Patent application title: Methods For Rapid Identification Of Pathogens In Humans And Animals
Inventors:
David J. Ecker (Encinitas, CA, US)
Richard H. Griffey (Vista, CA, US)
Rangarajan Sampath (San Diego, CA, US)
Steven A. Hofstadler (Oceanside, CA, US)
John Mcneil (La Jolla, CA, US)
Stanley T. Crooke (Carlsbad, CA, US)
Assignees:
Isis Pharmaceuticals, Inc.
IPC8 Class: AC12Q170FI
USPC Class:
435 5
Class name: Chemistry: molecular biology and microbiology measuring or testing process involving enzymes or micro-organisms; composition or test strip therefore; processes of forming such composition or test strip involving virus or bacteriophage
Publication date: 2008-12-18
Patent application number: 20080311558
Inventors list |
Agents list |
Assignees list |
List by place |
Classification tree browser |
Top 100 Inventors |
Top 100 Agents |
Top 100 Assignees |
Usenet FAQ Index |
Documents |
Other FAQs |
Patent application title: Methods For Rapid Identification Of Pathogens In Humans And Animals
Inventors:
Stanley T. Crooke
Richard H. Griffey
Rangarajan Sampath
David J. Ecker
Steven A. Hofstadler
John McNeil
Agents:
Casimir Jones, S.C.
Assignees:
Isis Pharmaceuticals, Inc.
Origin: MADISON, WI US
IPC8 Class: AC12Q170FI
USPC Class:
435 5
Abstract:
The present invention provides methods of: identifying pathogens in
biological samples from humans and animals, resolving a plurality of
etiologic agents present in samples obtained from humans and animals,
determining detailed genetic information about such pathogens or
etiologic agents, and rapid detection and identification of bioagents
from environmental, clinical or other samples.Claims:
1-22. (canceled)
23. A method of identifying a bioagent comprising the steps of: selecting at least one pair of oligonucleotide primers, wherein one member of said pair of primers hybridizes to a first conserved region of nucleic acid from said bioagent and the other member of said pair of primers hybridizes to a second conserved region of nucleic acid from said bioagent, wherein said first and second conserved regions flank a variable nucleic acid region that when amplified creates a unique base composition signature among at least five bioagents; amplifying nucleic acid from at least one said bioagent with said pair of oligonucleotide primers to produce an amplification product; determining the molecular mass of said amplification product by mass spectrometry; calculating the base composition of said amplification product from said molecular mass; comparing said base composition of at least one said bioagent to five or more calculated or measured base compositions of amplification products of known bioagents produced by using said at least one pair of oligonucleotide primers, thereby identifying said bioagent.
24. The method of claim 23, wherein said comparing comprises comparing said base composition of at least one said bioagent to nineteen or more calculated or measured base compositions of amplification products of known bioagents produced by using said at least one pair of oligonucleotide primers.
25. The method of claim 23, wherein said bioagent is a bacterium, virus, mold, fungus or parasite.
26. A method of identifying a bioagent comprising the steps of: selecting at least one pair of oligonucleotide primers, wherein one member of said pair of primers hybridizes to a first conserved region of a nucleic acid encoding a protein that participates in translation, replication, recombination, repair, transcription, nucleotide metabolism, amino acid metabolism, lipid metabolism, uptake, secretion, antibiotic resistance, virulence, or pathogenicity, and the other member of said pair of primers hybridizes to a second conserved region of said nucleic acid encoding a protein that participates in translation, replication, recombination, repair, transcription, nucleotide metabolism, amino acid metabolism, lipid metabolism, uptake, secretion, antibiotic resistance, virulence, or pathogenicity, wherein said first and second conserved regions flank a variable nucleic acid region that when amplified creates unique base composition signatures among at least five bioagents; amplifying nucleic acid from at least one said bioagent with said pair of oligonucleotide primers to produce an amplification product; determining the molecular mass of said amplification product by mass spectrometry; calculating the base composition of said amplification product from said molecular mass; comparing said base composition of at least one said bioagent to five or more calculated or measured base compositions of amplification products of known bioagents produced by using said at least one pair of oligonucleotide primers, thereby identifying said bioagent.
27. The method of claim 26, wherein said molecular masses of said amplification products are determined by ES1-TOF mass spectrometry.
28. The method of claim 26, wherein said bioagent is a bacterium, virus, mold, fungus or parasite.
29. The method of claim 26, wherein said comparing comprises comparing said base composition of at least one said bioagent to nineteen or more calculated or measured base compositions of amplification products of known bioagents produced by using said at least one pair of oligonucleotide primers.
Description:
CROSS-REFERENCE TO RELATED APPLICATIONS
[0001]The present application is a continuation of U.S. application Ser. No. 10/660,122, filed Sep. 11, 2003, which in turn: 1) is a continuation-in-part of U.S. application Ser. No. 10/323,233 filed Dec. 18, 2002; 2) is a continuation-in-part of U.S. application Ser. No. 10/326,051 filed Dec. 18, 2002; 3) is a continuation-in-part of U.S. application Ser. No. 10/325,526 filed Dec. 18, 2002; 4) is a continuation in part of U.S. application Ser. No. 10/325,527 filed Dec. 18, 2002; 5) is a continuation-in-part of U.S. application Ser. No. 09/798,007 filed Mar. 2, 2001; 6) claims the benefit of U.S. provisional application Ser. No. 60/431,319 filed Dec. 6, 2002; 7) claims the benefit of U.S. provisional application Ser. No. 60/443,443 filed Jan. 29, 2003; 8) claims the benefit of U.S. provisional application Ser. No. 60/443,788 filed Jan. 30, 2003; and 9) claims the benefit of U.S. provisional application Ser. No. 60/447,529 filed Feb. 14, 2003; each of which is incorporated herein by reference in its entirety.
FIELD OF THE INVENTION
[0003]The present invention relates generally to clinical applications of directed to the identification of pathogens in biological samples from humans and animals. The present invention is also directed to the resolution of a plurality of etiologic agents present in samples obtained from humans and animals. The invention is further directed to the determination of detailed genetic information about such pathogens or etiologic agents.
[0004]The identification of the bioagent is important for determining a proper course of treatment and/or eradication of the bioagent in such cases as biological warfare and natural infections. Furthermore, the determination of the geographic origin of a selected bioagent will facilitate the identification of potential criminal identity. The present invention also relates to methods for rapid detection and identification of bioagents from environmental, clinical or other samples. The methods provide for detection and characterization of a unique base composition signature (BCS) from any bioagent, including bacteria and viruses. The unique BCS is used to rapidly identify the bioagent.
BACKGROUND OF THE INVENTION
[0005]In the United States, hospitals report well over 5 million cases of recognized infectious disease-related illnesses annually. Significantly greater numbers remain undetected, both in the inpatient and community setting, resulting in substantial morbidity and mortality. Critical intervention for infectious disease relies on rapid, sensitive and specific detection of the offending pathogen, and is central to the mission of microbiology laboratories at medical centers. Unfortunately, despite the recognition that outcomes from infectious illnesses are directly associated with time to pathogen recognition, as well as accurate identification of the class and species of microbe, and ability to identify the presence of drug resistance isolates, conventional hospital laboratories often remain encumbered by traditional slow multi-step culture based assays. Other limitations of the conventional laboratory which have become increasingly apparent include: extremely prolonged wait-times for pathogens with long generation time (up to several weeks); requirements for additional testing and wait times for speciation and identification of antimicrobial resistance; diminished test sensitivity for patients who have received antibiotics; and absolute inability to culture certain pathogens in disease states associated with microbial infection.
[0006]For more than a decade, molecular testing has been heralded as the diagnostic tool for the new millennium, whose ultimate potential could include forced obsolescence of traditional hospital laboratories. However, despite the fact that significant advances in clinical application of PCR techniques have occurred, the practicing physician still relies principally on standard techniques. A brief discussion of several existing applications of PCR in the hospital-based setting follows.
[0007]Generally speaking molecular diagnostics have been championed for identifying organisms that cannot be grown in vitro, or in instances where existing culture techniques are insensitive and/or require prolonged incubation times. PCR-based diagnostics have been successfully developed for a wide variety of microbes. Application to the clinical arena has met with variable success, with only a few assays achieving acceptance and utility.
[0008]One of the earliest and perhaps most widely recognized applications of PCR for clinical practice is in detection of Mycobacterium tuberculosis. Clinical characteristics favoring development of a nonculture-based test for tuberculosis include week to month long delays associated with standard testing, occurrence of drug-resistant isolates and public health imperatives associated with recognition, isolation and treatment. Although frequently used as a diagnostic adjunctive, practical and routine clinical application of PCR remains problematic due to significant inter-laboratory variation in sensitivity, and inadequate specificity for use in low prevalence populations, requiring further development at the technical level. Recent advances in the laboratory suggest that identification of drug resistant isolates by amplification of mutations associated with specific antibiotic resistance (e.g., rpoB gene in rifampin resistant strains) may be forthcoming for clinical use, although widespread application will require extensive clinical validation.
[0009]One diagnostic assay, which has gained widespread acceptance, is for C. trachomatis. Conventional detection systems are limiting due to inadequate sensitivity and specificity (direct immunofluorescence or enzyme immunoassay) or the requirement for specialized culture facilities, due to the fastidious characteristics of this microbe. Laboratory development, followed by widespread clinical validation testing in a variety of acute and nonacute care settings have demonstrated excellent sensitivity (90-100%) and specificity (97%) of the PCR assay leading to its commercial development. Proven efficacy of the PCR assay from both genital and urine sampling, have resulted in its application to a variety of clinical setting, most recently including routine screening of patients considered at risk.
[0010]While the full potential for PCR diagnostics to provide rapid and critical information to physicians faced with difficult clinical-decisions has yet to be realized, one recently developed assay provides an example of the promise of this evolving technology. Distinguishing life-threatening causes of fever from more benign causes in children is a fundamental clinical dilemma faced by clinicians, particularly when infections of the central nervous system are being considered. Bacterial causes of meningitis can be highly aggressive, but generally cannot be differentiated on a clinical basis from aseptic meningitis, which is a relatively benign condition that can be managed on an outpatient basis. Existing blood culture methods often take several days to turn positive, and are often confounded by poor sensitivity or false-negative findings in patients receiving empiric antimicrobials. Testing and application of a PCR assay for enteroviral meningitis has been found to be highly sensitive. With reporting of results within 1 day, preliminary clinical trials have shown significant reductions in hospital costs, due to decreased duration of hospital stays and reduction in antibiotic therapy. Other viral PCR assays, now routinely available include those for herpes simplex virus, cytomegalovirus, hepatitis and HIV. Each has a demonstrated cost savings role in clinical practice, including detection of otherwise difficult to diagnose infections and newly realized capacity to monitor progression of disease and response to therapy, vital in the management of chronic infectious diseases.
[0011]The concept of a universal detection system has been forwarded for identification of bacterial pathogens, and speaks most directly to the possible clinical implications of a broad-based screening tool for clinical use. Exploiting the existence of highly conserved regions of DNA common to all bacterial species in a PCR assay would empower physicians to rapidly identify the presence of bacteremia, which would profoundly impact patient care. Previous empiric decision making could be abandoned in favor of educated practice, allowing appropriate and expeditious decision-making regarding need for antibiotic therapy and hospitalization.
[0012]Experimental work using the conserved features of the 16S rRNA common to almost all bacterial species, is an area of active investigation. Hospital test sites have focused on "high yield" clinical settings where expeditious identification of the presence of systemic bacterial infection has immediate high morbidity and mortality consequences. Notable clinical infections have included evaluation of febrile infants at risk for sepsis, detection of bacteremia in febrile neutropenic cancer patients, and examination of critically ill patients in the intensive care unit. While several of these studies have reported promising results (with sensitivity and specificity well over 90%), significant technical difficulties (described below) remain, and have prevented general acceptance of this assay in clinics and hospitals (which remain dependent on standard blood culture methodologies). Even the revolutionary advances of real-time PCR technique, which offers a quantitative more reproducible and technically simpler system, remains encumbered by inherent technical limitations of the PCR assay.
[0013]The principle shortcomings of applying PCR assays to the clinical setting include: inability to eliminate background DNA contamination; interference with the PCR amplification by substrates present in the reaction; and limited capacity to provide rapid reliable speciation, antibiotic resistance and subtype identification. Some laboratories have recently made progress in identifying and removing inhibitors; however background contamination remains problematic, and methods directed towards eliminating exogenous sources of DNA report significant diminution in assay sensitivity. Finally, while product identification and detailed characterization has been achieved using sequencing techniques, these approaches are laborious and time-intensive thus detracting from its clinical applicability.
[0014]Rapid and definitive microbial identification is desirable for a variety of industrial, medical, environmental, quality, and research reasons. Traditionally, the microbiology laboratory has functioned to identify the etiologic agents of infectious diseases through direct examination and culture of specimens. Since the mid-1980s, researchers have repeatedly demonstrated the practical utility of molecular biology techniques, many of which form the basis of clinical diagnostic assays. Some of these techniques include nucleic acid hybridization analysis, restriction enzyme analysis, genetic sequence analysis, and separation and purification of nucleic acids (See, e.g., J. Sambrook, E. F. Fritsch, and T. Maniatis, Molecular Cloning: A Laboratory Manual, 2nd Ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1989). These procedures, in general, are time-consuming and tedious. Another option is the polymerase chain reaction (PCR) or other amplification procedure that amplifies a specific target DNA sequence based on the flanking primers used. Finally, detection and data analysis convert the hybridization event into an analytical result.
[0015]Other not yet fully realized applications of PCR for clinical medicine is the identification of infectious causes of disease previously described as idiopathic (e.g. Bartonella henselae in bacillary angiomatosis, and Tropheryma whippellii as the uncultured bacillus associated with Whipple's disease). Further, recent epidemiological studies which suggest a strong association between Chlamydia pneumonia and coronary artery disease, serve as example of the possible widespread, yet undiscovered links between pathogen and host which may ultimately allow for new insights into pathogenesis and novel life sustaining or saving therapeutics.
[0016]For the practicing clinician, PCR technology offers a yet unrealized potential for diagnostic omnipotence in the arena of infectious disease. A universal reliable infectious disease detection system would certainly become a fundamental tool in the evolving diagnostic armamentarium of the 21st century clinician. For front line emergency physicians, or physicians working in disaster settings, a quick universal detection system, would allow for molecular triage and early aggressive targeted therapy. Preliminary clinical studies using species specific probes suggest that implementing rapid testing in acute care setting is feasible. Resources could thus be appropriately applied, and patients with suspected infections could rapidly be risk stratified to the different treatment settings, depending on the pathogen and virulence. Furthermore, links with data management systems, locally regionally and nationally, would allow for effective epidemiological surveillance, with obvious benefits for antibiotic selection and control of disease outbreaks.
[0017]For the hospitalists, the ability to speciate and subtype would allow for more precise decision-making regarding antimicrobial agents. Patients who are colonized with highly contagious pathogens could be appropriately isolated on entry into the medical setting without delay. Targeted therapy will diminish development of antibiotic resistance. Furthermore, identification of the genetic basis of antibiotic resistant strains would permit precise pharmacologic intervention. Both physician and patient would benefit with less need for repetitive testing and elimination of wait times for test results.
[0018]It is certain that the individual patient will benefit directly from this approach. Patients with unrecognized or difficult to diagnose infections would be identified and treated promptly. There will be reduced need for prolonged inpatient stays, with resultant decreases in iatrogenic events.
[0019]Mass spectrometry provides detailed information about the molecules being analyzed, including high mass accuracy. It is also a process that can be easily automated. Low-resolution MS may be unreliable when used to detect some known agents, if their spectral lines are sufficiently weak or sufficiently close to those from other living organisms in the sample. DNA chips with specific probes can only determine the presence or absence of specifically anticipated organisms. Because there are hundreds of thousands of species of benign bacteria, some very similar in sequence to threat organisms, even arrays with 10,000 probes lack the breadth needed to detect a particular organism.
[0020]Antibodies face more severe diversity limitations than arrays. If antibodies are designed against highly conserved targets to increase diversity, the false alarm problem will dominate, again because threat organisms are very similar to benign ones. Antibodies are only capable of detecting known agents in relatively uncluttered environments.
[0021]Several groups have described detection of PCR products using high resolution electrospray ionization-Fourier transform-ion cyclotron resonance mass spectrometry (ESI-FT-ICR MS). Accurate measurement of exact mass combined with knowledge of the number of at least one nucleotide allowed calculation of the total base composition for PCR duplex products of approximately 100 base pairs. (Aaserud et al., J. Am. Soc. Mass Spec., 1996, 7, 1266-1269; Muddiman et al., Anal. Chem., 1997, 69, 1543-1549; Wunschel et al., Anal. Chem., 1998, 70, 1203-1207; Muddiman et al., Rev. Anal. Chem., 1998, 17, 1-68). Electrospray ionization-Fourier transform-ion cyclotron resistance (ESI-FT-ICR) MS may be used to determine the mass of double-stranded, 500 base-pair PCR products via the average molecular mass (Hurst et al., Rapid Commun. Mass Spec. 1996, 10, 377-382). The use of matrix-assisted laser desorption ionization-time of flight (MALDI-TOF) mass spectrometry for characterization of PCR products has been described. (Muddiman et al., Rapid Commun. Mass Spec., 1999, 13, 1201-1204). However, the degradation of DNAs over about 75 nucleotides observed with MALDI limited the utility of this method.
[0022]U.S. Pat. No. 5,849,492 describes a method for retrieval of phylogenetically informative DNA sequences which comprise searching for a highly divergent segment of genomic DNA surrounded by two highly conserved segments, designing the universal primers for PCR amplification of the highly divergent region, amplifying the genomic DNA by PCR technique using universal primers, and then sequencing the gene to determine the identity of the organism.
[0023]U.S. Pat. No. 5,965,363 discloses methods for screening nucleic acids for polymorphisms by analyzing amplified target nucleic acids using mass spectrometric techniques and to procedures for improving mass resolution and mass accuracy of these methods.
[0024]WO 99/14375 describes methods, PCR primers and kits for use in analyzing preselected DNA tandem nucleotide repeat alleles by mass spectrometry.
[0025]WO 98/12355 discloses methods of determining the mass of a target nucleic acid by mass spectrometric analysis, by cleaving the target nucleic acid to reduce its length, making the target single-stranded and using MS to determine the mass of the single-stranded shortened target. Also disclosed are methods of preparing a double-stranded target nucleic acid for MS analysis comprising amplification of the target nucleic acid, binding one of the strands to a solid support, releasing the second strand and then releasing the first strand which is then analyzed by MS. Kits for target nucleic acid preparation are also provided.
[0026]PCT WO97/33000 discloses methods for detecting mutations in a target nucleic acid by nonrandomly fragmenting the target into a set of single-stranded nonrandom length fragments and determining their masses by MS.
[0027]U.S. Pat. No. 5,605,798 describes a fast and highly accurate mass spectrometer-based process for detecting the presence of a particular nucleic acid in a biological sample for diagnostic purposes.
[0028]WO 98/21066 describes processes for determining the sequence of a particular target nucleic acid by mass spectrometry. Processes for detecting a target nucleic acid present in a biological sample by PCR amplification and mass spectrometry detection are disclosed, as are methods for detecting a target nucleic acid in a sample by amplifying the target with primers that contain restriction sites and tags, extending and cleaving the amplified nucleic acid, and detecting the presence of extended product, wherein the presence of a DNA fragment of a mass different from wild-type is indicative of a mutation. Methods of sequencing a nucleic acid via mass spectrometry methods are also described.
[0029]WO 97/37041, WO 99/31278 and U.S. Pat. No. 5,547,835 describe methods of sequencing nucleic acids using mass spectrometry. U.S. Pat. Nos. 5,622,824, 5,872,003 and 5,691,141 describe methods, systems and kits for exonuclease-mediated mass spectrometric sequencing.
[0030]Thus, there is a need for a method for bioagent detection and identification which is both specific and rapid, and in which no nucleic acid sequencing is required. The present invention addresses this need.
SUMMARY OF THE INVENTION
[0031]The present invention is directed towards methods of identifying a pathogen in a biological sample by obtaining nucleic acid from a biological sample, selecting at least one pair of intelligent primers with the capability of amplification of nucleic acid of the pathogen, amplifying the nucleic acid with the primers to obtain at least one amplification product, determining the molecular mass of at least one amplification product from which the pathogen is identified. Further, this invention is directed to methods of epidemic surveillance. By identifying a pathogen from samples acquired from a plurality of geographic locations, the spread of the pathogen to a given geographic location can be determined.
