOS:9 and 89) and AMDP-3 (SEQ ID NOS:17 and 103-121). Preferred genes/proteins to be targeted for up-regulation are those showing decreased expression in AMD and related disorders, including, as demonstrated herein, SWI/SNF related OSA-1 nuclear protein (SEQ ID NOS:16 and 102) and ferritin heavy polypeptide 1 (SEQ ID NOS:10 and 101).
[0109] The AMDP-related and/or phagocytosis-related mRNA or protein can be the native, i.e., "wild-type" mRNA or protein, for example native MT1-MMP. In other embodiments, a polymorphic variant of an AMD-related or phagocytosis-related gene is targeted, for example one which results in an altered function of the expressed mRNA or protein. The altered mRNA or protein is inhibited while leaving expression of the wild type mRNA or protein intact.
[0110] The inhibitory agents used for down-regulation of expression can include, for example, antisense RNA molecules, ribozymes, small interfering RNA (RNAi) molecules and triple helix structures. Preferred embodiments of such agents are directed against PD2S (SEQ ID NO:2), CK1ε (SEQ ID NO:9), MT1-MMP (SEQ ID NO:15) and AMDP-3 (SEQ ID NO:17), or variants thereof. The inhibitory agents can also include antibody molecules that selectively bind to an over-expressed phagocytosis-related and/or AMDP-related protein, such as PD2S, MT1-MMP or AMDP-3.
[0111] Antisense nucleic acid molecules within the invention are those that specifically hybridize (for example bind) under cellular conditions to cellular mRNA and/or genomic DNA encoding a phagocytosis-related and/or AMDP-related protein in a manner that inhibits expression of the phagocytosis-related and/or AMDP-related protein, for example, by inhibiting transcription and/or translation. The binding may be by conventional base pair complementarity, or, for example, in the case of binding to DNA duplexes, through specific interactions in the major groove of the double helix. Methods for design of antisense molecules are well known to those of skill in the art. General approaches to constructing oligomers useful in antisense therapy have been reviewed, for example, by Van der Krol et al. (1988) Biotechniques 6:958-976; Stein et al. (1988) Cancer Res 48:2659-2668; and Narayanan, R. and Aktar, S. (1996): Antisense therapy. Curr. Opin. Oncol. 8(6):509-15. As non-limiting examples, antisense oligonucleotides may be targeted to hybridize to the following regions: mRNA cap region; translation initiation site; translational termination site; transcription initiation site; transcription termination site; polyadenylation signal; 3' untranslated region; 5' untranslated region; 5' coding region; mid coding region; and 3' coding region.
[0112] An antisense construct can be delivered, for example, as an expression plasmid which when transcribed in the cell produces RNA which is complementary to at least a unique portion of the cellular mRNA which encodes a phagocytosis-related and/or AMDP-related gene product. Alternatively, the antisense construct can take the form of an oligonucleotide probe generated ex vivo which, when introduced into a phagocytosis-related or AMDP-related gene expressing cell, causes selective inhibition of expression of the corresponding gene by hybridizing with an mRNA and/or genomic sequence coding for the phagocytosis-related or AMDP-related gene. Such oligonucleotide probes are preferably modified oligonucleotides that are resistant to endogenous nucleases, for example exonucleases and/or endonucleases, and are therefore stable in vivo. Exemplary nucleic acid molecules for use as antisense oligonucleotides are phosphoramidate, phosphothioate and methylphosphonate analogs of DNA (see, for example, U.S. Pat. Nos. 5,176,996; 5,264,564; and 5,256,775). With respect to antisense DNA, oligodeoxyribonucleotides derived from the translation initiation site, for example, between the -10 and +10 regions of a phagocytosis-related or AMDP-related gene encoding nucleotide sequence, are preferred.
[0113] Antisense approaches involve the design of oligonucleotides (either DNA or RNA) that are complementary to a phagocytosis-related and/or AMDP-related mRNA. The antisense oligonucleotides will bind to mRNA transcripts of the phagocytosis-related or AMDP-related gene and prevent translation. Absolute complementarity, although preferred, is not required. The ability to hybridize will depend on both the degree of complementarity and the length of the antisense nucleic acid. Generally, the longer the hybridizing nucleic acid, the more base mismatches with an RNA it may contain and still form a stable duplex (or triplex, as the case may be). One skilled in the art can ascertain a tolerable degree of mismatch by use of standard procedures to determine the melting point of the hybridized complex. Oligonucleotides that are complementary to the 5' end of the message, for example, the 5' untranslated sequence up to and including the AUG initiation codon, in general work most efficiently at inhibiting translation. However, sequences complementary to the 3' untranslated sequences of mRNAs have been shown to be effective at inhibiting translation of mRNAs as well. (See, for example, Wagner, R. (1994) Nature 372:333.) Therefore, oligonucleotides complementary to either the 5' or 3' untranslated non-coding regions of a phagocytosis-related or AMDP-related gene could be used in an antisense approach to inhibit translation of endogenous mRNA of a phagocytosis-related or AMDP-related gene. Oligonucleotides complementary to the 5' untranslated region of the mRNA should include the complement of the AUG start codon. Antisense oligonucleotides complementary to mRNA coding regions are less efficient inhibitors of translation but could be used in accordance with the invention. Whether designed to hybridize to the 5', 3' or coding region of the mRNA of a phagocytosis-related or AMDP-related gene, antisense nucleic acids should be at least six nucleotides in length, and are preferably less than about 100 and more preferably less than about 50, 25, 17 or 10 nucleotides in length.
[0114] Regardless of the choice of target sequence, it is preferred that in vitro studies are first performed to quantitate the ability of the antisense oligonucleotide to inhibit gene expression. It is preferred that these studies utilize controls that distinguish between antisense gene inhibition and nonspecific biological effects of oligonucleotides. It is preferred that the control oligonucleotide is of approximately the same length as the test oligonucleotide, and that the nucleotide sequence of the control oligonucleotide differs from that of the antisense sequence by no more than is necessary to prevent specific hybridization to the target sequence. It is also preferred that these studies compare levels of the target RNA or protein with that of an internal control RNA or protein.
[0115] Antisense oligonucleotides of the invention may comprise at least one modified base moiety which is selected from the group including but not limited to 5-fluorouracil, 5-bromouracil, 5-chlorouracil, 5-iodouracil, hypoxanthine, xanthine, 4-acetylcytosine, 5-(carboxyhydroxyethyl) uracil, 5-carboxymethylaminomethyl-2-thiouridine, 5-carboxymethylaminomethyluracil, dihydrouricil, beta-D-galactosylqueosine, inosine, N6-isopentenyladenine, 1-methylguanine, 1-methylinosine, 2,2-idimethylguanine, 2-methyladenine, 2-methylguanine, 3-methylcytosine, 5-methylcytosine, N6-adenine, 7-methylguanine, 5-methylaminomethyluracil, 5-methoxyaminomethyl-2-thiouracil, beta-D-mannosylqueosine, 5'-methoxycarboxymethyluracil, 5-methoxyuracil, 2-methylthio-N6-isopentenyladenine, uracil-5-oxyacetic acid (v), wybutoxosine, pseudouracil, queosine, 2-thiocytosine, 5-methyl-2-thiouracil, 2-thiouracil, 4-thiouracil, 5-methyluracil, uracil-5-oxyacetic acid methylester, uracil-5-oxyacetic acid (v), 5-methyl-2-thiouracil, 3-(3-amino-3-N2-carboxypropyl)uracil, (acp3)w, and 2,6-diaminopurine. Antisense oligonucleotides of the invention may also comprise at least one modified sugar moiety selected from the group including but not limited to arabinose, 2-fluoroarabinose, xylulose, and hexose, and may additionally include at least one modified phosphate backbone selected from the group consisting of a phosphorothioate, a phosphorodithioate, a phosphoramidothioate, a phosphoramidate, a phosphordiamidate, a methylphosphonate, an alkyl phosphotriester, and a formacetal or analog thereof.
[0116] Oligonucleotides of the invention may be synthesized by standard methods known in the art, for example, by use of an automated DNA synthesizer. As examples, phosphorothioate oligonucleotides may be synthesized by the method of Stein et al. (1988) Nucl. Acids Res. 16:3209), and methylphosphonate oligonucleotides can be prepared, for example, by use of controlled pore glass polymer supports (Sarin et al. (1988) Proc. Natl. Acad. Sci. U.S.A. 85:7448-7451).
[0117] The antisense molecules can be delivered into cells that express phagocytosis-related or AMDP-related genes in vivo. A number of methods have been developed for delivering antisense DNA or RNA into cells and are well known in the art. Because it is often difficult to achieve intracellular concentrations of the antisense sufficient to suppress translation of endogenous mRNAs, a preferred approach utilizes a recombinant DNA construct in which the antisense oligonucleotide is placed under the control of a strong promoter. The use of such a construct to transfect target cells in a subject preferably will result in the transcription of single-stranded RNAs that will hybridize with endogenous transcripts encoding the gene products of interest in sufficient amounts to prevent translation of the respective mRNAs. For example, a vector can be introduced in vivo such that it is taken up by a cell and directs the transcription of an antisense RNA. Such a vector can remain episomal or can become chromosomally integrated, as long as it can be transcribed to produce the desired antisense RNA. Such vectors can be constructed by recombinant DNA technology methods standard in the art and are further described below. Vectors can be plasmid, viral, or others known in the art, used for replication and expression in mammalian cells.
[0118] Expression of the sequence encoding the antisense RNA can be by any promoter known in the art to act in mammalian, and preferably human cells. Such promoters can be inducible or constitutive. Such promoters can include, but are not limited to: the SV40 early promoter region (Bernoist and Chambon, 1981, Nature 290:304-310), the promoter contained in the 3' long terminal repeat of Rous sarcoma virus (Yamamoto et al., 1980, Cell 22:787-797), the herpes thymidine kinase promoter (Wagner et al., 1981, Proc. Natl. Acad. Sci. U.S.A. 78:1441-1445), and the regulatory sequences of the metallothionein gene (Brinster et al., 1982, Nature 296:39-42). Promoters useful for tissue- or cell-specific expression, for example in photoreceptors, RPE cells, or choroidal cell types such as endothelial cells or melanocytes, are also known in the art, and are further described in Example 7 below.
[0119] A ribozyme is another preferred embodiment of an agent that can down-regulate expression of a phagocytosis-related and/or AMDP-related gene product. Ribozyme molecules are designed to catalytically cleave a transcript of a gene of interest, preventing its translation into a polypeptide. (See, for example, Sarver et al. (1990) Science 247:1222-1225 and U.S. Pat. No. 5,093,246). In general, ribozymes catalyze site-specific cleavage or ligation of phosphodiester bonds in RNA. While various forms of ribozymes that cleave mRNA at site-specific recognition sequences can be used to destroy phagocytosis-related or AMDP-related mRNAs, the use of hammerhead ribozymes is preferred. Hammerhead and hairpin ribozymes are RNA molecules that act by base pairing with complementary RNA target sequences, and carrying out cleavage reactions at particular sites. In the case of the hammerhead, the ribozyme cleaves after UX dinucleotides, where X can be any ribonucleotide except guanosine, although the rate of cleavage is highest if X is cytosine. The catalytic efficiency is further affected by the nucleotide preceding the uridine. In practice, NUX triplets (typically GUC, CUC or UUC) are required in the target mRNA. Such targets are used to design an antisense RNA of approximately 12 or 13 nucleotides surrounding that site, but skipping the C, which does not form a conventional base pair with the ribozyme.
[0120] Synthetic hammerhead ribozymes can be engineered to selectively bind and cleave a complementary mRNA molecule, then release the fragments, repeating the process with the efficiency of a protein enzyme. This can represent a significant advantage over, for example, antisense oligonucleotides which are not catalytic, but rather are stoichiometric, forming a 1:1 complex with target sequences. The hammerhead ribozymes of the invention can be designed in a 6-4-5 stem-loop-stem configuration, or any other configuration suitable for the purpose. In general, because the chemical cleavage step is rapid and the release step is rate-limiting, speed and specificity are enhanced if the hybridizing "arms" of the ribozyme (helices I and III) are relatively short, for example, about 5 or 6 nucleotides. Suitability of the design of a particular configuration can be determined empirically, using various assays known to those of skill in the art.
[0121] The construction and production of hammerhead ribozymes is well known in the art and is described more fully, for example, in Haseloff and Gerlach (1988) Nature 334:585-591. There are numerous potential hammerhead ribozyme cleavage sites within the nucleotide sequences of native phagocytosis-related or AMDP-related genes, for example, those encoded by SEQ ID NOS:1-17. Preferably the ribozyme is engineered so that the cleavage recognition site is located near the 5' end of the phagocytosis-related or AMDP-related mRNA, in order to increase efficiency and minimize the intracellular accumulation of non-functional mRNA transcripts. Ribozymes within the invention can be delivered to a cell using a vector as described below.
[0122] The ribozymes of the present invention also include RNA endoribonucleases (hereinafter "Cech-type ribozymes") such as the one which occurs naturally in Tetrahymena thermophila (known as the IVS, or L-19 IVS RNA) and which has been extensively described by Thomas Cech and collaborators (see, for example, Zaug et al., (1984), Science, 224:574-578; Been and Cech, (1986), Cell, 47:207-216). The Cech-type ribozymes have an eight base pair active site which hybridizes to a target RNA sequence whereafter cleavage of the target RNA takes place. The invention encompasses those Cech-type ribozymes which target eight base-pair active site sequences that are present in the mRNAs specific for the peptides and proteins of interest of the current invention.
[0123] Yet another preferred agent within the invention is an RNA-mediated interference (RNAi) molecule that down-regulates expression of a phagocytosis-related and/or AMDP-related gene. The RNAi mechanism involves the use of double-stranded RNA (dsRNA) to trigger the silencing of genes highly homologous in sequence to the dsRNA. RNAi is an evolutionarily conserved phenomenon common to such diverse organisms as plants, nematodes (Caenorhabditis elegans), fruit flies (Drosophila), amphibians, and mammals. It is thought to have evolved to protect the genome against invasion by mobile genetic elements such as transposons and viruses. In a multistep process, active small interfering RNA (siRNA) molecules are generated in vivo through the action of an RNase III endonuclease, termed Dicer. The resulting 21- to 23-nucleotide siRNA molecules mediate degradation of the complementary homologous RNA (Zamore et al., 2000; Grishok et al., 2000).
[0124] Non-naturally occurring RNAi molecules can be synthesized by methods known in the art and used advantageously to silence the expression of genes of interest. In mammalian cells, dsRNAs longer than 30 nucleotides are known to activate an antiviral response, leading to the nonspecific degradation of RNA transcripts and a general shutdown of host cell protein translation. However, gene-specific suppression in mammalian cells can be achieved by in vitro-synthesized siRNAs that are about 21 nucleotides in length, these molecules being long enough to induce gene-specific suppression, but short enough to evade the host interferon response (Elbashir, S. M. et al., 2001). Those of skill in the art will recognize that computer programs are available for the design of RNAi molecules directed against specific mRNA target sequences.
[0125] Small inhibitory RNA molecules act by binding to a protein complex within the cell, termed an RNA-induced silencing complex (RISC), which contains a helicase activity and an endonuclease activity. The helicase activity unwinds the two strands of RNA molecules, allowing the antisense strand of the siRNA to bind to the targeted RNA molecule (Zamore, 2002; Vickers et al., 2003). The endonuclease activity hydrolyzes the target RNA at the site where the antisense strand is bound.
[0126] RNAi strategies can be successfully combined with vector-based approaches to achieve synthesis in transfected cells of small RNAs from a DNA template under the control, for example, of an RNA polymerase III (Pol III) promoter. Use of Pol III provides the advantage of directing the synthesis of small, non-coding transcripts whose 3' ends are defined by termination within a stretch of 4-5 thymidines (Ts). These properties make it possible to use DNA templates to synthesize, in vivo, small RNAs with structural features close to those found to be required for active siRNAs synthesized in vitro. Using such templates, small RNAs targeting selected mRNAs of interest have been expressed in transfected cells, and shown to be able to efficiently and specifically inhibit the synthesis of the corresponding proteins (Sui et al., 2002).
[0127] For suppression of dominant gain-of-function mutations, or undesirable polymorphic variants of mRNAs of phagocytosis-related and/or AMDP-related genes which may differ from the wild type sequences by only a single base change (for example one of the AMD-associated variants of MT1-MMP, described herein), it may be desirable to selectively silence expression of the abnormal mRNA while permitting expression of the normal allele. A highly advantageous feature of the RNAi technology is the ability to selectively silence a mutation with single-nucleotide specificity. The feasibility of this approach has been demonstrated using RNAi to suppress the expression of a mutant allele of the Cu, Zn superoxide dismutase (SOD1) gene causing amyotrophic lateral sclerosis (ALS), while leaving expression of the normal allele intact (Ding et al., 2003).
[0128] The effectiveness of RNAi administration in vivo has been recently demonstrated in several mouse models of autoimmune hepatitis. Fas-mediated apoptosis is implicated in a broad spectrum of liver diseases. The in vivo silencing effect of siRNA duplexes targeting the Fas gene (also known as Tnfrsf6) encoding the Fas receptor was shown to protect mice from liver failure and fibrosis in these models. Intravenous injection of Fas siRNA specifically reduced Fas mRNA levels and expression of Fas protein in mouse hepatocytes, and the effects persisted without diminution for 10 days. In a fulminant hepatitis induced by injecting agonistic Fas-specific antibody, 82% of mice treated with siRNA that effectively silenced Fas survived for 10 days of observation, whereas all control mice died within 3 days (Song et al., 2003). A similar RNAi-based strategy is envisioned be useful in targeting or down-regulating abnormal or over-expressed genes in AMD patients.
[0129] Alternatively, expression of phagocytosis-related and/or AMDP-related genes can be reduced by targeting deoxyribonucleotide sequences complementary to regulatory regions of the phagocytosis-related or AMDP-related gene (i.e., the phagocytosis-related or AMDP-related gene promoters and/or enhancers) to form triple helical structures that prevent transcription of the phagocytosis-related or AMDP-related gene in target cells. (See generally, Helene, C. (1991) Anticancer Drug Des. 6(6):569-84; Helene, C., et al. (1992) Ann. N.Y. Acad. Sci. 660:27-36; and Maher, L. J. (1992) Bioassays 14(12):807-15). Nucleic acid molecules to be used in triple helix formation for the inhibition of transcription are preferably single-stranded and composed of deoxyribonucleotides. The base composition of these oligonucleotides should promote triple helix formation via Hoogsteen base pairing rules, which generally require sizable stretches of either purines or pyrimidines to be present on one strand of a duplex. Nucleotide sequences may be pyrimidine-based, which will result in TAT and CGC triplets across the three associated strands of the resulting triple helix. The pyrimidine-rich molecules provide base complementarity to a purine-rich region of a single strand of the duplex in a parallel orientation to that strand. In addition, nucleic acid molecules may be chosen that are purine-rich, for example, containing a stretch of G residues. These molecules will form a triple helix with a DNA duplex that is rich in GC pairs, in which the majority of the purine residues is located on a single strand of the targeted duplex, resulting in CGC triplets across the three strands in the triplex. Alternatively, the potential sequences that can be targeted for triple helix formation may be increased by creating a so-called "switchback" nucleic acid molecule. Switchback molecules are synthesized in an alternating 5'-3',3'-5' manner, such that they base pair with first one strand of a duplex and then the other, eliminating the necessity for a sizable stretch of either purines or pyrimidines to be present on one strand of a duplex.
[0130] Antisense RNA, ribozyme, RNAi, siRNA and triple helix molecules of the invention may be prepared by any method known in the art for the synthesis of such molecules. These include techniques for chemically synthesizing oligodeoxyribonucleotides and oligoribonucleotides well known in the art, such as for example solid phase phosphoramide chemical synthesis. Alternatively, RNA molecules may be generated by in vitro and in vivo transcription of DNA sequences encoding the antisense RNA molecule. Such DNA sequences may be incorporated into a wide variety of vectors which incorporate suitable RNA polymerase promoters. Alternatively, antisense cDNA constructs that synthesize antisense RNA constitutively or inducibly, depending on the promoter used, can be used.
[0131] Moreover, various well-known modifications to nucleic acid molecules may be introduced as a means of increasing intracellular stability and half-life. Possible modifications include but are not limited to the addition of flanking sequences of ribonucleotides or deoxyribonucleotides to the 5' and/or 3' ends of the molecule or the use of phosphorothioate or 2' O-methyl rather than phosphodiesterase linkages within the oligodeoxyribonucleotide backbone, as described above.
[0132] Other embodiments of agents that can down-regulate expression or neutralize the biological activity of the phagocytosis-related and/or AMDP-related genes of the invention are based on proteins. An example of a protein that can modulate expression and/or neutralize a biological function of a phagocytosis-related and/or AMDP-related gene product is an antibody that specifically binds a phagocytosis-related and/or AMDP-related polypeptide or peptide. Preferred polypeptides, for which mRNA levels are shown herein to be elevated in AMD, include those encoded by nucleic acids having SEQ ID NOS:2, 9, 15 and 17, i.e., polypeptides having amino acid sequences respectively identified herein as SEQ ID NOS:80, 89, 101, and 103-121. The antibodies of the invention can be used to interfere with the interaction of a phagocytosis-related and/or AMDP-related protein with one or more molecules that bind or otherwise interact with the phagocytosis-related and/or AMDP-related protein. For instance, an antibody directed against MT1-MMP protein is thought to neutralize the ability of this protein to activate progelatinase A. The results of a study described herein using an antibody directed against MT1-MMP showed delay of retinal degeneration in a rat model of RPE-based disease characterized by over-expression of MT1-MMP. Accordingly, inhibition of excessive production of MT1-MMP in the interphotoreceptor matrix using an anti-MT1-MMP antibody might be used in the eyes of patients with AMD to reduce destruction of the matrix and improve phagocytosis.
[0133] The proteins encoded by the nucleic acids of the invention (for example SEQ ID NOS:1-17, or immunogenic fragments or analogs thereof, and most preferably those encoded by nucleic acids found to be up-regulated in AMD (i.e., SEQ ID NOS:2, 9, 15 and 17) can be used to raise antibodies useful in the invention. Such proteins can be produced by purification from cells/tissues, recombinant techniques or chemical synthesis well known to those of skill in the art. Antibodies for use in the invention can include polyclonal antibodies, monoclonal antibodies, single chain antibodies, Fab fragments, F(ab')2 fragments, and molecules produced using a Fab expression library. See, for example, Kohler et al., Nature 256:495, 1975; Kohler et al., Eur. J. Immunol. 6:511, 1976; Kohler et al., Eur. J. Immunol. 6:292, 1976; Hammerling et al., "Monoclonal Antibodies and T Cell Hybridomas," Elsevier, N.Y., 1981; Ausubel et al., supra; U.S. Pat. Nos. 4,376,110, 4,704,692, and 4,946,778; Kosbor et al., Immunology Today 4:72, 1983; Cole et al., Proc. Natl. Acad. Sci. USA 80:2026, 1983; Cole et al., "Monoclonal Antibodies and Cancer Therapy," Alan R. Liss, Inc., pp. 77-96, 1983; and Huse et al., Science 246:1275, 1989.
[0134] Other protein-based agents that can modulate expression or activity of a phagocytosis-related and/or AMDP-related protein include variants of phagocytosis-related and/or AMDP-related proteins that can compete with the corresponding native proteins for binding ligands, for example naturally occurring ligands that bind prostaglandin D2 synthase (SEQ ID NO:2), CK1ε(SEQ ID NO:9), MT1-MMP (SEQ ID NO:15) and unknown gene AMDP-3 (SEQ ID NO:17). Such protein variants can be generated through various techniques known in the art. For example, a phagocytosis-related and/or AMDP-related protein variant can be made by mutagenesis, such as by introducing discrete point mutation(s), or by truncation. The mutation(s) can give rise to a phagocytosis-related and/or AMDP-related protein variant having substantially the same, or merely a subset of the functional activity of a native phagocytosis-related and/or AMDP-related protein. Alternatively, antagonistic forms of the protein can be generated which are able to inhibit the function of the naturally occurring form of the protein, such as by competitively binding to another molecule that interacts with a phagocytosis-related and/or AMDP-related protein. In addition, agonistic (or superagonistic) forms of the protein may be generated that constitutively express one or more phagocytosis-related and/or AMDP-related protein functional activities. Other variants of phagocytosis-related and/or AMDP-related proteins that can be generated include those that are resistant to proteolytic cleavage, as for example, due to mutations which alter protease target sequences. Whether a change in the amino acid sequence of a peptide results in a phagocytosis-related and/or AMDP-related protein variant having one or more functional activities of a native phagocytosis-related and/or AMDP-related protein can be readily determined by testing the variant for a native phagocytosis-related and/or AMDP-related gene protein functional activity (for example, binding a receptor or other ligand, or inducing a cellular response such as phagocytosis).
[0135] Another agent that can modulate expression or activity of a phagocytosis-related and/or AMDP-related gene product is a non-peptide mimetic or a chemically modified form of a phagocytosis-related and/or AMDP-related gene product that disrupts binding of a phagocytosis-related and/or AMDP-related protein to other proteins or molecules with which the native phagocytosis-related and/or AMDP-related gene product interacts. See, for example, Freidinger et al., in Peptides: Chemistry and Biology, G. R. Marshall ed., ESCOM Publisher: Leiden, Netherlands, 1988). Examples of such molecules include azepine (for example, see Huffman et al. in Peptides: Chemistry and Biology, G. R. Marshall ed., ESCOM Publisher: Leiden, Netherlands, 1988), substituted gamma lactam rings (Garvey et al. in Peptides: Chemistry and Biology, G. R. Marshall ed., ESCOM Publisher: Leiden, Netherlands, 1988), keto-methylene pseudopeptides (Ewenson et al. (1986) J. Med. Chem. 29:295; and Ewenson et al. in Peptides: Structure and Function (Proceedings of the 9th American Peptide Symposium) Pierce Chemical Co. Rockland, Ill., 1985), beta-turn dipeptide cores (Nagai et al. (1985) Tetrahedron Lett 26:647; and Sato et al. (1986) J. Chem. Soc. Perkin. Trans. 1:1231), and beta-amino alcohols (Gordon et al. (1985) Biochem. Biophys. Res. Commun. 126:419; and Dann et al. (1986) Biochem. Biophys. Res. Commun. 134:71).
[0136] A phagocytosis-related and/or AMDP-related protein may also be chemically modified to create a protein derivative by forming covalent or aggregate conjugates with other chemical moieties, such as glycosyl groups, lipids, phosphate, acetyl groups and the like. Covalent derivatives of phagocytosis-related and/or AMDP-related proteins can be prepared by linking the chemical moieties to functional groups on amino acid side chains of the protein or at the N-terminus or at the C-terminus of the polypeptide.
[0137] Yet other embodiments of agents that can modulate expression or activity of a phagocytosis-related or AMDP-related gene are small molecules. Small molecules from a wide range of chemical classes can interfere with the activity of a phagocytosis-related and/or AMDP-related protein, for example by binding to the protein and inactivating its activity, or alternatively by binding to a target of the phagocytosis-related and/or AMDP-related protein, thereby interfering with the interaction of the protein with its target. Depending upon the nature of the gene/protein of interest, inhibitory small molecules can be designed to achieve various purposes, such as 1) to occupy a binding site for a substrate or target interacting protein, 2) to bind to the phagocytosis and/or AMDP related protein so as to change its 3-dimensional conformation, thereby inhibiting its activity, or 3) to bind to a target molecule of the phago/AMDP protein, thereby inhibiting interaction of the protein with its normal target. For example, small molecule inhibitors of MT1-MMP protein (SEQ ID NO:100) are known, such as polyphenols extractable from green tea (i.e., Epigallocatechin 3-O-gallate (EGCG), (-)-epigallocatechin 3,5-di-O-gallate, and epitheaflagallin 3-O-gallate) that have potent and distinct inhibitory activity against this protein (Oku N. et al., Biol Pharm Bull. (2003) September; 26(9):1235-8). Other classes of inhibitors of metalloproteinases in general are disclosed, for example, in Beckett, R. et al. (2001), U.S. Pat. No. 6,310,084.
Gene Therapy for AMD and Other Retinal Degenerative Conditions Based on Phagocytosis-Related and AMD-Related Genes
[0138] In another aspect, the present invention provides for the delivery of natural or synthetic nucleic acids encoding phagocytosis-related and/or AMDP-related genes, or agents that modulate expression or activity of these genes. "Gene therapy" can be defined as the treatment of inherited or acquired diseases by the introduction and expression of genetic information in cells. Methods and compositions involving gene therapy vectors are described herein. Such techniques are generally known in the art and are described in methodology references such as Viral Vectors, eds. Yakov Gluzman and Stephen H. Hughes, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1988; Retroviruses, Cold Spring Harbor Laboratory Press, Plainview, N.Y., 2000; Gene Therapy Protocols (Methods in Molecular Medicine), ed. Jeffrey R. Morgan, Humana Press, Totawa, N.J., 2001.
[0139] In the various embodiments, the nucleic acids according to the invention are incorporated into recombinant nucleic acid constructs, typically DNA constructs, capable of introduction into and replication in a host cell. Such a construct preferably is a vector that includes a replication system and sequences that are capable of transcription and translation of a polypeptide-encoding sequence in a given host cell. For the present invention, conventional compositions and methods for preparing and using vectors and host cells can be employed, as described, for example, in Sambrook et al., supra, or Ausubel et al., supra.
[0140] Vectors useful in the practice of the invention comprise various types according to the purpose of the gene therapeutic approach. Some embodiments are vectors that include a nucleic acid encoding an agent that modulates (for example, down-regulates) expression of an AMDP-related or phagocytosis-related mRNA or protein. Other embodiments of the vectors include a wild-type or desirable polymorphic variant of a phagocytosis-related and/or AMDP-related gene of the invention. In various versions of the vectors of the former type, expression can be down-regulated by expressing, for example, an antisense RNA, ribozyme, RNAi molecule or triple helix molecule directed against an over-expressed mRNA, for example that of PD2S (SEQ ID NO:2), CK1ε (SEQ ID NO:9), MT1-MMP (SEQ ID NO:15), or AMDP-3 (SEQ ID NO:17).
[0141] Other embodiments of the vectors direct expression of a desired polymorphic form of an AMDP-related or phagocytosis-related gene, either a wild-type, or a variant form. For example, in one embodiment the nucleic acid encodes a normal (wild-type) form of MT1-MMP (for example, SEQ ID NO:15). Delivery of a wild type form can be useful, for example, for subjects who do not express the normal variant, but rather are homozygous for an undesirable polymorphic form (such as a D273N missense polymorphism of MT1-MMP described herein), or are heterozygous for two different undesirable allelic forms (for example, a D273N missense polymorphism and a P259P synonomous/splice variant polymorphism).
[0142] Natural or synthetic nucleic acids according to the present invention, including cDNAs, antisense, ribozyme and RNAi molecules can be incorporated into recombinant nucleic acid constructs, typically DNA constructs, capable of introduction into and replication in a host cell. For the present invention, conventional compositions and methods for preparing and using vectors and host cells can be employed, as described, for example, in Sambrook et al., supra, or Ausubel et al., supra. As used herein, an "expression vector" is a vector which (due to the presence of appropriate transcriptional and/or translational control sequences) is capable of expressing a DNA (encoding cDNA, antisense, ribozyme, or RNAi) molecule which has been cloned into the vector and of thereby producing an RNA or polypeptide/protein. Expression of the cloned sequences occurs when the expression vector is introduced into an appropriate host cell.
[0143] The precise nature of regulatory regions needed for gene expression may vary from organism to organism, and according to the nature of the cloned sequence and purpose for expressing the sequence in a cell, but in general these elements include a promoter which directs the initiation of RNA transcription. Such regions may include those 5' non-coding sequences involved with initiation of transcription, such as a TATA box. The promoter may be constitutive or regulatable. Constitutive promoters are those which cause an operably linked gene to be expressed essentially at all times. Regulatable promoters are those which can be activated or deactivated. Regulatable promoters include inducible promoters, which are usually "off," but which may be induced to turn "on," and "repressible" promoters, which are usually "on," but which may be turned "off." Many different regulators are known, including temperature, hormones, heavy metals, and regulatory proteins. These distinctions are not absolute; a constitutive promoter may be regulatable to some degree.
[0144] The promoter may be a "ubiquitous" promoter active in essentially all cells of the host organism, for example, the beta-actin or optomegalovirus promoters, or it may be a promoter whose expression is more or less specific to the target cell or tissue. Promoters suitable for cell-specific (for example, photoreceptor-specific, RPE-specific, and melanocyte-specific) expression in the eye, and inducible promoters used to initiate transgene expression in transgenic animals at specific ages are described in examples below.
[0145] A number of vectors suitable for stable transformation of animal cells or for the establishment of transgenic animals are known. See, for example, Pouwels et al., Cloning Vectors: A Laboratory Manual, 1985, Supp. 1987. Typically, animal expression vectors include (1) one or more cloned animal genes under the transcriptional control of 5' and 3' regulatory sequences and (2) a dominant selectable marker. Such animal expression vectors may also contain, if desired, a promoter regulatory region (for example, a regulatory region controlling inducible or constitutive, environmentally- or developmentally-regulated, or cell- or tissue-specific expression), a transcription initiation start site, a ribosome binding site, an RNA processing signal, a transcription termination site, and/or a polyadenylation signal. Animal expression vectors within the invention preferably contain a selectable marker gene used to identify the cells that have become transformed. Suitable selectable marker genes for animal systems include genes encoding enzymes that produce antibiotic resistance (for example, those conferring resistance to hygromycin, kanamycin, bleomycin, G418, or streptomycin). An example of a useful promoter which could be used to express a gene according to the invention is a cytomegalovirus (CMV) immediate early promoter (CMV IE) (Xu et al., Gene 272: 149-156, 2001). These promoters confer high levels of expression in most animal tissues, and are generally not dependent on the particular encoded proteins to be expressed. As an example, in most tissues of transgenic animals, the CMV IE promoter is a strong promoter. Examples of other promoters that are of use in the invention include SV40 early promoter, Rous sarcoma virus promoter, adenovirus major late promoter (MLP), Herpes Simplex Virus promoter, Mouse mammary tumor virus LTR promoter, HIV long terminal repeat (LTR) promoter, beta actin promoter (Genbank # K00790), or murine metallothionein promoter (Stratagene San Diego Calif.). Synthetic promoters, hybrid promoters, and the like are also useful in the invention and are known in the art.
[0146] Animal expression vectors may also include RNA processing signals such as introns, which have been shown to increase gene expression (Yu et al. (2002) 81: 155-163 and Gough et al. (2001) Immunology 103: 351-361). The location of the RNA splice sequences can influence the level of transgene expression in animals. In view of this fact, an intron may be positioned upstream or downstream of a phagocytosis-related or AMDP-related polypeptide-encoding sequence in the transgene to modulate levels of gene expression. Expression vectors within the invention may also include regulatory control regions which are generally present in the 5' regions of animal genes. Additionally, a 3' terminator region may be included in the expression vector to increase stability of the mRNA. See, for example, Jacobson et al. (1996) Annu. Rev. Biochem. 65:693-739; and Rajagopalan et al., (1997) Prog. Nucleic Acid Res. Mol. Biol. 56:257-286.
[0147] Adenovirus vectors have been shown to be capable of highly efficient gene expression in target cells and allow for a large coding capacity of heterologous DNA. "Heterologous DNA" in this context may be defined as any nucleotide sequence or gene which is not native to the adenovirus. Methods for use of recombinant adenoviruses as gene therapy vectors are discussed, for example, in W. C. Russell, Journal of General Virology 81:2573-2604, 2000, and Bramson et al., Curr. Opin. Biotechnol. 6:590-595, 1995.
[0148] A preferred form of recombinant adenovirus is a "gutless," "high-capacity," or "helper-dependent" adenovirus vector which has all viral coding sequences deleted, and contains the viral inverted terminal repeats (ITRs), therapeutic gene (including a natural or synthetic nucleic acid encoding a phagocytosis-related or AMDP-related gene, or an agent that modulates expression of a phagocytosis-related or AMDP-related gene, up to 28-32 kb) and the viral DNA packaging sequence. Variants of such recombinant adenovirus vectors such as vectors containing tissue-specific enhancers and promoters operably linked to a natural or synthetic nucleic acids encoding a phagocytosis-related or AMDP-related gene, or agent that modulates expression of such genes are also within the invention. More than one promoter can be present in a vector. Accordingly, more than one heterologous gene can be expressed by a vector.
[0149] The viral vectors of the present invention can also include Adeno-Associated Virus (AAV) vectors. AAV exhibits a high transduction efficiency of target cells and can integrate into the host genome in a site-specific manner. Methods for use of recombinant AAV vectors are discussed, for example, in Tal, J., J. Biomed. Sci. 7:279-291, 2000 and Monahan and Samulski, Gene Therapy 7:24-30, 2000. For cell-specific targeting, a preferred AAV vector comprises a pair of AAV inverted terminal repeats which flank at least one cassette containing a promoter which directs cell-specific (for example, photoreceptor, RPE, or melanocyte) expression, operably linked to the gene of interest. Using this vector, the DNA sequence of the AAV vector, including the ITRs, the promoter and natural or synthetic nucleic acid encoding a phagocytosis-related or AMDP-related genes, or agent that modulate expression of such a gene may be integrated into the host genome.
[0150] Another preferred vector for use in the invention is a herpes simplex virus (HSV) vector. Methods for use of HSV vectors are discussed, for example, in Cotter and Robertson, Curr. Opin. Mol. Ther. 1:633-644, 1999. HSV vectors, deleted of one or more immediate early genes (IE), are advantageously non-cytotoxic, persist in a state similar to latency in the host cell, and afford efficient host cell transduction. Recombinant HSV vectors allow for approximately 30 kb of coding capacity. A preferred HSV vector is engineered from HSV type I, deleted of the IE genes. HSV amplicon vectors may also be used according to the invention. Typically, HSV amplicon vectors are approximately 15 kb in length, possess a viral origin of replication and packaging sequences. More than one promoter can be present in the vector. Accordingly, more than one heterologous gene can be expressed by the vector. Further, the vector can comprise a sequence which encodes a signal peptide or other moiety which facilitates the secretion of the gene product from the host cell.
[0151] Viral vectors of the present invention may also include replication-defective lentiviral vectors, including HIV. Methods for use of lentiviral vectors are discussed, for example, in Vigna and Naldini, J. Gene Med. 5:308-316, 2000 and Miyoshi et al., J. Virol. 72:8150-8157, 1998. Lentiviral vectors are capable of infecting both dividing and non-dividing cells and of efficiently transducing epithelial tissues of humans. Lentiviral vectors according to the invention may be derived from human and non-human (including SIV) lentiviruses. These vectors may include the viral LTRs, primer binding site, polypurine tract, att sites and an encapsidation site. The lentiviral vector may be packaged into any suitable lentiviral capsid. The substitution of one particle protein by one from a different virus is referred to as "pseudotyping." The vector capsid may contain viral envelope proteins from other viruses, including Murine Leukemia Virus (MLV) or Vesicular Stomatitis Virus (VSV). The use of the VSV G-protein yields a high vector titer and results in greater stability of the vector virus particles. More than one promoter can be present in the lentiviral vector. Accordingly, more than one heterologous gene can be expressed by the vector.
[0152] The invention also provides for use of retroviral vectors, including Murine Leukemia Virus-based vectors. Methods for use of retrovirus-based vectors are discussed, for example, in Hu and Pathak, Pharmacol. Rev. 52:493-511, 2000 and Fong et al., Crit. Rev. Ther. Drug Carrier Syst. 17:1-60, 2000. Retroviral vectors according to the invention may contain up to 8 kb of heterologous (therapeutic) DNA, in place of the viral genes. Heterologous may be defined in this context as any nucleotide sequence or gene which is not native to the retrovirus. The heterologous DNA may include a tissue- or cell-specific promoter, as described above, and a phagocytosis-related and/or AMDP-related gene. The retroviral particle may be pseudotyped, and may contain a viral envelope glycoprotein from another virus, in place of the native retroviral glycoprotein. The retroviral vector of the present invention may integrate into the genome of the host cell. More than one promoter can be present in the retroviral vector. Accordingly, more than one heterologous gene can be expressed by the vector.
[0153] To combine advantageous properties of two viral vector systems, hybrid viral vectors may be used to deliver a phagocytosis-related or AMDP-related gene or an agent that modulate expression of such a gene, to a target tissue. Standard techniques for the construction of hybrid vectors are well known to those skilled in the art. Such techniques can be found, for example, in Sambrook, et al., In Molecular Cloning: A Laboratory Manual. Cold Spring Harbor, N.Y. or any number of laboratory manuals that discuss recombinant DNA technology. Double-stranded AAV genomes in adenoviral capsids containing a combination of AAV and adenoviral ITRs may be used to transduce cells. In another variation, an AAV vector may be placed into a "gutless," "helper-dependent," or "high-capacity" adenoviral vector. Adenovirus/AAV hybrid vectors are discussed, for example, in Lieber et al., J. Virol. 73:9314-, 1999. Retroviral/adenovirus hybrid vectors are discussed, for example, in Zheng et al., Nature Biotechnol. 18:176-186, 2000. Retroviral genomes contained within an adenovirus may integrate within the host cell genome and effect stable transgene expression. More than one promoter can be present in the hybrid viral vector. Accordingly, more than one heterologous gene can be expressed by the vector.
[0154] In accordance with the present invention, other nucleotide sequence elements which facilitate expression of a phagocytosis-related or AMDP-related gene, or agent that modulate expression or activity of such a gene, and cloning of the vector are further contemplated. The presence of enhancers upstream of the promoter, or terminators downstream of the coding region, for example, can facilitate expression.
[0155] Several non-viral methods are known for introducing a phagocytosis-related and/or AMDP-related nucleic acid, or an agent that modulates expression or activity of such a nucleic acid in a cell. For a review of non-viral methods, see, for example, Nishikawa and Huang, Human Gene Ther. 12:861-870, 2001. Various techniques employing plasmid DNA for the introduction into a cell of a phagocytosis-related and/or AMDP-related nucleic acid, or an agent that modulates expression of a phagocytosis-related and/or AMDP-related nucleic acid expressed within a cell are provided for according to the invention. Such techniques are generally known in the art and are described in references such as Ilan, Y., Curr. Opin. Mol. Ther. 1:116-120 (1999); and Wolff, J. A., Neuromuscular Disord. 7:314-318 (1997).
[0156] Methods involving physical techniques for the introduction into a host cell of a phagocytosis-related and/or AMDP-related nucleic acid, or an agent that modulates expression of such a nucleic acid in a cell can be adapted for use in the present invention. Cell electropermeabilization (also termed cell electroporation) may be employed for delivery of the selected nucleic acid into cells. This technique is discussed in Preat, V., Ann. Pharm. Fr. 59:239-244 (2001), and involves the application of pulsed electric fields to cells to enhance cell permeability, resulting in exogenous polynucleotide transit across the cytoplasmic membrane. Alternatively, the particle bombardment method of gene transfer involves an Accell device (gene gun) to accelerate DNA-coated microscopic gold particles into target tissue. This methodology is described, for example, in Yang et al., Mol. Med. Today 2:476-481 (1996); and Davidson et al., Rev. Wound Repair Regen. 6:452-459 (2000).
[0157] For construction of embodiments of the invention that are transgenic animals, several standard methods are known for introduction of recombinant genetic material into oocytes for the generation of a transgenic animal. Examples of such methods include: 1) particle delivery systems (see for example, Novakovic S et al. (1999) J Exp Clin Cancer Res 18:531-6; Tanigawa et al. (2000) Cancer Immunol Immunother 48:635-43); 2) microinjection protocols (see, for example, Krisher et al. (1994) Transgenic Res. 3: 226-231; Robinett C C and Dunaway M (1999), Modeling transcriptional regulation using microinjection into Xenopus oocytes. In: Methods: A Companion to Methods in Enzymology 17: 151-160; or Pinkert C A and Trounce I A (2002), Methods 26:348-57); (3) polyethylene glycol (PEG) procedures (see for example, Meyer O et al. (1998) J. Biol. Chem. 273:15621-7; or Park et al. (2002) Bioconj Chem, 13: 232-239); (4) liposome-mediated DNA uptake (see, for example, Hofland H E J and Sullivan S M (1997) J. Liposome Res. 7: 187-205; or Hui S W et al. (1996) Biophys. J. 71:590-599); and (5) electroporation protocols, described above.
[0158] Synthetic gene transfer molecules according to the invention can be designed to form multimolecular aggregates with plasmid DNA (harboring sequences encoding a phagocytosis-related and/or AMDP-related nucleic acid, or an agent that modulates expression or activity of such a nucleic acid in a cell, operably linked to a promoter) and to bind the resulting particles to a target cell surface in such a way as to trigger endocytosis and endosomal membrane disruption. Polymeric DNA-binding cations (including polylysine, protamine, and cationized albumin) can be linked to cell-targeting ligands to trigger receptor-mediated endocytosis. Methods involving polymeric DNA-binding cations are reviewed, for example, in Guy et al., Mol. Biotechnol. 3:237-248 (1995); and Garnett, M. C., Crit. Rev. Ther. Drug Carrier Syst. 16:147-207 (1999). Cationic amphiphiles, including lipopolyamines and cationic lipids, may provide receptor-independent gene transfer into target cells of phagocytosis-related and/or AMDP-related nucleic acids, or nucleic acids encoding an agent that modulates expression or activity of a phagocytosis-related and/or AMDP-related gene. Preformed cationic liposomes or cationic lipids may be mixed with plasmid DNA to generate cell transfecting complexes. Methods involving cationic lipid formulations are reviewed, for example, in Feigner et al., Ann. N.Y. Acad. Sci. 772:126-139 (1995); and Lasic and Templeton, Adv. Drug Delivery Rev. 20:221-266 (1996). Suitable methods can also include use of cationic liposomes as agents for introducing DNA or protein into cells. For therapeutic gene delivery, DNA may also be coupled to an amphipathic cationic peptide (Fominaya et al., J. Gene Med. 2:455-464, 2000).
[0159] Methods that involve both viral and non-viral based components may be used according to the invention. An Epstein Barr Virus (EBV) based plasmid for therapeutic gene delivery is described in Cui et al., Gene Therapy 8:1508-1513, 2001. A method involving a DNA/ligand/polycationic adjunct coupled to an adenovirus is described in Curiel, D. T., Nat. Immun. 13:141-164 (1994).
[0160] Protein transduction offers an alternative to gene therapy for the delivery of therapeutic proteins into target cells, and methods of protein transduction are within the scope of the invention. Protein transduction is the internalization of proteins into a host cell from the external environment. The internalization process relies on a protein or peptide which is able to penetrate the cell membrane. The transducing property of such a protein or peptide can be conferred upon proteins (phagocytosis-related and/or AMDP-related proteins, for example) which are expressed as fusion proteins. Commonly used protein transduction vehicles include the antennapedia peptide, the HIV TAT protein transduction domain and the herpes simplex virus VP22 protein. Such vehicles are reviewed, for example, in Ford et al., Gene Ther. 8:1-4 (2001).
[0161] Nucleic acids of the present invention may be expressed for any suitable length of time within the host cell, including transient expression and stable, long-term expression. In a preferred embodiment, a phagocytosis-related and/or AMDP-related nucleic acid, or an agent that modulates expression or activity of such a nucleic acid in a cell will be expressed in therapeutic amounts for a suitable and defined length of time. Methods of delivery that achieve either transient or long-term expression of a transgene are described herein. Episomally replicating vectors typically are maintained at intermediate to high copy number in the cell, which contributes to high levels of inserted DNA. Some vectors persist as episomes, and such vectors may behave as autonomous units replicating in the host independent of the host chromosome. DNA delivered via a plasmid or viral-based vector, including adenovirus, for example, exists in an episomal state within the host cell and is expressed in a transient manner.
[0162] Vectors according to the invention may contain nucleotide sequence elements which facilitate integration of DNA into host chromosomes. Integration is well tolerated by most transduced cells, and is preferred to ensure stability of newly introduced genetic information into a cell. Integration of a vector including a phagocytosis-related and/or AMDP-related nucleic acid, or a nucleic acid encoding an agent that modulates expression or activity of a phagocytosis-related and/or AMDP-related gene product in a cell may occur in a random or site-specific manner. Viral-based vectors that allow for integration into the host genome include those derived from AAV, retroviruses, and some AAV/adenovirus hybrids.
[0163] The compositions comprising nucleic acid molecules (including gene therapy vectors) of the invention may be administered to a mammalian subject by any suitable technique. For example, various techniques are known using viral vectors for the introduction of a natural or synthetic nucleic acid encoding a phagocytosis-related or AMDP-related gene, or in another aspect, an agent that modulates expression or activity of a natural or synthetic nucleic acid encoding a phagocytosis-related or AMDP-related gene. Viruses are naturally evolved vehicles which efficiently deliver their genes into host cells and therefore are desirable vector systems for the delivery of therapeutic genes. Preferred viral vectors exhibit low toxicity to the host cell and produce therapeutic quantities of the natural or synthetic nucleic acid encoding a phagocytosis-related or AMDP-related gene, or agent that modulates expression or activity of such a gene, for example in a tissue-specific manner. For delivery of the vectors of the invention to the eye, various approaches are known to those of skill in the art, including intraocular injection.
[0164] Association of MT1-MMP with AMD and Other Retinal Degenerations.
[0165] Some embodiments of the invention are methods of screening, animals models of retinal degeneration and treatment methods based on matrix metalloproteinase, membrane type 1 (MT1-MMP) (SEQ ID NO:15). Among the AMDP genes listed above, one gene, i.e., MT1-MMP, (herein also designated PHG-16 and AMDP-6), was initially selected for further evaluation as a candidate target for AMD therapy. As shown in the examples below, results of various confirmatory analyses clearly demonstrated that MT1-MMP is a phagogene, as evidenced by: 1) a diurnal pattern of expression, peaking in the early morning, the time of maximal OS shedding and phagoctytosis in vivo (FIG. 7); 2) localization to the tips of the OS in rat and human eyes (FIGS. 8, 9); and 3) inhibition of OS phagocytosis by an antibody to MT1-MMP, both in vitro (FIG. 10) and in vivo, following subretinal injection into rat eyes (FIG. 11).
[0166] A relationship of MT1-MMP with AMD was demonstrated by: 1) correlation of a graded increase in mRNA expression with severity of AMD-related changes in human donor eyes (FIGS. 12 and 13); 2) enhanced immunolocalization of MT1-MMP antibody in the interphotoreceptor matrix in a monkey model of AMD; and 3) increased incidence of a missense polymorphism (i.e., D273N) in the catalytic domain of MT1-MMP in human macular degenerative diseases including AMD, and increased incidence in AMD and macular degeneration patients of a synonymous polymorphism in MT1-MMP (i.e., P259P).
[0167] Additional studies of MT1-MMP provided evidence that overexpression of this gene is a common feature of at least one form of hereditary retinal degeneration besides AMD in which the primary etiology is in the RPE, i.e., that of the Royal College of Surgeons (RCS) rat. The RCS rat is a well known animal model of inherited retinal degeneration in which photoreceptor degeneration is due to a phagocytic defect in the RPE cells (Bok and Hall, 1971). The causative gene in this model is a mutated MERTK (D'Cruz et al. 2000). In studies described herein, MT1-MMP is shown to be overexpressed in the retina and RPE of the mutant RCS rat. Significantly, following injection of an anti-MT1-MMP antibody (2 μl volume) into the subretinal space of 7-day old RCS rats, the rate of photoreceptor degeneration relative to controls, is markedly slowed in anti-MT1-MMP antibody-injected animals observed at 30 and 60 days of age, whereas control antibodies or sham injection have no effect (FIG. 14). These results provide evidence that an agent directed against MT1-MMP protein present in the outer retina, for example within the interphotoreceptor matrix in the subretinal space, can provide a beneficial effect, such as slowing or reversing a retinal degenerative condition.
[0168] Previously recognized functions of MT1-MMP, which is expressed on invasive tumor cells, include an ability to activate progelatinase A, and to digest various ECM components (Sato et al., 1994; Cao et al., 1995; Pei and Weiss, 1996). Based on the discoveries described herein, it is now apparent that this gene provides an attractive new candidate gene to target therapeutically for AMD and other retinal and choroidal degenerative diseases.
[0169] Association of CK1ε with AMD, and with Cigarette Smoking, the Best Known Causative Environmental Factor for AMD
[0170] Some embodiments of the invention are methods of screening, animals models of retinal degeneration and treatment methods based on casein kinase 1 epsilon, CK1ε (SEQ ID NO:9). Among the AMDP genes listed above, this gene (herein also designated AMDP-4), was selected for further evaluation as a candidate target for AMD therapy. As shown in the examples below, results of various confirmatory analyses clearly demonstrated that CK1ε is a phagogene, as evidenced by: 1) changing levels of gene expression during in vitro phagocytosis assay (FIG. 4, CK1ε is represented as duplicate clones, identified in the graph as PHG-9 and PHG-10; see also Table 1); a diurnal pattern of expression in vivo (FIGS. 20A, B); 3) localization to the photoreceptor outer segments of the eyes.
[0171] A relationship of CK1ε with AMD was demonstrated by: 1) correlation of a graded increase in mRNA expression with severity of AMD-related changes in human donor eyes, demonstrated both by Northern analysis (FIG. 16) and real time quantitative PCR analysis (FIG. 17); 2) over-expression of CK1ε mRNA in retina and RPE/choroid (eyecup) of mice following laser photocoagulation treatment to produce choroidal neovascularization (CNV), a hallmark of the wet form of dry AMD (FIG. 19); and a phase shift in the peak time of expression of this gene in mice exposed to cigarette smoke, the number one environmental factor associated with incidence of AMD in humans. Additionally, exposure to smoke causes phase shifts of different durations in the retinas and eyecups of these mice (3 hours vs. 9 hours), suggesting a smoke-induced pathological mechanism involving "uncoupling" of the control of the circadian clocks governing diurnal gene expression in the two tissues (FIG. 20).
[0172] Previously recognized functions of CK1ε have demonstrated its intimate involvement as a regulator of circadian rhythms. In 1988, the golden hamster tau mutant, which has a free-running period of 22 hours, was the first mammalian circadian mutant discovered. Twelve years later, in 2000, the tau mutation was mapped to CK1ε (Lowrey, Phillip L.; Shimomura, Kazuhiro; Antoch, Marina P.; Yamakazi, Shin; Zemenides, Peter D.; Ralph, Martin R.; Menaker, Michael; Takahashi, Joseph S. (2000). Positional Syntenic Cloning and Functional Characterization of the Mammalian Circadian Mutation tau. Science 288 (5465): 483-491.) Since its discovery, the tau mutant has proven to be a valuable research tool in circadian biology.
