Patent application title: COX-2 function and wound healing
Inventors:
J. Patrick O'Connor (Fanwood, NJ, US)
IPC8 Class: AA61K3816FI
USPC Class:
514 186
Class name: Designated organic active ingredient containing (doai) peptide (e.g., protein, etc.) containing doai skin affecting
Publication date: 2011-03-03
Patent application number: 20110053860
Inventors list |
Agents list |
Assignees list |
List by place |
Classification tree browser |
Top 100 Inventors |
Top 100 Agents |
Top 100 Assignees |
Usenet FAQ Index |
Documents |
Other FAQs |
Patent application title: COX-2 function and wound healing
Inventors:
J. Patrick O'Connor
Agents:
Assignees:
Origin: ,
IPC8 Class: AA61K3816FI
USPC Class:
Publication date: 03/03/2011
Patent application number: 20110053860
Abstract:
The invention relates to compositions and methods for enhancing bone
healing, bone formation and wound healing. More specifically, it relates
to the use of cyclooxygenase 2 (COX-2) following bone fracture,
orthopaedic procedure or wound infliction to enhance healing.Claims:
1. A vector for use in enhancing wound healing comprising a promoter
linked to a cyclooxygenase expression cassette.
2. The vector of claim 1, wherein the promoter is constitutively induced.
3. The vector of claim 2, wherein the promoter is a cytomegalovirus promoter.
4. The vector of claim 1, wherein the cyclooxygenase expression cassette encodes a COX-2 gene product.
5. A method for enhancing wound healing comprising the step of delivering the vector of claim 1 to the location of the wound; wherein the cyclooxygenase expression cassette is expressed, thereby enhancing wound healing.
6. The method of claim 5, wherein the vector is a adenoviral vector, adeno-associated viral vector, a recombination-defective retrovirus or a plasmid.
7. The method of claim 5, wherein the vector is delivered by injection, electroporation, or inhalation of an aerosol.
8. The method of claim 5, wherein the vector is in a saline solution, encapsulated in liposomes, in a polymer solution, in a gel, or lyophilized.
9. The method of claim 5, wherein the promoter is cytomegalovirus.
10. The method of claim 5, wherein the cyclooxygenase expression cassette encodes for a COX-2 gene product.
11. The method of claim 5, wherein the wound is a bone fracture or a skin wound.
12. A method for enhancing wound healing following orthopaedic procedures comprising the step of administering the vector of claim 1 to the location of the procedure; wherein the cyclooxygenase expression cassette encodes for COX-2 gene product and wherein the cyclooxygenase expression cassette is expressed thereby enhancing wound healing.
13. A method for treating pathological heterotopic ossification conditions comprising the steps of: identifying locations of heterotopic ossification in a patient with a heterotopic ossification condition, and administering COX-2 selective NSAIDs to the patient.
14. The method of claim 13, wherein the heterotopic ossification condition is fibrodysplasia ossificans progressiva, occurs following a hip replacement procedure or after an acetabular fracture.
15. A method for inhibiting wound healing comprising the step of administering an effective amount of NSAIDs to the location of the wound, wherein the NSAIDs inhibit normal wound healing.
16. The method of claim 15, wherein the NSAIDs are COX-2 specific NSAIDs.
17. A method for treating osteoporosis, OI and brittle bone conditions comprising the step of administering the vector of claim 1, wherein expression of the vector enhances wound healing and bone formation.
18. A composition for use in wound healing comprising a formulated cyclooxygenase protein.
19. The composition of claim 18, further comprising a pharmaceutically acceptable carrier.
20. The composition of claim 18, wherein the cyclooxygenase protein is COX-1, COX-2 or a combination of the two.
21-22. (canceled)
Description:
INTRODUCTION
[0001]This application is a continuation of U.S. Ser. No. 09/953,067 filed Sep. 11, 2001, which is herein incorporated by reference in its entirety.
FIELD OF THE INVENTION
[0002]This invention relates to the field of cyclooxygenase activity and wound healing.
BACKGROUND OF THE INVENTION
[0003]Bones, along with a small number of cartilages, comprise the skeletal system that serves as the rigid supporting framework of the body in adult humans. Certain parts of the supporting framework form chambers, such as the skull and the thoracic cage, that are important for protecting the soft parts contained in the chambers. Bones also serve as attachments for muscles and act as levers in the joint system of the body.
[0004]Mature bone is comprised of an organic framework of fibrous tissue and inorganic salts known as crystalline hydroxyapatite (HA). HA is composed of calcium and phosphorous, which are derived from the blood plasma and ultimately from nutritional sources. HA represents about 60 percent of the weight of compact bone and is deposited on a fibrous structure of collagenous connective tissue. Without HA, bone loses most its weight and rigidity and is susceptible to damage.
[0005]The process of bone formation, also known as osteogenesis, involves three main steps: production of the extracellular organic matrix (osteoid); mineralization of the matrix to form bone; and bone remodeling by resorption and reformation. The cellular activities of osteoblasts, osteocytes, and osteoclasts are essential to the process.
[0006]Osteoblasts synthesize the collagenous precursors of bone matrix and also regulate its mineralization. During bone formation, osteoblasts line tiny spaces known as lacunae within the surrounding mineralized matrix. Osteoblasts that line the lacunae are called osteocytes. Osteocytes occupy minute canals (canaliculi) which permit the circulation of tissue fluids. Hormones, growth factors, physical activity, and other stimuli act mainly through osteoblasts to bring about effects on bone. Osteoclasts are derived from hematopoietic stem cells that also give rise to monocytes and macrophages. Osteoclasts adhere to the surface of bone undergoing resorption and lie in depressions referred to as resorption bays. Osteoclasts are apparently activated by signals from osteoblasts. Osteoclastic bone resorption does not occur in the absence of osteoblasts. To meet the requirements of skeletal growth and mechanical function, bone undergoes dynamic remodeling by a coupled process of bone resorption by osteoclasts and reformation by osteoblasts.
[0007]Bone is formed through one of two pathways, by replacement of cartilage or by direct elaboration from periosteum. These processes are known, respectively, as endochondral ossification and intramembraneous ossification. During endochondral ossification, a cartilaginous bone model is first formed. Then, a layer of bone on the surface of the cartilaginous shaft is formed by osteoblasts. Succeeding layers of bone follow. At the same time, the matrix of cartilage cells is calcified into a trabecular network of cartilage while the interstitial cartilage is degenerated. The combined processes of calcification and degeneration of the cartilage advance from the center toward the ends of the cartilage model. The osteoblasts penetrate the cartilage along with capillaries to produce bone on the cartilaginous trabeculae and advance from the center to the ends to progressively form bone on the cartilaginous trabeculae. Ultimately, the calcified cartilage is completely replaced by spongy bone.
[0008]In contrast, the process of intramembraneous ossification does not involve a cartilaginous template. Instead, mesenchymal cells become osteoblasts which begin to form the branching trabeculae of bone. The initial thin trabeculae are some times referred to as spicules. The trabecular bone becomes denser by widening of the trabeculae, and is then remodeled externally and internally. The mandibles, clavicles and certain bones of the skull are produced through intramembraneous ossification.
[0009]There are a number of diseases related to bone formation, deterioration and healing, including osteoporosis, osteogenesis imperfecta (OI) and fibrodysplasia ossificans progressiva (FOP). Osteoporosis, or porous bone, is a disease characterized by low bone mass and structural deterioration of bone tissue, leading to bone fragility and an increased susceptibility to fractures of the hip, spine, and wrist. Osteoporosis is a major public health threat for more than 28 million Americans, 80 percent of whom are women. The strength of bone depends on its mass and density. Bone density depends in part on the amount of calcium, phosphorus and other minerals bones contain. Bones that contain less mineral are weakened and lose internal supporting structure. A full cycle of bone remodeling takes about 2 to 3 months. Children tend to make new bone faster than old bone is broken down. As a result, bone mass increases. Peak bone mass is reached in an individual's mid-30s. Although bone remodeling continues, old bone is broken down faster than new bone is formed. As a result, adults lose slightly more bone than is gained--about 0.3 percent to 0.5 percent a year. Lack of vitamin D and calcium in an individual's diet can accelerate the process. In addition, for women at menopause, estrogen levels drop and bone loss accelerates to about 1 percent to 3 percent a year. Bone loss slows but doesn't stop at around age 60. Women may lose between 35 percent and 50 percent of their bone mass, while men may lose 20 percent to 35 percent of their bone mass. Development of osteoporosis depends on the bone mass attained between ages 25 and 35 (peak bone mass) and how rapidly it is lost as an individual get older. The higher an individual's peak bone mass, the less likely that individual will develop osteoporosis. Calcium, vitamin D and exercising regularly are important for maintaining bone strength. Nonetheless, methods for effectively treating osteoporosis are still desired.
[0010]Osteogenesis Imperfecta (OI) is a genetic disorder characterized by bones that break easily, often from little or no apparent cause. There are at least four distinct forms of the disorder, representing extreme variation in severity from one individual to another. For example, a person may have as few as ten or as many as several hundred fractures in a lifetime. While the number of persons affected with OI in the United States is unknown, the best estimate suggests a minimum of 20,000 and possibly as many as 50,000. OI can be dominantly or recessively inherited and can also occur as a mutation. A cure for OI has not yet been discovered. As a result, methods for treatment focus on preventing and controlling symptoms, strengthening bone mass and ensuring proper healing.
[0011]In addition, both osteoporosis and OI leave patients vulnerable to bone fractures. If these bone fractures do not heal properly, these patients may continue to suffer from pain and may be at increased risk for further fractures as well as other related complications. Methods or treatments that enhance and/or ensure proper fracture healing are important for patients with osteoporosis or brittle bone disease. Another group that will benefit from methods for enhanced wound healing are the elderly and patients who have undergone orthopaedic procedures.
[0012]Fracture healing is the culmination of a highly orchestrated series of physiological and cellular pathways to restore the function of broken bones. Fracture healing generally involves the following steps: the formation of a hematoma (collection of blood at the fracture site), development of a soft callus due to cell multiplication in the lining of the injured bone, growth of blood vessels and fibrocartilege in the middle of the fracture, formation of osteoblasts that migrate into the callus and deposit calcium to form a hard callus, and remodeling and strengthening of the bone through osteoblast and osteoclast formation.
[0013]Osteogenesis during fracture healing occurs by intramembraneous and endochondral ossification that histologically resembles fetal skeletogenesis (Einhorn 1998; Vortkamp et al. 1998; Ferguson et al. 1999). However, the localized tissue hypoxia, the fracture hematoma, subsequent inflammation at the fracture site, and the frank remodeling of the fracture callus at the later stages of healing are unique physiological and cellular responses to bone fractures that have no known corresponding counterpart during fetal development of the skeleton.
[0014]It has been hypothesized that the early physiological responses to a bone fracture, namely hypoxia and inflammation, induce gene expression pathways and promote cell proliferation and migration into the fracture site in order to promote healing (Brighton et al. 1991; Bolander 1992). Production or release of specific growth factors, cytokines, and local hormones at the fracture site by these physiological processes would create the appropriate microenvironment to (1) stimulate periosteal osteoblast proliferation and intramembraneous ossification to form the hard fracture callus, (2) stimulate cell proliferation and migration into the fracture site to form the soft callus, and (3) stimulate chondrocyte differentiation in the soft callus with subsequent endochondral ossification. Remodeling of the fracture callus by osteoclastic resorption and subsequent osteogenesis converts the fracture callus woven bone into cortical bone and thereby restores the shape and mechanical integrity of the fractured bone.
[0015]One potential class of factors that would mediate certain events of fracture healing is the prostaglandins. The effects of prostaglandins on bone metabolism are complex since prostaglandins can stimulate bone formation as well as bone resorption (Kawaguchi et al. 1995). However, because the in vivo half-life of purified or synthetic prostaglandins is very short, prostaglandins per se have a limited therapeutic value.
[0016]Prostaglandins are synthesized by osteoblasts and different cell stimuli can alter the amount and possibly the spectrum of prostaglandins produced by osteoblasts (Feyen et al. 1984; Klein-Nulend et al. 1997; Wadleigh and Herschman 1999). Therefore, signal transduction, mechanical perturbations, or other physiological signals can affect bone metabolism through altercation of prostaglandin production.
[0017]Prostaglandin synthesis begins with the release of arachidonic acid from membrane phospholipids by phospholipase activity. Arachidonic acid is subsequently converted into prostaglandin H2 (PGH2) by cyclooxygenase (COX) via two independent catalytic steps (Needleman et al. 1986). Synthase enzymes then convert PGH2 into the specific prostaglandins produced by that cell such as PGD2, PGE2, PGF2α, prostacyclin, and thromboxane. Thus, cyclooxygenase activity is essential for normal prostaglandin production and cyclooxygenase is believed to be the rate-limiting enzyme in the prostaglandin synthetic pathway.
[0018]There are two known forms of cyclooxygenase, COX-1 and COX-2, which are encoded by two genes (Xie et al. 1991; O'Banion et al. 1992). COX-1 is constitutively expressed by many tissues and provides a homeostatic level of prostaglandins for the body and specific organs, such as the stomach and kidneys (Vane et al. 1998). In contrast, COX-2 is inductively expressed in vitro by a diverse array of cell stimuli such as exposure to lipopolysaccharide (O'Sullivan et al. 1992a; O'Sullivan et al. 1992b), certain cytokines and growth factors (O'Banion et al. 1992; Wadleigh and Herschman 1999), or mechanical stress (Topper et al. 1996; Klein-Nulend et al. 1997). COX-2 expression can be stimulated in vivo by wounding and inflammation (Masferrer et al. 1994; Shigeta et al. 1998; Muscaret al. 2000).
[0019]Inhibiting the cyclooxygenase activity of COX-1 and COX-2 can reduce prostaglandin synthesis by preventing the conversion of arachidonic acid into PGG2, the precursor of PGH2. This is commonly done to reduce inflammation and pain with aspirin and non-steroidal anti-inflammatory drugs (NSAIDs), such as indomethacin. Most NSAIDs inhibit the cyclooxygenase activity of COX-1 and COX-2 with near equal potency, which often leads to detrimental gastro-intestinal or kidney side effects (Raskin 1999; Whelton 1999). Use of COX-2-selective NSAIDs has become very popular since these drugs, such as celecoxib (Celebrex) and rofecoxib (Vioxx) preferentially inhibit the cyclooxygenase activity of COX-2 with selectivity relative to COX-1 of approximately 8-fold for celecoxib and 35-fold for rofecoxib (Riendeau et al. 2001).
[0020]Prostaglandins are produced during fracture healing. Prostaglandin levels in and around the healing callus of rabbit tibia that had been severed by osteotomy showed that PGE and PGF levels were elevated between 1 and 14 and 7 and 14 days post-osteotomy, respectively (Dekel et al. 1981). No survey of the temporal pattern or variety of prostaglandins produced during fracture healing has been reported for other rodents or man. Non-specific NSAIDs have been shown to delay but not stop fracture healing in experimental animal models (Ro et al. 1976; Allen et al. 1980; Altman et al. 1995). In addition, non-specific NSAIDs have been shown to reduce the incidence and severity of heterotopic (abnormal or deviating from the natural position) bone formation in humans following certain fractures or orthopaedic surgical procedures (Pritchett 1995; Moore et al. 1998). These observation suggest that prostaglandins are necessary for bone formation but given the limitations of non-specific NSAID use, it is unknown whether prostaglandins produced by COX-1, COX-2 or both enzymes are essential for fracture healing.
SUMMARY OF THE INVENTION
[0021]The present invention relates to compositions and methods for use in wound healing and for use in enhancing fracture healing, bone formation and wound healing. The present invention further provides for methods for treating diseases related to bones, including osteoporosis, osteogenesis imperfecta and fibrodysplasia ossificans progressiva.
[0022]One embodiment of the present invention involves a vector for use in wound healing comprising a promoter linked to a cyclooxygenase expression cassette. In a further embodiment, the vectors of this invention may be used in gene therapy approaches to enhance wound healing. The wound conditions of this invention can include, bone fractures and skin lesions. The methods of this invention are particularly useful for wound healing in the elderly, patients with osteoporosis and OI, and patients that suffer from delayed wound healing.
[0023]In another embodiment, cyclooxygenase proteins, including COX-1, COX-2 or a combination of the two, are formulated as pharmaceutical compositions for use in wound healing. In a further embodiment of the invention, the pharmaceutical compositions are combined with a carrier for applications in wound healing.
[0024]Another embodiment of the invention provides for the use of the vectors of this invention in gene therapy approaches and/or the pharmaceutical compositions to treat osteoporosis, OI and other related brittle bone conditions. In a further embodiment, the vectors and compositions are used therapeutically to counteract conditions associated with osteoporosis, OI and brittle bones conditions. In a further embodiment, the vectors and compositions are used for wound healing and/or to enhance wound healing in patients with osteoporosis, OI or brittle bone conditions.
[0025]In another embodiment of the invention, COX-2 selective NSAIDs are used in the treatment of heterotopic ossification conditions. Such conditions can include fibrodysplasia ossificans progressiva. In addition, heterotopic ossification can occur following hip replacement procedures and after acetabular fractures. COX-2 selective NSAIDs can be used to treat heterotopic ossification under these circumstances as well.
BRIEF DESCRIPTION OF THE DRAWINGS
[0026]FIG. 1. Radiographic Analysis of Fracture Healing in NSAID Treated Rats
[0027]High resolution radiographs were made immediately post-fracture and then every week till the endpoint of the experiment (8 weeks) using a Hewlett-Packard Faxitron. Shown are radiographs of the fractured right femurs from the same rats taken at 2 (top), 4 (middle), and 8 weeks (bottom) post-fracture (dorsal-ventral view). As indicated, panels A-D shows radiographs from a no drug rat, an indomethacin treated rat, a celecoxib treated rat, and a rofecoxib treated rat, respectively. Note that the fracture is still clearly evident in the 8-week post-fracture radiographs of the celecoxib and rofecoxib treated rats.
[0028]FIG. 2. COX-2-Selective NSAIDs Alter the Mechanical Properties of Fractured Femurs
[0029]Mechanical testing data was obtained or derived as described in the Experimental Procedures. The data for each fractured femur was normalized as a percentage of the value obtained from that animal's contralateral, unfractured femur, except for shear modulus panel D). Shown are the mean normalized values at each time point and for each treatment group or the mean shear modulus values in panel D. The error bars represent standard errors of the mean. Pairwise t-test were made between the no drug and experimental treatments within a time point. Statistical significance differences (P<0.05) are noted with asterisks.
[0030]FIG. 3. COX-2-Selective NSAIDs Disrupt Endochondral Ossification During Fracture Healing
[0031]Shown are fracture calluses from no drug, indomethacin, celecoxib, and rofecoxib treated rats at 2, 3, and 4 weeks post-fracture as indicated. The specimens were embedded in polymethylmethacrylate (PMMA), sectioned, and stained with van Gieson's picrofuchsin and Stevenel's blue so that bone is red, calcified cartilage is orange to red, cartilage is deep blue to purple, and fibrous tissue and muscle is pale blue. Each section is oriented with the cortical bone on the bottom, fracture callus on top, and fracture site in the middle. Note the abnormal cartilage morphology in the calluses of the NSAID treated rats (panels D, G, and J) and the lack of cartilage in the celecoxib (panels H and I) and rofecoxib treated (panels K and L) rats at 3 and 4 weeks post-fracture.
[0032]FIG. 4. Abnormal Bone Resorption during Fracture Healing in COX-2-Selective NSAID Treated Animals
[0033]Shown are fracture calluses from rofecoxib treated rats at 3 (panels A and B) and 4 weeks (panel C) post-fracture. The orientation of panels A and B are the same with external callus on top and fracture site to the immediate left of the panel. In panel C, the external callus is to the left and the fracture site is at the immediate bottom of the panel. The specimens were embedded in PMMA, sectioned, and stained with van Gieson's picrofuchsin and Stevenel's blue so that bone is red, calcified cartilage is orange to red, cartilage is deep blue to purple, and other cell types are shades of blue. Original photographic magnification is indicated. CB: cortical bone; WB: woven bone; CC: calcified cartilage; Ca: cartilage; F: fracture site; ab: air bubble; M: area of magnification shown in panel B; and Oc: osteoclasts. The NSAID treated rats often developed areas of high bone resorption at the cortical bone, fracture site, external callus junction (M) as seen in panel A. The air bubble (ab) seen in panel A is an artifact of the PMMA embedding. At higher magnification, osteoclasts (Oc) can be seen lining the cortical bone surface of area M in the 3 week fracture callus as denoted by the arrows. Shown in panel C is an identical area of a 4 week post-fracture callus as shown in panel B. The extent and area of bone resorption appears to be greater at 4 weeks post-fracture and also often encompassed all surfaces of the cortical bone at the fracture site. Similar bone resorption patterns were seen in celecoxib treated rats and to a lesser extent in the indomethacin treated rats.