[0032]The present invention is also directed to methods of diagnosis of a plurality of etiologic agents of disease in an individual by obtaining a biological sample from an individual, isolating nucleic acid from the biological sample, selecting a plurality of amplification primers with the capability of amplification of nucleic acid of a plurality of etiologic agents of disease, amplifying the nucleic acid with a plurality of primers to obtain a plurality of amplification products corresponding to a plurality of etiologic agents, determining the molecular masses of the plurality of unique amplification products which identify the members of the plurality of etiologic agents.
[0033]The present invention is also directed to methods of in silico screening of primer sets to be used in identification of a plurality of bioagents by preparing a base composition probability cloud plot from a plurality of base composition signatures of the plurality of bioagents generated in silico, inspecting the base composition probability cloud plot for overlap of clouds from different bioagents, and choosing primer sets based on minimal overlap of the clouds.
[0034]The present invention is also directed to methods of predicting the identity of a bioagent with a heretofore unknown base composition signature by preparing a base composition probability cloud plot from a plurality of base composition signatures of the plurality of bioagents which includes the heretofore unknown base composition, inspecting the base composition probability cloud for overlap of the heretofore unknown base composition with the cloud of a known bioagent such that overlap predicts that the identity of the bioagent with a heretofore unknown base composition signature equals the identity of the known bioagent.
[0035]The present invention is also directed to methods for determining a subspecies characteristic for a given pathogen in a biological sample by identifying the pathogen in a biological sample using broad range survey primers or division-wide primers, selecting at least one pair of drill-down primers to amplify nucleic acid segments which provide a subspecies characteristic about the pathogen, amplifying the nucleic acid segments to produce at least one drill-down amplification product and determining the base composition signature of the drill-down amplification product wherein the base composition signature provides a subspecies characteristic about the pathogen.
[0036]The present invention is also directed to methods of pharmacogenetic analysis by obtaining a sample of genomic DNA from an individual, selecting a segment of the genomic DNA which provides pharmacogenetic information, using at least one pair of intelligent primers to produce an amplification product which comprises the segment of genomic DNA and determining the base composition signature of the amplification product, wherein the base composition signature provides pharmacogenetic information about said individual.
BRIEF DESCRIPTION OF THE DRAWINGS
[0037]FIGS. 1A-1H and FIG. 2 are consensus diagrams that show examples of conserved regions from 16S rRNA (FIG. 1A-1, 1A-2, 1A-3, 1A-4, and 1A-5), 23S rRNA (3'-half, FIG. 1B, 1C, and 1D; 5'-half, FIG. 1E-F), 23S rRNA Domain I (FIG. 1G), 23S rRNA Domain IV (FIG. 1H) and 16S rRNA Domain III (FIG. 2) which are suitable for use in the present invention. Lines with arrows are examples of regions to which intelligent primer pairs for PCR are designed. The label for each primer pair represents the starting and ending base number of the amplified region on the consensus diagram. Bases in capital letters are greater than 95% conserved; bases in lower case letters are 90-95% conserved, filled circles are 80-90% conserved; and open circles are less than 80% conserved. The label for each primer pair represents the starting and ending base number of the amplified region on the consensus diagram. The nucleotide sequence of the 16S rRNA consensus sequence is SEQ ID NO:3 and the nucleotide sequence of the 23S rRNA consensus sequence is SEQ ID NO:4.
[0038]FIG. 2 shows a typical primer amplified region from the 16S rRNA Domain III shown in FIG. 1A-1.
[0039]FIG. 3 is a schematic diagram showing conserved regions in RNase P. Bases in capital letters are greater than 90% conserved; bases in lower case letters are 80-90% conserved; filled circles designate bases which are 70-80% conserved; and open circles designate bases that are less than 70% conserved.
[0040]FIG. 4 is a schematic diagram of base composition signature determination using nucleotide analog "tags" to determine base composition signatures.
[0041]FIG. 5 shows the deconvoluted mass spectra of a Bacillus anthracis region with and without the mass tag phosphorothioate A (A*). The two spectra differ in that the measured molecular weight of the mass tag-containing sequence is greater than the unmodified sequence.
[0042]FIG. 6 shows base composition signature (BCS) spectra from PCR products from Staphylococcus aureus (S. aureus 16S--1337F) and Bacillus anthracis (B. anthr. 16S--1337F), amplified using the same primers. The two strands differ by only two (AT-->CG) substitutions and are clearly distinguished on the basis of their BCS.
[0043]FIG. 7 shows that a single difference between two sequences (A14 in B. anthracis vs. A15 in B. cereus) can be easily detected using ESI-TOF mass spectrometry.
[0044]FIG. 8 is an ESI-TOF of Bacillus anthracis spore coat protein sspE 56 mer plus calibrant. The signals unambiguously identify B. anthracis versus other Bacillus species.
[0045]FIG. 9 is an ESI-TOF of a B. anthracis synthetic 16S--1228 duplex (reverse and forward strands). The technique easily distinguishes between the forward and reverse strands.
[0046]FIG. 10 is an ESI-FTICR-MS of a synthetic B. anthracis 16S--1337 46 base pair duplex.
[0047]FIG. 11 is an ESI-TOF-MS of a 56 mer oligonucleotide (3 scans) from the B. anthracis saspB gene with an internal mass standard. The internal mass standards are designated by asterisks.
[0048]FIG. 12 is an ESI-TOF-MS of an internal standard with 5 mM TBA-TFA buffer showing that charge stripping with tributylammonium trifluoroacetate reduces the most abundant charge state from [M-8H+]8- to [M-3H+]3-.
[0049]FIG. 13 is a portion of a secondary structure defining database according to one embodiment of the present invention, where two examples of selected sequences are displayed graphically thereunder.
[0050]FIG. 14 is a three dimensional graph demonstrating the grouping of sample molecular weight according to species.
[0051]FIG. 15 is a three dimensional graph demonstrating the grouping of sample molecular weights according to species of virus and mammal infected.
[0052]FIG. 16 is a three dimensional graph demonstrating the grouping of sample molecular weights according to species of virus, and animal-origin of infectious agent.
[0053]FIG. 17 is a figure depicting how the triangulation method of the present invention provides for the identification of an unknown bioagent without prior knowledge of the unknown agent. The use of different primer sets to distinguish and identify the unknown is also depicted as primer sets I, II and III within this figure. A three dimensional graph depicts all of bioagent space (170), including the unknown bioagent, which after use of primer set I (171) according to a method according to the present invention further differentiates and classifies bioagents according to major classifications (176) which, upon further analysis using primer set II (172) differentiates the unknown agent (177) from other, known agents (173) and finally, the use of a third primer set (175) further specifies subgroups within the family of the unknown (174).
[0054]FIG. 18 shows a representative base composition probability cloud for a region of the RNA polymerase B gene from a cluster of enterobacteria. The dark spheres represent the actual base composition of the organisms. The lighter spheres represent the transitions among base compositions observed in different isolates of the same species of organism.
[0055]FIG. 19 shows resolution of enterobacteriae members with primers targeting RNA polymerase B (rpoB). A single pair of primers targeting a hyper-variable region within rpoB was sufficient to resolve most members of this group at the genus level (Salmonella from Escherichia from Yersinia) as well as the species/strain level (E. coli K12 from O157). All organisms with the exception of Y. pestis were tested in the lab and the measured base counts (shown with arrow) matched the predictions in every case.
[0056]FIG. 20 shows detection of S. aureus in blood. Spectra on the right indicate signals corresponding to S. aureus detection in spiked wells A1 and A4 with no detection in control wells A2 and A3.
[0057]FIG. 21 shows a representative base composition distribution of human adenovirus strain types for a single primer pair region on the hexon gene. The circles represent different adenovirus sequences in our database that were used for primer design. Measurement of masses and base counts for each of the unknown samples A, B, C and D matched one or more of the known groups of adenoviruses.
[0058]FIG. 22 shows a representative broad range survey/drill-down process as applied to emm-typing of streptococcus pyogenes (Group A Streptococcus: GAS). Genetic material is extracted (201) and amplified using broad range survey primers (202). The amplification products are analyzed (203) to determine the presence and identity of bioagents at the species level. If Streptococcus pyogenes is detected (204), the emm-typing "drill-down" primers are used to reexamine the extract to identify the emm-type of the sample (205). Different sets of drill down primers can be employed to determine a subspecies characteristic for various strains of various bioagents (206).
[0059]FIG. 23 shows a representative base composition distribution of bioagents detected in throat swabs from military personnel using a broad range primer pair directed to 16S rRNA.
[0060]FIG. 24 shows a representative deconvoluted ESI-FTICR spectra of the PCR products produced by the gtr primer for samples 12 (top) and 10 (bottom) corresponding to emm types 3 and 6, respectively. Accurate mass measurements were obtained by using an internal mass standard and post-calibrating each spectrum; the experimental mass measurement uncertainty on each strand is +0.035 Daltons (1 ppm). Unambiguous base compositions of the amplicons were determined by calculating all putative base compositions of each stand within the measured mass (and measured mass uncertainty) and selecting complementary pairs within the mass measurement uncertainty. In all cases there was only one base composition within 25 ppm. The measured mass difference of 15.985 Da between the strands shown on the left is in excellent agreement with the theoretical mass difference of 15.994 Da expected for an A to G substitution.
[0061]FIG. 25 shows representative results of the base composition analysis on throat swab samples using the six primer pairs, 5'-emm gene sequencing and the MLST gene sequencing method of the present invention for an outbreak of Streptococcus pyogenes (group A streptococcus; GAS) at a military training camp.
[0062]FIG. 26 shows: a) a representative ESI-FTICR mass spectrum of a restriction digest of a 986 bp region of the 16S ribosomal gene from E. coli K12 digested with a mixture of BstNI, BsmFI, BfaI, and NcoI; b) a deconvoluted representation (neutral mass) of the above spectrum showing the base compositions derived from accurate mass measurements of each fragment; and c) a representative reconstructed restriction map showing complete base composition coverage for nucleotides 1-856. The NcoI did not cut.
[0063]FIG. 27 shows a representative base composition distribution of poxviruses for a single primer pair region on the DNA-dependent polymerase B gene (DdDpB). The spheres represent different poxvirus sequences that were used for primer design.
DESCRIPTION OF EMBODIMENTS
A. Introduction
[0064]The present invention provides, inter alia, methods for detection and identification of bioagents in an unbiased manner using "bioagent identifying amplicons." "Intelligent primers" are selected to hybridize to conserved sequence regions of nucleic acids derived from a bioagent and which bracket variable sequence regions to yield a bioagent identifying amplicon which can be amplified and which is amenable to molecular mass determination. The molecular mass then provides a means to uniquely identify the bioagent without a requirement for prior knowledge of the possible identity of the bioagent. The molecular mass or corresponding "base composition signature" (BCS) of the amplification product is then matched against a database of molecular masses or base composition signatures. Furthermore, the method can be applied to rapid parallel "multiplex" analyses, the results of which can be employed in a triangulation identification strategy. The present method provides rapid throughput and does not require nucleic acid sequencing of the amplified target sequence for bioagent detection and identification.
B. Bioagents
[0065]In the context of this invention, a "bioagent" is any organism, cell, or virus, living or dead, or a nucleic acid derived from such an organism, cell or virus. Examples of bioagents include, but are not limited, to cells, including but not limited to, cells, including but not limited to human clinical samples, bacterial cells and other pathogens) viruses, fungi, and protists, parasites, and pathogenicity markers (including but not limited to: pathogenicity islands, antibiotic resistance genes, virulence factors, toxin genes and other bioregulating compounds). Samples may be alive or dead or in a vegetative state (for example, vegetative bacteria or spores) and may be encapsulated or bioengineered. In the context of this invention, a "pathogen" is a bioagent which causes a disease or disorder.
[0066]Despite enormous biological diversity, all forms of life on earth share sets of essential, common features in their genomes. Bacteria, for example have highly conserved sequences in a variety of locations on their genomes. Most notable is the universally conserved region of the ribosome. But there are also conserved elements in other non-coding RNAs, including RNAse P and the signal recognition particle (SRP) among others. Bacteria have a common set of absolutely required genes. About 250 genes are present in all bacterial species (Proc. Natl. Acad. Sci. U.S.A., 1996, 93, 10268; Science, 1995, 270, 397), including tiny genomes like Mycoplasma, Ureaplasma and Rickettsia. These genes encode proteins involved in translation, replication, recombination and repair, transcription, nucleotide metabolism, amino acid metabolism, lipid metabolism, energy generation, uptake, secretion and the like. Examples of these proteins are DNA polymerase III beta, elongation factor TU, heat shock protein groEL, RNA polymerase beta, phosphoglycerate kinase, NADH dehydrogenase, DNA ligase, DNA topoisomerase and elongation factor G. Operons can also be targeted using the present method. One example of an operon is the bfp operon from enteropathogenic E. coli. Multiple core chromosomal genes can be used to classify bacteria at a genus or genus species level to determine if an organism has threat potential. The methods can also be used to detect pathogenicity markers (plasmid or chromosomal) and antibiotic resistance genes to confirm the threat potential of an organism and to direct countermeasures.
C. Selection of "Bioagent Identifying Amplicons"
[0067]Since genetic data provide the underlying basis for identification of bioagents by the methods of the present invention, it is necessary to select segments of nucleic acids which ideally provide enough variability to distinguish each individual bioagent and whose molecular mass is amenable to molecular mass determination. In one embodiment of the present invention, at least one polynucleotide segment is amplified to facilitate detection and analysis in the process of identifying the bioagent. Thus, the nucleic acid segments which provide enough variability to distinguish each individual bioagent and whose molecular masses are amenable to molecular mass determination are herein described as "bioagent identifying amplicons." The term "amplicon" as used herein, refers to a segment of a polynucleotide which is amplified in an amplification reaction.
[0068]As used herein, "intelligent primers" are primers that are designed to bind to highly conserved sequence regions that flank an intervening variable region and yield amplification products which ideally provide enough variability to distinguish each individual bioagent, and which are amenable to molecular mass analysis. By the term "highly conserved," it is meant that the sequence regions exhibit between about 80-100%, or between about 90-100%, or between about 95-100% identity. The molecular mass of a given amplification product provides a means of identifying the bioagent from which it was obtained, due to the variability of the variable region. Thus design of intelligent primers requires selection of a variable region with appropriate variability to resolve the identity of a given bioagent.
[0069]In one embodiment, the bioagent identifying amplicon is a portion of a ribosomal RNA (rRNA) gene sequence. With the complete sequences of many of the smallest microbial genomes now available, it is possible to identify a set of genes that defines "minimal life" and identify composition signatures that uniquely identify each gene and organism. Genes that encode core life functions such as DNA replication, transcription, ribosome structure, translation, and transport are distributed broadly in the bacterial genome and are suitable regions for selection of bioagent identifying amplicons. Ribosomal RNA (rRNA) genes comprise regions that provide useful base composition signatures. Like many genes involved in core life functions, rRNA genes contain sequences that are extraordinarily conserved across bacterial domains interspersed with regions of high variability that are more specific to each species. The variable regions can be utilized to build a database of base composition signatures. The strategy involves creating a structure-based alignment of sequences of the small (16S) and the large (23S) subunits of the rRNA genes. For example, there are currently over 13,000 sequences in the ribosomal RNA database that has been created and maintained by Robin Gutell, University of Texas at Austin, and is publicly available on the Institute for Cellular and Molecular Biology web page on the world wide web of the Internet at, for example, "rna.icmb.utexas.edu/." There is also a publicly available rRNA database created and maintained by the University of Antwerp, Belgium on the world wide web of the Internet at, for example, "rrna.uia.ac.be."
[0070]These databases have been analyzed to determine regions that are useful as bioagent identifying amplicons. The characteristics of such regions include: a) between about 80 and 100%, or greater than about 95% identity among species of the particular bioagent of interest, of upstream and downstream nucleotide sequences which serve as sequence amplification primer sites; b) an intervening variable region which exhibits no greater than about 5% identity among species; and c) a separation of between about 30 and 1000 nucleotides, or no more than about 50-250 nucleotides, or no more than about 60-100 nucleotides, between the conserved regions.
[0071]As a non-limiting example, for identification of Bacillus species, the conserved sequence regions of the chosen bioagent identifying amplicon must be highly conserved among all Bacillus species while the variable region of the bioagent identifying amplicon is sufficiently variable such that the molecular masses of the amplification products of all species of Bacillus are distinguishable.
[0072]Bioagent identifying amplicons amenable to molecular mass determination are either of a length, size or mass compatible with the particular mode of molecular mass determination or compatible with a means of providing a predictable fragmentation pattern in order to obtain predictable fragments of a length compatible with the particular mode of molecular mass determination. Such means of providing a predictable fragmentation pattern of an amplification product include, but are not limited to, cleavage with restriction enzymes or cleavage primers, for example.
[0073]Identification of bioagents can be accomplished at different levels using intelligent primers suited to resolution of each individual level of identification. "Broad range survey" intelligent primers are designed with the objective of identifying a bioagent as a member of a particular division of bioagents. A "bioagent division" is defined as group of bioagents above the species level and includes but is not limited to: orders, families, classes, clades, genera or other such groupings of bioagents above the species level. As a non-limiting example, members of the Bacillus/Clostridia group or gamma-proteobacteria group may be identified as such by employing broad range survey intelligent primers such as primers which target 16S or 23S ribosomal RNA.
[0074]In some embodiments, broad range survey intelligent primers are capable of identification of bioagents at the species level. One main advantage of the detection methods of the present invention is that the broad range survey intelligent primers need not be specific for a particular bacterial species, or even genus, such as Bacillus or Streptomyces. Instead, the primers recognize highly conserved regions across hundreds of bacterial species including, but not limited to, the species described herein. Thus, the same broad range survey intelligent primer pair can be used to identify any desired bacterium because it will bind to the conserved regions that flank a variable region specific to a single species, or common to several bacterial species, allowing unbiased nucleic acid amplification of the intervening sequence and determination of its molecular weight and base composition. For example, the 16S--971-1062, 16S--1228-1310 and 16S--1100-1188 regions are 98-99% conserved in about 900 species of bacteria (16S=16S rRNA, numbers indicate nucleotide position). In one embodiment of the present invention, primers used in the present method bind to one or more of these regions or portions thereof.
[0075]Due to their overall conservation, the flanking rRNA primer sequences serve as good intelligent primer binding sites to amplify the nucleic acid region of interest for most, if not all, bacterial species. The intervening region between the sets of primers varies in length and/or composition, and thus provides a unique base composition signature. Examples of intelligent primers that amplify regions of the 16S and 23S rRNA are shown in FIGS. 1A-1H. A typical primer amplified region in 16S rRNA is shown in FIG. 2. The arrows represent primers that bind to highly conserved regions which flank a variable region in 16S rRNA domain III. The amplified region is the stem-loop structure under "1100-1188." It is advantageous to design the broad range survey intelligent primers to minimize the number of primers required for the analysis, and to allow detection of multiple members of a bioagent division using a single pair of primers. The advantage of using broad range survey intelligent primers is that once a bioagent is broadly identified, the process of further identification at species and sub-species levels is facilitated by directing the choice of additional intelligent primers.
[0076]Division-wide" intelligent primers are designed with an objective of identifying a bioagent at the species level. As a non-limiting example, a Bacillus anthracis, Bacillus cereus and Bacillus thuringiensis can be distinguished from each other using division-wide intelligent primers. Division-wide intelligent primers are not always required for identification at the species level because broad range survey intelligent primers may provide sufficient identification resolution to accomplishing this identification objective.
[0077]Drill-down" intelligent primers are designed with an objective of identifying a sub-species characteristic of a bioagent. A "sub-species characteristic" is defined as a property imparted to a bioagent at the sub-species level of identification as a result of the presence or absence of a particular segment of nucleic acid. Such sub-species characteristics include, but are not limited to, strains, sub-types, pathogenicity markers such as antibiotic resistance genes, pathogenicity islands, toxin genes and virulence factors. Identification of such sub-species characteristics is often critical for determining proper clinical treatment of pathogen infections.
Chemical Modifications of Intelligent Primers
[0078]Ideally, intelligent primer hybridization sites are highly conserved in order to facilitate the hybridization of the primer. In cases where primer hybridization is less efficient due to lower levels of conservation of sequence, intelligent primers can be chemically modified to improve the efficiency of hybridization.