[0173] At the time of our initial discovery, by the CHANGE expression profiling strategy, of the over-expression of clone 57-29 in human samples with AMD, this gene was unknown. By the year 2000, the CK1ε sequence was published in GenBank, and we were able to determine that our clone 57-29 encoded CK1ε. Based on the further discoveries described herein, it is now apparent that this gene provides an attractive new candidate gene to target therapeutically for AMD and other retinal and choroidal degenerative diseases.
[0174] Animal Models of AMD Based on Phagocytosis-Related and/or AMDP-Related Genes
[0175] In another aspect, the invention includes nonhuman transgenic animals (for example, mice) suitable for use as animal models of AMD and other degenerative conditions of the retina and choroid. Heretofore, testing of therapeutic compounds and treatment methods for AMD has been impeded by the lack of suitably short-lived animal models of the disease in which aging changes are practical to follow. Based on the discovery described herein of overexpression as determined by CHANGE of at least four AMD/phagogenes, i.e., PD2S (SEQ ID NO: 2), CK1ε(SEQ ID NO:9), MT1-MMP (SEQ ID NO:15) and AMDP-3 (SEQ ID NO:17) in human eyes with AMD, including demonstration of over-expression of MT1-MMP mRNA and protein in the retinas of humans with AMD, monkeys with AMD, RCS rats with inherited retinal degeneration, and a mouse laser model of AMD, and demonstration of the overexpression of CK1ε mRNA and protein in the retinas of humans with AMD and in two mouse models of AMD (laser photocoagulation model and mice subjected to cigarette smoke, a known causative factor in AMD), the invention provides as preferred embodiments transgenic animals that overexpress at least one of PD2S, CK1ε, MT1-MMP and AMDP-3.
[0176] Some of the transgenic models are engineered to conditionally overexpress the transgene only upon addition of an exogenous stimulus, such as doxycycline. Thus, the onset of transgene expression can be controlled in these animals by administration of doxycycline. As an example, transgene expression can be triggered at a particular time of life, such as after completion of postnatal development of the retina (occurring at around 30 days of age in a mouse). The feature of inducible expression is particularly advantageous with a gene such as MT1-MMP, which if overexpressed during the embryonic or early postnatal periods might be predicted to result in developmental abnormalities in the animals. Further details regarding how to make and use transgenic mouse models that inducibly over-express MT1-MMP and CK1ε are provided in Examples below. Other transgenic embodiments selectively overexpress a transgene, such as MT1-MMP, CK1ε, PD2S or AMDP-3 in particular cell types, for example in photoreceptors, RPE cells, or cell types of the choroid.
[0177] Yet other preferred embodiments of animal models of AMD/retinal and/or choroidal degenerations combine polymorphic variants of AMDP-related or phagocytosis-related genes, including those discovered and described herein. These models reflect the complex genetic inheritance pattern of AMD. A single genetic defect, such as a polymorphism present in MT1-MMP, may be unable to cause a disease in isolation. However, certain combinations of polymorphic variants of several genes, appropriate environmental factors, and the passage of time are likely to contribute jointly to dysfunction sufficient to tip the scale, the end result being AMD or another form of retinal, macular or choroidal degeneration. For example, other AMDP genes are likely to cooperate with polymorphic variants of MT1-MMP to produce the full spectrum of AMD.
[0178] Accordingly, some embodiments of the transgenic animal models of AMD and other retinal and choroidal degenerations express polymorphic variants of one or more genes with involvement in AMD and/or phagocytosis by RPE cells. Various preferred embodiments are polytransgenic models expressing MT1-MMP variants, for example in combination with polymorphic variants of one or more other AMD-related genes, including those AMDP genes disclosed herein (for example, genes having the wild type cDNA sequences shown herein as SEQ ID NOS: 2,9,10,16, 17), and AMD-related genes having polymorphic variants previously described to be correlated with AMD (for example, SEQ ID NOS:62, 63, 64, 65, 66, 67, 68, and 69). In other preferred embodiments of the polytransgenic models, polymorphic variants of MT1-MMP are expressed in combination with polymorphic variants of other phagocytosis-related genes (for example, genes having the wild type cDNA sequences shown herein as SEQ ID NOS:1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13 and 14).
EXAMPLES
[0179] The present invention is further illustrated by the following specific examples, which should not be construed as limiting the scope or content of the invention in any way.
Example 1
Research Tools for Isolation of Phagocytosis-Related and AMD-Related Genes
[0180] Described below are research tools developed during the course of the invention, including: 1) a simple and affordable method of simultaneously gauging expression in a large number of genes by hybridization; and 2) tools for identification of phagocytosis-related genes, based on a phagocytic RPE cell line and a vital assay of phagocytosis.
CHANGE Array System:
[0181] Referring to FIG. 1, a macroarray technique termed Comparative Hybridization Analysis of Gene Expression (CHANGE) was developed. λgt11 cDNA libraries were constructed using techniques well known to those of skill in the art of molecular biology, from rat RPE/choroid RNA and human retinal RNA. Rat RNA used for the library was obtained from the RPE/choroid of animals approximately 2-3 months of age, raised in cyclic light (12 h light:12 hr dark), and sacrificed at various times throughout the diurnal cycle. Approximately ten thousand clones from the libraries were individually picked, amplified on plates, and transferred to blots as arrays.
[0182] Total RNA from rat and human sources was used as a global expression hybridization probe, following conversion into cDNA, amplification by PCR, and testing to confirm its usefulness for detecting expression of specific genes on the arrays. Preliminary comparison of expression of a number of genes by CHANGE and Northern blot analysis confirmed the accuracy and demonstrated that a difference in mRNA expression as small as about 15-20% could be detected using the CHANGE method. It was apparent that the ability to readily perform iterative analysis with a combination of biologically related probes (for example, probes related on the basis of function, phenomenon, or pathology) was a very powerful aspect of this strategy.
[0183] Phagocytosis Gene Discovery Tools.
[0184] A preferred approach to identifying genes relevant to RPE phagocytosis in vivo is to analyze RPE gene expression in an in vitro system that performs the function of outer segment (OS) phagocytosis in a synchronous manner, as it occurs in vivo. In rodents and other mammals, shedding and phagocytosis of OS follows a circadian rhythm. Peak shedding by the photoreceptors and ingestion on a massive scale by the RPE cells is known to occur over a period of several hours beginning just before light onset (LaVail, 1976). To successfully identify phagogenes on the basis of differential expression in cultured RPE cells during the course of OS phagocytosis, it is preferable that the kinetics of the phagocytic process be uniform across the cultures, inter alia, to minimize "noise" from cells showing asynchronous phagocytosis with respect to their neighbors. Primary RPE cultures are generally unsuitable for this purpose, due to the marked phenotypic heterogeneity of RPE cells within primary cultures, and the corresponding heterogeneity in kinetics of phagocytosis displayed by cells of different phenotypes (McLaren, 1996).
[0185] The problem of heterogeneity can be circumvented by using an immortal RPE cell line that, like the RPE in vivo, demonstrates cobblestone morphology in culture, and is able to phagocytose fed OS with synchronous binding and ingestion. Methods for producing and maintaining immortal RPE cell lines from rodent and human sources are well known in the art. An exemplary cell line exhibiting the desired phagocytic characteristics is the BPEI-1 RPE cell line (McLaren et al., 1993b). BPEI-1 cultures were shown to follow the same kinetics of OS phagocytosis as "type 1" primary RPE cells, which most closely resemble RPE in vivo (McLaren et al., 1993a; McLaren, 1996). Use of such cell lines for isolation of phagocytosis-related genes is preferably carried out in large-scale phagocytosis assays having sufficient cells to yield RNA amounts (about 10-30 μg) needed for both probe preparation and Northern blotting. Accordingly, cells of a suitable RPE cell line, such as BPEI-1, are plated at high density (for example with approximately 106 cells per well in 6-well multi-well plates), and cultured for 1-2 days, for example in media as previously described (McLaren et al., 1993c; McLaren, 1996).
[0186] For preparation of probes for the CHANGE analysis representing specific stages of phagocytosis ("stage-specific" probes), it is advantageous to be able to follow the course of OS phagocytosis in living RPE cell cultures, to permit isolation of RNA at specific, documented, stages of the phagocytic process. To facilitate this, any suitable vital assay of OS phagocytosis can be used, for example, a double fluorescent assay previously described by McLaren et al. (1993c). Referring to FIG. 2, in this assay the lysosomes in the RPE cells are vitally stained with sulforhodamine (red fluorescence), and OS fed to the cells are prelabeled with fluorescein (FITC) (green fluorescence). The assay allows all stages of the phagocytic process (i.e., OS binding, ingestion, and digestion) to be followed by fluorescent microscopy in living cultures. FIG. 3 shows different stages of synchronous binding, ingestion and intracellular processing of OS typically observed in cultures of living BPEI-1 cells at various times after feeding the cells with FITC-stained OS.
[0187] Isolation of Phagogenes Using CHANGE.
[0188] To isolate phagogenes expressed at different stages of phagocytosis, stage-specific probes are prepared from total RNA extracted from the RPE cell cultures at various times (for example, 0, 1, 6, 12, 18, and 30 hours) after OS feeding, and at the same time points from control cultures not fed with OS. Following preparation of "+/-OS" phagocytosis probes by reverse transcription of the total RNA, pairs of such probes are used in a CHANGE analysis to screen a gene array, for example an array of approximately 10,000 RPE-expressed genes as disclosed herein, to identify those genes differentially expressed during OS phagocytosis by the RPE cells. Genes showing changes in expression during OS phagocytosis are subsequently identified by DNA sequence analysis using standard techniques and compared with sequences in databases such as GenBank.
Example 2
Isolation and Confirmation of Phagocytosis-Related Genes Expressed in RPE Cells
[0189] This example describes the isolation of genes showing changed expression during RPE phagocytosis, using the above-described methods.
[0190] From CHANGE analyses using "+/- OS" probes to screen arrays containing approximately 10,000 RPE-derived cDNAs, approximately 60 putative differentially expressed genes were initially obtained. Further detailed analyses, including confirmation of differential expression by Northern blot analysis, provided an initial subset of 16 confirmed phagocytosis-related genes selected for further investigation. Table 1 supra provides a listing of the identities and sequence listing notations (i.e., nucleic acids: SEQ ID NOS. 1-15 and amino acids: SEQ ID NOS:70-100) of confirmed phagogenes isolated as described herein by the CHANGE technique.
[0191] Detailed analysis of expression patterns of these genes during phagocytosis in vitro was examined in Northern blots of RNA extracted from BPEI cultures at various times after feeding the cells with OS. The particular stages of phagocytosis were observed in the living cells and documented photographically immediately prior to RNA extraction. As seen in FIG. 4, expression patterns of the 16 phagogenes were clustered into distinct groups that demonstrated peaks of expression at different times in the phagocytic process: i.e., early, early-mid, mid-late, and late.
Example 3
Isolation and Confirmation of RPE-Expressed Genes Exhibiting Differential Expression in AMD
[0192] Described herein are procedures used for isolation of putative AMD genes by CHANGE, and methods for confirming their relationship to AMD.
[0193] A similar approach to that described in Example 2 utilized the CHANGE technique to identify genes related to AMD, based on the assumption that genes playing a role in the pathogenesis of AMD show changes in expression during the course of the disease. Human donor eyes were obtained from a local eye bank. Generally, eyes were accepted that were enucleated within 3 hours of death and were available for processing within 12 hours. Regardless of time of death and time elapsed until processing, the actual quality of the tissue was assessed by several criteria, including appearance on gross examination, microscopic assessment of tissue sections, and the quantity and quality of the RNA obtained, as assessed by Northern blot analysis and RT-PCR.
[0194] Referring to FIG. 5, each eye was graded microscopically for AMD-related changes, on a scale of increasing severity of AMD changes from 0 to +5, in a strip of retina/choroid, approximately 3-4 mm wide, running from periphery to periphery and passing through the optic nerve head and the macula. In assigning a grade to each eye, several morphological criteria were taken into account, including: 1) degree of thickening of Bruch's membrane; 2) number, size, and location of any drusen; 3) presence or absence of neovascularization or choridal neovascular (CNV) membranes; and 4) RPE/photoreceptor atrophy, if any. RNA, DNA, and protein were isolated from the retina and RPE/choroid of each eye.
[0195] To prepare "+AMD" probes, total RNA was extracted from RPE/choroids of human donor eyes and pooled from multiple eyes with +3 to +5 (moderate to severe) AMD changes. Pooled RNAs from RPE/choroids of age-matched, unaffected eyes were used to prepare "- AMD" control probes. The +/- probes were used to identify differentially expressed genes by CHANGE, as described above. Approximately 200 RPE-expressed genes were initially identified that showed differential expression in subjects with AMD, compared to unaffected individuals.
[0196] To then obtain a subset of phagocytosis-related genes differentially expressed in AMD (i.e., "AMDP genes"), the results of the CHANGE screening for phagocytosis-related genes (Example 2 above) and the CHANGE screening for AMD-related genes (this example) were compared, to identify those RPE genes on the CHANGE panels demonstrating differential expression in both phagocytosis and AMD. The results of this analysis yielded an initial subset of 6 genes fitting both criteria, i.e., prostaglandin D2 synthase (SEQ ID NO:2), casein kinase epsilon 1 (CK1ε) (SEQ ID NO:9), ferritin heavy polypeptide 1 (SEQ ID NO:10), MT1-MMP (SEQ ID NO:15), SWI/SNF related/OSA-1 nuclear protein (SEQ ID NO:16) and human unknown cDNA AMDP-3 (SEQ ID NO:17). (See also Table 2, supra.)
Example 4
Isolation and Characterization of MT1-MMP as an AMD-Related and Phagocytosis-Related (AMDP) Gene
[0197] This example describes the identification of MT1-MMP (SEQ ID NO:15), an exemplary gene found by CHANGE to be differentially expressed in both phagocytosis and in AMD (i.e., an "AMD-related phagogene," or "AMDP gene"), and results of studies confirming that MT1-MMP is a phagogene and is upregulated in AMD eyes.
[0198] To identify genes related to both AMD and OS phagocytosis, the results of the two CHANGE analyses were compared as described above. Among the candidate genes differentially expressed in both screens, clone 91-40 stood out, as being a relatively new type of metalloproteinase, i.e., MT1-MMP (Sato et al., 1994) having functions that would reasonably fulfill the requirements of a gene with suspected involvement in AMD. These functions include a role in OS phagocytosis (as disclosed herein) as well as activation of progelatinase A and degradative activity against various extracellular matrix components (Sato et al., 1994; Cao et al., 1995; Pei and Weiss, 1996).
[0199] Northern blot analysis of expression of MT1-MMP in various tissues demonstrated highest levels of expression in the RPE, choroid, and retina, followed by lung and adrenal. The putative designation of MT1-MMP as a phagogene was based on its differential expression detected by CHANGE during OS phagocytosis in vitro. For functional confirmation, the pattern of expression of this gene was examined by Northern blot analysis in an independent assay of OS phagocytosis. Referring to FIG. 6, the result confirmed the increase in MT1-MMP expression at 13 hours after the initiation of phagocytosis, the same time of increase detected by CHANGE. The involvement of MT1-MMP in diurnally controlled OS phagocytosis in vivo was strongly supported by the further finding that expression of MT1-MMP mRNA, in both RPE and retina, follows a diurnal pattern with a peak at 6 AM, approximately 1-2 hours prior to the time of maximal shedding and phagocytosis of OS in vivo (FIG. 7).
[0200] Referring now to FIG. 8, immunofluorescent localization of MT1-MMP in the rat retina at several time points throughout the diurnal cycle demonstrated the strongest signal in the photoreceptor OS and RPE in retinas fixed at 6 AM. Immunolocalization of MT1-MMP protein in the human retina demonstrated signal in the tips of the rod, and especially cone, outer segments, consistent with activity at the interface between the photoreceptor OS membranes and the RPE apical processes, where it may be playing a role in preparing the OS tips for shedding and phagocytosis by the RPE (FIG. 9).
[0201] To obtain functional confirmation of the involvement of MT1-MMP in OS phagocytosis, an antibody against MT1-MMP (Chemicon International, Temecula, Calif.) was tested for its ability to inhibit OS phagocytosis by BPEI-1 cells in vitro. As seen in FIG. 10, the results clearly demonstrated inhibition of OS phagocytosis by this antibody, but not by an irrelevant (X-arrestin) antibody, confirming the functional requirement of MT1-MMP for the process of OS phagocytosis. Furthermore, in an in vivo functional assay, subretinal injection of the MT1-MMP antibody, but not X-arrestin antibody, into normal rat eyes resulted in marked structural disorganization and lengthening of the OS four days later, consistent with interference with the daily phagocytic process (FIGS. 11A, B). Thus, abundant evidence pointed to the involvement of MT1-MMP in OS phagocytosis by RPE cells.
[0202] MT1-MMP was also identified as a putative AMD gene by CHANGE on the basis of differential expression in AMD (i.e., an increase). The expression of this gene was examined independently by Northern blot analysis of RNAs from the RPE/choroid and retina of AMD-affected and normal human donor eyes. The result confirmed the increase and showed a greater increase in the retina than in the RPE (FIG. 12). As shown in FIG. 13, when a series of RNA samples from eyes with varying severity of AMD-related changes was tested, a positive correlation of increased expression of MT1-MMP in the retina was observed with increasing pathology in the eye (FIG. 13). This result strongly supported a possible role for this gene in the pathogenesis of AMD. Further, when tested in a monkey model of AMD that also showed increased expression of MT1-MMP by Northern analysis, MT1-MMP was found to be localized in the interphotoreceptor matrix (IPM) among highly disorganized OS.
[0203] Because MT1-MMP had been discovered to play a role in diurnally regulated OS phagocytosis, the inventors next tested whether the increased expression in AMD occurred at the time of maximal shedding and phagocytosis. The increase in MT1-MMP expression seen in the human eyes with AMD changes did not support this possibility, as the increase was present in eyes obtained at many different times of day after death. A plausible explanation for this result is that there may be dysregulation of MT1-MMP expression, which normally should peak only at approximately 6 AM, but in AMD may be highly active at other times as well. The functional consequence of dysregulation of MT1-MMP expression to the tightly controlled diurnal processes of OS shedding and phagocytosis could be profoundly deleterious over time.
Example 5
Isolation and Characterization of CK1ε as an AMD-Related and Phagocytosis-Related (AMDP) Gene
[0204] This example describes the identification of CK1ε (SEQ ID NO:9), an exemplary gene found by CHANGE to be differentially expressed in both phagocytosis and in AMD (i.e., an "AMD-related phagogene," or "AMDP gene"), and the results of studies confirming that CK1ε is a phagogene and is upregulated in AMD eyes.
[0205] To identify genes related to both AMD and OS phagocytosis, the results of the two CHANGE analyses were compared, as described above. Among the candidate genes determined to be differentially expressed in both screens, one clone of particular interest was clone 57-29 (identified in Table 1, supra, as two duplicate clones, i.e., PGH-9 and PHG-10, both determined upon sequencing to be casein kinase 1 epsilon, CK1ε, (SEQ ID NO:9)).
[0206] The putative designation of CK1ε as a phagogene was based on its differential expression detected by CHANGE during OS phagocytosis in vitro. For functional confirmation, the pattern of expression of this gene was examined by Northern blot analysis in an independent assay of OS phagocytosis. To determine the expression pattern of this gene by Northern analysis, a hybridization probe specific for rat and human CK1ε was created using a PCR-amplified sequence from the 57-29 rat CK1ε clone, obtained using 5' and 3' primers having the respective sequences:
TABLE-US-00003 (SEQ ID NO: 132) 5'-AAGTGTTGAATTTACCCTTC-3'; and (SEQ ID NO: 133) 5'-TACTTCAAATTAACAACCAC-3'.
[0207] As shown in FIG. 4 (data shown for genes identified as PHG-9 and PHG-10), this gene was confirmed by Northern blot analysis to be differentially expressed at various times during a 32 hour in vitro OS phagocytosis assay, peaking at the 16-hour timepoint. This timepoint corresponds to the period several hours after synchronous mass ingestion of ROS by the cultured RPE cells (see FIG. 3).
[0208] Immunolocalization of CK1ε protein in the mouse retina demonstrated signal in the photoreceptor outer segments (OS), a location that clearly would be consistent with a protein involved in phagocytois of photoreceptor OS by the RPE cells of the retina. Thus, the confirmatory evidence clearly pointed to the involvement of CK1ε in OS phagocytosis by RPE cells, as first revealed by the phagocytosis prong of the iterative CHANGE strategy.
[0209] As indicated above, CK1ε was also identified as a putative AMD gene in the second prong of the iterative CHANGE analysis (see Table 2), on the basis of differential expression in AMD (i.e., an increase). As discussed. in initial studies with this candidate AMD gene, conducted between the years 1998-2000, the expression studies were carried out by Northern blot analysis of mRNAs isolated from the RPE/choroid and retina of AMD-affected and normal human donor eyes, and the CK1ε mRNA transcript was detected using the hybridization probe described above. The Northern result confirmed the increase in expression revealed by CHANGE. As shown in FIG. 16, when a series of RNA samples from eyes with varying severity of AMD-related changes was tested, a positive correlation was observed of increased expression of CK1ε in the retina with increasing pathology in the eye, up to Grade 3 pathology. This result strongly supported a possible role for this gene in the pathogenesis of AMD. In more recent studies, conducted since the advent of real time PCR methodology and its predominance over Northern blot analysis for studies of gene expression, very similar results were obtained by real time PCR, using respective forward and reverse primers shown herein as:
TABLE-US-00004 (SEQ ID NO: 134) 5'-GCCTCTGGTGAGGAAGTCG-3'; and (SEQ ID NO: 135) 5'-CGGTAGGGAATGTGCTGGTG-3'
to amplify a 416 base pair fragment of the human CK1ε sequence. These results are further described in Example 6, infra, and are illustrated in the graph in FIG. 17A, which shows the results of real time PCR analysis of CK1ε expression in pooled RNA taken from a large number of samples of retina and eyecup (RPE/choroid) from human eyebank eyes sorted by grade level of pathology. As can be seen, the results obtained by real time PCR showed the identical pattern to those intially obtained by Northern analysis, confirming the initial Northern blot data (compare FIGS. 16 and 17A). Thus, by both methods of detection, the results confirmed the increase in expression of CK1ε in AMD, as initially detected by the CHANGE hybridization analysis.
Example 6
Evidence of Altered Expression and "Phase-shifting" of CK1ε in Animal Models of AMD
[0210] Example 5 supra provides evidence that CK1ε expression is upregulated in the eyes of humans with either the wet or dry forms of AMD. This example describes results of studies using two different animal models of AMD that show that CK1ε expression is increased in conditions in which AMD-like pathology is induced, consistent with the findings in the human eyebank samples described above.
[0211] Background.
[0212] Age-related macular degeneration (AMD) is the leading cause of blindness in the elderly population over 60 years old (Congdon et al. 2004; Klein et al. 1992; Friedman et al. 2004). Central vision necessary for reading, driving, and recognizing faces is lost in AMD, therefore making it a devastating condition for people who are otherwise still physically active. There are two types of AMD, the dry and the wet form (Bird et al. 1995). In both types, the retinal pigment epithelium (RPE), which is the single layer of epithelial cells in the back of the eye just behind the light-sensing photoreceptor cells, is thought to degenerate first (Green et al. 1985). The RPE in the macula, which is the region responsible for central vision, in the retina is affected most, thus, leading to the loss of central vision. This is because the RPE cells are critical to maintaining the homeostasis of photoreceptors, which are the light-sensing cells of the retina (Strauss 2005). When the RPE cells degenerate, the photoreceptors associated with the RPE degenerate, resulting in loss of vision.
[0213] In the dry form of AMD, focal degeneration of the RPE leading to photoreceptor degeneration occurs, especially in the macular region. This results in patches of RPE and photoreceptor degeneration and regionalized blindness in a pattern called geographic atrophy (GA) (Midena et al. 1997). The pattern of GA sometimes progresses to the wet form in which abnormal new blood vessels grow from the choroid, the layer of connective tissue containing blood vessels behind the RPE, into the space below or through the RPE layer into the subretinal space, i.e., in a process called neovascularization (Berger et al. 1999). These abnormal blood vessels leak, causing inflammation and cellular destruction of the RPE and photoreceptors in a process called choroidal neovascularization (CNV). The wet form of AMD and the CNV is responsible for ˜90% of the blindness that occurs in AMD (Ferris et al. 1984).
[0214] At this time there is no treatment for the dry form of AMD. Presently there is palliative treatment for the wet form of AMD, consisting of administration by intraocular injection of anti-vascular endothelial growth factor (VEGF) antibody to the eyes of patients suffering from wet AMD, at repeated intervals. Compared to the previously available treatment of photodynamic therapy for the wet form, the anti-VEGF treatment has shown substantial success in at least slowing down or stopping the CNV (Verteporfin in Photodynamic Therapy Study Group. 2001; Brown et al. 2006; Rosenfeld et al. 2006). It is, however, not a cure, and frequent, long-term injections of the antibody (Lucentis, Avastin) are required. When the injections are discontinued, abnormal blood vessels begin to grow again. Furthermore, up to 60% of the patients also do not respond adequately to anti-VEGF treatments (Rosenfeld et al. 2006). Thus, there is a continuing unmet need for better treatments to replace or augment anti-VEGF therapy for wet AMD.
[0215] The etiology of AMD is not known. Genetic and environmental factors are known to be important (Klein et al. 1994; Seddon et al. 1997). This condition is thought to be caused by the interaction of a number of genes gone awry, as well as environmental factors, the main ones being aging and cigarette smoking (Klein et al. 2004; Hammond et al. 1996; Smith et al. 1996; Tomany et al 2004). Because of the complexity of the likely etiology, despite great effort of many investigators, it has been difficult to elucidate the cause and the mechanism of this disease. Some of the likely causative factors proposed by the investigators include oxidative stress and damage, and inflammation (Winkler et al. 1999; Hageman and Mullins 1999; Frank et al. 1999).
[0216] The importance of inflammation as a causative factor in AMD has recently emerged from the demonstration of associations of various inflammatory proteins, such as complement factor H (CFH), factor B, and complement 2, with AMD (Edwards et al. 2005; Gold et al. 2006). However, a definitive answer on the cause and mechanism is not at hand yet. In this respect, the one environmental factor that has been definitively shown to increase the incidence of AMD, i.e., cigarette smoking, may be an extremely useful handle to uncover the cause of AMD. This is because some of the other candidate genes and mechanisms for AMD have already been identified through studying the AMD patients' DNA and tissues (Dewan et al. 2006; Edwards et al. 2005; Gold et al. 2006; Rivera et al. 2005; Justilien et al. 2007; Inana et al. 2007). Thus, if genes, proteins, and mechanisms that are affected by cigarette smoking are identified, and if any of them matches the already-identified candidates for AMD, it would go a long way towards strengthening the hypothesis that smoking contributes to AMD through the specific identified candidate(s).
[0217] The deleterious effects of tobacco smoke, which contains over 4000 different compounds, are well known with respect to cancer, lung diseases, and cardiovascular diseases (Pryor et al. 1992; Lyons et al. 1958; Cosgrove et al. 1985). The direct effects of tobacco smoke on reactive oxygen species (ROS) formation and oxidative stress, inflammation, and DNA damage have been demonstrated (Lyons et al. 1958; Cosgrove et al. 1985; Kalra et al. 1991; Asami et al. 1996; Piperakis et al. 1998). Although the direct negative effects of tobacco smoke on these processes have been demonstrated, surprisingly, the effects of smoking on the expression of genes in the target tissues have not been extensively investigated. Effects on such processes as ROS formation, inflammation, and DNA damage often start at the level of changes in the expression of genes involved in these processes. Since abnormal expression of specific genes is postulated to be central to the mechanism of AMD, it follows that it would be important to examine gene expression in an effort to find the commonality between smoking and AMD, and clearly this examination would best be carried out by determining gene expression profiles in the tissues most relevant to AMD, namely the RPE and the retina.
[0218] With this goal in mind, we set out to uncover genes common to smoking and AMD by performing a gene expression profiling analysis on RPE and retina from mice exposed to heavy smoke (HS) or second hand smoke (SHS) for different periods of time, and analyzing the results for matches in genes or mechanisms postulated to play a role in AMD.
[0219] Materials and Methods.
[0220] 1. Cigarette Smoke Exposure of Mice
[0221] All mice were entrained to a 12 hour Light: 12 hour Dark (12L:12D) lighting cycle with lights on at 7:00 AM. Groups of 10 adult C57BL6 mice (Charles River, Wilmington, Mass.) were exposed to second hand smoke (SHS) or heavy smoke (HS) from cigarettes, at a total suspended particulate (TSP) of 3-5 mg/m3 or 40-60 mg/m3, respectively, 6 hours/day, 5 days/week in a Teague TE10 smoking machine (Teague Enterprises, Davis, Calif.). Each group of mice was exposed to smoke for 3 or 6 weeks. All procedures using animals were conducted in accordance with the ARVO Statement for the Use of Animals in Ophthalmic and Vision Research. The 3R4F reference cigarettes from the University of Kentucky, College of Agriculture, Reference Cigarette Program were used. The TSP was measured using the 25 mm Pallflex membrane filter (EMFAB TX40H120-WW) (VWR) in a filter holder connected to the TE10 chamber sample port and the dry gas meter and weighing the filter before and after exposure to particulates. The mice did not require prior acclimatization to cigarette smoke at the low dose of SHS.
[0222] 2. Isolation of RNA from Mouse Eyes
[0223] After the scheduled smoke exposures, at eight diurnal time points (every 3 hours, starting at 12 AM), and at 13 time points after laser photocoagulation of the retina (0, 1, 3, 6, 12, 24, 48, 72 hours, 4, 6, 10, 12, 14 days) according to a published procedure (Espinosa-Heidmann et al. 2002), mice were euthanized and the eyes were enucleated. The retina and RPE/choroid were isolated from the eyes by microdissection and used for isolation of RNA with the Trizol Reagent (InVitrogen) (Chomczynski and Sacchi 1987). The smoke-exposed mouse RNA was further purified using the RNeasy mini-elute columns (Qiagen, Valencia, Calif.). The quality of the RNA was assessed by denaturing gel electrophoresis, spectrophotometric readings, and quantitative reverse-transcription polymerase chain reaction (qRT-PCR) amplification to test the functional integrity.
[0224] 3. Expression Profiling
[0225] Microarray. The smoked-exposed mouse RNA samples were used for expression profiling using the 38.5K Mouse Exonic Evidence Based Oligonucleotide (MEEBO) microarray (Lot 20595) by Ocean Ridge Biosciences (ORB, Palm Beach Gardens, Fla.). MEEBO microarrays were printed by Microarrays Inc. (Nashville, Tenn.) and contained 38,083 70-mer oligonucleotides probes complementary to constitutive exons of most mouse genes, as well as alternatively spliced exons, and control sequences. For more information on the MEEBO oligonucleotide set please refer to http://alizadehlab.stanford.edu/.
[0226] Sample Processing.
[0227] The RNA sample groups were made up of 1) control retina, 2) control eyecup, 3) 3-week retina, 4) 3-week eyecup, 5) 6-week retina, and 6) 6-week eyecup. Biotin-labeled complementary RNA (cRNA) was prepared from total RNA after two rounds of amplification by the method of Van Gelder et al. (Van Gelder, Multi-gene expression profile, U.S. Pat. No. 7,049,102). Briefly, an oligonucleotide containing a 5'-T7-promoter sequence and a 3' T24VN sequence was used to prime reverse transcription of RNA catalyzed by Superscript II (Invitrogen, Carlsbad, Calif.). Double-stranded cDNA was prepared from the 1st strand product by the method of Gubler and Hoffman (1983), and purified on a PCR purification column (Qiagen, Valencia, Calif.). The double-stranded cDNA was then used as a template for in vitro transcription with T7 RNA polymerase using a high yield transcription kit (Ambion, Austin, Tex.). A random primer (Invitrogen, Carlsbad, Calif.) was then used to prime the second round of reverse transcription of the purified cRNA catalyzed by Superscript II. Double-stranded cDNA was prepared from the 1st strand product after the hydrolysis of cRNA using RNAse H (Epicentre, Madison, Wis.). The column purified double-stranded cDNA was then used as a template for in vitro transcription with T7 RNA polymerase using the same high yield transcription kit (Ambion, Austin, Tex.) and including biotin-16-UTP (Epicentre, Madison, Wis.) in the reaction mixture. Biotinylated cRNA samples were purified on RNeasy MinElute purification column (Qiagen, Valencia, Calif.).
[0228] The cRNA samples were fragmented, diluted in a formamide-containing hybridization buffer, and loaded on to the surface of MEEBO microarray slides enclosed in custom hybridization chambers. The slides were hybridized for 16-18 hours under constant rotation in a Model 400 hybridization oven (Scigene, Sunnyvale, Calif.). After hybridization, the microarray slides were washed under stringent conditions, stained with Streptavidin-Alexa-647 (Invitrogen), and scanned using an Axon GenePix 4000B scanner (Molecular Devices, Sunnyvale, Calif.).
[0229] Data Pre-Processing.
[0230] Spot intensities for each probe were calculated by subtracting median local background from median local foreground for each spot. Spot intensities were transformed by taking the base 2 logarithm of each value. The spot intensities were then normalized by subtracting the 70th percentile of the spot intensities of probes against mouse constitutive exons and adding back a scaling factor (grand mean of 70th percentile). After removing data for low quality spots, control sequences, and non-mouse probes, 35,028 mouse probe intensities remained. The mouse probe intensities were filtered to identify all probes with intensity above a normalized threshold (log 2 (5*standard deviation of raw local background)+mean of log 2-transformed negative controls), to arrive at 23,204 probes above threshold in at least one sample.
[0231] Data Presentation.
[0232] The criteria considered for the gene ontology analyses were all) all probes, 0) 6 week retina vs. control retina, up>2 fold, 1) 6 week retina vs. control retina, down>2 fold, 2) 3 week retina vs. control retina, up>2 fold, 3) 3 week retina vs. control retina, down>2 fold, 4) 6 week eyecup vs. control eyecup, up>2 fold, 5) 6 week eyecup vs. control eyecup, down>2 fold, 6) 3 week eyecup vs. control eyecup, up>2 fold, and 7) 3 week eyecup vs. control eyecup, down>2 fold, 8) Principal Component (PC) number=3.
[0233] Fold changes between the treatment and control sample were computed and presented along with the log 2 transformed spot intensities of the 23,204 detectable probes. Principal Component Analysis was performed on the 23,204 detectable probes using the module built in to the National Institute of Ageing (NIA) Array Analysis software (Sharov et al. 2005).
[0234] Gene Ontology Analysis.
[0235] Gene ontology categories showing significant over-representation of differentially expressed genes were determined using GenMAPP software (Gladstone Institute, San Francisco, Calif.) for 23,204 detectable probes with current Entrez Gene IDs. Specifically, the MAPPfinder module of GenMAPP was first used to map all detectable probes, based on their gene targets, to GO and Local MAPP categories. Then MAPPfinder compared the relative representation in each functional group of genes associated with probes meeting one of nine differential expression criteria described above, to the relative representation of genes associated with the full set of 23,204 detectable probes. Significance was determined by permutation of Z scores with correction for multiple comparisons as described in the GenMAPP software manual.
[0236] WebGestalt software (Dr. Bing Zhang's group, Vanderbilt University) was utilized for a statistics-based pathway analysis in order to determine the distribution of differentially-regulated genes among functional biological pathways. The software compares the relative distribution of genes associated with probes that meet specific fold-change and significance criteria to the distribution of all genes associated with probes showing detectable signal on the array. The WebGestalt software was used to query two pathway databases including KEGG and Biocarta. Significance of the individual pathways was determined by the hyper geometric statistical method with criteria, adjusted to P<0.05 and more than 2 genes per pathway.
[0237] Hierarchical Clustering of Gene Expression Data.
[0238] Data for the 23,204 detectable probes were clustered using Cluster 3.0 software (DeHoon et al. 2004). The data was pre-processed by three consecutive rounds of gene median centering and gene median normalization, and then hierarchically clustered using uncentered correlation as the similarity metric and single linkage as clustering method.
[0239] 4. Quantitative RT-PCR
[0240] RNA samples from 1) the smoking experiment (eight diurnal time points), 2) the laser photocoagulated eyes, and 3) our archival normal control and AMD patient eyecups (Eye Bank eyes graded 1 to 4 according to a photomicrograph-based AREDS grading system, 3 samples per grade) were subjected to quantitative reverse transcription-polymerase chain reaction amplification (qRT-PCR) to determine the level of expression of CK1ε. The RNA (0.5-2 ug) was converted to cDNA with ImProm-Reverse Transcriptase (Promega Corp., Madison, Wis.) according to manufacturer's protocol. The equivalent of 1 ng of RNA/cDNA was used for qPCR with iQ SYBR Green Supermix (BioRad Corp., Hercules, Calif.) according to the manufacturer's protocol in the BioRad iCycler iQ Real-Time PCR Detection System. The PCR protocol consisted of incubation at 95° C. for 5 minutes, then 35 cycles of 95° C. for 1 minute, annealing temperature of 60° C. for 30 seconds, extension for 1 minute at 70° C., followed by 70° C. for 8 minutes. The PCR primers used were hck1eF (5'-GCCTCTGGTGAGGAAGTCG-3') (SEQ ID NO: 134) and hckleR (5'-CGGTAGGGAATGTGCTGGTG-3') (SEQ ID NO:135). The primer sequences were based on the human CK1ε mRNA sequence (GenBank: CU012990.1), but fortuitously, due to high sequence homology, they were also usable with rodent RNA. Since the Standard Curve method of calculating the relative expression of genes was used (ABI Prism 7700 Sequence Detection System, User Bulletin #2, October 2001), standard curves were determined for the gene primer set with the appropriate tissue, and the parameters from the standard curve were used in calculating the relative expressions from the results of the real-time PCR analysis.
[0241] Results.
[0242] Groups of 10 adult C57BL6 mice each were exposed to second hand smoke (SHS) or heavy smoke (HS) from cigarette smoke (3R4F Reference Cigarettes) in a Teague TE10 smoking machine chamber for 3 weeks and 6 weeks as described in Methods. The smoke exposure protocol was total suspended particulate (TSP) of 3-5 mg/m3 for SHS and 40-60 mg/m3 for HS, 6 hours/day, 5 days/week, consistent with most published protocols (Yuan et al. 2007; Zhu et al. 2003; Kuo et al. 2005; Churg et al. 2007). At 3 and 6 weeks of exposure, the mice, including a group of 10 unexposed control mice, were euthanized, and the eyes were obtained. Neuroretinas and RPE/choroid, or eyecups (EC), were isolated, and total RNA was prepared using Trizol. The quality of the RNA was examined by denaturing (formaldehyde) gel electrophoresis and spectrophotometric analysis. The former demonstrated the expected 28s and 18s ribosomal species without evidence of degradation (tail at the bottom), and the latter showed the OD260/280 ratio of the samples to be at least 1.70, consistent with a preparation of RNA of acceptable quality. The functional integrity of the RNA samples was tested by using them for quantitative reverse-transcription polymerase chain reaction (qRT-PCR) amplification of the cytochrome P450 enzyme, Cyp1b1, a gene related to and modulated by tobacco smoke (Grassman et al. 1998). Evidence of expression of Cyp1b1 in all of the samples was observed, confirming the functional integrity of the RNA samples (data not shown).
[0243] For gene expression profiling, approximately 5 μg each of the SHS RNA was used for preparation of biotinylated cRNA probes for hybridization to the 38.5K Mouse Exonic Evidence Based Oligonucleotide (MEEBO) microarray by Ocean Ridge Biosciences (ORB, Palm Beach Gardens, Fla.). Six samples were analyzed: 1) control unexposed mouse retina, 2) control unexposed mouse EC, 3) 3-week SHS-exposed mouse retina, 4) 3-week SHS-exposed mouse EC, 5) 6-week SHS-exposed mouse retina, and 6) 6-week SHS-exposed mouse EC.
[0244] The signals from the original 38,083 hybridized oligonucleotide spots were converted to log 2-transformed probe intensities and subjected to normalization and threshold selection as described in Methods. 23,204 mouse probes with positive signal detection above the threshold were obtained. The 3-week and 6-week SHS-exposed retinas, compared to unexposed control retinas showed differential expression of 2,248 and 1,089 probes, respectively. The 3-week and 6-week SHS-exposed ECs compared to unexposed control ECs showed differential expression in 4,391 and 3,529 probes, respectively.
[0245] Clustering of the 23,204 detectable probes was performed with the Cluster 3.0 program (DeHoon et al. 2004). For the clustering analysis, the data was pre-processed by three consecutive rounds of gene median centering and gene median normalization, and then hierarchically clustered using uncentered correlation as the similarity metric and single linkage as clustering method.
[0246] For gene ontology and pathway analysis, 23,199 of the 23,204 detectable probes had valid locus link identifiers (Entrez Gene), and the data for these probes were analyzed using the GenMAPP software (Gladstone Institute, San Francisco, Calif., to first map to Gene Ontology (GO) and Local MAPP categories. Then gene ontology categories showing significant over-representation of differentially expressed genes were determined by comparison of the relative representation in each functional group of genes associated with probes meeting one of nine differential expression criteria, described below, to the relative representation of genes associated with the full set of 23,204 detectable probes. Significance was determined by permutation of Z scores with correction for multiple comparisons as described in the GenMAPP software manual. The data were also analyzed by the WebGestalt software for a statistics-based pathway analysis in order to determine the distribution of differentially-regulated genes among functional biological pathways. The WebGestalt software was used to query two pathway databases including KEGG and Biocarta. Significance of the individual pathways was determined by the hyper geometric statistical method with criteria, adjusted P<0.05 and more than 2 genes per pathway. The criteria that were applied to the genes to uncover the pathways in which genes were significantly over-represented were: all) all probes, 0) 6 week retina vs control retina, up>2 fold, 1) 6 week retina vs control retina, down>2 fold, 2) 3 week retina vs control retina, up>2 fold, 3) 3 week retina vs control retina, down>2 fold, 4) 6 week eyecup vs control eyecup, up>2 fold, 5) 6 week eyecup vs control eyecup, down>2 fold, 6) 3 week eyecup vs control eyecup, up>2 fold, 7) 3 week eyecup vs control eyecup, down>2 fold, 8) Principal Component (PC) number=3.
[0247] The GenMAPP analysis of the data demonstrated three pathways in which differentially expressed genes linked to Entrez Gene IDs and meeting specific criteria were significantly over-represented. They were the Electron Transport Chain from criterion 5, the Structural Constituent of Eye Lens from criterion 6, and the Pheromone Receptor Activity from criterion 8. The WebGestalt analysis of the data revealed KEGG and Biocarta pathways that were enriched for genes fulfilling each of the nine criteria.
[0248] One pathway, Circadian Rhythm, enriched for genes correlated to criteria 8 (PC=3), caught our attention, since an important gene in this pathway, casein kinase 1 epsilon (CK1ε), was previously shown in our laboratory to be increased in the retina of AMD patients in a grade-dependent manner. Although CK1ε was not among the five genes originally selected by the software, a specific check on this gene in the raw profiling data showed a moderate increase (˜1.5×) in this gene in 3 and 6 week smoke-exposed retina, warranting further investigation.
[0249] As mentioned above, in order to confirm the increase in expression of CK1ε which was observed in AMD retina by the earlier Northern blot analysis (FIG. 16), normal human retina and AMD retina was first analyzed for CK1ε expression by real time qPCR. The results demonstrated a correlative increase in CK1ε expression with increasing grade of AMD pathology from Grade 1 to Grade 3 in the retina, with a fall-off of activity in Grade 4 (FIG. 17A). The same analysis for the normal human and AMD RPE/choroid (eyecup, EC) samples yielded the same result, confirming the increase in expression of this gene in AMD in both the retina and EC (FIG. 17B). Importantly, the pattern of variation in the expression seen by real time qPCR, including the dip in Grade 4, was identical to that observed by earlier Northern analysis.
[0250] Analysis of CK1ε expression by real time qPCR in the retina of mice exposed to smoke for 0 to 6 weeks showed an increase of >2 fold starting at 1 week and 5-6 fold by 3 to 6 weeks, confirming and expanding the results of the expression profiling (FIG. 18A). The same analysis for the eyecup (EC) also revealed a delayed increase of ˜4 fold in the expression of CK1ε at 5 weeks, attesting to the stimulatory effect of tobacco smoke on the expression of this gene (FIG. 18B).
[0251] The laser photocoagulation model to induce choroidal neovascularization (CNV) in the mouse retina is a well-established animal model of wet AMD (Espinosa-Heidmann et al. 2002). To further investigate the link between the increase in CK1ε expression and AMD, the expression of this gene was analyzed in the retina and EC of mice subjected to CNV induction by laser photocoagulation.
[0252] The results showed that a steady increase in CK1ε expression was observed in the retina during the 14-day period of CNV maturation (FIG. 19A), whereas a more variable pattern of increase (up, down, and up) was observed in the EC (FIG. 19B), consistent with the increase in this gene observed in AMD patients.
[0253] Finally, because CK1ε is an important regulator of circadian rhythm, its diurnal pattern of expression was examined to determine whether the expression of CK1ε itself is diurnally regulated in the retina and RPE/choroid of the eye. This is particularly a relevant question since the photoreceptors and RPE cells are involved in the critically important process of diurnally regulated daily shedding and phagocytosis of the outer tips of photoreceptor outer segments, which begins just before light onset.
[0254] The results showed that expression of CK1ε in the retina and EC in 12L:12D entrained mice is indeed diurnally regulated, with a peak of expression at 6 PM in the retina (FIG. 20A, Control) and at 3 AM in the EC (FIG. 20B, Control).
[0255] Significantly, when the diurnal pattern of CK1ε expression was examined in the group of mice exposed to smoke for 3 weeks, it was seen that the peaks were phase-shifted. The main peak in the retina was shifted from 6 PM to 9 PM (a 3-hour delay compared with control) with a second, smaller peak appearing at 3 AM (FIG. 20A). Unexpectedly, in the eyecup (EC), the peak was shifted from 3 AM to 12 PM, (a 9 hour delay compared with control) (FIG. 20B).
[0256] The observation of dramatic shifts in the time of peak expression of CK1ε in the retina and EC in mice exposed to cigarette smoke, and especially the differential nature of the smoking-induced shifts in CK1ε expression in the two tissues (3 hour delay in retina vs. 9 hour delay in EC) brings up the distinct and intriguing possibility that smoking may cause an uncoupling of normally coordinated patterns of diurnally-regulated gene expression that go on in the cells of retina and the eyecup that participate in the rhythmic joint process of retinal phagocytosis (see, for example, coordinated patterns of expression of phagocytosis-related genes shown in FIG. 4). Given the fact that the photoreceptors and the RPE cells are involved in a tightly-regulated collaborative function that follows a strict circadian rhythm (synchronous shedding of OS by the photoreceptors, and concomitant ingestion and digestion within several hours of masses of shed OS by the RPE cells), it would be expected that the expression of a large number of shedding-related genes in the photoreceptors would be coordinated with the expression of a large number of genes in the RPE cells that are required for the multiple steps involved in phagocytosis and digestion of the load of OS delivered each day to the RPE cells. For example, it would be expected that lysosomal enzymes involved in the digestion of the membranous material in the phagosomes would be in peak amounts in the RPE cells around the time of light onset, in anticipation of the requirement to break down and recycle components of the OS in the large numbers of phagosomes ingested by the cells at this time.
[0257] Accordingly, without intending to be bound by any particular theory, it would be expected that a phase shift such as the one discovered in the smoking experiments described herein, in which the circadian clock in the retina and the circadian clock in the EC appear to be "uncoupled," could have profound consequences. The results of the smoke exposure experiments indicate that circadian rhythms in the retina and the eyecup are out of synchrony with one another, relative to the respective controls, by a period of about 6 hours (as indicated by the changed peaks in expression of CK1ε, a major regulator of the circadian clock mechanism which controls diurnal expression of many genes). This uncoupling could have significant metabolic consequences for a demanding process such as the daily phagocytosis of OS by the RPE cells, and could result in additional stress on these hard-working caretaker cells if they were required to phagocytize OS at a time of day when they were not optimally primed to do so. Over the long term, such a dysregulation between circadian gene expression in the photoreceptors and circadian gene expression in the cells of the RPE/chorid, day in and day out for many years of adult life, would reasonably be expected to result in a condition such as AMD, in which undigested OS material accumulates in the RPE and is deposited first as lipofuschin and then as drusen in Bruch's membrane.
Example 7
Animal Models of AMD That Over-express Genes Upregulated in AMD
[0258] Studies of the pathogenesis of AMD are impeded by a lack of appropriate and practical animal models useful, for example, for studies designed to elucidate the underlying pathophysiological mechanisms leading to the AMD disease phenotype, and for in vivo testing of candidate therapeutic compounds and approaches. This example describes the construction of animal models of AMD in mice that over-express genes demonstrated herein to be upregulated in AMD. In preferred embodiments, the over-expressed genes are prostaglandin D2 synthase (PD2S), CK1ε, MT1-MMP, and AMDP-3, comprising respective cDNA sequences identified herein as SEQ ID NOS:2, 9, 15, and 17. In some embodiments, the genes are conditionally over-expressed, and in some versions, only in photoreceptors, RPE cells, and/or choroidal cells of the animals.
[0259] As described in examples above, over-expression or over-activity of MT1-MMP is observed in human and monkey eyes with AMD and in RCS rats with an RPE-based inherited retinal degeneration. To model the over-expression phenotype in a small laboratory rodent such as a mouse, transgenic mice over-expressing, for example, MT1-MMP are constructed. A particularly preferred embodiment is a transgenic mouse model featuring conditional over-expression of MT1-MMP in the fully developed, and aged retinas of these animals, which advantageously avoids deleterious effects that could result from over-expression of MT1-MMP during the embryonic or early postnatal stages of development.
[0260] For constructing an animal model that conditionally over-expresses MT1-MMP, a conditional expression system can be used, such as the Tet Gene Expression System (BD Biosciences, Palo Alto, Calif.). Over-expression of a transgene 1000-fold or more within hours of activation with doxycycline has been reported using this system (Gossen et al., 1995). Conditional expression systems are advantageous for temporal control of gene expression, such as the over-expression of MT1-MMP, to cause the expression of the MT1-MMP transgene to begin at a selected time in the life of the animal, for example only in adults with a fully developed retina.
[0261] Transgenic mice are constructed, using techniques well known to those of skill in the art, that over-express, for example, a human or a mouse MT1-MMP. Any suitable overexpression system can be used. In embodiments using the Tet system, a transgenic mouse is constructed that expresses a chimeric tetracycline-regulated transactivator rtTA (Tet-On) from a suitable promoter and a second transgene containing, for example, a human or mouse MT1-MMP cDNA connected to a Tet Response Element-silent promoter which responds to the transactivator. Administration of doxycycline to a double transgenic mouse thus constructed results in overexpression of the transgene, for example, MT1-MMP, through activation of the transactivator by doxycycline, and subsequent binding and activation of the silent promoter.
[0262] In some embodiments of transgenic mice overexpressing genes of interest such as PD2S, MT1-MMP, CK1ε and AMDP-3, expression of the transgene is limited to selected cell or tissue types. As is well known in the art of molecular biology, the cellular site of transgene expression can be controlled by selection of tissue- or cell-specific promoters. Accordingly, in one preferred embodiment, a transgenic model overexpresses a MT1-MMP transgene in a photoreceptor-specific manner. An exemplary promoter for this purpose is a bovine rhodopsin promoter (Zack et al., 1991), shown, for example, to be suitable for photoreceptor-specific expression of HRG4 (UNC119), in a transgenic mouse model (Kobayashi et al., 2000). Other embodiments of the transgenic mice selectively overexpress transgenes, such as MT1-MMP, CK1ε, PD2S, or AMDP-3, in RPE cells. RPE cell-specific expression is directed, for example, by an RPE-specific promoter such as one derived from promoter regions of the genes encoding RPE65 (Boulanger et al., 2000) or cellular retinaldehyde binding protein (Kennedy et al., 1998). Yet other embodiments are engineered to selectively express the transgenes in cell types of the choroid, for example in endothelial cells using an endothelial cell specific promoter (Cho et al., 2003), or in melanocytes and RPE cells using a promoter that drives expression of tyrosinase in pigmented cell types (Giraldo et al., 2003).
[0263] Transgenic mice are constructed by oocyte injection of a transgene-containing vector using techniques well known to those of skill in the art of molecular biology. (See, for example, Kobayashi et al., 2000). The over-expression of the selected transgene is confirmed in the appropriate tissues or cells of the transgenic animals (for example in the retina, or specifically in photoreceptors or RPE cells, or in one or more choroidal cell types) using techniques well known in the art and demonstrated in examples above, such as by Northern analysis or RT-PCR using appropriate probes or primers specific for the transgene, by Western blot analysis of proteins with an appropriate antibody, and by various immunolocalization techniques.
[0264] Pathology developing in the transgenic animals, for example in the retinas and/or RPE/choroid of these animals, is assessed by numerous known techniques, including, for example, examination of the retina by funduscopy, electroretinographic (ERG) testing, and light and electron microscopy at selected intervals throughout the lifetime of the animals, before and after activation of the transgene by administration of doxycycline, for example at 5, 10, 15, 20, and 30 days of age, (with administration of doxycline at age 30 days), and at 1, 2, 5, 10, 20, 30, 60, 80, 100, 120, 140, 160, 180, 200, 220, 240, 260, 280, 300, 320, 340, 360, 380, 400, 420, 440, 460, 480, 500, 520, 540, 560, 580, 600, 620, 640, 660, 680 and 700 days after activation with doxycycline. AMD-related pathology, such as lipofuscin accumulation, Bruch's membrane thickening, basal laminar and linear deposits, drusen formation, neovascularization, CNV membrane formation, photoreceptor/RPE atrophy or choroidal atrophy is monitored by standard techniques well known in the art.
Example 8
Animal Models of AMD That Express Polymorphic Variants of Phagocytosis-Related and/or AMD-Related Genes
[0265] This example describes the construction of mouse models of AMD and other retinal degenerations that express one or more polymorphic variants of a phagocytosis-related or AMD-related gene.
[0266] As shown above, certain polymorphic variants of genes, including MT1-MMP, are found at higher frequency in the DNA of patients with AMD. To model the human conditions, transgenic mouse models expressing polymorphic and wild-type human genes, for example MT1-MMP, are constructed as follows. First, the baseline status of the mouse MT1-MMP gene is preferably determined. For example, it has been determined that the wild-type amino acid residue located at the position of the D273N polymorphism in the human MT1-MMP DNA sequence is conserved in the human and mouse. A polymorphism at this residue has not been demonstrated in the mouse (Mouse Genome Project). Presence of the wild type residue is confirmed in the mice used for transgenic construction, by tail biopsy, DNA isolation, and genotyping.