[0034]FIG. 5. Experimental Complications Associated with NSAID Treatment during Fracture Healing
[0035]Complications that necessitated the pre-mature euthanization or resulted in the pre-mature death of a rat during the course of these experiments were compiled and used to determine the effects of NSAID treatment on anesthetic death, infection, and pin slippage. Experimental treatment group values were compared to the no drug values using a χ2 analysis. Significant differences are noted with an asterisk (P<0.01). As can be seen, pin slippage was by far the most common complication and was significantly different for each treatment group relative to the no drug rats with P-values of less than 1E-4, 1E-7, and 1E-24 for the indomethacin, celecoxib, and rofecoxib treated rats respectively. The rofecoxib treated rats were also found to have a statistically significant higher infection rate as compared to the no drug rats (P<0.0001). Death from anesthesia was not different between groups.
[0036]FIG. 6. Cox2 but not Cox1 is Essential for Normal Bone Fracture Healing
[0037]The right femora of Cox1.sup.-/- and Cox2.sup.-/- mice were fractured and examined radiographically and histologically at 2 weeks post-fracture. Panels A and D. Radiographs of fractured femurs from a Cox1.sup.-/- and a Cox2.sup.-/- mouse, respectively. Note the lack of mineralized tissue (X-ray dense) in the fracture callus region of the Cox2.sup.-/- mouse. Panels B and C. Sagital section through the fractured femur of a Cox1.sup.-/- stained with Masson's trichrome stain (cell nuclei-purple; muscle and cytoplasm=red; collagen and bone=blue). Panels E and F. Sagital section through the fractured femur of a Cox2.sup.-/- stained with Masson's trichrome stain. Note the presence of chondrocytes within the Cox2.sup.-/- callus but the lack of endochondral ossification relative to the Cox1.sup.-/- mouse fracture callus. Original photographic magnification is indicated. The Cox2.sup.-/- mouse fracture callus specimens are shown at higher magnification because the callus was smaller. B: bone; C: chondrocytes and cartilage; E: area of endochondral ossification; M: area of intramembraneous bone formation; F: fracture site.
DETAILED DESCRIPTION OF THE INVENTION
[0038]The present invention involves methods for enhancing wound healing and treating conditions of the bone with the use of cyclooxygenase (COX). COX is an enzyme that converts arachidonic acid into prostaglandin H2 (PGH2). PGH2 is then converted into specific prostaglandins, which affect bone metabolism and formation. There are two forms of cyclooxygenase, COX-1 and COX-2. COX-1 is constitutively expressed, while COX-2 is inductively expressed by various stimuli. Examples of such stimuli are wounding and inflammation. The subsequent expression of COX-2 in response to these stimuli results in the production of pro-inflammatory prostaglandins. The pain and inflammation associated with a wound are commonly treated with non-steroidal anti-inflammatory drugs (NSAIDs). Most NSAIDs inhibit the cyclooxygenase activity of both COX-1 and COX-2. However, due to the inhibition of COX-1 activity, gastro-intestinal and kidney side effects result from NSAIDs use. COX-2 selective NSAIDs have also been developed which preferentially inhibit COX-2 activity relative to COX-1 activity and thereby avoid the side effects associated with non-specific NSAIDs.
[0039]In addition to its inflammatory function, one aspect of the invention relates to the necessity of COX-2 activity in wound healing. As described in more detail below, COX-2 is essential for normal wound healing. In another aspect of the invention, treatment systems and methods employ COX-2 to enhance wound healing.
[0040]In a preferred embodiment of the invention, gene therapy techniques are employed to increase cyclooxygenase activity at the site of the wound to enhance wound healing. Gene therapy techniques allow an absent or faulty gene to be replaced with a working gene. They also allow for the delivery and controlled expression of therapeutic gene products. One embodiment of the invention provides for a vector containing a cyclooxygenase expression cassette. In a further embodiment, the vector containing the cyclooxygenase expression cassette is delivered to the wound using gene therapy techniques. The gene therapy techniques may use adenoviral vectors, adeno-associated viral vectors, recombination-defective retrovirus vectors or DNA vectors to deliver the cyclooxygenase expression cassette to the wound.
[0041]The vectors of this invention may be used to increase cyclooxygenase levels at the site of the wound and thereby enhance would healing. For example, these vectors may be used to enhance wound healing following bone fractures or orthopaedic procedures. The vectors may also be of particular use to the elderly and other patient groups that have delayed bone healing, including smokers, diabetics, and steroid users. In addition, the vectors may be used in the treatment of osteoporosis and osteogenesis imperfecta using gene therapy techniques for gene replacement and to enhance healing for wounds resulting from these diseases.
[0042]One aspect of the invention involves a vector that contains a cyclooxygenase expression cassette that encodes a cyclooxygenase gene product. The vector includes all necessary sequences for the expression of the cyclooxygenase expression cassette and any sequences that may be included to control the expression of the cassette. These sequences may include, but are not limited to, a promoter or initiation sequence, an enhancer sequence, termination sequence, RNA processing signals, and/or a polyadenylation signal sequence.
[0043]The term "vector" refers to a nucleic acid construct that encodes for a particular gene product. The vectors of the present invention are preferably adenoviral, adeno-associated viral, recombination-defective retrovirus, plasmid or DNA vectors.
[0044]The term "cyclooxygenase expression cassette" refers to nucleic acid which codes for cyclooxygenase enzyme, preferably COX-1 or COX-2. The expression cassette is positioned within the vector such that it can be transcribed into RNA and translated into the cyclooxygenase protein product. Table 3 provides for the human cDNA sequences for COX-1 (SEQ. ID. No. 1) and COX-2 (SEQ. ID. No. 2).
[0045]The term "necessary sequences for the expression of cyclooxygenase" refers to sequences necessary to ensure the transcription and translation of the expression cassette. The term "promoter" refers to a DNA sequence that is bound by RNA polymerase and is required to initiate transcription of a gene. There are a number of promoters that are known in the art, including those that can enhance or control expression of the gene or expression cassette. For example, cytomegalovirus promoter may be fused to the cyclooxygenase expression cassette to obtain constitutive expression of cassette or a COX-2 promoter may be linked to a cyclooxygenase cDNA to allow for expression of cyclooxygenase in a more normal fashion. For example, a COX-2 promoter may be linked to a COX-1 cDNA. This may allow a patient to remain on COX-2 selective NSAIDs but still have the ability to heal. In another aspect of the invention, the promoter may be induced in response to inflammation. Preferably, the inducible promoter employs promoter, or the complement system gene promoters. Other inflammatory gene promoters include promoters for TNF-α, or the NF-κB response element.
[0046]In another aspect of the invention, the vectors are delivered directly to the location of the wound by injection or direct application in order to enhance wound healing. The vectors of this invention may also be administered by electroporation or delivered as an aerosol. The vectors may be administered or delivered in saline solutions, encapsulated in liposomes, in polymer solutions, in gels or lyophilized. In an alternative aspect, targeted transfection may be used to deliver the vectors in vivo. The term "wound" refers to a bone fracture, the site of a surgical orthopaedic procedure, or a skin lesion. In another aspect of the invention, the vectors may be delivered ex vivo, wherein a patient's cells are transfected ex vivo with the vectors of this invention and the transfected cells are then returned to the patient.
[0047]A further aspect of this invention provides for pharmaceutical compositions and methods of their use in wound healing. These pharmaceutical compositions include cyclooxygenase protein formulations and/or pharmaceutically acceptable carriers. The cyclooxygenase protein formulations comprise COX-1 protein, COX-2 protein or a combination of the two that are formulated such that the proteins remain stable, retain their function, including their enzymatic activity, and are physiologically acceptable. The human amino acid sequences for COX-1 (SEQ. ID. NO. 3) and COX-2 (SEQ. ID. NO. 4) proteins are provided in Table 3. There are a variety of pharmaceutically acceptable carriers that are known in the art, including, but not limited to, saline, liposomes, gels and polymers. The formulation and/or carrier may also be lyophilized for aerosol delivery. In a further embodiment of the invention, these pharmaceutical compositions are administered topically, orally, intravenously, nasally or via inhalation, or locally for use in wound healing. Preferably, these composition are administered to enhance wound healing.
[0048]In another aspect of the invention, methods of enhancing wound healing in the elderly are provided. COX-2 gene expression is reduced with age as are wound healing and bone formation. In one embodiment, vectors containing cyclooxygenase expression cassettes are administered to elderly to enhance would healing by increasing COX-2 expression during wound healing. A further aspect of this invention provides for methods for enhancing wound healing in patient groups that have delayed wound healing by delivering the vectors of this invention to these patients during wound healing.
[0049]In another embodiment of the invention, the vectors of this invention are used to enhance wound healing in patients suffering from osteoporosis or OI by employing gene therapy techniques to deliver cyclooxygenase expression cassettes to the wound to enhance healing.
[0050]In another aspect of the invention, a method for treating pathological heterotopic ossification conditions is provided. Pathological heterotopic ossification conditions are diseases characterized by abnormal bone formation at locations of inflammation. An example of a pathological heterotopic ossification condition is fibrodysplasia ossificans progressiva (FOP). In one embodiment of the invention, COX-2 selective NSAIDs are administered to locations of inflammation that lack a wound. The COX-2 selective NSAIDs inhibit COX-2 activity thereby preventing abnormal bone formation.
[0051]Our experimental approach was to assess fracture healing using a standard rat closed femur fracture model in which COX-2 function was inhibited in vivo with the COX-2-selective NSAIDs, celecoxib and rofecoxib. The results were striking in that femur fracture healing in rats treated with celecoxib or rofecoxib was dramatically impaired. These observations were confirmed by examining fracture healing in Cox1 and Cox2 null mice. Histological observations indicated that the defect in fracture healing caused by the COX-2-selective NSAIDs or by lack of Cox2 occurred in the endochondral ossification pathway.
[0052]Using a standard rat closed femur fracture model, we have demonstrated that COX-2-selective NSAID treatment can stop normal fracture healing and induce the formation of mal-unions and non-unions. Non-unions refer to fractures that show no visible sign of healing. Mal-unions refer to bones that do not heal properly and result in misalignment of bones. These observations are in grim contrast to those observations we, and others have obtained by treating rats with non-selective NSAIDs such as indomethacin when it was observed that fracture healing was delayed but not prevented (Ro et al. 1976; Ro et al. 1978). Our observations suggest that COX-2 has an essential function during normal fracture healing and that COX-2-selective NSAID inhibition of prostaglandin synthesis stops normal fracture healing. This also suggests that the inflammatory phase is critical for normal fracture healing.
[0053]Consistent with the effects of COX-2-selective NSAIDS on rat femur fracture healing, mice homozygous for a targeted mutation in Cox2, but not Cox1, also showed inhibited fracture healing (FIG. 6). The cDNA sequences (SEQ. ID. No. 5 for COX-1 and SEQ. ID. No. 6 for COX-2) and corresponding amino acid sequences (SEQ. ID. NO. 7 for COX-1 and SEQ. ID. No. 8 for COX-2) for mouse COX-1 and COX-2 gene and protein are provided in Table 4. This excludes the possibility that any additional inhibitory activity against other cellular processes or proteins by celecoxib or rofecoxib (Jones et al. 1999; Hsu et al. 2000; Rossi et al. 2000) is primarily responsible for negatively affecting fracture healing.
[0054]The amount of rofecoxib used to treat the rats (3 mg/kg) in this study was approximately 4 times the nominal, maximum human daily dose of 50 mg (0.7 mg/kg) that is used to manage acute pain. In contrast, the celecoxib dose used to treat rats in this study was in the recommended dose range for humans. The rats received 280 mg/70 kg body weight of celecoxib once per day whereas the recommended human maximum daily dose of celecoxib is 200 mg twice a day. Additionally, the rats in this study received daily doses of each drug till the endpoint of the experiment, which is unlike common clinical scenarios when COX-2-selective NSAIDs are used to acutely manage pain, inflammation, and swelling following a fracture. In contrast, arthritis patients, in particular, do use COX-2-selective NSAIDs on a daily basis for extended periods. Based simply upon animal body weight and drug dose, and without accounting for pharmacokinetic variables, the indomethacin dose used (1 mg/kg) would be predicted to inhibit most COX-1 activity but to only partially inhibit COX-2 activity; the celecoxib dose used should inhibit most if not all COX-2 activity and possibly inhibit some COX-1 activity; and the rofecoxib dose used should completely inhibit COX-2 activity but not affect COX-1 (Warner et al. 1999). The estimated plasma half-lives for celecoxib and rofecoxib in male rats following a single drug dose are approximately 4 and 5 hours respectively (Halpin et al. 2000; Paulson et al. 2000). In humans, however, the estimated elimination half-life for celecoxib is 11 hours (Davies et al. 2000) and the plasma half-life for rofecoxib is approximately 10-17 hours depending upon drug dose (Depr et al. 2000). Unfortunately, these data were unknown to us when this study was initiated and consequently our rat drug dosing regime may actually be an under representation of the COX-2 inhibition level over a 24 hour cycle as that for humans receiving similar drug doses. Additionally, celecoxib and rofecoxib may have different inhibitory concentrations for rat versus human COX-2. Despite the pharmacokinetic variations between rats and humans receiving COX-2-selective NSAIDs, our data clearly indicate that these drugs have a dramatic negative effect on fracture healing in mammals, and thus caution in the use of these COX-2-selective NSAIDs in humans is warranted.
[0055]The fractured femurs of the celecoxib treated rats had increased mechanical properties and healed as mal-unions, rather than non-unions, in approximately 50% of the rats tested (Tables 1 and 2). Had the experimental endpoint been extended, some of the celecoxib treated rats may have gone on to heal their femur fractures. However, the delay in any such healing among the celecoxib treated rats would have been significantly longer than in no drug or indomethacin treated rats.
[0056]After mechanically testing the celecoxib and rofecoxib treated fractured femurs, we observed that the fracture callus had a shell-like morphology. The periphery of the fracture callus was bone that sometimes partly bridged the fracture gap and thus formed a mal-union in the celecoxib treated rats. However, little or no bone was present between the peripheral bone of the callus and the original femoral cortical bone ends. Often this space appeared to be filled with fatty marrow. Also strikingly apparent was the lack of new bone or primary bone healing at the cortical bone ends.
[0057]The indomethacin dose used in this study (1 mg/kg) was previously shown to delay fracture healing in rats and was used principally as a positive control (Altman et al. 1995). Increasing indomethacin doses to levels that which would completely inhibit COX-2 (and COX-1 activity) causes a steep increase in rat mortality from gastrointestinal bleeding (Allen et al. 1980; Wallace et al. 1998). Consequently, it would be difficult to directly compare the effects of COX-2-selective and traditional NSAIDs on fracture healing based solely upon COX-2 inhibition levels. An alternative approach would be to measure prostaglandin levels within and around the fracture callus during healing in control and drug treated rats and to then correlate those observations with different healing parameters.
[0058]The abnormal osteoclastic response observed at the fracture site in the NSAID treated rats is counter to experimental observations in which prostaglandins stimulate bone resorption (Klein and Raisz 1970) and COX-2 function promotes osteoclast formation (Okada et al. 2000). Lack of prostaglandins in the fracture callus could induce osteoclastic activity through an undescribed mechanism, or perhaps through an indirect mechanism such as increased mechanical instability at the fracture site. An additional possibility is that the amount and/or repertoire of prostaglandins produced at the fracture site in the NSAID treated rats is competent for inducing osteoclastic activity but insufficient for osteogenesis to proceed normally. The second possibility is favored since the relative short half-life of celecoxib and rofecoxib in rats should produce daily periods in which COX-2-dependent prostaglandin synthesis could occur and because a similar osteoclastic response was not observed in the Cox2.sup.-/- mouse fracture callus (FIG. 6). In addition, this large osteoclastic activity may be the causative factor involved in fracture destabilization by pin slippage, which was the major morbidity complicating these experiments (FIG. 5).
[0059]Chondrocyte differentiation and persistence in the fracture callus appears to be altered by COX-2-selective NSAID treatment and by lack of Cox2 (FIG. 6). Cartilage that is evident in the 2-week fracture calluses of COX-2-selective NSAID treated rats either disappears or is dramatically reduced in the 3 and 4-week fracture calluses (FIG. 3). In control rats, the fracture has bridged or is almost bridged by 4 weeks post-fracture as the cartilage present at early times had undergone normal endochondral ossification and is no longer evident. In contrast, cartilage within the indomethacin treated fracture callus does persist at 4 weeks post-fracture indicating a reduced rate of endochondral ossification. These observations suggest that the chondrocytes within the COX-2-selective NSAID treated rats deteriorated without forming a cartilage matrix in which endochondral ossification could occur. This phenomenon then leads to the development of fracture non-unions and mal-unions. Similar histological observations were seen in the Cox2.sup.-/- mouse fracture where chondrocytes present in the callus failed to form a mineralized matrix (FIG. 6). In support of this hypothesis, teratocarcinoma chondrocytes developed from Cox2.sup.-/- embryonic stem cells were found to be deteriorating, hypotrophic, and undergoing apoptosis (Zhang et al. 2000). Additionally, the Cox2.sup.-/- mouse fracture callus was much smaller than that from the Cox1.sup.-/- mouse suggesting that mesenchymal cell proliferation and migration into the Cox2.sup.-/- mouse fracture site is reduced or that the mesenchymal cells proportionately differentiate into fewer chondrocytes. Thus, COX-2 function is essential for normal progression of endochondral ossification during fracture healing.
[0060]There are several steps during endochondral ossification at which COX-2 could exert an essential regulatory function. Endochondral ossification is a complicated process that begins when chondrocytes mature to produce a cartilage matrix. Eventually the chondrocytes become hypertrophic and undergo apoptosis as the cartilage matrix matures and becomes calcified. Osteoclasts partially resorb the calcified cartilage with concurrent angiogenesis at the site of endochondral ossification. Osteoblasts proliferate and differentiate on the calcified cartilage and begin forming new bone. Remodeling of the new bone subsequently occurs to increase the mechanical properties of the bone and restore its normal architecture. Prostaglandins produced by COX-2 could be used to enhance osteoblast proliferation and differentiation (Kawaguchi et al. 1995). Prostaglandins may also be necessary to promote terminal differentiation of chondrocytes and formation of the cartilage matrix. Such an effect on chondrocytes could occur through a direct effect on differentiation or indirectly by preventing pre-mature apoptosis since inhibition of COX-2 function by COX-2-selective NSAIDs has been shown to induce apoptosis in cancer cell lines (Hsu et al. 2000). The potential effects of prostaglandins on osteoblasts and chondrocytes during endochondral ossification are not mutually exclusive. An additional possibility is that COX-2 dependent signaling occurs between osteoblasts and chondrocytes to initiate and maintain endochondral ossification. Prostaglandins are also known to promote osteoclast activity, which may be essential for the normal endochondral ossification process. However, our histological observations indicate an exuberant osteoclast response at the fracture site in the NSAID treated rats thus we do not favor this potential mechanism. Angiogenesis is also inhibited by COX-2-selective NSAIDs, at least in certain experimental models (Jones et al. 1999), and we failed to detect neovascularization of the Cox2.sup.-/- mouse fracture callus. Thus an additional possibility is that lack of angiogenesis precludes proper delivery of osteoclasts and osteoblasts to the cartilage matrix interface for continued endochondral ossification.
[0061]Our observations suggest that COX-2-selective NSAIDs may be effective in reducing or preventing pathological heterotopic ossification. One particular genetic disease for which COX-2-selective NSAIDs may be efficacious is fibrodysplasia ossificans progressiva (FOP) (Cohen et al. 1993). Children afflicted with FOP develop debilitating heterotopic bone through an endochondral ossification pathway that appears to initiate at sites of inflammation (Kaplan et al. 1993). Thus the COX-2-selective NSAIDs may be useful in reducing inflammation and stopping endochondral ossification at presumptive heterotopic ossification sites in children with FOP.
[0062]To determine if COX-2 functions in fracture healing, rats were treated with COX-2-selective non-steroidal anti-inflammatory drugs (NSAIDs) to stop COX-2-dependent prostaglandin production. Radiographic, histological, and mechanical testing demonstrated that fracture healing failed in rats treated with COX-2 selective NSAIDs. Normal fracture healing also failed in mice homozygous for a null mutation in the COX-2 gene. These results demonstrate that COX-2 activity is necessary for normal fracture healing and confirms that the effects of COX-2-selective NSAIDs on fracture healing is due to inhibition of COX-2 activity and not from a drug side effect. Furthermore, histological observations suggest that COX-2 is required for normal endochondral ossification during fracture healing. Since mice lacking Cox2 form normal skeletons, our observations indicate that fetal bone development and fracture healing are different and that COX-2 function is specifically essential for fracture healing.