[0079]For example, because any variation (due to codon wobble in the 3rd position) in these conserved regions among species is likely to occur in the third position of a DNA triplet, oligonucleotide primers can be designed such that the nucleotide corresponding to this position is a base which can bind to more than one nucleotide, referred to herein as a "universal base." For example, under this "wobble" pairing, inosine (I) binds to U, C or A; guanine (G) binds to U or C, and uridine (U) binds to U or C. Other examples of universal bases include nitroindoles such as 5-nitroindole or 3-nitropyrrole (Loakes et al., Nucleosides and Nucleotides, 1995, 14, 1001-1003), the degenerate nucleotides dP or dK (Hill et al.), an acyclic nucleoside analog containing 5-nitroindazole (Van Aerschot et al., Nucleosides and Nucleotides, 1995, 14, 1053-1056) or the purine analog 1-(2-deoxy-β-D-ribofuranosyl)-imidazole-4-carboxamide (Sala et al., Nucl. Acids Res., 1996, 24, 3302-3306).
[0080]In another embodiment of the invention, to compensate for the somewhat weaker binding by the "wobble" base, the oligonucleotide primers are designed such that the first and second positions of each triplet are occupied by nucleotide analogs which bind with greater affinity than the unmodified nucleotide. Examples of these analogs include, but are not limited to, 2,6-diaminopurine which binds to thymine, propyne T which binds to adenine and propyne C and phenoxazines, including G-clamp, which binds to G. Propynylated pyrimidines are described in U.S. Pat. Nos. 5,645,985, 5,830,653 and 5,484,908, each of which is commonly owned and incorporated herein by reference in its entirety. Propynylated primers are claimed in U.S. Ser. No. 10/294,203 which is also commonly owned and incorporated herein by reference in entirety. Phenoxazines are described in U.S. Pat. Nos. 5,502,177, 5,763,588, and 6,005,096, each of which is incorporated herein by reference in its entirety. G-clamps are described in U.S. Pat. Nos. 6,007,992 and 6,028,183, each of which is incorporated herein by reference in its entirety.
D. Characterization of Bioagent Identifying Amplicons
[0081]A theoretically ideal bioagent detector would identify, quantify, and report the complete nucleic acid sequence of every bioagent that reached the sensor. The complete sequence of the nucleic acid component of a pathogen would provide all relevant information about the threat, including its identity and the presence of drug-resistance or pathogenicity markers. This ideal has not yet been achieved. However, the present invention provides a straightforward strategy for obtaining information with the same practical value based on analysis of bioagent identifying amplicons by molecular mass determination.
[0082]In some cases, a molecular mass of a given bioagent identifying amplicon alone does not provide enough resolution to unambiguously identify a given bioagent. For example, the molecular mass of the bioagent identifying amplicon obtained using the intelligent primer pair "16S--971" would be 55622 Da for both E. coli and Salmonella typhimurium. However, if additional intelligent primers are employed to analyze additional bioagent identifying amplicons, a "triangulation identification" process is enabled. For example, the "16S--1100" intelligent primer pair yields molecular masses of 55009 and 55005 Da for E. coli and Salmonella typhimurium, respectively. Furthermore, the "23S--855" intelligent primer pair yields molecular masses of 42656 and 42698 Da for E. coli and Salmonella typhimurium, respectively. In this basic example, the second and third intelligent primer pairs provided the additional "fingerprinting" capability or resolution to distinguish between the two bioagents.
[0083]In another embodiment, the triangulation identification process is pursued by measuring signals from a plurality of bioagent identifying amplicons selected within multiple core genes. This process is used to reduce false negative and false positive signals, and enable reconstruction of the origin of hybrid or otherwise engineered bioagents. In this process, after identification of multiple core genes, alignments are created from nucleic acid sequence databases. The alignments are then analyzed for regions of conservation and variation, and bioagent identifying amplicons are selected to distinguish bioagents based on specific genomic differences. For example, identification of the three part toxin genes typical of B. anthracis (Bowen et al., J. Appl. Microbiol., 1999, 87, 270-278) in the absence of the expected signatures from the B. anthracis genome would suggest a genetic engineering event.
[0084]The triangulation identification process can be pursued by characterization of bioagent identifying amplicons in a massively parallel fashion using the polymerase chain reaction (PCR), such as multiplex PCR, and mass spectrometric (MS) methods. Sufficient quantities of nucleic acids should be present for detection of bioagents by MS. A wide variety of techniques for preparing large amounts of purified nucleic acids or fragments thereof are well known to those of skill in the art. PCR requires one or more pairs of oligonucleotide primers that bind to regions which flank the target sequence(s) to be amplified. These primers prime synthesis of a different strand of DNA, with synthesis occurring in the direction of one primer towards the other primer. The primers, DNA to be amplified, a thermostable DNA polymerase (e.g. Taq polymerase), the four deoxynucleotide triphosphates, and a buffer are combined to initiate DNA synthesis. The solution is denatured by heating, then cooled to allow annealing of newly added primer, followed by another round of DNA synthesis. This process is typically repeated for about 30 cycles, resulting in amplification of the target sequence.
[0085]Although the use of PCR is suitable, other nucleic acid amplification techniques may also be used, including ligase chain reaction (LCR) and strand displacement amplification (SDA). The high-resolution MS technique allows separation of bioagent spectral lines from background spectral lines in highly cluttered environments.
[0086]In another embodiment, the detection scheme for the PCR products generated from the bioagent(s) incorporates at least three features. First, the technique simultaneously detects and differentiates multiple (generally about 6-10) PCR products. Second, the technique provides a molecular mass that uniquely identifies the bioagent from the possible primer sites. Finally, the detection technique is rapid, allowing multiple PCR reactions to be run in parallel.
E. Mass Spectrometric Characterization of Bioagent Identifying Amplicons
[0087]Mass spectrometry (MS)-based detection of PCR products provides a means for determination of BCS which has several advantages. MS is intrinsically a parallel detection scheme without the need for radioactive or fluorescent labels, since every amplification product is identified by its molecular mass. The current state of the art in mass spectrometry is such that less than femtomole quantities of material can be readily analyzed to afford information about the molecular contents of the sample. An accurate assessment of the molecular mass of the material can be quickly obtained, irrespective of whether the molecular weight of the sample is several hundred, or in excess of one hundred thousand atomic mass units (amu) or Daltons. Intact molecular ions can be generated from amplification products using one of a variety of ionization techniques to convert the sample to gas phase. These ionization methods include, but are not limited to, electrospray ionization (ES), matrix-assisted laser desorption ionization (MALDI) and fast atom bombardment (FAB). For example, MALDI of nucleic acids, along with examples of matrices for use in MALDI of nucleic acids, are described in WO 98/54751 (Genetrace, Inc.).
[0088]In some embodiments, large DNAs and RNAs, or large amplification products therefrom, can be digested with restriction endonucleases prior to ionization. Thus, for example, an amplification product that was 10 kDa could be digested with a series of restriction endonucleases to produce a panel of, for example, 100 Da fragments. Restriction endonucleases and their sites of action are well known to the skilled artisan. In this manner, mass spectrometry can be performed for the purposes of restriction mapping.
[0089]Upon ionization, several peaks are observed from one sample due to the formation of ions with different charges. Averaging the multiple readings of molecular mass obtained from a single mass spectrum affords an estimate of molecular mass of the bioagent. Electrospray ionization mass spectrometry (ESI-MS) is particularly useful for very high molecular weight polymers such as proteins and nucleic acids having molecular weights greater than 10 kDa, since it yields a distribution of multiply-charged molecules of the sample without causing a significant amount of fragmentation.
[0090]The mass detectors used in the methods of the present invention include, but are not limited to, Fourier transform ion cyclotron resonance mass spectrometry (FT-ICR-MS), ion trap, quadrupole, magnetic sector, time of flight (TOF), Q-TOF, and triple quadrupole.
[0091]In general, the mass spectrometric techniques which can be used in the present invention include, but are not limited to, tandem mass spectrometry, infrared multiphoton dissociation and pyrolytic gas chromatography mass spectrometry (PGC-MS). In one embodiment of the invention, the bioagent detection system operates continually in bioagent detection mode using pyrolytic GC-MS without PCR for rapid detection of increases in biomass (for example, increases in fecal contamination of drinking water or of germ warfare agents). To achieve minimal latency, a continuous sample stream flows directly into the PGC-MS combustion chamber. When an increase in biomass is detected, a PCR process is automatically initiated. Bioagent presence produces elevated levels of large molecular fragments from, for example, about 100-7,000 Da which are observed in the PGC-MS spectrum. The observed mass spectrum is compared to a threshold level and when levels of biomass are determined to exceed a predetermined threshold, the bioagent classification process described hereinabove (combining PCR and MS, such as FT-ICR MS) is initiated. Optionally, alarms or other processes (halting ventilation flow, physical isolation) are also initiated by this detected biomass level.
[0092]The accurate measurement of molecular mass for large DNAs is limited by the adduction of cations from the PCR reaction to each strand, resolution of the isotopic peaks from natural abundance 13C and 15N isotopes, and assignment of the charge state for any ion. The cations are removed by in-line dialysis using a flow-through chip that brings the solution containing the PCR products into contact with a solution containing ammonium acetate in the presence of an electric field gradient orthogonal to the flow. The latter two problems are addressed by operating with a resolving power of >100,000 and by incorporating isotopically depleted nucleotide triphosphates into the DNA. The resolving power of the instrument is also a consideration. At a resolving power of 10,000, the modeled signal from the [M-14H+]14- charge state of an 84 mer PCR product is poorly characterized and assignment of the charge state or exact mass is impossible. At a resolving power of 33,000, the peaks from the individual isotopic components are visible. At a resolving power of 100,000, the isotopic peaks are resolved to the baseline and assignment of the charge state for the ion is straightforward. The [13C,15N]-depleted triphosphates are obtained, for example, by growing microorganisms on depleted media and harvesting the nucleotides (Batey et al., Nucl. Acids Res., 1992, 20, 4515-4523).
[0093]While mass measurements of intact nucleic acid regions are believed to be adequate to determine most bioagents, tandem mass spectrometry (MSn) techniques may provide more definitive information pertaining to molecular identity or sequence. Tandem MS involves the coupled use of two or more stages of mass analysis where both the separation and detection steps are based on mass spectrometry. The first stage is used to select an ion or component of a sample from which further structural information is to be obtained. The selected ion is then fragmented using, e.g., blackbody irradiation, infrared multiphoton dissociation, or collisional activation. For example, ions generated by electrospray ionization (ESI) can be fragmented using IR multiphoton dissociation. This activation leads to dissociation of glycosidic bonds and the phosphate backbone, producing two series of fragment ions, called the w-series (having an intact 3' terminus and a 5' phosphate following internal cleavage) and the α-Base series (having an intact 5' terminus and a 3' furan).
[0094]The second stage of mass analysis is then used to detect and measure the mass of these resulting fragments of product ions. Such ion selection followed by fragmentation routines can be performed multiple times so as to essentially completely dissect the molecular sequence of a sample.
[0095]If there are two or more targets of similar molecular mass, or if a single amplification reaction results in a product which has the same mass as two or more bioagent reference standards, they can be distinguished by using mass-modifying "tags." In this embodiment of the invention, a nucleotide analog or "tag" is incorporated during amplification (e.g., a 5-(trifluoromethyl) deoxythymidine triphosphate) which has a different molecular weight than the unmodified base so as to improve distinction of masses. Such tags are described in, for example, PCT WO97/33000, which is incorporated herein by reference in its entirety. This further limits the number of possible base compositions consistent with any mass. For example, 5-(trifluoromethyl)deoxythymidine triphosphate can be used in place of dTTP in a separate nucleic acid amplification reaction. Measurement of the mass shift between a conventional amplification product and the tagged product is used to quantitate the number of thymidine nucleotides in each of the single strands. Because the strands are complementary, the number of adenosine nucleotides in each strand is also determined.
[0096]In another amplification reaction, the number of G and C residues in each strand is determined using, for example, the cytidine analog 5-methylcytosine (5-meC) or propyne C. The combination of the A/T reaction and G/C reaction, followed by molecular weight determination, provides a unique base composition. This method is summarized in FIG. 4 and Table 1.
TABLE-US-00001 TABLE 1 Total Total Total Base Base base base Double Single mass info info comp. comp. strand strand this this other Top Bottom Mass tag sequence Sequence strand strand strand strand strand T*mass T*ACGT*ACGT* T*ACGT*ACGT* 3x 3T 3A 3T 3A (T*-T) = x AT*GCAT*GCA 2A 2T 2C 2G 2G 2C AT*GCAT*GCA 2x 2T 2A C*mass TAC*GTAC*GT TAC*GTAC*GT 2x 2C 2G (C*-C) = y ATGC*ATGC*A ATGC*ATGC*A 2x 2C 2G
[0097]The mass tag phosphorothioate A (A*) was used to distinguish a Bacillus anthracis cluster. The B. anthracis (A14G9C14T9) had an average MW of 14072.26, and the B. anthracis (A1A*13G9C14T9) had an average molecular weight of 14281.11 and the phosphorothioate A had an average molecular weight of +16.06 as determined by ESI-TOF MS. The deconvoluted spectra are shown in FIG. 5.
[0098]In another example, assume the measured molecular masses of each strand are 30,000.115Da and 31,000.115 Da respectively, and the measured number of dT and dA residues are (30,28) and (28,30). If the molecular mass is accurate to 100 ppm, there are 7 possible combinations of dG+dC possible for each strand. However, if the measured molecular mass is accurate to 10 ppm, there are only 2 combinations of dG+dC, and at 1 ppm accuracy there is only one possible base composition for each strand.
[0099]Signals from the mass spectrometer may be input to a maximum-likelihood detection and classification algorithm such as is widely used in radar signal processing. The detection processing uses matched filtering of BCS observed in mass-basecount space and allows for detection and subtraction of signatures from known, harmless organisms, and for detection of unknown bioagent threats. Comparison of newly observed bioagents to known bioagents is also possible, for estimation of threat level, by comparing their BCS to those of known organisms and to known forms of pathogenicity enhancement, such as insertion of antibiotic resistance genes or toxin genes.
[0100]Processing may end with a Bayesian classifier using log likelihood ratios developed from the observed signals and average background levels. The program emphasizes performance predictions culminating in probability-of-detection versus probability-of-false-alarm plots for conditions involving complex backgrounds of naturally occurring organisms and environmental contaminants. Matched filters consist of a priori expectations of signal values given the set of primers used for each of the bioagents. A genomic sequence database (e.g. GenBank) is used to define the mass basecount matched filters. The database contains known threat agents and benign background organisms. The latter is used to estimate and subtract the signature produced by the background organisms. A maximum likelihood detection of known background organisms is implemented using matched filters and a running-sum estimate of the noise covariance. Background signal strengths are estimated and used along with the matched filters to form signatures which are then subtracted. The maximum likelihood process is applied to this "cleaned up" data in a similar manner employing matched filters for the organisms and a running-sum estimate of the noise-covariance for the cleaned up data.
F. Base Composition Signatures as Indices of Bioagent Identifying Amplicons
[0101]Although the molecular mass of amplification products obtained using intelligent primers provides a means for identification of bioagents, conversion of molecular mass data to a base composition signature is useful for certain analyses. As used herein, a "base composition signature" (BCS) is the exact base composition determined from the molecular mass of a bioagent identifying amplicon. In one embodiment, a BCS provides an index of a specific gene in a specific organism.
[0102]Base compositions, like sequences, vary slightly from isolate to isolate within species. It is possible to manage this diversity by building "base composition probability clouds" around the composition constraints for each species. This permits identification of organisms in a fashion similar to sequence analysis. A "pseudo four-dimensional plot" can be used to visualize the concept of base composition probability clouds (FIG. 18). Optimal primer design requires optimal choice of bioagent identifying amplicons and maximizes the separation between the base composition signatures of individual bioagents. Areas where clouds overlap indicate regions that may result in a misclassification, a problem which is overcome by selecting primers that provide information from different bioagent identifying amplicons, ideally maximizing the separation of base compositions. Thus, one aspect of the utility of an analysis of base composition probability clouds is that it provides a means for screening primer sets in order to avoid potential misclassifications of BCS and bioagent identity. Another aspect of the utility of base composition probability clouds is that they provide a means for predicting the identity of a bioagent whose exact measured BCS was not previously observed and/or indexed in a BCS database due to evolutionary transitions in its nucleic acid sequence.
[0103]It is important to note that, in contrast to probe-based techniques, mass spectrometry determination of base composition does not require prior knowledge of the composition in order to make the measurement, only to interpret the results. In this regard, the present invention provides bioagent classifying information similar to DNA sequencing and phylogenetic analysis at a level sufficient to detect and identify a given bioagent. Furthermore, the process of determination of a previously unknown BCS for a given bioagent (for example, in a case where sequence information is unavailable) has downstream utility by providing additional bioagent indexing information with which to populate BCS databases. The process of future bioagent identification is thus greatly improved as more BCS indexes become available in the BCS databases.
[0104]Another embodiment of the present invention is a method of surveying bioagent samples that enables detection and identification of all bacteria for which sequence information is available using a set of twelve broad-range intelligent PCR primers. Six of the twelve primers are "broad range survey primers" herein defined as primers targeted to broad divisions of bacteria (for example, the Bacillus/Clostridia group or gamma-proteobacteria). The other six primers of the group of twelve primers are "division-wide" primers herein defined as primers which provide more focused coverage and higher resolution. This method enables identification of nearly 100% of known bacteria at the species level. A further example of this embodiment of the present invention is a method herein designated "survey/drill-down" wherein a subspecies characteristic for detected bioagents is obtained using additional primers. Examples of such a subspecies characteristic include but are not limited to: antibiotic resistance, pathogenicity island, virulence factor, strain type, sub-species type, and clade group. Using the survey/drill-down method, bioagent detection, confirmation and a subspecies characteristic can be provided within hours. Moreover, the survey/drill-down method can be focused to identify bioengineering events such as the insertion of a toxin gene into a bacterial species that does not normally make the toxin.
G. Fields of Application of the Present Invention
[0105]The present methods allow extremely rapid and accurate detection and identification of bioagents compared to existing methods. Furthermore, this rapid detection and identification is possible even when sample material is impure. The methods leverage ongoing biomedical research in virulence, pathogenicity, drug resistance and genome sequencing into a method which provides greatly improved sensitivity, specificity and reliability compared to existing methods, with lower rates of false positives. Thus, the methods are useful in a wide variety of fields, including, but not limited to, those fields discussed below.
[0106]1. Identification of Pathogens in Humans and Animals
[0107]In other embodiments of the invention, the methods disclosed herein can identify infectious agents in biological samples. At least a first biological sample containing at least a first unidentified infectious agent is obtained. An identification analysis is carried out on the sample, whereby the first infectious agent in the first biological sample is identified. More particularly, a method of identifying an infectious agent in a biological entity is provided. An identification analysis is carried out on a first biological sample obtained from the biological entity, whereby at least one infectious agent in the biological sample from the biological entity is identified. The obtaining and the performing steps are, optionally, repeated on at least one additional biological sample from the biological entity.
[0108]The present invention also provides methods of identifying an infectious agent that is potentially the cause of a health condition in a biological entity. An identification analysis is carried out on a first test sample from a first infectious agent differentiating area of the biological entity, whereby at least one infectious agent is identified. The obtaining and the performing steps are, optionally, repeated on an additional infectious agent differentiating area of the biological entity.
[0109]Biological samples include, but are not limited to, hair, mucosa, skin, nail, blood, saliva, rectal, lung, stool, urine, breath, nasal, ocular sample, or the like. In some embodiments, one or more biological samples are analyzed by the methods described herein. The biological sample(s) contain at least a first unidentified infectious agent and may contain more than one infectious agent. The biological sample(s) are obtained from a biological entity. The biological sample can be obtained by a variety of manners such as by biopsy, swabbing, and the like. The biological samples may be obtained by a physician in a hospital or other health care environment. The physician may then perform the identification analysis or send the biological sample to a laboratory to carry out the analysis.
[0110]Biological entities include, but are not limited to, a mammal, a bird, or a reptile. The biological entity may be a cow, horse, dog, cat, or a primate. The biological entity can also be a human. The biological entity may be living or dead.
[0111]An infectious agent differentiating area is any area or location within a biological entity that can distinguish between a harmful versus normal health condition. An infectious agent differentiating area can be a region or area of the biological entity whereby an infectious agent is more likely to predominate from another region or area of the biological entity. For example, infectious agent differentiating areas may include the blood vessels of the heart (heart disease, coronary artery disease, etc.), particular portions of the digestive system (ulcers, Crohn's disease, etc.), liver (hepatitis infections), and the like. In some embodiments, one or more biological samples from a plurality of infectious agent differentiating areas is analyzed the methods described herein.
[0112]Infectious agents of the invention may potentially cause a health condition in a biological entity. Health conditions include any condition, syndrome, illness, disease, or the like, identified currently or in the future by medical personnel. Infectious agents include, but are not limited to, bacteria, viruses, parasites, fungi, and the like.