[0267] To construct polymorphic and control (wild type) transgenic mouse models, expressing respectively, polymorphic and wild type variants of a human gene of interest, such as MT1-MMP, cDNAs containing human polymorphic variants and wild-type MT1-MMP residues are connected to a promoter sequence suitable for driving expression of the transgene in a desired tissue or cell. For expression of the transgene throughout the body, an exemplary promoter sequence is, for example, a 385 bp human MT1-MMP promoter sequence, prepared by PCR amplification from human genomic DNA and previously determined to drive robust expression of the gene (Lohi et al., 2000). To aid identification of the transgene, in some embodiments the MT1-MMP gene is expressed as a green fluorescent protein (GFP) fusion protein, using a suitable vector construct, such as a BioSignal vector (InVitrogen, Carlsbad, Calif.). Other embodiments selectively express the transgene in particular tissues or cell types, driven by tissue- or cell type-specific promoters as described above.
[0268] Transgenic mice are constructed by oocyte injection of the vector using known techniques. Expression of the human polymorphic and wild type variants, for example of MT1-MMP, is confirmed in the transgenics, such as by RT-PCR with allele-specific primers and, in versions expressing GFP fusion proteins, by analysis of GFP expression, for example by fluorescence microscopy, Western blotting analysis, or immunodetection. The transgenics are analyzed for the presence of AMD-related pathologies as described in Example 7 above.
[0269] Other embodiments of the animal models of AMD and other retinal degenerations are polytransgenic mice expressing polymorphic variants of at least two genes having a known association with AMD. In preferred embodiments, the animals express a polymorphic variant of MT1-MMP in combination with at least one other polymorphic gene variant showing a correlation with phagocytosis and/or AMD.
[0270] The polytransgenic versions of the animal models are based on the complex, multi-gene theory of AMD, which assumes that subtle mutations in a number of genes, commonly referred to as "polymorphisms," cooperate to cause, or create a susceptibility to, a disease. Accordingly, the full phenotype of AMD is likely to require the cooperation of at least two, and perhaps many, etiologic genes with the appropriate combination of polymorphisms. The causative genes may tip the scale toward development of AMD by contributing either "collectively" (for example, if related by a common function, such as involvement in the pathway of OS phagocytosis), or "cumulatively," for example, if unrelated by function, but each involved in a separate aspect of the pathogenic process leading to AMD.
[0271] A preferred embodiment of a polytransgenic model of AMD is a polytransgenic animal that co-expresses a first polymorphic variant of MT1-MMP and at least a second polymorphic variant of at least one other phagocytosis-related and/or AMD-related gene. Any other second or more gene showing a polymorphic variant correlated with AMD can be combined with any polymorphic variant of MT1-MMP. Genes presently reported to have variants correlated with AMD are listed in 3.
TABLE-US-00005 TABLE 3 Genes with Reported Polymorphisms or Mutations Correlated with AMD NUCLEIC AMINO ACID SEQ ACID SEQ GENE ID NO: ID NO: REFERENCE ABCR 62 124 Allikmets et al., 1997 Apolipoprotein 63 125 Klaver et al. 1998; E Simonelli et al. 2001 C-C chemokine 64 126 Ambati et al. 2003 receptor-2 Cystatin C 65 127 Zurdel et al. 2002 Hemicentin/ 66 128 Schultz et al. 2003 FIBL-6 Manganese 67 129 Kimura et al. 2000 superoxide dismutase C-C chemokine 68 130 Ambati et al. 2003 ligand/monocyte chemoattractant protein 1 Paraoxonase 69 131 Ikeda et al. 2001
[0272] Accordingly, in one form of the preferred embodiments, a polymorphic form of MT1-MMP is combined with a polymorphic form of at least one other gene, including ABCR, apolipoprotein E, C--C chemokine receptor-2, cystatin C, hemicentin/FIBL-6, manganese superoxide dismutase, C--C chemokine ligand/monocyte chemoattractant protein 1, and paraoxonase.
[0273] Similarly, a polytransgenic model reflecting the "collective" etiology theory of AMD combines polymorphic variants of genes with known involvement in the mechanism of an important function (for example OS phagocytosis) with polymorphic variants of MT1-MMP (a demonstrated phagocytosis-related gene as disclosed herein; wild type cDNA sequence: SEQ ID NO:15; wild type amino acid sequence: SEQ ID NO:100). Such genes include, for example, polymorphic variants of phagocytosis-related genes PHG-1 to PHG-15 (SEQ ID NOS:1-14) and AMDP-2 and 3 (SEQ ID NOS:16 and 17), disclosed herein (see Tables 1 and 2, supra).
[0274] For construction of the models, DNA containing the reported polymorphic variant(s) of a selected gene is first isolated using appropriate amplimers from DNA of patients with AMD and unaffected, age-matched individuals (for example, as described for MT1-MMP in Example 5 above), and is used to confirm the presence of the reported polymorphisms, for example, in ABCR (i.e., D2177N, G1961E); manganese superoxide dismutase (i.e., V47A); apolipoprotein E (i.e., epsilon2); cystatin C (i.e., A and B allele, including the Ala to Thr change); and paraoxonase (i.e., Q192R, L54M). Genotyping and mutational analyses are carried out using established methods (see for example, Mashima et al., 1994). The association of the polymorphism with AMD is confirmed and the statistical significance of any detected correlations with AMD is determined, for example by a chi-square test. For those polymorphic genes showing an association with AMD, their co-occurrence with a polymorphism in MT1-MMP is then confirmed.
[0275] Transgenic mice expressing a polymorphic variant of a selected gene, for example AMDP-3, are first constructed as generally described above. To construct polytransgenic models, transgenic mice expressing a polymorphic variant of the first gene of interest, for example, MT1-MMP, are crossed with transgenic mice expressing a polymorphic variant of a second phagocytosis/AMD-related gene of interest, such as AMDP-3. Expression of the various transgenes is confirmed in tissues of interest, for example the retina, RPE or choroid, by standard techniques known in the art, such as allele-specific RT-PCR of RNA and/or immunodetection of the polymorphic transgene protein of interest, for example by using antibodies specific for a particular polymorphic form of the protein. Alternatively, in embodiments in which a specific tag protein sequence is attached to the transgene protein, identification of the tag sequence is used to facilitate identification of the transgenic polymorphic variant protein and to distinguish it from the wild-type form. The polytransgenic mouse is analyzed for evidence of AMD-related changes as described above.
Example 9
Construction of Transgenic Conditional Over-Expression Mouse Models of AMD Based on MT1-MMP and CK1ε
[0276] This example further expands upon the description provided above, in Example 7, of how to make and use a nonhuman transgenic mouse in which overexpression of a transgene of interest is induced by administration of an inducer molecule such as doxycycline, which can be administered by several routes, including intraperitoneal or intraocular injection, or by inclusion in the drinking water of the animals.
[0277] The two specific embodiments described below were constructed based on the initial finding, by the CHANGE array hybridization strategy, of over-expression of two phagocytosis-related genes, i.e., MT1-MMP and CK1ε in human eyebank samples, with subsequent confirmation of the over-expression by Northern analysis and quantitative PCR, in mRNA from human AMD samples and mRNA from various rodent models of AMD, as described above.
[0278] 1. Construction of a Conditional MT1-MMP Over-Expression Transgenic Mouse
[0279] Transgenic mice were constructed to conditionally over-express human MT1-MMP using the techniques generally described in Example 7 above. Briefly, the Tet-On Inducible Over-expression system (BD, Palo Alto, Calif.) was used to construct a double transgenic mouse that expresses a modified tetracycline-regulated transactivator rtTA molecule (Tet-On) from a human MT1-MMP promoter (in the place of the CMV promoter in the pTet-On vector provided by the manufacturer), and a second transgene (pTRE2) containing a human MT1-MMP cDNA connected to a modified CMV promoter responsive to the doxycycline-bound transactivator. Conditional over-expression of MT1-MMP transgene is induced upon administration of doxycycline to the double transgenic mouse, through activation of the transactivator by doxycycline and subsequent binding and activation of the human MT1-MMP promoter. Expression of the transgene occurs in all tissues of the mice that naturally express MT1-MMP.
[0280] As further described below, induction of this single gene, i.e., MT1-MMP in mice with mature normal retinas (one month and older) results in a series of dramatic phenotypic changes that exhibit many of the pathological features of both the wet and dry forms of AMD. Thus, inter alia, this inducible over-expression transgenic model based on MT1-MMP provides a striking proof-of-concept for the entire CHANGE strategy for discovery of genes involved in the pathogenesis of AMD, in that MT1-MMP, which was initially discovered by CHANGE to be over-expressed in human eyes with AMD, indeed can be shown in the transgenic model to cause AMD-like pathology when this gene is inducibly over-expressed. Similarly, inducible transgenic models designed to over-express other genes found by the CHANGE strategy to be over-expressed in human eyes with AMD (i.e., CK1ε, prostaglandin D2 synthase, and AMDP-3) are expected to be very useful as animal models of AMD.
[0281] 2. Construction of a Conditional CK1ε Over-Expression Transgenic Mouse
[0282] It is shown in Example 8 above that CK1ε is over-expressed in human eyes with AMD, using real-time quantitative PCR, confirming the initial CHANGE and Northern blot data. Example 8 also describes results demonstrating that this gene is also upregulated in the laser photocoagulation model of AMD which features choroidal neovascualarization (CNV), the hallmark of the wet form of AMD. It is further demonstrated in Example 8 that the best known environmental factor associated with AMD, i.e., cigarette smoking, causes an increase in expression of CK1ε, in addition to phase shifting of the time of day for peaks in expression of this gene in the retina and in the RPE/choroid (eyecup). Most significantly, the duration of the smoking-induced phase shift is different in the retina than in the EC. Thus, the effect of cigarette smoking appears to be an uncoupling of the circadian clocks that control diurnal gene expression in the two tissues.
[0283] Thus, the various studies described herein using human and animal tissues provide abundant support for the likelihood that CK1ε, which we have shown to be upregulated in AMD, provides an excellent candidate as a therapeutic target for AMD. Accordingly, an inducible over-expression transgenic mouse that can conditionally over-express this gene is anticipated to be very informative for the study of the biological and molecular consequences of phase shifting the peaks of expression of CK1ε, such as occurs in smoking. Some preferred embodiments of the inducible CK1ε transgenic models may be based on an expression system that utilizes cloxacillin as the inducer molecule.
[0284] Transgenic mouse models that inducibly express CK1ε are expected to be very useful for the study of pathological features of the wet and dry forms of AMD associated with smoking, and as models for testing the efficacy of candidate therapeutic compounds for these conditions. For these reasons, we constructed a second inducible over-expression transgenic model, based on the CK1ε gene, as generally described in Example 7 above, and more particularly as described below.
[0285] Methods.
[0286] The Tet-On Inducible Gene Expression System vector, pTRE-Tight-BI (Clontech, Mountain View, Calif.) was used to construct an inducible over-expression vector of mouse CK1ε. Two 32-base primers, one ("Start") containing a Pvu II site and homologous to residues 114 to 145 of the mouse Ck1ε mRNA sequence (GenBank NM--013767.6), and the other ("Stop") containing a Hind III site and homologous to residues 1400 to 1432 of the mRNA sequence, were used to amplify up a 1319 bp cDNA using mouse eyecup (EC) cDNA as template by PCR. Primers used for this purpose are as shown:
TABLE-US-00006 Start: (SEQ ID NO: 136) 5'-ATTATCCAGCTGGCGCGCAAGGTTGAAGCATG-3' Stop: (SEQ ID NO: 137) 5'-TGAAGAAAGCTTGCAAACACTGGTCTGTGGTC-3'
[0287] The 1319 base pair PCR product was purified, digested with Pvu II and Hind III, purified again, and confirmed by DNA sequencing to be the correctly-digested mouse Ck1ε cDNA containing the translational initiation and termination codons. The cDNA was ligated into the Pvu II and Hind III sites of the MCS I region of the pTRE-Tight-BI vector, positioning it correctly with respect to the mini CMV promoters present in the vector for expression. This vector is shown schematically in FIG. 21.
[0288] The purified DNA was microinjected into fertilized mouse eggs, followed by transplantation into pregnant mice to generate the transgenics. Further procedures in production of the transgenic mice are generally as described above. The Tet-On system requires the presence of a second vector that expresses the transactivator, which when bound by doxycycline, interacts with the pTRE vector to induce the over-expression of the gene cloned downstream of the mini CMV promoters. To achieve this, mice positive for pTRE-TBI-mCK18 are crossed with transgenic mice containing the Rosa26 promoter-driven pTet-on vector, which expresses the transactivator ubiquitously in all tissues (Hochedlinger et al., 2005). Pups doubly positive for the pTRE-TBI-mCK1ε and the pTet-on-Rosa26 are identified among the progeny by tail biopsies and DNA screening. The double-positive transgenics respond to doxycycline administration by over-expressing CK1ε in all tissues, including retina and EC, the target tissues relevant to AMD.
Example 10
Inducible MT1-MMP Over-Expression Transgenic Mouse Model Exhibits Pathophysiological Features of Wet and Dry Forms of AMD
[0289] Several lines of inducible MT1-MMP over-expression transgenic mice were constructed as described in Example 9. Mice were raised under a 12 hour Light:Dark lighting cycle and were used for experiments starting at approximately one month of age, when the retina is mature. To induce over-expression of the MT1-MMP transgene, the inducer (doxycycline, "Dox") was administered daily to the test group, either by daily addition in the drinking water, or by daily intraperitoneal injection. Groups of Dox-treated mice, as well as age-matched untreated controls and Dox-treated control mice were sacrificed at various times from 1 to 60 days after Dox administration, and the eyes were analyzed in H& E stained paraffin sections for evidence of histopathological features of AMD.
[0290] The results showed that a dramatic phenotype, exhibiting many pathological features seen in human AMD, was rapidly induced, i.e., within one week of Dox administration, in the eyes of the Dox-treated transgenic mice groups, but not in groups of untreated or Dox-treated control mice. Features of the observed pathology, at various stages, are shown in FIGS. 22A-F. One of the earliest pathological features to develop in the Dox-treated transgenic eyes was the presence of large vacuoles within the RPE layer that protruded into the layer of photoreceptor outer segments, in some cases appearing as a series of regularly-spaced bubbles within the outer segment zone. Some of these bubbles were either partially filled with, or outlined by, pigment granules, and as such appeared to represent grossly swollen and degenerating RPE cells (FIG. 22A).
[0291] The intercellular junctions between the RPE cells (which in the normal retina are connected at their apical surfaces by tight junctions) are not usually visible in paraffin sections of pigmented mouse eyes, which feature a heavily pigmented RPE cell layer. However, in the Dox-treated transgenic mice, the junctions between the RPE cells in areas of pathology appeared much more prominent than in control eyes. This difference was especially appreciable in tangential sections, in which the junctions between the RPE cells could be visualized as clear gaps outlining the pigmented cells in the hexagonal RPE cell monolayer (FIG. 22B). Some of these spaces contained red blood cells that appeared to originate in the choroid. In the usual vertical sections through the retina, areas of pathology were seen in which red blood cells were pooled in the subretinal space, on the surface of the RPE cells (FIGS. 22C, D). In some such areas, abnormal new blood vessels were seen sprouting from the RPE layer, often in close proximity to a region of vacuolated RPE, and growing inward toward the vitreal surface of the retina (FIGS. 22C, D). In some instances, the abnormal new vessels coursed through the full thickness of the retina, and were associated with a mass of cellular debris deposited on the surface of the retina in the vitreous (FIG. 22E). In areas of pathology, the normally highly structured retinal tissue often appeared disorganized, presumably due to breakdown of matrix proteins in the extracellular matrix by the over-expressed MT1-MMP, a condition that may have facilitated the growth of the new vessels from the choroid right through the thickness of the retina, breaking through on the vitreal surface.
[0292] In areas of very advanced pathology, lesions were seen that bore a close resemblance to classic choroidal neovacular (CNV) membranes, which are the hallmark pathological feature seen in sections of human eyes with the advanced "wet" form of AMD (FIG. 22F).
[0293] Based on observations made in large numbers of transgenic animals from this inducible MT1-MMP over-expression model at various times after Dox administration, a classification system for AMD-like pathology was developed as follows:
[0294] Grade 1: vacuolar RPE degeneration
[0295] Grade 2: RPE migration and retinal structural disorganization
[0296] Grade 3: early blood vessel (tube) formation/migration
[0297] Grade 4: overt CNV
[0298] This model is useful inter alia for testing the efficacy of candidate drugs for the treatment of the wet and dry forms of AMD, in that it exhibits pathological features of both forms of AMD. It is remarkable that such an extensive pathological phenotype can be produced by over-expressing a single gene that was revealed by our CHANGE strategy, i.e., MT1-MMP. This finding suggests that this gene must be intimately involved in the pathogenesis of AMD, making it an excellent target for therapy for AMD.
Example 12
Delay of Retinal Degeneration by an Antibody that Binds MT1-MMP Polypeptides
[0299] This example describes studies demonstrating slowing of the rate of an inherited retinal degeneration in an animal (rat) model, using an antibody agent that neutralizes MT1-MMP protein. As described in Example 4 above, MT1-MMP was found to be overexpressed in human eyes with AMD, in a monkey model of AMD, and in the RCS rat, an animal model of an RPE-based inherited retinal degeneration. The mutant phenotype in the RCS rat, due to a mutation in the MERTK gene, is characterized by a defect in the ingestion phase of phagocytosis by the RPE cells. In separate studies, MT1-MMP was again isolated in a CHANGE analysis wherein +/- probes were prepared from retinal RNA of mutant and age-matched control RCS rats. Northern blot analysis of MT1-MMP expression in the RCS rat retina revealed that expression of MT1-MMP mRNA increased as the retinal degeneration progressed in this model. This result suggests that MT1-MMP may play a common role in the pathogenesis of multiple forms of retinal degeneration, particularly those based on a defect thought to affect primarily the RPE cells.
[0300] To test the functional involvement of MT1-MMP in the pathogenesis of the retinal degeneration in the RCS rat, a 2 μl volume, (as supplied by the manufacturer), of an antibody against MT1-MMP (Chemicon, Temecula, Calif.), was injected subretinally into the eyes of immature (7 day) RCS rats. The course of the retinal degeneration was followed for the subsequent two months. Referring to FIG. 15, the results showed a remarkable delay of up to a 50% in the retinal degeneration, as determined by the thickness of the outer nuclear layer, observed at 1 month post-injection. Sham injection, or injection of an unrelated (i.e., X-arrestin) antibody did not produce this effect. This result further reinforces the involvement of MT1-MMP in the pathogenesis of retinal degenerations, making it an attractive therapeutic target for retinal degenerative conditions involving over-expression of MT1-MMP.
Example 13
Reduction of AMD-like Pathology in Transgenic Mouse Model of MT1-MMP Over-Expression by RNAi Agents (siRNA) that Inhibit MT1-MMP
[0301] Methods.
[0302] Oligonucleotides. Double-stranded small inhibitory RNA (siRNA) (MT1-MMP inhibitory, designated "A") and unrelated RNA (control siRNA, designated "B") sequences were designed as shown, based on the human MT1-MMP gene:
TABLE-US-00007 Inhibitory MT1-MMP siRNA (Oligonucleotide A): (SEQ ID NO: 138) CCUCUCCACGCGCAGUACATT (SEQ ID NO: 139) UGUACUGCGCGUGGAGAGGTT Control siRNA (Oligonucleotide B): (SEQ ID NO: 140) CCGACAUCAUGAUCUUCUUTT (SEQ ID NO: 141) AAGAAGAUCAUGAUGUCGGTT
[0303] Animals. Groups of transgenic mice from the transgenic mouse model of MT1-MMP over-expression described above in Example 11 were entrained to a 12L:12 D lighting cycle from birth, and used at approximately one month of age. In a blind study, on Day 1, groups of animals, 3-4 per group, received an intraocular injection in one eye of either Oligonucleotide A, Oligonucleotide B (2 μl volume, 1 μg/μl, 200 μM), or saline solution. Starting on Day 1, all mice received daily administration of doxycycline, to induce over-expression of MT1-MMP in the animals, with concomitant AMD-like pathological features in the eye, as described above. On Day 8, the mice were euthanized and their eyes were processed for histological assessment of the observed degree of AMD-like pathology, using the grading criteria described in Example 11.
[0304] Results:
[0305] In 10 experiments performed with ˜40 transgenic mice, intraocular injection of siRNA directed against MT1-MMP (Oligonucleotide A) was shown to statistically significantly ameliorate the pathology (by ˜50%), with the average grade of pathology reduced to 2.2 vs. 4.0 in the control siRNA (Oligonucleotide B) and saline-treated groups (p=0.01).
[0306] These results provide an important independent confirmation that the pathology observed in the transgenic mouse model of inducible MT1-MMP over-expression described herein is indeed caused by over-expression of this gene. Most importantly, the results demonstrate the efficacy of a small inhibitory RNA (siRNA) therapeutic strategy based on MT-MMP to ameliorate AMD-like symptoms in a mouse model of AMD.
Example 14
Superior Reduction of Choroidal Neovascularization (CNV) in Mouse Laser Photocoagulation Model Using Antibody that Inhibits MT1-MMP vs. Antibody that Inhibits Vascular Endothelial Growth Factor (VEGF)
[0307] Background.
[0308] At present, the state-of-the-art treatment for the devastating neovascularization that occurs in the wet form of AMD is to administer by intravitreal injection an antibody (bevacizumab) directed against vascular endothelial growth factor (VEGF). This treatment has been well received around the world and represents a great step forward in the treatment of this blinding form of AMD. As the use of bevacizumab (or Avastin, another antibody based on VEGF) for treatment of AMD has continued, however, in recent years there have been reports of complications in some patients, as well as results from long-term studies indicating that not all patients respond well to anti-VEGF therapy, and that its efficacy can be lost with prolonged treatment. Thus, there is an ongoing unmet need for new therapies to augment or replace the VEGF-based strategy to control the neovascularization in wet AMD.
[0309] As described in Example 12 above, we have shown that an antibody directed against MT1-MMP is very effective in reducing the rate of retinal degeneration in the RCS rat, a rodent model having a defect in the MERTK gene, which is characterized by a functional defect in phagocytosis by the RPE cells, and in which we have determined that MT1-MMP is over-expressed. We have also shown, in Example 13, that MT1-MMP, when conditionally over-expressed in an inducible transgenic mouse model, results in striking AMD-like pathology after induction of this gene. Thus, we believe MT1-MMP to be a very important therapeutic target for the design of new drugs to treat AMD and other disorders affecting the RPE.
[0310] This Example describes the results of studies undertaken to compare the efficacy of an antibody directed against MT1-MMP with that of an antibody directed against VEGF. These studies were carried out using the well-established laser photocoagulation mouse model of wet AMD, discussed above, in which precisely-spaced discrete laser burns are delivered to the retina in order to induce choroidal neovascularization (CNV) (Espinosa-Heidmann et al. 2002).
[0311] Methods.
[0312] Normal mice were treated with laser (four burn sites per retina) following standard published procedures, and on the same day injected intraocularly with either an anti-MT1-MMP neutralizing antibody directed against the catalytic domain of MT1-MMP (Anti-MMP-14, catalytic domain, clone LEM-2/15.8, Chemicon, Temecula, Calif./Millipore), or a mouse anti-VEGF antibody (Mouse VEGF164 Antibody, affinity purified polyclonal goat IgG, Cat. No. AF-493-NA, R&D Systems). Two weeks later, the animals were euthanized, and the eyes were enucleated and processed in flatmount preparations for analysis by fluorescence microscopy and digital morphometry. For assessment of pathology, both the size and fluorescence of the CNV lesions were measured, to determine the CNV index ratios (with untreated control lesions being normalized to a value of 1.0). Fourteen separate experiments were performed.
[0313] Results.
[0314] The results of these experiments are shown in FIG. 23A-C and Table 4. Referring to FIG. 23A, a typical flatmount preparation of mouse retina after laser photocoagulation is shown. The arrows in the figure on the left indicate four laser-treated spots situated around the optic disc shortly after laser treatment. The photograph on the right (FIG. 23 B) is a higher magnification image showing a typical choroidal neovascularization (CNV) lesion that develops after approximately two weeks. The CNV lesions appears fluorescent due to labeling with fluorecein dextran, which is administered systemically immediately prior to sacrificing the animals, due to its selective binding to the leaky new blood vessels in the CNV.
[0315] FIG. 23C is a group of four images showing the results of a representative experiment comparing the effects of anti-MT1-MMP and anti-VEGF antibodies on CNV formation in the mouse laser photocoagulation model. As can be seen, the anti-MT1-MMP antibody showed a far superior ameliorative effect on CNV formation over the anti-VEGF antibody. More particularly, the upper left image in FIG. 23C shows a flatmount preparation of a lasered mouse eye treated with anti-MT1-MMP antibody. There is almost no fluorescence seen over the burn sites, indicating that typical CNV did not form (compare with uninjected eye, bottom left, showing fluorescent CNV lesion). By contrast, the lesions in the treated and untreated eyes essentially looked the same in the groups of animals treated with anti-VEGF antibody (FIG. 23C, upper and lower images on the right).
[0316] A comparison of the effect of the anti-MT1-MMP and anti-VEGF antibody treatments was carried out by digital morphometry in flatmounts from 14 independent experiments using the mouse laser model of CNV. It was determined that the observed effect of the anti-MT1-MMP antibody vs. untreated control eye was highly statistically significant (i.e., CNV index ratio of 0.52 for anti-MT1-MMP antibody (p=0.0000015)). whereas the CNV index ratio of 1.60 for the anti-VEGF antibody showed no difference from control (p=0.07). The difference between the two antibody treatments (i.e., MT1-MMP vs. anti-VEGF) was statistically significant by the paired test (p=0.016).
TABLE-US-00008 TABLE 4 Comparison of CNV ratio in lasered mouse eyes 14 days after induction of CNV following treatment with anti-MT1-MMP or anti- VEGF antibodies Degree of CNV (ratio compared to uninjected = 1) Ratio St Dev p value Anti-MT1-MMP Ab 0.52 0.282 1.4746E-06 Anti-VEGF Ab 1.6 0.777 0.066011402 Control (PBS) 0.83 0.656 0.697376005 Uninjected 1
[0317] These results strongly demonstrate the usefulness of an antibody directed against MT1-MMP as a therapeutic agent for the treatment of the AMD, a result that would in fact be predicted, based on our observation in the inducible transgenic MT1-MMP over-expression mouse model that turning on this single gene can result in a full range of AMD-like symptoms, including neovascularization. Importantly, these results point to the possibility that treatment of the wet form of AMD with an antibody directed against MT1-MMP could be more effective, either alone or in combination with anti-VEGF therapy, than treatment with an antibody directed against VEGF alone.
LITERATURE CITED
[0318] References cited herein are listed below for convenience and are hereby incorporated by reference in their entirety. [0319] Abdelsalam, A., Del Priore, L. & Zarbin, M. A. Drusen in age-related macular degeneration: pathogenesis, natural course, and laser photocoagulation-induced regression. Surv. Ophthalmol. 1999; 44:1-29. [0320] Algvere P V, Seregard S. Age-related maculopathy: pathogenetic features and new treatment modalities. Acta Ophthalmol Scand. 2002 April; 80(2):136-43. [0321] Allikmets R, Shroyer N F, Singh N, Seddon J M, Lewis R A, Bernstein P S, Peiffer A, abriskie N A, Li Y, Hutchinson A, Dean M, Lupski J R, Leppert M. Mutation of the Stargardt disease gene (ABCR) in age-related macular degeneration. Science. 1997 Sep. 19; 277(5333):1805-7. [0322] Ambati J, Anand A, Fernandez S, Sakurai E, Lynn B C, Kuziel W A, Rollins B J and Ambati B K. An animal model of age-related macular degeneration in senescent Cc1-2 or Ccr-2 deficient mice. Nature Med. 2003 Oct. 19 [Epub ahead of print]. [0323] Apte S S, Fukai N, Beier D R, Olsen B R. The matrix metalloproteinase-14 (MMP-14) gene is structurally distinct from other MMP genes and is co-expressed with the TIMP-2 gene during mouse embryogenesis. J Biol. Chem. 1997 Oct. 10; 272(41):25511-7. [0324] Berglin L, Sarman S, van der Ploeg I, Steen B, Ming Y, Itohara S, Seregard S, Kvanta A. Reduced choroidal neovascular membrane formation in matrix metalloproteinase-2-deficient mice. Invest Ophthalmol V is Sci. 2003 January; 44(1):403-8. [0325] Boulanger A, Liu S, Henningsgaard A A, Yu S, Redmond T M. The upstream region of the Rpe65 gene confers retinal pigment epithelium-specific expression in vivo and in vitro and contains critical octamer and E-box binding sites. J Biol. Chem. 2000 Oct. 6; 275(40):31274-82. [0326] Bok, D and Hall M O. The role of the pigment epithelium in the etiology of inherited retinal dystrophy in the rat. J. Cell Biol. 1971 June; 49(3):664-82. [0327] Boyle D, Tien L F, Cooper N G, Shepherd V, McLaughlin B J. A mannose receptor is involved in retinal phagocytosis. Invest Ophthalmol V is Sci. 1991 April; 32(5):1464-70. [0328] Bressler N M, Bressler S B, Fine S L. Age-related macular degeneration. Surv Ophthalmol 1988 May-June; 32(6):375-413 [0329] Bressler N M; Treatment of Age-Related Macular Degeneration with Photodynamic Therapy (TAP) Study Group. Photodynamic therapy of subfoveal choroidal neovascularization in age-related macular degeneration with verteporfin: two-year results of 2 randomized clinical trials-tap report 2. Arch Ophthalmol. 2001 February; 119(2):198-207. [0330] Cao J, Sato H, Takino T, Seiki M. The C-terminal region of membrane type matrix metalloproteinase is a functional transmembrane domain required for pro-gelatinase A activation. J Biol. Chem. 1995 Jan. 13; 270(2):801-5. [0331] Cho J, Lim W, Jang S, Lee Y. Development of an efficient endothelial cell specific vector using promoter and 5' untranslated sequences from the human preproendothelin-1 gene. Exp Mol. Med. 2003 Aug. 31; 35(4):269-74. [0332] Crabb J W, Miyagi M, Gu X, Shadrach K, West K A, Sakaguchi H, Kamei M, Hasan A, Yan L, Rayborn M E, Salomon R G, Hollyfield J G. Drusen proteome analysis: an approach to the etiology of age-related macular degeneration. Proc Natl Acad Sci USA. 2002 Nov. 12; 99(23):14682-7. [0333] D'Cruz P M, Yasumura D, Weir J, Matthes M T, Abderrahim H, LaVail M M, Vollrath D. Mutation of the receptor tyrosine kinase gene Mertk in the retinal dystrophic RCS rat. Hum Mol. Genet. 2000 Mar. 1; 9(4):645-51. [0334] De S, Sakmar T P. Interaction of A2E with model membranes. Implications to the pathogenesis of age-related macular degeneration. J Gen Physiol. 2002 August; 120(2):147-57. [0335] Ding H, Schwarz D S, Keene A, Affar el B, Fenton L, Xia X, Shi Y, Zamore P D, Xu Z. Selective silencing by RNAi of a dominant allele that causes amyotrophic lateral sclerosis. Cell. 2003 August; 2(4):209-17. [0336] Elbashir S M, Lendeckel W, Tuschl T. RNA interference is mediated by 21- and 22-nucleotide RNAs. Genes Dev. 2001 Jan. 15; 15(2):188-200. Evans, J. R. Risk factors for age-related macular degeneration. Prog. Retin. Eye. Res. 20, 227-253, 2001. [0337] Fine S L, Berger J W, Maguire M G, Ho A C. Age-related macular degeneration. N Engl J. Med. 2000 Feb. 17; 342(7):483-92. [0338] Flood V, Smith W, Wang J J, Manzi F, Webb K, Mitchell P. Dietary antioxidant intake and incidence of early age-related maculopathy: the Blue Mountains Eye Study. Ophthalmology. 2002 December; 109(12):2272-8. [0339] Gass J D, Jallow S, Davis B. Adult vitelliform macular detachment occurring in patients with basal laminar drusen. Am J. Ophthalmol. 1985 Apr. 15; 99(4):445-59. [0340] Gossen M, Freundlieb S, Bender G, Muller G, Hillen W, Bujard H. Transcriptional activation by tetracyclines in mammalian cells. Science. 1995 Jun. 23; 268(5218):1766-9. [0341] Giraldo P, Regales L, Lavado A, Tovar V, Garcia-Diaz A, Jimenez E, Montoliu L. IL-22 The mouse tyrosinase gene: structural and functional studies in transgenic mice. Pigment Cell Res. 2003 October; 16(5):582. [0342] Gottlieb J L. Age-related macular degeneration. JAMA 2002 Nov. 13; 288(18):2233-6 [0343] Green W R. Histopathology of age-related macular degeneration. Mol. Vis. 1999 Nov. 3; 5:27. [0344] Grishok A, Tabara H, Mello C C. Genetic requirements for inheritance of RNAi in C. elegans. Science. 2000 Mar. 31; 287(5462):2494-7. [0345] Guymer R. The genetics of age-related macular degeneration. Clin Exp Optom. 2001 July; 84(4):182-189. [0346] Hageman G S, Mullins R F, Russell S R, Johnson L V, Anderson D H. Vitronectin is a constituent of ocular drusen and the vitronectin gene is expressed in human retinal pigmented epithelial cells. FASEB J. 1999 March; 13(3):477-84. [0347] Hageman, G. S., Luthert, P. J., Chong, N. H. V., Johnson, L. V., Anderson, D. H. & Mullins, R. F. An integrated hypothesis that considers drusen as biomarkers of immune-mediated processes at the RPE-Bruch's membrane interface in aging and age-related macular degeneration. Prog. Retinal Eye Res. 2001; 20:705-732. [0348] Heiba I M, Elston R C, Klein B E, Klein R. Sibling correlations and segregation analysis of age-related maculopathy: the Beaver Dam Eye Study. Genet Epidemiol. 1994; 11(1):51-67. [0349] Hogan M J. Role of the retinal pigment epithelium in macular disease. Trans Am Acad Ophthalmol Otolaryngol. 1972 January-February; 76(1):64-80. [0350] Hochedlinger, K, Yamada Y, Beard C, Jaenisch R. Ectopic expression of Oct 4 blocks progenitor-cell differentiation and causes dysplasia in epithelial tissues. 2005 Cell 121(3):465-77. [0351] Husain D, Ambati B, Adamis A P, Miller J W. Mechanisms of age-related macular degeneration. Ophthalmol Clin North Am. 2002 March; 15(1):87-91. [0352] Hutchinson A, Dean M, Lupski J R, Leppert M. Mutation of the Stargardt disease gene (ABCR) in age-related macular degeneration. Science. 1997 Sep. 19; 277(5333):1805-7. [0353] Hyman L, Neborsky R. Risk factors for age-related macular degeneration: an update. Curr Opin Ophthalmol. 2002 June; 13(3):171-5. [0354] Ikeda T, Obayashi H, Hasegawa G, Nakamura N, Yoshikawa T, Imamura Y, Koizumi K, Kinoshita S. Paraoxonase gene polymorphisms and plasma oxidized low-density lipoprotein level as possible risk factors for exudative age-related macular degeneration. Am J. Ophthalmol. 2001 August; 132(2):191-5. [0355] Katz M L. Incomplete proteolysis may contribute to lipofuscin accumulation in the retinal pigment epithelium. Adv Exp Med. Biol. 1989; 266:109-16. [0356] Kennedy C J, Rakoczy P E, Constable I J. Lipofuscin of the retinal pigment epithelium: a review. Eye. 1995; 9 (Pt 6):763-71. [0357] Kennedy B N, Goldflam S, Chang M A, Campochiaro P, Davis A A, Zack D J, Crabb J W. Transcriptional regulation of cellular retinaldehyde-binding protein in the retinal pigment epithelium. A role for the photoreceptor consensus element. J Biol. Chem. 1998 Mar. 6; 273(10):5591-8. [0358] Kimura K, Isashiki Y, Sonoda S, Kakiuchi-Matsumoto T, Ohba N. Genetic association of manganese superoxide dismutase with exudative age-related macular degeneration. Am J Ophthalmol. 2000 December; 130(6):769-73. [0359] Klayer C C, Kliffen M, van Duijn C M, Hofman A, Cruts M, Grobbee D E, van Broeckhoven C, de Jong P T. Genetic association of apolipoprotein E with age-related macular degeneration. Am J Hum Genet. 1998 July; 63(1):200-6. [0360] Klein R, Klein B E, Linton K L. Prevalence of age-related maculopathy. The Beaver Dam Eye Study. Ophthalmology. 1992 June; 99(6):933-43. [0361] Klein M L, Mauldin W M, Stoumbos V D. Heredity and age-related macular degeneration. Observations in monozygotic twins. Arch Ophthalmol. 1994 July; 112(7):932-7. [0362] Klein M L, Schultz D W, Edwards A, Matise T C, Rust K, Berselli C B, Trzupek K, Weleber R G, Ott J, Wirtz M K, Acott T S. Age-related macular degeneration. Clinical features in a large family and linkage to chromosome 1q. Arch Ophthalmol. 1998 August; 116(8):1082-8. [0363] Kobayashi A, Higashide T, Hamasaki D, Kubota S, Sakuma H, An W, Fujimaki T, McLaren M J, Weleber R G, Inana G. HRG4 (UNC119) mutation found in cone-rod dystrophy causes retinal degeneration in a transgenic model. Invest Ophthalmol V is Sci. 2000 October; 41(11):3268-77. [0364] LaVail M M. Rod outer segment disk shedding in rat retina: relationship to cyclic lighting. Science. 1976 Dec. 3; 194(4269):1071-4. [0365] Lin H, Clegg D O. Integrin alphavbeta5 participates in the binding of photoreceptor rod outer segments during phagocytosis by cultured human retinal pigment epithelium. Invest Ophthalmol Vis Sci. 1998 August; 39(9):1703-12. [0366] Lohi J, Lehti K, Valtanen H, Parks W C, Keski-Oja J. Structural analysis and promoter characterization of the human membrane-type matrix metalloproteinase-1 (MT1-MMP) gene. Gene. 242(1-2):75-86, 2000 Jan. 25. [0367] Lowrey, P L; Shimomura, K, Antoch, M P; Yamakazi, S, Zemenides, P D, Ralph, M R, Menaker, M, Takahashi, J S. Positional Syntenic Cloning and Functional Characterization of the Mammalian Circadian Mutation tau. Science 2000, 288 (5465): 483-491 [0368] Mashima Y, Shiono T, Inana G. Rapid and efficient molecular analysis of gyrate atrophy using denaturing gradient gel electrophoresis. Invest Ophthalmol V is Sci. 1994 March; 35(3):1065-70. [0369] McLaren M J, Holderby M, Inana G. Phagocytosis of ROS by immortal rat RPE cell lines. Invest Ophthalmol V is Sci 34:A817, 1993a. [0370] McLaren M J, Sasabe T, Li C Y, Brown M E, Inana G. Spontaneously arising immortal cell line of rat retinal pigmented epithelial cells. Exp Cell Res. 1993b February; 204(2):311-20. [0371] McLaren M J, Sasabe T, Li C Y, Brown M E, Inana G. Double fluorescent vital assay of phagocytosis by cultured retinal pigment epithelial cells. Invest Ophthalmol V is Sci. 1993c February; 34(2):317-26. [0372] McLaren M J. Kinetics of rod outer segment phagocytosis by cultured retinal pigment epithelial cells. Relationship to cell morphology. Invest Ophthalmol V is Sci. 1996 June; 37(7):1213-24. [0373] Meyers S M, Greene T, Gutman F A. A twin study of age-related macular degeneration. Am J Ophthalmol. 1995 December; 120(6):757-66. [0374] Miceli M V, Newsome D A, Tate D J Jr. Vitronectin is responsible for serum-stimulated uptake of rod outer segments by cultured retinal pigment epithelial cells. Invest Ophthalmol V is Sci. 1997 July; 38(8):1588-97. [0375] Mitchell P, Wang J J, Smith W, Leeder S R. Smoking and the 5-year incidence of age-related maculopathy: the Blue Mountains Eye Study. Arch Ophthalmol. 2002 October; 120(10):1357-63. [0376] Oku N, Matsukawa M, Yamakawa S, Asai T, Yahara S, Hashimoto F, Akizawa T. Inhibitory effect of green tea polyphenols on membrane-type 1 matrix metalloproteinase, MT1-10 MMP. Biol Pharm Bull. 2003 September; 26(9):1235-8. [0377] Pei D, Weiss S J. Transmembrane-deletion mutants of the membrane-type matrix metalloproteinase-1 process progelatinase A and express intrinsic matrix-degrading activity. J Biol. Chem. 1996 Apr. 12; 271(15):9135-40. [0378] Sarks J P, Sarks S H, Killingsworth M C. Evolution of geographic atrophy of the retinal pigment epithelium. Eye. 1988; 2 (Pt 5):552-77. [0379] Sato H, Takino T, Okada Y, Cao J, Shinagawa A, Yamamoto E, Seiki M. A matrix metalloproteinase expressed on the surface of invasive tumour cells. Nature. 1994 Jul. 7; 370(6484):61-5. [0380] Schultz D W, Klein, M L, Humpert A J, Luzier C W, Persun V, /schain M, Mahan A, Runckel C, Cassera M, Vittal V, Doyle T M, Martin T M, Weleber R, Francis P J and Acott T S. Analysis of the ARMD1 locus: evidence that a mutation in hemicentin-1 is associated with age-related macular degeneration in a large family. Human Molecular Genetics Advance Access, published online Oct. 21, 2003. [0381] Shaban H, Borras C, Vina J, Richter C. Phosphatidylglycerol potently protects human retinal pigment epithelial cells against apoptosis induced by A2E, a compound suspected to cause age-related macula degeneration. Exp Eye Res. 2002 July; 75(1):99-108. [0382] Simonelli F, Margaglione M, Testa F, Cappucci G, Manitto M P, Brancato R, Rinaldi E. Apolipoprotein E polymorphisms in age-related macular degeneration in an Italian population. Ophthalmic Res. 2001 November-December; 33(6):325-8. [0383] Song E, Lee S K, Wang J, Ince N, Ouyang N, Min J, Chen J, Shankar P, Lieberman J. RNA interference targeting Fas protects mice from fulminant hepatitis. Nat. Med. 2003 March; 9(3):347-51. Epub 2003 Feb. 10. [0384] Stone E M, Webster A R, Vandenburgh K, Streb L M, Hockey R R, Lotery A J, Sheffield V C. Allelic variation in ABCR associated with Stargardt disease but not age-related [0385] Sui G, Soohoo C, Affar el B, Gay F, Shi Y, Forrester W C, Shi Y. A DNA vector-based RNAi technology to suppress gene expression in mammalian cells. Proc Natl Acad Sci USA. 2002 Apr. 16; 99(8):5515-20. [0386] Vickers T A, Koo S, Bennett C F, Crooke S T, Dean N M, Baker B F. Efficient reduction of target RNAs by small interfering RNA and RNase H-dependent antisense agents. A comparative analysis. J Biol. Chem. 2003 Feb. 28; 278(9):7108-18. Epub 2002 Dec. 23. [0387] Weeks D E, Conley Y P, Mah T S, Paul T O, Morse L, Ngo-Chang J, Dailey J P, Ferrell R E, Gorin M B. A full genome scan for age-related maculopathy. Hum Mol. Genet. 2000 May 22; 9(9):1329-49. [0388] Winkler B S, Boulton M E, Gottsch J D, Sternberg P. Oxidative damage and age-related macular degeneration. Mol. Vis. 1999 Nov. 3; 5:32. [0389] Young R W, Bok D. Participation of the retinal pigment epithelium in the rod outer segment renewal process. J. Cell Biol. 1969 August; 42(2):392-403. [0390] Zack D J, Bennett J, Wang Y, Davenport C, Klaunberg B, Gearhart J, Nathans J. Unusual topography of bovine rhodopsin promoter-lacZ fusion gene expression in transgenic mouse retinas. Neuron. 1991 February; 6(2):187-99. [0391] Zamore P D. Ancient pathways programmed by small RNAs. Science. 2002 May 17; 296(5571):1265-9. [0392] Zurdel J, Finckh U, Menzer G, Nitsch R M, Richard G. CST3 genotype associated with exudative age related macular degeneration. Br J. Ophthalmol.