Experimental Procedures
Animals, Drug Dosage, and Administration
[0063]A total of 253 male Sprague-Dawley rats (584.+-0.62 g) were fed a standard diet and kept caged separately in a constant temperature and humidity environment. All rats were 6-9 months old at the beginning of the experiment. Drugs were administered daily by gavage beginning two days prior to fracture. Animals were randomly selected for each treatment group. The rats were gavaged with aqueous suspensions of indomethacin (1 mg/kg), celecoxib (4 mg/kg), or rofecoxib (3 mg/kg). No drug (control) rats were not initially gavaged but later rats were gavaged with water and no difference was noted between the no drug rats that had been gavaged and those that had not. No statistically significant differences were found in animal weight changes during the experiments.
[0064]Retired breeder female Cox1.sup.-/- (B6; 129P2-Ptgs1.sup.tm1) and Cox2.sup.-/- (B6; 129P2-Ptgs2.sup.tm1) mice were obtained from Taconic Farms. Closed femur fracture production was done using a method similar to that described below.
Closed Femur Fracture Production
[0065]The rats were anesthetized by intraperitoneal injection of ketamine (40 mg/kg) and xylazine (5 mg/kg). Under aseptic conditions, a medial parapatellar incision (0.4-0.5 cm) was made in the right hindlimb and the patella was dislocated laterally. The medullary canal was entered through the intercondylar notch and reamed with an 18-gauge needle. A 1.1 mm stainless steel pin (Small Parts Inc., Miami Lakes, Fla.) was then inserted into the canal and secured in the proximal part of the greater trochanter by tamping. The distal portion of the pin was then cut flush with the femoral condyles and the patella dislocation was reduced. The soft tissue and skin were closed with 4-0 vicryl sutures. After closing, the diaphysis of the pinned femur was fractured by means of a three-point bending device as described by Bonnarens and Einhorn (Bonnarens and Einhorn 1984).
Radiography
[0066]Radiographs were made post-fracture to confirm the position and quality of each fracture and at sacrifice to determine the degree of healing. In addition several rats were selected randomly to produce serial radiographs (at least 2 rats per treatment group). Radiographs were made of these rats weekly under anesthesia until the experimental endpoint (8 weeks). Radiographs of mice were also made under anesthesia. All radiographs were made using a 43805N Faxitron (Hewlett-Packard, McMinnville, Oreg.) and Kodak MinR-2000 mammography film.
Mechanical Testing
[0067]Animals within each treatment group were sacrificed at 4, 6, and 8 weeks post-fracture by CO2 asphyxiation. Animals with oblique, comminuted, or infected fractures were not used for mechanical testing. Both femora were removed and cleaned of all soft tissue leaving the fracture callus undisturbed and then immediately processed for mechanical testing. The samples were wrapped in saline soaked gauze to prevent dehydration between steps. Measurements of the femora were taken using digital calipers to determine femur length and external callus dimensions. The intramedullary pin was removed from the fractured femur and a 1 mm-diameter stainless steel pin (˜0.8 cm length) was inserted at the proximal and distal end perpendicular to the long axis of the bone to prevent slipping in the potting material. The intact femur was also pinned as described above. The femoral ends were potted in 1-inch hexnuts using a low melt temperature metal (Wood's metal, Alfa Aesar, Ward Mill, Mass.). Once potted, the gage length (L) of each femur was measured. Torsional testing was conducted using a servohydraulic testing machine (MTS, Eden, Praire, Minn.) with a 20 Nm reaction torque cell (Interface, Scottsdale, Ariz.). The testing was carried out to failure at a rate of 2°/sec and a data recording rate of 20 Hz. Both the fractured and intact femora were tested in internal rotation in proper anatomic orientation. The peak torque and angle at failure were calculated from the load-deformation curves. Internal fracture callus dimensions were measured after mechanical testing. From the callus dimensions, the polar moment of inertia (J) was calculated based upon a hollow ellipse model (Bell et al. 1941; Engesaeter et al. 1978). The equations used to derive torsional rigidity, shear stress, shear modulus, and J were as follows (Popov 1968): (1) Torsional Rigidity: (TmaxL)/φ where Tmax is the peak torque value in Nmm, L is the gage length in mm, and φ is the angle at failure in radians. (2) Shear Stress: (TmaxRmax)/J where Rmax is the largest radial dimension of the fracture callus in mm (ao) and J is the polar moment of inertia. (3) Shear Modulus (G): (TmaxL)/J. (4) Polar Moment of Inertia (J): [π(ab3+a3b-(a-t)(b-t)3(-(a-t)3(b-t)/]4 where ai is [ai+[(ao-ai)/2]; b is [bi+[(bo-bi)/2]; t is the average bone thickness at the site of failure and is calculated as [(ao-ai)+(bo-bi)]/2 where ao is the callus maximum outside radius, a; is the maximum interior radius, bo is the least outside radius, and bi is the least interior radius in mm. Only torsional testing data for which the fractured and control femur tested without incident were used.
Histology
[0068]Rats were sacrificed at 2, 3, 4, 6, and 8 weeks post-fracture by CO2 asphyxiation. Both femora were resected and the stainless steel pin was removed from the medullary canal. The harvested femora were fixed in 10% buffered formalin and embedded in polymethylmethacrylate following standard histological techniques for calcified tissue. The samples were sectioned sagitally through the fracture callus using an Isomet diamond saw (Buehler Ltd., Lake Bluff, Ill.), mounted on plexiglass slides, and polished to a thickness of 100 μ.m. The slides were then stained with van Gieson's picrofuchsin and Stevenel's blue in order to identify new bone growth and cartilage formation (Maniatopoulos et al. 1986). Mice femora were fixed, decalcified, paraffin embedded, sectioned, and stained with Masson's trichrome stain. The samples were viewed and photomicrographs were taken using an Olympus BH2-RFCA microscope or an Olympus SZ40 microscope.
Treatment with COX-2-Selective NSAIDs Leads to Fracture Non-Unions and Mal-Unions
[0069]Femur fracture healing was followed by serial radiographic analysis of rats treated with celecoxib, rofecoxib, indomethacin, or gavaged daily with water (no drugs group). Radiographs were made immediately following fracture production and then every week till the end point of the experiments (8 weeks post-fracture). Representative results are shown in FIG. 1.
[0070]We found that femur fracture healing proceeded normally in the no drug rats as expected. At 1 week post-fracture, formation of the hard callus could be detected radiographically but was more evident at 2 weeks (FIG. 1A). By 4 weeks post-fracture, calcification of the soft callus was clearly evident indicating that endochondral ossification had occurred. Additionally, by 4 weeks post-fracture, the new bone formed during fracture repair had almost bridged the fracture gap. Bridging of the fracture and remodeling of the fracture callus were evident at 6 weeks post-fracture. Continued remodeling of the callus as well as remodeling of the original femoral cortical bone at the fracture site is clearly evident by 8 weeks post-fracture. These radiographic observations are typical of normal fracture healing.
[0071]Indomethacin treatment appeared to delay but not prevent fracture healing consistent with previous reports (Ro et al. 1976; Allen et al. 1980; Altman et al. 1995). By 2 weeks post-fracture, an X-ray dense hard callus is clearly evident in the indomethacin treated rats (FIG. 1B). However, bridging of the fracture gap did not appear to occur until 5-6 weeks post fracture as compared to approximately 4-5 weeks post-fracture in the untreated rats (compare FIGS. 1A and B). Bridging and remodeling were evident in the 8 week post-fracture radiographs of the indomethacin treated rats.
[0072]Celecoxib or rofecoxib treatment did not prevent formation of an X-ray dense hard callus as can be seen in the 2 and 4 week post-fracture radiographs (FIGS. 1C and D). However, the original fracture was still plainly evident in the celecoxib (FIG. 1C) and rofecoxib (FIG. 1D) treated rats even after 8 weeks. No rofecoxib treated rat was observed to have a normally bridged callus by radiography. However, non-unions, mal-unions, and unions of the fractured femurs were observed by radiography in the celecoxib treated rats. The mal-unions were typified by the radiograph seen in FIG. 1C in which one cortex of the fracture callus was bridged but in which the original cortical bone ends of the fractured femur had not joined and the fracture was still clearly evident.
[0073]In addition to the serial radiographs made for certain rats, all animals in this study were examined radiographically immediately post-fracture and when euthanized. A random, blinded sample of the 8-week post-fracture radiographs were independently examined by 7 observers and scored as a union (1 point), mal-union (0.5 points), or non-union (0 points). Control rats had an average score of 0.71. In contrast, the indomethacin, celecoxib and rofecoxib treated rats had average scores of 0.54, 0.49, and 0.32, respectively. Despite the known difficulties associated with judging fracture healing from radiographs (Nicholls et al. 1979; Panjabi et al. 1989), a statistical comparison between treatment groups showed that the no drug and rofecoxib groups were significantly different (P<0.007 using a Fisher's PLSD test at a 5% significance level).
[0074]These observations clearly indicate that rofecoxib treatment inhibits fracture healing in rats leading to non-unions. It would also appear that at least by radiographic examination, celecoxib treatment negatively affects fracture healing to an extent similar to, if not worse than, indomethacin treatment.
The Mechanical Properties of the Healing Femur Fracture Callus are Diminished by NSAID Treatment
[0075]In conjunction with the radiographic analysis, torsional mechanical testing of fractured femurs was also performed. The fractured femur and contralateral control femur from rats at 4, 6, and 8 weeks post-fracture were tested to failure in torsion for each treatment group (no drugs, indomethacin, celecoxib, and rofecoxib). The data from these tests is summarized in FIG. 2 and Table 1. Peak torque is the maximum twisting force generated during torsional testing of the femur. Torsional rigidity is a measure of a structure's resistance to torque. Thus a bone with high torsional rigidity would fail after only a few degrees of rotation, but soft tissue would not reach its peak torque until after a large angular deflection. Maximum shear stress is a measure of the ultimate shearing force withstood by the femur prior to failure and is a function of the applied torque and polar moment of inertia, which is dictated by callus geometry. Shear modulus measures the elastic resistance to deformation by a shearing stress for a given material and is constant for a given material.
[0076]We found in the no drug rats that the normalized peak torque (101%) and torsional rigidity (88%) of the fractured femur was restored by 8-weeks post-fracture as compared to the contralateral control femurs from each animal (FIG. 2). However, the shear modulus (1.3 GPa) and normalized shear stress (48%) of the fractured femurs at 8 weeks post-fracture were still less than the contralateral control femurs (FIG. 2, Table 1). This is the expected result because during fracture healing, the ultimate mechanical integrity of the fractured bone, that is peak torque, is maintained at a high level by increasing bone diameter via the fracture callus. Since the mechanical properties of the initially soft tissue within the callus and later the newly formed bone are much weaker than the mechanical properties of mature cortical bone; shear stress and shear modulus were, as expected, less than the contralateral control femurs. As the newly formed bone within the fracture callus matures by remodeling, the mechanical properties of the fractured bone increase. This is evident in our results as increases in shear stress and shear modulus with time (FIG. 2). The high normalized torsional rigidity found for the fractured femurs in the no drug rats at 6 (89%) and 8 (88%) weeks post-fracture indicates that the fracture had been bridged by new bone as would be expected.
[0077]We also observed that all of the 6 and 8 week post-fracture no drug femurs and all the contralateral control femurs failed as predicted, mid-diaphyseal spiral fractures during the torsional mechanical testing.
[0078]Indomethacin treatment reduced the mechanical properties of the healing femur fractures at earlier time points (FIG. 2). However by 8 weeks post-fracture, the normalized peak torque, torsional rigidity, and shear stress values obtained from the indomethacin treated rats were not significantly different from the no drug rats. In contrast, at 6 weeks post-fracture, the normalized peak torque, torsional rigidity, and shear stress values (46, 34, and 15%, respectively) obtained from the indomethacin treated rats were less than the no drug rats at 6 weeks post-fracture (77, 89, and 36%, respectively). Pointedly, the significantly low torsional rigidity of the fractured femurs from the indomethacin treated rats at 6 weeks post-fracture indicates that the fracture had not been bridged by bone. Of the eight fractured femurs tested at 8 weeks post-fracture, 6 failed as unions, 1 failed as a mal-union, and 1 failed as a non-union (Table 2). These observations indicate that the non-selective NSAID, indomethacin, delays, but does not prevent fracture healing, which is consistent with previous studies and demonstrates the validity of our assay methods (Ro et al. 1976; Allen et al. 1980; Altman et al. 1995).
[0079]Rofecoxib treatment had a drastic effect on the mechanical properties of the fractured femurs. At 8 weeks post-fracture, for all values measured or derived, the mechanical properties of the fractured femurs from the rofecoxib treated rats were significantly less than the no drug rat fractured femurs (FIG. 2, Table 1). At 8 weeks post-fracture, the fractured femurs of the rofecoxib treated rats had only obtained 50%, 31%, and 18% of peak torque, torsional rigidity, or maximum shear stress of the contralateral unfractured femurs, respectively. The low torsional rigidity and shear modulus (0.5 GPa) values obtained from the fractured femurs of the rofecoxib treated rats are consistent with healing failure and the formation of non-unions. In addition, whereas all of the contralateral control femurs from the rofecoxib treated rats failed as mid-diaphyseal spiral fractures, 4 of the 5 fractured femurs at 8 weeks post-fracture failed as non-unions and the other failed as a malunion (Table 2). These observations demonstrate that, at the dose and treatment regime employed, the COX-2-selective NSAID rofecoxib stops fracture healing.
[0080]Unlike rofecoxib treatment, no significant differences were found in the mechanical properties of the healing fractured femurs from the celecoxib treated rats as compared to no drug rats. Despite the overall similarities in the mechanical values obtained between no drug rats and celecoxib treated rats, 3 of 6 fractured femurs from the celecoxib treated rats at 8 weeks post-fracture failed as non-unions during the mechanical testing procedure and the other 3 failed as mal-unions (Table 2). The relatively low normalized torsional rigidity (50%) and shear stress (22%) found for fractured femurs from the celecoxib treated rats at 6 weeks post-fracture indicates that the fracture site had not been bridged with bone (FIG. 2). Even though not statistically different from the no drug rat fractured femurs, the data obtained from the celecoxib treated rat fractured femurs parallels closely the patterns obtained from the femurs of the indomethacin treated rats. Together these observations suggest that, at the celecoxib dose and treatment regime used, fracture healing is delayed and to a lesser extent than that found for the rofecoxib treatment regime, inhibited.
[0081]A χ2 analysis was performed on visual inspection data obtained from the 8 week post-fracture femurs following mechanical testing (Table 2). The fractured femurs were considered to have failed as (a) unions if a spiral fracture developed through the diaphysis of the femur, (b) non-unions if the femur failed completely along the original fracture site, and (c) mal-unions if some new bone bridging of the fracture site was evident but that the femur still failed primarily along the original fracture site. The data from the no drug, celecoxib, and rofecoxib treatment groups were compared to that from the indomethacin treatment group. Our analysis indicates that no statistical difference exists between the no drug and the indomethacin treatment groups but that the celecoxib and rofecoxib treatment groups are significantly different from the indomethacin treatment group. Again, these observations strongly indicate that inhibition of COX-2 dramatically inhibits fracture healing.
[0082]No significant differences in the mechanical properties of the contralateral femurs were found between treatment groups. This indicates that the experimental treatment regimes did not alter the intrinsic properties of the rat bone by enhanced bone resorption or deposition at least for the time frame examined.
COX-2-Selective NSAID Treatment Alters Cartilage Formation During Fracture Healing
[0083]The radiographic and torsional mechanic testing analyses of the COX-2-selective NSAID treated rats demonstrated that these drugs dramatically inhibit fracture healing. However, hard callus formation appeared not to be impaired in the COX-2-selective NSAID treated rats. This suggests that COX-2-selective NSAIDs impair fracture healing in the soft callus where endochondral ossification occurs. Since radiography cannot assess the early stages of endochondral ossification in the soft callus, we undertook a histological analysis of fracture healing in the drug treated rats.
[0084]At 2 weeks post-fracture, gross abnormalities were present in the histology of the healing femur fractures of the indomethacin, celecoxib, or rofecoxib treated rats as compared to an untreated control rat (FIG. 3). In all four experimental groups, significant periosteal intramembraneous ossification was evident at the fracture site as expected from our radiographic data. Endochondral ossification also appeared to be proceeding normally in the no drug rat specimens. In contrast, the histological specimens from the indomethacin and COX-2-selective NSAID treated rats had abnormally formed cartilage elements within the callus. The positional extent of new bone formed in the callus of the NSAID treated rats also appeared to be abnormal in that it did not fully extend to the ends of the fractured bone. In the no drug rats, new bone in the callus extends to the very ends of the cortical bone fracture site. This is not so in the NSAID treated rats where this region of the callus is generally occupied by cartilage.
[0085]NSAID treatment grossly altered fracture callus morphology at 3 and 4 weeks post-fracture (FIG. 3). Fracture healing proceeded normally in the no drug rats with near bridging of the callus apparent by 4 weeks post-fracture in concurrence with our radiographic data (FIG. 1). However, healing was clearly delayed in the NSAID treated rats. The fracture calluses of the indomethacin treated rats were not bridged at 4 weeks but still appeared to be undergoing endochondral ossification based upon the presence of cartilage within the soft callus at 3 and 4 weeks post-fracture. In contrast, little or no cartilage was evident in the fracture calluses of the celecoxib or rofecoxib treated rats at 3 and 4 weeks post-fracture indicating that endochondral ossification had ceased. Additionally massive resorption of the woven bone in the hard callus of the NSAID treated rats appeared to leave a shell-like callus on the ends of the fractured bone.
[0086]Celecoxib and rofecoxib treatment often caused a massive bone resorption event at the distal ends of the fracture bones leaving what appear to be indentations into the hard callus (FIGS. 3 and 4). The magnitude of this bone resorption phase is indicated by the large number of osteoclasts that were found on the femur periosteal surface at the distal ends of the fractured bone near the apparent indentation (FIG. 4).
[0087]At 6 and 8 weeks post-fracture, the fractured femurs of the no drug rats appeared to be healing normally with active remodeling of the cortical bone ends and fracture callus. Fractured femurs from indomethacin treated rats also appeared to be healing at 6 and 8 weeks post-fracture with evident bridging and active remodeling. In contrast, no further healing was evident in the celecoxib or rofecoxib treated rat fractured femurs. The callus in the COX-2-selective NSAID treated rats was smaller at 6 and 8 weeks post-fracture but the fracture gap was still clearly evident and often filled with fibrous tissue. These observations are consistent with our radiographic and torsional mechanical testing data demonstrating that celecoxib and rofecoxib inhibit fracture healing.
Complications Associated with Use of COX-2-Selective NSAIDs
[0088]As can be seen in FIG. 5, pin slippage was a severe complication with as many as 30% of the rofecoxib treated rats having to be euthanized prior to the endpoint. Pin slippage is dislodgement of the intramedullary stainless steel rod used to stabilize the fracture and permit the rat to weight-bear on the fractured femur. Once fracture stability was lost, the rat was euthanized since the rat could no longer weight-bear on the femur and since the callus would be re-injured and thus alter healing. The etiology of the pin slippage is unknown. Animals with these complications were excluded. As such, final data may be skewed in favor of rats that had healed. Other complications included anesthesia death during weekly radiographs and infections that excluded specimens from further analysis. It was found using a χ2 analysis to compare each experimental group value to the no drug rat value that the pin slippage rate was significant for all NSAID treatment groups (P<0.0001) and that the infection rate for the rofecoxib treated rats was also significant (P<0.0001).
Normal Fracture Healing Fails in Cox2 Null Mice
[0089]Our data clearly demonstrate that COX-2-selective NSAIDs are detrimental to fracture healing. Unfortunately, these observations do not distinguish between a specific effect on COX-2 and a non-specific effect of the NSAIDs on fracture healing. Therefore to specifically address whether fracture healing requires Cox1 or Cox2 gene function, femur fracture healing was assessed in Cox1 and Cox2 knock-out mice (Langenbach et al. 1995; Morham et al. 1995). Using a modified method, closed femur fractures were produced in three female Cox1.sup.-/- (Cox1 knock-out) and three female Cox2.sup.-/- (Cox2 knock-out) mice. The animals were examined radiographically immediately post-fracture and then at 7, 10, 14, 21, 28, and 42 days post-fracture. Fracture healing appeared to proceed normally in the Cox1.sup.-/- mice relative to our previous observations in outbred and inbred strains of mice (Manigrasso and O'Connor, unpublished). In contrast, only a slight periosteal hard callus was detected in any of the Cox2.sup.-/- mice, which is indicative of healing failure. As can be seen in FIG. 6, the apparent difference in fracture callus size was most obvious at 2 weeks post-fracture when the Cox1.sup.-/- mice had formed a large fracture callus but little or no callus was evident in the Cox2.sup.-/- mice.