[0113]In other embodiments of the invention, the methods disclosed herein can be used to screen blood and other bodily fluids and tissues for pathogenic and non-pathogenic bacteria, viruses, parasites, fungi and the like. Animal samples, including but not limited to, blood and other bodily fluid and tissue samples, can be obtained from living animals, who are either known or not known to or suspected of having a disease, infection, or condition. Alternately, animal samples such as blood and other bodily fluid and tissue samples can be obtained from deceased animals. Blood samples can be further separated into plasma or cellular fractions and further screened as desired. Bodily fluids and tissues can be obtained from any part of the animal or human body. Animal samples can be obtained from, for example, mammals and humans.
[0114]Clinical samples are analyzed for disease causing bioagents and biowarfare pathogens simultaneously with detection of bioagents at levels as low as 100-1000 genomic copies in complex backgrounds with throughput of approximately 100-300 samples with simultaneous detection of bacteria and viruses. Such analyses provide additional value in probing bioagent genomes for unanticipated modifications. These analyses are carried out in reference labs, hospitals and the LRN laboratories of the public health system in a coordinated fashion, with the ability to report the results via a computer network to a common data-monitoring center in real time. Clonal propagation of specific infectious agents, as occurs in the epidemic outbreak of infectious disease, can be tracked with base composition signatures, analogous to the pulse field gel electrophoresis fingerprinting patterns used in tracking the spread of specific food pathogens in the Pulse Net system of the CDC (Swaminathan, B., et al., Emerging Infectious Diseases, 2001, 7, 382-389). The present invention provides a digital barcode in the form of a series of base composition signatures, the combination of which is unique for each known organism. This capability enables real-time infectious disease monitoring across broad geographic locations, which may be essential in a simultaneous outbreak or attack in different cities.
[0115]In other embodiments of the invention, the methods disclosed herein can be used for detecting the presence of pathogenic and non-pathogenic bacteria, viruses, parasites, fungi and the like in organ donors and/or in organs from donors. Such examination can result in the prevention of the transfer of, for example, viruses such as West Nile virus, hepatitis viruses, human immunodeficiency virus, and the like from a donor to a recipient via a transplanted organ. The methods disclosed herein can also be used for detection of host versus graft or graft versus host rejection issues related to organ donors by detecting the presence of particular antigens in either the graft or host known or suspected of causing such rejection. In particular, the bioagents in this regard are the antigens of the major histocompatibility complex, such as the HLA antigens. The present methods can also be used to detect and track emerging infectious diseases, such as West Nile virus infection, HIV-related diseases.
[0116]In other embodiments of the invention, the methods disclosed herein can be used for pharmacogenetic analysis and medical diagnosis including, but not limited to, cancer diagnosis based on mutations and polymorphisms, drug resistance and susceptibility testing, screening for and/or diagnosis of genetic diseases and conditions, and diagnosis of infectious diseases and conditions. In context of the present invention, pharmacogenetics is defined as the study of variability in drug response due to genetic factors. Pharmacogenetic investigations are often based on correlating patient outcome with variations in genes involved in the mode of action of a given drug. For example, receptor genes, or genes involved in metabolic pathways. The methods of the present invention provide a means to analyze the DNA of a patient to provide the basis for pharmacogenetic analysis.
[0117]The present method can also be used to detect single nucleotide polymorphisms (SNPs), or multiple nucleotide polymorphisms, rapidly and accurately. A SNP is defined as a single base pair site in the genome that is different from one individual to another. The difference can be expressed either as a deletion, an insertion or a substitution, and is frequently linked to a disease state. Because they occur every 100-1000 base pairs, SNPs are the most frequently bound type of genetic marker in the human genome.
[0118]For example, sickle cell anemia results from an A-T transition, which encodes a valine rather than a glutamic acid residue. Oligonucleotide primers may be designed such that they bind to sequences that flank a SNP site, followed by nucleotide amplification and mass determination of the amplified product. Because the molecular masses of the resulting product from an individual who does not have sickle cell anemia is different from that of the product from an individual who has the disease, the method can be used to distinguish the two individuals. Thus, the method can be used to detect any known SNP in an individual and thus diagnose or determine increased susceptibility to a disease or condition.
[0119]In one embodiment, blood is drawn from an individual and peripheral blood mononuclear cells (PBMC) are isolated and simultaneously tested, such as in a high-throughput screening method, for one or more SNPs using appropriate primers based on the known sequences which flank the SNP region. The National Center for Biotechnology Information maintains a publicly available database of SNPs on the world wide web of the Internet at, for example, "ncbi.nlm.nih.gov/SNP/."
[0120]The method of the present invention can also be used for blood typing. The gene encoding A, B or O blood type can differ by four single nucleotide polymorphisms. If the gene contains the sequence CGTGGTGACCCTT (SEQ ID NO:5), antigen A results. If the gene contains the sequence CGTCGTCACCGCTA (SEQ ID NO:6) antigen B results. If the gene contains the sequence CGTGGT-ACCCCTT (SEQ ID NO:7), blood group 0 results ("-" indicates a deletion). These sequences can be distinguished by designing a single primer pair which flanks these regions, followed by amplification and mass determination.
[0121]The method of the present invention can also be used for detection and identification of blood-borne pathogens such as Staphylococcus aureus for example.
[0122]The method of the present invention can also be used for strain typing of respiratory pathogens in epidemic surveillance. Group A streptococci (GAS), or Streptococcus pyogenes, is one of the most consequential causes of respiratory infections because of prevalence and ability to cause disease with complications such as acute rheumatic fever and acute glomerulonephritis. GAS also causes infections of the skin (impetigo) and, in rare cases, invasive disease such as necrotizing fasciitis and toxic shock syndrome. Despite many decades of study, the underlying microbial ecology and natural selection that favors enhanced virulence and explosive GAS outbreaks is still poorly understood. The ability to detect GAS and multiple other pathogenic and non-pathogenic bacteria and viruses in patient samples would greatly facilitate our understanding of GAS epidemics. It is also essential to be able to follow the spread of virulent strains of GAS in populations and to distinguish virulent strains from less virulent or avirulent streptococci that colonize the nose and throat of asymptomatic individuals at a frequency ranging from 5-20% of the population (Bisno, A. L. (1995) in Principles and Practice of Infectious Diseases, eds. Mandell, G. L., Bennett, J. E. & Dolin, R. (Churchill Livingston, New York), Vol. 2, pp. 1786-1799). Molecular methods have been developed to type GAS based upon the sequence of the emm gene that encodes the M-protein virulence factor (Beall, B., Facklam, R. & Thompson, T. (1996) J. Clin. Micro. 34, 953-958; Beall, B., et al. (1997) J. Clin. Micro. 35, 1231-1235; Facklam, R., et al. (1999) Emerging Infectious Diseases 5, 247-253). Using this molecular classification, over 150 different emm-types are defined and correlated with phenotypic properties of thousands of GAS isolates (www.cdc.gov/ncidod/biotech/strep/strepindex.html) (Facklam, R., et al. (2002) Clinical Infectious Diseases 34, 28-38). Recently, a strategy known as Multi Locus Sequence Typing (MLST) was developed to follow the molecular Epidemiology of GAS (13). In MLST, internal fragments of seven housekeeping genes are amplified, sequenced, and compared to a database of previously studied isolates (www.test.mlst.net/).
[0123]The present invention enables an emm-typing process to be carried out directly from throat swabs for a large number of samples within 12 hours, allowing strain tracking of an ongoing epidemic, even if geographically dispersed, on a larger scale than ever before achievable.
[0124]In another embodiment, the present invention can be employed in the serotyping of viruses including, but not limited to, adenoviruses. Adenoviruses are DNA viruses that cause over 50% of febrile respiratory illnesses in military recruits. Human adenoviruses are divided into six major serogroups (A through F), each containing multiple strain types. Despite the prevalence of adenoviruses, there are no rapid methods for detecting and serotyping adenoviruses.
[0125]In another embodiment, the present invention can be employed in distinguishing between members of the Orthopoxvirus genus. Smallpox is caused by the Variola virus. Other members of the genus include Vaccinia, Monkeypox, Camelpox, and Cowpox. All are capable of infecting humans, thus, a method capable of identifying and distinguishing among members of the Orthopox genus is a worthwhile objective.
[0126]In another embodiment, the present invention can be employed in distinguishing between viral agents of viral hemorrhagic fevers (VHF). VHF agents include, but are not limited to, Filoviridae (Marburg virus and Ebola virus), Arenaviridae (Lassa, Junin, Machupo, Sabia, and Guanarito viruses), Bunyaviridae (Crimean-Congo hemorrhagic fever virus (CCHFV), Rift Valley fever virus, and Hanta viruses), and Flaviviridae (yellow fever virus and dengue virus). Infections by VHF viruses are associated with a wide spectrum of clinical manifestations such as diarrhea, myalgia, cough, headache, pneumonia, encephalopathy, and hepatitis. Filoviruses, arenaviruses, and CCHFV are of particular relevance because they can be transmitted from human to human, thus causing epidemics with high mortality rates (Khan, A. S., et al., Am. J. Trop. Med. Hyg, 1997, 57, 519-525). In the absence of bleeding or organ manifestation, VHF is clinically difficult to diagnose, and the various etiologic agents can hardly be distinguished by clinical tests. Current approaches to PCR detection of these agents are time-consuming, as they include a separate cDNA synthesis step prior to PCR, agarose gel analysis of PCR products, and in some instances a second round of nested amplification or Southern hybridization. PCRs for different pathogens have to be run assay by assay due to differences in cycling conditions, which complicate broad-range testing in a short period. Moreover, post-PCR processing or nested PCR steps included in currently used assays increase the risk of false positive results due to carryover contamination (Kwok, S, and R. Higuchi, Nature 1989, 339, 237-238).
[0127]In another embodiment, the present invention, can be employed in the diagnosis of a plurality of etiologic agents of a disease. An "etiologic agent" is herein defined as a pathogen acting as the causative agent of a disease. Diseases may be caused by a plurality of etiologic agents. For example, recent studies have implicated both human herpesvirus 6 (HHV-6) and the obligate intracellular bacterium Chlamydia pneumoniae in the etiology of multiple sclerosis (Swanborg, R. H. Microbes and Infection 2002, 4, 1327-1333). The present invention can be applied to the identification of multiple etiologic agents of a disease by, for example, the use of broad range bacterial intelligent primers and division-wide primers (if necessary) for the identification of bacteria such as Chlamydia pneumoniae followed by primers directed to viral housekeeping genes for the identification of viruses such as HHV-6, for example.
[0128]In other embodiments of the invention, the methods disclosed herein can be used for detection and identification of pathogens in livestock. Livestock includes, but is not limited to, cows, pigs, sheep, chickens, turkeys, goats, horses and other farm animals. For example, conditions classified by the California Department of Food and Agriculture as emergency conditions in livestock (www.cdfa.ca.gov/ahfss/ah/pdfs/CA_reportable_disease_list--05292002.- pdf) include, but are not limited to: Anthrax (Bacillus anthracis), Screwworm myiasis (Cochliomyia hominivorax or Chrysomya bezziana), African trypanosomiasis (Tsetse fly diseases), Bovine babesiosis (piroplasmosis), Bovine spongiform encephalopathy (Mad Cow), Contagious bovine pleuropneumonia (Mycoplasma mycoides mycoides small colony), Foot-and-mouth disease (Hoof-and-mouth), Heartwater (Cowdria ruminantium), Hemorrhagic septicemia (Pasteurella multocida serotypes B:2 or E:2), Lumpy skin disease, Malignant catarrhal fever (African type), Rift Valley fever, Rinderpest (Cattle plague), Theileriosis (Corridor disease, East Coast fever), Vesicular stomatitis, Contagious agalactia (Mycoplasma species), Contagious caprine pleuropneumonia (Mycoplasma capricolumn capripneumoniae), Nairobi sheep disease, Peste des petits ruminants (Goat plague), Pulmonary adenomatosis (Viral neoplastic pneumonia), Salmonella abortus ovis, Sheep and goat pox, African swine fever, Classical swine fever (Hog cholera), Japanese encephalitis, Nipah virus, Swine vesicular disease, Teschen disease (Enterovirus encephalomyelitis), Vesicular exanthema, Exotic Newcastle disease (Viscerotropic velogenic Newcastle disease), Highly pathogenic avian influenza (Fowl plague), African horse sickness, Dourine (Trypanosoma equiperdum), Epizootic lymphangitis (equine blastomycosis, equine histoplasmosis), Equine piroplasmosis (Babesia equi, B. caballi), Glanders (Farcy) (Pseudomonas mallei), Hendra virus (Equine morbillivirus), Horse pox, Surra (Trypanosoma evansi), Venezuelan equine encephalomyelitis, West Nile Virus, Chronic wasting disease in cervids, and Viral hemorrhagic disease of rabbits (calicivirus)
[0129]Conditions classified by the California Department of Food and Agriculture as regulated conditions in livestock include, but are not limited to: rabies, Bovine brucellosis (Brucella abortus), Bovine tuberculosis (Mycobacterium bovis), Cattle scabies (multiple types), Trichomonosis (Tritrichomonas fetus), Caprine and ovine brucellosis (excluding Brucella ovis), Scrapie, Sheep scabies (Body mange) (Psoroptes ovis), Porcine brucellosis (Brucella suis), Pseudorabies (Aujeszky's disease), Ornithosis (Psittacosis or avian chlamydiosis) (Chlamydia psittaci), Pullorum disease (Fowl typhoid) (Salmonella gallinarum and pullorum), Contagious equine metritis (Taylorella equigenitalis), Equine encephalomyelitis (Eastern and Western equine encephalitis), Equine infectious anemia (Swamp fever), Duck viral enteritis (Duck plague), and Tuberculosis in cervids.
[0130]Additional conditions monitored by the California Department of Food and Agriculture include, but are not limited to: Avian tuberculosis (Mycobacterium avium), Echinococcosis/Hydatidosis (Echinococcus species), Leptospirosis, Anaplasmosis (Anaplasma marginale or A. centrale), Bluetongue, Bovine cysticercosis (Taenia saginata in humans), Bovine genital campylobacteriosis (Campylobacter fetus venerealis), Dermatophilosis (Streptothricosis, mycotic dermatitis) (Dermatophilus congolensis), Enzootic bovine leukosis (Bovine leukemia virus), Infectious bovine rhinotracheitis (Bovine herpesvirus-1), Johne's disease (Paratuberculosis) (Mycobacterium avium paratuberculosis), Malignant catarrhal fever (North American), Q Fever (Coxiella burnetii), Caprine (contagious) arthritis/encephalitis, Enzootic abortion of ewes (Ovine chlamydiosis) (Chlamydia psittaci), Maedi-Visna (Ovine progressive pneumonia), Atrophic rhinitis (Bordetella bronchiseptica, Pasteurella multocida), Porcine cysticercosis (Taenia solium in humans), Porcine reproductive and respiratory syndrome, Transmissible gastroenteritis (coronavirus), Trichinellosis (Trichinella spiralis), Avian infectious bronchitis, Avian infectious laryngotracheitis, Duck viral hepatitis, Fowl cholera (Pasteurella multocida), Fowl pox, Infectious bursal disease (Gumboro disease), Low pathogenic avian influenza, Marek's disease, Mycoplasmosis (Mycoplasma gallisepticum), Equine influenza Equine rhinopneumonitis (Equine herpesvirus-1), Equine viral arteritis, and Horse mange (multiple types).
[0131]2. Identification of Bioagents of Biological Warfare
[0132]A key problem in determining that an infectious outbreak is the result of a bioterrorist attack is the sheer variety of organisms that might be used by terrorists. According to a recent review (Taylor, L. H. et al. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2001, 356, 983-989), there are over 1400 organisms infectious to humans; most of these have the potential to be used in a deliberate, malicious attack. These numbers do not include numerous strain variants of each organism, bioengineered versions, or pathogens that infect plants or animals. Paradoxically, most of the new technology being developed for detection of biological weapons incorporates a version of quantitative PCR, which is based upon the use of highly specific primers and probes designed to selectively identify specific pathogenic organisms. This approach requires assumptions about the type and strain of bacteria or virus which is expected to be detected. Although this approach will work for the most obvious organisms, like smallpox and anthrax, experience has shown that it is very difficult to anticipate what a terrorist will do.
[0133]The present invention can be used to detect and identify any biological agent, including bacteria, viruses, fungi and toxins without prior knowledge of the organism being detected and identified. As one example, where the agent is a biological threat, the information obtained such as the presence of toxin genes, pathogenicity islands and antibiotic resistance genes for example, is used to determine practical information needed for countermeasures. In addition, the methods can be used to identify natural or deliberate engineering events including chromosome fragment swapping, molecular breeding (gene shuffling) and emerging infectious diseases. The present invention provides broad-function technology that may be the only practical means for rapid diagnosis of disease caused by a biowarfare or bioterrorist attack, especially an attack that might otherwise be missed or mistaken for a more common infection.
[0134]Bacterial biological warfare agents capable of being detected by the present methods include, but are not limited to, Bacillus anthracis (anthrax), Yersinia pestis (pneumonic plague), Franciscella tularensis (tularemia), Brucella suis, Brucella abortus, Brucella melitensis (undulant fever), Burkholderia mallei (glanders), Burkholderia pseudomalleii (melioidosis), Salmonella typhi (typhoid fever), Rickettsia typhii (epidemic typhus), Rickettsia prowasekii (endemic typhus) and Coxiella burnetii (Q fever), Rhodobacter capsulatus, Chlamydia pneumoniae, Escherichia coli, Shigella dysenteriae, Shigella flexneri, Bacillus cereus, Clostridium botulinum, Coxiella burnetti, Pseudomonas aeruginosa, Legionella pneumophila, and Vibrio cholerae.
[0135]Besides 16S and 23S rRNA, other target regions suitable for use in the present invention for detection of bacteria include, but are not limited to, 5S rRNA and RNase P (FIG. 3).
[0136]Fungal biowarfare agents include, but are not limited to, Coccidioides immitis (Coccidioidomycosis), and Magnaporthe grisea.
[0137]Biological warfare toxin genes capable of being detected by the methods of the present invention include, but are not limited to, botulinum toxin, T-2 mycotoxins, ricin, staph enterotoxin B, shigatoxin, abrin, aflatoxin, Clostridium perfringens epsilon toxin, conotoxins, diacetoxyscirpenol, tetrodotoxin and saxitoxin.
[0138]Parasites that could be used in biological warfare include, but are not limited to: Ascaris suum, Giardia lamblia, Cryptosporidium, and Schistosoma.
[0139]Biological warfare viral threat agents are mostly RNA viruses (positive-strand and negative-strand), with the exception of smallpox. Every RNA virus is a family of related viruses (quasispecies). These viruses mutate rapidly and the potential for engineered strains (natural or deliberate) is very high. RNA viruses cluster into families that have conserved RNA structural domains on the viral genome (e.g., virion components, accessory proteins) and conserved housekeeping genes that encode core viral proteins including, for single strand positive strand RNA viruses, RNA-dependent RNA polymerase, double stranded RNA helicase, chymotrypsin-like and papain-like proteases and methyltransferases. "Housekeeping genes" refers to genes that are generally always expressed and thought to be involved in routine cellular metabolism.
[0140]Examples of (-)-strand RNA viruses include, but are not limited to, arenaviruses (e.g., sabia virus, lassa fever, Machupo, Argentine hemorrhagic fever, flexal virus), bunyaviruses (e.g., hantavirus, nairovirus, phlebovirus, hantaan virus, Congo-crimean hemorrhagic fever, rift valley fever), and mononegavirales (e.g., filovirus, paramyxovirus, ebola virus, Marburg, equine morbillivirus).
[0141]Examples of (+)-strand RNA viruses include, but are not limited to, picornaviruses (e.g., coxsackievirus, echovirus, human coxsackievirus A, human echovirus, human enterovirus, human poliovirus, hepatitis A virus, human parechovirus, human rhinovirus), astroviruses (e.g., human astrovirus), calciviruses (e.g., chiba virus, chitta virus, human calcivirus, norwalk virus), nidovirales (e.g., human coronavirus, human torovirus), flaviviruses (e.g., dengue virus 1-4, Japanese encephalitis virus, Kyanasur forest disease virus, Murray Valley encephalitis virus, Rocio virus, St. Louis encephalitis virus, West Nile virus, yellow fever virus, hepatitis c virus) and togaviruses (e.g., Chikugunya virus, Eastern equine encephalitis virus, Mayaro virus, O'nyong-nyong virus, Ross River virus, Venezuelan equine encephalitis virus, Rubella virus, hepatitis E virus). The hepatitis C virus has a 5'-untranslated region of 340 nucleotides, an open reading frame encoding 9 proteins having 3010 amino acids and a 3'-untranslated region of 240 nucleotides. The 5'-UTR and 3'-UTR are 99% conserved in hepatitis C viruses.