2002 February; 86(2):214-9.
Sequence CWU
1
14112110DNAHomo sapiens 1gggatcgttc gatttaagcc atcatcagct taatttaagt
ttgtagtttt tgctgaagga 60ttatatgtat taatacttac ggttttaaat gtgttgcttt
ggatacacac atagtttctt 120ttttaataga atatactgtc ttgtctcact ttggactggg
acagtggatg cccatctaaa 180agttaagtgt catttctttt agatgtttac cttcagccat
agcttgattg ctcagagaaa 240tatgcagaag gcaggatcaa agacacacag gagtcctttc
ttttgaaatg ccacgtgcca 300ttgtctttcc tcccttcttt gcttcttttt cttaccctct
ctttcaattg cagatgccaa 360aaaagatgcc aacagacact acattaccct aatggctgct
acccagaacc tttttatagg 420ttgttcttaa tttttttgtt gttgttgttc aagcttttcc
tttctttttt ttcttggtgt 480ttgggccacg attttaaaat gacttttatt atgggtatgt
gttgccaaag ctggcttttt 540gtcaaataaa atgaatacga acttaaaaaa taaaagctgg
tatcttaaaa tgtaagagag 600taagactgtg aagcctaaaa tgactggctg agaatgaacc
agaaatgcca tttgccaaac 660agttgtaact agaaatttga ttctcacggt ccattctttt
ctttgtcctt aagatgacat 720tgttagtgtt cacgtcccat gttcagtgtc caaaccggca
atgtaaaaag tatcctgtgt 780ggtttaacag gaaatctgtt tatgtctctt tatttgaaac
cagttttact ctcagtggtt 840ctttaagttc aatgaagtct gccaggaaca ttggttggta
gtattattcc gacaccttta 900atttccaaaa tctgaagttc ctgctagttt accaccttca
tgatcttctt gaactggtaa 960ctgattaggt tgaacttatg gaagatttgt ggacttaact
caaaagtaac ctctcagtgt 1020tctatagaac atgtatttgt gtaactgaac ctaccaggag
aaatgtttgg aattctatat 1080gtgcaatttt tcaacaaatg caaaaaaaat acagcacatg
tattgacaag cttctgtcaa 1140gcagcttgag ttgaaatttg atttaagaaa ataaatcatg
attgttcaaa gctgctggga 1200cgttagaatt aggccatgat actggtctca ttttaactac
agtggtattt ggcactagtg 1260taaacttcca tataaatcac tcttttggaa caacaaaggg
ggagggagaa aaatcacggc 1320ctgttaaatg agtaccaaag ccgcccaaca gtaatgagat
gttctcatcc ttgattctcc 1380cagcctcaaa caacacagct tacttttttt ttcccttgct
cagaaagtac ctgtaattta 1440acaaacagac tgcctgtagg tatagtgcaa ttacaaatgc
tctaatcatt gtacatacat 1500ctctcttgat attgcagcat ccatactggc tttgtaatca
ttaatttttt ggcagattga 1560atgtgctgta ttgatatgta tctatgtaat tgtattgtat
gtctatagct aattcacgtt 1620ttgaataatg ttattttatt tactttttta agagaggaga
atgtaaattt gtcagtttat 1680ttctgactag ggatattttc tttccattta gaaaagaaga
aaaaaaaaaa accttactgt 1740catacagagc ggtactagcg tcgtgctgta taaaatcatt
tgcacattcc tgagtagagg 1800tatactgatt ataagaccca aaggtaattt catagcaaaa
tacataaaat cagtcggagc 1860ttttatacaa acatggaaac caactttgta gaacttttgc
catttgatct aggattggaa 1920tatgagcttt tatacaattc atattcttat ttggcaaatg
cacagtttag tattacctct 1980ctgatggcct ttactagaaa ggcagtttta gaagctattg
tgatccacta aggaaatgtt 2040ttaacagcta gagaccactg cttgcctgaa agggcgttct
taaatttggt gcagcaaaaa 2100aaagaaaaaa
21102775DNAHomo sapiens 2tgcaggagaa tggctactca
tcacacgctg tggatgggac tggccctgct gggggtgctg 60ggcgacctgc aggcagcacc
ggaggcccag gtctccgtgc agcccaactt ccagcaggac 120aagttcctgg ggcgctggtt
cagcgcgggc ctcgcctcca actcgagctg gctccgggag 180aagaaggcgg cgttgtccat
gtgcaagtct gtggtggccc ctgccacgga tggtggcctc 240aacctgacct ccaccttcct
caggaaaaac cagtgtgaga cccgaaccat gctgctgcag 300cccgcggggt ccctcggctc
ctacagctac cggagtcccc actggggcag cacctactcc 360gtgtcagtgg tggagaccga
ctacgaccag tacgcgctgc tgtacagcca gggcagcaag 420ggccctggcg aggacttccg
catggccacc ctctacagcc gaacccagac ccccagggct 480gagttaaagg agaaattcac
cgccttctgc aaggcccagg gcttcacaga ggataccatt 540gtcttcctgc cccaaaccga
taagtgcatg acggaacaat aggactcccc agggctgaag 600ctgggatccc ggccagccag
gtgaccccca cgctctggat gtctctgctc tgttccttcc 660ccgagcccct gccccggctc
cccgccaaag cacccctgcc cactcgggct tcatcctgca 720caataaactc cggaagcaag
tcagttaaaa aaaaaaaaaa aaaaaaaaaa aaaaa 77532139DNAHomo sapiens
3gaaaacagtg cagccacctc cgagagcctg gatgtgatgg cgtcacagaa gagaccctcc
60cagaggcacg gatccaagta cctggccaca gcaagtacca tggaccatgc caggcatggc
120ttcctcccaa ggcacagaga cacgggcatc cttgactcca tcgggcgctt ctttggcggt
180gacaggggtg cgccaaagcg gggctctggc aaggactcac accacccggc aagaactgct
240cactatggct ccctgcccca gaagtcacac ggccggaccc aagatgaaaa ccccgtagtc
300cacttcttca agaacattgt gacgcctcgc acaccacccc cgtcgcaggg aaaggggaga
360ggactgtccc tgagcagatt tagctggggg gccgaaggcc agagaccagg atttggctac
420ggaggcagag cgtccgacta taaatcggct cacaagggat tcaagggagt cgatgcccag
480ggcacgcttt ccaaaatttt taagctggga ggaagagata gtcgctctgg atcacccatg
540gctagacgct gaaaacccac ctggttccgg aatcctgtcc tcagcttctt aatataactg
600ccttaaaact ttaatcccac ttgcccctgt tacctaatta gagcagatga cccctcccct
660aatgcctgcg gagttgtgca cgtagtaggg tcaggccacg gcagcctacc ggcaatttcc
720ggccaacagt taaatgagaa catgaaaaca gaaaacggtt aaaactgtcc ctttctgtgt
780gaagatcacg ttccttcccc cgcaatgtgc ccccagacgc acgtgggtct tcagggggcc
840aggtgcacag acgtccctcc acgttcaccc ctccaccctt ggactttctt ttcgccgtgg
900ctcggcaccc ttgcgctttt gctggtcact gccatggagg cacacagctg cagagacaga
960gaggacgtgg gcggcagaga ggactgttga catccaagct tcctttgttt ttttttcctg
1020tccttctctc acctcctaaa gtagacttca tttttcctaa caggattaga cagtcaagga
1080gtggcttact acatgtggga gctttttggt atgtgacatg cgggctgggc agctgttaga
1140gtccaacgtg gggcagcaca gagagggggc cacctcccca ggccgtggct gcccacacac
1200cccaattagc tgaattcgcg tgtggcagag ggaggaaaag gaggcaaacg tgggctgggc
1260aatggcctca cataggaaac agggtcttcc tggagatttg gtgatggaga tgtcaagcag
1320gtggcctctg gacgtcaccg ttgccctgca tggtggcccc agagcagcct ctatgaacaa
1380cctcgtttcc aaaccacagc ccacagccgg agagtccagg aagacttgcg cactcagagc
1440agaagggtag gagtcctcta gacagcctcg cagccgcgcc agtcgcccat agacactggc
1500tgtgaccggg cgtgctggca gcggcagtgc acagtggcca gcactaaccc tccctgagaa
1560gataaccggc tcattcactt cctcccagaa gacgcgtggt agcgagtagg cacaggcgtg
1620cacctgctcc cgaattactc accgagacac acgggctgag cagacggccc ctgtgatgga
1680gacaaagagc tcttctgacc atatccttct taacacccgc tggcatctcc tttcgcgcct
1740ccctccctaa cctactgacc caccttttga ttttagcgca cctgtgattg ataggccttc
1800caaagagtcc cacgctggca tcaccctccc cgaggacgga gatgaggagt agtcagcgtg
1860atgccaaaac gcgtcttctt aatccaattc taattctgaa tgtttcgtgt gggcttaata
1920ccatgtctat taatatatag cctcgatgat gagagagtta caaagaacaa aactccagac
1980acaaacctcc aaatttttca gcagaagcac tctgcgtcgc tgagctgagg tcggctctgc
2040gatccatacg tggccgcacc cacacagcac gtgctgtgac gatggctgaa cggaaagtgt
2100acactgttcc tgaatattga aataaaacaa taaactttt
21394166DNAHomo sapiens 4ttcatataca aaaagataaa acttgaaata gttctagatt
tttcctccta ttgttggggt 60gtaactgctt cttcacacag ggggaaaaaa ctacattcac
atcggtttat ttgaggaccc 120agtgcagagt tcaagcagca aaaccccaac ttagcagatc
taattt 16651618DNAHomo sapiens 5ggcttggtca ccgcattaag
gcattcccgc tctccgcgga actgctctgc cgtctcggcg 60gtgaaagtgt gagagggtcc
gtagttgggt caactttgac tcctctcgcc tgcccggatc 120cttaagggcc tcctcgtcct
cccggtctcc ggtcgctgcc gggtctgtgc gccggtccgc 180gcccgccctc gctctgccat
gggcgcttcc agctcctccg cgctggcccg cctcggcctc 240ccagcccggc cctggcccag
gtggctcggg gtcgccgcgc taggactggc cgccgtggcc 300ctggggactg tcgcctggcg
ccgcgcatgg cccaggcggc gccggcggct gcagcaggtg 360ggcaccgtgg cgaagctctg
gatctacccg gtgaaatcct gcaaaggggt gccggtgagc 420gaggctgagt gcacggccat
ggggctgcgc agcggcaacc tgcgggacag gttttggctg 480gtgattaagg aagatggaca
catggtcact gcccgacagg agcctcgcct cgtgctcatc 540tccatcattt atgagaataa
ctgcctgatc ttcagggctc cagacatgga ccagctggtt 600ttgcctagca agcagccttc
ctcaaacaaa ctccacaact gcaggatatt tggccttgac 660attaaaggca gagactgtgg
caatgaggca gctaagtggt tcaccaactt cttgaaaact 720gaagcgtata gattggttca
atttgagaca aacatgaagg gaagaacatc aagaaaactt 780ctccccactc ttgatcagaa
tttccaggtg gcctacccag actactgccc gctcctgatc 840atgacagatg cctccctggt
agatttgaat accaggatgg agaagaaaat gaaaatggag 900aatttcaggc caaatattgt
ggtgaccggc tgtgatgctt ttgaggagga tacctgggat 960gaactcctaa ttggtagtgt
agaagtgaaa aaggtaatgg catgccccag gtgtattttg 1020acaacggtgg acccagacac
tggagtcata gacaggaaac agccactgga caccctgaag 1080agctaccgcc tgtgtgatcc
ttctgagagg gaattgtaca agttgtctcc actttttggg 1140atctattatt cagtggaaaa
aattggaagc ctgagagttg gtgaccctgt gtatcggatg 1200gtgtagtgat gagtgatgga
tccactaggg tgatatggct tcagcaacca ggagggattg 1260actgagatct taacaacagc
agcaacgata catcagcaaa tccttattat ccagccttca 1320actatcttta ccctggaaaa
caatctcgat ttttgacttt tcaaagttgt gtatgctcca 1380ggttaatgca aggaaagtat
tagagggggg aatatgaaag tatatatata aattttaggt 1440actgaaggct ttaaaaataa
ttaagatcat caaaaatgct attttgaatg ttatcatggc 1500tattacactt ttacttcctg
actttaatat tgatgaataa agcaagttta atgaatcaac 1560taaaaagctg caaaaaaaaa
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaa 161861669DNAHomo sapiens
6cggcggtgct gcgaggtcgg cgcgcagctc cgccgcgggt cgctcgggcg ctgtccaggc
60ggagccggcc ccgcccgggc tgcagccatg atcaagcgtt tcctggagga caccacggat
120gatggagaac tgagcaagtt cgtgaaggat ttctcaggaa atgcgagctg ccacccacca
180gaggctaaga cctgggcatc caggccccaa gtcccggagc caaggcccca ggccccggac
240ctctatgatg atgacctgga gttcagaccc ccctcgcggc cccagtcctc tgacaaccag
300cagtacttct gtgccccagc ccctctcagc ccatctgcca ggccccgcag cccatggggc
360aagcttgatc cctatgattc ctctgaggat gacaaggagt atgtgggctt tgcaaccctc
420cccaaccaag tccaccgaaa gtccgtgaag aaaggctttg actttaccct catggtggca
480ggagagtctg gcctgggcaa atccacactt gtcaatagcc tcttcctcac tgatctgtac
540cgggaccgga aacttcttgg tgctgaagag aggatcatgc aaactgtgga gatcactaag
600catgcagtgg acatagaaga gaagggtgtg aggctgcggc tcaccattgt ggacacacca
660ggttttgggg atgcagtcaa caacacagag tgctggaagc ctgtggcaga atacattgat
720cagcagtttg agcagtattt ccgagacgag agtggcctga accgaaagaa catccaagac
780aacagggtgc actgctgcct gtacttcatc tcacccttcg gccatgggct ccggccattg
840gatgttgaat tcatgaaggc cctgcatcag cgggtcaaca tcgtgcctat cctggctaag
900gcagacacac tgacacctcc cgaagtggac cacaagaaac gcaaaatccg ggaggagatt
960gagcattttg gaatcaagat ctatcaattc ccagactgtg actctgatga ggatgaggac
1020ttcaaattgc aggaccaagc cctaaaggaa agcatcccat ttgcagtaat tggcagcaac
1080actgtagtag aggccagagg gcggcgagtt cggggtcgac tctacccctg gggcatcgtg
1140gaagtggaaa acccagggca ctgcgacttt gtgaagctga ggacaatgct ggtacgtacc
1200cacatgcagg acctgaagga tgtgacgcgg gagacacatt atgagaacta ccgggcacag
1260tgcatccaga gcatgacccg cctggtggtg aaggaacgga atcgcaacaa actgactcgg
1320gaaagtggta ccgacttccc catccctgct gtcccaccag ggacagatcc agaaactgag
1380aagcttatcc gagagaaaga tgaggagctg cggcggatgc aggagatgct acacaaaata
1440caaaaacaga tgaaggagaa ctattaactg gctttcagcc ctggatattt aaatctcctc
1500ctcttcttcc tgtccatgcc ggcccctccc agcaccagct ctgctcaggc cccttcagct
1560actgccactt cgccttacat ccctgctgac tgcccagaga ctcagaggaa ataaagttta
1620ataaatctgt aggtggctaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaa
166971850DNAHomo sapiens 7cgcgctcgca gctcgcaggc gccgcgtagc cgtcgccacc
gccgccagcc cgtgcgccct 60cggcgcgtac ccgccgcgct cccatccccg ccgccggcca
ggggcgcgct cggccgcccc 120ggacagtgtc ccgctgcggc tccgcggcga tggccaccaa
gatcgacaaa gaggcttgcc 180gggcggcgta caacctggtg cgcgacgacg gctcggccgt
catctgggtg acttttaaat 240atgacggctc caccatcgtc cccggcgagc agggagcgga
gtaccagcac ttcatccagc 300agtgcacaga tgacgtccgg ttgtttgcct tcgtgcgctt
caccaccggg gatgccatga 360gcaagaggtc caagtttgcc ctcatcacgt ggatcggtga
gaacgtcagc gggctgcagc 420gcgccaaaac cgggacggac aagaccctgg tgaaggaggt
cgtacagaat ttcgctaagg 480agtttgtgat cagtgatcgg aaggagctgg aggaagattt
catcaagagc gagctgaaga 540aggcgggggg agccaattac gacgcccaga cggagtaacc
ccagcccccg ccacaccacc 600ccttgccaaa gtcatctgcc tgctccccgg gggagaggac
cgccggcctc agctactagc 660ccaccagccc accagggaga aaagaagcca tgagaggcag
cgcccgccac cctgtgtcca 720cagcccccac cttcccgctt cccttagaac cctgccgtgt
cctatctcat gacgctcatg 780gaacctcttt ctttgatctt ctttttcttt tctccccctc
ttttttgttc taaagaaaag 840tcattttgat gcaaggtcct gcctgccatc agatccgagg
tgcctcctgc agtgacccct 900tttcctggca tttctcttcc acgcgacgag gtctgcctag
tgagatctgc atgacctcac 960gttgctttcc agagcccggg cctattttgc catctcagtt
ttcctggacc ctgcttcctg 1020tgtaccactg aggggcagct gggccaggag ctgtgcccgg
tgcctgcagc cttcataagc 1080acacacgtcc attccctact aaggcccaga cctcctggta
tctgccccgg gctccctcat 1140cccacctcca tccggagttg cctaagatgc atgtccagca
taggcaggat tgctcggtgg 1200tgagaaggtt aggtccggct cagactgaat aagaagagat
aaaatttgcc ttaaaactta 1260cctggcagtg gctttgctgc acggtctgaa accacctgtt
cccaccctct tgaccgaaat 1320ttccttgtga cacagagaag ggcaaaggtc tgagcccaga
gttgacggag ggagtatttc 1380agggttcact tcaggggctc ccaaagcgac aagatcgtta
gggagagagg cccagggtgg 1440ggactgggaa tttaaggaga gctgggaacg gatcccttag
gttcaggaag cttctgtgta 1500agctgcgagg atggcttggg ccgaagggtt gctctgcccg
ccgcgctagc tgtgagctga 1560gcaaagccct gggctcacag caccccaaaa gcctgtggct
tcagtcctgc gtctgcacca 1620cacattcaaa aggatcgttt tgttttgttt ttaaagaaag
gtgagattgg cttggttctt 1680catgagcaca tttgatatag ctctttttct gtttttcctt
gctcatttcg ttttggggaa 1740gaaatctgta ctgtattggg attgtaaaga acatctctgc
actcagacag tttacagaaa 1800taaatgtttt ttttgttttt cagaaaaaaa aaaaaaaaaa
aaaaaaaaaa 185081646DNAHomo sapiens 8ctgaccgagg cgtgcaaaga
ctccagaatt ggaggcatga tgaagactct gctgctgttt 60gtggggctgc tgctgacctg
ggagagtggg caggtcctgg gggaccagac ggtctcagac 120aatgagctcc aggaaatgtc
caatcaggga agtaagtacg tcaataagga aattcaaaat 180gctgtcaacg gggtgaaaca
gataaagact ctcatagaaa aaacaaacga agagcgcaag 240acactgctca gcaacctaga
agaagccaag aagaagaaag aggatgccct aaatgagacc 300agggaatcag agacaaagct
gaaggagctc ccaggagtgt gcaatgagac catgatggcc 360ctctgggaag agtgtaagcc
ctgcctgaaa cagacctgca tgaagttcta cgcacgcgtc 420tgcagaagtg gctcaggcct
ggttggccgc cagcttgagg agttcctgaa ccagagctcg 480cccttctact tctggatgaa
tggtgaccgc atcgactccc tgctggagaa cgaccggcag 540cagacgcaca tgctggatgt
catgcaggac cacttcagcc gcgcgtccag catcatagac 600gagctcttcc aggacaggtt
cttcacccgg gagccccagg atacctacca ctacctgccc 660ttcagcctgc cccaccggag
gcctcacttc ttctttccca agtcccgcat cgtccgcagc 720ttgatgccct tctctccgta
cgagcccctg aacttccacg ccatgttcca gcccttcctt 780gagatgatac acgaggctca
gcaggccatg gacatccact tccacagccc ggccttccag 840cacccgccaa cagaattcat
acgagaaggc gacgatgacc ggactgtgtg ccgggagatc 900cgccacaact ccacgggctg
cctgcggatg aaggaccagt gtgacaagtg ccgggagatc 960ttgtctgtgg actgttccac
caacaacccc tcccaggcta agctgcggcg ggagctcgac 1020gaatccctcc aggtcgctga
gaggttgacc aggaaataca acgagctgct aaagtcctac 1080cagtggaaga tgctcaacac
ctcctccttg ctggagcagc tgaacgagca gtttaactgg 1140gtgtcccggc tggcaaacct
cacgcaaggc gaagaccagt actatctgcg ggtcaccacg 1200gtggcttccc acacttctga
ctcggacgtt ccttccggtg tcactgaggt ggtcgtgaag 1260ctctttgact ctgatcccat
cactgtgacg gtccctgtag aagtctccag gaagaaccct 1320aaatttatgg agaccgtggc
ggagaaagcg ctgcaggaat accgcaaaaa gcaccgggag 1380gagtgagatg tggatgttgc
ttttgcacct acgggggcat ctgagtccag ctccccccaa 1440gatgagctgc agccccccag
agagagctct gcacgtcacc aagtaaccag gccccagcct 1500ccaggccccc aactccgccc
agcctctccc cgctctggat cctgcactct aacactcgac 1560tctgctgctc atgggaagaa
cagaattgct cctgcatgca actaattcaa taaaactgtc 1620ttgtgagctg aaaaaaaaaa
aaaaaa 164691559DNAHomo sapiens
9gggaggcggc ggcggcggcg gcggcggcgg cgagagccca gagccagagc ccggccgggg
60ccgagcggag cgcggcggcg gcggcggcgg cggcggctgg gccgggagag gctggcgcgc
120cgggcggctc cgcgaatcct ccggcatccg ccccggcggg ccgcccccgc ccgcggcagc
180cccccgagca gtggcccggc atcggcgcct tcccggcggg caagagtgag ccatggagct
240acgtgtgggg aacaagtacc gcctgggacg gaagatcggg agcgggtcct tcggagatat
300ctacctgggt gccaacatcg cctctggtga ggaagtcgcc atcaagctgg agtgtgtgaa
360gacaaagcac ccccagctgc acatcgagag caagttctac aagatgatgc agggtggcgt
420ggggatcccg tccatcaagt ggtgcggagc tgagggcgac tacaacgtga tggtcatgga
480gctgctgggg cctagcctcg aggacctgtt caacttctgt tcccgcaaat tcagcctcaa
540gacggtgctg ctcttggccg accagatgat cagccgcatc gagtatatcc actccaagaa
600cttcatccac cgggacgtca agcccgacaa cttcctcatg gggctgggga agaagggcaa
660cctggtctac atcatcgact tcggcctggc caagaagtac cgggacgccc gcacccacca
720gcacattccc taccgggaaa acaagaacct gaccggcacg gcccgctacg cttccatcaa
780cacgcacctg ggcattgagc aaagccgtcg agatgacctg gagagcctgg gctacgtgct
840catgtacttc aacctgggct ccctgccctg gcaggggctc aaagcagcca ccaagcgcca
900gaagtatgaa cggatcagcg agaagaagat gtcaacgccc atcgaggtcc tctgcaaagg
960ctatccctcc gaattctcaa catacctcaa cttctgccgc tccctgcggt ttgacgacaa
1020gcccgactac tcttacctac gtcagctctt ccgcaacctc ttccaccggc agggcttctc
1080ctatgactac gtctttgact ggaacatgct gaaattcggt gcagcccgga atcccgagga
1140tgtggaccgg gagcggcgag aacacgaacg cgaggagagg atggggcagc tacgggggtc
1200cgcgacccga gccctgcccc ctggcccacc cacgggggcc actgccaacc ggctccgcag
1260tgccgccgag cccgtggctt ccacgccagc ctcccgcatc cagccggctg gcaatacttc
1320tcccagagcg atctcgcggg tcgaccggga gaggaaggtg agtatgaggc tgcacagggg
1380tgcgcccgcc aacgtctcct cctcagacct cactgggcgg caagaggtct cccggatccc
1440agcctcacag acaagtgtgc catttgacca tctcgggaag tgaggagagc ccccattgga
1500ccagtgtttg cttagtgtct tcactgtatt ttctttaaaa aaaaaaaaaa aaaaaaaaa
155910910DNAHomo sapiens 10cctgcttcaa cagtgcttgg acggaacccg gcgctcgttc
cccaccccgg ccggccgccc 60atagccagcc ctccgtcacc tcttcaccgc accctcggac
tgccccaagg cccccgccgc 120cgctccagcg ccgcgcagcc accgccgccg ccgccgcctc
tccttagtcg ccgccatgac 180gaccgcgtcc acctcgcagg tgcgccagaa ctaccaccag
gactcagagg ccgccatcaa 240ccgccagatc aacctggagc tctacgcctc ctacgtttac
ctgtccatgt cttactactt 300tgaccgcgat gatgtggctt tgaagaactt tgccaaatac
tttcttcacc aatctcatga 360ggagagggaa catgctgaga aactgatgaa gctgcagaac
caacgaggtg gccgaatctt 420ccttcaggat atcaagaaac cagactgtga tgactgggag
agcgggctga atgcaatgga 480gtgtgcatta catttggaaa aaatgtgaat cagtcactac
tggaactgca caaactggcc 540actgacaaaa atgaccccca tttgtgtgac ttcattgaga
cacattacct gaatgagcag 600gtgaaagcca tcaaagaatt gggtgaccac gtgaccaact
tgcgcaagat gggagcgccc 660gaatctggct tggcggaata tctctttgac aagcacaccc
tgggagacag tgataatgaa 720agctaagcct cgggctaatt tccccatagc cgtggggtga
cttccctggt caccaaggca 780gtgcatgcat gttggggttt cctttacctt ttctataagt
tgtaccaaaa catccactta 840agttctttga tttgtaccat tccttcaaat aaagaaattt
ggtacccaaa aaaaaaaaaa 900aaaaaaaaaa
910112740DNAHomo sapiens 11cgctgccatg cggctggcgc
tgctctgggc cctggggctc ctgggcgcgg gcagccctct 60gccttcctgg ccgctcccaa
atataggtgg cactgaggag cagcaggcag agtcagagaa 120ggccccgagg gagcccttgg
agccccaggt ccttcaggac gatctcccaa ttagcctcaa 180aaaggtgctt cagaccagtc
tgcctgagcc cctgaggatc aagttggagc tggacggtga 240cagtcatatc ctggagctgc
tacagaatag ggagttggtc ccaggccgcc caaccctggt 300gtggtaccag cccgatggca
ctcgggtggt cagtgaggga cacactttgg agaactgctg 360ctaccaggga agagtgcggg
gatatgcagg ctcctgggtg tccatctgca cctgctctgg 420gctcagaggc ttggtggtcc
tgaccccaga gagaagctat accctggagc aggggcctgg 480ggaccttcag ggtcctccca
ttatttcgcg aatccaagat ctccacctgc caggccacac 540ctgtgccctg agctggcggg
aatctgtaca cactcagacg ccaccagagc accccctggg 600acagcgccac attcgccgga
ggcgggatgt ggtaacagag accaagactg tggagttggt 660gattgtggct gatcactcgg
aggcccagaa ataccgggac ttccagcacc tgctaaaccg 720cacactggaa gtggccctct
tgctggacac attcttccgg cccctgaatg tacgagtggc 780actagtgggc ctggaggcct
ggacccagcg tgacctggtg gagatcagcc caaacccagc 840tgtcaccctc gaaaacttcc
tccactggcg cagggcacat ttgctgcctc gattgcccca 900tgacagtgcc cagctggtga
ctggtacttc attctctggg cctacggtgg gcatggccat 960tcagaactcc atctgttctc
ctgacttctc aggaggtgtg aacatggacc actccaccag 1020catcctggga gtcgcctcct
ccatagccca tgagttgggc cacagcctgg gcctggacca 1080tgatttgcct gggaatagct
gcccctgtcc aggtccagcc ccagccaaga cctgcatcat 1140ggaggcctcc acagacttcc
taccaggcct gaacttcagc aactgcagcc gacgggccct 1200ggagaaagcc ctcctggatg
gaatgggcag ctgcctcttc gaacggctgc ctagcctacc 1260ccctatggct gctttctgcg
gaaatatgtt tgtggagccg ggcgagcagt gtgactgtgg 1320cttcctggat gactgcgtcg
atccctgctg tgattctttg acctgccagc tgaggccagg 1380tgcacagtgt gcatctgacg
gaccctgttg tcaaaattgc cagctgcgcc cgtctggctg 1440gcagtgtcgt cctaccagag
gggattgtga cttgcctgaa ttctgcccag gagacagctc 1500ccagtgtccc cctgatgtca
gcctagggga tggcgagccc tgcgctggcg ggcaagctgt 1560gtgcatgcac gggcgttgtg
cctcctatgc ccagcagtgc cagtcacttt ggggacctgg 1620agcccagccc gctgcgccac
tttgcctcca gacagctaat actcggggaa atgcttttgg 1680gagctgtggg cgcaacccca
gtggcagtta tgtgtcctgc acccctagag atgccatttg 1740tgggcagctc cagtgccaga
caggtaggac ccagcctctg ctgggctcca tccgggatct 1800actctgggag acaatagatg
tgaatgggac tgagctgaac tgcagctggg tgcacctgga 1860cctgggcagt gatgtggccc
agcccctcct gactctgcct ggcacagcct gtggccctgg 1920cctggtgtgt atagaccatc
gatgccagcg tgtggatctc ctgggggcac aggaatgtcg 1980aagcaaatgc catggacatg
gggtctgtga cagcaacagg cactgctact gtgaggaggg 2040ctgggcaccc cctgactgca
ccactcagct caaagcaacc agctccctga ccacagggct 2100gctcctcagc ctcctggtct
tattggtcct ggtgatgctt ggtgccggct actggtaccg 2160tgcccgcctg caccagcgac
tctgccagct caagggaccc acctgccagt acagggcagc 2220ccaatctggt ccctctgaac
ggccaggacc tccgcagagg gccctgctgg cacgaggcac 2280taagtctcag gggccagcca
agcccccacc cccaaggaag ccactgcctg ccgaccccca 2340gggccggtgc ccatcgggtg
acctgcccgg cccaggggct ggaatcccgc ccctagtggt 2400accctccaga ccagcgccac
cgcctccgac agtgtcctcg ctctacctct gacctctccg 2460gaggttccgc tgcctccaag
ccggacttag ggcttcaaga ggcgggcgtg ccctctggag 2520tcccctacca tgactgaagg
cgccagagac tggcggtgtc ttaagactcc gggcaccgcc 2580acgcgctgtc aagcaacact
ctgcggacct gccggcgtag ttgcagcggg ggcttgggga 2640ggggctgggg gttggacggg
attgaggaag gtccgcacag cctgtctctg ctcagttgca 2700ataaacgtga catcttggga
gcgttaaaaa aaaaaaaaaa 2740121476DNAHomo sapiens
12gtttaatagc ttgaggaagg gagactttaa aaggacgtgt gtgagtgaaa taggatatag
60ccattaccac ggtgccagga cctgacagcg ttccaattct ttttgcagca tggggaatca
120aaggtggcat gccaagttca actcagggct gaggtatcca cattgtccac atcaggcaag
180ccctgcactg acggttgagc ctcatggaga ggagcatgtg ttggaaagag atccctttgt
240taactgtttt gtggtgttct cttcaatgaa ttagagctca tgcccctttt ctggctttgc
300tgttgatttt ggatggtaga gaatattcct gagagccttc cttttggccc ccagcttatg
360ccacccactc tcttctcttg gttgaattct ctgaaggaaa ggttcatgtg ctattgtcct
420gttagtcaat agtcttcata tataattgtg ttacatatat tgctgtagac tctcagaaat
480cagggtagag cttttccttt gagcagttta atgagtgaat tcagcagcaa agtcgcaaga
540aatggttctc cagccaggag aggttatgtt tatcctctga ttgcccgttt tctctgcaca
600cagtgatatc gtattcagtg agaggtgctg ttggcaccca gcagcaccct gggcacacag
660catttcatgt catgtcacag tgtacaagct accctctaat tcagaaagaa gagcattttg
720cacagagaaa aataaaaaga tccatgaatg tcatctttta tcttttattt tcagttggct
780gatgttggaa tttttgttct tgtcatgaac ttgtaaacca atcttgccaa gatacaagtt
840gttttggttt ttcactacaa tgacctcttg ttcctcctgt cttgactgct gacgttcctc
900aatgattcta ttgtctattt tatgggaagc agccttccca taggtttcct tttacacact
960gcagggctat ctttatactt taaaaaaaaa aaaaaaaaaa aaaaggacaa gaactgtcac
1020taacctcatg gaggggtttg cgtaaaacca tttagcccac cttgagcaaa gggtagattc
1080cgtgttgttt ttttaagctc actgtaataa aatagatcta attcagcatt attgtgctac
1140ctcaaaggta aaaaatgttt taaggtcttc ttttggtcct gagttctata tacagtgttt
1200gaaatgtctt tcatttggaa ttatttttta aattcttgga gtgaatttta ttttaatctg
1260ttttaatctt gtattttaat ctcagaagaa taagtgattg aaacgtgatc aattcttgct
1320ctgtggtgtt aaacatataa tgaacagtca ttaagaatta agtcactgtt tgccataaac
1380aaggttgatg ttctttttgt tgttgttaag gaaaccctag ggctcggctt tactcttgat
1440taataaaggc tgacaaatca aaaaaaaaaa aaaaaa
1476131679DNAHomo sapiens 13ggcacgaggt agagctccag gacattcagg taccaggtag
ccccaaggag gagctgccga 60cctggcaggg aacaaccaag actggggtta aatctcacag
cctgcaagtg gaagagaaga 120acttgaaccc aggtccaact tttgcgccac agcaggctgc
ctcttggtcc tgacaggaag 180tcacaacttg gccctgactt cctatcctag ggaaggggcc
ggctggagag gccaggacag 240agaaagcaga tcccttcttt ttccaaggac tctgtgtctt
ccataggcaa catgtcagaa 300ggggtgggca cgttccgcat ggtacctgaa gaggaacagg
agctccgtgc ccaactggag 360cagctcacaa ccaaggacca tggacctgtc tttggcccgt
gcagccagct gccccgccac 420accttgcaga aggccaagga tgagctgaac gagagagagg
agacccggga ggaggcagtg 480cgagagctgc aggagatggt gcaggcgcag gcggcctcgg
gggaggagct ggcggtggcc 540gtggcggaga gggtgcaaga gaaggacagc ggcttcttcc
tgcgcttcat ccgcgcacgg 600aagttcaacg tgggccgtgc ctatgagctg ctcagaggct
atgtgaattt ccggctgcag 660taccctgagc tctttgacag cctgtcccca gaggctgtcc
gctgcaccat tgaagctggc 720taccctggtg tcctctctag tcgggacaag tatggccgag
tggtcatgct cttcaacatt 780gagaactggc aaagtcaaga aatcaccttt gatgagatct
tgcaggcata ttgcttcatc 840ctggagaagc tgctggagaa tgaggaaact caaatcaatg
gcttctgcat cattgagaac 900ttcaagggct ttaccatgca gcaggctgct agtctccgga
cttcagatct caggaagatg 960gtggacatgc tccaggattc cttcccagcc cggttcaaag
ccatccactt catccaccag 1020ccatggtact tcaccacgac ctacaatgtg gtcaagccct
tcttgaagag caagctgctt 1080gagagggtct ttgtccacgg ggatgacctt tctggtttct
accaggagat cgatgagaac 1140atcctgccct ctgacttcgg gggcacgctg cccaagtatg
atggcaaggc cgttgctgag 1200cagctctttg gcccccaggc ccaagctgag aacacagcct
tctgaaaaca tctcctgcca 1260gctgaactgt agttagaatc tctgggcctc tcctcaactg
tcctggaccc aaggctagga 1320aagggctgct tgagatgact gtggtccccc cttagactcc
ctaagcccga gtgagctcag 1380gtgtcaccct gttctcaagt tgggggatgg ggaataaagg
agggggaatt cccttgaaca 1440agaagaactg gggatagtta tatttccacc tgcccttgaa
gctttaagac agtgattttt 1500gtgtaaggtt gtatttcaaa gactcgaatt cattttctca
atcatttcct ttgtaacaga 1560gttttacgac ttagagtctg tgaaaacagg caaggagccc
gggttaaaat atccccctat 1620tcgcccccaa aatgcaataa aagaagataa aagagagagg
aaaaaaaaaa aaaaaaaaa 1679141962DNAHomo sapiens 14agctctcgca ctctgttctt
ccgccgctcc gccgtcgcgt ttctctgccg gtcgcaatgg 60aagaagagat cgccgcgctg
gtcattgaca atggctccgg catgtgcaaa gctggttttg 120ctggggacga cgctccccga
gccgtgtttc cttccatcgt cgggcgcccc agacaccagg 180gcgtcatggt gggcatgggc
cagaaggact cctacgtggg cgacgaggcc cagagcaagc 240gtggcatcct gaccctgaag
taccccattg agcatggcat cgtcaccaac tgggacgaca 300tggagaagat ctggcaccac
accttctaca acgagctgcg cgtggccccg gaggagcacc 360cagtgctgct gaccgaggcc
cccctgaacc ccaaggccaa cagagagaag atgactcaga 420ttatgtttga gaccttcaac
accccggcca tgtacgtggc catccaggcc gtgctgtccc 480tctacgcctc tgggcgcacc
actggcattg tcatggactc tggagacggg gtcacccaca 540cggtgcccat ctacgagggc
tacgccctcc cccacgccat cctgcgtctg gacctggctg 600gccgggacct gaccgactac
ctcatgaaga tcctcactga gcgaggctac agcttcacca 660ccacggccga gcgggaaatc
gtgcgcgaca tcaaggagaa gctgtgctac gtcgccctgg 720acttcgagca ggagatggcc
accgccgcat cctcctcttc tctggagaag agctacgagc 780tgcccgatgg ccaggtcatc
accattggca atgagcggtt ccggtgtccg gaggcgctgt 840tccagccttc cttcctgggt
atggaatctt gcggcatcca cgagaccacc ttcaactcca 900tcatgaagtg tgacgtggac
atccgcaaag acctgtacgc caacacggtg ctgtcgggcg 960gcaccaccat gtacccgggc
attgccgaca ggatgcagaa ggagatcacc gccctggcgc 1020ccagcaccat gaagatcaag
atcatcgcac ccccagagcg caagtactcg gtgtggatcg 1080gtggctccat cctggcctca
ctgtccacct tccagcagat gtggattagc aagcaggagt 1140acgacgagtc gggcccctcc
atcgtccacc gcaaatgctt ctaaacggac tcagcagatg 1200cgtagcattt gctgcatggg
ttaattgaga atagaaattt gcccctggca aatgcacaca 1260cctcatgcta gcctcacgaa
actggaataa gccttcgaaa agaaattgtc cttgaagctt 1320gtatctgata tcagcactgg
attgtagaac ttgttgctga ttttgacctt gtattgaagt 1380taactgttcc ccttggtatt
tgtttaatac cctgtacata tctttgagtt caacctttag 1440tacgtgtggc ttggtcactt
cgtggctaag gtaagaacgt gcttgtggaa gacaagtctg 1500tggcttggtg agtctgtgtg
gccagcagcc tctgatctgt gcagggtatt aacgtgtcag 1560ggctgagtgt tctgggattt
ctctagaggc tggcaagaac cagttgtttt gtcttgcggg 1620tctgtcaggg ttggaaagtc
caagccgtag gacccagttt cctttcttag ctgatgtctt 1680tggccagaac accgtgggct
gttacttgct ttgagttgga agcggtttgc atttacgcct 1740gtaaatgtat tcattcttaa
tttatgtaag gttttttttg tacgcaattc tcgattcttt 1800gaagagatga caacaaattt
tggttttcta ctgttatgtg agaacattag gccccagcaa 1860cacgtcattg tgtaaggaaa
aataaaagtg ctgccgtaac caaaaaaaaa aaaaaaaaaa 1920aaaaaaaaaa aaaaaaaaaa
aaaaaaaaaa aaaaaaaaaa aa 1962152365DNAHomo sapiens
15gaattcaagt tcagtgccta ccgaagacaa aggcgccccg agggagtggc ggtgcgaccc
60cagggcgtgg gcccggccgc ggagcccaca ctgcccggct gacccggtgg tctcggacca
120tgtctcccgc cccaagaccc tcccgttgtc tcctgctccc cctgctcacg ctcggcaccg
180cgctcgcctc cctcggctcg gcccaaagca gcagcttcag ccccgaagcc tggctacagc
240aatatggcta cctgcctccc ggggacctac gtacccacac acagcgctca ccccagtcac
300tctcagcggc catcgctgcc atgcagaagt tttacggctt gcaagtaaca ggcaaagctg
360atgcagacac catgaaggcc atgaggcgcc cccgatgtgg tgttccagac aagtttgggg
420ctgagatcaa ggccaatgtt cgaaggaagc gctacgccat ccagggtctc aaatggcaac
480ataatgaaat cactttctgc atccagaatt acacccccaa ggtgggcgag tatgccacat
540acgaggccat tcgcaaggcg ttccgcgtgt gggagagtgc cacaccactg cgcttccgcg
600aggtgcccta tgcctacatc cgtgagggcc atgagaagca ggccgacatc atgatcttct
660ttgccgaggg cttccatggc gacagcacgc ccttcgatgg tgagggcggc ttcctggccc
720atgcctactt cccaggcccc aacattggag gagacaccca ctttgactct gccgagcctt
780ggactgtcag gaatgaggat ctgaatggaa atgacatctt cctggtggct gtgcacgagc
840tgggccatgc cctggggctc gagcattcca gtgacccctc ggccatcatg gcaccctttt
900accagtggat ggacacggag aattttgtgc tgcccgatga tgaccgccgg ggcatccagc
960aactttatgg gggtgagtca gggttcccca ccaagatgcc ccctcaaccc aggactacct
1020cccggccttc tgttcctgat aaacccaaaa accccaccta tgggcccaac atctgtgacg
1080ggaactttga caccgtggcc atgctccgag gggagatgtt tgtcttcaag gagcgctggt
1140tctggcgggt gaggaataac caagtgatgg atggataccc aatgcccatt ggccagttct
1200ggcggggcct gcctgcgtcc atcaacactg cctacgagag gaaggatggc aaattcgtct
1260tcttcaaagg agacaagcat tgggtgtttg atgaggcgtc cctggaacct ggctacccca
1320agcacattaa ggagctgggc cgagggctgc ctaccgacaa gattgatgct gctctcttct
1380ggatgcccaa tggaaagacc tacttcttcc gtggaaacaa gtactaccgt ttcaacgaag
1440agctcagggc agtggatagc gagtacccca agaacatcaa agtctgggaa gggatccctg
1500agtctcccag agggtcattc atgggcagcg atgaagtctt cacttacttc tacaagggga
1560acaaatactg gaaattcaac aaccagaagc tgaaggtaga accgggctac cccaagtcag
1620ccctgaggga ctggatgggc tgcccatcgg gaggccggcc cgatgagggg actgaggagg
1680agacggaggt gatcatcatt gaggtggacg aggagggcgg cggggcggtg agcgctgctg
1740ccgtggtgct gcccgtgctg ctgctgctcc tggtgctggc ggtgggacta gcagtcttct
1800tcttcagacg ccatgggacc cccaggcgac tgctctactg ccagcgttcc ctgctggaca
1860aggtctgacg cccaccgccg gcccgcccac tcctaccaca aggactttgc ctctgaagac
1920cagtgtcagc aaggtggtgg tgggtgggct gctcccatcc gtccggagcc ccctccccgc
1980agcctccttg cttctctcag tcccctggct ggcctccttc accctcaccg cctgtagctt
2040gtgtctgtcc agccccatct gaatgtgttg ggggctctgc acttgaaggc aggaccctca
2100gacctcgctg gtaaaggtca aatggggtca tctgctcctt ttccatcccc tgacatacct
2160taacctctga actctgacct caggaggctc tgggcactcc agccctgaaa gccccaagtg
2220tacccagttg gcagcctccc gtcactctga ctaaaaagaa tcttcagagt gcatatttgg
2280aggtggaaag attgttcagt taccctaaag actttgaaag aaagaaagaa agaaagaaaa
2340aaaaaaaaaa aaaaaaaaaa aaaaa
2365168595DNAHomo sapiens 16aaagcggaga gtcacagcgg ggccaggccc tggggagcgg
agcctccacc gcccccctca 60ttcccaggca agggcttggg gggaatgagc cgggagagcc
gggtcccgag cctacagagc 120cgggagcagc tgagccgccg gcgcctcggc cgccgccgcc
gcctcctcct cctccgccgc 180cgccagcccg gagcctgagc cggcggggcg ggggggagag
gagcgagcgc agcgcagcag 240cggagccccg cgaggcccgc ccgggcgggt ggggagggca
gcccggggga ctgggccccg 300gggcggggtg ggaggggggg agaagacgaa gacagggccg
ggtctctccg cggacgagac 360agcggggatc atggccgcgc aggtcgcccc cgccgccgcc
agcagcctgg gcaacccgcc 420gccgccgccg ccctcggagc tgaagaaagc cgagcagcag
cagcgggagg aggcgggggg 480cgaggcggcg gcggcggcag cggccgagcg cggggaaatg
aaggcagccg ccgggcagga 540aagcgagggc cccgccgtgg ggccgccgca gccgctggga
aaggagctgc aggacggggc 600cgagagcaat gggggtggcg gcggcggcgg agccggcagc
ggcggcgggc ccggcgcgga 660gccggacctg aagaactcga acgggaacgc gggccctagg
cccgccctga acaataacct 720cacggagccg cccggcggcg gcggtggcgg cagcagcgat
ggggtggggg cgcctcctca 780ctcagccgcg gccgccttgc cgcccccagc ctacggcttc
gggcaaccct acggccggag 840cccgtctgcc gtcgccgccg ccgcggccgc cgtcttccac
caacaacatg gcggacaaca 900aagccctggc ctggcagcgc tgcagagcgg cggcggcggg
ggcctggagc cctacgcggg 960gccccagcag aactctcacg accacggctt ccccaaccac
cagtacaact cctactaccc 1020caaccgcagc gcctaccccc cgcccgcccc ggcctacgcg
ctgagctccc cgagaggtgg 1080cactccgggc tccggcgcgg cggcggctgc cggctccaag
ccgcctccct cctccagcgc 1140ctccgcctcc tcgtcgtctt cgtccttcgc tcagcagcgc
ttcggggcca tggggggagg 1200cggcccctcc gcggccggcg ggggaactcc ccagcccacc
gccaccccca ccctcaacca 1260actgctcacg tcgcccagct cggcccgggg ctaccagggc
taccccgggg gcgactacag 1320tggcgggccc caggacgggg gcgccggcaa gggcccggcg
gacatggcct cgcagtgttg 1380gggggctgcg gcggcggcag ctgcggcggc ggccgcctcg
ggaggggccc aacaaaggag 1440ccaccacgcg cccatgagcc ccgggagcag cggcggcggg
gggcagccgc tcgcccggac 1500ccctcagcca tccagtccaa tggatcagat gggcaagatg
agacctcagc catatggcgg 1560gactaaccca tactcgcagc aacagggacc tccgtcagga
ccgcagcaag gacatgggta 1620cccagggcag ccatacgggt cccagacccc gcagcggtac
ccgatgacca tgcagggccg 1680ggcgcagagt gccatgggcg gcctctctta tacacagcag
attcctcctt atggacaaca 1740aggccccagc gggtatggtc aacagggcca gactccatat
tacaaccagc aaagtcctca 1800ccctcagcag cagcagccac cctactccca gcaaccaccg
tcccagaccc ctcatgccca 1860accttcgtat cagcagcagc cacagtctca accaccacag
ctccagtcct ctcagcctcc 1920atactcccag cagccatccc agcctccaca tcagcagtcc
ccggctccat acccctccca 1980gcagtcgacg acacagcagc acccccagag ccagcccccc
tactcacagc cacaggctca 2040gtctccttac cagcagcagc aacctcagca gccagcaccc
tcgacgctct cccagcaggc 2100tgcgtatcct cagccccagt ctcagcagtc ccagcaaact
gcctattccc agcagcgctt 2160ccctccaccg caggagctat ctcaagattc atttgggtct
caggcatcct cagccccctc 2220aatgacctcc agtaagggag ggcaagaaga tatgaacctg
agccttcagt caagaccctc 2280cagcttgcct gatctatctg gttcaataga tgacctcccc
atggggacag aaggagctct 2340gagtcctgga gtgagcacat cagggatttc cagcagccaa
ggagagcaga gtaatccagc 2400tcagtctcct ttctctcctc atacctcccc tcacctgcct
ggcatccgag gcccttcccc 2460gtcccctgtt ggctctcccg ccagtgttgc tcagtctcgc
tcaggaccac tctcgcctgc 2520tgcagtgcca ggcaaccaga tgccacctcg gccacccagt
ggccagtcgg acagcatcat 2580gcatccttcc atgaaccaat caagcattgc ccaagatcga
ggttatatgc agaggaaccc 2640ccagatgccc cagtacagtt ccccccagcc cggctcagcc
ttatctccgc gtcagccttc 2700cggaggacag atacacacag gcatgggctc ctaccagcag
aactccatgg ggagctatgg 2760tccccagggg ggtcagtatg gcccacaagg tggctacccc
aggcagccaa actataatgc 2820cttgcccaat gccaactacc ccagtgcagg catggctgga
ggcataaacc ccatgggtgc 2880cggaggtcaa atgcatggac agcctggcat cccaccttat
ggcacactcc ctccagggag 2940gatgagtcac gcctccatgg gcaaccggcc ttatggccct
aacatggcca atatgccacc 3000tcaggttggg tcagggatgt gtcccccacc agggggcatg
aaccggaaaa cccaagaaac 3060tgctgtcgcc atgcatgttg ctgccaactc tatccaaaac
aggccgccag gctaccccaa 3120tatgaatcaa gggggcatga tgggaactgg acctccttat
ggacaaggga ttaatagtat 3180ggctggcatg atcaaccctc agggaccccc atattccatg
ggtggaacca tggccaacaa 3240ttctgcaggg atggcagcca gcccagagat gatgggcctt
ggggatgtaa agttaactcc 3300agccaccaaa atgaacaaca aggcagatgg gacacccaag
acagaatcca aatccaagaa 3360atccagttct tctactacaa ccaatgagaa gatcaccaag
ttgtatgagc tgggtggtga 3420gcctgagagg aagatgtggg tggaccgtta tctggccttc
actgaggaga aggccatggg 3480catgacaaat ctgcctgctg tgggtaggaa acctctggac
ctctatcgcc tctatgtgtc 3540tgtgaaggag attggtggat tgactcaggt caacaagaac
aaaaaatggc gggaacttgc 3600aaccaacctc aatgtgggca catcaagcag tgctgccagc
tccttgaaaa agcagtatat 3660ccagtgtctc tatgcctttg aatgcaagat tgaacgggga
gaagaccctc ccccagacat 3720ctttgcagct gctgattcca agaagtccca gcccaagatc
cagcctccct ctcctgcggg 3780atcaggatct atgcaggggc cccagactcc ccagtcaacc
agcagttcca tggcagaagg 3840aggagactta aagccaccaa ctccagcatc cacaccacac
agtcagatcc ccccattgcc 3900aggcatgagc aggagcaatt cagttgggat ccaggatgcc
tttaatgatg gaagtgactc 3960cacattccag aagcggaatt ccatgactcc aaaccctggg
tatcagccca gtatgaatac 4020ctctgacatg atggggcgca tgtcctatga gccaaataag
gatccttatg gcagcatgag 4080gaaagctcca gggagtgatc ccttcatgtc ctcagggcag
ggccccaacg gcgggatggg 4140tgacccctac agtcgtgctg ccggccctgg gctaggaaat
gtggcgatgg gaccacgaca 4200gcactatccc tatggaggtc cttatgacag agtgaggacg
gagcctggaa tagggcctga 4260gggaaacatg agcactgggg ccccacagcc gaatctcatg
ccttccaacc cagactcggg 4320gatgtattct cctagccgct accccccgca gcagcagcag
cagcagcagc aacgacatga 4380ttcctatggc aatcagttct ccacccaagg caccccttct
ggcagcccct tccccagcca 4440gcagactaca atgtatcaac agcaacagca gaattacaag
cggccaatgg atggcacata 4500tggccctcct gccaagcggc acgaagggga gatgtacagc
gtgccataca gcactgggca 4560ggggcagcct cagcagcagc agttgccccc agcccagccc
cagcctgcca gccagcaaca 4620agctgcccag ccttcccctc agcaagatgt atacaaccag
tatggcaatg cctatcctgc 4680cactgccaca gctgctactg agcgccgacc agcaggcggc
ccccagaacc aatttccatt 4740ccagtttggc cgagaccgtg tctctgcacc ccctggcacc
aatgcccagc aaaacatgcc 4800accacaaatg atgggcggcc ccatacaggc atcagctgag
gttgctcagc aaggcaccat 4860gtggcagggg cgtaatgaca tgacctataa ttatgccaac
aggcagagca cgggctctgc 4920cccccagggc cccgcctatc atggcgtgaa ccgaacagat
gaaatgctgc acacagatca 4980gagggccaac cacgaaggct cgtggccttc ccatggcaca
cgccagcccc catatggtcc 5040ctctgcccct gtgcccccca tgacaaggcc ccctccatct
aactaccagc ccccaccaag 5100catgcagaat cacattcctc aggtatccag ccctgctccc
ctgccccggc caatggagaa 5160ccgcacctct cctagcaagt ctccattcct gcactctggg
atgaaaatgc agaaggcagg 5220tcccccagta cctgcctcgc acatagcacc tgcccctgtg
cagcccccca tgattcggcg 5280ggatatcacc ttcccacctg gctctgttga agccacacag