[0090]One mouse of each genotype was euthanized at 2 weeks post-fracture and the fractured femur examined histologically (FIG. 6). New bone and differentiating chondrocytes were abundant within the Cox1.sup.-/- callus indicating that COX-1 activity is not essential for fracture healing. In contrast, there was a plainly evident lack of new bone formation in the Cox2.sup.-/- fracture callus with only some apparent intramembraneous bone formation occurring at the edges of the callus (hard callus). Chondrocytes at different stages of differentiation were observed throughout the Cox2.sup.-/- soft fracture callus. However, the Cox2.sup.-/- chondrocytes failed to form a mineralized matrix as evident by the radiolucency and histological appearance of the soft callus. The amount of endochondral ossification at the hard callus-soft callus boundary appeared greatly reduced in the Cox2.sup.-/- specimen. In addition, neovascularization of the Cox2.sup.-/- fracture callus was not observed. These observations clearly indicate that normal fracture healing and endochondral ossification are stopped or dramatically reduced in the Cox2.sup.-/- mice and confirms our observations made in the COX-2-selective NSAID treated rats.
[0091]Our data indicate that COX-2 function is essential for fracture healing. In contrast, adult mice homozygous for targeted mutation of Cox2 appear to have normal skeletons. Together these observations demonstrate that fetal osteogenesis and fracture healing, though similar in many ways, are different and are probably initiated and maintained through different molecular mechanisms. Cox2 is the first gene to be identified that is specifically essential for fracture healing but not fetal osteogenesis. Targeted mutation of other mouse genes involved in prostaglandin synthesis and signaling should enable further analysis of the prostaglandin pathway(s) involved in fracture healing and skeletal biology in general.
[0092]One skilled in the art will readily appreciate that the present invention is well adapted to carry out the objects and obtain the ends and advantages mentioned as well as those inherent therein. The cyclooxygenase vectors along with the methods, procedures and treatments described herein are presently representative of preferred embodiments and are exemplary and not intended as limitations on the scope of the invention. Changes therein and other uses will occur to those skilled in the art which are encompassed within the spirit of the invention or defined by this scope with the claims.
[0093]It will be readily apparent to one skilled in the art that varying substitutions and modifications may be made to the invention disclosed herein without departing from the scope and spirit of the invention.
[0094]All patents and publications referenced herein are incorporated by reference to the same extent as if each individual publication was specifically and individually indicated to be incorporated by reference. The following references are likewise incorporated by reference in order to more fully describe the state of the art.
REFERENCES
[0095]Allen, H. L., A. Wase, and W. T. Bear. 1980. Indomethacin and aspirin: effect of nonsteroidal anti-inflammatory agents on the rate of fracture repair in the rat. Acta. Orthop. Scand. 51: 595-600. [0096]Altman, R. D., L. L. Latta, R. Keer, K. Renfree, F. J. Hornicek, and K. Banovac. 1995. Effect of nonsteroidal antiinflammatory drugs on fracture healing a laboratory study in rats. J. Orthop. Trauma 9: 392-400. [0097]Bell, G. H., D. P. Cuthbertson, and J. On. 1941. Strength and size of bone in relation to calcium intake. Journal of Physiology 100: 299-317. [0098]Bolander, M. E. 1992. Regulation of fracture repair by growth factors. Proc. Soc. Exp. Biol. Med. 200: 165-170. [0099]Bonnarens, F. and T. A. Einhorn. 1984. Production of a standard closed fracture in laboratory animal bone. J. Orthop. Res. 2: 97-101. [0100]Brighton, C. T., J. L. Schaffer, D. B. Shapiro, J. J. S. Tang, and C. C. Clark. 1991. Proliferation and macromolecular synthesis by rat calvarial bone cells grown in various oxygen tensions. J. Orthop. Res. 9: 847-854. [0101]Cohen, R. B., G. V. Hahn, J. A. Tabas, J. Peeper, C. L. Levitz, A. Sando, N. Sando, M. Zasloff, and F. S. Kaplan. 1993. The natural history of heterotopic ossification in patients who have fibrodysplasia ossificans progressive. J. Bone and Joint Surg. 75-A: 215-219. [0102]Davies, N. M., A. J. McLachlan, R. O. Day, and K. M. Williams. 2000. Clinical pharmacokinetics and pharmacodynamics of celecoxib: a selective cyclo-oxygenase-2 inhibitor. Clinical Pharmacokinetics 38: 225-242. [0103]Dekel, S., G. Lenthall, and M. J. O. Francis. 1981. Release of prostaglandins from bone and muscle after tibial fracture. J. Bone and Joint Surg. 63-B: 185-189. [0104]Depr, M., E. Ehrich, A. van Hecken, I. De Lepeleire, A. Dallob, P. Wong, A. Porras, B. J. Gertz, and P. J. De Schepper. 2000. Pharmacokinetics, COX-2 specificity, and tolerability of supratherapeutic doses of rofecoxib in humans European Journal of Pharmacology 56: 167-174. [0105]Einhorn, T. A. 1998. The cell and molecular biology of fracture healing. Clin Orthop: S7-21. [0106]Engesaeter, L. B., A. Ekeland, and N. Langeland. 1978. Methods for testing the mechanical properties of the rat femur. Acta. Orthop. Scand. 49: 512-518. [0107]Ferguson, C., E. Alpern, T. Miclau, and J. A. Helms. 1999. Does adult fracture repair recapitulate embryonic skeletal formation? Mech. Dev. 87: 57-66. [0108]Feyen, J. H., G. van der Wilt, P. Moonen, A. Di Bon, and P. J. Nijweide. 1984. Stimulation of arachidonic acid metabolism in primary cultures of osteoblast-like cells by hormones and drugs. Prostaglandins 28: 769-781. [0109]Halpin, R. A., L. A. Geer, K. E. Zhang, T. M. Marks, D. C. Dean, A. N. Jones, D. Melillo, G. Doss, and K. P. Vyas. 2000. The absorption, distribution, metabolism and excretion of rofecoxib, a potent and selective cyclooxygenase-2 inhibitor, in rats and dogs. Drug Metabolism and Disposition 28: 1244-1254. [0110]Hsu, A.-L., T.-T. Ching, D.-S. Wang, X. Song, V. M. Rangnekar, and C.-S. Chen. 2000. The cyclooxygenase-2 inhibitor celecoxib induces apoptosis by blocking Akt activation in human prostate cancer cells independently of Bc1-2. J. Biol. Chem. 275: 11397-11403. [0111]Jones, M. K., H. Wang, B. M. Peskar, E. Levin, R. M. Itani, I. J. Sarfeh, and A. S. Tamawski. 1999. Inhibition of angiogenesis by nonsteroidal anti-inflammatory drugs: insight into the mechanisms and implications for cancer growth and ulcer healing. Nat. Med. 5: 1418-1423. [0112]Kaplan, F. S., J. A. Tabas, F. H. Gannon, G. Finkel, G. V. Hahn, and M. A. Zasloff. 1993. The histopathology of fibrodysplasia ossificans progressiva: an endochondral process. J. Bone and Joint Surg. 75-A: 220-230. [0113]Kawaguchi, H., C. C. Pilbeam, J. R. Harrison, and L. G. Raisz. 1995. The role of prostaglandins in the regulation of bone metabolism. Clin. Orth. Rel. Res. 313: 36-46. [0114]Klein, D. C. and L. G. Raisz. 1970. Prostaglandins: stimulation of bone resorption in tissue culture. Endocrinol 86: 1436-1440. [0115]Klein-Nulend, J., E. H. Burger, C. M. Semeins, L. G. Raisz, and C. C. Pilbeam. 1997. Pulsating fluid flow stimulates prostaglandin release and inducible prostaglandin G/H synthase mRNA expression in primary mouse bone cells. J. Bone Miner. Res. 12: 45-51. [0116]Langenbach, R., S. G. Morham, H. F. Tiano, C. D. Loftin, B. I. Ghanayem, P. C. Chulada, J. F. Mahler, C. A. Lee, E. H. Goulding, K. D. Kluckman, H. S. Kim, and O, Smithies. 1995. Prostaglandin synthase 1 gene disruption in mice reduces arachidonic acid-induced inflammation and indomethacin-induced gastric ulceration. Cell 83: 483-492. [0117]Maniatopoulos, C., A. Rodriguez, D. A. Deporter, and A. H. Melcher. 1986. An improved method for preparing histological sections of metallic implants. International Journal of Oral and Maxillofacial Implants 1: 31-37. [0118]Masferrer, J. L., B. S. Zweifel, P. T. Manning, S. D. Hauser, K. M. Leahy, W. G. Smith, P. C. Isakson, and K. Seibert. 1994. Selective inhibition of inducible cyclooxygenase 2 in vivo is antiinflammatory and nonulcerogenic. Proc. Natl. Acad. Sci. USA 91: 3228-3232. [0119]Moore, K. D., K. Goss, and J. O. Anglen. 1998. Indomethacin versus radiation therapy for prophylaxis against heterotopic ossification in acetabular fractures. J. Bone and Joint Surg. 80-B: 259-263. [0120]Morham, S. G., R. Langenbach, C. D. Loftin, H. F. Tiano, N. Vouloumanos, J. C. Jennette, J. F. Mahler, K. D. Kluckman, A. Ledford, C. A. Lee, and O, Smithies. 1995. Prostaglandin synthase 2 gene disruption causes severe renal pathology in the mouse. Cell 83: 473-482. [0121]Muscar, M. N., W. McKnight, S. Asfaha, and J. L. Wallace. 2000. Wound collagen deposition in rats: effects of an NO-NSAID and a selective COX-2 inhibitor. British Journal of Pharmacology 129: 681-686. [0122]Needleman, P., J. Turk, B. A. Jakschik, A. R. Morrison, and J. B. Lefkowith. 1986. Arachidonic acid metabolism. Ann. Rev. Biochem. 55: 69-102. [0123]Nicholls, P. J., E. Berg, F. E. Bliven, Jr., and J. M. Kling. 1979. X-ray diagnosis of healing fractures in rabbits. Clin. Orth. Rel. Res. 142: 234-236. [0124]O'Banion, M. K., V. D. Winn, and D. A. Young. 1992. cDNA cloning and functional activity of a glucocorticoid-regulated inflammatory cyclooxygenase. Proc. Natl. Acad. Sci. USA 89: 4888-4892. [0125]Okada, Y., J. A. Lorenzo, A. M. Freeman, M. Tomita, S. G. Morham, L. G. Raisz, and C. C. Pilbeam. 2000. Prostaglandin G/H synthase-2 is required for maximal formation of osteoclast-like cells in culture. J. Clin. Invest. 105: 823-832. [0126]O'Sullivan, M. G., F. H. Chilton, E. M. Huggins, Jr., and C. E. McCall. 1992a. Lipopolysaccharide priming of alveolar macrophages for enhanced synthesis of prostanoids involves induction of a novel prostaglandin H synthase. J. Biol. Chem. 267: 14547-14550. [0127]O'Sullivan, M. G., E. M. Huggins, Jr., E. A. Meade, D. L. DeWitt, and C. E. McCall. 1992b. Lipopolysaccharide induces prostaglandin H synthase-2 in alveolar macrophages. Biochem. Biophys. Res. Comm. 187: 1123-1127. [0128]Panjabi, M. M., R. W. Lindsey, S. D. Walter, and A. A. White, 3rd. 1989. The clinician's ability to evaluate the strength of healing fractures from plain radiographs. J. Orthop. Trauma 3: 29-32. [0129]Paulson, S. K., J. Y. Zhang, A. P. Breau, J. D. Hribar, N. W. K. Liu, S. M. Jessen, Y. M. Lawal, J. N. Cogburn, C. J. Gresk, C. S. Markos, T. J. Maziasz, G. L. Schoenhard, and E. G. Burton. 2000. Pharmacokinetics, tissue distribution, metabolism, and excretion of celecoxib in rats. Drug Metabolism and Disposition 28: 514-521. [0130]Popov, E. P. 1968. Introduction to mechanics of solids. Prentice-Hall, Inc., Englewood Cliffs, N.J. [0131]Pritchett, J. W. 1995. Ketorolac prophylaxis against heterotopic ossification after hip replacement. Clin. Orth. Rel. Res. 314: 162-165. [0132]Raskin, J. B. 1999. Gastrointestinal effects of nonsteroidal anti-inflammatory therapy. Am. J. Med. 106 (5B): 3S-12S. [0133]Riendeau, D., M. D. Percival, C. Brideau, S. Charleson, D. Dube, D. Ethier, J.-P. Falgueyret, R. W. Friesen, R. Gordon, G. Greig, J. Guay, J. Mancini, M. Ouellet, E. Wong, L. Xu, S. Boyce, D. Visco, Y. Girard, P. Prasit, R. Zamboni, I. W. Rodger, M. Gresser, A. W. Ford-Hutchinson, R. N. Young, and C.-C. Chan. 2001. Etoricoxib (MK-0663): preclinical profile and comparison with other agents that selectively inhibit cyclooxygenase-2. Journal of Pharmacology and Experimental Therapeutics 296: 558-566. [0134]Ro, J., N. Langeland, and J. Sander. 1978. Effect of indomethacin on collagen metabolism of rat fracture callus in vitro. Acta. Orthop. Scand. 49: 323-328. [0135]Ro, J., E. Sudmann, and P. F. Marton. 1976. Effect of indomethacin on fracture healing in rats. Acta. Orthop. Scand. 47: 588-599. [0136]Rossi, A., P. Kapahi, G. Natoli, T. Takahashi, Y. Chen, M. Karin, and M. G. Santoro. 2000. Anti-inflammatory cylopentenone prostaglandins are direct inhibitors of IκB kinase. Nature 403: 103-108. [0137]Shigeta, J.-I., S. Takahashi, and S. Okabe. 1998. Role of cyclooxygenase-2 in the healing of gastric ulcers in rats. Journal of Pharmacology and Experimental Therapeutics 286: 1383-1390. [0138]Topper, J. N., J. Cai, D. Falb, and M. A. Gimbrone, Jr. 1996. Identification of vascular endothelial genes differentially responsive to fluid mechanical stimuli: cyclooxygenase-2, manganese superoxide dismutase, and endothelial cell nitric oxide synthase are selectively up-regulated by steady laminar shear stress. Proc. Natl. Acad. Sci. USA 93: 10417-10422. [0139]Vane, J. R., Y. S. Bakhle, and R. M. Botting. 1998. Cyclooxygenase 1 and 2. Annual Review of Pharmacology and Toxicology 38: 97-120. [0140]Vortkamp, A., S. Pathi, G. M. Peretti, E. M. Caruso, D. J. Zaleske, and C. J. Tabin. 1998. Recapitulation of signals regulating embryonic bone formation during postnatal growth and fracture repair. Mech. Dev. 71: 65-76. [0141]Wadleigh, D. J. and H. R. Herschman. 1999. Transcriptional regulation of the cyclooxygenase-2 gene by diverse ligands in murine osteoblasts. Biochem. Biophys. Res. Comm. 264: 865-870. [0142]Wallace, J. L., A. Bak, W. McKnight, S. Asfaha, K. A. Sharkey, and W. K. MacNaughton. 1998. Cyclooxygenase 1 contributes to inflammatory responses in rats and mice: implications for gastrointestinal toxicity. Gastroenterology 115: 101-109.
[0143]Warner, T. D., F. Giuliano, I. Vojnovic, A. Bukasa, J. A. Mitchell, and J. R. Vane. 1999. Nonsteroid drug selectivities for cyclo-oxygenase-1 rather than cyclo-oxygenase-2 are associated with human gastrointestinal toxicity: a full in vitro analysis. Proc. Natl. Acad. Sci. USA 96: 7563-7568. [0144]Whelton, A. 1999. Nephrotoxicity of nonsteroidal anti-inflammatory drugs: physiological foundations and clinical implications. Am. J. Med. 106 (5B): 13S-24S. [0145]White, A. A., III, M. M. Panjabi, and W. O. Southwick. 1977. The four biomechanical stages of fracture repair. J. Bone and Joint Surg. 59-A: 188-192. [0146]Xie, W., J. G. Chipman, D. L. Robertson, R. L. Erikson, and D. L. Simmons. 1991. Expression of a mitogen-responsive gene encoding prostaglandin synthase is regulated by mRNA splicing. Proc. Natl. Acad. Sci. USA 88: 2692-2696. [0147]Zhang, X., S. G. Morham, R. Langenbach, R. B. Baggs, and D. A. Young. 2000. Lack of cyclooxygenase-2 inhibits growth of teratocarcinomas in mice. Exp. Cell Res. 254: 232-240.
TABLE-US-00001 [0147]TABLE 1 Torsional Mechanical Testing Data Fractured Femur Data (not normalized to contralateral control femur) All Contralateral No Drug Indomethacin Control Femurs 4 wks 6 wks 8 wks 4 wks 6 wks 8 wks Peak Torque (Tmax) in Nmm Mean 716 452 430 643 417 357 469 CV 28 21 29 28 52 37 56 Significance -- -- -- -- 0.69 0.36 0.16 Torsional Rigidity [(Tmax L )/φ] Mean 83,050 27,200 66,068 56,506 21,913 28,658 57,630 CV 35 48 17 20 46 57 55 Significance -- -- -- -- 0.43 <0.01 0.93 Shear Stress [(Tmax φ)/J] mean 145 33 35 56 25 22 47 CV 38 42 34 27 85 47 66 Significance -- -- -- -- 0.42 0.08 0.48 Shear Modulus (GPa) Mean 6.9 0.4 1.5 1.3 0.3 0.4 1.8 CV 54 58 71 38 78 73 99 Significance -- -- -- -- 0.33 0.08 0.46 Sample Size 81 6 5 7 8 7 8 Fractured Femur Data (not normalized to contralateral control femur) All Contralateral Celecoxib Rofecoxib Control Femurs 4 wks 6 wks 8 wks 4 wks 6 wks 8 wks Peak Torque (Tmax) in Nmm Mean 302 567 620 250 383 338 CV 44 34 49 53 33 36 Significance 0.03 0.15 0.88 0.01 0.54 0.01 Torsional Rigidity [(Tmax L )/φ] Mean 13,866 43,189 59,256 12,338 21,231 25,026 CV 53 77 22 59 51 69 Significance 0.06 0.11 0.69 0.04 <0.01 0.01 Shear Stress [(Tmax φ)/J] mean 25 44 67 15 24 25 CV 73 73 41 58 41 48 Significance 0.37 0.47 0.40 0.03 0.13 <0.01 Shear Modulus (GPa) Mean 0.3 0.9 1.7 0.2 0.3 0.5 CV 77 116 27 49 56 9.1 Significance 0.26 0.40 0.16 0.04 0.07 0.01 Sample Size 8 8 6 6 7 5 CV: Coefficient of Variation (standard deviation as a percentage). Significance: Comparisons were made using a standard two-tailed T-test between the no drug fractured femur values and the experimental fractured femur values within a time point. These comparisons were not made between data normalized with the contralateral control femur as shown in FIG. 3.
TABLE-US-00002 TABLE 2 Frequency of Unions, Mal-Unions, and Non-Unions in the Mechanically Tested Fractured Femurs at 8 weeks Post-Fracture Union Mal-Union Non-Union χ2 (Stage IV)* (Stage III) (Stage I/II) P-value** No Drugs 7 0 0 0.338 Indomethacin 6 1 1 -- Celecoxib 0 3 3 <0.001 Rofecoxib 0 1 4 <0.001 *Classification scheme as described by White et al. (1977) **Comparisons were made to the indomethacin treatment group data.