[0142]In one embodiment, the target gene is an RNA-dependent RNA polymerase or a helicase encoded by (+)-strand RNA viruses, or RNA polymerase from a (-)-strand RNA virus. (+)-strand RNA viruses are double stranded RNA and replicate by RNA-directed RNA synthesis using RNA-dependent RNA polymerase and the positive strand as a template. Helicase unwinds the RNA duplex to allow replication of the single stranded RNA. These viruses include viruses from the family picornaviridae (e.g., poliovirus, coxsackievirus, echovirus), togaviridae (e.g., alphavirus, flavivirus, rubivirus), arenaviridae (e.g., lymphocytic choriomeningitis virus, lassa fever virus), cononaviridae (e.g., human respiratory virus) and Hepatitis A virus. The genes encoding these proteins comprise variable and highly conserved regions which flank the variable regions.
[0143]In one embodiment, the method can be used to detect the presence of antibiotic resistance and/or toxin genes in a bacterial species. For example, Bacillus anthracis comprising a tetracycline resistance plasmid and plasmids encoding one or both anthracis toxins (px01 and/or px02) can be detected by using antibiotic resistance primer sets and toxin gene primer sets. If the B. anthracis is positive for tetracycline resistance, then a different antibiotic, for example quinalone, is used.
[0144]While the present invention has been described with specificity in accordance with certain of its embodiments, the following examples serve only to illustrate the invention and are not intended to limit the same.
EXAMPLES
Example 1
Nucleic Acid Isolation and PCR
[0145]In one embodiment, nucleic acid is isolated from the organisms and amplified by PCR using standard methods prior to BCS determination by mass spectrometry. Nucleic acid is isolated, for example, by detergent lysis of bacterial cells, centrifugation and ethanol precipitation. Nucleic acid isolation methods are described in, for example, Current Protocols in Molecular Biology (Ausubel et al.) and Molecular Cloning; A Laboratory Manual (Sambrook et al.). The nucleic acid is then amplified using standard methodology, such as PCR, with primers which bind to conserved regions of the nucleic acid which contain an intervening variable sequence as described below.
[0146]General Genomic DNA Sample Prep Protocol: Raw samples are filtered using Supor-200 0.2 μm membrane syringe filters (VWR International). Samples are transferred to 1.5 ml eppendorf tubes pre-filled with 0.45 g of 0.7 mm Zirconia beads followed by the addition of 350 μl of ATL buffer (Qiagen, Valencia, Calif.). The samples are subjected to bead beating for 10 minutes at a frequency of 19 l/s in a Retsch Vibration Mill (Retsch). After centrifugation, samples are transferred to an S-block plate (Qiagen) and DNA isolation is completed with a BioRobot 8000 nucleic acid isolation robot (Qiagen).
[0147]Swab Sample Protocol: Allegiance S/P brand culture swabs and collection/transport system are used to collect samples. After drying, swabs are placed in 17×100 mm culture tubes (VWR
[0148]International) and the genomic nucleic acid isolation is carried out automatically with a Qiagen Mdx robot and the Qiagen QIAamp DNA Blood BioRobot Mdx genomic preparation kit (Qiagen, Valencia, Calif.).
Example 2
Mass Spectrometry
[0149]FTICR Instrumentation: The FTICR instrument is based on a 7 tesla actively shielded superconducting magnet and modified Bruker Daltonics Apex II 70e ion optics and vacuum chamber. The spectrometer is interfaced to a LEAP PAL autosampler and a custom fluidics control system for high throughput screening applications. Samples are analyzed directly from 96-well or 384-well microtiter plates at a rate of about 1 sample/minute. The Bruker data-acquisition platform is supplemented with a lab-built ancillary NT datastation which controls the autosampler and contains an arbitrary waveform generator capable of generating complex rf-excite waveforms (frequency sweeps, filtered noise, stored waveform inverse Fourier transform (SWIFT), etc.) for sophisticated tandem MS experiments. For oligonucleotides in the 20-30-mer regime typical performance characteristics include mass resolving power in excess of 100,000 (FWHM), low ppm mass measurement errors, and an operable m/z range between 50 and 5000 m/z.
[0150]Modified ESI Source: In sample-limited analyses, analyte solutions are delivered at 150 mL/minute to a 30 mm i.d. fused-silica ESI emitter mounted on a 3-D micromanipulator. The ESI ion optics consists of a heated metal capillary, an rf-only hexapole, a skimmer cone, and an auxiliary gate electrode. The 6.2 cm rf-only hexapole is comprised of 1 mm diameter rods and is operated at a voltage of 380 Vpp at a frequency of 5 MHz. A lab-built electro-mechanical shutter can be employed to prevent the electrospray plume from entering the inlet capillary unless triggered to the "open" position via a TTL pulse from the data station. When in the "closed" position, a stable electrospray plume is maintained between the ESI emitter and the face of the shutter. The back face of the shutter arm contains an elastomeric seal that can be positioned to form a vacuum seal with the inlet capillary. When the seal is removed, a 1 mm gap between the shutter blade and the capillary inlet allows constant pressure in the external ion reservoir regardless of whether the shutter is in the open or closed position. When the shutter is triggered, a "time slice" of ions is allowed to enter the inlet capillary and is subsequently accumulated in the external ion reservoir. The rapid response time of the ion shutter (<25 ms) provides reproducible, user defined intervals during which ions can be injected into and accumulated in the external ion reservoir.
[0151]Apparatus for Infrared Multiphoton Dissociation: A 25 watt CW CO2 laser operating at 10.6 μm has been interfaced to the spectrometer to enable infrared multiphoton dissociation (IRMPD) for oligonucleotide sequencing and other tandem MS applications. An aluminum optical bench is positioned approximately 1.5 m from the actively shielded superconducting magnet such that the laser beam is aligned with the central axis of the magnet. Using standard IR-compatible mirrors and kinematic mirror mounts, the unfocused 3 mm laser beam is aligned to traverse directly through the 3.5 mm holes in the trapping electrodes of the FTICR trapped ion cell and longitudinally traverse the hexapole region of the external ion guide finally impinging on the skimmer cone. This scheme allows IRMPD to be conducted in an m/z selective manner in the trapped ion cell (e.g. following a SWIFT isolation of the species of interest), or in a broadband mode in the high pressure region of the external ion reservoir where collisions with neutral molecules stabilize IRMPD-generated metastable fragment ions resulting in increased fragment ion yield and sequence coverage.
Example 3
Identification of Bioagents
[0152]Table 2 shows a small cross section of a database of calculated molecular masses for over 9 primer sets and approximately 30 organisms. The primer sets were derived from rRNA alignment. Examples of regions from rRNA consensus alignments are shown in FIGS. 1A-1C. Lines with arrows are examples of regions to which intelligent primer pairs for PCR are designed. The primer pairs are >95% conserved in the bacterial sequence database (currently over 10,000 organisms). The intervening regions are variable in length and/or composition, thus providing the base composition "signature" (BCS) for each organism. Primer pairs were chosen so the total length of the amplified region is less than about 80-90 nucleotides. The label for each primer pair represents the starting and ending base number of the amplified region on the consensus diagram.
[0153]Included in the short bacterial database cross-section in Table 2 are many well known pathogens/biowarfare agents (shown in bold/red typeface) such as Bacillus anthracis or Yersinia pestis as well as some of the bacterial organisms found commonly in the natural environment such as Streptomyces. Even closely related organisms can be distinguished from each other by the appropriate choice of primers. For instance, two low G+C organisms, Bacillus anthracis and Staph aureus, can be distinguished from each other by using the primer pair defined by 16S--1337 or 23S--855 (ΔM of 4 Da).
TABLE-US-00002 TABLE 2 Cross Section Of A Database Of Calculated Molecular Masses1 Primer Regions Bug Name 16S_971 16S_1100 16S_1337 16S_1294 16S_1228 23S_1021 23S_855 23S_193 23S_115 Acinetobacter calcoaceticus 55619.1 55004 28446.7 35854.9 51295.4 30299 42654 39557.5 54999 55005 54388 28448 35238 51296 30295 42651 39560 56850 Bacillus cereus 55622.1 54387.9 28447.6 35854.9 51296.4 30295 42651 39560.5 56850.3 Bordetella bronchiseptica 56857.3 51300.4 28446.7 35857.9 51307.4 30299 42653 39559.5 51920.5 Borrelia burgdorferi 56231.2 55621.1 28440.7 35852.9 51295.4 30297 42029.9 38941.4 52524.6 58098 55011 28448 35854 50683 Campylobacter jejuni 58088.5 54386.9 29061.8 35856.9 50674.3 30294 42032.9 39558.5 45732.5 55000 55007 29063 35855 50676 30295 42036 38941 56230 55006 53767 28445 35855 51291 30300 42656 39562 54999 Clostridium difficile 56855.3 54386.9 28444.7 35853.9 51296.4 30294 41417.8 39556.5 55612.2 Enterococcus faecalis 55620.1 54387.9 28447.6 35858.9 51296.4 30297 42652 39559.5 56849.3 55622 55009 28445 35857 51301 30301 42656 39562 54999 53769 54385 28445 35856 51298 Haemophilus influenzae 55620.1 55006 28444.7 35855.9 51298.4 30298 42656 39560.5 55613.1 Klebsiella pneumoniae 55622.1 55008 28442.7 35856.9 51297.4 30300 42655 39562.5 55000 55618 55626 28446 35857 51303 Mycobacterium avium 54390.9 55631.1 29064.8 35858.9 51915.5 30298 42656 38942.4 56241.2 Mycobacterium leprae 54389.9 55629.1 29064.8 35860.9 51917.5 30298 42656 39559.5 56240.2 Mycobacterium tuberculosis 54390.9 55629.1 29064.8 35860.9 51301.4 30299 42656 39560.5 56243.2 Mycoplasma genitalium 53143.7 45115.4 29061.8 35854.9 50671.3 30294 43264.1 39558.5 56842.4 Mycoplasma pneumoniae 53143.7 45118.4 29061.8 35854.9 50673.3 30294 43264.1 39559.5 56843.4 Neisseria gonorrhoeae 55627.1 54389.9 28445.7 35855.9 51302.4 30300 42649 39561.5 55000 55623 55010 28443 35858 51301 30298 43272 39558 55619 58093 55621 28448 35853 50677 30293 42650 39559 53139 58094 55623 28448 35853 50679 30293 42648 39559 53755 55622 55005 28445 35857 51301 30301 42658 55623 55009 28444 35857 51301 Staphylococcus aureus 56854.3 54386.9 28443.7 35852.9 51294.4 30298 42655 39559.5 57466.4 Streptomyces 54389.9 59341.6 29063.8 35858.9 51300.4 39563.5 56864.3 Treponema pallidum 56245.2 55631.1 28445.7 35851.9 51297.4 30299 42034.9 38939.4 57473.4 55625 55626 28443 35857 52536 29063 30303 35241 50675 Vibrio parahaemolyticus 54384.9 55626.1 28444.7 34620.7 50064.2 55620 55626 28443 35857 51299 1Molecular mass distribution of PCR amplified regions for a selection of organisms (rows) across various primer pairs (columns). Pathogens are shown in bold. Empty cells indicate presently incomplete or missing data.
[0154]FIG. 6 shows the use of ESI-FT-ICR MS for measurement of exact mass. The spectra from 46 mer PCR products originating at position 1337 of the 16S rRNA from S. aureus (upper) and B. anthracis (lower) are shown. These data are from the region of the spectrum containing signals from the [M-8H+]8- charge states of the respective 5'-3' strands. The two strands differ by two (AT→CG) substitutions, and have measured masses of 14206.396 and 14208.373+0.010 Da, respectively. The possible base compositions derived from the masses of the forward and reverse strands for the B. anthracis products are listed in Table 3.
TABLE-US-00003 TABLE 3 Possible base composition for B. anthracis products Calc. Mass Error Base Comp. 14208.2935 0.079520 A1 G17 C10 T18 14208.3160 0.056980 A1 G20 C15 T10 14208.3386 0.034440 A1 G23 C20 T2 14208.3074 0.065560 A6 G11 C3 T26 14208.3300 0.043020 A6 G14 C8 T18 14208.3525 0.020480 A6 G17 C13 T10 14208.3751 0.002060 A6 G20 C18 T2 14208.3439 0.029060 A11 G8 C1 T26 14208.3665 0.006520 A11 G11 C6 T18 14208.3890 0.016020 A11 G14 C11 T10 14208.4116 0.038560 A11 G17 C16 T2 14208.4030 0.029980 A16 G8 C4 T18 14208.4255 0.052520 A16 G11 C9 T10 14208.4481 0.075060 A16 G14 C14 T2 14208.4395 0.066480 A21 G5 C2 T18 14208.4620 0.089020 A21 G8 C7 T10 14079.2624 0.080600 A0 G14 C13 T19 14079.2849 0.058060 A0 G17 C18 T11 14079.3075 0.035520 A0 G20 C23 T3 14079.2538 0.089180 A5 G5 C1 T35 14079.2764 0.066640 A5 G8 C6 T27 14079.2989 0.044100 A5 G11 C11 T19 14079.3214 0.021560 A5 G14 C16 T11 14079.3440 0.000980 A5 G17 C21 T3 14079.3129 0.030140 A10 G5 C4 T27 14079.3354 0.007600 A10 G8 C9 T19 14079.3579 0.014940 A10 G11 C14 T11 14079.3805 0.037480 A10 G14 C19 T3 14079.3494 0.006360 A15 G2 C2 T27 14079.3719 0.028900 A15 G5 C7 T19 14079.3944 0.051440 A15 G8 C12 T11 14079.4170 0.073980 A15 G11 C17 T3 14079.4084 0.065400 A20 G2 C5 T19 14079.4309 0.087940 A20 G5 C10 T13
[0155]Among the 16 compositions for the forward strand and the 18 compositions for the reverse strand that were calculated, only one pair (shown in bold) are complementary, corresponding to the actual base compositions of the B. anthracis PCR products.
Example 4
BCS of Region from Bacillus anthracis and Bacillus cereus
[0156]A conserved Bacillus region from B. anthracis (A14G9C14T9) and B. cereus (A15G9C13T9) having a C to A base change was synthesized and subjected to ESI-TOF MS. The results are shown in FIG. 7 in which the two regions are clearly distinguished using the method of the present invention (MW=14072.26 vs. 14096.29).
Example 5
Identification of Additional Bioagents
[0157]In other examples of the present invention, the pathogen Vibrio cholera can be distinguished from Vibrio parahemolyticus with ΔM>600 Da using one of three 16S primer sets shown in Table 2 (16S--971, 16S--1228 or 16S--1294) as shown in Table 4. The two mycoplasma species in the list (M. genitalium and M. pneumoniae) can also be distinguished from each other, as can the three mycobacteriae. While the direct mass measurements of amplified products can identify and distinguish a large number of organisms, measurement of the base composition signature provides dramatically enhanced resolving power for closely related organisms. In cases such as Bacillus anthracis and Bacillus cereus that are virtually indistinguishable from each other based solely on mass differences, compositional analysis or fragmentation patterns are used to resolve the differences. The single base difference between the two organisms yields different fragmentation patterns, and despite the presence of the ambiguous/unidentified base N at position 20 in B. anthracis, the two organisms can be identified.
[0158]Tables 4a-b show examples of primer pairs from Table 1 which distinguish pathogens from background.
TABLE-US-00004 TABLE 4a Organism name 23S_855 16S_1337 23S_1021 Bacillus anthracis 42650.98 28447.65 30294.98 Staphylococcus aureus 42654.97 28443.67 30297.96
TABLE-US-00005 TABLE 4b Organism name 16S_971 16S_1294 16S_1228 Vibrio cholerae 55625.09 35856.87 52535.59 Vibrio parahaemolyticus 54384.91 34620.67 50064.19
[0159]Table 5 shows the expected molecular weight and base composition of region 16S--1100-1188 in Mycobacterium avium and Streptomyces sp.
TABLE-US-00006 TABLE 5 Organism Molecular Region name Length weight Base comp. 16S_1100-1188 Mycobac- 82 25624.1728 A16G32C18T16 terium avium 16S_1100-1188 Strepto- 96 29904.871 A17G38C27T14 myces sp.
[0160]Table 6 shows base composition (single strand) results for 16S--1100-1188 primer amplification reactions different species of bacteria. Species which are repeated in the table (e.g., Clostridium botulinum) are different strains which have different base compositions in the 16S--1100-1188 region.
TABLE-US-00007 TABLE 6 Organism name Base comp. Mycobacterium avium A16G32C18T16 Streptomyces sp. A17G38C27T14 Ureaplasma urealyticum A18G30C17T17 Streptomyces sp. A19G36C24T18 Mycobacterium leprae A20G32C22T16 A20G33C21T16 A20G33C21T16 Fusobacterium necroforum A21G26C22T18 Listeria monocytogenes A21G27C19T19 Clostridium botulinum A21G27C19T21 Neisseria gonorrhoeae A21G28C21T18 Bartonella quintana A21G30C22T16 Enterococcus faecalis A22G27C20T19 Bacillus megaterium A22G28C20T18 Bacillus subtilis A22G28C21T17 Pseudomonas aeruginosa A22G29C23T15 Legionella pneumophila A22G32C20T16 Mycoplasma pneumoniae A23G20C14T16 Clostridium botulinum A23G26C20T19 Enterococcus faecium A23G26C21T18 Acinetobacter calcoaceti A23G26C21T19 A23G26C24T15 A23G26C24T15 Clostridium perfringens A23G27C19T19 A23G27C20T18 A23G27C20T18 A23G27C20T18 Aeromonas hydrophila A23G29C21T16 Escherichia coli A23G29C21T16 Pseudomonas putida A23G29C21T17 A23G29C22T15 A23G29C22T15 Vibrio cholerae A23G30C21T16 A23G31C21T15 A23G31C21T15 Mycoplasma genitalium A24G19C12T18 Clostridium botulinum A24G25C18T20 Bordetella bronchiseptica A24G26C19T14 Francisella tularensis A24G26C19T19 A24G26C20T18 A24G26C20T18 A24G26C20T18 Helicobacter pylori A24G26C20T19 Helicobacter pylori A24G26C21T18 Moraxella catarrhalis A24G26C23T16 Haemophilus influenzae Rd A24G28C20T17 A24G28C21T16 A24G28C21T16 A24G28C21T16 Pseudomonas putida A24G29C21T16 A24G30C21T15 A24G30C21T15 A24G30C21T15 Clostridium botulinum A25G24C18T21 Clostridium tetani A25G25C18T20 Francisella tularensis A25G25C19T19 Acinetobacter calcoacetic A25G26C20T19 Bacteriodes fragilis A25G27C16T22 Chlamydophila psittaci A25G27C21T16 Borrelia burgdorferi A25G29C17T19 Streptobacillus monilifor A26G26C20T16 Rickettsia prowazekii A26G28C18T18 Rickettsia rickettsii A26G28C20T16 Mycoplasma mycoides A28G23C18T20
[0161]The same organism having different base compositions are different strains. Groups of organisms which are highlighted or in italics have the same base compositions in the amplified region. Some of these organisms can be distinguished using multiple primers. For example, Bacillus anthracis can be distinguished from Bacillus cereus and Bacillus thuringiensis using the primer 16S--971-1062 (Table 7). Other primer pairs which produce unique base composition signatures are shown in Table 6 (bold). Clusters containing very similar threat and ubiquitous non-threat organisms (e.g. anthracis cluster) are distinguished at high resolution with focused sets of primer pairs. The known biowarfare agents in Table 6 are Bacillus anthracis, Yersinia pestis, Francisella tularensis and Rickettsia prowazekii.