cctgtgttga agcagaggag 5340gcggctcaca atgaaagaca ttggaacccc ggaggcatgg
cgggtaatga tgtccctcaa 5400gtctggtctc ctggcagaga gcacatgggc attagatacc
atcaacatcc tgctgtatga 5460tgacaacagc atcatgacct tcaacctcag tcagctccca
gggttgctag agctccttgt 5520agaatatttc cgacgatgcc tgattgagat ctttggcatt
ttaaaggagt atgaggtggg 5580tgacccagga cagagaacgc tactggatcc tgggaggttc
agcaaggtgt ctagtccagc 5640tcccatggag ggtggggaag aagaagaaga acttctaggt
cctaaactag aagaggaaga 5700agaagaggaa gtagttgaaa atgatgagga gatagccttt
tcaggcaagg acaagccagc 5760ttcagagaat agtgaggaga agctgatcag taagtttgac
aagcttccag taaagatcgt 5820acagaagaat gatccatttg tggtggactg ctcagataag
cttgggcgtg tgcaggagtt 5880tgacagtggc ctgctgcact ggcggattgg tgggggggac
accactgagc atatccagac 5940ccacttcgag agcaagacag agctgctgcc ttcccggcct
cacgcaccct gcccaccagc 6000ccctcggaag catgtgacaa cagcagaggg tacaccaggg
acaacagacc aggaggggcc 6060cccacctgat ggacctccag aaaaacggat cacagccact
atggatgaca tgttgtctac 6120tcggtctagc accttgaccg aggatggagc taagagttca
gaggccatca aggagagcag 6180caagtttcca tttggcatta gcccagcaca gagccaccgg
aacatcaaga tcctagagga 6240cgaaccccac agtaaggatg agaccccact gtgtaccctt
ctggactggc aggattctct 6300tgccaagcgc tgcgtctgtg tgtccaatac cattcgaagc
ctgtcatttg tgccaggcaa 6360tgactttgag atgtccaaac acccagggct gctgctcatc
ctgggcaagc tgatcctgct 6420gcaccacaag cacccagaac ggaagcaggc accactaact
tatgaaaagg aggaggaaca 6480ggaccaaggg gtgagctgca acaaagtgga gtggtggtgg
gactgcttgg agatgctccg 6540ggaaaacacc ttggttacac tcgccaacat ctcggggcag
ttggacctat ctccataccc 6600cgagagcatt tgcctgcctg tcctggacgg actcctacac
tgggcagttt gcccttcagc 6660tgaagcccag gacccctttt ccaccctggg ccccaatgcc
gtcctttccc cgcagagact 6720ggtcttggaa accctcagca aactcagcat ccaggacaac
aatgtggacc tgattctggc 6780cacacccccc ttcagccgcc tggagaagtt gtatagcact
atggtgcgct tcctcagtga 6840ccgaaagaac ccggtgtgcc gggagatggc tgtggtactg
ctggccaacc tggctcaggg 6900ggacagcctg gcagctcgtg ccattgcagt gcagaagggc
agtatcggca acctcctggg 6960cttcctagag gacagccttg ccgccacaca gttccagcag
agccaggcca gcctcctcca 7020catgcagaac ccaccctttg agccaactag tgtggacatg
atgcggcggg ctgcccgcgc 7080gctgcttgcc ttggccaagg tggacgagaa ccactcagag
tttactctgt acgaatcacg 7140gctgttggac atctcggtat caccgttgat gaactcattg
gtttcacaag tcatttgtga 7200tgtactgttt ttgattggcc agtcatgaca gccgtgggac
acctcccccc cccgtgtgtg 7260tgtgcgtgtg tggagaactt agaaactgac tgttgccctt
tatttatgca aaaccacctc 7320agaatccagt ttaccctgtg ctgtccagct tctcccttgg
gaaaaagtct ctcctgtttc 7380tctctcctcc ttccacctcc cctccctcca tcacctcacg
cctttctgtt ccttgtcctc 7440accttactcc cctcaggacc ctaccccacc ctctttgaaa
agacaaagct ctgcctacat 7500agaagacttt ttttatttta accaaagtta ctgttgttta
cagtgagttt ggggaaaaaa 7560aataaaataa aaatggcttt cccagtcctt gctggctttc
ccagtccttg catcaacggg 7620atgccacatt tcataactgt ttttaatggt aaaaaaaaaa
aaaaaaaata caaaaaaaaa 7680ttctgaagga caaaaaaggt gactgctgaa ctgtgtgtgg
tttattgttg tacattcaca 7740atcttgcagg agccaagaag ttcgcagttg tgaacagacc
ctgttcactg gagaggcctg 7800tgcagtagag tgtagaccct ttcatgtact gtactgtaca
cctgatactg taaacatact 7860gtaataataa tgtctcacat ggaaacagaa aacgctgggt
cagcagcaag ctgtagtttt 7920taaaaatgtt tttagttaaa cgttgaggag aaaaaaaaaa
aaggcttttc ccccaaagta 7980tcatgtgtga acctacaaca ccctgacctc tttctctcct
ccttgattgt atgaataacc 8040ctgagatcac ctcttagaac tggttttaac ctttagctgc
agcggctacg ctgccacgtg 8100tgtatatata tgacgttgta cattgcacat acccttggat
ccccacagtt tggtcctcct 8160cccagctacc cctttatagt atgacgagtt aacaagttgg
tgacctgcac aaagcgagac 8220acagctattt aatctcttgc cagatatcgc ccctcttggt
gcgatgctgt acaggtctct 8280gtaaaaagtc cttgctgtct cagcagccaa tcaacttata
gtttattttt ttctgggttt 8340ttgttttgtt ttgttttctt tctaatcgag gtgtgaaaaa
gttctaggtt cagttgaagt 8400tctgatgaag aaacacaatt gagatttttt cagtgataaa
atctgcatat ttgtatttca 8460acaatgtagc taaaacttga tgtaaattcc tccttttttt
ccttttttgg cttaatgaat 8520atcatttatt cagtatgaaa tctttatact atatgttcca
cgtgttaaga ataaatgtac 8580attaaatctt ggtaa
8595173488DNAHomo sapiens 17taaaaagcat taggcatata
aatgtataaa tatattttat catgtacagt acaaaaatgg 60aaccttatgc atgggcctta
ggaatacagg ctagtatttc agcacagact tccctgcttg 120agttcttgct gatgcttgca
ccgtgacagt gggcaccaac acagacgtgc cacccaaccc 180cctgcacaca ccaccggcca
ccaggggccc ccttgtgcgc cttggcttta taactcctct 240gggggtgata ttggtggtga
tcacagctcc tagcataatg agagttccat ttggtattgt 300cacacgtctc ctgcctcgct
tgggttgcca tgtttgagcg atggccctgt tgatttcacc 360ctgcctttta ctgaatctgt
aaattgttgt gcaattgtgg ttatagtaga ctgtagcaca 420ttgccttttc taaactgcta
catgtttata atcttcattt ttaaagtatg tgtaattttt 480ttaagtatgt attctattca
tatggtctgc ttgtcagtga gccagacttg cttactatat 540tcctttataa taatgctagc
cacttcctgg attctttagt aatgtgctgc atgcaagaac 600tttccagtag cagtgaagga
gggctgcctc tccaagcttc ctaagggatg ctgccctgtg 660tggggatgca ttgcagaggc
actagtagca tgggggctag agtggggagc gagatgtaaa 720agggtggggg gataggagaa
ttccagagtg cttccagcat tagggtcctg agaacttctg 780agttcagaga aacatgcaaa
gtgactaaca aaatagctac ttacctttgc agttctacag 840accctgggag ctgctttggg
agtgagaaag gcaaccctcc aatgtgtttc aactttaaaa 900tgttgaattc ttttcagaca
tggtatctca tttattctcc ttttctagcg tttgttgaat 960ttcaggcaga atgtcttaca
gactgtccta gaaccagatt atcatttaat ctgaaacagc 1020tgaggaaggg acagagaagg
tacaagggca aggcagcaca aaacagatca ggagaatgaa 1080gagggaatgc tttggttttt
tgttttgttt tgttttttct ttttcaagta actaaaacaa 1140catctacatg tagagtgttg
tggagagctg agaccagggt aaagtcaagt gcagcatcag 1200tactgcgaga cccaccagcc
cctggagagg gtcagccgag aatctggtag tgaagcctgt 1260ctagggtccc ggcaccctca
ccctcagcca cctgcagaga ggccagggcc ccagagacta 1320gcctggttct gaagtgggca
ggggtgctgc cagagccctc tgccccttat gttgagaccc 1380tgctttcagg acaggccagc
cgttggccac catgtcacat tctgagtgag tgtcacaggt 1440ccctaacaat aattttctga
tctggagcat atcagcagaa tgcttagcct caaggggcct 1500ggaagctgta atgtttgatt
tatgatgaga actatccgag gccacccttg gcctctaaat 1560aagctgctct agggagccgc
ctactttttg atgagaaatt agaagagtac ctaatgttga 1620aaacatgaca tgcgctcttg
ggatctgctg ttctctccag ggctccagaa cctgatacct 1680gttaccaaag ctaggaaaga
gctttatcac aagccttcac tgtcctggca tgagaactgg 1740ctgccaggct cagtgtaccc
cattaactgt gaatgaatct gagcttggtt tcctttattg 1800cttcctctgc aatatgattg
ctgaaacaca ttttaaaaat tcagaagctt gtcactcctg 1860ttaatgggag gatcagtcac
acatgtgtag tacaaggcgg actttgtgtt tgtttttggt 1920gttaattttt agcattgtgt
gtgttgcttc cccaccctga ggagaggaca ccatggctta 1980ctactcagga caagtatgcc
ccgctcaggg tgtgatttca ggtggcttcc aaacttgtac 2040gcagtttaaa gatggtgggg
acagactttg cctctaccta gtgaacccca cttaaagaat 2100aaggagcatt tgaatctctt
ggaaaaggcc atgaagaata aagcagtcaa aaagaagtcc 2160tccatgttgg tgccaaggac
ttgcgagggg aaataaaaat gttatccagc ctgaccaaca 2220tggagaaacc ccgtctccat
taaaaataca aaattagcct ggcatggtgg cgcatgcctg 2280taatcccagc tactctggag
gctgaggcag gagaatcgct tgaacccagg aggcggaggt 2340cgcagtgagc cgagatcatg
ccagtgcact ccagcctggg taacaagagt gaaactccgt 2400gtcaaaaaaa aaaaaaaaaa
atgttactca tcctctctga aagcaaaaag gaaaccctaa 2460cagctctgaa ctctggtttt
atttttcttg ctgtatttgg gtgaacattg tatgattagg 2520cataatgtta aaaaaaaaaa
attttttttt ggtagaaatg caatcaccag taaagaggta 2580cgaaaaagct agcctctctc
agagaccggg gaggcagagt actactagag gaagtgaagt 2640tctgatggaa tcatgcctgt
caaatgaggt cttgaagcgg atgcccaaat aaaagagtat 2700attttatcta aatcttaagt
gggtaacatt ttatgcagtt taaatgaatg gaatattttc 2760ctcttgttta gttgtatctg
tttgtatttt tctttgatga atgattggtc atgaggcctc 2820ttgccacact ccagaaatac
gtgtgcggct gcttttaaga actatgtgtc tggtcactta 2880tttctctaaa attatctcat
tgcctggcaa tcagtcttct cttgtatact tgtcctagca 2940cattatgtac atgggaaatg
taaacaaatg tgaaggagga ccagaaaaat tagttaatat 3000ttaaaaaaat gtattgtgca
ttttggcttc acatgtttaa ctttttttaa gaaaaaagtt 3060gcatgaatgg aaaaaaaaat
ctgtatacag tatctgtaaa aactatctta tctgtttcaa 3120ttccttgctc atatcccata
taatctagaa ctaaatatgg tgtgtggcca tatttaaaca 3180cctgagagtc aagcagttga
gactttgatt tgaagcacct catccttctt tcaatgcgaa 3240cactatcata tggcattctt
actgaggatt ttgtctaacc atatgttgcc atgaattaac 3300tctgccgcct ttcttaagga
tcaaaaccag tttgatttgg gaatcttccc ctttccaaat 3360gaaatagaga tgcagtactt
aactttcctt ggtgtttgta gatattgcct tgtgtattcc 3420acttaaaacc gtaatctagt
ttgtaaaaga gatggtgacg catgtaaata aagcatcagt 3480gacactct
34881820DNAHomo sapiens
18gcctaccgaa gacaaaggcg
201920DNAHomo sapiens 19tagaggctgt cccctaggag
202020DNAHomo sapiens 20agaggcaccc tatgggccag
202120DNAHomo sapiens
21catctctggc gctggcattg
202220DNAHomo sapiens 22gcactgatcc caatcctcgc
202320DNAHomo sapiens 23ccctgcataa gcacaatggg
202420DNAHomo sapiens
24gggaaggaga atgttgcccc
202520DNAHomo sapiens 25gaggagggaa ccacccctac
202620DNAHomo sapiens 26gggaggctga gggaagggac
202720DNAHomo sapiens
27ggggaaatgc gtagaccagg
202820DNAHomo sapiens 28cccgcctcct cctaagtctg
202920DNAHomo sapiens 29cagcatgagc caccatgccc
203020DNAHomo sapiens
30gaaccagaga cctaggccgc
203120DNAHomo sapiens 31cagctcctct agggagaccc
203220DNAHomo sapiens 32ctagagccta agttgaaccc
203320DNAHomo sapiens
33gtggtggtgg tttatgaggg
203420DNAHomo sapiens 34taggacatgc ccatgtccgc
203520DNAHomo sapiens 35tccgctcttc ctcaactccc
203620DNAHomo sapiens
36ctctttgggt cttcccttcc
203720DNAHomo sapiens 37cttcagaggc aaagtccttg
203820DNAHomo sapiens 38ctcggctcgg cccaaagcag
203920DNAHomo sapiens
39gtaggtcccc gggaggcagg
204021DNAHomo sapiens 40gttttacggc ttgcaagtaa c
214120DNAHomo sapiens 41ccaaacttgt ctggaacacc
204220DNAHomo sapiens
42ccagggtctc aaatggcaac
204320DNAHomo sapiens 43atgtggcata ctcgcccacc
204420DNAHomo sapiens 44ctctgccgag ccttggactg
204520DNAHomo sapiens
45gcatggccca gctcgtgcac
204620DNAHomo sapiens 46tgcccgatga tgaccgccgg
204720DNAHomo sapiens 47gggttgaggg ggcatcttgg
204820DNAHomo sapiens
48caccgtggcc atgctccgag
204920DNAHomo sapiens 49ccatcacttg gttattcctc
205020DNAHomo sapiens 50cctacgagag gaaggatggc
205120DNAHomo sapiens
51ggttccaggg acgcctcatc
205220DNAHomo sapiens 52ggatgcccaa tggaaagacc
205320DNAHomo sapiens 53cgctatccac tgccctgagc
205420DNAHomo sapiens
54gggatccctg agtctcccag
205520DNAHomo sapiens 55tgttgaattt ccagtatttg
205620DNAHomo sapiens 56tattagtaaa ctggcccttc
205720DNAHomo sapiens
57atctttcttc tgcttagtcg
205820DNAHomo sapiens 58tagaggtgga actaaacccc
2059286DNAHomo sapiens 59gggaggctga gggaagggac
tcaggctgct atcgtcactg tccccatcct tccaggaaat 60gacatcttcc tggtggctgt
gcacgagctg ggccatgccc tggggctcga gcattccagt 120gacccctcgg ccatcatggc
acccttttac cagtggatgg acacggagaa ttttgtgctg 180cccgatgatg accgccgggg
catccagcaa ctttatggcg agtagtctac acccacgcct 240gctccctcct ctgctgcttg
ttccctcctg gtctacgcat ttcccc 28660286DNAHomo sapiens
60gggaggctga gggaagggac tcaggctgct atcgtcactg tccccatcct tccaggaaat
60gacatcttcc tggtggctgt gcacgagctg ggccatgccc tggggctcga gcattccagt
120gacccctcgg ccatcatggc accgttttac cagtggatgg acacggagaa ttttgtgctg
180cccgatgatg accgccgggg catccagcaa ctttatggcg agtagtctac acccacgcct
240gctccctcct ctgctgcttg ttccctcctg gtctacgcat ttcccc
28661286DNAHomo sapiens 61gggaggctga gggaagggac tcaggctgct atcgtcactg
tccccatcct tccaggaaat 60gacatcttcc tggtggctgt gcacgagctg ggccatgccc
tggggctcga gcattccagt 120gacccctcgg ccatcatggc accgttttac cagtggatgg
acacggagaa ttttgtgctg 180cccaatgatg accgccgggg catccagcaa ctttatggcg
agtagtctac acccacgcct 240gctccctcct ctgctgcttg ttccctcctg gtctacgcat
ttcccc 286627318DNAHomo sapiens 62ctggctctta acggcgttta
tgtcctttgc tgtctgaggg gcctcagctc tgaccaatct 60ggtcttcgtg tggtcattag
catgggcttc gtgagacaga tacagctttt gctctggaag 120aactggaccc tgcggaaaag
gcaaaagatt cgctttgtgg tggaactcgt gtggccttta 180tctttatttc tggtcttgat
ctggttaagg aatgccaacc cgctctacag ccatcatgaa 240tgccatttcc ccaacaaggc
gatgccctca gcaggaatgc tgccgtggct ccaggggatc 300ttctgcaatg tgaacaatcc
ctgttttcaa agccccaccc caggagaatc tcctggaatt 360gtgtcaaact ataacaactc
catcttggca agggtatatc gagattttca agaactcctc 420atgaatgcac cagagagcca
gcaccttggc cgtatttgga cagagctaca catcttgtcc 480caattcatgg acaccctccg
gactcacccg gagagaattg caggaagagg aatacgaata 540agggatatct tgaaagatga
agaaacactg acactatttc tcattaaaaa catcggcctg 600tctgactcag tggtctacct
tctgatcaac tctcaagtcc gtccagagca gttcgctcat 660ggagtcccgg acctggcgct
gaaggacatc gcctgcagcg aggccctcct ggagcgcttc 720atcatcttca gccagagacg
cggggcaaag acggtgcgct atgccctgtg ctccctctcc 780cagggcaccc tacagtggat
agaagacact ctgtatgcca acgtggactt cttcaagctc 840ttccgtgtgc ttcccacact
cctagacagc cgttctcaag gtatcaatct gagatcttgg 900ggaggaatat tatctgatat
gtcaccaaga attcaagagt ttatccatcg gccgagtatg 960caggacttgc tgtgggtgac
caggcccctc atgcagaatg gtggtccaga gacctttaca 1020aagctgatgg gcatcctgtc
tgacctcctg tgtggctacc ccgagggagg tggctctcgg 1080gtgctctcct tcaactggta
tgaagacaat aactataagg cctttctggg gattgactcc 1140acaaggaagg atcctatcta
ttcttatgac agaagaacaa catccttttg taatgcattg 1200atccagagcc tggagtcaaa
tcctttaacc aaaatcgctt ggagggcggc aaagcctttg 1260ctgatgggaa aaatcctgta
cactcctgat tcacctgcag cacgaaggat actgaagaat 1320gccaactcaa cttttgaaga
actggaacac gttaggaagt tggtcaaagc ctgggaagaa 1380gtagggcccc agatctggta
cttctttgac aacagcacac agatgaacat gatcagagat 1440accctgggga acccaacagt
aaaagacttt ttgaataggc agcttggtga agaaggtatt 1500actgctgaag ccatcctaaa
cttcctctac aagggccctc gggaaagcca ggctgacgac 1560atggccaact tcgactggag
ggacatattt aacatcactg atcgcaccct ccgcctggtc 1620aatcaatacc tggagtgctt
ggtcctggat aagtttgaaa gctacaatga tgaaactcag 1680ctcacccaac gtgccctctc
tctactggag gaaaacatgt tctgggccgg agtggtattc 1740cctgacatgt atccctggac
cagctctcta ccaccccacg tgaagtataa gatccgaatg 1800gacatagacg tggtggagaa
aaccaataag attaaagaca ggtattggga ttctggtccc 1860agagctgatc ccgtggaaga
tttccggtac atctggggcg ggtttgccta tctgcaggac 1920atggttgaac aggggatcac
aaggagccag gtgcaggcgg aggctccagt tggaatctac 1980ctccagcaga tgccctaccc
ctgcttcgtg gacgattctt tcatgatcat cctgaaccgc 2040tgtttcccta tcttcatggt
gctggcatgg atctactctg tctccatgac tgtgaagagc 2100atcgtcttgg agaaggagtt
gcgactgaag gagaccttga aaaatcaggg tgtctccaat 2160gcagtgattt ggtgtacctg
gttcctggac agcttctcca tcatgtcgat gagcatcttc 2220ctcctgacga tattcatcat
gcatggaaga atcctacatt acagcgaccc attcatcctc 2280ttcctgttct tgttggcttt
ctccactgcc accatcatgc tgtgctttct gctcagcacc 2340ttcttctcca aggccagtct
ggcagcagcc tgtagtggtg tcatctattt caccctctac 2400ctgccacaca tcctgtgctt
cgcctggcag gaccgcatga ccgctgagct gaagaaggct 2460gtgagcttac tgtctccggt
ggcatttgga tttggcactg agtacctggt tcgctttgaa 2520gagcaaggcc tggggctgca
gtggagcaac atcgggaaca gtcccacgga aggggacgaa 2580ttcagcttcc tgctgtccat
gcagatgatg ctccttgatg ctgcgtgcta tggcttactc 2640gcttggtacc ttgatcaggt
gtttccagga gactatggaa ccccacttcc ttggtacttt 2700cttctacaag agtcgtattg
gcttagcggt gaagggtgtt caaccagaga agaaagagcc 2760ctggaaaaga ccgagcccct
aacagaggaa acggaggatc cagagcaccc agaaggaata 2820cacgactcct tctttgaacg
tgagcatcca gggtgggttc ctggggtatg cgtgaagaat 2880ctggtaaaga tttttgagcc
ctgtggccgg ccagctgtgg accgtctgaa catcaccttc 2940tacgagaacc agatcaccgc
attcctgggc cacaatggag ctgggaaaac caccaccttg 3000tccatcctga cgggtctgtt
gccaccaacc tctgggactg tgctcgttgg gggaagggac 3060attgaaacca gcctggatgc
agtccggcag agccttggca tgtgtccaca gcacaacatc 3120ctgttccacc acctcacggt
ggctgagcac atgctgttct atgcccagct gaaaggaaag 3180tcccaggagg aggcccagct
ggagatggaa gccatgttgg aggacacagg cctccaccac 3240aagcggaatg aagaggctca
ggacctatca ggtggcatgc agagaaagct gtcggttgcc 3300attgcctttg tgggagatgc
caaggtggtg attctggacg aacccacctc tggggtggac 3360ccttactcga gacgctcaat
ctgggatctg ctcctgaagt atcgctcagg cagaaccatc 3420atcatgccca ctcaccacat
ggacgaggcc gaccaccaag gggaccgcat tgccatcatt 3480gcccagggaa ggctctactg
ctcaggcacc ccactcttcc tgaagaactg ctttggcaca 3540ggcttgtact taaccttggt
gcgcaagatg aaaaacatcc agagccaaag gaaaggcagt 3600gaggggacct gcagctgctc
gtctaagggt ttctccacca cgtgtccagc ccacgtcgat 3660gacctaactc cagaacaagt
cctggatggg gatgtaaatg agctgatgga tgtagttctc 3720caccatgttc cagaggcaaa
gctggtggag tgcattggtc aagaacttat cttccttctt 3780ccaaataaga acttcaagca
cagagcatat gccagccttt tcagagagct ggaggagacg 3840ctggctgacc ttggtctcag
cagttttgga atttctgaca ctcccctgga agagattttt 3900ctgaaggtca cggaggattc
tgattcagga cctctgtttg cgggtggcgc tcagcagaaa 3960agagaaaacg tcaacccccg
acacccctgc ttgggtccca gagagaaggc tggacagaca 4020ccccaggact ccaatgtctg
ctccccaggg gcgccggctg ctcacccaga gggccagcct 4080cccccagagc cagagtgccc
aggcccgcag ctcaacacgg ggacacagct ggtcctccag 4140catgtgcagg cgctgctggt
caagagattc caacacacca tccgcagcca caaggacttc 4200ctggcgcaga tcgtgctccc
ggctaccttt gtgtttttgg ctctgatgct ttctattgtt 4260atccttcctt ttggcgaata
ccccgctttg acccttcacc cctggatata tgggcagcag 4320tacaccttct tcagcatgga
tgaaccaggc agtgagcagt tcacggtact tgcagacgtc 4380ctcctgaata agccaggctt
tggcaaccgc tgcctgaagg aagggtggct tccggagtac 4440ccctgtggca actcaacacc
ctggaagact ccttctgtgt ccccaaacat cacccagctg 4500ttccagaagc agaaatggac
acaggtcaac ccttcaccat cctgcaggtg cagcaccagg 4560gagaagctca ccatgctgcc
agagtgcccc gagggtgccg ggggcctccc gcccccccag 4620agaacacagc gcagcacgga
aattctacaa gacctgacgg acaggaacat ctccgacttc 4680ttggtaaaaa cgtatcctgc
tcttataaga agcagcttaa agagcaaatt ctgggtcaat 4740gaacagaggt atggaggaat
ttccattgga ggaaagctcc cagtcgtccc catcacgggg 4800gaagcacttg ttgggttttt
aagcgacctt ggccggatca tgaatgtgag cgggggccct 4860atcactagag aggcctctaa
agaaatacct gatttcctta aacatctaga aactgaagac 4920aacattaagg tgtggtttaa
taacaaaggc tggcatgccc tggtcagctt tctcaatgtg 4980gcccacaacg ccatcttacg
ggccagcctg cctaaggaca ggagccccga ggagtatgga 5040atcaccgtca ttagccaacc
cctgaacctg accaaggagc agctctcaga gattacagtg 5100ctgaccactt cagtggatgc
tgtggttgcc atctgcgtga ttttctccat gtccttcgtc 5160ccagccagct ttgtccttta
tttgatccag gagcgggtga acaaatccaa gcacctccag 5220tttatcagtg gagtgagccc
caccacctac tgggtgacca acttcctctg ggacatcatg 5280aattattccg tgagtgctgg
gctggtggtg ggcatcttca tcgggtttca gaagaaagcc 5340tacacttctc cagaaaacct
tcctgccctt gtggcactgc tcctgctgta tggatgggcg 5400gtcattccca tgatgtaccc
agcatccttc ctgtttgatg tccccagcac agcctatgtg 5460gctttatctt gtgctaatct
gttcatcggc atcaacagca gtgctattac cttcatcttg 5520gaattatttg ataataaccg
gacgctgctc aggttcaacg ccgtgctgag gaagctgctc 5580attgtcttcc cccacttctg
cctgggccgg ggcctcattg accttgcact gagccaggct 5640gtgacagatg tctatgcccg
gtttggtgag gagcactctg caaatccgtt ccactgggac 5700ctgattggga agaacctgtt
tgccatggtg gtggaagggg tggtgtactt cctcctgacc 5760ctgctggtcc agcgccactt
cttcctctcc caatggattg ccgagcccac taaggagccc 5820attgttgatg aagatgatga
tgtggctgaa gaaagacaaa gaattattac tggtggaaat 5880aaaactgaca tcttaaggct
acatgaacta accaagattt atctgggcac ctccagccca 5940gcagtggaca ggctgtgtgt
cggagttcgc cctggagagt gctttggcct cctgggagtg 6000aatggtgccg gcaaaacaac
cacattcaag atgctcactg gggacaccac agtgacctca 6060ggggatgcca ccgtagcagg
caagagtatt ttaaccaata tttctgaagt ccatcaaaat 6120atgggctact gtcctcagtt
tgatgcaatc gatgagctgc tcacaggacg agaacatctt 6180tacctttatg cccggcttcg
aggtgtacca gcagaagaaa tcgaaaaggt tgcaaactgg 6240agtattaaga gcctgggcct
gactgtctac gccgactgcc tggctggcac gtacagtggg 6300ggcaacaagc ggaaactctc
cacagccatc gcactcattg gctgcccacc gctggtgctg 6360ctggatgagc ccaccacagg
gatggacccc caggcacgcc gcatgctgtg gaacgtcatc 6420gtgagcatca tcagaaaagg
gagggctgtg gtcctcacat cccacagcat ggaagaatgt 6480gaggcactgt gtacccggct
ggccatcatg gtaaagggcg cctttcgatg tatgggcacc 6540attcagcatc tcaagtccaa
atttggagat ggctatatcg tcacaatgaa gatcaaatcc 6600ccgaaggacg acctgcttcc
tgacctgaac cctgtggagc agttcttcca ggggaacttc 6660ccaggcagtg tgcagaggga
gaggcactac aacatgctcc agttccaggt ctcctcctcc 6720tccctggcga ggatcttcca
gctcctcctc tcccacaagg acagcctgct catcgaggag 6780tactcagtca cacagaccac
actggaccag gtgtttgtaa attttgctaa acagcagact 6840gaaagtcatg acctccctct
gcaccctcga gctgctggag ccagtcgaca agcccaggac 6900tgatctttca caccgctcgt
tcctgcagcc agaaaggaac tctgggcagc tggaggcgca 6960ggagcctgtg cccatatggt
catccaaatg gactggccca gcgtaaatga ccccactgca 7020gcagaaaaca aacacacgag
gagcatgcag cgaattcaga aagaggtctt tcagaaggaa 7080accgaaactg acttgctcac
ctggaacacc tgatggtgaa accaaacaaa tacaaaatcc 7140ttctccagac cccagaacta
gaaaccccgg gccatcccac tagcagcttt ggcctccata 7200ttgctctcat ttcaagcaga
tctgcttttc tgcatgtttg tctgtgtgtc tgcgttgtgt 7260gtgattttca tggaaaaata
aaatgcaaat gcactcatca caaaaaaaaa aaaaaaaa 7318631156DNAHomo sapiens
63cgcagcggag gtgaaggacg tccttcccca ggagccgact ggccaatcac aggcaggaag
60atgaaggttc tgtgggctgc gttgctggtc acattcctgg caggatgcca ggccaaggtg
120gagcaagcgg tggagacaga gccggagccc gagctgcgcc agcagaccga gtggcagagc
180ggccagcgct gggaactggc actgggtcgc ttttgggatt acctgcgctg ggtgcagaca
240ctgtctgagc aggtgcagga ggagctgctc agctcccagg tcacccagga actgagggcg
300ctgatggacg agaccatgaa ggagttgaag gcctacaaat cggaactgga ggaacaactg
360accccggtgg cggaggagac gcgggcacgg ctgtccaagg agctgcaggc ggcgcaggcc
420cggctgggcg cggacatgga ggacgtgtgc ggccgcctgg tgcagtaccg cggcgaggtg
480caggccatgc tcggccagag caccgaggag ctgcgggtgc gcctcgcctc ccacctgcgc
540aagctgcgta agcggctcct ccgcgatgcc gatgacctgc agaagcgcct ggcagtgtac
600caggccgggg cccgcgaggg cgccgagcgc ggcctcagcg ccatccgcga gcgcctgggg
660cccctggtgg aacagggccg cgtgcgggcc gccactgtgg gctccctggc cggccagccg
720ctacaggagc gggcccaggc ctggggcgag cggctgcgcg cgcggatgga ggagatgggc
780agccggaccc gcgaccgcct ggacgaggtg aaggagcagg tggcggaggt gcgcgccaag
840ctggaggagc aggcccagca gatacgcctg caggccgagg ccttccaggc ccgcctcaag
900agctggttcg agcccctggt ggaagacatg cagcgccagt gggccgggct ggtggagaag
960gtgcaggctg ccgtgggcac cagcgccgcc cctgtgccca gcgacaatca ctgaacgccg
1020aagcctgcag ccatgcgacc ccacgccacc ccgtgcctcc tgcctccgcg cagcctgcag
1080cgggagaccc tgtccccgcc ccagccgtcc tcctggggtg gaccctagtt taataaagat
1140tcaccaagtt tcacgc
1156642273DNAHomo sapiens 64caggactgcc tgagacaagc cacaagctga acagagaaag
tggattgaac aaggacgcat 60ttccccagta catccacaac atgctgtcca catctcgttc
tcggtttatc agaaatacca 120acgagagcgg tgaagaagtc accacctttt ttgattatga
ttacggtgct ccctgtcata 180aatttgacgt gaagcaaatt ggggcccaac tcctgcctcc
gctctactcg ctggtgttca 240tctttggttt tgtgggcaac atgctggtcg tcctcatctt
aataaactgc aaaaagctga 300agtgcttgac tgacatttac ctgctcaacc tggccatctc
tgatctgctt tttcttatta 360ctctcccatt gtgggctcac tctgctgcaa atgagtgggt
ctttgggaat gcaatgtgca 420aattattcac agggctgtat cacatcggtt attttggcgg
aatcttcttc atcatcctcc 480tgacaatcga tagatacctg gctattgtcc atgctgtgtt
tgctttaaaa gccaggacgg 540tcacctttgg ggtggtgaca agtgtgatca cctggttggt
ggctgtgttt gcttctgtcc 600caggaatcat ctttactaaa tgccagaaag aagattctgt
ttatgtctgt ggcccttatt 660ttccacgagg atggaataat ttccacacaa taatgaggaa
cattttgggg ctggtcctgc 720cgctgctcat catggtcatc tgctactcgg gaatcctgaa
aaccctgctt cggtgtcgaa 780acgagaagaa gaggcatagg gcagtgagag tcatcttcac
catcatgatt gtttactttc 840tcttctggac tccctataac attgtcattc tcctgaacac
cttccaggaa ttcttcggcc 900tgagtaactg tgaaagcacc agtcaactgg accaagccac
gcaggtgaca gagactcttg 960ggatgactca ctgctgcatc aatcccatca tctatgcctt
cgttggggag aagttcagaa 1020gcctttttca catagctctt ggctgtagga ttgccccact
ccaaaaacca gtgtgtggag 1080gtccaggagt gagaccagga aagaatgtga aagtgactac
acaaggactc ctcgatggtc 1140gtggaaaagg aaagtcaatt ggcagagccc ctgaagccag
tcttcaggac aaagaaggag 1200cctagagaca gaaatgacag atctctgctt tggaaatcac
acgtctggct tcacagatgt 1260gtgattcaca gtgtgaatct tggtgtctac gttaccaggc
aggaaggctg agaggagaga 1320gactccagct gggttggaaa acagtatttt ccaaactacc
ttccagttcc tcatttttga 1380atacaggcat agagttcaga ctttttttaa atagtaaaaa
taaaattaaa gctgaaaact 1440gcaacttgta aatgtggtaa agagttagtt tgagttgcta
tcatgtcaaa cgtgaaaatg 1500ctgtattagt cacagagata attctagctt tgagcttaag
aattttgagc aggtggtatg 1560tttgggagac tgctgagtca acccaatagt tgttgattgg
caggagttgg aagtgtgtga 1620tctgtgggca cattagccta tgtgcatgca gcatctaagt
aatgatgtcg tttgaatcac 1680agtatacgct ccatcgctgt catctcagct ggatctccat
tctctcaggc ttgctgccaa 1740aagccttttg tgttttgttt tgtatcatta tgaagtcatg
cgtttaatca cattcgagtg 1800tttcagtgct tcgcagatgt ccttgatgct catattgttc
cctaatttgc cagtgggaac 1860tcctaaatca aattggcttc taatcaaagc ttttaaaccc
tattggtaaa gaatggaagg 1920tggagaagct ccctgaagta agcaaagact ttcctcttag
tcgagccaag ttaagaatgt 1980tcttatgttg cccagtgtgt ttctgatctg atgcaagcaa
gaaacactgg gcttctagaa 2040ccaggcaact tgggaactag actcccaagc tggactatgg
ctctactttc aggccacatg 2100gctaaagaag gtttcagaaa gaagtgggga cagagcagaa
ctttcacctt catatatttg 2160tatgatccta atgaatgcat aaaatgttaa gttgatggtg
atgaaatgta aatactgttt 2220ttaacaacta tgatttggaa aataaatcaa tgctataact
atgttgataa aag 227365818DNAHomo sapiens 65cgcagcgggt cctctctatc
tagctccagc ctctcgcctg cgccccactc cccgcgtccc 60gcgtcctagc cgaccatggc
cgggcccctg cgcgccccgc tgctcctgct ggccatcctg 120gccgtggccc tggccgtgag
ccccgcggcc ggctccagtc ccggcaagcc gccgcgcctg 180gtgggaggcc ccatggacgc
cagcgtggag gaggagggtg tgcggcgtgc actggacttt 240gccgtcggcg agtacaacaa
agccagcaac gacatgtacc acagccgcgc gctgcaggtg 300gtgcgcgccc gcaagcagat
cgtagctggg gtgaactact tcttggacgt ggagctgggc 360cgaaccacgt gtaccaagac
ccagcccaac ttggacaact gccccttcca tgaccagcca 420catctgaaaa ggaaagcatt
ctgctctttc cagatctacg ctgtgccttg gcagggcaca 480atgaccttgt cgaaatccac
ctgtcaggac gcctaggggt ctgtaccggg ctggcctgtg 540cctatcacct cttatgcaca
cctcccaccc cctgtattcc cacccctgga ctggtggccc 600ctgccttggg gaaggtctcc
ccatgtgcct gcaccaggag acagacagag aaggcagcag 660gcggcctttg ttgctcagca
aggggctctg ccctccctcc ttccttcttg cttctcatag 720ccccggtgtg cggtgcatac
acccccacct cctgcaataa aatagtagca tcggcaaaaa 780aaaaaaaaaa aaaaaaaaaa
aaaaaaaaaa aaaaaaaa 8186618209DNAHomo sapiens
66gaagccgcat ccagacaaaa gctgccgcat ccctgccctg cccaacccct ggagggattc
60gagtttggtg cttgtccccg tctgattctc agcgccaaac tttttgctag ttcagagatt
120ccaagagtct gatgagttac tctgagagga aaccctctgc ctgttgttga ggaggactga
180gcacagtgct taggcgctga gggggaaaaa gagggggaaa aaaaagaaaa tgatttcctg
240ggaagttgtc catacagtat tcctgtttgc tcttctttat tcttccctag ctcaagatgc
300gagcccccag tcagagatca gagctgagga aattcccgag ggggcctcca cgttggcttt
360tgtgtttgat gtgactggtt ctatgtatga tgatttagtt caggtgattg aaggggcttc
420caaaattttg gagacgtctt tgaaaagacc taaaagacct cttttcaact ttgcgttggt
480gcctttccat gatccagaaa ttggcccagt gacaattacc acagatccca agaaatttca
540atatgaactc agagaactgt atgttcaggg tggtggtgat tgcccagaaa tgagtattgg
600agctataaaa attgccttgg aaatttctct tcctggttct ttcatctatg ttttcactga
660tgctcggtcc aaagattacc ggctcaccca tgaggtgctg caacttatcc aacagaaaca
720gtcacaagtc gtatttgttc tgactggaga ttgtgatgac aggacccata ttggatataa
780agtctatgaa gaaattgcct ctacaagttc tggtcaagtg ttccatctgg acaaaaaaca
840agttaatgag gtattaaaat gggtagaaga agcagtacag gcctccaaag ttcacctttt
900atccacagat catttggaac aggctgtaaa tacttggaga attccttttg atcccagcct
960gaaagaggtc actgtgtctt tgagtgggcc ttctccaatg attgaaattc gcaatccttt
1020agggaagctg ataaaaaagg gatttggcct gcatgagcta ttaaatatcc ataactctgc
1080caaagtagtg aatgtgaaag agccagaggc tggaatgtgg acagtgaaga cctcaagcag
1140tggaaggcac tctgttcgca ttactggcct cagtactatt gatttccgag ctggcttttc
1200tcgaaagccc accctggact tcaaaaaaac agtcagcaga ccagtgcaag gaatacctac
1260ctatgtactg ctcaatactt ctggaatttc cactccagct agaatagatc ttcttgaact
1320tttgagtatc tcaggaagtt ctcttaagac tattcctgtt aaatattacc cacatcgaaa
1380accttatggc atatggaata tttctgactt tgtaccacca aatgaagctt tctttctcaa
1440agtaacaggc tatgataaag atgattacct cttccagaga gtatcaagtg tttccttttc
1500tagtattgtc ccagatgctc ccaaagttac gatgcctgag aaaaccccag gatactatct
1560gcagccgggc caaattccct gctctgttga cagtcttttg ccctttacct tgagctttgt
1620cagaaatgga gttacacttg gagtagacca gtatttgaaa gaatctgcca gtgtgaactt
1680agatattgca aaggtcactt tgtctgacga aggtttctat gaatgcattg ctgtcagcag
1740tgcaggtact ggacgggcac agacattttt tgacgtatca gagccccctc cggtcatcca
1800agtgcctaac aatgttacag tcactcctgg agagagagca gttttaacat gtctcatcat
1860cagtgcggtg gattacaatc taacctggca gaggaatgac agagatgtca gactggcaga
1920gccagcgaga attaggacct tggctaatct gtcattggag ctaaagagtg tgaaattcaa
1980cgatgctgga gagtatcatt gtatggtttc tagtgaaggt ggatcatcag ccgcttcagt
2040tttcctcaca gtgcaagaac cacccaaagt cactgtgatg cccaagaatc agtctttcac
2100aggagggtct gaggtctcca tcatgtgttc tgcaacaggt tatcccaaac caaagattgc
2160ctggaccgtt aacgatatgt ttatcgtggg ttcacacagg tataggatga cctcagatgg
2220taccttattt atcaaaaatg cagctcccaa agatgcaggg atctatggtt gcctagcaag
2280taattcagct ggaacagata aacagaattc tactctcaga tacattgaag cccctaagtt
2340gatggtagtt cagagtgagc tcttggttgc ccttggggat ataaccgtta tggaatgcaa
2400aacctctggt attcctccac ctcaagttaa atggttcaaa ggagatcttg agttgaggcc
2460ctcaacattc ctcattattg accctctctt gggacttttg aagattcaag aaacacaaga
2520tctggatgct ggcgattata cctgtgtagc catcaatgag gctggaagag caactggcaa
2580gataactctg gatgttggct cacctccagt tttcatacaa gaacctgctg atgtgtctat
2640ggaaattggc tcaaatgtga cattaccttg ttatgttcag ggttatccag aaccaacaat
2700caaatggcga agattagaca acatgccaat tttctcaaga cctttttcag ttagttccat
2760cagccaacta agaacaggag ctctctttat tttaaactta tgggcaagtg ataaaggaac
2820ctatatttgt gaagctgaaa accagtttgg aaagatccag tcagagacaa cagtaacagt
2880gaccggactt gttgctccac ttattggaat cagcccttca gtggccaatg ttattgaagg
2940acagcagctt actttgccct gtactctgtt agctggaaat cccattccag aacgtcggtg
3000gattaagaat tcagctatgt tgctccaaaa tccttacatc actgtgcgca gtgatgggag
3060cctccatatt gaaagagttc agcttcagga tggtggtgaa tatacttgtg tggccagtaa
3120cgttgctggg accaataaca aaactacctc tgtggttgtg catgttctgc caaccattca
3180gcatgggcag cagatactca gtacaattga aggcattcca gtaactttac catgcaaagc
3240aagtggaaat cccaaaccgt ctgtcatctg gtccaagaaa ggagagctga tttcaaccag
3300cagtgctaag ttttcagcag gagctgatgg tagtctgtat gtggtatcac ctggaggaga
3360ggagagtggg gagtatgtct gcactgccac caatacagcc ggctacgcca aaaggaaagt
3420gcagctaaca gtctatgtaa ggcccagagt gtttggagat caacgaggac tgtcccagga
3480taagcctgtt gagatctccg tccttgcagg ggaagaggta acacttccat gtgaagtgaa
3540gagcttacct ccacccataa ttacttgggc caaagaaacc cagctcatct caccgttctc
3600tccaagacac acattcctcc cttctggttc aatgaagatc actgaaaccc gcacttcaga
3660tagtgggatg tatctttgtg ttgccacaaa tattgctggg aatgtgactc aggctgtcaa
3720attaaatgtc catgttcctc caaagataca gcgtggacct aaacatctca aagtccaagt
3780tggtcaaaga gtggatattc catgtaatgc tcaagggact cctcttcctg taatcacctg
3840gtccaaaggt ggaagcacta tgctggttga tggagagcac catgttagca atccagacgg
3900aactttaagc atcgaccaag ccacgccctc agatgctggc atatatacat gtgttgctac
3960taacatagca ggcactgatg aaacagagat aacgctacat gtccaagaac cacccacagt
4020ggaagatcta gaacctccat ataacactac tttccaagaa agagtggcca atcaacgcat
4080tgaatttcca tgtcctgcaa aaggtacccc taaaccaacc atcaaatggt tacacaatgg
4140tagagagttg acaggcagag agcctggcat ttctatcttg gaagatggca cattgctggt
4200tattgcttct gttacaccct atgacaatgg ggagtacatc tgtgtggcag tcaatgaagc
4260tggaaccaca gaaagaaaat ataacctcaa agtccatgtt cctccagtaa ttaaagataa
4320agaacaagtt acaaatgtgt cggtgttgtt aaatcagctg accaatctct tctgtgaagt
4380ggaaggcact ccatctccca tcattatgtg gtataaagat aatgtccagg tgactgaaag
4440cagcactatt cagactgtga acaatgggaa gatactgaag ctcttcagag ccactccaga
4500ggatgcagga agatattcct gcaaagcaat taatattgca ggcacttctc agaagtactt
4560taacattgat gtgctagttc cacccaccat aataggtacc aacttcccaa atgaagtctc
4620agttgtcctc aaccgtgacg tcgcccttga atgccaggtc aaaggcactc cctttcctga
4680tattcattgg ttcaaagatg gcaagccttt atttttgggc gatcctaatg ttgaacttct
4740agacagagga caagtcttac atttaaagaa tgcacggaga aatgacaagg ggcgctacca
4800atgtactgtg tctaatgcag ctggcaaaca agccaaggat ataaaactga ctatctatat
4860tccacctagt attaaaggag gaaatgtcac cacagacata tcagtattga tcaacagcct
4920tattaaactg gaatgtgaaa cacggggact tccaatgcct gccattactt ggtataagga
4980cgggcagcca atcatgtcca gctcacaagc actttatatt gataaaggac aatatcttca
5040tattcctcga gcacaggtct ctgattcagc aacatatacg tgtcacgtag ccaatgttgc
5100tggaactgct gaaaaatcat tccatgtgga tgtctatgtt cctccaatga ttgaaggcaa
5160cttggccacg cctttgaata agcaagtagt tattgctcat tctctgacac tggagtgcaa
5220agctgctgga aacccttctc ccattctcac ctggttgaaa gatggtgtac ctgtgaaagc
5280taatgacaat atccgcatag aagctggtgg gaagaaactc gaaatcatga gtgcccaaga
5340aattgatcga ggacagtaca tatgcgtggc taccagtgtg gcaggagaaa aggaaatcaa
5400atatgaagtt gatgtcttgg tgccaccagc tatagaagga ggagatgaaa catcttactt
5460cattgtgatg gttaataact tactggagct agattgtcat gtgacaggct ctcccccacc
5520aactatcatg tggctgaagg atggccagtt aattgatgaa agggatggat tcaagatttt
5580attaaatgga cgcaaactgg ttattgctca ggctcaagtg tcaaacacag gcctttatcg
5640gtgcatggca gcaaatactg ctggagacca caagaaggaa tttgaagtga ctgttcatgt
5700tcctccaaca atcaagtcct caggcctttc tgagagagtt gtggtaaaat acaagcctgt
5760cgccttgcag tgcatagcca atgggattcc aaatccttcc attacatggt taaaagatga
5820ccagcctgtg aacactgccc aaggaaacct taaaatacag tcttctggtc gagttctaca
5880aattgccaaa accctgttgg aagatgctgg cagatacaca tgtgtggcta ccaacgcagc
5940tggagaaaca caacagcaca ttcaactgca tgttcatgaa ccacctagtc tggaagatgc
6000tggaaaaatg ctgaatgaga ctgtgttggt gagcaaccct gtacagctgg agtgtaaggc
6060agctggaaat cctgtgcctg ttattacatg gtacaaagat aatcgtctac tctcaggttc
6120caccagcatg actttcttga acagaggaca gatcattgat attgaaagtg cccagatctc
6180agatgctggc atatataaat gcgtggccat caactcagct ggagctacag agttatttta
6240cagtctgcaa gttcatgtgg ccccatcaat ttctggcagc aataacatgg tggcagtggt
6300ggttaataac ccggtgaggt tagaatgtga agccagaggt attcctgccc caagtctgac
6360ctggttgaaa gatgggagtc ctgtttctag tttttctaat ggattacagg ttctctctgg
6420tggtcgaatc ctagcattga ccagtgcaca aatcagcgac acaggaaggt acacctgcgt
6480ggcagtgaat gctgctggag aaaagcaaag ggacattgac ctccgagtat atgttccgcc
6540aaatattatg ggagaagaac agaatgtctc tgtcctcatt agccaagctg tggaattact
6600atgtcaaagt gatgctattc ccccacctac tcttacttgg ttaaaagacg gccacccctt
6660gctgaagaaa ccaggcctca gtatatctga aaatagaagt gtgttaaaga ttgaagatgc
6720tcaggttcaa gacactggtc gttacacttg tgaagcaaca aatgttgctg gaaaaactga
6780aaaaaaaaac tacaatgtca acatttgggt ccccccaaat attggtggtt ctgatgaact
6840tactcaactt acagtcattg aagggaatct cattagtctg ttgtgtgaat caagtggtat
6900tccaccccca aatctcatct ggaagaagaa aggctctcca gtgctgactg attccatggg
6960gcgagttaga attttatctg ggggcaggca attacaaatt tcaattgctg aaaagtctga
7020tgcagcactc tattcatgtg tggcgtcgaa tgttgctggg actgcaaaga aagaatacaa
7080tctgcaagtt tacattagac caaccataac caacagtggc agccacccta ctgaaattat
7140tgtgacccga gggaagagta tctccttgga gtgtgaggtg cagggtattc caccaccaac
7200agtgacctgg atgaaagatg gccacccctt gatcaaggca aagggagtag aaatactgga
7260tgaaggtcac atccttcagc tgaagaacat tcatgtatct gacacaggcc gttatgtgtg
7320tgttgctgtg aatgtagcag gaatgactga caaaaaatat gacttaagtg tccatgctcc
7380tccaagcatc ataggaaacc acaggtcacc tgaaaatatt agtgtggtag aaaagaactc
7440agtatctttg acttgtgaag cttctggaat tcccctgcct tccataacct ggttcaaaga
7500tgggtggcct gtcagcctta gcaattctgt gaggattctt tcaggaggca ggatgctacg
7560gctgatgcag accacaatgg aagatgctgg ccaatatact tgcgttgtaa ggaatgcagc
7620tggtgaagaa agaaaaatct ttgggctttc agtattagta ccacctcata ttgtgggtga
7680aaatacattg gaagatgtga aggtaaaaga gaaacagagt gttacgctga cttgtgaagt
7740gacagggaat ccagtgccag aaattacatg gcacaaagat gggcagcccc tccaagaaga
7800tgaagcccat cacattatat ctggtggccg ttttcttcaa attaccaatg tccaggtgcc
7860acacactgga agatatacat gtttggcttc cagtccagct ggccacaaga gcaggagctt
7920cagtcttaat gtatttgtat ctcctacaat tgctggtgta ggtagtgatg gcaaccctga
7980agatgtcact gtcatcctta acagccctac atctttggtc tgtgaagctt attcatatcc
8040tccagctacc atcacctggt ttaaggatgg cactccttta gaatctaacc gaaatattcg
8100tattcttcca ggaggcagaa ctctgcagat cctcaatgca caggaggaca atgctggaag
8160atactcttgt gtagccacga atgaggctgg agaaatgata aagcactatg aagtgaaggt
8220gtacattcca cccataatca ataaagggga cctttggggg ccaggtcttt cccctaaaga
8280agtgaagatc aaagtaaaca acactctgac cttggaatgt gaagcgtatg caattccttc
8340tgcctccctc agctggtaca aggatggaca gccccttaaa tccgatgatc atgttaatat
8400tgctgcgaat ggacacacac ttcaaataaa ggaggctcaa atatcagaca ccggacgata
8460tacttgtgta gcatctaaca ttgcaggtga agatgagttg gattttgatg tgaatattca
8520agttcctcca agttttcaga aactctggga aataggaaac atgctagata ctggcaggaa
8580tggtgaagcc aaagatgtga tcatcaacaa tcccatttct ctttactgtg agacaaatgc
8640tgctccccct cctacactga catggtacaa agatggccac cctctgacct caagtgataa
8700agtattgatt ttgccaggag ggcgagtgtt gcagattcct cgggctaaag tagaagatgc
8760tgggagatac acatgtgtgg ctgtgaatga ggctggagaa gattcccttc aatatgatgt
8820ccgtgtactc gtgccgccaa ttatcaaggg agcaaatagt gatctccctg aagaggtcac
8880cgtgctggtg aacaagagtg cactgataga gtgtttatcc agtggcagcc cagcaccaag
8940gaattcctgg cagaaagatg gacagccctt gctagaagat gaccatcata aatttctatc
9000taatggacga attctgcaga ttctgaatac tcaaataaca gatatcggca ggtatgtgtg
9060tgttgctgag aacacagctg ggagtgccaa aaaatatttt aacctcaatg ttcatgttcc
9120tccaagtgtc attggtccta aatctgaaaa tcttaccgtc gtggtgaaca atttcatctc
9180tttgacctgt gaggtctctg gttttccacc tcctgacctc agctggctca agaatgaaca
9240gcccatcaaa ctgaacacaa atactctcat tgtgcctggt ggtcgaactc tacagattat