TABLE-US-00003 TABLE 3 I. HUMAN COX-1 cDNA SEQUENCE (SEQ. ID. NO. 1) GCGCCATGAGCCGGAGTCTCTTGCTCCGGTTCTTGCTGTTCCTGCTCCTG CTCCCGCCGCTCCCCGTCCTGCTCGCGGACCCAGGGGCGCCCACGCCAGT GAATCCCTGTTGTTACTATCCATGCCAGCACCAGGGCATCTGTGTCCGCT TCGGCCTTGACCGCTACCAGTGTGACTGCACCCGCACGGGCTATTCCGGC CCCAACTGCACCATCCCTGGCCTGTGGACCTGGCTCCGGAATTCACTGCG GCCCAGCCCCTCTTTCACCCACTTCCTGCTCACTCACGGGCGCTGGTTCT GGGAGTTTGTCAATGCCACCTTCATCCGAGAGATGCTCATGCGCCTGGTA CTCACAGTGCGCTCCAACCTTATCCCCAGTCCCCCCACCTACAACTCAGC ACATGACTACATCAGCTGGGAGTCTTTCTCCAACGTGAGCTATTACACTC GTATTCTGCCCTCTGTGCCTAAAGATTGCCCCACACCCATGGGAACCAAA GGGAAGAAGCAGTTGCCAGATGCCCAGCTCCTGGCCCGCCGCTTCCTGCT CAGGAGGAAGTTCATACCTGACCCCCAAGGCACCAACCTCATGTTTGCCT TCTTTGCACAACACTTCACCCACCAGTTCTTCAAAACTTCTGGCAAGATG GGTCCTGGCTTCACCAAGGCCTTGGGCCATGGGGTAGACCTCGGCCACAT TTATGGAGACAATCTGGAGCGTCAGTATCAACTGCGGCTCTTTAAGGATG GGAAACTCAAGTACCAGGTGCTGGATGGAGAAATGTACCCGCCCTCGGTA GAAGAGGCGCCTGTGTTGATGCACTACCCCCGAGGCATCCCGCCCCAGAG CCAGATGGCTGTGGGCCAGGAGGTGTTTGGGCTGCTTCCTGGGCTCATGC TGTATGCCACGCTCTGGCTACGTGAGCACAACCGTGTGTGTGACCTGCTG AAGGCTGAGCACCCCACCTGGGGCGATGAGCAGCTTTTCCAGACGACCCG CCTCATCCTCATAGGGGAGACCATCAAGATTGTCATCGAGGAGTACGTGC AGCAGCTGAGTGGCTATTTCCTGCAGCTGAAATTTGACCCAGAGCTGCTG TTCGGTGTCCAGTTCCAATACCGCAACCGCATTGCCATGGAGTTCAACCA TCTCTACCACTGGCACCCCCTCATGCCTGACTCCTTCAAGGTGGGCTCCC AGGAGTACAGCTACGAGCAGTTCTTGTTCAACACCTCCATGTTGGTGGAC TATGGGGTTGAGGCCCTGGTGGATGCCTTCTCTCGCCAGATTGCTGGCCG GATCGGTGGGGGCAGGAACATGGACCACCACATCCTGCATGTGGCTGTGG ATGTCATCAGGGAGTCTCGGGAGATGCGGCTGCAGCCCTTCAATGAGTAC CGCAAGAGGTTTGGCATGAAACCCTACACCTCCTTCCAGGAGCTCGTAGG AGAGAAGGAGATGGCAGCAGAGTTGGAGGAATTGTATGGAGACATTGATG CGTTGGAGTTCTACCCTGGACTGCTTCTTGAAAAGTGCCATCCAAACTCT ATCTTTGGGGAGAGTATGATAGAGATTGGGGCTCCCTTTTCCCTCAAGGG TCTCCTAGGGAATCCCATCTGTTCTCCGGAGTACTGGAAGCCGAGCACAT TTGGCGGCGAGGTGGGCTTTAACATTGTCAAGACGGCCACACTGAAGAAG CTGGTCTGCCTCAACACCAAGACCTGTCCCTACGTTTCCTTCCGTGTGCC GGATGCCAGTCAGGATGATGGGCCTGCTGTGGAGCGACCATCCACAGAGC TCTGAGGGGCAGGAAAGCAGCATTCTGGAGGGGAGAGCTTTGTGCTTGTC ATTCCAGAGTGCTGAGGCCAGGGCTGATGGTCTTAAATGCTCATTTTCTG GTTTGGCATGGTGAGTGTTGGGGTTGACATTTAGAACTTTAAGTCTCACC CATTATCTGGAATATTGTGATTCTGTTTATTCTTCCAGAATGCTGAACTC CTTGTTAGCCCTTCAGATTGTTAGGAGTGGTTCTCATTTGGTCTGCCAGA ATACTGGGTTCTTAGTTGACAACCTAGAATGTCAGATTTCTGGTTGATTT GTAACACAGTCATTCTAGGATGTGGAGCTACTGATGAAATCTGCTAGAAA GTTAGGGGGTTCTTATTTTGCATTCCAGAATCTTGACTTTCTGATTGGTG ATTCAAAGTGTTGTGTTCCCTGGCTGATGATCCAGAACAGTGGCTCGTAT CCCAAATCTGTCAGCATCTGGCTGTCTAGAATGTGGATTTGATTCATTTT CCTGTTCAGTGAGATATCATAGAGACGGAGATCCTAAGGTCCAACAAGAA TGCATTCCCTGAATCTGTGCCTGCACTGAGAGGGCAAGGAAGTGGGGTGT TCTTCTTGGGACCCCCACTAAGACCCTGGTCTGAGGATGTAGAGAGAACA GGTGGGCTGTATTCACGCCATTGGTTGGAAGCTACCAGAGCTCTATCCCC ATCCAGGTCTTGACTCATGGCAGCTGTTTCTCATGAAGCTAATAAAATTC GCCC II. HUMAN COX-2 cDNA SEQUENCE (SEQ. ID. NO. 2) CAATTGTCATACGACTTGCAGTGAGCGTCAGGAGCACGTCCAGGAACTCC TCAGCAGCGCCTCCTTCAGCTCCACAGCCAGACGCCCTCAGACAGCAAAG CCTACCCCCGCGCCGCGCCCTGCCCGCCGCTCGGATGCTCGCCCGCGCCC TGCTGCTGTGCGCGGTCCTGGCGCTCAGCCATACAGCAAATCCTTGCTGT TCCCACCCATGTCAAAACCGAGGTGTATGTATGAGTGTGGGATTTGACCA GTATAAGTGCGATTGTACCCGGACAGGATTCTATGGAGAAAACTGCTCAA CACCGGAATTTTTGACAAGAATAAAATTATTTCTGAAACCCACTCCAAAC ACAGTGCACTACATACTTACCCACTTCAAGGGATTTTGGAACGTTGTGAA TAACATTCCCTTCCTTCGAAATGCAATTATGAGTTATGTCTTGACATCCA GATCACATTTGATTGACAGTCCACCAACTTACAATGCTGACTATGGCTAC AAAAGCTGGGAAGCCTTCTCTAACCTCTCCTATTATACTAGAGCCCTTCC TCCTGTGCCTGATGATTGCCCGACTCCCTTGGGTGTCAAAGGTAAAAAGC AGCTTCCTGATTCAAATGAGATTGTGGAAAAATTGCTTCTAAGAAGAAAG TTCATCCCTGATCCCCAGGGCTCAAACATGATGTTTGCATTCTTTGCCCA GCACTTCACGCATCAGTTTTTCAAGACAGATCATAAGCGAGGGCCAGCTT TCACCAACGGGCTGGGCCATGGGGTGGACTTAAATCATATTTACGGTGAA ACTCTGGCTAGACAGCGTAAACTGCGCCTTTTCAAGGATGGAAAAATGAA ATATCAGATAATTGATGGAGAGATGTATCCTCCCACAGTCAAAGATACTC AGGCAGAGATGATCTACCCTCCTCAAGTCCCTGAGCATCTACGGTTTGCT GTGGGGCAGGAGGTCTTTGGTCTGGTGCCTGGTCTGATGATGTATGCCAC AATCTGGCTGCGGGAACACAACAGAGTATGCGATGTGCTTAAACAGGAGC ATCCTGAATGGGGTGATGAGCAGTTGTTCCAGACAAGCAGGCTAATACTG ATAGGAGAGACTATTAAGATTGTGATTGAAGATTATGTGCAACACTTGAG TGGCTATCACTTCAAACTGAAATTTGACCCAGAACTACTTTTCAACAAAC AATTCCAGTACCAAAATCGTATTGCTGCTGAATTTAACACCCTCTATCAC TGGCATCCCCTTCTGCCTGACACCTTTCAAATTCATGACCAGAAATACAA CTATCAACAGTTTATCTACAACAACTCTATATTGCTGGAACATGGAATTA CCCAGTTTGTTGAATCATTCACCAGGCAAATTGCTGGCAGGGTTGCTGGT GGTAGGAATGTTCCACCCGCAGTACAGAAAGTATCACAGGCTTCCATTGA CCAGAGCAGGCAGATGAAATACCAGTCTTTTAATGAGTACCGCAAACGCT TTATGCTGAAGCCCTATGAATCATTTGAAGAACTTACAGGAGAAAAGGAA ATGTCTGCAGAGTTGGAAGCACTCTATGGTGACATCGATGCTGTGGAGCT GTATCCTGCCCTTCTGGTAGAAAAGCCTCGGCCAGATGCCATCTTTGGTG AAACCATGGTAGAAGTTGGAGCACCATTCTCCTTGAAAGGACTTATGGGT AATGTTATATGTTCTCCTGCCTACTGGAAGCCAAGCACTTTTGGTGGAGA AGTGGGTTTTCAAATCATCAACACTGCCTCAATTCAGTCTCTCATCTGCA ATAACGTGAAGGGCTGTCCCTTTACTTCATTCAGTGTTCCAGATCCAGAG CTCATTAAAACAGTCACCATCAATGCAAGTTCTTCCCGCTCCGGACTAGA TGATATCAATCCCACAGTACTACTAAAAGAACGTTCGACTGAACTGTAGA AGTCTAATGATCATATTTATTTATTTATATGAACCATGTCTATTAATTTA ATTATTTAATAATATTTATATTAAACTCCTTATGTTACTTAACATCTTCT GTAACAGAAGTCAGTACTCCTGTTGCGGAGAAAGGAGTCATACTTGTGAA GACTTTTATGTCACTACTCTAAAGATTTTGCTGTTGCTGTTAAGTTTGGA AAACAGTTTTTATTCTGTTTTATAAACCAGAGAGAAATGAGTTTTGACGT CTTTTTACTTGAATTACTATCACAAGATGGCAAAATGCTGAAAGTTTTTA CACTGTCGATGTTTCCAATGCATCTTCCATGATGCATTAGAAGTAACTAA TGTTTGAAATTTTAAAGTACTTTTGGTTATTTTTCTGTCATCAAACAAAA ACAGGTATCAGTGCATTATTAAATGAATATTTAAATTAGACATTACCAGT AATTTCATGTCTACTTTTTAAAATCAGCAATGAAACAATAATTTGAAATT TCTAAATTCATAGGGTAGAATCACCTGTAAAAGCTTGTTTGATTTCTTAA AGTTATTAAACTTGTACATATACCAAAAAGAAGCTGTCTTGGATTTAAAT CTGTAAAATCAGATGAAATTTTACTACAATTGCTTGTTAAAATATTTTAT AAGTGATGTTCCTTTTTCACCAAGAGTATAAACCTTTTTAGTGTGACTGT TAAAACTTCCTTTTAAATCAAAATGCCAAATTTATTAAGGTGGTGGAGCC ACTGCAGTGTTATCTCAAAATAAGAATATTTTGTTGAGATATTCCAGAAT TTGTTTATATGGCTGGTAACATGTAAAATCTATATCAGCAAAAGGGTCTA CCTTTAAAATAAGCAATAACAAAGAAGAAAACCAAATTATTGTTCAAATT TAGGTTTAAACTTTTGAAGCAAACTTTTTTTTATCCTTGTGCACTGCAGG CCTGGTACTCAGATTTTGCTATGAGGTTAATGAAGTACCAAGCTGTGCTT GAATAACGATATGTTTTCTCAGATTTTCTGTTGTACAGTTTAATTTAGCA GTCCATATCACATTGCAAAAGTAGCAATGACCTCATAAAATACCTCTTCA AAATGCTTAAATTCATTTCACACATTAATTTTATCTCAGTCTTGAAGCCA ATTCAGTAGGTGCATTGGAATCAAGCCTGGCTACCTGCATGCTGTTCCTT TTCTTTTCTTCTTTTAGCCATTTTGCTAAGAGACACAGTCTTCTCATCAC TTCGTTTCTCCTATTTTGTTTTACTAGTTTTAAGATCAGAGTTCACTTTC TTTGGACTCTGCCTATATTTTCTTACCTGAACTTTTGCAAGTTTTCAGGT AAACCTCAGCTCAGGACTGCTATTTAGCTCCTCTTAAGAAGATTAAAAGA GAAAAAAAAAGGCCCTTTTAAAAATAGTATACACTTATTTTAAGTGAAAA GCAGAGAATTTTATTTATAGCTAATTTTAGCTATCTGTAACCAAGATGGA TGCAAAGAGGCTAGTGCCTCAGAGAGAACGCCCTTTCTTATTTAAAAACA AAACCAAATGATATCTAAGTAGTTCTCAGCAATAATAATAATGACGATAA
TACTTCTTTTCCACATCTCATTGTCACTGACATTTAATGGTACTGTATAT TACTTAATTTATTGAAGATTATTATTTATGTCTTATTAGGACACTATGGT TATAAACTGTGTTTAAGCCTACAATCATTGATTTTTTTTTGTTATGTCAC AATCAGTATATTTTCTTTGGGGTTACCTCTCTGAATATTATGTAAACAAT CCAAAGAAATGATTGTATTAAGATTTGTGAATAAATTTTTAGAAATCTGA TTGGCATATTGAGATATTTAAGGTTGAATGTTTGTCCTTAGGATAGGCCT ATGTGCTAGCCCACAAAGAATATTGTCTCATTAGCCTGAATGTGCCATAA GACTGACCTTTTAAAATGTTTTGAGGGATCTGTGGATGCTTCGTTAATTT GTTCAGCCACAATTTATTGAGAAAATATTCTGTGTCAAGCACTGTGGGTT TTAATATTTTTAAATCAAACGCTGATTACAGATAATAGTATTTATATAAA TAATTGAAAAAATTTTCTTTTGGGAAGAGGGAGAAAATGAAATAAATATC ATTAAAGATAACTCAGGAGAATCTTCTTTACAATTTTACGTTTAGAATGT TTAAGGTTAAGAAAGAAATAGTCAATATGCTTGTATAAAACACTGTTCAC TGTTTTTTTTAAAAAAAAAACTTGATTTGTTATTAACATTGATCTGCTGA CAAAACCTGGGAATTTGGGTTGTGTATGCGAATGTTTCAGTGCCTCAGAC AAATGTGTATTTAACTTATGTAAAAGATAAGTCTGGAAATAAATGTCTGT TTATTTTTGTACTATTTA III. HUMAN COX-1 AMINO ACID SEQUENCE (SEQ. ID. NO. 3) MSRSLLLRFLLFLLLLPPLPVLLADPGAPTPVNPCCYYPCQHQGICVRFG LDRYQCDCTRTGYSGPNCTIPGLWTWLRNSLRPSPSFTHFLLTFGRWFWE FVNATFIREMLMRLVLTVRSNLIPSPPTYNSAHDYISWESFSNVSYYTRI LPSVPKDCPTPMGTKGKKQLPDAQLLARRFLLRRKFIPDPQGTNLMFAFF AQHFTHQFFKTSGKMGPGFTKALGHGVDLGHIYGDNLERQYQLRLFKDGK LKYQVLDGEMYPPSVEEAPVLMHYPRGIPPQSQMAVGQEVFGLLPGLMLY ATLWLREHNRVCDLLKAEHPTWGDEQLFQTTRLILIGETIKIVIEEYVQQ LSGYFLQLKFDPELLFGVQFQYRNRIAMEFNHLYHWHPLMPDSFKVGSQE YSYEQFLFNTSMLVDYGVEALVDAFSRQIAGRIGGGRNMDHHILHVAVDV IRESREMRLQPFNEYRKRFGMKPYTSFQELVGEKEMAAELEELYGDIDAL EFYPGLLLEKCHPNSIFGESMIEIGAPFSLKGLLGNPICSPEYWKPSTFG GEVGFNIVKTATLKKLVCLNTKTCPYVSFRVPDASQDDGPAVERPSTEL IV. HUMAN COX-2 AMINO ACID SEQUENCE (SEQ. ID. NO. 4) MLARALLLCAVLALSHTANPCCSHPCQNRGVCMSVGFDQYKCDCTRTGFY GENCSTPEFLTRIKLFLKPTPNTVHYILTHFKGFWNVVNNIPFLRNAIMS YVLTSRSHLIDSPPTYNADYGYKSWEAFSNLSYYTRALPPVPDDCPTPLG VKGKKQLPDSNEIVEKLLLRRKFIPDPQGSNMMFAFFAQHFTHQFFKTDH KRGPAFTNGLGHGVDLNHIYGETLARQRKLRLFKDGKMKYQIIDGEMYPP TVKDTQAEMIYPPQVPEHLRFAVGQEVFGLVPGLMMYATIWLREHNRVCD VLKQEHPEWGDEQLFQTSRLILIGETIKIVIEDYVQHLSGYHFKLKFDPE LLFNKQFQYQNRIAAEFNTLYHWHPLLPDTFQIHDQKYNYQQFIYNNSIL LEHGITQFVESFTRQIAGRVAGGRNVPPAVQKVSQASIDQSRQMKYQSFN EYRKRFMLKPYESFEELTGEKEMSAELEALYGDIDAVELYPALLVEKPRP DAIFGETMVEVGAPFSLKGLMGNVICSPAYWKPSTFGGEVGFQIINTASI QSLICNNVKGCPFTSFSVPDPELIKTVTINASSSRSGLDDINPTVLLKER STEL
TABLE-US-00004 TABLE 4 I. MOUSE COX-1 cDNA SEQUENCE (SEQ. ID. NO. 5) GCCGTTGGCATTGCACATCCATCCACTCCCAGAGTCATGAGTCGAAGGAG TCTCTCGCTCTGGTTTCCCCTGCTGCTGCTCCTGCTGCTGCCGCCGACAC CCTCGGTCCTGCTCGCAGATCCTGGGGTGCCCTCACCAGTCAATCCCTGT TGTTACTATCCGTGCCAGAACCAGGGTGTCTGTGTCCGCTTTGGCCTCGA CAACTACCAGTGTGATTGTACTCGCACGGGCTACTCAGGCCCCAACTGTA CCATCCCTGAGATCTGGACCTGGCTTCGGAATTCTCTGCGGCCCAGCCCC TCGTTCACCCATTTCCTGCTGACACATGGATACTGGCTCTGGGAATTTGT GAATGCCACCTTCATCCGAGAAGTACTCATGCGCCTGGTACTCACAGTGC GGTCCAACCTTATCCCCAGCCCTCCGACCTACAACTCAGCGCATGACTAC ATCAGCTGGGAGTCCTTCTCCAATGTGAGCTACTATACTCGCATTCTGCC CTCTGTACCCAAAGACTGCCCCACACCCATGGGGACCAAAGGGAAGAAAC AGTTACCAGATGTTCAGCTTCTGGCCCAACAGCTGCTGCTGAGAAGGGAG TTCATTCCTGCCCCCCAGGGCACCAACATCCTGTTTGCCTTCTTTGCACA ACACTTCACCCACCAGTTCTTCAAGACCTCTGGAAAGATGGGTCCTGGCT TTACCAAGGCCTTAGGCCACGGGGTAGACCTTGGCCACATTTATGGAGAT AATCTGGAACGACAGTATCACCTGCGGCTCTTCAAGGATGGGAAACTTAA GTACCAGGTGCTGGACGGAGAGGTGTACCCACCTTCCGTGGAACAGGCGT CCGTGTTGATGCGCTACCCACCAGGTGTCCCGCCTGAAAGGCAGATGGCT GTGGGCCAGGAGGTGTTTGGGTTGCTTCCGGGGCTGATGCTCTTCTCCAC GATCTGGCTTCGTGAACATAACCGCGTGTGCGACCTGCTGAAGGAGGAGC ATCCCACGTGGGATGATGAGCAGCTCTTCCAGACCACTCGCCTCATCCTT ATAGGAGAAACCATCAAAATTGTCATTGAGGAGTATGTGCAGCACTTGAG TGGCTATTTCCTGCAGCTCAAGTTTGACCCGGAGCTGCTGTTCCGAGCCC AGTTCCAATATCGAAACCGCATCGCCATGGAATTTAACCATCTCTATCAC TGGCATCCACTCATGCCCAACTCCTTCCAAGTGGGCTCACAAGAGTACAG CTACGAGCAGTTTTTATTTAACACTTCTATGCTGGTGGACTATGGGGTTG AGGCACTGGTGGATGCCTTCTCTCGCCAGAGGGCTGGCCGGATTGGTGGA GGTAGGAACTTTGACTATCATGTTCTGCATGTGGCTGTGGATGTCATCAA GGAGTCCCGAGAGATGCGCCTACAGCCCTTCAATGAATACCGAAAGAGGT TTGGCTTGAAGCCTTACACCTCTTTCCAGGAGCTCACAGGAGAGAAGGAG ATGGCTGCTGAGTTGGAGGAGCTGTACGGTGACATCGATGCTTTAGAGTT CTACCCGGGGTTGCTGCTGGAGAAGTGCCAGCCCAACTCCATCTTTGGAG AAAGTATGATAGAGATGGGGGCTCCCTTTTCCCTCAAGGGCCTCCTAGGG AATCCCATCTGTTCCCCAGAGTACTGGAAACCCAGCACGTTCGGTGGTGA CGTGGGCTTCAACCTTGTCAACACAGCCTCACTGAAGAAACTGGTCTGCC TCAACACCAAGACCTGCCCCTATGTTTCCTTCCGTGTGCCAGATTACCCT GGAGATGACGGGTCTGTCTTAGTGAGACGCTCCACTGAGCTCTGAGGGAG CTGGAAAGCAGCCTCTGGAGGGAGGAGTTTTGTTCCTGATGAAGACAAGT CCTTGATGTGGGTTTTCGTGGCTTGGCATTGTGAGAGCTGATGCTCACAT TTGAAACTTTGGGTCTTACCCTTGCCTAGAAAATTGTGATTTTGCCACTT TCGGATGTTGAATTCTTTGTTAACTAAGAAAGTTAGAAGTGGTTTTGTCT GCCTCCTCAGAACTTGGCTCTTTGTTGGCAACTCAGAAAGTCAGATTTCT GGTTGATTTGGAATATAGGCTTAAAACTTTATATTATAGGGTAGGGTGTG GTTGCACACACCTTAATCCCAGCACTTGGAAGGCAGAGGCAGTTGGATCT CTGGGAGTTTGAGGCCAGTTTGGCCTATATAGTGAGTTCTAGGCCAGCCA TGGATGCATAGTGAGACTCTTTCTCAAAACAAACAAACAAACAAACAAAC AAACAACTTTTAGAATGTAGAATTCCGTAAAAAAAAAAAATCCCTTGAAA GTTAATGGGGTCCTCAATTTTCTTCCTAGAATTTGGAGGCCTCTTCAGAA TGTTGACTATCTGACAGGTGACTCAGAAGGTCCTGTTCCTGGTCAATGAT CTATAACATGGGCCAAAACATTCCCAACTTGAATGTCTAGAATGTGGAAT TGGTTCATTTTCCTGTTCAGTGAAATGGACACAGAACAAAAGAACCCAGT GTCCAGCAAGAATTGCCTTGCCCAAACCTACGTCTACGCCAAAGGTCAAG GCAGTAAGGTGTTCTTGGGAGCCACACTTAGACTCTTTCCAAAGATGTGG AGGGAACAGATGGACTCATCTATGATCTTGGTTGGAAACCACCACAGTTC TATCCCCATCCAGATCTTTGCTTGTGGCAGCTGTTTCTCATGAAGCTAAT AAAATTC II. MOUSE COX-2 cDNA SEQUENCE (SEQ. ID. NO. 6) AGTTGTCAAACTGCGAGCTAAGAGCTTCAGGAGTCAGTCAGGACTCTGCT CACGAAGAATCTCAGCACTGCATCCTGCCAGCTCCACCGCCACCACCACT GCCACCTCCGCTGCCACCTCTGCGATGCTCTTCCGAGCTGTGCTGCTCTG CGCTGCCCTGGGGCTCAGCCAGGCAGCAAATCCTTGCTGTTCCAATCCAT