TABLE-US-00008 TABLE 7 Organism 16S_971-1062 16S_1228-1310 16S_1100-1188 Aeromonas hydrophila A21G29C22T20 A22G27C21T13 A23G31C21T15 Aeromonas salmonicida A21G29C22T20 A22G27C21T13 A23G31C21T15 Bacillus anthracis A21G27C22T22 A24G22C19T18 A23G27C20T18 Bacillus cereus A22G27C22T22 A24G22C19T18 A23G27C20T18 Bacillus thuringiensis A22G27C21T22 A24G22C19T18 A23G27C20T18 Chlamydia trachomatis A22G26C20T23 A24G23C19T16 A24G28C21T16 Chlamydia pneumoniae AR39 A26G23C20T22 A26G22C16T18 A24G28C21T16 Leptospira borgpetersenii A22G26C20T21 A22G25C21T15 A23G26C24T15 Leptospira interrogans A22G26C20T21 A22G25C21T15 A23G26C24T15 Mycoplasma genitalium A28G23C15T22 A30G18C15T19 A24G19C12T18 Mycoplasma pneumoniae A28G23C15T22 A27G19C16T20 A23G20C14T16 Escherichia coli A22G28C20T22 A24G25C21T13 A23G29C22T15 Shigella dysenteriae A22G28C21T21 A24G25C21T13 A23G29C22T15 Proteus vulgaris A23G26C22T21 A26G24C19T14 A24G30C21T15 Yersinia pestis A24G25C21T22 A25G24C20T14 A24G30C21T15 Yersinia pseudotuberculosis A24G25C21T22 A25G24C20T14 A24G30C21T15 Francisella tularensis A20G25C21T23 A23G26C17T17 A24G26C19T19 Rickettsia prowazekii A21G26C24T25 A24G23C16T19 A26G28C18T18 Rickettsia rickettsii A21G26C25T24 A24G24C17T17 A26G28C20T16
[0162]The sequence of B. anthracis and B. cereus in region 16S--971 is shown below. Shown in bold is the single base difference between the two species which can be detected using the methods of the present invention. B. anthracis has an ambiguous base at position 20.
TABLE-US-00009 B.anthracis_16S_971 (SEQ ID NO: 1) GCGAAGAACCUUACCAGGUNUUGACAUCCUCUGACAACCCUAGAGAUAGG GCUUCUCCUUCGGGAGCAGAGUGACAGGUGGUGCAUGGUU B.cereus_16S_971 (SEQ ID NO: 2) GCGAAGAACCUUACCAGGUCUUGACAUCCUCUGAAAACCCUAGAGAUAGG GCUUCUCCUUCGGGAGCAGAGUGACAGGUGGUGCAUGGUU
Example 6
ESI-TOF MS of sspE 56-mer Plus Calibrant
[0163]The mass measurement accuracy that can be obtained using an internal mass standard in the ESI-MS study of PCR products is shown in FIG. 8. The mass standard was a 20-mer phosphorothioate oligonucleotide added to a solution containing a 56-mer PCR product from the B. anthracis spore coat protein sspE. The mass of the expected PCR product distinguishes B. anthracis from other species of Bacillus such as B. thuringiensis and B. cereus.
Example 7
B. anthracis ESI-TOF Synthetic 16S--1228 Duplex
[0164]An ESI-TOF MS spectrum was obtained from an aqueous solution containing 5 μM each of synthetic analogs of the expected forward and reverse PCR products from the nucleotide 1228 region of the B. anthracis 16S rRNA gene. The results (FIG. 9) show that the molecular weights of the forward and reverse strands can be accurately determined and easily distinguish the two strands. The [M-21H.sup.+].sup.21- and [M-20H.sup.+].sup.20+ charge states are shown.
Example 8
ESI-FTICR-MS of Synthetic B. anthracis 16S--1337 46 Base Pair Duplex
[0165]An ESI-FTICR-MS spectrum was obtained from an aqueous solution containing 5 μM each of synthetic analogs of the expected forward and reverse PCR products from the nucleotide 1337 region of the B. anthracis 16S rRNA gene. The results (FIG. 10) show that the molecular weights of the strands can be distinguished by this method. The [M-16H.sup.+]16- through [M-10H.sup.+]10- charge states are shown. The insert highlights the resolution that can be realized on the FTICR-MS instrument, which allows the charge state of the ion to be determined from the mass difference between peaks differing by a single 13C substitution.
Example 9
ESI-TOF MS of 56-mer Oligonucleotide from saspB Gene of B. anthracis with Internal Mass Standard
[0166]ESI-TOF MS spectra were obtained on a synthetic 56-mer oligonucleotide (5 μM) from the saspB gene of B. anthracis containing an internal mass standard at an ESI of 1.7 μL/min as a function of sample consumption. The results (FIG. 11) show that the signal to noise is improved as more scans are summed, and that the standard and the product are visible after only 100 scans.
Example 10
ESI-TOF MS of an Internal Standard with Tributylammonium (TBA)-trifluoroacetate (TFA) Buffer
[0167]An ESI-TOF-MS spectrum of a 20-mer phosphorothioate mass standard was obtained following addition of 5 mM TBA-TFA buffer to the solution. This buffer strips charge from the oligonucleotide and shifts the most abundant charge state from [M-8H.sup.+]8- to [M-3H.sup.+]3- (FIG. 12).
Example 11
Master Database Comparison
[0168]The molecular masses obtained through Examples 1-10 are compared to molecular masses of known bioagents stored in a master database to obtain a high probability matching molecular mass.
Example 12
Master Data Base Interrogation over the Internet
[0169]The same procedure as in Example 11 is followed except that the local computer did not store the Master database. The Master database is interrogated over an internet connection, searching for a molecular mass match.
Example 13
Master Database Updating
[0170]The same procedure as in example 11 is followed except the local computer is connected to the internet and has the ability to store a master database locally. The local computer system periodically, or at the user's discretion, interrogates the Master database, synchronizing the local master database with the global Master database. This provides the current molecular mass information to both the local database as well as to the global Master database. This further provides more of a globalized knowledge base.
Example 14
Global Database Updating
[0171]The same procedure as in example 13 is followed except there are numerous such local stations throughout the world. The synchronization of each database adds to the diversity of information and diversity of the molecular masses of known bioagents.
[0172]Various modifications of the invention, in addition to those described herein, will be apparent to those skilled in the art from the foregoing description. Such modifications are also intended to fall within the scope of the appended claims. Each reference cited in the present application is incorporated herein by reference in its entirety.
Example 15
Demonstration of Detection and Identification of Five Species of Bacteria in a Mixture
[0173]Broad range intelligent primers were chosen following analysis of a large collection of curated bacterial 16S rRNA sequences representing greater than 4000 species of bacteria. Examples of primers capable of priming from greater than 90% of the organisms in the collection include, but are not limited to, those exhibited in Table 8 wherein Tp=5'propynylated uridine and Cp=5'propynylated cytidine.
TABLE-US-00010 TABLE 8 Intelligent Primer Pairs for Identification of Bacteria Forward Reverse Primer Forward Primer SEQ ID Reverse Primer SEQ ID Pair Name Sequence NO: Sequence NO: 16S_EC_107 GTGAGATGTTGGGTTAAGTCCC 8 GACGTCATCCCCACCTTCCTC 9 7_1195 GTAACGAG 16S_EC_108 ATGTTGGGTTAAGTCCCGCAAC 10 TTGACGTCATCCCCACCTTCCT 11 2_1197 GAG C 16S_EC_109 TTAAGTCCCGCAACGATCGCAA 12 TGACGTCATCCCCACCTTCCTC 13 0_1196 16S_EC_122 GCTACACACGTGCTACAATG 14 CGAGTTGCAGACTGCGATCCG 15 2_1323 16S_EC_133 AAGTCGGAATCGCTAGTAATCG 16 GACGGGCGGTGTGTACAAG 17 2_1407 16S_EC_30-- TGAACGCTGGTGGCATGCTTAA 18 TACGCATTACTCACCCGTCCGC 19 126 CAC 16S_EC_38-- GTGGCATGCCTAATACATGCAA 20 TTACTCACCCGTCCGCCGCT 21 120 GTCG 16S_EC_49-- TAACACATGCAAGTCGAACG 22 TTACTCACCCGTCCGCC 23 120 16S_EC_683 GTGTAGCGGTGAAATGCG 24 GTATCTAATCCTGTTTGCTCCC 25 795 16S_EC_713 AGAACACCGATGGCGAAGGC 26 CGTGGACTACCAGGGTATCTA 27 809 16S_EC_785 GGATTAGAGACCCTGGTAGTCC 28 GGCCGTACTCCCCAGGCG 29 897 16S_EC_785 GGATTAGATACCCTGGTAGTCC 30 GGCCGTACTCCCCAGGCG 31 _897_2 ACGC 16S_EC_789 TAGATACCCTGGTAGTCCACGC 32 CGTACTCCCCAGGCG 33 894 16S_EC_960 TTCGATGCAACGCGAAGAACCT 34 ACGAGCTGACGACAGCCATG 35 1073 16S_EC_969 ACGCGAAGAACCTTACC 36 ACGACACGAGCTGACGAC 37 1078 23S_EC_182 CTGACACCTGCCCGGTGC 38 GACCGTTATAGTTACGGCC 39 6_1924 23S_EC_264 TCTGTCCCTAGTACGAGAGGAC 40 TGCTTAGATGCTTTCAGC 41 5_2761 CGG 23S_EC_264 CTGTCCCTAGTACGAGACGACC 42 GTTTCATGCTTAGATGCTTTCA 43 5_2767 GG GC 23S_EC_493 GGGGAGTGAAAGAGATCCTGAA 44 ACAAAACGTACGCCGTCACCC 45 _571 ACCG 23S_EC_493 GGGGAGTGAAAGAGATCCTGAA 46 ACAAAAGGCACGCCATCACCC 47 _571_2 ACCG 23S_EC_971 CGAGAGGGAAACAACCCAGACC 48 TGGCTGCTTCTAAGCCAAC 49 1077 INFB_EC_13 TGCTCGTGGTGCACAAGTAACG 50 TGCTCCTTTCGCATGGTTAATT 51 65_1467 GATATTA GCTTCAA RPOC_EC_10 CAAAACTTATTAGGTAAGCGTG 52 TCAAGCGCCATTTCTTTTGGTA 53 18_1124 TTGACT AACCACAT RPOC_EC_10 CAAAACTTATTAGGTAAGCGTG 54 TCAAGCGCCATCTCTTTCGGTA 55 18_1124_2 TTGACT ATCCACAT RPOC_EC_11 TAAGAAGCCGGAAACCATCAAC 56 GGCGCTTGTACTTACCGCAC 57 4_232 TACCG RPOC_EC_21 TGATTCTCGTGCCCGTGGT 58 TTGGCCATCAGGCCACGCATAC 59 78_2246 RPOC_EC_21 TGATTCCGGTGCCCGTGGT 60 TTGGCCATCAGACCACGCATAC 61 78_2246_2 RPOC_EC_22 CTGGCAGGTATGCGTGGTCTGA 62 CGCACCGTGGGTTGAGATGAAG 63 18_2337 TG TAC RPOC_EC_22 CTTGCTGGTATGCGTGGTCTGA 64 CGCACCATGCGTAGAGATGAAG 65 18_2337_2 TG TAC RPOC_EC_80 CGTCGGCTGATTAACCGTAACA 66 GTTTTTCGTTGCGTACGATGAT 67 8_889 ACCG GTC RPOC_EC_80 CGTCGTGTAATTAACCGTAACA 68 ACGTTTTTCGTTTTGAACGATA 69 8_891 ACCG ATGCT RPOC_EC_99 CAAAGGTAAGCAAGGTCGTTTC 70 CGAACGGCCTGAGTAGTCAACA 71 3_1059 CGTCA CG RPOC_EC_99 CAAAGGTAAGCAAGGACGTTTC 72 CGAACGGCCAGAGTAGTCAACA 73 3_1059_2 CGTCA CG TUFB_EC_23 TAGACTGCCCAGGACACGCTG 74 GCCGTCCATCTGAGCAGCACC 75 9_303 TUFB_EC_23 TTGACTGCCCAGGTCACGCTG 76 GCCGTCCATTTGAGCAGCACC 77 9_303_2 TUFB_EC_97 AACTACCGTCCGCAGTTCTACT 78 GTTGTCGCCACGCATAACCATT 79 6_1068 TCC TC TUFB_EC_97 AACTACCGTCCTCACTTCTACT 80 GTTGTCACCAGGCATTACCATT 81 6_1068_2 TCC TC TUFB_EC_98 CCACAGTTCTACTTCCGTACTA 82 TCCAGGCATTACCATTTCTACT 83 5_1062 CTGACG CCTTCTCG RPLB_EC_65 GACCTACAGTAAGAGGTTCTGT 84 TCCAAGTGCTGGTTTACCCCAT 85 0_762 AATGAACC GG RPLB_EC_68 CATCCACACGGTGGTGGTGAAG 86 GTGCTGCTTTACCCCATGGAGT 87 8_757 G RPOC_EC_10 CGTGTTGACTATTCGGGGCGTT 88 ATTCAAGAGCCATTTCTTTTGG 89 36_1126 CAG TAAACCAC RPOB_EC_37 TCAACAACCTCTTGGAGGTAAA 90 TTTCTTGAAGAGTATGAGCTGC 91 62_3865 GCTCAGT TCCGTAAG RBLB_EC_68 CATCCACACGGTGGTGGTGAAG 92 TGTTTTGTATCCAAGTGCTGGT 93 8_771 G TTACCCC VALS_EC_11 CGTGGCGGCGTGGTTATCGA 94 CGGTACGAACTGGATGTCGCCG 95 05_1218 TT RPOB_EC_18 TATCGCTCAGGCGAACTCCAAC 96 GCTGGATTCGCCTTTGCTACG 97 45_1929 RPLB_EC_66 TGTAATGAACCCTAATGACCAT 98 CCAAGTGCTGGTTTACCCCATG 99 9_761 CCACACGG GAGTA RPLB_EC_67 TAATGAACCCTAATGACCATCC 100 TCCAAGTGCTGGTTTACCCCAT 101 1_762 ACACGGTG GGAG RPOB_EC_37 CTTGGAGGTAAGTCTCATTTTG 102 CGTATAAGCTGCACCATAAGCT 103 75_3858 GTGGGCA TGTAATGC VALS_EC_18 CGACGCGCTGCGCTTCAC 104 GCGTTCCACAGCTTGTTGCAGA 105 33_1943 AG RPOB_EC_13 GACCACCTCGGCAACCGT 106 TTCGCTCTCGGCCTGGCC 107 36_1455 TUFB_EC_22 GCACTATGCACACGTAGATTGT 108 TATAGCACCATCCATCTGAGCG 109 5_309 CCTGG GCAC DNAK_EC_42 CGGCGTACTTCAACGACAGCCA 110 CGCGGTCGGCTCGTTGATGA 111 8_522 VALS_EC_19 CTTCTGCAACAAGCTGTGGAAC 112 TCGCAGTTCATCAGCACGAAGC 113 20_1970 GC G TUFB_EC_75 AAGACGACCTGCACGGGC 114 GCGCTCCACGTCTTCACGC 115 7_867 23S_EC_264 CTGTTCTTAGTACGAGAGGACC 116 TTCGTGCTTAGATGCTTTCAG 117 6_2765 16S_EC_969 ACGCGAAGAACCTTACpC 118 ACGACACGAGCpTpGACGAC 119 _1078_3P 16S_EC_972 CGAAGAACpCpTTACC 120 ACACGAGCpTpGAC 121 _1075_4P 16S_EC_972 CGAAGAACCTTACC 122 ACACGAGCTGAC 123 _1075 23S_EC_- CCTGATAAGGGTGAGGTCG 124 ACGTCCTTCATCGCCTCTGA 125 347_59 23S_EC_- GTTGTGAGGTTAAGCGACTAAG 126 CTATCGGTCAGTCAGGAGTAT 127 7_450 23S_EC_- GTTGTGAGGTTAAGCGACTAAG 128 TTGCATCGGGTTGGTAAGTC 129 7_910 23S_EC_430 ATACTCCTGACTGACCGATAG 130 AACATAGCCTTCTCCGTCC 131 _1442 23S_EC_891 GACTTACCAACCCGATGCAA 132 TACCTTAGGACCGTTATAGTTA 133 _1931 CG 23S_EC_142 GGACGGAGAAGGCTATGTT 134 CCAAACACCGCCGTCGATAT 135 4_2494 23S_EC_190 CGTAACTATAACGGTCCTAAGG 136 GCTTACACACCCGGCCTATC 137 8_2852 TA 23S_EC_247 ATATCGACGGCGGTGTTTGG 138 GCGTGACAGGCAGGTATTC 139 5_3209 16S_EC_- AGTCTCAAGAGTGAACACGTAA 140 GCTGCTGGCACGGAGTTA 141 60_525 16S_EC_326 GACACGGTCCAGACTCCTAC 142 CCATGCAGCACCTGTCTC 143 _1058 16S_EC_705 GATCTGGAGGAATACCGGTG 144 ACGGTTACCTTGTTACGACT 145 _1512 16S_EC_126 GAGAGCAAGCGGACCTCATA 146 CCTCCTGCGTGCAAAGC 147 8_1775 GROL_EC_94 TGGAAGATCTGGGTCAGGC 148 CAATCTGCTGACGGATCTGAGC 149 1_1060 INFB_EC_11 GTCGTGAAAACGAGCTGGAAGA 150 CATGATGGTCACAACCGG 151 03_1191 HFLB_EC_10 TGGCGAACCTGGTGAACGAAGC 152 CTTTCGCTTTCTCGAACTCAAC 153 82_1168 CAT INFB_EC_19 CGTCAGGGTAAATTCCGTGAAG 154 AACTTCGCCTTCGGTCATGTT 155 69_2058 TTAA GROL_EC_21 GGTGAAAGAAGTTGCCTCTAAA 156 TTCAGGTCCATCGGGTTCATGC 157 9_350 GC C VALS_EC_11 CGTGGCGGCGTGGTTATCGA 158 ACGAACTGGATGTCGCCGTT 159 05_1214 16S_EC_556 CGGAATTACTGGGCGTAAAG 160 CGCATTTCACCGCTACAC 161 _700 RPOC_EC_12 ACCCAGTGCTGCTGAACCGTGC 162 GTTCAAATGCCTGGATACCCA 163 56_1315 16S_EC_774 GGGAGCAAACAGGATTAGATAC 164 CGTACTCCCCAGGCG 165 _894 RPOC_EC_15 TGGCCCGAAAGAAGCTGAGCG 166 ACGCGGGCATGCAGAGATGCC 167 84_1643 16S_EC_108 ATGTTGGGTTAAGTCCCGC 168 TGACGTCATCCCCACCTTCC 169 2_1196
16S_EC_138 CTTGTACACACCGCCCGTC 170 AAGGAGGTGATCCAGCC 171 9_1541 16S_EC_130 CGGATTGGAGTCTGCAACTCG 172 GACGGGCGGTGTGTACAAG 173 3_1407 23S_EC_23-- GGTGGATGCCTTGGC 174 GGGTTTCCCCATTCGG 175 130 23S_EC_187 GGGAACTGAAACATCTAAGTA 176 TTCGCTCGCCGCTAC 177 _256 23S_EC_160 TACCCCAAACCGACACAGG 178 CCTTCTCCCGAAGTTACG 179 2_1703 23S_EC_168 CCGTAACTTCGGGAGAAGG 180 CACCGGGCAGGCGTC 181 5_1842 23S_EC_182 GACGCCTGCCCGGTGC 182 CCGACAAGGAATTTCGCTACC 183 7_1949 23S_EC_243 AAGGTACTCCGGGGATAACAGG 184 AGCCGACATCGAGGTGCCAAAC 185 4_2511 C 23S_EC_259 GACAGTTCGGTCCCTATC 186 CCGGTCCTCTCGTACTA 187 9_2669 23S_EC_265 TAGTACGAGAGGACCGG 188 TTAGATGCTTTCAGCACTTATC 189 3_2758 23S_BS_- AAACTAGATAACAGTAGACATC 190 GTGCGCCCTTTCTAACTT 191 68_21 AC 16S_EC_8_3 AGAGTTTGATCATGGCTCAG 192 ACTGCTGCCTCCCGTAG 193 58 16S_EC_314 CACTGGAACTGAGACACGG 194 CTTTACGCCCAGTAATTCCG 195 _575 16S_EC_518 CCAGCAGCCGCGGTAATAC 196 GTATCTAATCCTGTTTGCTCCC 197 _795 16S_EC_683 GTGTAGCGGTGAAATGCG 198 GGTAAGGTTCTTCGCGTTG 199 _985 16S_EC_937 AAGCGGTGGAGCATGTGG 200 ATTGTAGCACGTGTGTAGCCC 201 _1240 16S_EC_119 CAAGTCATCATGGCCCTTA 202 AAGGAGGTGATCCAGCC 203 5_1541 16S_EC_8_1 ACAGTTTGATCATGGCTCAG 204 AAGGAGGTGATCCAGCC 205 541 23S_EC_183 ACCTGCCCAGTGCTGGAAG 206 TCGCTACCTTAGGACCGT 207 1_1936 16S_EC_138 GCCTTGTACACACCTCCCGTC 208 CACGGCTACCTTGTTACGAC 209 7_1513 16S_EC_139 TTGTACACACCGCCCGTCATAC 210 CCTTGTTACGACTTCACCCC 211 0_1505 16S_EC_136 TACGGTGAATACGTTCCCGGG 212 ACCTTGTTACGACTTCACCCCA 213 7_1506 16S_EC_804 ACCACGCCGTAAACGATGA 214 CCCCCGTCAATTCCTTTGAGT 215 _929 16S_EC_791 GATACCCTGGTAGTCCACACCG 216 GCCTTGCGACCGTACTCCC 217 _904 16S_EC_789 TAGATACCCTGGTAGTCCACGC 218 GCGACCGTACTCCCCAGG 219 _899 16S_EC_109 TAGTCCCGCAACGAGCGC 220 GACGTCATCCCCACCTTCCTCC 221 2_1195 23S_EC_258 TAGAACGTCGCGAGACAGTTCG 222 AGTCCATCCCGGTCCTCTCG 223 6_2677 HEXAMER_EC GAGGAAAGTCCGGGCTC 224 ATAAGCCGGGTTCTGTCG 225 61_362 RNASEP_BS-- GAGGAAAGTCCATGCTCGC 226 GTAAGCCATGTTTTGTTCCATC 227 43_384 RNASEP_EC-- GAGGAAAGTCCGGGCTC 228 ATAAGCCGGGTTCTGTCG 229 61_362 YAED_TRNA-- GCGGGATCCTCTAGAGGTGTTA 230 GCGGGATCCTCTAGAAGACCTC 231 ALA- AATAGCCTGGCAG CTGCGTGCAAAGC RRNH_EC_51 3_49 RNASEP_SA-- GAGGAAAGTCCATGCTCAC 232 ATAAGCCATGTTCTGTTCCATC 233 31_379 16S_EC_108 ATGTTGGGTTAAGTCCCGC 234 AAGGAGGTGATCCAGCC 235 2_1541 16S_EC_556 CGGAATTACTGGGCGTAAAG 236 GTATCTAATCCTGTTTGCTCCC 237 _795 16S_EC_108 ATGTTGGGTTAAGTCCCGC 238 TGACGTCATGCCCACCTTCC 239 2_1196_10G 16S_EC_108 ATGTTGGGTTAAGTCCCGC 240 TGACGTCATGGCCACCTTCC 241 2_1196_10G _11G TRNA_ILERR GCGGGATCCTCTAGACCTGATA 242 GCGGGATCCTCTAGAGCGTGAC 243 NH_ASPRRNH AGGGTGAGGTCG AGGCAGGTATTC _EC_32_41 16S_EC_969 ACGCGAAGAACCTTACC 244 GACGGGCGGTGTGTACAAG 245 _1407 16S_EC_683 GTGTAGCGGTGAAATGCG 246 CGAGTTGCAGACTGCGATCCG 247 _1323 16S_EC_49-- TAACACATGCAAGTCGAACG 248 CGTACTCCCCAGGCG 249 894 16S_EC_49-- TAACACATGCAAGTCGAACG 250 ACGACACGAGCTGACGAC 251 1078 CYA_BA_134 ACAACGAAGTACAATACAAGAC 252 CTTCTACATTTTTAGCCATCAC 253 9_1447 16S_EC_109 TTAAGTCCCGCAACGAGCGCAA 254 TGACGTCATCCCCACCTTCCTC 255 0_1196_2 16S_EC_405 TGAGTGATGAAGGCCTTACGGT 256 CGGCTGCTGGCACGAAGTTAG 257 _527 TGTAAA GROL_EC_49 ATGGACAAGGTTGGCAAGGAAG 258 TAGCCGCGGTCGAATTGCAT 259 6_596 G GROL_EC_51 AAGGAAGGCGTGATCACCGTTG 260 CCGCGGTCGAATTGCATGCCTT 261 1_593 AAGA C VALS_EC_18 ACGCGCTGCGCTTCAC 262 TTGCAGAAGTTGCGGTAGCC 263 35_1928 RPOB_EC_13 TCGACCACCTGGGCAACC 264 ATCAGGTCGTGCGGCATCA 265 34_1478 DNAK_EC_42 CACGGTGCCGGCGTACT 266 GCGGTCGGCTCGTTGATGAT 267 0_521 RPOB_EC_37 TTGGAGGTAAGTCTCATTTTGG 268 AAGCTGCACCATAAGCTTGTAA 269 76_3853 TGG TGC RPOB_EC_38 CAGCGTTTCGGCGAAATGGA 270 CGACTTGACGGTTAACATTTCC 271 02_3885 TG RPOB_EC_37 GGGCAGCGTTTCGGCGAAATGG 272 GTCCGACTTGACGGTCAACATT 273 99_3888 A TCCTG RPOC_EC_21 CAGGAGTCGTTCAACTCGATCT 274 ACGCCATCAGGCCACGCAT 275 46_2245 ACATGAT ASPS_EC_40 GCACAACCTGCGGCTGCG 276 ACGGCACGAGGTAGTCGC 277 5_538 RPOC_EC_13 CGCCGACTTCGACGGTGACC 278 GAGCATCAGCGTGCGTGCT 279 74_1455 TUFB_EC_95 CCACACGCCGTTCTTCAACAAC 280 GGCATCACCATTTCCTTGTCCT 281 7_1058 T TCG 16S_EC_7_1 GAGAGTTTGATCCTGGCTCAGA 282 TGTTACTCACCCGTCTGCCACT 283 22 ACGAA VALS_EC_61 ACCGAGCAAGGAGACCAGC 284 TATAACGCACATCGTCAGGGTG 285 0_727 A
[0174]For evaluation in the laboratory, five species of bacteria were selected including three γ-proteobacteria (E. coli, K pneumoniae and P. auergiosa) and two low G+C gram positive bacteria (B. subtilitis and S. aureus). The identities of the organisms were not revealed to the laboratory technicians.