9300tcgggccaag gtatcagatg gtggtgaata cacttgtata gctatcaatc aagctggcga
9360aagcaagaaa aagttttccc tgactgttta tgtgccccca agcattaaag accatgacag
9420tgaatctctt tctgtagtta atgtaagaga gggaacttct gtgtctttgg agtgtgagtc
9480gaacgctgtg ccacctccag tcatcacttg gtataagaat gggcggatga taacagagtc
9540tactcatgtg gagattttag ctgatggaca aatgctacac attaagaaag ctgaggtatc
9600tgacacaggc cagtatgtat gtagagctat aaatgtagca ggacgggatg ataaaaattt
9660ccacctcaat gtatatgtgc cacccagtat tgaaggacct gaaagagaag tgattgtgga
9720gacgatcagc aatcctgtga cattaacatg tgatgccact gggatcccac ctcccacgat
9780agcatggtta aagaaccaca agcgcataga aaattctgac tcactggaag ttcgtatttt
9840gtctggaggt agcaaactcc agattgcccg gtctcagcat tcagatagtg gaaactatac
9900atgtattgct tcaaatatgg agggaaaagc ccagaaatat tactttcttt caattcaagt
9960tcctccaagt gttgctggtg ctgaaattcc aagtgatgtc agtgtccttc taggagaaaa
10020tgttgagctg gtctgcaatg caaatggcat tcctactcca cttattcaat ggcttaaaga
10080tggaaagccc atagctagtg gtgaaacaga aagaatccga gtgagtgcaa atggcagcac
10140attaaacatt tatggagctc ttacatctga cacggggaaa tacacatgtg ttgctactaa
10200tcccgctgga gaagaagacc gaatttttaa cttgaatgtc tatgttacac ctacaattag
10260gggtaataaa gatgaagcag agaaactaat gactttagtg gatacttcaa taaatattga
10320atgcagagcc acagggacgc ctccaccaca gataaactgg ctgaagaatg gacttcctct
10380gcctctctcc tcccatatcc ggttactggc agcaggacaa gttatcagga ttgtgagagc
10440tcaggtgtct gatgtcgctg tgtatacttg tgtggcctcc aacagagctg gggtggataa
10500taagcattac aatcttcaag tgtttgcacc accaaatatg gacaattcaa tggggacaga
10560ggaaatcaca gttctcaaag gtagttccac ctctatggca tgcattactg atggaacccc
10620agctcccagt atggcctggc ttagagatgg ccagcctctg gggcttgatg cccatctgac
10680agtcagcacc catggaatgg tcctgcagct cctcaaagca gagactgaag attcgggaaa
10740gtacacctgc attgcctcaa atgaagctgg agaagtcagc aagcacttta tcctcaaggt
10800cctagaacca cctcacatta atggatctga agaacatgaa gagatatcag taattgttaa
10860taacccactt gaacttacct gcattgcttc tggaatccca gcccctaaaa tgacctggat
10920gaaagatggc cggccccttc cacagacgga tcaagtgcaa actctaggag gaggagaggt
10980tcttcgaatt tctactgctc aggtggagga tacaggaaga tatacatgtc tggcatccag
11040tcctgcagga gatgatgata aggaatatct agtgagagtg catgtacctc ctaatattgc
11100tggaactgat gagccccggg atatcactgt gttacggaac agacaagtga cattggaatg
11160caagtcagat gcagtgcccc cacctgtaat tacttggctc agaaatggag aacggttaca
11220ggcaacacct cgagtgcgaa tcctatctgg agggagatac ttgcaaatca acaatgctga
11280cctaggtgat acagccaatt atacctgtgt tgccagcaac attgcaggaa agactacaag
11340agaatttatt ctcactgtaa atgttcctcc aaacataaag gggggccccc agagccttgt
11400aattctttta aataagtcaa ctgtattgga atgcatcgct gaaggtgtgc caactccaag
11460gataacatgg agaaaggatg gagctgttct agctgggaat catgcaagat attccatctt
11520ggaaaatgga ttccttcata ttcaatcagc acatgtcact gacactggac ggtatttgtg
11580tatggccacc aatgctgctg gaacagatcg caggcgaata gatttacagg tccatgttcc
11640tccatctatt gctccgggtc ctaccaacat gactgtaata gtaaatgttc aaactactct
11700ggcttgtgag gctactggga taccaaaacc atcaatcaat tggagaaaaa atgggcatct
11760tcttaatgtg gatcaaaatc agaactcata caggctcctt tcttcaggtt cactagtaat
11820tatttcccct tctgtggatg acactgcaac ctatgaatgt actgtgacaa acggtgctgg
11880agatgataaa agaactgtgg atctcactgt ccaagttcca ccttccatag ctgatgagcc
11940tacagatttc ctagtaacca aacatgcccc agcagtaatt acctgcactg cttcgggagt
12000tccatttccc tcaattcact ggaccaaaaa tggtataaga ctgcttccca ggggagatgg
12060ctatagaatt ctgtcctcag gagcaattga aatacttgcc acccaattaa accatgctgg
12120aagatacact tgtgtcgcta ggaatgcggc tggctctgca catcgacacg tgacccttca
12180tgttcatgag cctccagtca ttcagcccca accaagtgaa ctacacgtca ttctgaacaa
12240tcctatttta ttaccatgtg aagcaacagg gacacccagt cctttcatta cttggcaaaa
12300agaaggcatc aatgttaaca cttcaggcag aaaccatgca gttcttccta gtggcggctt
12360acagatctcc agagctgtcc gagaggatgc tggcacttac atgtgtgtgg cccagaaccc
12420ggctggtaca gccttgggca aaatcaagtt aaatgtccaa gttcctccag tcattagccc
12480tcatctaaag gaatatgtta ttgctgtgga caagcccatc acgttatcct gtgaagcaga
12540tggcctccct ccgcctgaca ttacatggca taaagatggg cgtgcaattg tggaatctat
12600ccgccagcgc gtcctcagct ctggctctct gcaaatagca tttgtccagc ctggtgatgc
12660tggccattac acgtgcatgg cagccaatgt agcaggatca agcagcacaa gcaccaagct
12720caccgtccat gtaccaccca ggatcagaag tacagaagga cactacacgg tcaatgagaa
12780ttcacaagcc attcttccat gcgtagctga tggaatcccc acaccagcaa ttaactggaa
12840aaaagacaat gttcttttag ctaacttgtt aggaaaatac actgctgaac catatggaga
12900actcatttta gaaaatgttg tgctggagga ttctggcttc tatacctgtg ttgctaacaa
12960tgctgcaggt gaagatacac acactgtcag cctgactgtg catgttctcc ccacttttac
13020tgaacttcct ggagacgtgt cattaaataa aggagaacag ctacgattaa gctgtaaagc
13080tactggtatt ccattgccca aattaacatg gaccttcaat aacaatatta ttccagccca
13140ctttgacagt gtgaatggac acagtgaact tgttattgaa agagtgtcaa aagaggattc
13200aggtacttat gtgtgcaccg cagagaacag cgttggcttt gtgaaggcaa ttggatttgt
13260ttatgtgaaa gaacctccag tcttcaaagg tgattatcct tctaactgga ttgaaccact
13320tggtgggaat gcaatcctga attgtgaggt gaaaggagac cccaccccaa ccatccagtg
13380gaacagaaag ggagtggata ttgaaattag ccacagaatc cggcaactgg gcaatggctc
13440cctggccatc tatggcactg ttaatgaaga tgccggtgac tatacatgtg tagctaccaa
13500tgaagctggg gtggtggagc gcagcatgag tctgactctg caaagtcctc ctattatcac
13560tcttgagcca gtggaaactg ttattaatgc tggtggcaaa atcatattga attgtcaggc
13620aactggagag cctcaaccaa ccattacatg gtcccgtcaa gggcactcta tttcctggga
13680tgaccgggtt aacgtgttgt ccaacaactc attatatatt gctgatgctc agaaagaaga
13740tacctctgaa tttgaatgtg ttgctcgaaa cttaatgggt tctgtccttg tcagagtgcc
13800agtcatagtc caggttcatg gtggattttc ccagtggtct gcatggagag cctgcagtgt
13860cacctgtgga aaaggcatcc aaaagaggag tcgtctgtgc aaccagcccc ttccagccaa
13920tggtgggaag ccctgccaag gttcagattt ggaaatgcga aactgtcaaa ataagccttg
13980tccagtggat ggtagctggt cggaatggag tctttgggaa gaatgcacaa ggagctgtgg
14040acgcggcaac caaaccagga ccaggacttg caataatcca tcagttcagc atggtgggcg
14100gccatgtgaa gggaatgctg tggaaataat tatgtgcaac attaggcctt gcccagttca
14160tggagcatgg agcgcttggc agccttgggg aacatgcagc gaaagttgtg ggaaaggtac
14220tcagacaaga gcaagacttt gtaataaccc accaccagcg tttggtgggt cctactgtga
14280tggagcagaa acacagatgc aagtttgcaa tgaaagaaat tgtccaattc atggcaagtg
14340ggcgacttgg gccagttgga gtgcctgttc tgtgtcatgt ggaggaggtg ccagacagag
14400aacaaggggc tgctccgacc ctgtgcccca gtatggagga aggaaatgcg aagggagtga
14460tgtccagagt gatttttgca acagtgaccc ttgcccaacc catggtaact ggagtccttg
14520gagtggctgg ggaacatgca gccggacgtg taacggaggg cagatgcggc ggtaccgcac
14580atgtgataac cctcctccct ccaatggggg aagagcttgt gggggaccag actcccagat
14640ccagaggtgc aacactgaca tgtgtcctgt ggatggaagt tggggaagct ggcatagttg
14700gagccagtgc tctgcctcct gtggaggagg tgaaaagact cggaagcggc tgtgcgacca
14760tcctgtgcca gttaaaggtg gccgtccctg tcccggagac actactcagg tgaccaggtg
14820caatgtacaa gcatgtccag gtgggcccca gcgagccaga ggaagtgtta ttggaaatat
14880taatgatgtt gaatttggaa ttgctttcct taatgccaca ataactgata gccctaactc
14940tgatactaga ataatacgtg ccaaaattac caatgtacct cgtagtcttg gttcagcaat
15000gagaaagata gtttctattc taaatcccat ttattggaca acagcaaagg aaataggaga
15060agcagtcaat ggctttaccc tcaccaatgc agtcttcaaa agagaaactc aagtggaatt
15120tgcaactgga gaaatcttgc agatgagtca tattgcccgg ggcttggatt ccgatggttc
15180tttgctgcta gatatcgttg tgagtggcta tgtcctacag cttcagtcac ctgctgaagt
15240cactgtaaag gattacacag aggactacat tcaaacaggt cctgggcagc tgtacgccta
15300ctcaacccgg ctgttcacca ttgatggcat cagcatccca tacacatgga accacaccgt
15360tttctatgat caggcacagg gaagaatgcc tttcttggtt gaaacacttc atgcatcctc
15420tgtggaatct gactataacc agatagaaga gacactgggt tttaaaattc atgcttcaat
15480atccaaagga gatcgcagta atcagtgccc ctccgggttt accttagact cagttggacc
15540tttttgtgct gatgaggatg aatgtgcagc agggaatccc tgctcccata gctgccacaa
15600tgccatgggg acttactact gctcctgccc taaaggcctc accatagctg cagatggaag
15660aacttgtcaa gatattgatg agtgtgcttt gggtaggcat acctgccacg ctggtcagga
15720ctgtgacaat acgattggat cttatcgctg tgtggtccgt tgtggaagtg gctttcgaag
15780aacctctgat gggctgagtt gtcaagatat taatgaatgt caagaatcca gcccctgtca
15840ccagcgctgt ttcaatgcca taggaagttt ccattgtgga tgtgaacctg ggtatcagct
15900caaaggcaga aaatgcatgg atgtgaacga gtgtagacaa aatgtatgca gaccagatca
15960gcactgtaag aacacccgtg gtggctataa gtgcattgat ctttgtccaa atggaatgac
16020caaggcagaa aatggaacct gtattgatat tgatgaatgt aaagatggga cccatcagtg
16080cagatataac cagatatgtg agaatacaag aggcagctat cgttgtgtat gcccaagagg
16140ttatcggtct caaggagttg gaagaccctg catggacatt aatgaatgtg aacaagtgcc
16200taaaccttgt gcacatcagt gctccaacac ccccggcagc ttcaagtgta tctgtccacc
16260aggacaacat ttattagggg acgggaaatc ttgcgctgga ttggagaggc tgccaaatta
16320tggcactcaa tacagtagct ataaccttgc acggttctcc cctgtgagaa acaactatca
16380acctcaacag cattacagac agtactcaca tctctacagc tcctactcag agtatagaaa
16440cagcagaaca tctctctcca ggactagaag gactattagg aaaacttgcc ctgaaggctc
16500tgaggcaagc catgacacat gtgtagatat tgatgaatgt gaaaatacag atgcctgcca
16560gcatgagtgt aagaatacct ttggaagtta tcagtgcatc tgcccacctg gctatcaact
16620cacacacaat ggaaagacat gccaagatat cgatgaatgt ctggagcaga atgtgcactg
16680tggacccaat cgcatgtgct tcaacatgag aggaagctac cagtgcatcg atacaccctg
16740tccacccaac taccaacggg atcctgtttc agggttctgc ctcaagaact gtccacccaa
16800tgatttggaa tgtgccttga gcccatatgc cttggaatac aaactcgtct ccctcccatt
16860tggaatagcc accaatcaag atttaatccg gctggttgca tacacacagg atggagtgat
16920gcatcccagg acaactttcc tcatggtaga tgaggaacag actgttcctt ttgccttgag
16980ggatgaaaac ctgaaaggag tggtgtatac aacacgacca ctacgagaag cagagaccta
17040ccgcatgagg gtccgagcct catcctacag tgccaatggg accattgaat atcagaccac
17100attcatagtt tatatagctg tgtccgccta tccatactaa ggaactctcc aaagcctatt
17160ccacatattt aaaccgcatt aatcatggca atcaagcccc cttccagatt actgtctctt
17220gaacagttgc aatcttggca gcttgaaaat ggtgctacac tctgttttgt gtgccttcct
17280tggtacttct gaggtatttt catgatccca ccatggtcat atcttgaagt atggtctaga
17340aaagtccctt attattttat ttattacact ggagcagtta cttcccaaag attattctga
17400acatctaaca ggacatatca gtgatggttt acagtagtgt agtacctaag atcattttcc
17460tgaaagccaa accaaacaac gaaaaacaag aacaactaat tcagaatcaa atagagtttt
17520tgagcatttg actattttta gaatcataaa attagttact aagtattttg atcaaagctt
17580ataaaataac ttacggagat ttttgtaagt attgatacat tataatagga cttgcctatt
17640ttcattttta agaagaaaaa caccactcat tttataaaat atagtacagc tactataagg
17700cttgtttgat cccaaatggt gcttatcttg attgaacatt cagaacaagg atattatttt
17760cagtgatttt gtgagatcag ctgaaccact tatgataata ataataaaaa agactgcttt
17820gccctcacgt cagttgtaca tggcatggaa ctttaaaaat tttaatataa actttcatcc
17880agttagcttc ataactttta cgttccagaa ttttgtttat tttcctgtca atgaaagcaa
17940tttttaaaga taccagtggg acaggtttgg ttttttaaaa atctcatgtg ttcaaattaa
18000cataaatatt acacgtcaat acactgtaca tggtggtaat agactctaag caattgccaa
18060gatgtattct atttttatga agtgtatata tattacctta gtgtgcattt tctatataat
18120atcttgatgg actcttttat aaaattattt tataaaaaac aatgttacac taaaatcagc
18180ctaaataaat tttcacaact ttttttcat
18209671026DNAHomo sapiens 67cagcatgttg agccgggcag tgtgcggcac cagcaggcag
ctgcctccgg ttttggggta 60tctgggctcc aggcagaagc acagcctccc cgacctgccc
tacgactacg gcgccctgga 120acctcacatc aacgcgcaga tcatgcagct gcaccacagc
aagcaccacg cggcctacgt 180gaacaacctg aacgtcaccg aggagaagta ccaggaggcg
ttggccaagg gagatgttac 240agcccagata gctcttcagc ctgcactgaa gttcaatggt
ggtggtcata tcaatcatag 300cattttctgg acaaacctca gccctaacgg tggtggagaa
cccaaagggg agttgctgga 360agccatcaaa ctggactttg gttcctttga caagtttaag
gagaagctga cggctgcatc 420tgttggtgtc caaggctcag gttggggttg gcttggtttc
aataaggaac ggggacactt 480acaaattgct gcttgtccaa atcaggatcc actgcaagga
acaacaggcc ttattccact 540gctggggatt gatgtgtggg agcacgctta ctaccttcag
tataaaaatg tcaggcctga 600ttatctaaaa gctatttgga atgtaatcaa ctgggagaat
gtaactgaaa gatacatggc 660ttgcaaaaag taaaccacga tcgttatgct gagtatgtta
agctctttat gactgttttt 720gtagtggtat agagtactgc agaatacagt aagctgctct
attgtagcat ttcttgatgt 780tgcttagtca cttatttcat aaacaactta atgttctgaa
taatttctta ctaaacattt 840tgttattggg caagtgattg aaaatagtaa atgctttgtg
tgattgaatc tgattggaca 900ttttcttcag agagctaaat tacaattgtc atttataaaa
ccatcaaaaa tattccatcc 960atatactttg gggacttgta gggatgcctt tctagtccta
ttctattgca gttatagaaa 1020atctag
102668757DNAHomo sapiens 68ggaaccgaga ggctgagact
aacccagaaa catccaattc tcaaactgaa gctcgcactc 60tcgcctccag catgaaagtc
tctgccgccc ttctgtgcct gctgctcata gcagccacct 120tcattcccca agggctcgct
cagccagatg caatcaatgc cccagtcacc tgctgttata 180acttcaccaa taggaagatc
tcagtgcaga ggctcgcgag ctatagaaga atcaccagca 240gcaagtgtcc caaagaagct
gtgatcttca agaccattgt ggccaaggag atctgtgctg 300accccaagca gaagtgggtt
caggattcca tggaccacct ggacaagcaa acccaaactc 360cgaagacttg aacactcact
ccacaaccca agaatctgca gctaacttat tttcccctag 420ctttccccag acaccctgtt
ttattttatt ataatgaatt ttgtttgttg atgtgaaaca 480ttatgcctta agtaatgtta
attcttattt aagttattga tgttttaagt ttatctttca 540tggtactagt gttttttaga
tacagagact tggggaaatt gcttttcctc ttgaaccaca 600gttctacccc tgggatgttt
tgagggtctt tgcaagaatc attaatacaa agaatttttt 660ttaacattcc aatgcattgc
taaaatatta ttgtggaaat gaatattttg taactattac 720accaaataaa tatatttttg
tacaaaaaaa aaaaaaa 757692395DNAHomo sapiens
69agagcctcct agcccgtcgg tgtctgcgcc catcgatccc tttgtctatc cccgaccatg
60gcgaagctga ttgcgctcac cctcttgggg atgggactgg cactcttcag gaaccaccag
120tcttcttacc aaacacgact taatgctctc cgagaggtac aacccgtaga acttcctaac
180tgtaatttag ttaaaggaat cgaaactggc tctgaagact tggagatact gcctaatgga
240ctggctttca ttagctctgg attaaagtat cctggaataa agagcttcaa ccccaacagt
300cctggaaaaa tacttctgat ggacctgaat gaagaagatc caacagtgtt ggaattgggg
360atcactggaa gtaaatttga tgtatcttca tttaaccctc atgggattag cacattcaca
420gatgaagata atgccatgta cctcctggtg gtgaaccatc cagatgccaa gtccacagtg
480gagttgttta aatttcaaga agaagaaaaa tcgcttttgc atctaaaaac catcagacat
540aaacttctgc ctaatttgaa tgatattgtt gctgtgggac ctgagcactt ttatggcaca
600aatgatcact attttcttga cccctactta caatcctggg agatgtattt gggtttagcg
660tggtcgtatg ttgtctacta tagtccaagt gaagttcgag tggtggcaga aggatttgat
720tttgctaatg gaatcaacat ttcacccgat ggcaagtatg tctatatagc tgagttgctg
780gctcataaga ttcatgtgta tgaaaagcat gctaattgga ctttaactcc attgaagtcc
840cttgacttta ataccctcgt ggataacata tctgtggatc ctgagacagg agacctttgg
900gttggatgcc atcccaatgg catgaaaatc ttcttctatg actcagagaa tcctcctgca
960tcagaggtgc ttcgaatcca gaacattcta acagaagaac ctaaagtgac acaggtttat
1020gcagaaaatg gcacagtgtt gcaaggcagt acagttgcct ctgtgtacaa agggaaactg
1080ctgattggca cagtgtttca caaagctctt tactgtgagc tctaacagac cgatttgcac
1140ccatgccata gaaactgagg ccattatttc aaccgcttgc catattccga ggacccagtg
1200ttcttagctg aacaatgaat gctgacccta aatgtggaca tcatgaagca tcaaagcact
1260gtttaactgg gagtgatatg atgtgtaggg cttttttttg agaatacact atcaaatcag
1320tcttggaata cttgaaaacc tcatttacca taaaaatcct tctcactaaa atggataaat
1380cagttatgtc aattgtcaga tattaaataa cagtgtgtga ccccaaaagt acttacccta
1440aaacatgtgt tgcctgaaag cacatgtgtg tatcgctgcc ttgccatgtc ttgttcagaa
1500gacacagggg agcagggtta gctcacgtgt ctttagaact ccagtactca cccagggact
1560ccagttcaca ggccagaaaa catatgcatt atgaagttcc cctctactcc atgcacatag
1620taagtctgac tatggcagtc agacttactt actcccattt tcccttcgat atatgacttt
1680ttctcagtaa atattaacct gaactattcc aactcccctt gtactcttgc tttttcaatt
1740ctcctgttgc aatgacacat aggaaaatct taaaattctt gggagtgttg tcacacctga
1800aaattatgag tctctatgat cttggcacaa attgtacatt tgagtgtctt tgacttggtt
1860aaaggaagtt tgttcacttc gatgactgga tacagaatga atcccataat tgacatgggc
1920gacagtaaaa gtgtccccaa agactacact gttgttgagg tggtggtagt gctggtgggt
1980ttttgtttaa tatttaaact tcttgttgtg gaggctgaaa agaaaaaaaa taatagaaag
2040gtaaacaaac aaataaatag aaaagatcaa caaccccttt ggctatctac tgagacatga
2100ctaggaagaa aacatgactt tatcattttg ttatagaagc tgatatataa ggttacacat
2160tttcatttat ttgtttttct gatttgaagg tataaccttc atgatgaatt acttcttcag
2220ggtgttaagg cagtgacttt agaaacaaat ttttttcttg cttttgtttt gtttttgaga
2280ccgaatctca ctctgttgcc caggctggag tgcagtggtg cgatcttggc tcactgcaac
2340ttctacctcc gaggttcaag agattcttgt gcctcagcct cccggatagc tgccg
23957074PRTHomo sapiens 70Met Ala Phe Leu Val His Ser Gln Pro Val Ile Leu
Gly Phe Thr Val1 5 10
15Leu Leu Ser Tyr Ile Leu Arg Tyr Gln Leu Leu Phe Phe Lys Phe Val
20 25 30Phe Ile Leu Phe Asp Lys Lys
Pro Ala Leu Ala Thr His Thr His Asn 35 40
45Lys Ser His Phe Lys Ile Val Ala Gln Thr Pro Arg Lys Lys Arg
Lys 50 55 60Glu Lys Leu Glu Gln Gln
Gln Gln Lys Asn65 707155PRTHomo sapiens 71Met Thr Leu
Leu Val Phe Thr Ser His Val Gln Cys Pro Asn Arg Gln1 5
10 15Cys Lys Lys Tyr Pro Val Trp Phe Asn
Arg Lys Ser Val Tyr Val Ser 20 25
30Leu Phe Glu Thr Ser Phe Thr Leu Ser Gly Ser Leu Ser Ser Met Lys
35 40 45Ser Ala Arg Asn Ile Gly Trp
50 557252PRTHomo sapiens 72Met Glu Thr Asn Phe Val
Glu Leu Leu Pro Phe Asp Leu Gly Leu Glu1 5
10 15Tyr Glu Leu Leu Tyr Asn Ser Tyr Ser Tyr Leu Ala
Asn Ala Gln Phe 20 25 30Ser
Ile Thr Ser Leu Met Ala Phe Thr Arg Lys Ala Val Leu Glu Ala 35
40 45Ile Val Ile His 507345PRTHomo
sapiens 73Met Tyr Phe Ala Met Lys Leu Pro Leu Gly Leu Ile Ile Ser Ile
Pro1 5 10 15Leu Leu Arg
Asn Val Gln Met Ile Leu Tyr Ser Thr Thr Leu Val Pro 20
25 30Leu Cys Met Thr Val Arg Phe Phe Phe Phe
Leu Leu Phe 35 40 457443PRTHomo
sapiens 74Met Asp Arg Glu Asn Gln Ile Ser Ser Tyr Asn Cys Leu Ala Asn
Gly1 5 10 15Ile Ser Gly
Ser Phe Ser Ala Ser His Phe Arg Leu His Ser Leu Thr 20
25 30Leu Leu His Phe Lys Ile Pro Ala Phe Ile
Phe 35 407537PRTHomo sapiens 75Met Cys Cys Phe
Gly Tyr Thr His Ser Phe Phe Phe Asn Arg Ile Tyr1 5
10 15Cys Leu Val Ser Leu Trp Thr Gly Thr Val
Asp Ala His Leu Lys Val 20 25
30Lys Cys His Phe Phe 357635PRTHomo sapiens 76Met Phe Ser Val Gln
Thr Gly Asn Val Lys Ser Ile Leu Cys Gly Leu1 5
10 15Thr Gly Asn Leu Phe Met Ser Leu Tyr Leu Lys
Pro Val Leu Leu Ser 20 25
30Val Val Leu 357734PRTHomo sapiens 77Met Ile Tyr Phe Leu Lys Ser
Asn Phe Asn Ser Ser Cys Leu Thr Glu1 5 10
15Ala Cys Gln Tyr Met Cys Cys Ile Phe Phe Ala Phe Val
Glu Lys Leu 20 25 30His
Ile7834PRTHomo sapiens 78Met Pro Arg Ala Ile Val Phe Pro Pro Phe Phe Ala
Ser Phe Ser Tyr1 5 10
15Pro Leu Phe Gln Leu Gln Met Pro Lys Lys Met Pro Thr Asp Thr Thr
20 25 30Leu Pro79190PRTHomo sapiens
79Met Ala Thr His His Thr Leu Trp Met Gly Leu Ala Leu Leu Gly Val1
5 10 15Leu Gly Asp Leu Gln Ala
Ala Pro Glu Ala Gln Val Ser Val Gln Pro 20 25
30Asn Phe Gln Gln Asp Lys Phe Leu Gly Arg Trp Phe Ser
Ala Gly Leu 35 40 45Ala Ser Asn
Ser Ser Trp Leu Arg Glu Lys Lys Ala Ala Leu Ser Met 50
55 60Cys Lys Ser Val Val Ala Pro Ala Thr Asp Gly Gly
Leu Asn Leu Thr65 70 75
80Ser Thr Phe Leu Arg Lys Asn Gln Cys Glu Thr Arg Thr Met Leu Leu85
90 95Gln Pro Ala Gly Ser Leu Gly Ser Tyr Ser
Tyr Arg Ser Pro His Trp100 105 110Gly Ser
Thr Tyr Ser Val Ser Val Val Glu Thr Asp Tyr Asp Gln Tyr115
120 125Ala Leu Leu Tyr Ser Gln Gly Ser Lys Gly Pro Gly
Glu Asp Phe Arg130 135 140Met Ala Thr Leu
Tyr Ser Arg Thr Gln Thr Pro Arg Ala Glu Leu Lys145 150
155 160Glu Lys Phe Thr Ala Phe Cys Lys Ala
Gln Gly Phe Thr Glu Asp Thr165 170 175Ile
Val Phe Leu Pro Gln Thr Asp Lys Cys Met Thr Glu Gln180
185 19080170PRTHomo sapiens 80Met Ala Ser Gln Lys Arg Pro
Ser Gln Arg His Gly Ser Lys Tyr Leu1 5 10
15Ala Thr Ala Ser Thr Met Asp His Ala Arg His Gly Phe
Leu Pro Arg 20 25 30His Arg
Asp Thr Gly Ile Leu Asp Ser Ile Gly Arg Phe Gly Gly Asp 35
40 45Arg Gly Ala Pro Lys Arg Gly Ser Gly Lys
Asp Ser His His Pro Ala 50 55 60Arg
Thr Ala His Tyr Gly Ser Leu Pro Gln Lys Ser His Gly Arg Thr65
70 75 80Gln Asp Glu Asn Pro Val
Val His Phe Phe Lys Asn Ile Val Thr Pro85 90
95Arg Thr Pro Pro Pro Ser Gln Gly Lys Gly Arg Gly Leu Ser Leu Ser100
105 110Arg Phe Ser Trp Gly Ala Glu Gly
Gln Arg Pro Gly Phe Gly Tyr Gly115 120
125Gly Arg Ala Ser Asp Tyr Lys Ser Ala His Lys Gly Phe Lys Gly Val130
135 140Asp Ala Gln Gly Thr Leu Ser Lys Ile
Phe Lys Leu Gly Gly Arg Asp145 150 155
160Ser Arg Ser Gly Ser Pro Met Ala Arg Arg165
1708138PRTHomo sapiens 81Ile Arg Ser Ala Lys Leu Gly Phe Cys Cys Leu Asn
Ser Ala Leu Gly1 5 10
15Pro Gln Ile Asn Arg Cys Glu Cys Ser Phe Phe Pro Leu Cys Glu Glu
20 25 30Ala Val Thr Pro Gln Gln
358238PRTHomo sapiens 82Leu Leu Gly Cys Asn Cys Phe Phe Thr Gln Gly Glu
Lys Thr Thr Phe1 5 10
15Thr Ser Val Tyr Leu Arg Thr Gln Cys Arg Val Gln Ala Ala Lys Pro
20 25 30Gln Leu Ser Arg Ser Asn
358337PRTHomo sapiens 83Phe Ile Tyr Lys Lys Ile Lys Leu Glu Ile Val Leu
Asp Phe Ser Ser1 5 10
15Tyr Cys Trp Gly Val Thr Ala Ser Ser His Arg Gly Lys Lys Leu His
20 25 30Ser His Arg Phe Ile
3584335PRTHomo sapiens 84Met Gly Ala Ser Ser Ser Ser Ala Leu Ala Arg Leu
Gly Leu Pro Ala1 5 10
15Arg Pro Trp Pro Arg Trp Leu Gly Val Ala Ala Leu Gly Leu Ala Ala
20 25 30Val Ala Leu Gly Thr Val Ala
Trp Arg Arg Ala Trp Pro Arg Arg Arg 35 40
45Arg Arg Leu Gln Gln Val Gly Thr Val Ala Lys Leu Trp Ile Tyr
Pro 50 55 60Val Lys Ser Cys Lys Gly
Val Pro Val Ser Glu Ala Glu Cys Thr Ala65 70
75 80Met Gly Leu Arg Ser Gly Asn Leu Arg Asp Arg
Phe Trp Leu Val Ile85 90 95Lys Glu Asp
Gly His Met Val Thr Ala Arg Gln Glu Pro Arg Leu Val100
105 110Leu Ile Ser Ile Ile Tyr Glu Asn Asn Cys Leu Ile
Phe Arg Ala Pro115 120 125Asp Met Asp Gln
Leu Val Leu Pro Ser Lys Gln Pro Ser Ser Asn Lys130 135
140Leu His Asn Cys Arg Ile Phe Gly Leu Asp Ile Lys Gly Arg
Asp Cys145 150 155 160Gly
Asn Glu Ala Ala Lys Trp Phe Thr Asn Phe Leu Lys Thr Glu Ala165
170 175Tyr Arg Leu Val Gln Phe Glu Thr Asn Met Lys
Gly Arg Thr Ser Arg180 185 190Lys Leu Leu
Pro Thr Leu Asp Gln Asn Phe Gln Val Ala Tyr Pro Asp195
200 205Tyr Cys Pro Leu Leu Ile Met Thr Asp Ala Ser Leu
Val Asp Leu Asn210 215 220Thr Arg Met Glu
Lys Lys Met Lys Met Glu Asn Phe Arg Pro Asn Ile225 230
235 240Val Val Thr Gly Cys Asp Ala Phe Glu
Glu Asp Thr Trp Asp Glu Leu245 250 255Leu
Ile Gly Ser Val Glu Val Lys Lys Val Met Ala Cys Pro Arg Cys260
265 270Ile Leu Thr Thr Val Asp Pro Asp Thr Gly Val
Ile Asp Arg Lys Gln275 280 285Pro Leu Asp
Thr Leu Lys Ser Tyr Arg Leu Cys Asp Pro Ser Glu Arg290
295 300Glu Leu Tyr Lys Leu Ser Pro Leu Phe Gly Ile Tyr
Tyr Ser Val Glu305 310 315
320Lys Ile Gly Ser Leu Arg Val Gly Asp Pro Val Tyr Arg Met Val325
330 33585459PRTHomo sapiens 85Met Ile Lys Arg
Phe Leu Glu Asp Thr Thr Asp Asp Gly Glu Leu Ser1 5
10 15Lys Phe Val Lys Asp Phe Ser Gly Asn Ala
Ser Cys His Pro Pro Glu 20 25
30Ala Lys Thr Trp Ala Ser Arg Pro Gln Val Pro Glu Pro Arg Pro Gln
35 40 45Ala Pro Asp Leu Tyr Asp Asp Asp
Leu Glu Phe Arg Pro Pro Ser Arg 50 55
60Pro Gln Ser Ser Asp Asn Gln Gln Tyr Phe Cys Ala Pro Ala Pro Leu65
70 75 80Ser Pro Ser Ala Arg
Pro Arg Ser Pro Trp Gly Lys Leu Asp Pro Tyr85 90
95Asp Ser Ser Glu Asp Asp Lys Glu Tyr Val Gly Phe Ala Thr Leu
Pro100 105 110Asn Gln Val His Arg Lys Ser
Val Lys Lys Gly Phe Asp Phe Thr Leu115 120
125Met Val Ala Gly Glu Ser Gly Leu Gly Lys Ser Thr Leu Val Asn Ser130
135 140Leu Phe Leu Thr Asp Leu Tyr Arg Asp
Arg Lys Leu Leu Gly Ala Glu145 150 155
160Glu Arg Ile Met Gln Thr Val Glu Ile Thr Lys His Ala Val
Asp Ile165 170 175Glu Glu Lys Gly Val Arg
Leu Arg Leu Thr Ile Val Asp Thr Pro Gly180 185
190Phe Gly Asp Ala Val Asn Asn Thr Glu Cys Trp Lys Pro Val Ala
Glu195 200 205Tyr Ile Asp Gln Gln Phe Glu
Gln Tyr Phe Arg Asp Glu Ser Gly Leu210 215
220Asn Arg Lys Asn Ile Gln Asp Asn Arg Val His Cys Cys Leu Tyr Phe225
230 235 240Ile Ser Pro Phe
Gly His Gly Leu Arg Pro Leu Asp Val Glu Phe Met245 250
255Lys Ala Leu His Gln Arg Val Asn Ile Val Pro Ile Leu Ala
Lys Ala260 265 270Asp Thr Leu Thr Pro Pro
Glu Val Asp His Lys Lys Arg Lys Ile Arg275 280
285Glu Glu Ile Glu His Phe Gly Ile Lys Ile Tyr Gln Phe Pro Asp
Cys290 295 300Asp Ser Asp Glu Asp Glu Asp
Phe Lys Leu Gln Asp Gln Ala Leu Lys305 310
315 320Glu Ser Ile Pro Phe Ala Val Ile Gly Ser Asn Thr
Val Val Glu Ala325 330 335Arg Gly Arg Arg
Val Arg Gly Arg Leu Tyr Pro Trp Gly Ile Val Glu340 345
350Val Glu Asn Pro Gly His Cys Asp Phe Val Lys Leu Arg Thr
Met Leu355 360 365Val Arg Thr His Met Gln
Asp Leu Lys Asp Val Thr Arg Glu Thr His370 375
380Tyr Glu Asn Tyr Arg Ala Gln Cys Ile Gln Ser Met Thr Arg Leu
Val385 390 395 400Val Lys
Glu Arg Asn Arg Asn Lys Leu Thr Arg Glu Ser Gly Thr Asp405
410 415Phe Pro Ile Pro Ala Val Pro Pro Gly Thr Asp Pro
Glu Thr Glu Lys420 425 430Leu Ile Arg Glu
Lys Asp Glu Glu Leu Arg Arg Met Gln Glu Met Leu435 440
445His Lys Ile Gln Lys Gln Met Lys Glu Asn Tyr450
45586142PRTHomo sapiens 86Met Ala Thr Lys Ile Asp Lys Glu Ala Cys
Arg Ala Ala Tyr Asn Leu1 5 10
15Val Arg Asp Asp Gly Ser Ala Val Ile Trp Val Thr Phe Lys Tyr Asp
20 25 30Gly Ser Thr Ile Val Pro
Gly Glu Gln Gly Ala Glu Tyr Gln His Phe 35 40
45Ile Gln Gln Cys Thr Asp Asp Val Arg Leu Phe Ala Phe Val
Arg Phe 50 55 60Thr Thr Gly Asp Ala
Met Ser Lys Arg Ser Lys Phe Ala Leu Ile Thr65 70
75 80Trp Ile Gly Glu Asn Val Ser Gly Leu Gln
Arg Ala Lys Thr Gly Thr85 90 95Asp Lys
Thr Leu Val Lys Glu Val Val Gln Asn Phe Ala Lys Glu Phe100
105 110Val Ile Ser Asp Arg Lys Glu Leu Glu Glu Asp Phe
Ile Lys Ser Glu115 120 125Leu Lys Lys Ala
Gly Gly Ala Asn Tyr Asp Ala Gln Thr Glu130 135
14087449PRTHomo sapiens 87Met Met Lys Thr Leu Leu Leu Phe Val Gly
Leu Leu Leu Thr Trp Glu1 5 10
15Ser Gly Gln Val Leu Gly Asp Gln Thr Val Ser Asp Asn Glu Leu Gln
20 25 30Glu Met Ser Asn Gln Gly
Ser Lys Tyr Val Asn Lys Glu Ile Gln Asn 35 40
45Ala Val Asn Gly Val Lys Gln Ile Lys Thr Leu Ile Glu Lys
Thr Asn 50 55 60Glu Glu Arg Lys Thr
Leu Leu Ser Asn Leu Glu Glu Ala Lys Lys Lys65 70
75 80Lys Glu Asp Ala Leu Asn Glu Thr Arg Glu
Ser Glu Thr Lys Leu Lys85 90 95Glu Leu
Pro Gly Val Cys Asn Glu Thr Met Met Ala Leu Trp Glu Glu100
105 110Cys Lys Pro Cys Leu Lys Gln Thr Cys Met Lys Phe
Tyr Ala Arg Val115 120 125Cys Arg Ser Gly
Ser Gly Leu Val Gly Arg Gln Leu Glu Glu Phe Leu130 135
140Asn Gln Ser Ser Pro Phe Tyr Phe Trp Met Asn Gly Asp Arg
Ile Asp145 150 155 160Ser
Leu Leu Glu Asn Asp Arg Gln Gln Thr His Met Leu Asp Val Met165
170 175Gln Asp His Phe Ser Arg Ala Ser Ser Ile Ile
Asp Glu Leu Phe Gln180 185 190Asp Arg Phe
Phe Thr Arg Glu Pro Gln Asp Thr Tyr His Tyr Leu Pro195
200 205Phe Ser Leu Pro His Arg Arg Pro His Phe Phe Phe
Pro Lys Ser Arg210 215 220Ile Val Arg Ser
Leu Met Pro Phe Ser Pro Tyr Glu Pro Leu Asn Phe225 230
235 240His Ala Met Phe Gln Pro Phe Leu Glu
Met Ile His Glu Ala Gln Gln245 250 255Ala
Met Asp Ile His Phe His Ser Pro Ala Phe Gln His Pro Pro Thr260
265 270Glu Phe Ile Arg Glu Gly Asp Asp Asp Arg Thr
Val Cys Arg Glu Ile275 280 285Arg His Asn
Ser Thr Gly Cys Leu Arg Met Lys Asp Gln Cys Asp Lys290
295 300Cys Arg Glu Ile Leu Ser Val Asp Cys Ser Thr Asn
Asn Pro Ser Gln305 310 315
320Ala Lys Leu Arg Arg Glu Leu Asp Glu Ser Leu Gln Val Ala Glu Arg325
330 335Leu Thr Arg Lys Tyr Asn Glu Leu Leu
Lys Ser Tyr Gln Trp Lys Met340 345 350Leu
Asn Thr Ser Ser Leu Leu Glu Gln Leu Asn Glu Gln Phe Asn Trp355
360 365Val Ser Arg Leu Ala Asn Leu Thr Gln Gly Glu
Asp Gln Tyr Tyr Leu370 375 380Arg Val Thr
Thr Val Ala Ser His Thr Ser Asp Ser Asp Val Pro Ser385
390 395 400Gly Val Thr Glu Val Val Val
Lys Leu Phe Asp Ser Asp Pro Ile Thr405 410
415Val Thr Val Pro Val Glu Val Ser Arg Lys Asn Pro Lys Phe Met Glu420
425 430Thr Val Ala Glu Lys Ala Leu Gln Glu
Tyr Arg Lys Lys His Arg Glu435 440
445Glu88416PRTHomo sapiens 88Met Glu Leu Arg Val Gly Asn Lys Tyr Arg Leu
Gly Arg Lys Ile Gly1 5 10
15Ser Gly Ser Phe Gly Asp Ile Tyr Leu Gly Ala Asn Ile Ala Ser Gly
20 25 30Glu Glu Val Ala Ile Lys Leu
Glu Cys Val Lys Thr Lys His Pro Gln 35 40
45Leu His Ile Glu Ser Lys Phe Tyr Lys Met Met Gln Gly Gly Val
Gly 50 55 60Ile Pro Ser Ile Lys Trp
Cys Gly Ala Glu Gly Asp Tyr Asn Val Met65 70
75 80Val Met Glu Leu Leu Gly Pro Ser Leu Glu Asp
Leu Phe Asn Phe Cys85 90 95Ser Arg Lys
Phe Ser Leu Lys Thr Val Leu Leu Leu Ala Asp Gln Met100
105 110Ile Ser Arg Ile Glu Tyr Ile His Ser Lys Asn Phe
Ile His Arg Asp115 120 125Val Lys Pro Asp
Asn Phe Leu Met Gly Leu Gly Lys Lys Gly Asn Leu130 135
140Val Tyr Ile Ile Asp Phe Gly Leu Ala Lys Lys Tyr Arg Asp
Ala Arg145 150 155 160Thr
His Gln His Ile Pro Tyr Arg Glu Asn Lys Asn Leu Thr Gly Thr165
170 175Ala Arg Tyr Ala Ser Ile Asn Thr His Leu Gly
Ile Glu Gln Ser Arg180 185 190Arg Asp Asp
Leu Glu Ser Leu Gly Tyr Val Leu Met Tyr Phe Asn Leu195
200 205Gly Ser Leu Pro Trp Gln Gly Leu Lys Ala Ala Thr
Lys Arg Gln Lys210 215 220Tyr Glu Arg Ile
Ser Glu Lys Lys Met Ser Thr Pro Ile Glu Val Leu225 230
235 240Cys Lys Gly Tyr Pro Ser Glu Phe Ser
Thr Tyr Leu Asn Phe Cys Arg245 250 255Ser
Leu Arg Phe Asp Asp Lys Pro Asp Tyr Ser Tyr Leu Arg Gln Leu260
265 270Phe Arg Asn Leu Phe His Arg Gln Gly Phe Ser
Tyr Asp Tyr Val Phe275 280 285Asp Trp Asn
Met Leu Lys Phe Gly Ala Ala Arg Asn Pro Glu Asp Val290
295 300Asp Arg Glu Arg Arg Glu His Glu Arg Glu Glu Arg
Met Gly Gln Leu305 310 315
320Arg Gly Ser Ala Thr Arg Ala Leu Pro Pro Gly Pro Pro Thr Gly Ala325
330 335Thr Ala Asn Arg Leu Arg Ser Ala Ala
Glu Pro Val Ala Ser Thr Pro340 345 350Ala
Ser Arg Ile Gln Pro Ala Gly Asn Thr Ser Pro Arg Ala Ile Ser355
360 365Arg Val Asp Arg Glu Arg Lys Val Ser Met Arg
Leu His Arg Gly Ala370 375 380Pro Ala Asn
Val Ser Ser Ser Asp Leu Thr Gly Arg Gln Glu Val Ser385
390 395 400Arg Ile Pro Ala Ser Gln Thr
Ser Val Pro Phe Asp His Leu Gly Lys405 410
41589110PRTHomo sapiens 89Met Thr Thr Ala Ser Thr Ser Gln Val Arg Gln
Asn Tyr His Gln Asp1 5 10
15Ser Glu Ala Ala Ile Asn Arg Gln Ile Asn Leu Glu Leu Tyr Ala Ser
20 25 30Tyr Val Tyr Leu Ser Met Ser
Tyr Tyr Phe Asp Arg Asp Asp Val Ala 35 40
45Leu Lys Asn Phe Ala Lys Tyr Phe Leu His Gln Ser His Glu Glu
Arg 50 55 60Glu His Ala Glu Lys Leu
Met Lys Leu Gln Asn Gln Arg Gly Gly Arg65 70
75 80Ile Phe Leu Gln Asp Ile Lys Lys Pro Asp Cys
Asp Asp Trp Glu Ser85 90 95Gly Leu Asn
Ala Met Glu Cys Ala Leu His Leu Glu Lys Met100 105
11090814PRTHomo sapiens 90Met Arg Leu Ala Leu Leu Trp Ala Leu
Gly Leu Leu Gly Ala Gly Ser1 5 10
15Pro Leu Pro Ser Trp Pro Leu Pro Asn Ile Gly Gly Thr Glu Glu
Gln 20 25 30Gln Ala Glu Ser
Glu Lys Ala Pro Arg Glu Pro Leu Glu Pro Gln Val 35
40 45Leu Gln Asp Asp Leu Pro Ile Ser Leu Lys Lys Val
Leu Gln Thr Ser 50 55 60Leu Pro Glu
Pro Leu Arg Ile Lys Leu Glu Leu Asp Gly Asp Ser His65 70
75 80Ile Leu Glu Leu Leu Gln Asn Arg
Glu Leu Val Pro Gly Arg Pro Thr85 90
95Leu Val Trp Tyr Gln Pro Asp Gly Thr Arg Val Val Ser Glu Gly His100
105 110Thr Leu Glu Asn Cys Cys Tyr Gln Gly Arg
Val Arg Gly Tyr Ala Gly115 120 125Ser Trp
Val Ser Ile Cys Thr Cys Ser Gly Leu Arg Gly Leu Val Val130
135 140Leu Thr Pro Glu Arg Ser Tyr Thr Leu Glu Gln Gly
Pro Gly Asp Leu145 150 155
160Gln Gly Pro Pro Ile Ile Ser Arg Ile Gln Asp Leu His Leu Pro Gly165
170 175His Thr Cys Ala Leu Ser Trp Arg Glu
Ser Val His Thr Gln Thr Pro180 185 190Pro
Glu His Pro Leu Gly Gln Arg His Ile Arg Arg Arg Arg Asp Val195
200 205Val Thr Glu Thr Lys Thr Val Glu Leu Val Ile
Val Ala Asp His Ser210 215 220Glu Ala Gln
Lys Tyr Arg Asp Phe Gln His Leu Leu Asn Arg Thr Leu225
230 235 240Glu Val Ala Leu Leu Leu Asp
Thr Phe Phe Arg Pro Leu Asn Val Arg245 250
255Val Ala Leu Val Gly Leu Glu Ala Trp Thr Gln Arg Asp Leu Val Glu260
265 270Ile Ser Pro Asn Pro Ala Val Thr Leu
Glu Asn Phe Leu His Trp Arg275 280 285Arg
Ala His Leu Leu Pro Arg Leu Pro His Asp Ser Ala Gln Leu Val290
295 300Thr Gly Thr Ser Phe Ser Gly Pro Thr Val Gly
Met Ala Ile Gln Asn305 310 315
320Ser Ile Cys Ser Pro Asp Phe Ser Gly Gly Val Asn Met Asp His
Ser325 330 335Thr Ser Ile Leu Gly Val Ala
Ser Ser Ile Ala His Glu Leu Gly His340 345
350Ser Leu Gly Leu Asp His Asp Leu Pro Gly Asn Ser Cys Pro Cys Pro355
360 365Gly Pro Ala Pro Ala Lys Thr Cys Ile
Met Glu Ala Ser Thr Asp Phe370 375 380Leu
Pro Gly Leu Asn Phe Ser Asn Cys Ser Arg Arg Ala Leu Glu Lys385
390 395 400Ala Leu Leu Asp Gly Met
Gly Ser Cys Leu Phe Glu Arg Leu Pro Ser405 410
415Leu Pro Pro Met Ala Ala Phe Cys Gly Asn Met Phe Val Glu Pro
Gly420 425 430Glu Gln Cys Asp Cys Gly Phe
Leu Asp Asp Cys Val Asp Pro Cys Cys435 440
445Asp Ser Leu Thr Cys Gln Leu Arg Pro Gly Ala Gln Cys Ala Ser Asp450
455 460Gly Pro Cys Cys Gln Asn Cys Gln Leu
Arg Pro Ser Gly Trp Gln Cys465 470 475
480Arg Pro Thr Arg Gly Asp Cys Asp Leu Pro Glu Phe Cys Pro
Gly Asp485 490 495Ser Ser Gln Cys Pro Pro
Asp Val Ser Leu Gly Asp Gly Glu Pro Cys500 505
510Ala Gly Gly Gln Ala Val Cys Met His Gly Arg Cys Ala Ser Tyr
Ala515 520 525Gln Gln Cys Gln Ser Leu Trp
Gly Pro Gly Ala Gln Pro Ala Ala Pro530 535
540Leu Cys Leu Gln Thr Ala Asn Thr Arg Gly Asn Ala Phe Gly Ser Cys545
550 555 560Gly Arg Asn Pro
Ser Gly Ser Tyr Val Ser Cys Thr Pro Arg Asp Ala565 570
575Ile Cys Gly Gln Leu Gln Cys Gln Thr Gly Arg Thr Gln Pro
Leu Leu580 585 590Gly Ser Ile Arg Asp Leu
Leu Trp Glu Thr Ile Asp Val Asn Gly Thr595 600
605Glu Leu Asn Cys Ser Trp Val His Leu Asp Leu Gly Ser Asp Val
Ala610 615 620Gln Pro Leu Leu Thr Leu Pro
Gly Thr Ala Cys Gly Pro Gly Leu Val625 630
635 640Cys Ile Asp His Arg Cys Gln Arg Val Asp Leu Leu
Gly Ala Gln Glu645 650 655Cys Arg Ser Lys
Cys His Gly His Gly Val Cys Asp Ser Asn Arg His660 665
670Cys Tyr Cys Glu Glu Gly Trp Ala Pro Pro Asp Cys Thr Thr
Gln Leu675 680 685Lys Ala Thr Ser Ser Leu
Thr Thr Gly Leu Leu Leu Ser Leu Leu Val690 695
700Leu Leu Val Leu Val Met Leu Gly Ala Gly Tyr Trp Tyr Arg Ala
Arg705 710 715 720Leu His
Gln Arg Leu Cys Gln Leu Lys Gly Pro Thr Cys Gln Tyr Arg725
730 735Ala Ala Gln Ser Gly Pro Ser Glu Arg Pro Gly Pro
Pro Gln Arg Ala740 745 750Leu Leu Ala Arg
Gly Thr Lys Ser Gln Gly Pro Ala Lys Pro Pro Pro755 760
765Pro Arg Lys Pro Leu Pro Ala Asp Pro Gln Gly Arg Cys Pro
Ser Gly770 775 780Asp Leu Pro Gly Pro Gly
Ala Gly Ile Pro Pro Leu Val Val Pro Ser785 790
795 800Arg Pro Ala Pro Pro Pro Pro Thr Val Ser Ser
Leu Tyr Leu805 8109155PRTHomo sapiens 91Met Asn Leu Ser
Phe Arg Glu Phe Asn Gln Glu Lys Arg Val Gly Gly1 5
10 15Ile Ser Trp Gly Pro Lys Gly Arg Leu Ser
Gly Ile Phe Ser Thr Ile 20 25
30Gln Asn Gln Gln Gln Ser Gln Lys Arg Gly Met Ser Ser Asn Ser Leu
35 40 45Lys Arg Thr Pro Gln Asn Ser
50 559254PRTHomo sapiens 92Met Gly Asn Gln Arg Trp His
Ala Lys Phe Asn Ser Gly Leu Arg Tyr1 5 10
15Pro His Cys Pro His Gln Ala Ser Pro Ala Leu Thr Val
Glu Pro His 20 25 30Gly Glu
Glu His Val Leu Glu Arg Asp Pro Phe Val Asn Cys Phe Val 35
40 45Val Phe Ser Ser Met Asn 509351PRTHomo
sapiens 93Met Leu Cys Ala Gln Gly Ala Ala Gly Cys Gln Gln His Leu Ser
Leu1 5 10 15Asn Thr Ile
Ser Leu Cys Ala Glu Lys Thr Gly Asn Gln Arg Ile Asn 20
25 30Ile Thr Ser Pro Gly Trp Arg Thr Ile Ser
Cys Asp Phe Ala Ala Glu 35 40
45Phe Thr His 509443PRTHomo sapiens 94Met Pro Pro Leu Ile Pro His Ala
Ala Lys Arg Ile Gly Thr Leu Ser1 5 10
15Gly Pro Gly Thr Val Val Met Ala Ile Ser Tyr Phe Thr His
Thr Arg 20 25 30Pro Phe Lys
Val Ser Leu Pro Gln Ala Ile Lys 35 409539PRTHomo
sapiens 95Met Val Glu Asn Ile Pro Glu Ser Leu Pro Phe Gly Pro Gln Leu
Met1 5 10 15Pro Pro Thr
Leu Phe Ser Trp Leu Asn Ser Leu Lys Glu Arg Phe Met 20
25 30Cys Tyr Cys Pro Val Ser Gln
359636PRTHomo sapiens 96Met Ser Gln Cys Thr Ser Tyr Pro Leu Ile Gln Lys
Glu Glu His Phe1 5 10
15Ala Gln Arg Lys Ile Lys Arg Ser Met Asn Val Ile Phe Tyr Leu Leu
20 25 30Phe Ser Val Gly
359733PRTHomo sapiens 97Met Gly Ser Ser Leu Pro Ile Gly Phe Leu Leu His
Thr Ala Gly Leu1 5 10
15Ser Leu Tyr Phe Lys Lys Lys Lys Lys Lys Lys Lys Asp Lys Asn Cys
20 25 30His 98317PRTHomo sapiens
98Met Ser Glu Gly Val Gly Thr Phe Arg Met Val Pro Glu Glu Glu Gln1
5 10 15Glu Leu Arg Ala Gln Leu
Glu Gln Leu Thr Thr Lys Asp His Gly Pro 20 25
30Val Phe Gly Pro Cys Ser Gln Leu Pro Arg His Thr Leu
Gln Lys Ala 35 40 45Lys Asp Glu
Leu Asn Glu Arg Glu Glu Thr Arg Glu Glu Ala Val Arg 50
55 60Glu Leu Gln Glu Met Val Gln Ala Gln Ala Ala Ser
Gly Glu Glu Leu65 70 75
80Ala Val Ala Val Ala Glu Arg Val Gln Glu Lys Asp Ser Gly Phe Phe85
90 95Leu Arg Phe Ile Arg Ala Arg Lys Phe Asn
Val Gly Arg Ala Tyr Glu100 105 110Leu Leu
Arg Gly Tyr Val Asn Phe Arg Leu Gln Tyr Pro Glu Leu Phe115
120 125Asp Ser Leu Ser Pro Glu Ala Val Arg Cys Thr Ile
Glu Ala Gly Tyr130 135 140Pro Gly Val Leu
Ser Ser Arg Asp Lys Tyr Gly Arg Val Val Met Leu145 150
155 160Phe Asn Ile Glu Asn Trp Gln Ser Gln
Glu Ile Thr Phe Asp Glu Ile165 170 175Leu
Gln Ala Tyr Cys Phe Ile Leu Glu Lys Leu Leu Glu Asn Glu Glu180
185 190Thr Gln Ile Asn Gly Phe Cys Ile Ile Glu Asn
Phe Lys Gly Phe Thr195 200 205Met Gln Gln
Ala Ala Ser Leu Arg Thr Ser Asp Leu Arg Lys Met Val210
215 220Asp Met Leu Gln Asp Ser Phe Pro Ala Arg Phe Lys
Ala Ile His Phe225 230 235
240Ile His Gln Pro Trp Tyr Phe Thr Thr Thr Tyr Asn Val Val Lys Pro245