GTCAAAACCGTGGGGAATGTATGAGCACAGGATTTGACCAGTATAAGTGT GACTGTACCCGGACTGGATTCTATGGTGAAAACTGTACTACACCTGAATT TCTGACAAGAATCAAATTACTGCTGAAGCCCACCCCAAACACAGTGCACT ACATCCTGACCCACTTCAAGGGAGTCTGGAACATTGTGAACAACATCCCC TTCCTGCGAAGTTTAACTATGAAATATGTGCTGACATCCAGATCATATTT GATTGACAGTCCACCTACTTACAATGTGCACTATGGTTACAAAAGCTGGG AAGCCTTCTCCAACCTCTCCTACTACACCAGGGCCCTTCCTCCAGTAGCA GATGACTGCCCAACTCCCATGGGTGTGAAGGGAAATAAGGAGCTTCCTGA TTCAAAAGAAGTGCTGGAAAAGGTTCTTCTACGGAGAGAGTTCATCCCTG ACCCCCAAGGCTCAAATATGATGTTTGCATTCTTTGCCCAGCACTTCACC CATCAGTTTTTCAAGACAGATCATAAGCGAGGACCTGGGTTCACCCGAGG ACTGGGCCATGGAGTGGACTTAAATCACATTTATGGTGAAACTCTGGACA GACAACATAAACTGCGCCTTTTCAAGGATGGAAAATTGAAATATCAGGTC ATTGGTGGAGAGGTGTATCCCCCCACAGTCAAAGACACTCAGGTAGAGAT GATCTACCCTCCTCACATCCCTGAGAACCTGCAGTTTGCTGTGGGGCAGG AAGTCTTTGGTCTGGTGCCTGGTCTGATGATGTATGCCACCATCTGGCTT CGGGAGCACAACAGAGTGTGCGACATACTCAAGCAGGAGCATCCTGAGTG GGGTGATGAGCAACTATTCCAAACCAGCAGACTCATACTCATAGGAGAGA CTATCAAGATAGTGATCGAAGACTACGTGCAACACCTGAGCGGTTACCAC TTCAAACTCAAGTTTGACCCAGAGCTCCTTTTCAACCAGCAGTTCCAGTA TCAGAACCGCATTGCCTCTGAATTCAACACACTCTATCACTGGCACCCCC TGCTGCCCGACACCTTCAACATTGAAGACCAGGAGTACAGCTTCAAACAG TTTCTCTACAACAACTCCATCCTCCTGGAACATGGACTCACTCAGTTTGT TGAGTCATTCACCAGACAGATTGCTGGCCGGGTTGCTGGGGGAAGAAATG TGCCAATTGCTGTACAAGCAGTGGCAAAGGCCTCCATTGACCAGAGCAGA GAGATGAAATACCAGTCTCTCAATGAGTACCGCAAACGCTTCTCCCTGAA GCCGTACACATCATTTGAAGAACTTACAGGAGAGAAGGAAATGGCTGCAG AATTGAAAGCCCTCTACAGTGACATCGATGTCATGGAACTGTACCCTGCC CTGCTGGTGGAAAAACCTCGTCCAGATGCTATCTTTGGGGAGACCATGGT AGAGCTTGGAGCACCATTCTCCTTGAAAGGACTTATGGGAAATCCCATCT GTTCTCCTCAATACTGGAAGCCGAGCACCTTTGGAGGCGAAGTGGGTTTT AAGATCATCAATACTGCCTCAATTCAGTCTCTCATCTGCAATAATGTGAA GGGGTGTCCCTTCACTTCTTTCAATGTGCAAGATCCACAGCCTACCAAAA CAGCCACCATCAATGCAAGTGCCTCCCACTCCAGACTAGATGACATTAAC CCTACAGTACTAATCAAAAGGCGTTCAACTGAGCTGTAAAAGTCTACTGA CCATATTTATTTATTTATGTGAAGGAATTTAATTTAATTATTTAATATTT ATACTGAATTTTTTTTCATGTAACATCTTCCATAACAGAAGGCAATGTTC TTGAACAATGTTACATTTGTGAAGATTCCTCCGGTGTTTGTCCTTTAAAT ATGTGTTACCTGAAACTGAAAGGAAATCAGCATTCATTCCTCTACATAAG CCAGTGAGAAGGGAAATGAATTTTGATATCTTTATACTTGAATTTCAGAT CATGAATTAGCTTAACAAGAACCAAGGAAAAATGTATGAATATGTGAGTG TTGTTACAAGATGAAAAATGCTGCAGGTATCAACACTGTTGGTTACACTG TGTCTTCTTTACCTATGATAGGAGCATGTAATGTGGAATTCGTCTTAAAT CCTTGCATATCTTTATCTCATCAAACAAAGGGGTCCAAGTTCAGTTTTAA ATAAGCATTTAAGGCAGATACTGACAACAATCTCATTTTTTTAAAATGTT GTCTTGAGACAAATAATTTGAAATTTCTAAATTGGGACGTTTGAATCACT TTTGAAAGCTCTTACTTTCTTAAGCTGTCAGGTTTGTACCGACATGGAGT AAACAGCTATCATAAACGTAAATCTCCAAAACTAGTAGAAATTATGTCAT GATTGATGGTTAAGATACCATGTCAGGGATTGTCTTTTCTTAGAAGTAGT GAAAGCTACTTACTATGACAATCAGACCTTCCTTGTATGTCAAAATGCTG GTGTGGAAGGTGGTGGAGCCCGTGCTGCTCTGTCTTAACTATGAGTGTGA GCTTTAAAGCTCGTTGATGAGTGGTAGCCAGCAAAGCCTAGAGCAACAAA AGCGTTCTACAAAGGAACTAACCAAGAACAAAGAAGGGTTCCCAATTAAA GATCACATTCAGGGTTAAACTTCCAAAGGAGACATCCTGATCCTGGTTTT GTGCTGGCCTGGTACTCAGTAGGTTTTTGCTGTGAGGTTAAAGACTTGCC AGGGCTGAACTTCGAAACAGTTTTTCTGTTGCACAGTATGATGTAACAGT CCATCTCTCAATGCAATAGGTATCAGTGGCCTCGTGAGCTTCTTCACAAT ATTTGATATGTCTTCCAGCCCATTGAACCTGGACTGCAGAAGGCCCCATG TCATGTGTGAGCTCAGCCTGGATGCCAGCATTTGCTGCTCCTCTTAGTTC CGTTTCTCGTGGTCACTTTACTACGAGAAACGCTGATTGGGTTTTCGTAG CTGTGTTACCAGGTTTTTAGTATCAGAACTATTCTTTCTTTAACCTCTAT TCATATTTTCTCTACTTGAAGTTTTACATTCAGGAAAACCTCAGCGCTCA GGACTACTATGTACCTCCCCTTTGGAGGGAAAAATTCATTTTTAGGTAAA
AGGCAAAAATTTTTTAAAAATATTTTTTATTTATAATTATATGGAAGGGC CCTACCAAGATGCTAGAAATATAGGGAGTTCCTGACAAGAAATTTCCATT CTTATTCTGAAGAATTGCTTTCTTACTTAAAAACAAAGACAGTTTGTGAG TAGTTCTGGGCAATAGGGATAAATATAAAACAATAATGATGATCATTTTC TACATCTCATTATCAGCTGAGGTACTGTATATTACTGAATTTATTGAAGA TAGTTATGTCTTTTAGACATTGTTGTTATAAACTATGTTTAAGCCTACTA CAAGTGTTTCTTTTTTGCATTATGTTGGAATTGATGTACCTTTTTTATGA TTACCTCTCTGAACTATGGTGTGAACAATCAAACAAAATGATGAGATTAA CGTTCATGGATAAATTCTAAGAAAACTAGTGTATTTTTTTGAAAAGTTTG AAGTTAGAACTTAGGCTGTTGGAATTTACGCATAAAGCAGACTGCATAGG ATCCAATATTGACTGACCCAAGCATGTTATAAAGACTGACATTTTAGACA TTTTGAAGGCCCTGTAAGTGTTTATTAATTAGTTAGAACTTAATTGATTA AAAAATATATCCAAAGCACTATAGGCATTAGAATTC III. MOUSE COX-1 AMINO ACID SEQUENCE (SEQ. ID. NO. 7) MSRRSLSLWFPLLLLLLLPPTPSVLLADPGVPSPVNPCCYYPCQNQGVCV RFGLDNYQCDCTRTGYSGPNCTIPEIWTWLRNSLRPSPSFTHFLLTHGYW LWEFVNATFIREVLMRLVLTVRSNLIPSPPTYNSAHDYISWESFSNVSYY TRILPSVPKDCPTPMGTKGKKQLPDVQLLAQQLLLRREFIPAPQGTNI- LFAFFAQHFTHQFFKTSGKMGPGFTKALGHGVDLGHIYGDNLERQYHLRL FKDGKLKYQVLDGEVYPPSVEQASVLMRYPPGVPPERQMAVGQEVFGLLP GLMLFSTIWLREHNRVCDLLKEEHPTWDDEQLFQTTRLILIGETIKIVIE EYVQHLSGYFLQLKFDPELLFRAQFQYRNRIAMEFNHLYHWHPLMPNSFQ VGSQEYSYEQFLFNTSMLVDYGVEALVDAFSRQRAGRIGGGRNFDYHVLH VAVDVIKESREMRLQPFNEYRKRFGLKPYTSFQELTGEKEMAAELEELYG DIDALEFYPGLLLEKCQPNSIFGESMIEMGAPFSLKGLLGNPICSPEYWK PSTFGGDVGFNLVNTASLKKLVCLNTKTCPYVSFRVPDYPGDDGSVLVRR STEL IV. MOUSE COX-2 AMINO ACID SEQUENCE (SEQ. ID. NO. 8) MLFRAVLLCAALGLSQAANPCCSNPCQNRGECMSTGFDQYKCDCTRTGFY GENCTTPEFLTRIKLLLKPTPNTVHYILTHFKGVWNIVNNIPFLRSLTMK YVLTSRSYLIDSPPTYNVHYGYKSWEAFSNLSYYTRALPPVADDCPTPMG VKGNKELPDSKEVLEKVLLRREFIPDPQGSNMMFAFFAQHFTHQFFKTDH KRGPGFTRGLGHGVDLNHIYGETLDRQHKLRLFKDGKLKYQVIGGEVYPP TVKDTQVEMIYPPHIPENLQFAVGQEVFGLVPGLMMYATIWLREHNRVCD ILKQEHPEWGDEQLFQTSRLILIGETIKIVIEDYVQHLSGYHFKLKFDPE LLFNQQFQYQNRIASEFNTLYHWHPLLPDTFNIEDQEYSFKQFLYNNSIL LEHGLTQFVESFTRQIAGRVAGGRNVPIAVQAVAKASIDQSREMKYQSLN EYRKRFSLKPYTSFEELTGEKEMAAELKALYSDIDVMELYPALLVEKPRP DAIFGETMVELGAPFSLKGLMGNPICSPQYWKPSTFGGEVGFKIINTASI QSLICNNVKGCPFTSFNVQDPQPT
Sequence CWU
1
812554DNAArtificial sequenceHuman COX-1 cDNA 1gcgccatgag ccggagtctc
ttgctccggt tcttgctgtt cctgctcctg ctcccgccgc 60tccccgtcct gctcgcggac
ccaggggcgc ccacgccagt gaatccctgt tgttactatc 120catgccagca ccagggcatc
tgtgtccgct tcggccttga ccgctaccag tgtgactgca 180cccgcacggg ctattccggc
cccaactgca ccatccctgg cctgtggacc tggctccgga 240attcactgcg gcccagcccc
tctttcaccc acttcctgct cactcacggg cgctggttct 300gggagtttgt caatgccacc
ttcatccgag agatgctcat gcgcctggta ctcacagtgc 360gctccaacct tatccccagt
ccccccacct acaactcagc acatgactac atcagctggg 420agtctttctc caacgtgagc
tattacactc gtattctgcc ctctgtgcct aaagattgcc 480ccacacccat gggaaccaaa
gggaagaagc agttgccaga tgcccagctc ctggcccgcc 540gcttcctgct caggaggaag
ttcatacctg acccccaagg caccaacctc atgtttgcct 600tctttgcaca acacttcacc
caccagttct tcaaaacttc tggcaagatg ggtcctggct 660tcaccaaggc cttgggccat
ggggtagacc tcggccacat ttatggagac aatctggagc 720gtcagtatca actgcggctc
tttaaggatg ggaaactcaa gtaccaggtg ctggatggag 780aaatgtaccc gccctcggta
gaagaggcgc ctgtgttgat gcactacccc cgaggcatcc 840cgccccagag ccagatggct
gtgggccagg aggtgtttgg gctgcttcct gggctcatgc 900tgtatgccac gctctggcta
cgtgagcaca accgtgtgtg tgacctgctg aaggctgagc 960accccacctg gggcgatgag
cagcttttcc agacgacccg cctcatcctc ataggggaga 1020ccatcaagat tgtcatcgag
gagtacgtgc agcagctgag tggctatttc ctgcagctga 1080aatttgaccc agagctgctg
ttcggtgtcc agttccaata ccgcaaccgc attgccatgg 1140agttcaacca tctctaccac
tggcaccccc tcatgcctga ctccttcaag gtgggctccc 1200aggagtacag ctacgagcag
ttcttgttca acacctccat gttggtggac tatggggttg 1260aggccctggt ggatgccttc
tctcgccaga ttgctggccg gatcggtggg ggcaggaaca 1320tggaccacca catcctgcat
gtggctgtgg atgtcatcag ggagtctcgg gagatgcggc 1380tgcagccctt caatgagtac
cgcaagaggt ttggcatgaa accctacacc tccttccagg 1440agctcgtagg agagaaggag
atggcagcag agttggagga attgtatgga gacattgatg 1500cgttggagtt ctaccctgga
ctgcttcttg aaaagtgcca tccaaactct atctttgggg 1560agagtatgat agagattggg
gctccctttt ccctcaaggg tctcctaggg aatcccatct 1620gttctccgga gtactggaag
ccgagcacat ttggcggcga ggtgggcttt aacattgtca 1680agacggccac actgaagaag
ctggtctgcc tcaacaccaa gacctgtccc tacgtttcct 1740tccgtgtgcc ggatgccagt
caggatgatg ggcctgctgt ggagcgacca tccacagagc 1800tctgaggggc aggaaagcag
cattctggag gggagagctt tgtgcttgtc attccagagt 1860gctgaggcca gggctgatgg
tcttaaatgc tcattttctg gtttggcatg gtgagtgttg 1920gggttgacat ttagaacttt
aagtctcacc cattatctgg aatattgtga ttctgtttat 1980tcttccagaa tgctgaactc
cttgttagcc cttcagattg ttaggagtgg ttctcatttg 2040gtctgccaga atactgggtt
cttagttgac aacctagaat gtcagatttc tggttgattt 2100gtaacacagt cattctagga
tgtggagcta ctgatgaaat ctgctagaaa gttagggggt 2160tcttattttg cattccagaa
tcttgacttt ctgattggtg attcaaagtg ttgtgttccc 2220tggctgatga tccagaacag
tggctcgtat cccaaatctg tcagcatctg gctgtctaga 2280atgtggattt gattcatttt
cctgttcagt gagatatcat agagacggag atcctaaggt 2340ccaacaagaa tgcattccct
gaatctgtgc ctgcactgag agggcaagga agtggggtgt 2400tcttcttggg acccccacta
agaccctggt ctgaggatgt agagagaaca ggtgggctgt 2460attcacgcca ttggttggaa
gctaccagag ctctatcccc atccaggtct tgactcatgg 2520cagctgtttc tcatgaagct
aataaaattc gccc 255424465DNAArtificial
sequenceHuman COX-2 cDNA 2caattgtcat acgacttgca gtgagcgtca ggagcacgtc
caggaactcc tcagcagcgc 60ctccttcagc tccacagcca gacgccctca gacagcaaag
cctacccccg cgccgcgccc 120tgcccgccgc tcggatgctc gcccgcgccc tgctgctgtg
cgcggtcctg gcgctcagcc 180atacagcaaa tccttgctgt tcccacccat gtcaaaaccg
aggtgtatgt atgagtgtgg 240gatttgacca gtataagtgc gattgtaccc ggacaggatt
ctatggagaa aactgctcaa 300caccggaatt tttgacaaga ataaaattat ttctgaaacc
cactccaaac acagtgcact 360acatacttac ccacttcaag ggattttgga acgttgtgaa
taacattccc ttccttcgaa 420atgcaattat gagttatgtc ttgacatcca gatcacattt
gattgacagt ccaccaactt 480acaatgctga ctatggctac aaaagctggg aagccttctc
taacctctcc tattatacta 540gagcccttcc tcctgtgcct gatgattgcc cgactccctt
gggtgtcaaa ggtaaaaagc 600agcttcctga ttcaaatgag attgtggaaa aattgcttct
aagaagaaag ttcatccctg 660atccccaggg ctcaaacatg atgtttgcat tctttgccca
gcacttcacg catcagtttt 720tcaagacaga tcataagcga gggccagctt tcaccaacgg
gctgggccat ggggtggact 780taaatcatat ttacggtgaa actctggcta gacagcgtaa
actgcgcctt ttcaaggatg 840gaaaaatgaa atatcagata attgatggag agatgtatcc
tcccacagtc aaagatactc 900aggcagagat gatctaccct cctcaagtcc ctgagcatct
acggtttgct gtggggcagg 960aggtctttgg tctggtgcct ggtctgatga tgtatgccac
aatctggctg cgggaacaca 1020acagagtatg cgatgtgctt aaacaggagc atcctgaatg
gggtgatgag cagttgttcc 1080agacaagcag gctaatactg ataggagaga ctattaagat
tgtgattgaa gattatgtgc 1140aacacttgag tggctatcac ttcaaactga aatttgaccc
agaactactt ttcaacaaac 1200aattccagta ccaaaatcgt attgctgctg aatttaacac
cctctatcac tggcatcccc 1260ttctgcctga cacctttcaa attcatgacc agaaatacaa
ctatcaacag tttatctaca 1320acaactctat attgctggaa catggaatta cccagtttgt
tgaatcattc accaggcaaa 1380ttgctggcag ggttgctggt ggtaggaatg ttccacccgc
agtacagaaa gtatcacagg 1440cttccattga ccagagcagg cagatgaaat accagtcttt
taatgagtac cgcaaacgct 1500ttatgctgaa gccctatgaa tcatttgaag aacttacagg
agaaaaggaa atgtctgcag 1560agttggaagc actctatggt gacatcgatg ctgtggagct
gtatcctgcc cttctggtag 1620aaaagcctcg gccagatgcc atctttggtg aaaccatggt
agaagttgga gcaccattct 1680ccttgaaagg acttatgggt aatgttatat gttctcctgc
ctactggaag ccaagcactt 1740ttggtggaga agtgggtttt caaatcatca acactgcctc
aattcagtct ctcatctgca 1800ataacgtgaa gggctgtccc tttacttcat tcagtgttcc
agatccagag ctcattaaaa 1860cagtcaccat caatgcaagt tcttcccgct ccggactaga
tgatatcaat cccacagtac 1920tactaaaaga acgttcgact gaactgtaga agtctaatga
tcatatttat ttatttatat 1980gaaccatgtc tattaattta attatttaat aatatttata
ttaaactcct tatgttactt 2040aacatcttct gtaacagaag tcagtactcc tgttgcggag
aaaggagtca tacttgtgaa 2100gacttttatg tcactactct aaagattttg ctgttgctgt
taagtttgga aaacagtttt 2160tattctgttt tataaaccag agagaaatga gttttgacgt
ctttttactt gaatttcaac 2220ttatattata agaacgaaag taaagatgtt tgaatactta
aacactatca caagatggca 2280aaatgctgaa agtttttaca ctgtcgatgt ttccaatgca
tcttccatga tgcattagaa 2340gtaactaatg tttgaaattt taaagtactt ttggttattt
ttctgtcatc aaacaaaaac 2400aggtatcagt gcattattaa atgaatattt aaattagaca
ttaccagtaa tttcatgtct 2460actttttaaa atcagcaatg aaacaataat ttgaaatttc
taaattcata gggtagaatc 2520acctgtaaaa gcttgtttga tttcttaaag ttattaaact
tgtacatata ccaaaaagaa 2580gctgtcttgg atttaaatct gtaaaatcag atgaaatttt
actacaattg cttgttaaaa 2640tattttataa gtgatgttcc tttttcacca agagtataaa
cctttttagt gtgactgtta 2700aaacttcctt ttaaatcaaa atgccaaatt tattaaggtg
gtggagccac tgcagtgtta 2760tctcaaaata agaatatttt gttgagatat tccagaattt
gtttatatgg ctggtaacat 2820gtaaaatcta tatcagcaaa agggtctacc tttaaaataa
gcaataacaa agaagaaaac 2880caaattattg ttcaaattta ggtttaaact tttgaagcaa
actttttttt atccttgtgc 2940actgcaggcc tggtactcag attttgctat gaggttaatg