[0175]Bacteria were grown in culture, DNA was isolated and processed, and PCR performed using standard protocols. Following PCR, all samples were desalted, concentrated, and analyzed by Fourier Transform Ion Cyclotron Resonance (FTICR) mass spectrometry. Due to the extremely high precision of the FTICR, masses could be measured to within 1 Da and unambiguously deconvoluted to a single base composition. The measured base compositions were compared with the known base composition signatures in our database. As expected when using broad range survey 16S primers, several phylogenetic near-neighbor organisms were difficult to distinguish from our test organisms. Additional non-ribosomal primers were used to triangulate and further resolve these clusters.
[0176]An example of the use of primers directed to regions of RNA polymerase B (rpoB) is shown in FIG. 19. This gene has the potential to provide broad priming and resolving capabilities. A pair of primers directed against a conserved region of rpoB provided distinct base composition signatures that helped resolve the tight enterobacteriae cluster. Joint probability estimates of the signatures from each of the primers resulted in the identification of a single organism that matched the identity of the test sample. Therefore a combination of a small number of primers that amplify selected regions of the 16S ribosomal RNA gene and a few additional primers that amplify selected regions of protein encoding genes provide sufficient information to detect and identify all bacterial pathogens.
Example 16
Detection of Staphylococcus aureus in Blood Samples
[0177]Blood samples in an analysis plate were spiked with genomic DNA equivalent of 103 organisms/ml of Staphylococcus aureus. A single set of 16S rRNA primers was used for amplification. Following PCR, all samples were desalted, concentrated, and analyzed by Fourier Transform Ion Cyclotron Resonance (FTICR) mass spectrometry. In each of the spiked wells, strong signals were detected which are consistent with the expected BCS of the S. aureus amplicon (FIG. 20). Furthermore, there was no robotic carryover or contamination in any of the blood only or water blank wells. Methods similar to this one will be applied for other clinically relevant samples including, but not limited to: urine and throat or nasal swabs.
Example 17
Detection and Serotyping of Viruses
[0178]The virus detection capability of the present invention was demonstrated in collaboration with Naval health officers using adenoviruses as an example.
[0179]All available genomic sequences for human adenoviruses available in public databases were surveyed. The hexon gene was identified as a candidate likely to have broad specificity across all serotypes. Four primer pairs were selected from a group of primers designed to yield broad coverage across the majority of the adenoviral strain types (Table 9) wherein Tp=5'propynylated uridine and Cp=5'propynylated cytidine.
TABLE-US-00011 TABLE 9 Intelligent Primer Pairs for Serotyping of Adenoviruses Forward Reverse Primer Pair Forward Primer SEQ ID Reverse Primer SEQ ID Name Sequence NO: Sequence NO: HEX_HAD7 + 4 + 2 AGACCCAATTACATTGGCTT 286 CCACTGCTGTTGTAGTACAT 287 1_934_995 HEX_HAD7 + 4 + 2 ATGTACTACAACAGTACTGG 288 CAAGTCAACCACAGCATTCA 289 1_976_1050 HEX_HAD7 + 4 + 2 GGGCTTATGTACTACAACAG 290 TCTGTCTTGCAAGTCAACCAC 291 1_970_1059 HEX_HAD7 + 3_7 GGAATTTTTTGATGGTAGAGA 292 TAAAGCACAATTTCAGGCG 293 71_827 HEX_HAD4 + 16-- TAGATCTGGCTTTCTTTGAC 294 ATATGAGTATCTGGAGTCTGC 295 746_848 HEX_HAD7_509 GGAAAGACATTACTGCAGACA 296 CCAACTTGAGGCTCTGGCTG 297 _578 HEX_HAD4_121 ACAGACACTTACCAGGGTG 298 ACTGTGGTGTCATCTTTGTC 299 6_1289 HEX_HAD21_51 TCACTAAAGACAAAGGTCTTCC 300 GGCTTCGCCGTCTGTAATTTC 301 5_567 HEX_HAD_1342 CGGATCCAAGCTAATCTTTGG 302 GGTATGTACTCATAGGTGTTG 303 _1469 GTG HEX_HAD7 + 4 + 2 AGACpCpCAATTpACpATpTGG 304 CpCpAGTGCTGTpTpGTAGTA 305 1_934_995P CTT CAT HEX_HAD7 + 4 + 2 ATpGTpACTpACAACAGTACpT 306 CAAGTpCpAACCACAGCATpT 307 1_976_1050P pGG pCA HEX_HAD7 + 4 + 2 GGGCpTpTATpGTpACTACAAC 308 TCTGTpCpTTGCAAGTpCpAA 309 1_970_1059P pAG CCAC HEX_HAD7 + 3_7 GGAATTpTpTpTpTGATGGTAG 310 TAAAGCACAATpTpTpCpAGG 311 71_827P AGA CG HEX_HAD4 + 16-- TAGATCTGGCTpTpTpCpTTTG 312 ATATGAGTATpCpTpGGAGTp 313 746_848P AC CpTGC HEX_HAD_1342 CGGATpCCAAGCpTAATCpTpT 314 GGTATGTACTCATAGGTCTpT 315 _1469P TGG pGGTG HEX_HAD7 + 21 + AACAGACCCAATTACATTGGCT 316 GAGGCACTTGTATGTGGAAAG 317 3_931_1645 T G HEX_HAD4 + 2_9 ATGCCTAACAGACCCAATTACA 318 TTCATGTAGTCGTAGGTGTTG 319 25_1469 T G HEX_HAD7 + 21 + CGCGCCTAATACATCTCAGTGG 320 AAGCCAATGTAATTGCGTCTG 321 3_384_953 AT TT HEX_HAD4 + 2_3 CTACTCTGGCACTGCCTACAAC 322 ATGTAATTGGGTCTGTTAGGC 323 45_947 AT HEX_HAD2_772 CAATCCGTTCTGGTTCCGGATG 324 CTTGCCGGTCGTTCAAAGAGG 325 _865 AA TAG HEX_HAD7 + 4 + 2 AGTCCGGGTCTGGTGCAG 326 CGGTCGGTGGTCACATC 327 1_73_179 HEX_HAD7 + 4 + 2 ATGGCCACCCCATCGATG 328 CTGTCCGGCGATGTGCATG 329 1_1_54 HEX_HAD7 + 4 + 2 GGTCGTTATGTGCCTTTCCACA 330 TCCTTTCTGAAGTTCCACTCA 331 1_1612_1718 T TAGG HEX_HAD7 + 4 + 2 ACAACATTGGCTACCAGGGCTT 332 CCTGCCTGCTCATAGGCTGGA 333 1_2276_2368 AGTT
[0180]These primers also served to clearly distinguish those strains responsible for most disease (types 3, 4, 7 and 21) from all others. DNA isolated from field samples known to contain adenoviruses were tested using the hexon gene PCR primers, which provided unambiguous strain identification for all samples. A single sample was found to contain a mixture of two viral DNAs belonging to strains 7 and 21.
[0181]Test results (FIG. 21) showed perfect concordance between predicted and observed base composition signatures for each of these samples. Classical serotyping results confirmed each of these observations. Processing of viral samples directly from collection material such as throat swabs rather than from isolated DNA, will result in a significant increase in throughput, eliminating the need for virus culture.
Example 18
Broad Rapid Detection and Strain Typing of Respiratory Pathogens for Epidemic Surveillance
[0182]Genome preparation: Genomic materials from culture samples or swabs were prepared using a modified robotic protocol using DNeasy® 96 Tissue Kit, Qiagen). Cultures of Streptococcus pyogenes were pelleted and transferred to a 1.5 mL tube containing 0.45 g of 0.7 mm Zirconia beads (Biospec Products, Inc.). Cells were lysed by shaking for 10 minutes at a speed of 19 l/s using a MM300 Vibration Mill (Retsch, Germany). The samples were centrifuged for 5 min and the supernatants transferred to deep well blocks and processed using the manufacture's protocol and a Qiagen 8000 BioRobot.
[0183]PCR: PCR reactions were assembled using a Packard MPII liquid handling platform and were performed in 50 μL volume using 1.8 units each of Platinum Taq (Invitrogen) and Hotstart PFU Turbo (Stratagene) polymerases. Cycling was performed on a DNA Engine Dyad (MJ Research) with cycling conditions consisting of an initial 2 min at 95° C. followed by 45 cycles of 20 at 95° C., 15 s at 58° C., and 15 s at 72° C.
[0184]Broad-range primers: PCR primer design for base composition analysis from precise mass measurements is constrained by an upper limit where ionization and accurate deconvolution can be achieved. Currently, this limit is approximately 140 base pairs. Primers designed to broadly conserved regions of bacterial ribosomal RNAs (16 and 23 S) and the gene encoding ribosomal protein L3 (rpoC) are shown in Table 10.
TABLE-US-00012 TABLE 10 Broad Range Primer Pairs SEQ Length Target Direc- ID of Gene tion Primer NO Amplicon 16S_1 F GGATTAGAGACCCTGGTAGTCC 334 116 16S_1 R GGCCGTACTCCCCAGGCG 335 116 16S_2 F TTCGATGCAACGCGAAGAACCT 336 115 16S_2 R ACGAGCTGACGACAGCCATG 337 115 23S F TCTGTCCCTAGTACGAGAGGACCGG 338 118 23S R TGCTTAGATGCTTTCAGC 339 118 rpoC F CTGGCAGGTATGCGTGGTCTGATG 340 121 rpoC R CGCACCGTGGGTTGAGATGAAGTAC 341 121
[0185]Emm-typing primers: The allelic profile of a GAS strain by Multilocus Sequencing Technique (MLST) can be obtained by sequencing the internal fragments of seven housekeeping genes. The nucleotide sequences for each of these housekeeping genes, for 212 isolates of GAS (78 distinct emm types), are available (www.mlst.net). This corresponds to one hundred different allelic profiles or unique sequence types, referred to by Enright et al. as ST1-ST100 (Enright, M. C., et al., Infection and Immunity 2001, 69, 2416-2427). For each sequence type, we created a virtual transcript by concatenating sequences appropriate to their allelic profile from each of the seven genes. MLST primers were designed using these sequences and were constrained to be within each gene loci. Twenty-four primer pairs were initially designed and tested against the sequenced GAS strain 700294. A final subset of six primer pairs Table 11 was chosen based on a theoretical calculation of minimal number of primer pairs that maximized resolution of between emm types.
TABLE-US-00013 TABLE 11 Drill-Down Primer Pairs Used in Determining emm-type Length SEQ of Target Direc- ID Ampli- Gene tion Primer NO con gki F GGGGATTCAGCCATCAAAGCAGCTATTGA 342 116 C gki R CCAACCTTTTCCACAACAGAATCAGC 343 116 gtr F CCTTACTTCGAACTATGAATCTTTTGGAA 344 115 G gtr R CCCATTTTTTCACGCATGCTGAAAATATC 345 115 murI F CGCAAAAAAATCCAGCTATTAGC 346 118 murI R AAACTATTTTTTTAGCTATACTCGAACAC 347 118 mutS F ATGATTACAATTCAAGAAGGTCGTCACGC 348 121 mutS R TTGGACCTGTAATCAGCTGAATACTGG 349 121 xpt F GATGACTTTTTAGCTAATGGTCAGGCAGC 350 122 xpt R AATCGACGACCATCTTGGAAAGATTTCTC 351 122 yqiL F GCTTCAGGAATCAATGATGGAGCAG 352 119 yqiL R GGGTCTACACCTGCACTTGCATAAC 353 119
[0186]Microbiology: GAS isolates were identified from swabs on the basis of colony morphology and beta-hemolysis on blood agar plates, gram stain characteristics, susceptibility to bacitracin, and positive latex agglutination reactivity with group A-specific antiserum.
[0187]Sequencing: Bacterial genomic DNA samples of all isolates were extracted from freshly grown GAS strains by using QIAamp DNA Blood Mini Kit (Qiagen, Valencia, Calif.) according to the procedures described by the manufacture. Group A streptococcal cells were subjected to PCR and sequence analysis using emm-gene specific PCR as previously described (Beall, B., et al. J. Clin. Micro., 1996, 34, 953-958; Facklam, R., et al. Emerg. Infect. Dis. 1999, 5, 247-253). Homology searches on DNA sequences were conducted against known emm sequences present in (www.cdc.gov/ncidod/biotech/infotech_hp.html). For MLST analysis, internal fragments of seven housekeeping genes, were amplified by PCR and analyzed as previously described (Enright, M. C., et al., Infection and Immunity 2001, 69, 2416-2427). The emm-type was determined from comparison to the MLST database.
[0188]Broad Range Survey/Drill-Down Process (100): For Streptococcus pyogenes, the objective was the identification of a signature of the virulent epidemic strain and determination of its emm-type. Emm-type information is useful both for treatment considerations and epidemic surveillance. A total of 51 throat swabs were taken both from healthy recruits and from hospitalized patients in December 2002, during the peak of a GAS outbreak at a military training camp. Twenty-seven additional isolates from previous infections ascribed to GAS were also examined. Initially, isolated colonies were examined both from throat culture samples and throat swabs directly without the culture step. The latter path can be completed within 6-12 hours providing information on a significant number of samples rapidly enough to be useful in managing an ongoing epidemic.
[0189]The process of broad range survey/drill-down (200) is shown in FIG. 22. A clinical sample such as a throat swab is first obtained from an individual (201). Broad range survey primers are used to obtain amplification products from the clinical sample (202) which are analyzed to determine a BCS (203) from which a species is identified (204). Drill-down primers are then employed to obtain PCR products (205) from which specific information is obtained about the species (such as Emm-type) (206).
[0190]Broad Range Survey Priming: Genomic regions targeted by the broad range survey primers were selected for their ability to allow amplification of virtually all known species of bacteria and for their capability to distinguish bacterial species from each other by base composition analysis. Initially, four broad-range PCR target sites were selected and the primers were synthesized and tested. The targets included universally conserved regions of 16S and 23S rRNA, and the gene encoding ribosomal protein L3 (rpoC).
[0191]While there was no special consideration of Streptococcus pyogenes in the selection of the broad range survey primers (which were optimized for distinguishing all important pathogens from each other), analysis of genomic sequences showed that the base compositions of these regions distinguished Streptococcus pyogenes from other respiratory pathogens and normal flora, including closely related species of streptococci, staphylococci, and bacilli (FIG. 23).
[0192]Drill Down Priming (Emm-Typing): In order to obtain strain-specific information about the epidemic, a strategy was designed to measure the base compositions of a set of fast clock target genes to generate strain-specific signatures and simultaneously correlate with emm-types. In classic MLST analysis, internal fragments of seven housekeeping genes (gki, gtr, muri, mutS, recP, xpt, yqiL) are amplified, sequenced and compared to a database of previously studied isolates whose emm-types have been determined (Horner, M. J., et al. Fundamental and Applied Toxicology, 1997, 36, 147). Since the analysis enabled by the present embodiment of the present invention provides base composition data rather than sequence data, the challenge was to identify the target regions that provide the highest resolution of species and least ambiguous emm-classification. The data set from Table 2 of Enright et al. (Enright, M. C., et al. Infection and Immunity, 2001, 69, 2416-2427) to bioinformatically construct an alignment of concatenated alleles of the seven housekeeping genes from each of 212 previously emm-typed strains, of which 101 were unique sequences that represented 75 distinct emm-types. This alignment was then analyzed to determine the number and location of the optimal primer pairs that would maximize strain discrimination strictly on base composition data.