250 255Phe Leu Lys Ser Lys Leu Leu Glu Arg
Val Phe Val His Gly Asp Asp260 265 270Leu
Ser Gly Phe Tyr Gln Glu Ile Asp Glu Asn Ile Leu Pro Ser Asp275
280 285Phe Gly Gly Thr Leu Pro Lys Tyr Asp Gly Lys
Ala Val Ala Glu Gln290 295 300Leu Phe Gly
Pro Gln Ala Gln Ala Glu Asn Thr Ala Phe305 310
31599375PRTHomo sapiens 99Met Glu Glu Glu Ile Ala Ala Leu Val Ile
Asp Asn Gly Ser Gly Met1 5 10
15Cys Lys Ala Gly Phe Ala Gly Asp Asp Ala Pro Arg Ala Val Phe Pro
20 25 30Ser Ile Val Gly Arg Pro
Arg His Gln Gly Val Met Val Gly Met Gly 35 40
45Gln Lys Asp Ser Tyr Val Gly Asp Glu Ala Gln Ser Lys Arg
Gly Ile 50 55 60Leu Thr Leu Lys Tyr
Pro Ile Glu His Gly Ile Val Thr Asn Trp Asp65 70
75 80Asp Met Glu Lys Ile Trp His His Thr Phe
Tyr Asn Glu Leu Arg Val85 90 95Ala Pro
Glu Glu His Pro Val Leu Leu Thr Glu Ala Pro Leu Asn Pro100
105 110Lys Ala Asn Arg Glu Lys Met Thr Gln Ile Met Phe
Glu Thr Phe Asn115 120 125Thr Pro Ala Met
Tyr Val Ala Ile Gln Ala Val Leu Ser Leu Tyr Ala130 135
140Ser Gly Arg Thr Thr Gly Ile Val Met Asp Ser Gly Asp Gly
Val Thr145 150 155 160His
Thr Val Pro Ile Tyr Glu Gly Tyr Ala Leu Pro His Ala Ile Leu165
170 175Arg Leu Asp Leu Ala Gly Arg Asp Leu Thr Asp
Tyr Leu Met Lys Ile180 185 190Leu Thr Glu
Arg Gly Tyr Ser Phe Thr Thr Thr Ala Glu Arg Glu Ile195
200 205Val Arg Asp Ile Lys Glu Lys Leu Cys Tyr Val Ala
Leu Asp Phe Glu210 215 220Gln Glu Met Ala
Thr Ala Ala Ser Ser Ser Ser Leu Glu Lys Ser Tyr225 230
235 240Glu Leu Pro Asp Gly Gln Val Ile Thr
Ile Gly Asn Glu Arg Phe Arg245 250 255Cys
Pro Glu Ala Leu Phe Gln Pro Ser Phe Leu Gly Met Glu Ser Cys260
265 270Gly Ile His Glu Thr Thr Phe Asn Ser Ile Met
Lys Cys Asp Val Asp275 280 285Ile Arg Lys
Asp Leu Tyr Ala Asn Thr Val Leu Ser Gly Gly Thr Thr290
295 300Met Tyr Pro Gly Ile Ala Asp Arg Met Gln Lys Glu
Ile Thr Ala Leu305 310 315
320Ala Pro Ser Thr Met Lys Ile Lys Ile Ile Ala Pro Pro Glu Arg Lys325
330 335Tyr Ser Val Trp Ile Gly Gly Ser Ile
Leu Ala Ser Leu Ser Thr Phe340 345 350Gln
Gln Met Trp Ile Ser Lys Gln Glu Tyr Asp Glu Ser Gly Pro Ser355
360 365Ile Val His Arg Lys Cys Phe370
375100582PRTHomo sapiens 100Met Ser Pro Ala Pro Arg Pro Ser Arg Cys Leu
Leu Leu Pro Leu Leu1 5 10
15Thr Leu Gly Thr Ala Leu Ala Ser Leu Gly Ser Ala Gln Ser Ser Ser
20 25 30Phe Ser Pro Glu Ala Trp Leu
Gln Gln Tyr Gly Tyr Leu Pro Pro Gly 35 40
45Asp Leu Arg Thr His Thr Gln Arg Ser Pro Gln Ser Leu Ser Ala
Ala 50 55 60Ile Ala Ala Met Gln Lys
Phe Tyr Gly Leu Gln Val Thr Gly Lys Ala65 70
75 80Asp Ala Asp Thr Met Lys Ala Met Arg Arg Pro
Arg Cys Gly Val Pro85 90 95Asp Lys Phe
Gly Ala Glu Ile Lys Ala Asn Val Arg Arg Lys Arg Tyr100
105 110Ala Ile Gln Gly Leu Lys Trp Gln His Asn Glu Ile
Thr Phe Cys Ile115 120 125Gln Asn Tyr Thr
Pro Lys Val Gly Glu Tyr Ala Thr Tyr Glu Ala Ile130 135
140Arg Lys Ala Phe Arg Val Trp Glu Ser Ala Thr Pro Leu Arg
Phe Arg145 150 155 160Glu
Val Pro Tyr Ala Tyr Ile Arg Glu Gly His Glu Lys Gln Ala Asp165
170 175Ile Met Ile Phe Phe Ala Glu Gly Phe His Gly
Asp Ser Thr Pro Phe180 185 190Asp Gly Glu
Gly Gly Phe Leu Ala His Ala Tyr Phe Pro Gly Pro Asn195
200 205Ile Gly Gly Asp Thr His Phe Asp Ser Ala Glu Pro
Trp Thr Val Arg210 215 220Asn Glu Asp Leu
Asn Gly Asn Asp Ile Phe Leu Val Ala Val His Glu225 230
235 240Leu Gly His Ala Leu Gly Leu Glu His
Ser Ser Asp Pro Ser Ala Ile245 250 255Met
Ala Pro Phe Tyr Gln Trp Met Asp Thr Glu Asn Phe Val Leu Pro260
265 270Asp Asp Asp Arg Arg Gly Ile Gln Gln Leu Tyr
Gly Gly Glu Ser Gly275 280 285Phe Pro Thr
Lys Met Pro Pro Gln Pro Arg Thr Thr Ser Arg Pro Ser290
295 300Val Pro Asp Lys Pro Lys Asn Pro Thr Tyr Gly Pro
Asn Ile Cys Asp305 310 315
320Gly Asn Phe Asp Thr Val Ala Met Leu Arg Gly Glu Met Phe Val Phe325
330 335Lys Glu Arg Trp Phe Trp Arg Val Arg
Asn Asn Gln Val Met Asp Gly340 345 350Tyr
Pro Met Pro Ile Gly Gln Phe Trp Arg Gly Leu Pro Ala Ser Ile355
360 365Asn Thr Ala Tyr Glu Arg Lys Asp Gly Lys Phe
Val Phe Phe Lys Gly370 375 380Asp Lys His
Trp Val Phe Asp Glu Ala Ser Leu Glu Pro Gly Tyr Pro385
390 395 400Lys His Ile Lys Glu Leu Gly
Arg Gly Leu Pro Thr Asp Lys Ile Asp405 410
415Ala Ala Leu Phe Trp Met Pro Asn Gly Lys Thr Tyr Phe Phe Arg Gly420
425 430Asn Lys Tyr Tyr Arg Phe Asn Glu Glu
Leu Arg Ala Val Asp Ser Glu435 440 445Tyr
Pro Lys Asn Ile Lys Val Trp Glu Gly Ile Pro Glu Ser Pro Arg450
455 460Gly Ser Phe Met Gly Ser Asp Glu Val Phe Thr
Tyr Phe Tyr Lys Gly465 470 475
480Asn Lys Tyr Trp Lys Phe Asn Asn Gln Lys Leu Lys Val Glu Pro
Gly485 490 495Tyr Pro Lys Ser Ala Leu Arg
Asp Trp Met Gly Cys Pro Ser Gly Gly500 505
510Arg Pro Asp Glu Gly Thr Glu Glu Glu Thr Glu Val Ile Ile Ile Glu515
520 525Val Asp Glu Glu Gly Gly Gly Ala Val
Ser Ala Ala Ala Val Val Leu530 535 540Pro
Val Leu Leu Leu Leu Leu Val Leu Ala Val Gly Leu Ala Val Phe545
550 555 560Phe Phe Arg Arg His Gly
Thr Pro Arg Arg Leu Leu Tyr Cys Gln Arg565 570
575Ser Leu Leu Asp Lys Val5801012285PRTHomo sapiens 101Met Ala Ala
Gln Val Ala Pro Ala Ala Ala Ser Ser Leu Gly Asn Pro1 5
10 15Pro Pro Pro Pro Pro Ser Glu Leu Lys
Lys Ala Glu Gln Gln Gln Arg 20 25
30Glu Glu Ala Gly Gly Glu Ala Ala Ala Ala Ala Ala Ala Glu Arg Gly
35 40 45Glu Met Lys Ala Ala Ala Gly
Gln Glu Ser Glu Gly Pro Ala Val Gly 50 55
60Pro Pro Gln Pro Leu Gly Lys Glu Leu Gln Asp Gly Ala Glu Ser Asn65
70 75 80Gly Gly Gly Gly
Gly Gly Gly Ala Gly Ser Gly Gly Gly Pro Gly Ala85 90
95Glu Pro Asp Leu Lys Asn Ser Asn Gly Asn Ala Gly Pro Arg
Pro Ala100 105 110Leu Asn Asn Asn Leu Thr
Glu Pro Pro Gly Gly Gly Gly Gly Gly Ser115 120
125Ser Asp Gly Val Gly Ala Pro Pro His Ser Ala Ala Ala Ala Leu
Pro130 135 140Pro Pro Ala Tyr Gly Phe Gly
Gln Pro Tyr Gly Arg Ser Pro Ser Ala145 150
155 160Val Ala Ala Ala Ala Ala Ala Val Phe His Gln Gln
His Gly Gly Gln165 170 175Gln Ser Pro Gly
Leu Ala Ala Leu Gln Ser Gly Gly Gly Gly Gly Leu180 185
190Glu Pro Tyr Ala Gly Pro Gln Gln Asn Ser His Asp His Gly
Phe Pro195 200 205Asn His Gln Tyr Asn Ser
Tyr Tyr Pro Asn Arg Ser Ala Tyr Pro Pro210 215
220Pro Ala Pro Ala Tyr Ala Leu Ser Ser Pro Arg Gly Gly Thr Pro
Gly225 230 235 240Ser Gly
Ala Ala Ala Ala Ala Gly Ser Lys Pro Pro Pro Ser Ser Ser245
250 255Ala Ser Ala Ser Ser Ser Ser Ser Ser Phe Ala Gln
Gln Arg Phe Gly260 265 270Ala Met Gly Gly
Gly Gly Pro Ser Ala Ala Gly Gly Gly Thr Pro Gln275 280
285Pro Thr Ala Thr Pro Thr Leu Asn Gln Leu Leu Thr Ser Pro
Ser Ser290 295 300Ala Arg Gly Tyr Gln Gly
Tyr Pro Gly Gly Asp Tyr Ser Gly Gly Pro305 310
315 320Gln Asp Gly Gly Ala Gly Lys Gly Pro Ala Asp
Met Ala Ser Gln Cys325 330 335Trp Gly Ala
Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala Ser Gly Gly340
345 350Ala Gln Gln Arg Ser His His Ala Pro Met Ser Pro
Gly Ser Ser Gly355 360 365Gly Gly Gly Gln
Pro Leu Ala Arg Thr Pro Gln Pro Ser Ser Pro Met370 375
380Asp Gln Met Gly Lys Met Arg Pro Gln Pro Tyr Gly Gly Thr
Asn Pro385 390 395 400Tyr
Ser Gln Gln Gln Gly Pro Pro Ser Gly Pro Gln Gln Gly His Gly405
410 415Tyr Pro Gly Gln Pro Tyr Gly Ser Gln Thr Pro
Gln Arg Tyr Pro Met420 425 430Thr Met Gln
Gly Arg Ala Gln Ser Ala Met Gly Gly Leu Ser Tyr Thr435
440 445Gln Gln Ile Pro Pro Tyr Gly Gln Gln Gly Pro Ser
Gly Tyr Gly Gln450 455 460Gln Gly Gln Thr
Pro Tyr Tyr Asn Gln Gln Ser Pro His Pro Gln Gln465 470
475 480Gln Gln Pro Pro Tyr Ser Gln Gln Pro
Pro Ser Gln Thr Pro His Ala485 490 495Gln
Pro Ser Tyr Gln Gln Gln Pro Gln Ser Gln Pro Pro Gln Leu Gln500
505 510Ser Ser Gln Pro Pro Tyr Ser Gln Gln Pro Ser
Gln Pro Pro His Gln515 520 525Gln Ser Pro
Ala Pro Tyr Pro Ser Gln Gln Ser Thr Thr Gln Gln His530
535 540Pro Gln Ser Gln Pro Pro Tyr Ser Gln Pro Gln Ala
Gln Ser Pro Tyr545 550 555
560Gln Gln Gln Gln Pro Gln Gln Pro Ala Pro Ser Thr Leu Ser Gln Gln565
570 575Ala Ala Tyr Pro Gln Pro Gln Ser Gln
Gln Ser Gln Gln Thr Ala Tyr580 585 590Ser
Gln Gln Arg Phe Pro Pro Pro Gln Glu Leu Ser Gln Asp Ser Phe595
600 605Gly Ser Gln Ala Ser Ser Ala Pro Ser Met Thr
Ser Ser Lys Gly Gly610 615 620Gln Glu Asp
Met Asn Leu Ser Leu Gln Ser Arg Pro Ser Ser Leu Pro625
630 635 640Asp Leu Ser Gly Ser Ile Asp
Asp Leu Pro Met Gly Thr Glu Gly Ala645 650
655Leu Ser Pro Gly Val Ser Thr Ser Gly Ile Ser Ser Ser Gln Gly Glu660
665 670Gln Ser Asn Pro Ala Gln Ser Pro Phe
Ser Pro His Thr Ser Pro His675 680 685Leu
Pro Gly Ile Arg Gly Pro Ser Pro Ser Pro Val Gly Ser Pro Ala690
695 700Ser Val Ala Gln Ser Arg Ser Gly Pro Leu Ser
Pro Ala Ala Val Pro705 710 715
720Gly Asn Gln Met Pro Pro Arg Pro Pro Ser Gly Gln Ser Asp Ser
Ile725 730 735Met His Pro Ser Met Asn Gln
Ser Ser Ile Ala Gln Asp Arg Gly Tyr740 745
750Met Gln Arg Asn Pro Gln Met Pro Gln Tyr Ser Ser Pro Gln Pro Gly755
760 765Ser Ala Leu Ser Pro Arg Gln Pro Ser
Gly Gly Gln Ile His Thr Gly770 775 780Met
Gly Ser Tyr Gln Gln Asn Ser Met Gly Ser Tyr Gly Pro Gln Gly785
790 795 800Gly Gln Tyr Gly Pro Gln
Gly Gly Tyr Pro Arg Gln Pro Asn Tyr Asn805 810
815Ala Leu Pro Asn Ala Asn Tyr Pro Ser Ala Gly Met Ala Gly Gly
Ile820 825 830Asn Pro Met Gly Ala Gly Gly
Gln Met His Gly Gln Pro Gly Ile Pro835 840
845Pro Tyr Gly Thr Leu Pro Pro Gly Arg Met Ser His Ala Ser Met Gly850
855 860Asn Arg Pro Tyr Gly Pro Asn Met Ala
Asn Met Pro Pro Gln Val Gly865 870 875
880Ser Gly Met Cys Pro Pro Pro Gly Gly Met Asn Arg Lys Thr
Gln Glu885 890 895Thr Ala Val Ala Met His
Val Ala Ala Asn Ser Ile Gln Asn Arg Pro900 905
910Pro Gly Tyr Pro Asn Met Asn Gln Gly Gly Met Met Gly Thr Gly
Pro915 920 925Pro Tyr Gly Gln Gly Ile Asn
Ser Met Ala Gly Met Ile Asn Pro Gln930 935
940Gly Pro Pro Tyr Ser Met Gly Gly Thr Met Ala Asn Asn Ser Ala Gly945
950 955 960Met Ala Ala Ser
Pro Glu Met Met Gly Leu Gly Asp Val Lys Leu Thr965 970
975Pro Ala Thr Lys Met Asn Asn Lys Ala Asp Gly Thr Pro Lys
Thr Glu980 985 990Ser Lys Ser Lys Lys Ser
Ser Ser Ser Thr Thr Thr Asn Glu Lys Ile995 1000
1005Thr Lys Leu Tyr Glu Leu Gly Gly Glu Pro Glu Arg Lys Met
Trp1010 1015 1020Val Asp Arg Tyr Leu Ala
Phe Thr Glu Glu Lys Ala Met Gly Met1025 1030
1035Thr Asn Leu Pro Ala Val Gly Arg Lys Pro Leu Asp Leu Tyr
Arg1040 1045 1050Leu Tyr Val Ser Val Lys
Glu Ile Gly Gly Leu Thr Gln Val Asn1055 1060
1065Lys Asn Lys Lys Trp Arg Glu Leu Ala Thr Asn Leu Asn Val
Gly1070 1075 1080Thr Ser Ser Ser Ala Ala
Ser Ser Leu Lys Lys Gln Tyr Ile Gln1085 1090
1095Cys Leu Tyr Ala Phe Glu Cys Lys Ile Glu Arg Gly Glu Asp
Pro1100 1105 1110Pro Pro Asp Ile Phe Ala
Ala Ala Asp Ser Lys Lys Ser Gln Pro1115 1120
1125Lys Ile Gln Pro Pro Ser Pro Ala Gly Ser Gly Ser Met Gln
Gly1130 1135 1140Pro Gln Thr Pro Gln Ser
Thr Ser Ser Ser Met Ala Glu Gly Gly1145 1150
1155Asp Leu Lys Pro Pro Thr Pro Ala Ser Thr Pro His Ser Gln
Ile1160 1165 1170Pro Pro Leu Pro Gly Met
Ser Arg Ser Asn Ser Val Gly Ile Gln1175 1180
1185Asp Ala Phe Asn Asp Gly Ser Asp Ser Thr Phe Gln Lys Arg
Asn1190 1195 1200Ser Met Thr Pro Asn Pro
Gly Tyr Gln Pro Ser Met Asn Thr Ser1205 1210
1215Asp Met Met Gly Arg Met Ser Tyr Glu Pro Asn Lys Asp Pro
Tyr1220 1225 1230Gly Ser Met Arg Lys Ala
Pro Gly Ser Asp Pro Phe Met Ser Ser1235 1240
1245Gly Gln Gly Pro Asn Gly Gly Met Gly Asp Pro Tyr Ser Arg
Ala1250 1255 1260Ala Gly Pro Gly Leu Gly
Asn Val Ala Met Gly Pro Arg Gln His1265 1270
1275Tyr Pro Tyr Gly Gly Pro Tyr Asp Arg Val Arg Thr Glu Pro
Gly1280 1285 1290Ile Gly Pro Glu Gly Asn
Met Ser Thr Gly Ala Pro Gln Pro Asn1295 1300
1305Leu Met Pro Ser Asn Pro Asp Ser Gly Met Tyr Ser Pro Ser
Arg1310 1315 1320Tyr Pro Pro Gln Gln Gln
Gln Gln Gln Gln Gln Arg His Asp Ser1325 1330
1335Tyr Gly Asn Gln Phe Ser Thr Gln Gly Thr Pro Ser Gly Ser
Pro1340 1345 1350Phe Pro Ser Gln Gln Thr
Thr Met Tyr Gln Gln Gln Gln Gln Asn1355 1360
1365Tyr Lys Arg Pro Met Asp Gly Thr Tyr Gly Pro Pro Ala Lys
Arg1370 1375 1380His Glu Gly Glu Met Tyr
Ser Val Pro Tyr Ser Thr Gly Gln Gly1385 1390
1395Gln Pro Gln Gln Gln Gln Leu Pro Pro Ala Gln Pro Gln Pro
Ala1400 1405 1410Ser Gln Gln Gln Ala Ala
Gln Pro Ser Pro Gln Gln Asp Val Tyr1415 1420
1425Asn Gln Tyr Gly Asn Ala Tyr Pro Ala Thr Ala Thr Ala Ala
Thr1430 1435 1440Glu Arg Arg Pro Ala Gly
Gly Pro Gln Asn Gln Phe Pro Phe Gln1445 1450
1455Phe Gly Arg Asp Arg Val Ser Ala Pro Pro Gly Thr Asn Ala
Gln1460 1465 1470Gln Asn Met Pro Pro Gln
Met Met Gly Gly Pro Ile Gln Ala Ser1475 1480
1485Ala Glu Val Ala Gln Gln Gly Thr Met Trp Gln Gly Arg Asn
Asp1490 1495 1500Met Thr Tyr Asn Tyr Ala
Asn Arg Gln Ser Thr Gly Ser Ala Pro1505 1510
1515Gln Gly Pro Ala Tyr His Gly Val Asn Arg Thr Asp Glu Met
Leu1520 1525 1530His Thr Asp Gln Arg Ala
Asn His Glu Gly Ser Trp Pro Ser His1535 1540
1545Gly Thr Arg Gln Pro Pro Tyr Gly Pro Ser Ala Pro Val Pro
Pro1550 1555 1560Met Thr Arg Pro Pro Pro
Ser Asn Tyr Gln Pro Pro Pro Ser Met1565 1570
1575Gln Asn His Ile Pro Gln Val Ser Ser Pro Ala Pro Leu Pro
Arg1580 1585 1590Pro Met Glu Asn Arg Thr
Ser Pro Ser Lys Ser Pro Phe Leu His1595 1600
1605Ser Gly Met Lys Met Gln Lys Ala Gly Pro Pro Val Pro Ala
Ser1610 1615 1620His Ile Ala Pro Ala Pro
Val Gln Pro Pro Met Ile Arg Arg Asp1625 1630
1635Ile Thr Phe Pro Pro Gly Ser Val Glu Ala Thr Gln Pro Val
Leu1640 1645 1650Lys Gln Arg Arg Arg Leu
Thr Met Lys Asp Ile Gly Thr Pro Glu1655 1660
1665Ala Trp Arg Val Met Met Ser Leu Lys Ser Gly Leu Leu Ala
Glu1670 1675 1680Ser Thr Trp Ala Leu Asp
Thr Ile Asn Ile Leu Leu Tyr Asp Asp1685 1690
1695Asn Ser Ile Met Thr Phe Asn Leu Ser Gln Leu Pro Gly Leu
Leu1700 1705 1710Glu Leu Leu Val Glu Tyr
Phe Arg Arg Cys Leu Ile Glu Ile Phe1715 1720
1725Gly Ile Leu Lys Glu Tyr Glu Val Gly Asp Pro Gly Gln Arg
Thr1730 1735 1740Leu Leu Asp Pro Gly Arg
Phe Ser Lys Val Ser Ser Pro Ala Pro1745 1750
1755Met Glu Gly Gly Glu Glu Glu Glu Glu Leu Leu Gly Pro Lys
Leu1760 1765 1770Glu Glu Glu Glu Glu Glu
Glu Val Val Glu Asn Asp Glu Glu Ile1775 1780
1785Ala Phe Ser Gly Lys Asp Lys Pro Ala Ser Glu Asn Ser Glu
Glu1790 1795 1800Lys Leu Ile Ser Lys Phe
Asp Lys Leu Pro Val Lys Ile Val Gln1805 1810
1815Lys Asn Asp Pro Phe Val Val Asp Cys Ser Asp Lys Leu Gly
Arg1820 1825 1830Val Gln Glu Phe Asp Ser
Gly Leu Leu His Trp Arg Ile Gly Gly1835 1840
1845Gly Asp Thr Thr Glu His Ile Gln Thr His Phe Glu Ser Lys
Thr1850 1855 1860Glu Leu Leu Pro Ser Arg
Pro His Ala Pro Cys Pro Pro Ala Pro1865 1870
1875Arg Lys His Val Thr Thr Ala Glu Gly Thr Pro Gly Thr Thr
Asp1880 1885 1890Gln Glu Gly Pro Pro Pro
Asp Gly Pro Pro Glu Lys Arg Ile Thr1895 1900
1905Ala Thr Met Asp Asp Met Leu Ser Thr Arg Ser Ser Thr Leu
Thr1910 1915 1920Glu Asp Gly Ala Lys Ser
Ser Glu Ala Ile Lys Glu Ser Ser Lys1925 1930
1935Phe Pro Phe Gly Ile Ser Pro Ala Gln Ser His Arg Asn Ile
Lys1940 1945 1950Ile Leu Glu Asp Glu Pro
His Ser Lys Asp Glu Thr Pro Leu Cys1955 1960
1965Thr Leu Leu Asp Trp Gln Asp Ser Leu Ala Lys Arg Cys Val
Cys1970 1975 1980Val Ser Asn Thr Ile Arg
Ser Leu Ser Phe Val Pro Gly Asn Asp1985 1990
1995Phe Glu Met Ser Lys His Pro Gly Leu Leu Leu Ile Leu Gly
Lys2000 2005 2010Leu Ile Leu Leu His His
Lys His Pro Glu Arg Lys Gln Ala Pro2015 2020
2025Leu Thr Tyr Glu Lys Glu Glu Glu Gln Asp Gln Gly Val Ser
Cys2030 2035 2040Asn Lys Val Glu Trp Trp
Trp Asp Cys Leu Glu Met Leu Arg Glu2045 2050
2055Asn Thr Leu Val Thr Leu Ala Asn Ile Ser Gly Gln Leu Asp
Leu2060 2065 2070Ser Pro Tyr Pro Glu Ser
Ile Cys Leu Pro Val Leu Asp Gly Leu2075 2080
2085Leu His Trp Ala Val Cys Pro Ser Ala Glu Ala Gln Asp Pro
Phe2090 2095 2100Ser Thr Leu Gly Pro Asn
Ala Val Leu Ser Pro Gln Arg Leu Val2105 2110
2115Leu Glu Thr Leu Ser Lys Leu Ser Ile Gln Asp Asn Asn Val
Asp2120 2125 2130Leu Ile Leu Ala Thr Pro
Pro Phe Ser Arg Leu Glu Lys Leu Tyr2135 2140
2145Ser Thr Met Val Arg Phe Leu Ser Asp Arg Lys Asn Pro Val
Cys2150 2155 2160Arg Glu Met Ala Val Val
Leu Leu Ala Asn Leu Ala Gln Gly Asp2165 2170
2175Ser Leu Ala Ala Arg Ala Ile Ala Val Gln Lys Gly Ser Ile
Gly2180 2185 2190Asn Leu Leu Gly Phe Leu
Glu Asp Ser Leu Ala Ala Thr Gln Phe2195 2200
2205Gln Gln Ser Gln Ala Ser Leu Leu His Met Gln Asn Pro Pro
Phe2210 2215 2220Glu Pro Thr Ser Val Asp
Met Met Arg Arg Ala Ala Arg Ala Leu2225 2230
2235Leu Ala Leu Ala Lys Val Asp Glu Asn His Ser Glu Phe Thr
Leu2240 2245 2250Tyr Glu Ser Arg Leu Leu
Asp Ile Ser Val Ser Pro Leu Met Asn2255 2260
2265Ser Leu Val Ser Gln Val Ile Cys Asp Val Leu Phe Leu Ile
Gly2270 2275 2280Gln Ser228510288PRTHomo
sapiens 102Met Ala Thr Gln Ala Arg Gln Glu Thr Cys Asp Asn Thr Lys Trp
Asn1 5 10 15Ser His Tyr
Ala Arg Ser Cys Asp His His Gln Tyr His Pro Gln Arg 20
25 30Ser Tyr Lys Ala Lys Ala His Lys Gly Ala
Pro Gly Gly Arg Trp Cys 35 40
45Val Gln Gly Val Gly Trp His Val Cys Val Gly Ala His Cys His Gly 50
55 60Ala Ser Ile Ser Lys Asn Ser Ser Arg
Glu Val Cys Ala Glu Ile Leu65 70 75
80Ala Cys Ile Pro Lys Ala His Ala
8510369PRTHomo sapiens 103Met Pro Tyr Asp Ser Val Arg Ile Glu Arg Arg Met
Arg Cys Phe Lys1 5 10
15Ser Lys Ser Gln Leu Leu Asp Ser Gln Val Phe Lys Tyr Gly His Thr
20 25 30Pro Tyr Leu Val Leu Asp Tyr
Met Gly Tyr Glu Gln Gly Ile Glu Thr 35 40
45Asp Lys Ile Val Phe Thr Asp Thr Val Tyr Arg Phe Phe Phe Pro
Phe 50 55 60Met Gln Leu Phe
Ser6510465PRTHomo sapiens 104Met Cys Phe Asn Phe Lys Met Leu Asn Ser Phe
Gln Thr Trp Tyr Leu1 5 10
15Ile Tyr Ser Pro Phe Leu Ala Phe Val Glu Phe Gln Ala Glu Cys Leu
20 25 30Thr Asp Cys Pro Arg Thr Arg
Leu Ser Phe Asn Leu Lys Gln Leu Arg 35 40
45Lys Gly Gln Arg Arg Tyr Lys Gly Lys Ala Ala Gln Asn Arg Ser
Gly 50 55 60Glu6510561PRTHomo
sapiens 105Met Leu Gly Ala Val Ile Thr Thr Asn Ile Thr Pro Arg Gly Val
Ile1 5 10 15Lys Pro Arg
Arg Thr Arg Gly Pro Leu Val Ala Gly Gly Val Cys Arg 20
25 30Gly Leu Gly Gly Thr Ser Val Leu Val Pro
Thr Val Thr Val Gln Ala 35 40
45Ser Ala Arg Thr Gln Ala Gly Lys Ser Val Leu Lys Tyr 50
55 6010658PRTHomo sapiens 106Met Cys Asn Phe Phe Lys
Tyr Val Phe Tyr Ser Tyr Gly Leu Leu Val1 5
10 15Ser Glu Pro Asp Leu Leu Thr Ile Phe Leu Tyr Asn
Asn Ala Ser His 20 25 30Phe
Leu Asp Ser Leu Val Met Cys Cys Met Gln Glu Leu Ser Ser Ser 35
40 45Ser Glu Gly Gly Leu Pro Leu Gln Ala
Ser 50 5510755PRTHomo sapiens 107Met Leu Lys Lys Lys
Asn Phe Phe Leu Val Glu Met Gln Ser Pro Val1 5
10 15Lys Arg Tyr Glu Lys Ala Ser Leu Ser Gln Arg
Pro Gly Arg Gln Ser 20 25
30Thr Thr Arg Gly Ser Glu Val Leu Met Glu Ser Cys Leu Ser Asn Glu
35 40 45Val Leu Lys Arg Met Pro Lys
50 5510850PRTHomo sapiens 108Met Leu Gln Ile Arg Lys Leu
Leu Leu Gly Thr Cys Asp Thr His Ser1 5 10
15Glu Cys Asp Met Val Ala Asn Gly Trp Pro Val Leu Lys
Ala Gly Ser 20 25 30Gln His
Lys Gly Gln Arg Ala Leu Ala Ala Pro Leu Pro Thr Ser Glu 35
40 45Pro Gly 5010949PRTHomo sapiens 109Met
Arg His His Leu Phe Tyr Lys Leu Asp Tyr Gly Phe Lys Trp Asn1
5 10 15Thr Gln Gly Asn Ile Tyr Lys
His Gln Gly Lys Leu Ser Thr Ala Ser 20 25
30Leu Phe His Leu Glu Arg Gly Arg Phe Pro Asn Gln Thr Gly
Phe Asp 35 40 45Pro 11048PRTHomo
sapiens 110Met Pro Val His Ser Ser Leu Gly Asn Lys Ser Glu Thr Pro Cys
Gln1 5 10 15Lys Lys Lys
Lys Lys Met Leu Leu Ile Leu Ser Glu Ser Lys Lys Glu 20
25 30Thr Leu Thr Ala Leu Asn Ser Gly Phe Ile
Phe Leu Ala Val Phe Gly 35 40
4511148PRTHomo sapiens 111Met Arg Ser Trp Asp Leu Leu Phe Ser Pro Gly Leu
Gln Asn Leu Ile1 5 10
15Pro Val Thr Lys Ala Arg Lys Glu Leu Tyr His Lys Pro Ser Leu Ser
20 25 30Trp His Glu Asn Trp Leu Pro
Gly Ser Val Tyr Pro Ile Asn Cys Glu 35 40
4511245PRTHomo sapiens 112Met Ile Gly His Glu Ala Ser Cys His
Thr Pro Glu Ile Arg Val Arg1 5 10
15Leu Leu Leu Arg Thr Met Cys Leu Val Thr Tyr Phe Ser Lys Ile
Ile 20 25 30Ser Leu Pro Gly
Asn Gln Ser Ser Leu Val Tyr Leu Ser 35 40
4511345PRTHomo sapiens 113Met Phe Ile Ile Phe Ile Phe Lys Val
Cys Val Ile Phe Leu Ser Met1 5 10
15Tyr Ser Ile His Met Val Cys Leu Ser Val Ser Gln Thr Cys Leu
Leu 20 25 30Tyr Ser Phe Ile
Ile Met Leu Ala Thr Ser Trp Ile Leu 35 40
4511444PRTHomo sapiens 114Met Arg Thr Gly Cys Gln Ala Gln Cys
Thr Pro Leu Thr Val Asn Glu1 5 10
15Ser Glu Leu Gly Phe Leu Tyr Cys Phe Leu Cys Asn Met Ile Ala
Glu 20 25 30Thr His Phe Lys
Asn Ser Glu Ala Cys His Ser Cys 35 4011541PRTHomo
sapiens 115Val Met Ala Tyr Tyr Ser Gly Gln Val Cys Pro Ala Gln Gly Val
Ile1 5 10 15Ser Gly Gly
Phe Gln Thr Cys Thr Gln Phe Lys Asp Gly Gly Asp Arg 20
25 30Leu Cys Leu Tyr Leu Val Asn Pro Thr
35 4011639PRTHomo sapiens 116Met Ile Ser Ala His Cys Asp
Leu Arg Leu Leu Gly Ser Ser Asp Ser1 5 10
15Pro Ala Ser Ala Ser Arg Val Ala Gly Ile Thr Gly Met
Arg His His 20 25 30Ala Arg
Leu Ile Leu Tyr Phe 3511739PRTHomo sapiens 117Met Glu Asp Phe Phe
Leu Thr Ala Leu Phe Phe Met Ala Phe Ser Lys1 5
10 15Arg Phe Lys Cys Ser Leu Phe Phe Lys Trp Gly
Ser Leu Gly Arg Gly 20 25
30Lys Val Cys Pro His His Leu 3511839PRTHomo sapiens 118Met Leu
Glu Ala Leu Trp Asn Ser Pro Ile Pro Pro Pro Phe Tyr Ile1 5
10 15Ser Leu Pro Thr Leu Ala Pro Met
Leu Leu Val Pro Leu Gln Cys Ile 20 25
30Pro Thr Gln Gly Ser Ile Pro 3511934PRTHomo sapiens
119Met Tyr Ser Thr Lys Met Glu Pro Tyr Ala Trp Ala Leu Gly Ile Gln1
5 10 15Ala Ser Ile Ser Ala Gln
Thr Ser Leu Leu Glu Phe Leu Leu Met Leu 20 25
30Ala Pro 12033PRTHomo sapiens 120Met Val Ser Ser Pro
Gln Gly Gly Glu Ala Thr His Thr Met Leu Lys1 5
10 15Ile Asn Thr Lys Asn Lys His Lys Val Arg Leu
Val Leu His Met Cys 20 25
30Asp12153PRTHomo sapiens 121Asn Asp Ile Phe Leu Val Ala Val His Glu Leu
Gly His Ala Leu Gly1 5 10
15Leu Glu His Ser Ser Asp Pro Ser Ala Ile Met Ala Pro Phe Tyr Gln
20 25 30Trp Met Asp Thr Glu Asn Phe
Val Leu Pro Asp Asp Asp Arg Arg Gly 35 40
45Ile Gln Gln Leu Tyr 50122260PRTHomo sapiens 122Met Ser Pro
Ala Pro Arg Pro Ser Arg Cys Leu Leu Leu Pro Leu Leu1 5
10 15Thr Leu Gly Thr Ala Leu Ala Ser Leu
Gly Ser Ala Gln Ser Ser Ser 20 25
30Phe Ser Pro Glu Ala Trp Leu Gln Gln Tyr Gly Tyr Leu Pro Pro Gly
35 40 45Asp Leu Arg Thr His Thr Gln
Arg Ser Pro Gln Ser Leu Ser Ala Ala 50 55
60Ile Ala Ala Met Gln Lys Phe Tyr Gly Leu Gln Val Thr Gly Lys Ala65
70 75 80Asp Ala Asp Thr
Met Lys Ala Met Arg Arg Pro Arg Cys Gly Val Pro85 90
95Asp Lys Phe Gly Ala Glu Ile Lys Ala Asn Val Arg Arg Lys
Arg Tyr100 105 110Ala Ile Gln Gly Leu Lys
Trp Gln His Asn Glu Ile Thr Phe Cys Ile115 120
125Gln Asn Tyr Thr Pro Lys Val Gly Glu Tyr Ala Thr Tyr Glu Ala
Ile130 135 140Arg Lys Ala Phe Arg Val Trp
Glu Ser Ala Thr Pro Leu Arg Phe Arg145 150
155 160Glu Val Pro Tyr Ala Tyr Ile Arg Glu Gly His Glu
Lys Gln Ala Asp165 170 175Ile Met Ile Phe
Phe Ala Glu Gly Phe His Gly Asp Ser Thr Pro Phe180 185
190Asp Gly Glu Gly Gly Phe Leu Ala His Ala Tyr Phe Pro Gly
Pro Asn195 200 205Ile Gly Gly Asp Thr His
Phe Asp Ser Ala Glu Pro Trp Thr Val Arg210 215
220Asn Glu Asp Leu Asn Gly Asn Asp Ile Phe Leu Val Ala Val His
Glu225 230 235 240Leu Gly
His Ala Leu Gly Leu Glu His Ser Ser Asp Pro Ser Ala Ile245
250 255Met Ala Pro Gly26012353PRTHomo sapiens 123Asn Asp
Ile Phe Leu Val Ala Val His Glu Leu Gly His Ala Leu Gly1 5
10 15Leu Glu His Ser Ser Asp Pro Ser
Ala Ile Met Ala Pro Phe Tyr Gln 20 25
30Trp Met Asp Thr Glu Asn Phe Val Leu Pro Asn Asp Asp Arg Arg
Gly 35 40 45Ile Gln Gln Leu Tyr
501242273PRTHomo sapiens 124Met Gly Phe Val Arg Gln Ile Gln Leu Leu Leu
Trp Lys Asn Trp Thr1 5 10
15Leu Arg Lys Arg Gln Lys Ile Arg Phe Val Val Glu Leu Val Trp Pro
20 25 30Leu Ser Leu Phe Leu Val Leu
Ile Trp Leu Arg Asn Ala Asn Pro Leu 35 40
45Tyr Ser His His Glu Cys His Phe Pro Asn Lys Ala Met Pro Ser
Ala 50 55 60Gly Met Leu Pro Trp Leu
Gln Gly Ile Phe Cys Asn Val Asn Asn Pro65 70
75 80Cys Phe Gln Ser Pro Thr Pro Gly Glu Ser Pro
Gly Ile Val Ser Asn85 90 95Tyr Asn Asn
Ser Ile Leu Ala Arg Val Tyr Arg Asp Phe Gln Glu Leu100
105 110Leu Met Asn Ala Pro Glu Ser Gln His Leu Gly Arg
Ile Trp Thr Glu115 120 125Leu His Ile Leu
Ser Gln Phe Met Asp Thr Leu Arg Thr His Pro Glu130 135
140Arg Ile Ala Gly Arg Gly Ile Arg Ile Arg Asp Ile Leu Lys
Asp Glu145 150 155 160Glu
Thr Leu Thr Leu Phe Leu Ile Lys Asn Ile Gly Leu Ser Asp Ser165
170 175Val Val Tyr Leu Leu Ile Asn Ser Gln Val Arg
Pro Glu Gln Phe Ala180 185 190His Gly Val
Pro Asp Leu Ala Leu Lys Asp Ile Ala Cys Ser Glu Ala195
200 205Leu Leu Glu Arg Phe Ile Ile Phe Ser Gln Arg Arg
Gly Ala Lys Thr210 215 220Val Arg Tyr Ala
Leu Cys Ser Leu Ser Gln Gly Thr Leu Gln Trp Ile225 230
235 240Glu Asp Thr Leu Tyr Ala Asn Val Asp
Phe Phe Lys Leu Phe Arg Val245 250 255Leu
Pro Thr Leu Leu Asp Ser Arg Ser Gln Gly Ile Asn Leu Arg Ser260
265 270Trp Gly Gly Ile Leu Ser Asp Met Ser Pro Arg
Ile Gln Glu Phe Ile275 280 285His Arg Pro
Ser Met Gln Asp Leu Leu Trp Val Thr Arg Pro Leu Met290
295 300Gln Asn Gly Gly Pro Glu Thr Phe Thr Lys Leu Met
Gly Ile Leu Ser305 310 315
320Asp Leu Leu Cys Gly Tyr Pro Glu Gly Gly Gly Ser Arg Val Leu Ser325
330 335Phe Asn Trp Tyr Glu Asp Asn Asn Tyr
Lys Ala Phe Leu Gly Ile Asp340 345 350Ser
Thr Arg Lys Asp Pro Ile Tyr Ser Tyr Asp Arg Arg Thr Thr Ser355
360 365Phe Cys Asn Ala Leu Ile Gln Ser Leu Glu Ser
Asn Pro Leu Thr Lys370 375 380Ile Ala Trp
Arg Ala Ala Lys Pro Leu Leu Met Gly Lys Ile Leu Tyr385
390 395 400Thr Pro Asp Ser Pro Ala Ala
Arg Arg Ile Leu Lys Asn Ala Asn Ser405 410
415Thr Phe Glu Glu Leu Glu His Val Arg Lys Leu Val Lys Ala Trp Glu420
425 430Glu Val Gly Pro Gln Ile Trp Tyr Phe
Phe Asp Asn Ser Thr Gln Met435 440 445Asn
Met Ile Arg Asp Thr Leu Gly Asn Pro Thr Val Lys Asp Phe Leu450
455 460Asn Arg Gln Leu Gly Glu Glu Gly Ile Thr Ala
Glu Ala Ile Leu Asn465 470 475
480Phe Leu Tyr Lys Gly Pro Arg Glu Ser Gln Ala Asp Asp Met Ala
Asn485 490 495Phe Asp Trp Arg Asp Ile Phe
Asn Ile Thr Asp Arg Thr Leu Arg Leu500 505
510Val Asn Gln Tyr Leu Glu Cys Leu Val Leu Asp Lys Phe Glu Ser Tyr515
520 525Asn Asp Glu Thr Gln Leu Thr Gln Arg
Ala Leu Ser Leu Leu Glu Glu530 535 540Asn
Met Phe Trp Ala Gly Val Val Phe Pro Asp Met Tyr Pro Trp Thr545
550 555 560Ser Ser Leu Pro Pro His
Val Lys Tyr Lys Ile Arg Met Asp Ile Asp565 570
575Val Val Glu Lys Thr Asn Lys Ile Lys Asp Arg Tyr Trp Asp Ser
Gly580 585 590Pro Arg Ala Asp Pro Val Glu
Asp Phe Arg Tyr Ile Trp Gly Gly Phe595 600
605Ala Tyr Leu Gln Asp Met Val Glu Gln Gly Ile Thr Arg Ser Gln Val610
615 620Gln Ala Glu Ala Pro Val Gly Ile Tyr
Leu Gln Gln Met Pro Tyr Pro625 630 635
640Cys Phe Val Asp Asp Ser Phe Met Ile Ile Leu Asn Arg Cys
Phe Pro645 650 655Ile Phe Met Val Leu Ala
Trp Ile Tyr Ser Val Ser Met Thr Val Lys660 665
670Ser Ile Val Leu Glu Lys Glu Leu Arg Leu Lys Glu Thr Leu Lys
Asn675 680 685Gln Gly Val Ser Asn Ala Val
Ile Trp Cys Thr Trp Phe Leu Asp Ser690 695
700Phe Ser Ile Met Ser Met Ser Ile Phe Leu Leu Thr Ile Phe Ile Met705
710 715 720His Gly Arg Ile
Leu His Tyr Ser Asp Pro Phe Ile Leu Phe Leu Phe725 730
735Leu Leu Ala Phe Ser Thr Ala Thr Ile Met Leu Cys Phe Leu
Leu Ser740 745 750Thr Phe Phe Ser Lys Ala
Ser Leu Ala Ala Ala Cys Ser Gly Val Ile755 760
765Tyr Phe Thr Leu Tyr Leu Pro His Ile Leu Cys Phe Ala Trp Gln
Asp770 775 780Arg Met Thr Ala Glu Leu Lys
Lys Ala Val Ser Leu Leu Ser Pro Val785 790
795 800Ala Phe Gly Phe Gly Thr Glu Tyr Leu Val Arg Phe
Glu Glu Gln Gly805 810 815Leu Gly Leu Gln
Trp Ser Asn Ile Gly Asn Ser Pro Thr Glu Gly Asp820 825
830Glu Phe Ser Phe Leu Leu Ser Met Gln Met Met Leu Leu Asp
Ala Ala835 840 845Cys Tyr Gly Leu Leu Ala
Trp Tyr Leu Asp Gln Val Phe Pro Gly Asp850 855
860Tyr Gly Thr Pro Leu Pro Trp Tyr Phe Leu Leu Gln Glu Ser Tyr
Trp865 870 875 880Leu Ser
Gly Glu Gly Cys Ser Thr Arg Glu Glu Arg Ala Leu Glu Lys885
890 895Thr Glu Pro Leu Thr Glu Glu Thr Glu Asp Pro Glu
His Pro Glu Gly900 905 910Ile His Asp Ser
Phe Phe Glu Arg Glu His Pro Gly Trp Val Pro Gly915 920
925Val Cys Val Lys Asn Leu Val Lys Ile Phe Glu Pro Cys Gly
Arg Pro930 935 940Ala Val Asp Arg Leu Asn
Ile Thr Phe Tyr Glu Asn Gln Ile Thr Ala945 950
955 960Phe Leu Gly His Asn Gly Ala Gly Lys Thr Thr
Thr Leu Ser Ile Leu965 970 975Thr Gly Leu
Leu Pro Pro Thr Ser Gly Thr Val Leu Val Gly Gly Arg980
985 990Asp Ile Glu Thr Ser Leu Asp Ala Val Arg Gln Ser
Leu Gly Met Cys995 1000 1005Pro Gln His
Asn Ile Leu Phe His His Leu Thr Val Ala Glu His1010
1015 1020Met Leu Phe Tyr Ala Gln Leu Lys Gly Lys Ser
Gln Glu Glu Ala1025 1030 1035Gln Leu
Glu Met Glu Ala Met Leu Glu Asp Thr Gly Leu His His1040
1045 1050Lys Arg Asn Glu Glu Ala Gln Asp Leu Ser Gly
Gly Met Gln Arg1055 1060 1065Lys Leu
Ser Val Ala Ile Ala Phe Val Gly Asp Ala Lys Val Val1070
1075 1080Ile Leu Asp Glu Pro Thr Ser Gly Val Asp Pro
Tyr Ser Arg Arg1085 1090 1095Ser Ile
Trp Asp Leu Leu Leu Lys Tyr Arg Ser Gly Arg Thr Ile1100
1105 1110Ile Met Pro Thr His His Met Asp Glu Ala Asp
His Gln Gly Asp1115 1120 1125Arg Ile
Ala Ile Ile Ala Gln Gly Arg Leu Tyr Cys Ser Gly Thr1130
1135 1140Pro Leu Phe Leu Lys Asn Cys Phe Gly Thr Gly
Leu Tyr Leu Thr1145 1150 1155Leu Val
Arg Lys Met Lys Asn Ile Gln Ser Gln Arg Lys Gly Ser1160
1165 1170Glu Gly Thr Cys Ser Cys Ser Ser Lys Gly Phe
Ser Thr Thr Cys1175 1180 1185Pro Ala
His Val Asp Asp Leu Thr Pro Glu Gln Val Leu Asp Gly1190
1195 1200Asp Val Asn Glu Leu Met Asp Val Val Leu His
His Val Pro Glu1205 1210 1215Ala Lys
Leu Val Glu Cys Ile Gly Gln Glu Leu Ile Phe Leu Leu1220
1225 1230Pro Asn Lys Asn Phe Lys His Arg Ala Tyr Ala
Ser Leu Phe Arg1235 1240 1245Glu Leu
Glu Glu Thr Leu Ala Asp Leu Gly Leu Ser Ser Phe Gly1250
1255 1260Ile Ser Asp Thr Pro Leu Glu Glu Ile Phe Leu
Lys Val Thr Glu1265 1270 1275Asp Ser
Asp Ser Gly Pro Leu Phe Ala Gly Gly Ala Gln Gln Lys1280
1285 1290Arg Glu Asn Val Asn Pro Arg His Pro Cys Leu
Gly Pro Arg Glu1295 1300 1305Lys Ala
Gly Gln Thr Pro Gln Asp Ser Asn Val Cys Ser Pro Gly1310
1315 1320Ala Pro Ala Ala His Pro Glu Gly Gln Pro Pro
Pro Glu Pro Glu1325 1330 1335Cys Pro
Gly Pro Gln Leu Asn Thr Gly Thr Gln Leu Val Leu Gln1340
1345 1350His Val Gln Ala Leu Leu Val Lys Arg Phe Gln
His Thr Ile Arg1355 1360 1365Ser His
Lys Asp Phe Leu Ala Gln Ile Val Leu Pro Ala Thr Phe1370
1375 1380Val Phe Leu Ala Leu Met Leu Ser Ile Val Ile
Leu Pro Phe Gly1385 1390 1395Glu Tyr
Pro Ala Leu Thr Leu His Pro Trp Ile Tyr Gly Gln Gln1400
1405 1410Tyr Thr Phe Phe Ser Met Asp Glu Pro Gly Ser
Glu Gln Phe Thr1415 1420 1425Val Leu
Ala Asp Val Leu Leu Asn Lys Pro Gly Phe Gly Asn Arg1430
1435 1440Cys Leu Lys Glu Gly Trp Leu Pro Glu Tyr Pro
Cys Gly Asn Ser1445 1450 1455Thr Pro
Trp Lys Thr Pro Ser Val Ser Pro Asn Ile Thr Gln Leu1460
1465 1470Phe Gln Lys Gln Lys Trp Thr Gln Val Asn Pro
Ser Pro Ser Cys1475 1480 1485Arg Cys
Ser Thr Arg Glu Lys Leu Thr Met Leu Pro Glu Cys Pro1490
1495 1500Glu Gly Ala Gly Gly Leu Pro Pro Pro Gln Arg
Thr Gln Arg Ser1505 1510 1515Thr Glu
Ile Leu Gln Asp Leu Thr Asp Arg Asn Ile Ser Asp Phe1520
1525 1530Leu Val Lys Thr Tyr Pro Ala Leu Ile Arg Ser
Ser Leu Lys Ser1535 1540 1545Lys Phe
Trp Val Asn Glu Gln Arg Tyr Gly Gly Ile Ser Ile Gly1550
1555 1560Gly Lys Leu Pro Val Val Pro Ile Thr Gly Glu
Ala Leu Val Gly1565 1570 1575Phe Leu
Ser Asp Leu Gly Arg Ile Met Asn Val Ser Gly Gly Pro1580
1585 1590Ile Thr Arg Glu Ala Ser Lys Glu Ile Pro Asp
Phe Leu Lys His1595 1600 1605Leu Glu
Thr Glu Asp Asn Ile Lys Val Trp Phe Asn Asn Lys Gly1610
1615 1620Trp His Ala Leu Val Ser Phe Leu Asn Val Ala
His Asn Ala Ile1625 1630 1635Leu Arg
Ala Ser Leu Pro Lys Asp Arg Ser Pro Glu Glu Tyr Gly1640
1645 1650Ile Thr Val Ile Ser Gln Pro Leu Asn Leu Thr
Lys Glu Gln Leu1655 1660 1665Ser Glu
Ile Thr Val Leu Thr Thr Ser Val Asp Ala Val Val Ala1670
1675 1680Ile Cys Val Ile Phe Ser Met Ser Phe Val Pro
Ala Ser Phe Val1685 1690 1695Leu Tyr
Leu Ile Gln Glu Arg Val Asn Lys Ser Lys His Leu Gln1700
1705 1710Phe Ile Ser Gly Val Ser Pro Thr Thr Tyr Trp
Val Thr Asn Phe1715 1720 1725Leu Trp
Asp Ile Met Asn Tyr Ser Val Ser Ala Gly Leu Val Val1730
1735 1740Gly Ile Phe Ile Gly Phe Gln Lys Lys Ala Tyr
Thr Ser Pro Glu1745 1750 1755Asn Leu
Pro Ala Leu Val Ala Leu Leu Leu Leu Tyr Gly Trp Ala1760
1765 1770Val Ile Pro Met Met Tyr Pro Ala Ser Phe Leu
Phe Asp Val Pro1775 1780 1785Ser Thr
Ala Tyr Val Ala Leu Ser Cys Ala Asn Leu Phe Ile Gly1790
1795 1800Ile Asn Ser Ser Ala Ile Thr Phe Ile Leu Glu
Leu Phe Asp Asn1805 1810 1815Asn Arg
Thr Leu Leu Arg Phe Asn Ala Val Leu Arg Lys Leu Leu1820
1825 1830Ile Val Phe Pro His Phe Cys Leu Gly Arg Gly
Leu Ile Asp Leu1835 1840 1845Ala Leu
Ser Gln Ala Val Thr Asp Val Tyr Ala Arg Phe Gly Glu1850
1855 1860Glu His Ser Ala Asn Pro Phe His Trp Asp Leu
Ile Gly Lys Asn1865 1870 1875Leu Phe
Ala Met Val Val Glu Gly Val Val Tyr Phe Leu Leu Thr1880
1885 1890Leu Leu Val Gln Arg His Phe Phe Leu Ser Gln
Trp Ile Ala Glu1895 1900 1905Pro Thr
Lys Glu Pro Ile Val Asp Glu Asp Asp Asp Val Ala Glu1910
1915 1920Glu Arg Gln Arg Ile Ile Thr Gly Gly Asn Lys
Thr Asp Ile Leu1925 1930 1935Arg Leu
His Glu Leu Thr Lys Ile Tyr Leu Gly Thr Ser Ser Pro1940
1945 1950Ala Val Asp Arg Leu Cys Val Gly Val Arg Pro
Gly Glu Cys Phe1955 1960 1965Gly Leu
Leu Gly Val Asn Gly Ala Gly Lys Thr Thr Thr Phe Lys1970
1975 1980Met Leu Thr Gly Asp Thr Thr Val Thr Ser Gly
Asp Ala Thr Val1985 1990 1995Ala Gly
Lys Ser Ile Leu Thr Asn Ile Ser Glu Val His Gln Asn2000
2005 2010Met Gly Tyr Cys Pro Gln Phe Asp Ala Ile Asp
Glu Leu Leu Thr2015 2020 2025Gly Arg
Glu His Leu Tyr Leu Tyr Ala Arg Leu Arg Gly Val Pro2030
2035 2040Ala Glu Glu Ile Glu Lys Val Ala Asn Trp Ser
Ile Lys Ser Leu2045 2050 2055Gly Leu
Thr Val Tyr Ala Asp Cys Leu Ala Gly Thr Tyr Ser Gly2060
2065 2070Gly Asn Lys Arg Lys Leu Ser Thr Ala Ile Ala
Leu Ile Gly Cys2075 2080 2085Pro Pro
Leu Val Leu Leu Asp Glu Pro Thr Thr Gly Met Asp Pro2090
2095 2100Gln Ala Arg Arg Met Leu Trp Asn Val Ile Val
Ser Ile Ile Arg2105 2110 2115Lys Gly
Arg Ala Val Val Leu Thr Ser His Ser Met Glu Glu Cys2120
2125 2130Glu Ala Leu Cys Thr Arg Leu Ala Ile Met Val
Lys Gly Ala Phe2135 2140 2145Arg Cys
Met Gly Thr Ile Gln His Leu Lys Ser Lys Phe Gly Asp2150
2155 2160Gly Tyr Ile Val Thr Met Lys Ile Lys Ser Pro
Lys Asp Asp Leu2165 2170 2175Leu Pro
Asp Leu Asn Pro Val Glu Gln Phe Phe Gln Gly Asn Phe2180
2185 2190Pro Gly Ser Val Gln Arg Glu Arg His Tyr Asn
Met Leu Gln Phe2195 2200 2205Gln Val
Ser Ser Ser Ser Leu Ala Arg Ile Phe Gln Leu Leu Leu2210
2215 2220Ser His Lys Asp Ser Leu Leu Ile Glu Glu Tyr
Ser Val Thr Gln2225 2230 2235Thr Thr
Leu Asp Gln Val Phe Val Asn Phe Ala Lys Gln Gln Thr2240
2245 2250Glu Ser His Asp Leu Pro Leu His Pro Arg Ala
Ala Gly Ala Ser2255 2260 2265Arg Gln
Ala Gln Asp2270125317PRTHomo sapiens 125Met Lys Val Leu Trp Ala Ala Leu
Leu Val Thr Phe Leu Ala Gly Cys1 5 10
15Gln Ala Lys Val Glu Gln Ala Val Glu Thr Glu Pro Glu Pro
Glu Leu 20 25 30Arg Gln Gln
Thr Glu Trp Gln Ser Gly Gln Arg Trp Glu Leu Ala Leu 35
40 45Gly Arg Phe Trp Asp Tyr Leu Arg Trp Val Gln
Thr Leu Ser Glu Gln 50 55 60Val Gln
Glu Glu Leu Leu Ser Ser Gln Val Thr Gln Glu Leu Arg Ala65
70 75 80Leu Met Asp Glu Thr Met Lys
Glu Leu Lys Ala Tyr Lys Ser Glu Leu85 90
95Glu Glu Gln Leu Thr Pro Val Ala Glu Glu Thr Arg Ala Arg Leu Ser100
105 110Lys Glu Leu Gln Ala Ala Gln Ala Arg
Leu Gly Ala Asp Met Glu Asp115 120 125Val
Cys Gly Arg Leu Val Gln Tyr Arg Gly Glu Val Gln Ala Met Leu130
135 140Gly Gln Ser Thr Glu Glu Leu Arg Val Arg Leu
Ala Ser His Leu Arg145 150 155
160Lys Leu Arg Lys Arg Leu Leu Arg Asp Ala Asp Asp Leu Gln Lys
Arg165 170 175Leu Ala Val Tyr Gln Ala Gly
Ala Arg Glu Gly Ala Glu Arg Gly Leu180 185
190Ser Ala Ile Arg Glu Arg Leu Gly Pro Leu Val Glu Gln Gly Arg Val195
200 205Arg Ala Ala Thr Val Gly Ser Leu Ala
Gly Gln Pro Leu Gln Glu Arg210 215 220Ala
Gln Ala Trp Gly Glu Arg Leu Arg Ala Arg Met Glu Glu Met Gly225
230 235 240Ser Arg Thr Arg Asp Arg
Leu Asp Glu Val Lys Glu Gln Val Ala Glu245 250
255Val Arg Ala Lys Leu Glu Glu Gln Ala Gln Gln Ile Arg Leu Gln
Ala260 265 270Glu Ala Phe Gln Ala Arg Leu
Lys Ser Trp Phe Glu Pro Leu Val Glu275 280
285Asp Met Gln Arg Gln Trp Ala Gly Leu Val Glu Lys Val Gln Ala Ala290
295 300Val Gly Thr Ser Ala Ala Pro Val Pro
Ser Asp Asn His305 310 315126374PRTHomo
sapiens 126Met Leu Ser Thr Ser Arg Ser Arg Phe Ile Arg Asn Thr Asn Glu
Ser1 5 10 15Gly Glu Glu
Val Thr Thr Phe Phe Asp Tyr Asp Tyr Gly Ala Pro Cys 20
25 30His Lys Phe Asp Val Lys Gln Ile Gly Ala
Gln Leu Leu Pro Pro Leu 35 40
45Tyr Ser Leu Val Phe Ile Phe Gly Phe Val Gly Asn Met Leu Val Val 50
55 60Leu Ile Leu Ile Asn Cys Lys Lys Leu
Lys Cys Leu Thr Asp Ile Tyr65 70 75
80Leu Leu Asn Leu Ala Ile Ser Asp Leu Leu Phe Leu Ile Thr
Leu Pro85 90 95Leu Trp Ala His Ser Ala
Ala Asn Glu Trp Val Phe Gly Asn Ala Met100 105
110Cys Lys Leu Phe Thr Gly Leu Tyr His Ile Gly Tyr Phe Gly Gly
Ile115 120 125Phe Phe Ile Ile Leu Leu Thr