aagtaccaag ctgtgcttga 3000ataacgatat gttttctcag attttctgtt gtacagttta
atttagcagt ccatatcaca 3060ttgcaaaagt agcaatgacc tcataaaata cctcttcaaa
atgcttaaat tcatttcaca 3120cattaatttt atctcagtct tgaagccaat tcagtaggtg
cattggaatc aagcctggct 3180acctgcatgc tgttcctttt cttttcttct tttagccatt
ttgctaagag acacagtctt 3240ctcatcactt cgtttctcct attttgtttt actagtttta
agatcagagt tcactttctt 3300tggactctgc ctatattttc ttacctgaac ttttgcaagt
tttcaggtaa acctcagctc 3360aggactgcta tttagctcct cttaagaaga ttaaaagaga
aaaaaaaagg cccttttaaa 3420aatagtatac acttatttta agtgaaaagc agagaatttt
atttatagct aattttagct 3480atctgtaacc aagatggatg caaagaggct agtgcctcag
agagaactgt acggggtttg 3540tgactggaaa aagttacgtt cccattctaa ttaatgccct
ttcttattta aaaacaaaac 3600caaatgatat ctaagtagtt ctcagcaata ataataatga
cgataatact tcttttccac 3660atctcattgt cactgacatt taatggtact gtatattact
taatttattg aagattatta 3720tttatgtctt attaggacac tatggttata aactgtgttt
aagcctacaa tcattgattt 3780ttttttgtta tgtcacaatc agtatatttt ctttggggtt
acctctctga atattatgta 3840aacaatccaa agaaatgatt gtattaagat ttgtgaataa
atttttagaa atctgattgg 3900catattgaga tatttaaggt tgaatgtttg tccttaggat
aggcctatgt gctagcccac 3960aaagaatatt gtctcattag cctgaatgtg ccataagact
gaccttttaa aatgttttga 4020gggatctgtg gatgcttcgt taatttgttc agccacaatt
tattgagaaa atattctgtg 4080tcaagcactg tgggttttaa tatttttaaa tcaaacgctg
attacagata atagtattta 4140tataaataat tgaaaaaaat tttcttttgg gaagagggag
aaaatgaaat aaatatcatt 4200aaagataact caggagaatc ttctttacaa ttttacgttt
agaatgttta aggttaagaa 4260agaaatagtc aatatgcttg tataaaacac tgttcactgt
tttttttaaa aaaaaaactt 4320gatttgttat taacattgat ctgctgacaa aacctgggaa
tttgggttgt gtatgcgaat 4380gtttcagtgc ctcagacaaa tgtgtattta acttatgtaa
aagataagtc tggaaataaa 4440tgtctgttta tttttgtact attta
44653599PRTHomo sapiens 3Met Ser Arg Ser Leu Leu
Leu Arg Phe Leu Leu Phe Leu Leu Leu Leu1 5
10 15Pro Pro Leu Pro Val Leu Leu Ala Asp Pro Gly Ala
Pro Thr Pro Val 20 25 30Asn
Pro Cys Cys Tyr Tyr Pro Cys Gln His Gln Gly Ile Cys Val Arg 35
40 45Phe Gly Leu Asp Arg Tyr Gln Cys Asp
Cys Thr Arg Thr Gly Tyr Ser 50 55
60Gly Pro Asn Cys Thr Ile Pro Gly Leu Trp Thr Trp Leu Arg Asn Ser65
70 75 80Leu Arg Pro Ser Pro
Ser Phe Thr His Phe Leu Leu Thr His Gly Arg 85
90 95Trp Phe Trp Glu Phe Val Asn Ala Thr Phe Ile
Arg Glu Met Leu Met 100 105
110Arg Leu Val Leu Thr Val Arg Ser Asn Leu Ile Pro Ser Pro Pro Thr
115 120 125Tyr Asn Ser Ala His Asp Tyr
Ile Ser Trp Glu Ser Phe Ser Asn Val 130 135
140Ser Tyr Tyr Thr Arg Ile Leu Pro Ser Val Pro Lys Asp Cys Pro
Thr145 150 155 160Pro Met
Gly Thr Lys Gly Lys Lys Gln Leu Pro Asp Ala Gln Leu Leu
165 170 175Ala Arg Arg Phe Leu Leu Arg
Arg Lys Phe Ile Pro Asp Pro Gln Gly 180 185
190Thr Asn Leu Met Phe Ala Phe Phe Ala Gln His Phe Thr His
Gln Phe 195 200 205Phe Lys Thr Ser
Gly Lys Met Gly Pro Gly Phe Thr Lys Ala Leu Gly 210
215 220His Gly Val Asp Leu Gly His Ile Tyr Gly Asp Asn
Leu Glu Arg Gln225 230 235
240Tyr Gln Leu Arg Leu Phe Lys Asp Gly Lys Leu Lys Tyr Gln Val Leu
245 250 255Asp Gly Glu Met Tyr
Pro Pro Ser Val Glu Glu Ala Pro Val Leu Met 260
265 270His Tyr Pro Arg Gly Ile Pro Pro Gln Ser Gln Met
Ala Val Gly Gln 275 280 285Glu Val
Phe Gly Leu Leu Pro Gly Leu Met Leu Tyr Ala Thr Leu Trp 290
295 300Leu Arg Glu His Asn Arg Val Cys Asp Leu Leu
Lys Ala Glu His Pro305 310 315
320Thr Trp Gly Asp Glu Gln Leu Phe Gln Thr Thr Arg Leu Ile Leu Ile
325 330 335Gly Glu Thr Ile
Lys Ile Val Ile Glu Glu Tyr Val Gln Gln Leu Ser 340
345 350Gly Tyr Phe Leu Gln Leu Lys Phe Asp Pro Glu
Leu Leu Phe Gly Val 355 360 365Gln
Phe Gln Tyr Arg Asn Arg Ile Ala Met Glu Phe Asn His Leu Tyr 370
375 380His Trp His Pro Leu Met Pro Asp Ser Phe
Lys Val Gly Ser Gln Glu385 390 395
400Tyr Ser Tyr Glu Gln Phe Leu Phe Asn Thr Ser Met Leu Val Asp
Tyr 405 410 415Gly Val Glu
Ala Leu Val Asp Ala Phe Ser Arg Gln Ile Ala Gly Arg 420
425 430Ile Gly Gly Gly Arg Asn Met Asp His His
Ile Leu His Val Ala Val 435 440
445Asp Val Ile Arg Glu Ser Arg Glu Met Arg Leu Gln Pro Phe Asn Glu 450
455 460Tyr Arg Lys Arg Phe Gly Met Lys
Pro Tyr Thr Ser Phe Gln Glu Leu465 470
475 480Val Gly Glu Lys Glu Met Ala Ala Glu Leu Glu Glu
Leu Tyr Gly Asp 485 490
495Ile Asp Ala Leu Glu Phe Tyr Pro Gly Leu Leu Leu Glu Lys Cys His
500 505 510Pro Asn Ser Ile Phe Gly
Glu Ser Met Ile Glu Ile Gly Ala Pro Phe 515 520
525Ser Leu Lys Gly Leu Leu Gly Asn Pro Ile Cys Ser Pro Glu
Tyr Trp 530 535 540Lys Pro Ser Thr Phe
Gly Gly Glu Val Gly Phe Asn Ile Val Lys Thr545 550
555 560Ala Thr Leu Lys Lys Leu Val Cys Leu Asn
Thr Lys Thr Cys Pro Tyr 565 570
575Val Ser Phe Arg Val Pro Asp Ala Ser Gln Asp Asp Gly Pro Ala Val
580 585 590Glu Arg Pro Ser Thr
Glu Leu 5954604PRTHomo sapiens 4Met Leu Ala Arg Ala Leu Leu Leu
Cys Ala Val Leu Ala Leu Ser His1 5 10
15Thr Ala Asn Pro Cys Cys Ser His Pro Cys Gln Asn Arg Gly
Val Cys 20 25 30Met Ser Val
Gly Phe Asp Gln Tyr Lys Cys Asp Cys Thr Arg Thr Gly 35
40 45Phe Tyr Gly Glu Asn Cys Ser Thr Pro Glu Phe
Leu Thr Arg Ile Lys 50 55 60Leu Phe
Leu Lys Pro Thr Pro Asn Thr Val His Tyr Ile Leu Thr His65
70 75 80Phe Lys Gly Phe Trp Asn Val
Val Asn Asn Ile Pro Phe Leu Arg Asn 85 90
95Ala Ile Met Ser Tyr Val Leu Thr Ser Arg Ser His Leu
Ile Asp Ser 100 105 110Pro Pro
Thr Tyr Asn Ala Asp Tyr Gly Tyr Lys Ser Trp Glu Ala Phe 115
120 125Ser Asn Leu Ser Tyr Tyr Thr Arg Ala Leu
Pro Pro Val Pro Asp Asp 130 135 140Cys
Pro Thr Pro Leu Gly Val Lys Gly Lys Lys Gln Leu Pro Asp Ser145
150 155 160Asn Glu Ile Val Glu Lys
Leu Leu Leu Arg Arg Lys Phe Ile Pro Asp 165
170 175Pro Gln Gly Ser Asn Met Met Phe Ala Phe Phe Ala
Gln His Phe Thr 180 185 190His
Gln Phe Phe Lys Thr Asp His Lys Arg Gly Pro Ala Phe Thr Asn 195
200 205Gly Leu Gly His Gly Val Asp Leu Asn
His Ile Tyr Gly Glu Thr Leu 210 215
220Ala Arg Gln Arg Lys Leu Arg Leu Phe Lys Asp Gly Lys Met Lys Tyr225
230 235 240Gln Ile Ile Asp
Gly Glu Met Tyr Pro Pro Thr Val Lys Asp Thr Gln 245
250 255Ala Glu Met Ile Tyr Pro Pro Gln Val Pro
Glu His Leu Arg Phe Ala 260 265
270Val Gly Gln Glu Val Phe Gly Leu Val Pro Gly Leu Met Met Tyr Ala
275 280 285Thr Ile Trp Leu Arg Glu His
Asn Arg Val Cys Asp Val Leu Lys Gln 290 295
300Glu His Pro Glu Trp Gly Asp Glu Gln Leu Phe Gln Thr Ser Arg
Leu305 310 315 320Ile Leu
Ile Gly Glu Thr Ile Lys Ile Val Ile Glu Asp Tyr Val Gln
325 330 335His Leu Ser Gly Tyr His Phe
Lys Leu Lys Phe Asp Pro Glu Leu Leu 340 345
350Phe Asn Lys Gln Phe Gln Tyr Gln Asn Arg Ile Ala Ala Glu
Phe Asn 355 360 365Thr Leu Tyr His
Trp His Pro Leu Leu Pro Asp Thr Phe Gln Ile His 370
375 380Asp Gln Lys Tyr Asn Tyr Gln Gln Phe Ile Tyr Asn
Asn Ser Ile Leu385 390 395
400Leu Glu His Gly Ile Thr Gln Phe Val Glu Ser Phe Thr Arg Gln Ile
405 410 415Ala Gly Arg Val Ala
Gly Gly Arg Asn Val Pro Pro Ala Val Gln Lys 420
425 430Val Ser Gln Ala Ser Ile Asp Gln Ser Arg Gln Met
Lys Tyr Gln Ser 435 440 445Phe Asn
Glu Tyr Arg Lys Arg Phe Met Leu Lys Pro Tyr Glu Ser Phe 450
455 460Glu Glu Leu Thr Gly Glu Lys Glu Met Ser Ala
Glu Leu Glu Ala Leu465 470 475
480Tyr Gly Asp Ile Asp Ala Val Glu Leu Tyr Pro Ala Leu Leu Val Glu
485 490 495Lys Pro Arg Pro
Asp Ala Ile Phe Gly Glu Thr Met Val Glu Val Gly 500
505 510Ala Pro Phe Ser Leu Lys Gly Leu Met Gly Asn
Val Ile Cys Ser Pro 515 520 525Ala
Tyr Trp Lys Pro Ser Thr Phe Gly Gly Glu Val Gly Phe Gln Ile 530
535 540Ile Asn Thr Ala Ser Ile Gln Ser Leu Ile
Cys Asn Asn Val Lys Gly545 550 555
560Cys Pro Phe Thr Ser Phe Ser Val Pro Asp Pro Glu Leu Ile Lys
Thr 565 570 575Val Thr Ile
Asn Ala Ser Ser Ser Arg Ser Gly Leu Asp Asp Ile Asn 580
585 590Pro Thr Val Leu Leu Lys Glu Arg Ser Thr
Glu Leu 595 60052757DNAArtificial sequenceMouse
COX-1 cDNA 5gccgttggca ttgcacatcc atccactccc agagtcatga gtcgaaggag
tctctcgctc 60tggtttcccc tgctgctgct cctgctgctg ccgccgacac cctcggtcct
gctcgcagat 120cctggggtgc cctcaccagt caatccctgt tgttactatc cgtgccagaa
ccagggtgtc 180tgtgtccgct ttggcctcga caactaccag tgtgattgta ctcgcacggg
ctactcaggc 240cccaactgta ccatccctga gatctggacc tggcttcgga attctctgcg
gcccagcccc 300tcgttcaccc atttcctgct gacacatgga tactggctct gggaatttgt
gaatgccacc 360ttcatccgag aagtactcat gcgcctggta ctcacagtgc ggtccaacct
tatccccagc 420cctccgacct acaactcagc gcatgactac atcagctggg agtccttctc
caatgtgagc 480tactatactc gcattctgcc ctctgtaccc aaagactgcc ccacacccat
ggggaccaaa 540gggaagaaac agttaccaga tgttcagctt ctggcccaac agctgctgct
gagaagggag 600ttcattcctg ccccccaggg caccaacatc ctgtttgcct tctttgcaca
acacttcacc 660caccagttct tcaagacctc tggaaagatg ggtcctggct ttaccaaggc
cttaggccac 720ggggtagacc ttggccacat ttatggagat aatctggaac gacagtatca
cctgcggctc 780ttcaaggatg ggaaacttaa gtaccaggtg ctggacggag aggtgtaccc
accttccgtg 840gaacaggcgt ccgtgttgat gcgctaccca ccaggtgtcc cgcctgaaag
gcagatggct 900gtgggccagg aggtgtttgg gttgcttccg gggctgatgc tcttctccac
gatctggctt 960cgtgaacata accgcgtgtg cgacctgctg aaggaggagc atcccacgtg
ggatgatgag 1020cagctcttcc agaccactcg cctcatcctt ataggagaaa ccatcaaaat
tgtcattgag 1080gagtatgtgc agcacttgag tggctatttc ctgcagctca agtttgaccc
ggagctgctg 1140ttccgagccc agttccaata tcgaaaccgc atcgccatgg aatttaacca
tctctatcac 1200tggcatccac tcatgcccaa ctccttccaa gtgggctcac aagagtacag
ctacgagcag 1260tttttattta acacttctat gctggtggac tatggggttg aggcactggt
ggatgccttc 1320tctcgccaga gggctggccg gattggtgga ggtaggaact ttgactatca
tgttctgcat 1380gtggctgtgg atgtcatcaa ggagtcccga gagatgcgcc tacagccctt
caatgaatac 1440cgaaagaggt ttggcttgaa gccttacacc tctttccagg agctcacagg
agagaaggag 1500atggctgctg agttggagga gctgtacggt gacatcgatg ctttagagtt
ctacccgggg 1560ttgctgctgg agaagtgcca gcccaactcc atctttggag aaagtatgat
agagatgggg 1620gctccctttt ccctcaaggg cctcctaggg aatcccatct gttccccaga
gtactggaaa 1680cccagcacgt tcggtggtga cgtgggcttc aaccttgtca acacagcctc
actgaagaaa 1740ctggtctgcc tcaacaccaa gacctgcccc tatgtttcct tccgtgtgcc
agattaccct 1800ggagatgacg ggtctgtctt agtgagacgc tccactgagc tctgagggag
ctggaaagca 1860gcctctggag ggaggagttt tgttcctgat gaagacaagt ccttgatgtg
ggttttcgtg 1920gcttggcatt gtgagagctg atgctcacat ttgaaacttt gggtcttacc
cttgcctaga 1980aaattgtgat tttgccactt tcggatgttg aattctttgt taactaagaa
agttagaagt 2040ggttttgtct gcctcctcag aacttggctc tttgttggca actcagaaag
tcagatttct 2100ggttgatttg gaatataggc ttaaaacttt atattatagg gtagggtgtg
gttgcacaca 2160ccttaatccc agcacttgga aggcagaggc agttggatct ctgggagttt
gaggccagtt 2220tggcctatat agtgagttct aggccagcca tggatgcata gtgagactct
ttctcaaaac 2280aaacaaacaa acaaacaaac aaacaacttt tagaatgtag aattccgtaa
aaaaaaaaaa 2340tcccttgaaa gttaatgggg tcctcaattt tcttcctaga atttggaggc
ctcttcagaa 2400tgttgactat ctgacaggtg actcagaagg tcctgttcct ggtcaatgat
ctataacatg 2460ggccaaaaca ttcccaactt gaatgtctag aatgtggaat tggttcattt
tcctgttcag 2520tgaaatggac acagaacaaa agaacccagt gtccagcaag aattgccttg
cccaaaccta 2580cgtctacgcc aaaggtcaag gcagtaaggt gttcttggga gccacactta
gactctttcc 2640aaagatgtgg agggaacaga tggactcatc tatgatcttg gttggaaacc
accacagttc 2700tatccccatc cagatctttg cttgtggcag ctgtttctca tgaagctaat
aaaattc 275763986DNAArtificial sequenceMouse COX-2 cDNA 6agttgtcaaa
ctgcgagcta agagcttcag gagtcagtca ggactctgct cacgaagaat 60ctcagcactg
catcctgcca gctccaccgc caccaccact gccacctccg ctgccacctc 120tgcgatgctc
ttccgagctg tgctgctctg cgctgccctg gggctcagcc aggcagcaaa 180tccttgctgt
tccaatccat gtcaaaaccg tggggaatgt atgagcacag gatttgacca 240gtataagtgt
gactgtaccc ggactggatt ctatggtgaa aactgtacta cacctgaatt 300tctgacaaga
atcaaattac tgctgaagcc caccccaaac acagtgcact acatcctgac 360ccacttcaag
ggagtctgga acattgtgaa caacatcccc ttcctgcgaa gtttaactat 420gaaatatgtg
ctgacatcca gatcatattt gattgacagt ccacctactt acaatgtgca 480ctatggttac
aaaagctggg aagccttctc caacctctcc tactacacca gggcccttcc 540tccagtagca
gatgactgcc caactcccat gggtgtgaag ggaaataagg agcttcctga 600ttcaaaagaa
gtgctggaaa aggttcttct acggagagag ttcatccctg acccccaagg 660ctcaaatatg
atgtttgcat tctttgccca gcacttcacc catcagtttt tcaagacaga 720tcataagcga
ggacctgggt tcacccgagg actgggccat ggagtggact taaatcacat 780ttatggtgaa