Ile Asp Arg Tyr Leu Ala Ile Val His130 135
140Ala Val Phe Ala Leu Lys Ala Arg Thr Val Thr Phe Gly Val Val Thr145
150 155 160Ser Val Ile Thr
Trp Leu Val Ala Val Phe Ala Ser Val Pro Gly Ile165 170
175Ile Phe Thr Lys Cys Gln Lys Glu Asp Ser Val Tyr Val Cys
Gly Pro180 185 190Tyr Phe Pro Arg Gly Trp
Asn Asn Phe His Thr Ile Met Arg Asn Ile195 200
205Leu Gly Leu Val Leu Pro Leu Leu Ile Met Val Ile Cys Tyr Ser
Gly210 215 220Ile Leu Lys Thr Leu Leu Arg
Cys Arg Asn Glu Lys Lys Arg His Arg225 230
235 240Ala Val Arg Val Ile Phe Thr Ile Met Ile Val Tyr
Phe Leu Phe Trp245 250 255Thr Pro Tyr Asn
Ile Val Ile Leu Leu Asn Thr Phe Gln Glu Phe Phe260 265
270Gly Leu Ser Asn Cys Glu Ser Thr Ser Gln Leu Asp Gln Ala
Thr Gln275 280 285Val Thr Glu Thr Leu Gly
Met Thr His Cys Cys Ile Asn Pro Ile Ile290 295
300Tyr Ala Phe Val Gly Glu Lys Phe Arg Ser Leu Phe His Ile Ala
Leu305 310 315 320Gly Cys
Arg Ile Ala Pro Leu Gln Lys Pro Val Cys Gly Gly Pro Gly325
330 335Val Arg Pro Gly Lys Asn Val Lys Val Thr Thr Gln
Gly Leu Leu Asp340 345 350Gly Arg Gly Lys
Gly Lys Ser Ile Gly Arg Ala Pro Glu Ala Ser Leu355 360
365Gln Asp Lys Glu Gly Ala370127146PRTHomo sapiens 127Met
Ala Gly Pro Leu Arg Ala Pro Leu Leu Leu Leu Ala Ile Leu Ala1
5 10 15Val Ala Leu Ala Val Ser Pro
Ala Ala Gly Ser Ser Pro Gly Lys Pro 20 25
30Pro Arg Leu Val Gly Gly Pro Met Asp Ala Ser Val Glu Glu
Glu Gly 35 40 45Val Arg Arg Ala
Leu Asp Phe Ala Val Gly Glu Tyr Asn Lys Ala Ser 50 55
60Asn Asp Met Tyr His Ser Arg Ala Leu Gln Val Val Arg
Ala Arg Lys65 70 75
80Gln Ile Val Ala Gly Val Asn Tyr Phe Leu Asp Val Glu Leu Gly Arg85
90 95Thr Thr Cys Thr Lys Thr Gln Pro Asn Leu
Asp Asn Cys Pro Phe His100 105 110Asp Gln
Pro His Leu Lys Arg Lys Ala Phe Cys Ser Phe Gln Ile Tyr115
120 125Ala Val Pro Trp Gln Gly Thr Met Thr Leu Ser Lys
Ser Thr Cys Gln130 135 140Asp
Ala1451285622PRTHomo sapiens 128Met Ile Ser Trp Glu Val Val His Thr Val
Phe Leu Phe Ala Leu Leu1 5 10
15Tyr Ser Ser Leu Ala Gln Asp Ala Ser Pro Gln Ser Glu Ile Arg Ala
20 25 30Glu Glu Ile Pro Glu Gly
Ala Ser Thr Leu Ala Phe Val Phe Asp Val 35 40
45Thr Gly Ser Met Tyr Asp Asp Leu Val Gln Val Ile Glu Gly
Ala Ser 50 55 60Lys Ile Leu Glu Thr
Ser Leu Lys Arg Pro Lys Arg Pro Leu Phe Asn65 70
75 80Phe Ala Leu Val Pro Phe His Asp Pro Glu
Ile Gly Pro Val Thr Ile85 90 95Thr Thr
Asp Pro Lys Lys Phe Gln Tyr Glu Leu Arg Glu Leu Tyr Val100
105 110Gln Gly Gly Gly Asp Cys Pro Glu Met Ser Ile Gly
Ala Ile Lys Ile115 120 125Ala Leu Glu Ile
Ser Leu Pro Gly Ser Phe Ile Tyr Val Phe Thr Asp130 135
140Ala Arg Ser Lys Asp Tyr Arg Leu Thr His Glu Val Leu Gln
Leu Ile145 150 155 160Gln
Gln Lys Gln Ser Gln Val Val Phe Val Leu Thr Gly Asp Cys Asp165
170 175Asp Arg Thr His Ile Gly Tyr Lys Val Tyr Glu
Glu Ile Ala Ser Thr180 185 190Ser Ser Gly
Gln Val Phe His Leu Asp Lys Lys Gln Val Asn Glu Val195
200 205Leu Lys Trp Val Glu Glu Ala Val Gln Ala Ser Lys
Val His Leu Leu210 215 220Ser Thr Asp His
Leu Glu Gln Ala Val Asn Thr Trp Arg Ile Pro Phe225 230
235 240Asp Pro Ser Leu Lys Glu Val Thr Val
Ser Leu Ser Gly Pro Ser Pro245 250 255Met
Ile Glu Ile Arg Asn Pro Leu Gly Lys Leu Ile Lys Lys Gly Phe260
265 270Gly Leu His Glu Leu Leu Asn Ile His Asn Ser
Ala Lys Val Val Asn275 280 285Val Lys Glu
Pro Glu Ala Gly Met Trp Thr Val Lys Thr Ser Ser Ser290
295 300Gly Arg His Ser Val Arg Ile Thr Gly Leu Ser Thr
Ile Asp Phe Arg305 310 315
320Ala Gly Phe Ser Arg Lys Pro Thr Leu Asp Phe Lys Lys Thr Val Ser325
330 335Arg Pro Val Gln Gly Ile Pro Thr Tyr
Val Leu Leu Asn Thr Ser Gly340 345 350Ile
Ser Thr Pro Ala Arg Ile Asp Leu Leu Glu Leu Leu Ser Ile Ser355
360 365Gly Ser Ser Leu Lys Thr Ile Pro Val Lys Tyr
Tyr Pro His Arg Lys370 375 380Pro Tyr Gly
Ile Trp Asn Ile Ser Asp Phe Val Pro Pro Asn Glu Ala385
390 395 400Phe Phe Leu Lys Val Thr Gly
Tyr Asp Lys Asp Asp Tyr Leu Phe Gln405 410
415Arg Val Ser Ser Val Ser Phe Ser Ser Ile Val Pro Asp Ala Pro Lys420
425 430Val Thr Met Pro Glu Lys Thr Pro Gly
Tyr Tyr Leu Gln Pro Gly Gln435 440 445Ile
Pro Cys Ser Val Asp Ser Leu Leu Pro Phe Thr Leu Ser Phe Val450
455 460Arg Asn Gly Val Thr Leu Gly Val Asp Gln Tyr
Leu Lys Glu Ser Ala465 470 475
480Ser Val Asn Leu Asp Ile Ala Lys Val Thr Leu Ser Asp Glu Gly
Phe485 490 495Tyr Glu Cys Ile Ala Val Ser
Ser Ala Gly Thr Gly Arg Ala Gln Thr500 505
510Phe Phe Asp Val Ser Glu Pro Pro Pro Val Ile Gln Val Pro Asn Asn515
520 525Val Thr Val Thr Pro Gly Glu Arg Ala
Val Leu Thr Cys Leu Ile Ile530 535 540Ser
Ala Val Asp Tyr Asn Leu Thr Trp Gln Arg Asn Asp Arg Asp Val545
550 555 560Arg Leu Ala Glu Pro Ala
Arg Ile Arg Thr Leu Ala Asn Leu Ser Leu565 570
575Glu Leu Lys Ser Val Lys Phe Asn Asp Ala Gly Glu Tyr His Cys
Met580 585 590Val Ser Ser Glu Gly Gly Ser
Ser Ala Ala Ser Val Phe Leu Thr Val595 600
605Gln Glu Pro Pro Lys Val Thr Val Met Pro Lys Asn Gln Ser Phe Thr610
615 620Gly Gly Ser Glu Val Ser Ile Met Cys
Ser Ala Thr Gly Tyr Pro Lys625 630 635
640Pro Lys Ile Ala Trp Thr Val Asn Asp Met Phe Ile Val Gly
Ser His645 650 655Arg Tyr Arg Met Thr Ser
Asp Gly Thr Leu Phe Ile Lys Asn Ala Ala660 665
670Pro Lys Asp Ala Gly Ile Tyr Gly Cys Leu Ala Ser Asn Ser Ala
Gly675 680 685Thr Asp Lys Gln Asn Ser Thr
Leu Arg Tyr Ile Glu Ala Pro Lys Leu690 695
700Met Val Val Gln Ser Glu Leu Leu Val Ala Leu Gly Asp Ile Thr Val705
710 715 720Met Glu Cys Lys
Thr Ser Gly Ile Pro Pro Pro Gln Val Lys Trp Phe725 730
735Lys Gly Asp Leu Glu Leu Arg Pro Ser Thr Phe Leu Ile Ile
Asp Pro740 745 750Leu Leu Gly Leu Leu Lys
Ile Gln Glu Thr Gln Asp Leu Asp Ala Gly755 760
765Asp Tyr Thr Cys Val Ala Ile Asn Glu Ala Gly Arg Ala Thr Gly
Lys770 775 780Ile Thr Leu Asp Val Gly Ser
Pro Pro Val Phe Ile Gln Glu Pro Ala785 790
795 800Asp Val Ser Met Glu Ile Gly Ser Asn Val Thr Leu
Pro Cys Tyr Val805 810 815Gln Gly Tyr Pro
Glu Pro Thr Ile Lys Trp Arg Arg Leu Asp Asn Met820 825
830Pro Ile Phe Ser Arg Pro Phe Ser Val Ser Ser Ile Ser Gln
Leu Arg835 840 845Thr Gly Ala Leu Phe Ile
Leu Asn Leu Trp Ala Ser Asp Lys Gly Thr850 855
860Tyr Ile Cys Glu Ala Glu Asn Gln Phe Gly Lys Ile Gln Ser Glu
Thr865 870 875 880Thr Val
Thr Val Thr Gly Leu Val Ala Pro Leu Ile Gly Ile Ser Pro885
890 895Ser Val Ala Asn Val Ile Glu Gly Gln Gln Leu Thr
Leu Pro Cys Thr900 905 910Leu Leu Ala Gly
Asn Pro Ile Pro Glu Arg Arg Trp Ile Lys Asn Ser915 920
925Ala Met Leu Leu Gln Asn Pro Tyr Ile Thr Val Arg Ser Asp
Gly Ser930 935 940Leu His Ile Glu Arg Val
Gln Leu Gln Asp Gly Gly Glu Tyr Thr Cys945 950
955 960Val Ala Ser Asn Val Ala Gly Thr Asn Asn Lys
Thr Thr Ser Val Val965 970 975Val His Val
Leu Pro Thr Ile Gln His Gly Gln Gln Ile Leu Ser Thr980
985 990Ile Glu Gly Ile Pro Val Thr Leu Pro Cys Lys Ala
Ser Gly Asn Pro995 1000 1005Lys Pro Ser
Val Ile Trp Ser Lys Lys Gly Glu Leu Ile Ser Thr1010
1015 1020Ser Ser Ala Lys Phe Ser Ala Gly Ala Asp Gly
Ser Leu Tyr Val1025 1030 1035Val Ser
Pro Gly Gly Glu Glu Ser Gly Glu Tyr Val Cys Thr Ala1040
1045 1050Thr Asn Thr Ala Gly Tyr Ala Lys Arg Lys Val
Gln Leu Thr Val1055 1060 1065Tyr Val
Arg Pro Arg Val Phe Gly Asp Gln Arg Gly Leu Ser Gln1070
1075 1080Asp Lys Pro Val Glu Ile Ser Val Leu Ala Gly
Glu Glu Val Thr1085 1090 1095Leu Pro
Cys Glu Val Lys Ser Leu Pro Pro Pro Ile Ile Thr Trp1100
1105 1110Ala Lys Glu Thr Gln Leu Ile Ser Pro Phe Ser
Pro Arg His Thr1115 1120 1125Phe Leu
Pro Ser Gly Ser Met Lys Ile Thr Glu Thr Arg Thr Ser1130
1135 1140Asp Ser Gly Met Tyr Leu Cys Val Ala Thr Asn
Ile Ala Gly Asn1145 1150 1155Val Thr
Gln Ala Val Lys Leu Asn Val His Val Pro Pro Lys Ile1160
1165 1170Gln Arg Gly Pro Lys His Leu Lys Val Gln Val
Gly Gln Arg Val1175 1180 1185Asp Ile
Pro Cys Asn Ala Gln Gly Thr Pro Leu Pro Val Ile Thr1190
1195 1200Trp Ser Lys Gly Gly Ser Thr Met Leu Val Asp
Gly Glu His His1205 1210 1215Val Ser
Asn Pro Asp Gly Thr Leu Ser Ile Asp Gln Ala Thr Pro1220
1225 1230Ser Asp Ala Gly Ile Tyr Thr Cys Val Ala Thr
Asn Ile Ala Gly1235 1240 1245Thr Asp
Glu Thr Glu Ile Thr Leu His Val Gln Glu Pro Pro Thr1250
1255 1260Val Glu Asp Leu Glu Pro Pro Tyr Asn Thr Thr
Phe Gln Glu Arg1265 1270 1275Val Ala
Asn Gln Arg Ile Glu Phe Pro Cys Pro Ala Lys Gly Thr1280
1285 1290Pro Lys Pro Thr Ile Lys Trp Leu His Asn Gly
Arg Glu Leu Thr1295 1300 1305Gly Arg
Glu Pro Gly Ile Ser Ile Leu Glu Asp Gly Thr Leu Leu1310
1315 1320Val Ile Ala Ser Val Thr Pro Tyr Asp Asn Gly
Glu Tyr Ile Cys1325 1330 1335Val Ala
Val Asn Glu Ala Gly Thr Thr Glu Arg Lys Tyr Asn Leu1340
1345 1350Lys Val His Val Pro Pro Val Ile Lys Asp Lys
Glu Gln Val Thr1355 1360 1365Asn Val
Ser Val Leu Leu Asn Gln Leu Thr Asn Leu Phe Cys Glu1370
1375 1380Val Glu Gly Thr Pro Ser Pro Ile Ile Met Trp
Tyr Lys Asp Asn1385 1390 1395Val Gln
Val Thr Glu Ser Ser Thr Ile Gln Thr Val Asn Asn Gly1400
1405 1410Lys Ile Leu Lys Leu Phe Arg Ala Thr Pro Glu
Asp Ala Gly Arg1415 1420 1425Tyr Ser
Cys Lys Ala Ile Asn Ile Ala Gly Thr Ser Gln Lys Tyr1430
1435 1440Phe Asn Ile Asp Val Leu Val Pro Pro Thr Ile
Ile Gly Thr Asn1445 1450 1455Phe Pro
Asn Glu Val Ser Val Val Leu Asn Arg Asp Val Ala Leu1460
1465 1470Glu Cys Gln Val Lys Gly Thr Pro Phe Pro Asp
Ile His Trp Phe1475 1480 1485Lys Asp
Gly Lys Pro Leu Phe Leu Gly Asp Pro Asn Val Glu Leu1490
1495 1500Leu Asp Arg Gly Gln Val Leu His Leu Lys Asn
Ala Arg Arg Asn1505 1510 1515Asp Lys
Gly Arg Tyr Gln Cys Thr Val Ser Asn Ala Ala Gly Lys1520
1525 1530Gln Ala Lys Asp Ile Lys Leu Thr Ile Tyr Ile
Pro Pro Ser Ile1535 1540 1545Lys Gly
Gly Asn Val Thr Thr Asp Ile Ser Val Leu Ile Asn Ser1550
1555 1560Leu Ile Lys Leu Glu Cys Glu Thr Arg Gly Leu
Pro Met Pro Ala1565 1570 1575Ile Thr
Trp Tyr Lys Asp Gly Gln Pro Ile Met Ser Ser Ser Gln1580
1585 1590Ala Leu Tyr Ile Asp Lys Gly Gln Tyr Leu His
Ile Pro Arg Ala1595 1600 1605Gln Val
Ser Asp Ser Ala Thr Tyr Thr Cys His Val Ala Asn Val1610
1615 1620Ala Gly Thr Ala Glu Lys Ser Phe His Val Asp
Val Tyr Val Pro1625 1630 1635Pro Met
Ile Glu Gly Asn Leu Ala Thr Pro Leu Asn Lys Gln Val1640
1645 1650Val Ile Ala His Ser Leu Thr Leu Glu Cys Lys
Ala Ala Gly Asn1655 1660 1665Pro Ser
Pro Ile Leu Thr Trp Leu Lys Asp Gly Val Pro Val Lys1670
1675 1680Ala Asn Asp Asn Ile Arg Ile Glu Ala Gly Gly
Lys Lys Leu Glu1685 1690 1695Ile Met
Ser Ala Gln Glu Ile Asp Arg Gly Gln Tyr Ile Cys Val1700
1705 1710Ala Thr Ser Val Ala Gly Glu Lys Glu Ile Lys
Tyr Glu Val Asp1715 1720 1725Val Leu
Val Pro Pro Ala Ile Glu Gly Gly Asp Glu Thr Ser Tyr1730
1735 1740Phe Ile Val Met Val Asn Asn Leu Leu Glu Leu
Asp Cys His Val1745 1750 1755Thr Gly
Ser Pro Pro Pro Thr Ile Met Trp Leu Lys Asp Gly Gln1760
1765 1770Leu Ile Asp Glu Arg Asp Gly Phe Lys Ile Leu
Leu Asn Gly Arg1775 1780 1785Lys Leu
Val Ile Ala Gln Ala Gln Val Ser Asn Thr Gly Leu Tyr1790
1795 1800Arg Cys Met Ala Ala Asn Thr Ala Gly Asp His
Lys Lys Glu Phe1805 1810 1815Glu Val
Thr Val His Val Pro Pro Thr Ile Lys Ser Ser Gly Leu1820
1825 1830Ser Glu Arg Val Val Val Lys Tyr Lys Pro Val
Ala Leu Gln Cys1835 1840 1845Ile Ala
Asn Gly Ile Pro Asn Pro Ser Ile Thr Trp Leu Lys Asp1850
1855 1860Asp Gln Pro Val Asn Thr Ala Gln Gly Asn Leu
Lys Ile Gln Ser1865 1870 1875Ser Gly
Arg Val Leu Gln Ile Ala Lys Thr Leu Leu Glu Asp Ala1880
1885 1890Gly Arg Tyr Thr Cys Val Ala Thr Asn Ala Ala
Gly Glu Thr Gln1895 1900 1905Gln His
Ile Gln Leu His Val His Glu Pro Pro Ser Leu Glu Asp1910
1915 1920Ala Gly Lys Met Leu Asn Glu Thr Val Leu Val
Ser Asn Pro Val1925 1930 1935Gln Leu
Glu Cys Lys Ala Ala Gly Asn Pro Val Pro Val Ile Thr1940
1945 1950Trp Tyr Lys Asp Asn Arg Leu Leu Ser Gly Ser
Thr Ser Met Thr1955 1960 1965Phe Leu
Asn Arg Gly Gln Ile Ile Asp Ile Glu Ser Ala Gln Ile1970
1975 1980Ser Asp Ala Gly Ile Tyr Lys Cys Val Ala Ile
Asn Ser Ala Gly1985 1990 1995Ala Thr
Glu Leu Phe Tyr Ser Leu Gln Val His Val Ala Pro Ser2000
2005 2010Ile Ser Gly Ser Asn Asn Met Val Ala Val Val
Val Asn Asn Pro2015 2020 2025Val Arg
Leu Glu Cys Glu Ala Arg Gly Ile Pro Ala Pro Ser Leu2030
2035 2040Thr Trp Leu Lys Asp Gly Ser Pro Val Ser Ser
Phe Ser Asn Gly2045 2050 2055Leu Gln
Val Leu Ser Gly Gly Arg Ile Leu Ala Leu Thr Ser Ala2060
2065 2070Gln Ile Ser Asp Thr Gly Arg Tyr Thr Cys Val
Ala Val Asn Ala2075 2080 2085Ala Gly
Glu Lys Gln Arg Asp Ile Asp Leu Arg Val Tyr Val Pro2090
2095 2100Pro Asn Ile Met Gly Glu Glu Gln Asn Val Ser
Val Leu Ile Ser2105 2110 2115Gln Ala
Val Glu Leu Leu Cys Gln Ser Asp Ala Ile Pro Pro Pro2120
2125 2130Thr Leu Thr Trp Leu Lys Asp Gly His Pro Leu
Leu Lys Lys Pro2135 2140 2145Gly Leu
Ser Ile Ser Glu Asn Arg Ser Val Leu Lys Ile Glu Asp2150
2155 2160Ala Gln Val Gln Asp Thr Gly Arg Tyr Thr Cys
Glu Ala Thr Asn2165 2170 2175Val Ala
Gly Lys Thr Glu Lys Lys Asn Tyr Asn Val Asn Ile Trp2180
2185 2190Val Pro Pro Asn Ile Gly Gly Ser Asp Glu Leu
Thr Gln Leu Thr2195 2200 2205Val Ile
Glu Gly Asn Leu Ile Ser Leu Leu Cys Glu Ser Ser Gly2210
2215 2220Ile Pro Pro Pro Asn Leu Ile Trp Lys Lys Lys
Gly Ser Pro Val2225 2230 2235Leu Thr
Asp Ser Met Gly Arg Val Arg Ile Ile Ala Glu Lys Ser2240
2245 2250Asp Ala Ala Leu Tyr Ser Cys Val Ala Ser Asn
Val Ala Gly Thr2255 2260 2265Ala Lys
Lys Glu Tyr Asn Leu Gln Val Tyr Ile Arg Pro Thr Ile2270
2275 2280Thr Asn Ser Gly Ser His Pro Thr Glu Ile Ile
Val Thr Arg Gly2285 2290 2295Lys Ser
Ile Ser Leu Glu Cys Glu Val Gln Gly Ile Pro Pro Pro2300
2305 2310Thr Val Thr Trp Met Lys Asp Gly His Pro Leu
Ile Lys Ala Lys2315 2320 2325Gly Val
Glu Ile Leu Asp Glu Gly His Ile Leu Gln Leu Lys Asn2330
2335 2340Ile His Val Ser Asp Thr Gly Arg Tyr Val Cys
Val Ala Val Asn2345 2350 2355Val Ala
Gly Met Thr Asp Lys Lys Tyr Asp Leu Ser Val His Ala2360
2365 2370Pro Pro Ser Ile Ile Gly Asn His Arg Ser Pro
Glu Asn Ile Ser2375 2380 2385Val Val
Glu Lys Asn Ser Val Ser Leu Thr Cys Glu Ala Ser Gly2390
2395 2400Ile Pro Leu Pro Ser Ile Thr Trp Phe Lys Asp
Gly Trp Pro Val2405 2410 2415Ser Leu
Ser Asn Ser Val Arg Ile Leu Ser Gly Gly Arg Met Leu2420
2425 2430Arg Leu Met Gln Thr Thr Met Glu Asp Ala Gly
Gln Tyr Thr Cys2435 2440 2445Val Val
Arg Asn Ala Ala Gly Glu Glu Arg Lys Ile Phe Gly Leu2450
2455 2460Ser Val Leu Val Pro Pro His Ile Val Gly Glu
Asn Thr Leu Glu2465 2470 2475Asp Val
Lys Val Lys Glu Lys Gln Ser Val Thr Leu Thr Cys Glu2480
2485 2490Val Thr Gly Asn Pro Val Pro Glu Ile Thr Trp
His Lys Asp Gly2495 2500 2505Gln Pro
Leu Gln Glu Asp Glu Ala His His Ile Ile Ser Gly Gly2510
2515 2520Arg Phe Leu Gln Ile Thr Asn Val Gln Val Pro
His Thr Gly Arg2525 2530 2535Tyr Thr
Cys Leu Ala Ser Ser Pro Ala Gly His Lys Ser Arg Ser2540
2545 2550Phe Ser Leu Asn Val Phe Val Ser Pro Thr Ile
Ala Gly Val Gly2555 2560 2565Ser Asp
Gly Asn Pro Glu Asp Val Thr Val Ile Leu Asn Ser Pro2570
2575 2580Thr Ser Leu Val Cys Glu Ala Tyr Ser Tyr Pro
Pro Ala Thr Ile2585 2590 2595Thr Trp
Phe Lys Asp Gly Thr Pro Leu Glu Ser Asn Arg Asn Ile2600
2605 2610Arg Ile Leu Pro Gly Gly Arg Thr Leu Gln Ile
Leu Asn Ala Gln2615 2620 2625Glu Asp
Asn Ala Gly Arg Tyr Ser Cys Val Ala Thr Asn Glu Ala2630
2635 2640Gly Glu Met Ile Lys His Tyr Glu Val Lys Val
Tyr Ile Pro Pro2645 2650 2655Ile Ile
Asn Lys Gly Asp Leu Trp Gly Pro Gly Leu Ser Pro Lys2660
2665 2670Glu Val Lys Ile Lys Val Asn Asn Thr Leu Thr
Leu Glu Cys Glu2675 2680 2685Ala Tyr
Ala Ile Pro Ser Ala Ser Leu Ser Trp Tyr Lys Asp Gly2690
2695 2700Gln Pro Leu Lys Ser Asp Asp His Val Asn Ile
Ala Ala Asn Gly2705 2710 2715His Thr
Leu Gln Ile Lys Glu Ala Gln Ile Ser Asp Thr Gly Arg2720
2725 2730Tyr Thr Cys Val Ala Ser Asn Ile Ala Gly Glu
Asp Glu Leu Asp2735 2740 2745Phe Asp
Val Asn Ile Gln Val Pro Pro Ser Phe Gln Lys Leu Trp2750
2755 2760Glu Ile Gly Asn Met Leu Asp Thr Gly Arg Asn
Gly Glu Ala Lys2765 2770 2775Asp Val
Ile Ile Asn Asn Pro Ile Ser Leu Tyr Cys Glu Thr Asn2780
2785 2790Ala Ala Pro Pro Pro Thr Leu Thr Trp Tyr Lys
Asp Gly His Pro2795 2800 2805Leu Thr
Ser Ser Asp Lys Val Leu Ile Leu Pro Gly Gly Arg Val2810
2815 2820Leu Gln Ile Pro Arg Ala Lys Val Glu Asp Ala
Gly Arg Tyr Thr2825 2830 2835Cys Val
Ala Val Asn Glu Ala Gly Glu Asp Ser Leu Gln Tyr Asp2840
2845 2850Val Arg Val Leu Val Pro Pro Ile Ile Lys Gly
Ala Asn Ser Asp2855 2860 2865Leu Pro
Glu Glu Val Thr Val Leu Val Asn Lys Ser Ala Leu Ile2870
2875 2880Glu Cys Leu Ser Ser Gly Ser Pro Ala Pro Arg
Asn Ser Trp Gln2885 2890 2895Lys Asp
Gly Gln Pro Leu Leu Glu Asp Asp His His Lys Phe Leu2900
2905 2910Ser Asn Gly Arg Ile Leu Gln Ile Leu Asn Thr
Gln Ile Thr Asp2915 2920 2925Ile Gly
Arg Tyr Val Cys Val Ala Glu Asn Thr Ala Gly Ser Ala2930
2935 2940Lys Lys Tyr Phe Asn Leu Asn Val His Val Pro
Pro Ser Val Ile2945 2950 2955Gly Pro
Lys Ser Glu Asn Leu Thr Val Val Val Asn Asn Phe Ile2960
2965 2970Ser Leu Thr Cys Glu Val Ser Gly Phe Pro Pro
Pro Asp Leu Ser2975 2980 2985Trp Leu
Lys Asn Lys Leu Asn Thr Asn Thr Leu Ile Val Pro Gly2990
2995 3000Gly Arg Thr Leu Gln Ile Ile Arg Ala Lys Val
Ser Asp Gly Gly3005 3010 3015Glu Tyr
Thr Cys Ile Ala Ile Asn Gln Ala Gly Glu Ser Lys Lys3020
3025 3030Lys Phe Ser Leu Thr Val Tyr Val Pro Pro Ser
Ile Lys Asp His3035 3040 3045Asp Ser
Glu Ser Leu Ser Val Val Asn Val Arg Glu Gly Thr Ser3050
3055 3060Val Ser Leu Glu Cys Glu Ser Asn Ala Val Pro
Pro Pro Val Ile3065 3070 3075Thr Trp
Tyr Lys Asn Gly Arg Met Ile Thr Glu Ser Thr His Val3080
3085 3090Glu Ile Leu Ala Asp Gly Gln Met Leu His Ile
Lys Lys Ala Glu3095 3100 3105Val Ser
Asp Thr Gly Gln Tyr Val Cys Arg Ala Ile Asn Val Ala3110
3115 3120Gly Arg Asp Asp Lys Asn Phe His Leu Asn Val
Tyr Val Pro Pro3125 3130 3135Ser Ile
Glu Gly Pro Glu Arg Glu Val Ile Val Glu Thr Ile Ser3140
3145 3150Asn Pro Val Thr Leu Thr Cys Asp Ala Thr Gly
Ile Pro Pro Pro3155 3160 3165Thr Ile
Ala Trp Leu Lys Asn His Lys Arg Ile Glu Asn Ser Asp3170
3175 3180Ser Leu Glu Val Arg Ile Leu Ser Gly Gly Ser
Lys Leu Gln Ile3185 3190 3195Ala Arg
Ser Gln His Ser Asp Ser Gly Asn Tyr Thr Cys Ile Ala3200
3205 3210Ser Asn Met Glu Gly Lys Ala Gln Lys Tyr Tyr
Phe Leu Ser Ile3215 3220 3225Gln Val
Pro Pro Ser Val Ala Gly Ala Glu Ile Pro Ser Asp Val3230
3235 3240Ser Val Leu Leu Gly Glu Asn Val Glu Leu Val
Cys Asn Ala Asn3245 3250 3255Gly Ile
Pro Thr Pro Leu Ile Gln Trp Leu Lys Asp Gly Lys Pro3260
3265 3270Ile Ala Ser Gly Glu Thr Glu Arg Ile Arg Val
Ser Ala Asn Gly3275 3280 3285Ser Thr
Leu Asn Ile Tyr Gly Ala Leu Thr Ser Asp Thr Gly Lys3290
3295 3300Tyr Thr Cys Val Ala Thr Asn Pro Ala Gly Glu
Glu Asp Arg Ile3305 3310 3315Phe Asn
Leu Asn Val Tyr Val Thr Pro Thr Ile Arg Gly Asn Lys3320
3325 3330Asp Glu Ala Glu Lys Leu Met Thr Leu Val Asp
Thr Ser Ile Asn3335 3340 3345Ile Glu
Cys Arg Ala Thr Gly Thr Pro Pro Pro Gln Ile Asn Trp3350
3355 3360Leu Lys Asn Gly Leu Pro Leu Pro Leu Ser Ser
His Ile Arg Leu3365 3370 3375Leu Ala
Ala Gly Gln Val Ile Arg Ile Val Arg Ala Gln Val Ser3380
3385 3390Asp Val Ala Val Tyr Thr Cys Val Ala Ser Asn
Arg Ala Gly Val3395 3400 3405Asp Asn
Lys His Tyr Asn Leu Gln Val Phe Ala Pro Pro Asn Met3410
3415 3420Asp Asn Ser Met Gly Thr Glu Glu Ile Thr Val
Leu Lys Gly Ser3425 3430 3435Ser Thr
Ser Met Ala Cys Ile Thr Asp Gly Thr Pro Ala Pro Ser3440
3445 3450Met Ala Trp Leu Arg Asp Gly Gln Pro Leu Gly
Leu Asp Ala His3455 3460 3465Leu Thr
Val Ser Thr His Gly Met Val Leu Gln Leu Leu Lys Ala3470
3475 3480Glu Thr Glu Asp Ser Gly Lys Tyr Thr Cys Ile
Ala Ser Asn Glu3485 3490 3495Ala Gly
Glu Val Ser Lys His Phe Ile Leu Lys Val Leu Glu Pro3500
3505 3510Pro His Ile Asn Gly Ser Glu Glu His Glu Glu
Ile Ser Val Ile3515 3520 3525Val Asn
Asn Pro Leu Glu Leu Thr Cys Ile Ala Ser Gly Ile Pro3530
3535 3540Ala Pro Lys Met Thr Trp Met Lys Asp Gly Arg
Pro Leu Pro Gln3545 3550 3555Thr Asp
Gln Val Gln Thr Leu Gly Gly Gly Glu Val Leu Arg Ile3560
3565 3570Ser Thr Ala Gln Val Glu Asp Thr Gly Arg Tyr
Thr Cys Leu Ala3575 3580 3585Ser Ser
Pro Ala Gly Asp Asp Asp Lys Glu Tyr Leu Val Arg Val3590
3595 3600His Val Pro Pro Asn Ile Ala Gly Thr Asp Glu
Pro Arg Asp Ile3605 3610 3615Thr Val
Leu Arg Asn Arg Gln Val Thr Leu Glu Cys Lys Ser Asp3620
3625 3630Ala Val Pro Pro Pro Val Ile Thr Trp Leu Arg
Asn Gly Glu Arg3635 3640 3645Leu Gln
Ala Thr Pro Arg Val Arg Ile Leu Ser Gly Gly Arg Tyr3650
3655 3660Leu Gln Ile Asn Asn Ala Asp Leu Gly Asp Thr
Ala Asn Tyr Thr3665 3670 3675Cys Val
Ala Ser Asn Ile Ala Gly Lys Thr Thr Arg Glu Phe Ile3680
3685 3690Leu Thr Val Asn Val Pro Pro Asn Ile Lys Gly
Gly Pro Gln Ser3695 3700 3705Leu Val
Ile Leu Leu Asn Lys Ser Thr Val Leu Glu Cys Ile Ala3710
3715 3720Glu Gly Val Pro Thr Pro Arg Ile Thr Trp Arg
Lys Asp Gly Ala3725 3730 3735Val Leu
Ala Gly Asn His Ala Arg Tyr Ser Ile Leu Glu Asn Gly3740
3745 3750Phe Leu His Ile Gln Ser Ala His Val Thr Asp
Thr Gly Arg Tyr3755 3760 3765Leu Cys
Met Ala Thr Asn Ala Ala Gly Thr Asp Arg Arg Arg Ile3770
3775 3780Asp Leu Gln Val His Val Pro Pro Ser Ile Ala
Pro Gly Pro Thr3785 3790 3795Asn Met
Thr Val Ile Val Asn Val Gln Thr Thr Leu Ala Cys Glu3800
3805 3810Ala Thr Gly Ile Pro Lys Pro Ser Ile Asn Trp
Arg Lys Asn Gly3815 3820 3825His Leu
Leu Asn Val Asp Gln Asn Gln Asn Ser Tyr Arg Leu Leu3830
3835 3840Ser Ser Gly Ser Leu Val Ile Ile Ser Pro Ser
Val Asp Asp Thr3845 3850 3855Ala Thr
Tyr Glu Cys Thr Val Thr Asn Gly Ala Gly Asp Asp Lys3860
3865 3870Arg Thr Val Asp Leu Thr Val Gln Val Pro Pro
Ser Ile Ala Asp3875 3880 3885Glu Pro
Thr Asp Phe Leu Val Thr Lys His Ala Pro Ala Val Ile3890
3895 3900Thr Cys Thr Ala Ser Gly Val Pro Phe Pro Ser
Ile His Trp Thr3905 3910 3915Lys Asn
Gly Ile Arg Leu Leu Pro Arg Gly Asp Gly Tyr Arg Ile3920
3925 3930Leu Ser Ser Gly Ala Ile Glu Ile Leu Ala Thr
Gln Leu Asn His3935 3940 3945Ala Gly
Arg Tyr Thr Cys Val Ala Arg Asn Ala Ala Gly Ser Ala3950
3955 3960His Arg His Val Thr Leu His Val His Glu Pro
Pro Val Ile Gln3965 3970 3975Pro Gln
Pro Ser Glu Leu His Val Ile Leu Asn Asn Pro Ile Leu3980
3985 3990Leu Pro Cys Glu Ala Thr Gly Thr Pro Ser Pro
Phe Ile Thr Trp3995 4000 4005Gln Lys
Glu Gly Ile Asn Val Asn Thr Ser Gly Arg Asn His Ala4010
4015 4020Val Leu Pro Ser Gly Gly Leu Gln Ile Ser Arg
Ala Val Arg Glu4025 4030 4035Asp Ala
Gly Thr Tyr Met Cys Val Ala Gln Asn Pro Ala Gly Thr4040
4045 4050Ala Leu Gly Lys Ile Lys Leu Asn Val Gln Val
Pro Pro Val Ile4055 4060 4065Ser Pro
His Leu Lys Glu Tyr Val Ile Ala Val Asp Lys Pro Ile4070
4075 4080Thr Leu Ser Cys Glu Ala Asp Gly Leu Pro Pro
Pro Asp Ile Thr4085 4090 4095Trp His
Lys Asp Gly Arg Ala Ile Val Glu Ser Ile Arg Gln Arg4100
4105 4110Val Leu Ser Ser Gly Ser Leu Gln Ile Ala Phe
Val Gln Pro Gly4115 4120 4125Asp Ala
Gly His Tyr Thr Cys Met Ala Ala Asn Val Ala Gly Ser4130
4135 4140Ser Ser Thr Ser Thr Lys Leu Thr Val His Val
Pro Pro Arg Ile4145 4150 4155Arg Ser
Thr Glu Gly His Tyr Thr Val Asn Glu Asn Ser Gln Ala4160
4165 4170Ile Leu Pro Cys Val Ala Asp Gly Ile Pro Thr
Pro Ala Ile Asn4175 4180 4185Trp Lys
Lys Asp Asn Val Leu Leu Ala Asn Leu Leu Gly Lys Tyr4190
4195 4200Thr Ala Glu Pro Tyr Gly Glu Leu Ile Leu Glu
Asn Val Val Leu4205 4210 4215Glu Asp
Ser Gly Phe Tyr Thr Cys Val Ala Asn Asn Ala Ala Gly4220
4225 4230Glu Asp Thr His Thr Val Ser Leu Thr Val His
Val Leu Pro Thr4235 4240 4245Phe Thr
Glu Leu Pro Gly Asp Val Ser Leu Asn Lys Gly Glu Gln4250
4255 4260Leu Arg Leu Ser Cys Lys Ala Thr Gly Ile Pro
Leu Pro Lys Leu4265 4270 4275Thr Trp
Thr Phe Asn Asn Asn Ile Ile Pro Ala His Phe Asp Ser4280
4285 4290Val Asn Gly His Ser Glu Leu Val Ile Glu Arg
Val Ser Lys Glu4295 4300 4305Asp Ser
Gly Thr Tyr Val Cys Thr Ala Glu Asn Ser Val Gly Phe4310
4315 4320Val Lys Ala Ile Gly Phe Val Tyr Val Lys Glu
Pro Pro Val Phe4325 4330 4335Lys Gly
Asp Tyr Pro Ser Asn Trp Ile Glu Pro Leu Gly Gly Asn4340
4345 4350Ala Ile Leu Asn Cys Glu Val Lys Gly Asp Pro
Thr Pro Thr Ile4355 4360 4365Gln Trp
Asn Arg Lys Gly Val Asp Ile Glu Ile Ser His Arg Ile4370
4375 4380Arg Gln Leu Gly Asn Gly Ser Leu Ala Ile Tyr
Gly Thr Val Asn4385 4390 4395Glu Asp
Ala Gly Asp Tyr Thr Cys Val Ala Thr Asn Glu Ala Gly4400
4405 4410Val Val Glu Arg Ser Met Ser Leu Thr Leu Gln
Ser Pro Pro Ile4415 4420 4425Ile Thr
Leu Glu Pro Val Glu Thr Val Ile Asn Ala Gly Gly Lys4430
4435 4440Ile Ile Leu Asn Cys Gln Ala Thr Gly Glu Pro
Gln Pro Thr Ile4445 4450 4455Thr Trp
Ser Arg Gln Gly His Ser Ile Ser Trp Asp Asp Arg Val4460
4465 4470Asn Val Leu Ser Asn Asn Ser Leu Tyr Ile Ala
Asp Ala Gln Lys4475 4480 4485Glu Asp
Thr Ser Glu Phe Glu Cys Val Ala Arg Asn Leu Met Gly4490
4495 4500Ser Val Leu Val Arg Val Pro Val Ile Val Gln
Val His Gly Gly4505 4510 4515Phe Ser
Gln Trp Ser Ala Trp Arg Ala Cys Ser Val Thr Cys Gly4520
4525 4530Lys Gly Ile Gln Lys Arg Ser Arg Leu Cys Asn
Gln Pro Leu Pro4535 4540 4545Ala Asn
Gly Gly Lys Pro Cys Gln Gly Ser Asp Leu Glu Met Arg4550
4555 4560Asn Cys Gln Asn Lys Pro Cys Pro Val Asp Gly
Ser Trp Ser Glu4565 4570 4575Trp Ser
Leu Trp Glu Glu Cys Thr Arg Ser Cys Gly Arg Gly Asn4580
4585 4590Gln Thr Arg Thr Arg Thr Cys Asn Asn Pro Ser
Val Gln His Gly4595 4600 4605Gly Arg
Pro Cys Glu Gly Asn Ala Val Glu Ile Ile Met Cys Asn4610
4615 4620Ile Arg Pro Cys Pro Val His Gly Ala Trp Ser
Ala Trp Gln Pro4625 4630 4635Trp Gly
Thr Cys Ser Glu Ser Cys Gly Lys Gly Thr Gln Thr Arg4640
4645 4650Ala Arg Leu Cys Asn Asn Pro Pro Pro Ala Phe
Gly Gly Ser Tyr4655 4660 4665Cys Asp
Gly Ala Glu Thr Gln Met Gln Val Cys Asn Glu Arg Asn4670
4675 4680Cys Pro Ile His Gly Lys Trp Ala Thr Trp Ala
Ser Trp Ser Ala4685 4690 4695Cys Ser
Val Ser Cys Gly Gly Gly Ala Arg Gln Arg Thr Arg Gly4700
4705 4710Cys Ser Asp Pro Val Pro Gln Tyr Gly Gly Arg
Lys Cys Glu Gly4715 4720 4725Ser Asp
Val Gln Ser Asp Phe Cys Asn Ser Asp Pro Cys Pro Thr4730
4735 4740His Gly Asn Trp Ser Pro Trp Ser Gly Trp Gly
Thr Cys Ser Arg4745 4750 4755Thr Cys
Asn Gly Gly Gln Met Arg Arg Tyr Arg Thr Cys Asp Asn4760
4765 4770Pro Pro Pro Ser Asn Gly Gly Arg Ala Cys Gly
Gly Pro Asp Ser4775 4780 4785Gln Ile
Gln Arg Cys Asn Thr Asp Met Cys Pro Val Asp Gly Ser4790
4795 4800Trp Gly Ser Trp His Ser Trp Ser Gln Cys Ser
Ala Ser Cys Gly4805 4810 4815Gly Gly
Glu Lys Thr Arg Lys Arg Leu Cys Asp His Pro Val Pro4820
4825 4830Val Lys Gly Gly Arg Pro Cys Pro Gly Asp Thr
Thr Gln Val Thr4835 4840 4845Arg Cys
Asn Val Gln Ala Cys Pro Gly Gly Pro Gln Arg Ala Arg4850
4855 4860Gly Ser Val Ile Gly Asn Ile Asn Asp Val Glu
Phe Gly Ile Ala4865 4870 4875Phe Leu
Asn Ala Thr Ile Thr Asp Ser Pro Asn Ser Asp Thr Arg4880
4885 4890Ile Ile Arg Ala Lys Ile Thr Asn Val Pro Arg
Ser Leu Gly Ser4895 4900 4905Ala Met
Arg Lys Ile Val Ser Ile Leu Asn Pro Ile Tyr Trp Thr4910
4915 4920Thr Ala Lys Glu Ile Gly Glu Ala Val Asn Gly
Phe Thr Leu Thr4925 4930 4935Asn Ala
Val Phe Lys Arg Glu Thr Gln Val Glu Phe Ala Thr Gly4940
4945 4950Glu Ile Leu Gln Met Ser His Ile Ala Arg Gly
Leu Asp Ser Asp4955 4960 4965Gly Ser
Leu Leu Leu Asp Ile Val Val Ser Gly Tyr Val Leu Gln4970
4975 4980Leu Gln Ser Pro Ala Glu Val Thr Val Lys Asp
Tyr Thr Glu Asp4985 4990 4995Tyr Ile
Gln Thr Gly Pro Gly Gln Leu Tyr Ala Tyr Ser Thr Arg5000
5005 5010Leu Phe Thr Ile Asp Gly Ile Ser Ile Pro Tyr
Thr Trp Asn His5015 5020 5025Thr Val
Phe Tyr Asp Gln Ala Gln Gly Arg Met Pro Phe Leu Val5030
5035 5040Glu Thr Leu His Ala Ser Ser Val Glu Ser Asp
Tyr Asn Gln Ile5045 5050 5055Glu Glu
Thr Leu Gly Phe Lys Ile His Ala Ser Ile Ser Lys Gly5060
5065 5070Asp Arg Ser Asn Gln Cys Pro Ser Gly Phe Thr
Leu Asp Ser Val5075 5080 5085Gly Pro
Phe Cys Ala Asp Glu Asp Glu Cys Ala Ala Gly Asn Pro5090
5095 5100Cys Ser His Ser Cys His Asn Ala Met Gly Thr
Tyr Tyr Cys Ser5105 5110 5115Cys Pro
Lys Gly Leu Thr Ile Ala Ala Asp Gly Arg Thr Cys Gln5120
5125 5130Asp Ile Asp Glu Cys Ala Leu Gly Arg His Thr
Cys His Ala Gly5135 5140 5145Gln Asp
Cys Asp Asn Thr Ile Gly Ser Tyr Arg Cys Val Val Arg5150
5155 5160Cys Gly Ser Gly Phe Arg Arg Thr Ser Asp Gly
Leu Ser Cys Gln5165 5170 5175Asp Ile
Asn Glu Cys Gln Glu Ser Ser Pro Cys His Gln Arg Cys5180
5185 5190Phe Asn Ala Ile Gly Ser Phe His Cys Gly Cys
Glu Pro Gly Tyr5195 5200 5205Gln Leu
Lys Gly Arg Lys Cys Met Asp Val Asn Glu Cys Arg Gln5210
5215 5220Asn Val Cys Arg Pro Asp Gln His Cys Lys Asn
Thr Arg Gly Gly5225 5230 5235Tyr Lys
Cys Ile Asp Leu Cys Pro Asn Gly Met Thr Lys Ala Glu5240
5245 5250Asn Gly Thr Cys Ile Asp Ile Asp Glu Cys Lys
Asp Gly Thr His5255 5260 5265Gln Cys
Arg Tyr Asn Gln Ile Cys Glu Asn Thr Arg Gly Ser Tyr5270
5275 5280Arg Cys Val Cys Pro Arg Gly Tyr Arg Ser Gln
Gly Val Gly Arg5285 5290 5295Pro Cys
Met Asp Ile Asn Glu Cys Glu Gln Val Pro Lys Pro Cys5300
5305 5310Ala His Gln Cys Ser Asn Thr Pro Gly Ser Phe
Lys Cys Ile Cys5315 5320 5325Pro Pro
Gly Gln His Leu Leu Gly Asp Gly Lys Ser Cys Ala Gly5330
5335 5340Leu Glu Arg Leu Pro Asn Tyr Gly Thr Gln Tyr
Ser Ser Tyr Asn5345 5350 5355Leu Ala
Arg Phe Ser Pro Val Arg Asn Asn Tyr Gln Pro Gln Gln5360
5365 5370His Tyr Arg Gln Tyr Ser His Leu Tyr Ser Ser
Tyr Ser Glu Tyr5375 5380 5385Arg Asn
Ser Arg Thr Ser Leu Ser Arg Thr Arg Arg Thr Ile Arg5390
5395 5400Lys Thr Cys Pro Glu Gly Ser Glu Ala Ser His
Asp Thr Cys Val5405 5410 5415Asp Ile
Asp Glu Cys Glu Asn Thr Asp Ala Cys Gln His Glu Cys5420
5425 5430Lys Asn Thr Phe Gly Ser Tyr Gln Cys Ile Cys
Pro Pro Gly Tyr5435 5440 5445Gln Leu
Thr His Asn Gly Lys Thr Cys Gln Asp Ile Asp Glu Cys5450
5455 5460Leu Glu Gln Asn Val His Cys Gly Pro Asn Arg
Met Cys Phe Asn5465 5470 5475Met Arg
Gly Ser Tyr Gln Cys Ile Asp Thr Pro Cys Pro Pro Asn5480
5485 5490Tyr Gln Arg Asp Pro Val Ser Gly Phe Cys Leu
Lys Asn Cys Pro5495 5500 5505Pro Asn
Asp Leu Glu Cys Ala Leu Ser Pro Tyr Ala Leu Glu Tyr5510
5515 5520Lys Leu Val Ser Leu Pro Phe Gly Ile Ala Thr
Asn Gln Asp Leu5525 5530 5535Ile Arg
Leu Val Ala Tyr Thr Gln Asp Gly Val Met His Pro Arg5540
5545 5550Thr Thr Phe Leu Met Val Asp Glu Glu Gln Thr
Val Pro Phe Ala5555 5560 5565Leu Arg
Asp Glu Asn Leu Lys Gly Val Val Tyr Thr Thr Arg Pro5570
5575 5580Leu Arg Glu Ala Glu Thr Tyr Arg Met Arg Val
Arg Ala Ser Ser5585 5590 5595Tyr Ser
Ala Asn Gly Thr Ile Glu Tyr Gln Thr Thr Phe Ile Val5600
5605 5610Tyr Ile Ala Val Ser Ala Tyr Pro Tyr5615
5620129222PRTHomo sapiens 129Met Leu Ser Arg Ala Val Cys Gly Thr
Ser Arg Gln Leu Pro Pro Val1 5 10
15Leu Gly Tyr Leu Gly Ser Arg Gln Lys His Ser Leu Pro Asp Leu
Pro 20 25 30Tyr Asp Tyr Gly
Ala Leu Glu Pro His Ile Asn Ala Gln Ile Met Gln 35
40 45Leu His His Ser Lys His His Ala Ala Tyr Val Asn
Asn Leu Asn Val 50 55 60Thr Glu Glu
Lys Tyr Gln Glu Ala Leu Ala Lys Gly Asp Val Thr Ala65 70
75 80Gln Ile Ala Leu Gln Pro Ala Leu
Lys Phe Asn Gly Gly Gly His Ile85 90
95Asn His Ser Ile Phe Trp Thr Asn Leu Ser Pro Asn Gly Gly Gly Glu100
105 110Pro Lys Gly Glu Leu Leu Glu Ala Ile Lys
Leu Asp Phe Gly Ser Phe115 120 125Asp Lys
Phe Lys Glu Lys Leu Thr Ala Ala Ser Val Gly Val Gln Gly130
135 140Ser Gly Trp Gly Trp Leu Gly Phe Asn Lys Glu Arg
Gly His Leu Gln145 150 155
160Ile Ala Ala Cys Pro Asn Gln Asp Pro Leu Gln Gly Thr Thr Gly Leu165
170 175Ile Pro Leu Leu Gly Ile Asp Val Trp
Glu His Ala Tyr Tyr Leu Gln180 185 190Tyr
Lys Asn Val Arg Pro Asp Tyr Leu Lys Ala Ile Trp Asn Val Ile195
200 205Asn Trp Glu Asn Val Thr Glu Arg Tyr Met Ala
Cys Lys Lys210 215 22013099PRTHomo
sapiens 130Met Lys Val Ser Ala Ala Leu Leu Cys Leu Leu Leu Ile Ala Ala
Thr1 5 10 15Phe Ile Pro
Gln Gly Leu Ala Gln Pro Asp Ala Ile Asn Ala Pro Val 20
25 30Thr Cys Cys Tyr Asn Phe Thr Asn Arg Lys
Ile Ser Val Gln Arg Leu 35 40
45Ala Ser Tyr Arg Arg Ile Thr Ser Ser Lys Cys Pro Lys Glu Ala Val 50
55 60Ile Phe Lys Thr Ile Val Ala Lys Glu
Ile Cys Ala Asp Pro Lys Gln65 70 75
80Lys Trp Val Gln Asp Ser Met Asp His Leu Asp Lys Gln Thr
Gln Thr85 90 95Pro Lys Thr131355PRTHomo
sapiens 131Met Ala Lys Leu Ile Ala Leu Thr Leu Leu Gly Met Gly Leu Ala
Leu1 5 10 15Phe Arg Asn
His Gln Ser Ser Tyr Gln Thr Arg Leu Asn Ala Leu Arg 20
25 30Glu Val Gln Pro Val Glu Leu Pro Asn Cys
Asn Leu Val Lys Gly Ile 35 40
45Glu Thr Gly Ser Glu Asp Leu Glu Ile Leu Pro Asn Gly Leu Ala Phe 50
55 60Ile Ser Ser Gly Leu Lys Tyr Pro Gly
Ile Lys Ser Phe Asn Pro Asn65 70 75
80Ser Pro Gly Lys Ile Leu Leu Met Asp Leu Asn Glu Glu Asp
Pro Thr85 90 95Val Leu Glu Leu Gly Ile
Thr Gly Ser Lys Phe Asp Val Ser Ser Phe100 105
110Asn Pro His Gly Ile Ser Thr Phe Thr Asp Glu Asp Asn Ala Met
Tyr115 120 125Leu Leu Val Val Asn His Pro
Asp Ala Lys Ser Thr Val Glu Leu Phe130 135
140Lys Phe Gln Glu Glu Glu Lys Ser Leu Leu His Leu Lys Thr Ile Arg145
150 155 160His Lys Leu Leu
Pro Asn Leu Asn Asp Ile Val Ala Val Gly Pro Glu165 170
175His Phe Tyr Gly Thr Asn Asp His Tyr Phe Leu Asp Pro Tyr
Leu Gln180 185 190Ser Trp Glu Met Tyr Leu
Gly Leu Ala Trp Ser Tyr Val Val Tyr Tyr195 200
205Ser Pro Ser Glu Val Arg Val Val Ala Glu Gly Phe Asp Phe Ala
Asn210 215 220Gly Ile Asn Ile Ser Pro Asp
Gly Lys Tyr Val Tyr Ile Ala Glu Leu225 230
235 240Leu Ala His Lys Ile His Val Tyr Glu Lys His Ala
Asn Trp Thr Leu245 250 255Thr Pro Leu Lys
Ser Leu Asp Phe Asn Thr Leu Val Asp Asn Ile Ser260 265
270Val Asp Pro Glu Thr Gly Asp Leu Trp Val Gly Cys His Pro
Asn Gly275 280 285Met Lys Ile Phe Phe Tyr
Asp Ser Glu Asn Pro Pro Ala Ser Glu Val290 295
300Leu Arg Ile Gln Asn Ile Leu Thr Glu Glu Pro Lys Val Thr Gln
Val305 310 315 320Tyr Ala
Glu Asn Gly Thr Val Leu Gln Gly Ser Thr Val Ala Ser Val325
330 335Tyr Lys Gly Lys Leu Leu Ile Gly Thr Val Phe His
Lys Ala Leu Tyr340 345 350Cys Glu
Leu35513220DNAArtificial SequenceDescription of Artificial Sequence
Synthetic primer 132aagtgttgaa tttacccttc
2013320DNAArtificial SequenceDescription of
Artificial Sequence Synthetic primer 133tacttcaaat taacaaccac
2013419DNAArtificial
SequenceDescription of Artificial Sequence Synthetic primer
134gcctctggtg aggaagtcg
1913520DNAArtificial SequenceDescription of Artificial Sequence Synthetic
primer 135cggtagggaa tgtgctggtg
2013632DNAArtificial SequenceDescription of Artificial
Sequence Synthetic primer 136attatccagc tggcgcgcaa ggttgaagca tg
3213732DNAArtificial SequenceDescription of
Artificial Sequence Synthetic primer 137tgaagaaagc ttgcaaacac
tggtctgtgg tc 3213821DNAArtificial
SequenceDescription of Artificial Sequence Synthetic oligonucleotide
138ccucuccacg cgcaguacat t
2113921DNAArtificial SequenceDescription of Artificial Sequence Synthetic
oligonucleotide 139uguacugcgc guggagaggt t
2114021DNAArtificial SequenceDescription of
Artificial Sequence Synthetic oligonucleotide 140ccgacaucau
gaucuucuut t
2114121DNAArtificial SequenceDescription of Artificial Sequence Synthetic
oligonucleotide 141aagaagauca ugaugucggt t
21
User Contributions:
Comment about this patent or add new information about this topic:
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 | 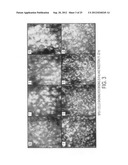 |
 |  |
 |  |
 | 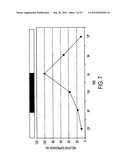 |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
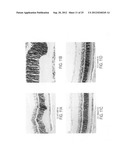 |  |
 | 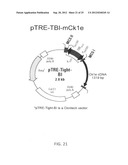 |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 | 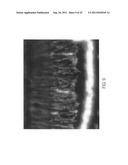 |
 |  |
 |  |
 |  |
 |  |
 |  |
| Similar patent applications: | |
| Date | Title |
|---|---|
| 2012-02-23 | Hco32 and hco27 and related examples |
| 2013-12-12 | Hox compositions and methods |
| 2010-06-10 | Method of monitoring retinopathy |
| 2012-02-23 | Estrous cycle monitoring by color response |
| 2013-09-12 | Estrous cycle monitoring by color response |
| New patent applications in this class: | |
| Date | Title |
|---|---|
| 2016-12-29 | Transgenic non-human mammal that expresses human mmp2 |
| 2016-07-07 | Humanized mice expressing the pyrin domain only protein 2 |
| 2016-07-07 | Method for creating endometriotic cells and endometriosis model animal |
| 2016-06-23 | Genetically modified rat models for pharmacokinetics |
| 2016-06-23 | Genetically modified rat models for pain |
| New patent applications from these inventors: | |
| Date | Title |
|---|---|
| 2011-12-01 | Transforming container candles and uses thereof |
| 2011-01-20 | Methods and compositions for detecting and treating retinal diseases based on metargidin (adam-15) |
| 2009-06-04 | Methods and compositions for detecting and treating retinal diseases |
| Top Inventors for class "Multicellular living organisms and unmodified parts thereof and related processes" | |
| Rank | Inventor's name |
|---|---|
| 1 | Gregory J. Holland |
| 2 | William H. Eby |
| 3 | Richard G. Stelpflug |
| 4 | Laron L. Peters |
| 5 | Justin T. Mason |