actctggaca gacaacataa actgcgcctt ttcaaggatg gaaaattgaa 840atatcaggtc
attggtggag aggtgtatcc ccccacagtc aaagacactc aggtagagat 900gatctaccct
cctcacatcc ctgagaacct gcagtttgct gtggggcagg aagtctttgg 960tctggtgcct
ggtctgatga tgtatgccac catctggctt cgggagcaca acagagtgtg 1020cgacatactc
aagcaggagc atcctgagtg gggtgatgag caactattcc aaaccagcag 1080actcatactc
ataggagaga ctatcaagat agtgatcgaa gactacgtgc aacacctgag 1140cggttaccac
ttcaaactca agtttgaccc agagctcctt ttcaaccagc agttccagta 1200tcagaaccgc
attgcctctg aattcaacac actctatcac tggcaccccc tgctgcccga 1260caccttcaac
attgaagacc aggagtacag cttcaaacag tttctctaca acaactccat 1320cctcctggaa
catggactca ctcagtttgt tgagtcattc accagacaga ttgctggccg 1380ggttgctggg
ggaagaaatg tgccaattgc tgtacaagca gtggcaaagg cctccattga 1440ccagagcaga
gagatgaaat accagtctct caatgagtac cgcaaacgct tctccctgaa 1500gccgtacaca
tcatttgaag aacttacagg agagaaggaa atggctgcag aattgaaagc 1560cctctacagt
gacatcgatg tcatggaact gtaccctgcc ctgctggtgg aaaaacctcg 1620tccagatgct
atctttgggg agaccatggt agagcttgga gcaccattct ccttgaaagg 1680acttatggga
aatcccatct gttctcctca atactggaag ccgagcacct ttggaggcga 1740agtgggtttt
aagatcatca atactgcctc aattcagtct ctcatctgca ataatgtgaa 1800ggggtgtccc
ttcacttctt tcaatgtgca agatccacag cctaccaaaa cagccaccat 1860caatgcaagt
gcctcccact ccagactaga tgacattaac cctacagtac taatcaaaag 1920gcgttcaact
gagctgtaaa agtctactga ccatatttat ttatttatgt gaaggaattt 1980aatttaatta
tttaatattt atactgaatt ttttttcatg taacatcttc cataacagaa 2040ggcaatgttc
ttgaacaatg ttacatttgt gaagattcct ccggtgtttg tcctttaaat 2100atgtgttacc
tgaaactgaa aggaaatcag cattcattcc tctacataag ccagtgagaa 2160gggaaatgaa
ttttgatatc tttatacttg aatttcagat catgaattag cttaacaaga 2220accaaggaaa
aatgtatgaa tatgtgagtg ttgttacaag atgaaaaatg ctgcaggtat 2280caacactgtt
ggttacactg tgtcttcttt acctatgata ggagcatgta atgtggaatt 2340cgtcttaaat
ccttgcatat ctttatctca tcaaacaaag gggtccaagt tcagttttaa 2400ataagcattt
aaggcagata ctgacaacaa tctcattttt ttaaaatgtt gtcttgagac 2460aaataatttg
aaatttctaa attgggacgt ttgaatcact tttgaaagct cttactttct 2520taagctgtca
ggtttgtacc gacatggagt aaacagctat cataaacgta aatctccaaa 2580actagtagaa
attatgtcat gattgatggt taagatacca tgtcagggat tgtcttttct 2640tagaagtagt
gaaagctact tactatgaca atcagacctt ccttgtatgt caaaatgctg 2700gtgtggaagg
tggtggagcc cgtgctgctc tgtcttaact atgagtgtga gctttaaagc 2760tcgttgatga
gtggtagcca gcaaagccta gagcaacaaa agcgttctac aaaggaacta 2820accaagaaca
aagaagggtt cccaattaaa gatcacattc agggttaaac ttccaaagga 2880gacatcctga
tcctggtttt gtgctggcct ggtactcagt aggtttttgc tgtgaggtta 2940aagacttgcc
agggctgaac ttcgaaacag tttttctgtt gcacagtatg atgtaacagt 3000ccatctctca
atgcaatagg tatcagtggc ctcgtgagct tcttcacaat atttgatatg 3060tcttccagcc
cattgaacct ggactgcaga aggccccatg tcatgtgtga gctcagcctg 3120gatgccagca
tttgctgctc ctcttagttc cgtttctcgt ggtcacttta ctacgagaaa 3180cgctgattgg
gttttcgtag ctgtgttacc aggtttttag tatcagaact attctttctt 3240taacctctat
tcatattttc tctacttgaa gttttacatt caggaaaacc tcagcgctca 3300ggactactat
gtacctcccc tttggaggga aaaattcatt tttaggtaaa aggcaaaaat 3360tttttaaaaa
tattttttat ttataattat atggaagggc cctaccaaga tgctagaaat 3420atagggagtt
cctgacaaga aatttccatt cttattctga agaattgctt tcttacttaa 3480aaacaaagac
agtttgtgag tagttctggg caatagggat aaatataaaa caataatgat 3540gatcattttc
tacatctcat tatcagctga ggtactgtat attactgaat ttattgaaga 3600tagttatgtc
ttttagacat tgttgttata aactatgttt aagcctacta caagtgtttc 3660ttttttgcat
tatgttggaa ttgatgtacc ttttttatga ttacctctct gaactatggt 3720gtgaacaatc
aaacaaaatg atgagattaa cgttcatgga taaattctaa gaaaactagt 3780gtattttttt
gaaaagtttg aagttagaac ttaggctgtt ggaatttacg cataaagcag 3840actgcatagg
atccaatatt gactgaccca agcatgttat aaagactgac attttagaca 3900ttttgaaggc
cctgtaagtg tttattaatt agttagaact taattgatta aaaaatatat 3960ccaaagcact
ataggcatta gaattc 39867602PRTMus
musculus 7Met Ser Arg Arg Ser Leu Ser Leu Trp Phe Pro Leu Leu Leu Leu
Leu1 5 10 15Leu Leu Pro
Pro Thr Pro Ser Val Leu Leu Ala Asp Pro Gly Val Pro 20
25 30Ser Pro Val Asn Pro Cys Cys Tyr Tyr Pro
Cys Gln Asn Gln Gly Val 35 40
45Cys Val Arg Phe Gly Leu Asp Asn Tyr Gln Cys Asp Cys Thr Arg Thr 50
55 60Gly Tyr Ser Gly Pro Asn Cys Thr Ile
Pro Glu Ile Trp Thr Trp Leu65 70 75
80Arg Asn Ser Leu Arg Pro Ser Pro Ser Phe Thr His Phe Leu
Leu Thr 85 90 95His Gly
Tyr Trp Leu Trp Glu Phe Val Asn Ala Thr Phe Ile Arg Glu 100
105 110Val Leu Met Arg Leu Val Leu Thr Val
Arg Ser Asn Leu Ile Pro Ser 115 120
125Pro Pro Thr Tyr Asn Ser Ala His Asp Tyr Ile Ser Trp Glu Ser Phe
130 135 140Ser Asn Val Ser Tyr Tyr Thr
Arg Ile Leu Pro Ser Val Pro Lys Asp145 150
155 160Cys Pro Thr Pro Met Gly Thr Lys Gly Lys Lys Gln
Leu Pro Asp Val 165 170
175Gln Leu Leu Ala Gln Gln Leu Leu Leu Arg Arg Glu Phe Ile Pro Ala
180 185 190Pro Gln Gly Thr Asn Ile
Leu Phe Ala Phe Phe Ala Gln His Phe Thr 195 200
205His Gln Phe Phe Lys Thr Ser Gly Lys Met Gly Pro Gly Phe
Thr Lys 210 215 220Ala Leu Gly His Gly
Val Asp Leu Gly His Ile Tyr Gly Asp Asn Leu225 230
235 240Glu Arg Gln Tyr His Leu Arg Leu Phe Lys
Asp Gly Lys Leu Lys Tyr 245 250
255Gln Val Leu Asp Gly Glu Val Tyr Pro Pro Ser Val Glu Gln Ala Ser
260 265 270Val Leu Met Arg Tyr
Pro Pro Gly Val Pro Pro Glu Arg Gln Met Ala 275
280 285Val Gly Gln Glu Val Phe Gly Leu Leu Pro Gly Leu
Met Leu Phe Ser 290 295 300Thr Ile Trp
Leu Arg Glu His Asn Arg Val Cys Asp Leu Leu Lys Glu305
310 315 320Glu His Pro Thr Trp Asp Asp
Glu Gln Leu Phe Gln Thr Thr Arg Leu 325
330 335Ile Leu Ile Gly Glu Thr Ile Lys Ile Val Ile Glu
Glu Tyr Val Gln 340 345 350His
Leu Ser Gly Tyr Phe Leu Gln Leu Lys Phe Asp Pro Glu Leu Leu 355
360 365Phe Arg Ala Gln Phe Gln Tyr Arg Asn
Arg Ile Ala Met Glu Phe Asn 370 375
380His Leu Tyr His Trp His Pro Leu Met Pro Asn Ser Phe Gln Val Gly385
390 395 400Ser Gln Glu Tyr
Ser Tyr Glu Gln Phe Leu Phe Asn Thr Ser Met Leu 405
410 415Val Asp Tyr Gly Val Glu Ala Leu Val Asp
Ala Phe Ser Arg Gln Arg 420 425
430Ala Gly Arg Ile Gly Gly Gly Arg Asn Phe Asp Tyr His Val Leu His
435 440 445Val Ala Val Asp Val Ile Lys
Glu Ser Arg Glu Met Arg Leu Gln Pro 450 455
460Phe Asn Glu Tyr Arg Lys Arg Phe Gly Leu Lys Pro Tyr Thr Ser
Phe465 470 475 480Gln Glu
Leu Thr Gly Glu Lys Glu Met Ala Ala Glu Leu Glu Glu Leu
485 490 495Tyr Gly Asp Ile Asp Ala Leu
Glu Phe Tyr Pro Gly Leu Leu Leu Glu 500 505
510Lys Cys Gln Pro Asn Ser Ile Phe Gly Glu Ser Met Ile Glu
Met Gly 515 520 525Ala Pro Phe Ser
Leu Lys Gly Leu Leu Gly Asn Pro Ile Cys Ser Pro 530
535 540Glu Tyr Trp Lys Pro Ser Thr Phe Gly Gly Asp Val
Gly Phe Asn Leu545 550 555
560Val Asn Thr Ala Ser Leu Lys Lys Leu Val Cys Leu Asn Thr Lys Thr
565 570 575Cys Pro Tyr Val Ser
Phe Arg Val Pro Asp Tyr Pro Gly Asp Asp Gly 580
585 590Ser Val Leu Val Arg Arg Ser Thr Glu Leu
595 6008604PRTMus musculus 8Met Leu Phe Arg Ala Val Leu
Leu Cys Ala Ala Leu Gly Leu Ser Gln1 5 10
15Ala Ala Asn Pro Cys Cys Ser Asn Pro Cys Gln Asn Arg
Gly Glu Cys 20 25 30Met Ser
Thr Gly Phe Asp Gln Tyr Lys Cys Asp Cys Thr Arg Thr Gly 35
40 45Phe Tyr Gly Glu Asn Cys Thr Thr Pro Glu
Phe Leu Thr Arg Ile Lys 50 55 60Leu
Leu Leu Lys Pro Thr Pro Asn Thr Val His Tyr Ile Leu Thr His65
70 75 80Phe Lys Gly Val Trp Asn
Ile Val Asn Asn Ile Pro Phe Leu Arg Ser 85
90 95Leu Thr Met Lys Tyr Val Leu Thr Ser Arg Ser Tyr
Leu Ile Asp Ser 100 105 110Pro
Pro Thr Tyr Asn Val His Tyr Gly Tyr Lys Ser Trp Glu Ala Phe 115
120 125Ser Asn Leu Ser Tyr Tyr Thr Arg Ala
Leu Pro Pro Val Ala Asp Asp 130 135
140Cys Pro Thr Pro Met Gly Val Lys Gly Asn Lys Glu Leu Pro Asp Ser145
150 155 160Lys Glu Val Leu
Glu Lys Val Leu Leu Arg Arg Glu Phe Ile Pro Asp 165
170 175Pro Gln Gly Ser Asn Met Met Phe Ala Phe
Phe Ala Gln His Phe Thr 180 185
190His Gln Phe Phe Lys Thr Asp His Lys Arg Gly Pro Gly Phe Thr Arg
195 200 205Gly Leu Gly His Gly Val Asp
Leu Asn His Ile Tyr Gly Glu Thr Leu 210 215
220Asp Arg Gln His Lys Leu Arg Leu Phe Lys Asp Gly Lys Leu Lys
Tyr225 230 235 240Gln Val
Ile Gly Gly Glu Val Tyr Pro Pro Thr Val Lys Asp Thr Gln
245 250 255Val Glu Met Ile Tyr Pro Pro
His Ile Pro Glu Asn Leu Gln Phe Ala 260 265
270Val Gly Gln Glu Val Phe Gly Leu Val Pro Gly Leu Met Met
Tyr Ala 275 280 285Thr Ile Trp Leu
Arg Glu His Asn Arg Val Cys Asp Ile Leu Lys Gln 290
295 300Glu His Pro Glu Trp Gly Asp Glu Gln Leu Phe Gln
Thr Ser Arg Leu305 310 315
320Ile Leu Ile Gly Glu Thr Ile Lys Ile Val Ile Glu Asp Tyr Val Gln
325 330 335His Leu Ser Gly Tyr
His Phe Lys Leu Lys Phe Asp Pro Glu Leu Leu 340
345 350Phe Asn Gln Gln Phe Gln Tyr Gln Asn Arg Ile Ala
Ser Glu Phe Asn 355 360 365Thr Leu
Tyr His Trp His Pro Leu Leu Pro Asp Thr Phe Asn Ile Glu 370
375 380Asp Gln Glu Tyr Ser Phe Lys Gln Phe Leu Tyr
Asn Asn Ser Ile Leu385 390 395
400Leu Glu His Gly Leu Thr Gln Phe Val Glu Ser Phe Thr Arg Gln Ile
405 410 415Ala Gly Arg Val
Ala Gly Gly Arg Asn Val Pro Ile Ala Val Gln Ala 420
425 430Val Ala Lys Ala Ser Ile Asp Gln Ser Arg Glu
Met Lys Tyr Gln Ser 435 440 445Leu
Asn Glu Tyr Arg Lys Arg Phe Ser Leu Lys Pro Tyr Thr Ser Phe 450
455 460Glu Glu Leu Thr Gly Glu Lys Glu Met Ala
Ala Glu Leu Lys Ala Leu465 470 475
480Tyr Ser Asp Ile Asp Val Met Glu Leu Tyr Pro Ala Leu Leu Val
Glu 485 490 495Lys Pro Arg
Pro Asp Ala Ile Phe Gly Glu Thr Met Val Glu Leu Gly 500
505 510Ala Pro Phe Ser Leu Lys Gly Leu Met Gly
Asn Pro Ile Cys Ser Pro 515 520
525Gln Tyr Trp Lys Pro Ser Thr Phe Gly Gly Glu Val Gly Phe Lys Ile 530
535 540Ile Asn Thr Ala Ser Ile Gln Ser
Leu Ile Cys Asn Asn Val Lys Gly545 550
555 560Cys Pro Phe Thr Ser Phe Asn Val Gln Asp Pro Gln
Pro Thr Lys Thr 565 570
575Ala Thr Ile Asn Ala Ser Ala Ser His Ser Arg Leu Asp Asp Ile Asn
580 585 590Pro Thr Val Leu Ile Lys
Arg Arg Ser Thr Glu Leu 595 600
User Contributions:
comments("1"); ?> comment_form("1"); ?>Inventors list |
Agents list |
Assignees list |
List by place |
Classification tree browser |
Top 100 Inventors |
Top 100 Agents |
Top 100 Assignees |
Usenet FAQ Index |
Documents |
Other FAQs |
User Contributions:
Comment about this patent or add new information about this topic:
| People who visited this patent also read: | |
| Patent application number | Title |
|---|---|
| 20130166975 | APPARATUS FOR PROTECTING AGAINST EXTERNAL ATTACK FOR PROCESSOR BASED ON ARM CORE AND METHOD USING THE SAME |
| 20130166974 | METHODS AND SYSTEMS FOR LOGIC DEVICE DEFECT TOLERANT REDUNDANCY |
| 20130166973 | STORAGE-MEDIUM DIAGNOSIS DEVICE, STORAGE-MEDIUM DIAGNOSIS METHOD |
| 20130166972 | Apparatus and Methods of Programming Memory Cells using Adjustable Charge State Level(s) |
| 20130166971 | Method And Apparatus For Encoding Channel Quality Indicator And Precoding Control Information Bits |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 | 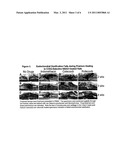 |
 |  |
 |  |
 |  |
 |  |
 |  |
| Similar patent applications: | |
| Date | Title |
|---|---|
| 2008-09-04 | Method for cell adhesion and wound healing |
| 2009-09-03 | Purification and use of a factor for supporting wound healing |
| 2009-09-03 | Use of activators of soluble guanylate cyclase for promoting wound healing |
| 2009-08-27 | Reelin deficiency or dysfunction and methods related thereto |
| 2008-12-25 | Method to promote wound healing |
| New patent applications in this class: | |
| Date | Title |
|---|---|
| 2017-08-17 | Formulations for histatin therapeutics |
| 2016-07-07 | Ointment containing a hydrolyzed fibroin and manufacturing method of same |
| 2016-05-12 | Use of recombinant ganoderma immunoregulatory protein (rlz-8) in preparation of drug for treating melanoma |
| 2016-04-21 | Carbohydrate compounds for nutritional and therapeutic use |
| 2016-04-07 | Compositions containing hc-ha complex and methods of use thereof |
| Top Inventors for class "Drug, bio-affecting and body treating compositions" | |
| Rank | Inventor's name |
|---|---|
| 1 | Anthony W. Czarnik |
| 2 | Ulrike Wachendorff-Neumann |
| 3 | Ken Chow |
| 4 | John E. Donello |
| 5 | Rajinder Singh |