[0193]An example of assignment of BCSs of PCR products is shown in FIG. 24 where PCR products obtained using the gtr primer (a drill-down emm-typing primer) from two different swab samples were analyzed (sample 12--top and sample 10--bottom). The deconvoluted ESI-FCTIR spectra provide accurate mass measurements of both strands of the PCR products, from which a series of candidate BCSs were calculated from the measured mass (and within the measured mass uncertainty). The identification of complementary candidate BCSs from each strand provides a means for unambiguous assignment of the BCS of the PCR product. BCSs and molecular masses for each strand of the PCR product from the two different samples are also shown in FIG. 24. In this case, the determination of BCSs for the two samples resulted in the identification of the emm-type of Streptococcus pyogenes-sample 12 was identified as emm-type 3 and sample 10 was identified as emm-type 6.
[0194]The results of the composition analysis using the six primer pairs, 5'-emm gene sequencing and MLST gene sequencing method for the GAS epidemic at a military training facility are compared in FIG. 25. The base composition results for the six primer pairs showed a perfect concordance with 5'-emm gene sequencing and MLST sequencing methods. Of the 51 samples taken during the peak of the epidemic, all but three had identical compositions and corresponded to emm-type 3. The three outliers, all from healthy individuals, probably represent non-epidemic strains harbored by asymptomatic carriers. Samples 52-80, which were archived from previous infections from Marines at other naval training facilities, showed a much greater heterogeneity of composition signatures and emm-types.
Example 19
Base Composition Probability Clouds
[0195]FIG. 18 illustrates the concept of base composition probability clouds via a pseudo-four dimensional plot of base compositions of enterobacteria including Y. pestis, Y. psuedotuberculosis, S. typhimurium, S. typhi, Y. enterocolitica, E. coli K12, and E. coli O157:H7. In the plot of FIG. 18, A, C and G compositions correspond to the x, y and z axes respectively whereas T compositions are represented by the size of the sphere at the junction of the x, y and z coordinates. There is no absolute requirement for having a particular nucleobase composition associated with a particular axis. For example, a plot could be designed wherein G, T and C compositions correspond to the x, y and z axes respectively whereas the A composition corresponds to the size of the sphere at the junction of the x, y and z coordinates. Furthermore, a different representation can be made of the "pseudo fourth" dimension i.e.: other than the size of the sphere at junction of the x, y and z coordinates. For example, a symbol having vector information such as an arrow or a cone can be rotated at an angle which varies proportionally with the composition of the nucleobase corresponding to the pseudo fourth dimension. The choice of axes and pseudo fourth dimensional representation is typically made with the aim of optimal visualization of the data being presented.
[0196]A similar base composition probability cloud analysis has been presented for a series of viruses in U.S. provisional patent application Ser. No. 60/431,319, which is commonly owned and incorporated herein by reference in its entirety. In this base composition probability cloud analysis, the closely related Dengue virus types 1-4 are clearly distinguishable from each other. This example is indicative of a challenging scenario for species identification based on BCS analysis because RNA viruses have a high mutation rate, it would be expected to be difficult to resolve closely related species. However, as this example illustrates, BCS analysis, aided by base composition probability cloud analysis is capable of resolution of closely related viral species.
[0197]A base composition probability cloud can also be represented as a three dimensional plot instead of a pseudo-four dimensional plot. An example of such a three dimensional plot is a plot of G, A and C compositions correspond to the x, y and z axes respectively, while the composition of T is left out of the plot. Another such example is a plot where the compositions of all four nucleobases is included: G, A and C+T compositions correspond to the x, y and z axes respectively. As for the pseudo-four dimensional plots, the choice of axes for a three dimensional plot is typically made with the aim of optimal visualization of the data being presented.
Example 20
Biochemical Processing of Large Amplification Products for Analysis by Mass Spectrometry
[0198]In the example illustrated in FIG. 26, a primer pair which amplifies a 986 bp region of the 16S ribosomal gene in E. coli (K12) was digested with a mixture of 4 restriction enzymes: BstNI, BsmFI, Bfal, and NcoI. FIG. 26(a) illustrates the complexity of the resulting ESI-FTICR mass spectrum which contains multiple charge states of multiple restriction fragments. Upon mass deconvolution to neutral mass, the spectrum is significantly simplified and discrete oligonucleotide pairs are evident (FIG. 26b). When base compositions are derived from the masses of the restriction fragments, perfect agreement is observed for the known sequence of nucleotides 1-856 (FIG. 26c); the batch of NcoI enzyme used in this experiment was inactive and resulted in a missed cleavage site and a 197-mer fragment went undetected as it is outside the mass range of the mass spectrometer under the conditions employed. Interestingly however, both a forward and reverse strand were detected for each fragment measured (solid and dotted lines in, respectively) within 2 ppm of the predicted molecular weights resulting in unambiguous determination of the base composition of 788 nucleotides of the 985 nucleotides in the amplicon. The coverage map offers redundant coverage as both 5' to 3' and 3' to 5' fragments are detected for fragments covering the first 856 nucleotides of the amplicon.
[0199]This approach is in many ways analogous to those widely used in MS-based proteomics studies in which large intact proteins are digested with trypsin, or other proteolytic enzyme(s), and the identity of the protein is derived by comparing the measured masses of the tryptic peptides with theoretical digests. A unique feature of this approach is that the precise mass measurements of the complementary strands of each digest product allow one to derive a de novo base composition for each fragment, which can in turn be "stitched together" to derive a complete base composition for the larger amplicon. An important distinction between this approach and a gel-based restriction mapping strategy is that, in addition to determination of the length of each fragment, an unambiguous base composition of each restriction fragment is derived. Thus, a single base substitution within a fragment (which would not be resolved on a gel) is readily observed using this approach. Because this study was performed on a 7 Tesla ESI-FTICR mass spectrometer, better than 2 ppm mass measurement accuracy was obtained for all fragments. Interestingly, calculation of the mass measurement accuracy required to derive unambiguous base compositions from the complementary fragments indicates that the highest mass measurement accuracy actually required is only 15 ppm for the 139 bp fragment (nucleotides 525-663). Most of the fragments were in the 50-70 bp size-range which would require mass accuracy of only ˜50 ppm for unambiguous base composition determination. This level of performance is achievable on other more compact, less expensive MS platforms such as the ESI-TOF suggesting that the methods developed here could be widely deployed in a variety of diagnostic and human forensic arenas.
[0200]This example illustrates an alternative approach to derive base compositions from larger PCR products. Because the amplicons of interest cover many strain variants, for some of which complete sequences are not known, each amplicon can be digested under several different enzymatic conditions to ensure that a diagnostically informative region of the amplicon is not obscured by a "blind spot" which arises from a mutation in a restriction site. The extent of redundancy required to confidently map the base composition of amplicons from different markers, and determine which set of restriction enzymes should be employed and how they are most effectively used as mixtures can be determined. These parameters will be dictated by the extent to which the area of interest is conserved across the amplified region, the compatibility of the various restriction enzymes with respect to digestion protocol (buffer, temperature, time) and the degree of coverage required to discriminate one amplicon from another.
Example 21
Identification of members of the Viral Genus Orthopoxvirus
[0201]Primer sites were identified on three essential viral genes--the DNA-dependent polymerase (DdDp), and two sub-units of DNA-dependent RNA polymerases A and B (DdRpA and DdRpB). These intelligent primers designed to identify members of the viral genus Orthopoxvirus are shown in Table 12 wherein Tp=5'propynylated uridine and Cp=5'propynylated cytidine.
TABLE-US-00014 TABLE 12 Intelligent Primer Pairs for Identification of members of the Viral Genus Orthopoxvirus Forward Reverse Primer Pair Forward Primer SEQ ID Reverse Primer SEQ ID Name Sequence NO: Sequence NO: A25L_NC00161 GTACTGAATCCGCCTAAG 354 GTGAATAAAGTATCGCCCTAA 355 1_28_127 TA A18R_NC00161 GAAGTTGAACCGGGATCA 356 ATTATCGGTCGTTGTTAATGT 357 1_100_207 A18R_NC00161 CTGTCTGTAGATAAACTAGGAT 358 CGTTCTTCTCTGGAGGAT 359 1_1348_1445 T E9L_NC001611 CGATACTACGGACGC 360 CTTTATGAATTACTTTACATA 361 _1119_1222 T K8R_NC001611 CTCCTCCATCACTAGGAA 362 CTATAACATTGAAAGCTTATT 363 _221_311 G A24R_NC00161 CGCGATAATAGATAGTGCTAAA 364 GCTTCCACCAGGTCATTAA 365 1_795_878 C A25L_NC00161 GTACpTpGAATpCpCpGCpCpT 366 GTGAATAAAGTATpCpGCpCp 367 1_28_127P AAG CpTpAATA A18R_NC00161 GAAGTpTpGAACpCpGGGATCA 368 ATTATCGGTpCpGTpTpGTpT 369 1_100_207P pAATGT A18R_NC00161 CTGTpCpTpGTAGATAAACpTp 370 CGTTCpTpTpCpTpCpTpGGA 371 1_1348_1445P AGGATT GGAT E9L_NC001611 CGATACpTpACpGGACGC 372 CTTTATGAATpTpACpTpTpT 373 _1119_1222P pACATAT K8R_NC001611 CTpCpCpTCpCpATCACpTpAG 374 CTATAACATpTpCpAAAGCpT 375 _221_311P GAA pTpATTG A24R_NC00161 CGCGATpAATpAGATAGTpGCp 376 GCTTCpCpACpCAGGTpCATp 377 1_795_8782 TpAAAC TAA
[0202]As illustrated in FIG. 27, members of the Orthopoxvirus genus group can be identified, distinguished from one another, and distinguished from other members of the Poxvirus family using a single pair of primers designed against the DdRpB gene.
[0203]Since the primers were designed across regions of high conservation within this genus, the likelihood of missed detection due to sequence variations at these sites is minimized. Further, none of the primers is expected to amplify other viruses or any other DNA, based on the data available in GenBank. This method can be used for all families of viral threat agents and is not limited to members of the Orthopoxvirus genus.
Example 22
Identification of Viruses that Cause Viral Hemorrhagic Fevers
[0204]In accordance with the present invention an approach of broad PCR priming across several different viral species is employed using conserved regions in the various viral genomes, amplifying a small, yet highly informative region in these organisms, and then analyzing the resultant amplicons with mass spectrometry and data analysis. These regions will be tested with live agents, or with genomic constructs thereof.
[0205]Detection of RNA viruses will necessitate a reverse transcription (RT) step prior to the PCR amplification of the TIGER reporter amplicon. To maximize throughput and yield while minimizing the handling of the samples, commercial one-step reverse transcription polymerase chain reaction (RT-PCR) kits will be evaluated for use. If necessary, a one-step RT-PCR mix using our selected DNA polymerase for the PCR portion of the reaction will be developed. To assure there is no variation in our reagent performance all new lots of enzymes, nucleotides and buffers will be individually tested prior to use.
[0206]Various modifications of the invention, in addition to those described herein, will be apparent to those skilled in the art from the foregoing description. Such modifications are also intended to fall within the scope of the appended claims. Each reference cited in the present application is incorporated herein by reference in its entirety
Sequence CWU
1
382190RNABacillus anthracismisc_feature(20)..(20)N = A, U, G or C
1gcgaagaacc uuaccaggun uugacauccu cugacaaccc uagagauagg gcuucuccuu
60cgggagcaga gugacaggug gugcaugguu
90290RNABacillus cereus 2gcgaagaacc uuaccagguc uugacauccu cugaaaaccc
uagagauagg gcuucuccuu 60cgggagcaga gugacaggug gugcaugguu
9031542RNAArtificial Sequencemisc_feature16S rRNA
consensus sequence 3nnnnnnnaga guuugaucnu ggcucagnnn gaacgcuggc
ggnnngcnun anacaugcaa 60gucgancgnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn
agnggcnnac gggugaguaa 120nncnunnnna nnunccnnnn nnnnnggnan annnnnnnga
aannnnnnnu aauaccnnau 180nnnnnnnnnn nnnnaaagnn nnnnnnnnnn nnnnnnnnnn
nnnnnngann nnnnnnngnn 240nnaunagnun guuggunngg uaanggcnna ccaagncnnn
gannnnuagc ngnncugaga 300ggnngnncng ccacanuggn acugaganac ggnccanacu
ccuacgggag gcagcagunn 360ggaaunuunn ncaauggnng naanncugan nnagcnannc
cgcgugnnng anganggnnu 420nnngnungua aannncunun nnnnnngang annnnnnnnn
nnnnnnnnnn nnnnnnnnnu 480gacnnuannn nnnnannaag nnncggcnaa cuncgugcca
gcagccgcgg uaauacgnag 540gnngcnagcg uunnncggan unanugggcg uaaagngnnn
gnaggnggnn nnnnnngunn 600nnngunaaan nnnnnngcun aacnnnnnnn nnncnnnnnn
nacnnnnnnn cungagnnnn 660nnagnggnnn nnngaauunn nnguguagng gugnaauncg
naganaunng nangaanacc 720nnungcgaag gcnnnnnncu ggnnnnnnac ugacncunan
nnncgaaagc nugggnagcn 780aacaggauua gauacccugg uaguccangc nnuaaacgnu
gnnnnnunnn ngnnngnnnn 840nnnnnnnnnn nnnnnnnnna nnnaacgnnn uaannnnncc
gccuggggag uacgnncgca 900agnnunaaac ucaaangaau ugacggggnc cngcacaagc
ngnggagnau guggnuuaau 960ucgangnnac gcgnanaacc uuaccnnnnn uugacaunnn
nnnnnnnnnn nnganannnn 1020nnnnnnnnnn nnnnnnnnnn nnnacaggug nugcauggnu
gucgucagcu cgugnnguga 1080gnuguugggu uaagucccgn aacgagcgca acccnnnnnn
nnnguuncna ncnnnnnnnn 1140ngngnacucn nnnnnnacug ccnnngnnaa nnnggaggaa
ggnggggang acgucaanuc 1200nucaugnccc uuangnnnng ggcuncacac nuncuacaau
ggnnnnnaca nngngnngcn 1260annnngnnan nnnnagcnaa ncnnnnaaan nnnnucnnag
uncggaungn nnncugcaac 1320ucgnnnncnu gaagnnggan ucgcuaguaa ucgnnnauca
gnangnnncg gugaauacgu 1380ucncgggncu uguacacacc gcccgucann ncangnnagn
nnnnnnnncc nnaagnnnnn 1440nnnnnnncnn nnnngnnnnn nnnnncnang gnnnnnnnnn
nganugggnn naagucguaa 1500caagguancc nuannngaan nugnggnugg aucaccuccu
un 154242904RNAArtificial Sequencemisc_feature23S
rRNA consensus sequence 4nnnnaagnnn nnaagngnnn nngguggaug ccunggcnnn
nnnagncgan gaaggangnn 60nnnnncnncn nnanncnnng gnnagnngnn nnnnnncnnn
nnanccnnng nunuccgaau 120ggggnaaccc nnnnnnnnnn nnnnnnnnan nnnnnnnnnn
nnnnnnnnnn nnnnnnngnn 180nacnnnnnga anugaaacau cunaguannn nnaggaanag
aaannaannn ngauuncnnn 240nguagnggcg agcgaannng nannagncnn nnnnnnnnnn
nnnnnnnnnn nnnannngaa 300nnnnnuggna agnnnnnnnn nannngguna nannccngua
nnnnaaannn nnnnnnnnnn 360nnnnnnnnnn aguannncnn nncncgngnn annnngunng
aannngnnnn gaccannnnn 420naagncuaaa uacunnnnnn ngaccnauag ngnannagua
cngugangga aaggngaaaa 480gnacccnnnn nangggagug aaanagnncc ugaaaccnnn
nncnuanaan nngunnnagn 540nnnnnnnnnn nnnuganngc gunccuuuug nannaugnnn
cngnganuun nnnunnnnng 600cnagnuuaan nnnnnnnngn agncgnagng aaancgagun
nnaanngngc gnnnagunnn 660nngnnnnaga cncgaancnn ngugancuan nnaugnncag
gnugaagnnn nnguaanann 720nnnuggaggn ccgaacnnnn nnnnguugaa aannnnnngg
augannugug nnungnggng 780aaanncnaan cnaacnnngn nauagcuggu ucucnncgaa
annnnuuuag gnnnngcnun 840nnnnnnnnnn nnnnggnggu agagcacugn nnnnnnnnng
gnnnnnnnnn nnnnuacnna 900nnnnnnnnaa acuncgaaun ccnnnnnnnn nnnnnnnngn
agnnanncnn ngngngnuaa 960nnuncnnngu nnanagggna acancccaga ncnncnnnua
aggncccnaa nnnnnnnnua 1020aguggnaaan gangugnnnn nncnnanaca nnnaggangu
uggcuuagaa gcagccancn 1080uunaaagann gcguaanagc ucacunnucn agnnnnnnng
cgcngannau nuancgggnc 1140uaannnnnnn nccgaannnn nngnnnnnnn nnnnnnnnnn
nnnnngguag nngagcgunn 1200nnnnnnnnnn ngaagnnnnn nngnnannnn nnnuggannn
nnnnnnagug ngnaugnngn 1260naunaguanc gannnnnnnn gugananncn nnnncnccgn
annncnaagg nuuccnnnnn 1320nangnunnuc nnnnnngggu nagucgnnnc cuaagnngag
ncnganangn nuagnngaug 1380gnnannnggu nnauauuccn nnacnnnnnn nnnnnnnnnn
nnnnngacgn nnnnngnnnn 1440nnnnnnnnnn nnnnggnnnn nnnnnnnnnn nnnnnnnnnn
nnnnnnnnnn nnnnnnnnnn 1500nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn
nnnnnnnnnn nnnnnnnnnn 1560nnnncnngaa aannnnnnnn nnnnnnnnnn nnnnnnnnnc
guaccnnaaa ccgacacagg 1620ungnnnngnn gagnanncnn aggngnnngn nnnaannnnn
nnnaaggaac unngcaaanu 1680nnnnccguan cuucggnana aggnnnncnn nnnnnnnnnn
nnnnnnnnnn nnnnnnnnnn 1740nnnnnnnnng nnnnannnan nngnnnnnnn cnacuguuua
nnaaaaacac agnncnnugc 1800naanncgnaa gnnganguau anggnnugac nccugcccng
ugcnngaagg uuaanngnnn 1860nnnnnngnnn nngnnnnnnn nnnnannnaa gcccnnguna
acggcggnng uaacuauaac 1920nnuccuaagg uagcgaaauu ccuugucggg uaaguuccga
ccngcacgaa nggngnaang 1980annnnnnnnc ugucucnnnn nnnnncncng ngaanuunna
nunnnnguna agaugcnnnn 2040uncncgcnnn nngacggaaa gaccccnngn ancuuuacun
nannnunnna nugnnnnnnn 2100nnnnnnnnug unnagnauag gunggagncn nngannnnnn
nncgnnagnn nnnnnggagn 2160cnnnnnugnn auacnacncu nnnnnnnnnn nnnnucuaac
nnnnnnnnnn nancnnnnnn 2220nnngacanug nnngnngggn aguuunacug gggcggunnc
cuccnaaann guaacggagg 2280ngnncnaagg unnncunann nnggnnggnn aucnnnnnnn
nagunnaann gnanaagnnn 2340gcnunacugn nagnnnnacn nnncgagcag nnncgaaagn
nggnnnuagu gauccggngg 2400unnnnnnugg aagngccnuc gcucaacgga uaaaagnuac
ncnggggaua acaggcunau 2460nnnncccaag aguncanauc gacggnnnng uuuggcaccu
cgaugucggc ucnucncauc 2520cuggggcugn agnngguccc aagggunngg cuguucgccn
nuuaaagngg nacgngagcu 2580ggguunanaa cgucgugaga caguungguc ccuaucngnn
gngngngnnn gannnuugan 2640nngnnnugnn cnuaguacga gaggaccggn nngnacnnan
cncuggugnn ncnguugunn 2700ngccannngc anngcngnnu agcuannunn ggnnnngaua
anngcugaan gcaucuaagn 2760nngaancnnn cnnnnagann agnnnucncn nnnnnnnnnn
nnnnnnnnna gnnncnnnnn 2820agannannnn gungauaggn nngnnnugna agnnnngnna
nnnnunnagn nnacnnnuac 2880uaaunnnncn nnnnncuunn nnnn
2904513DNAArtificial Sequencemisc_featurePrimer
5cgtggtgacc ctt
13614DNAArtificial Sequencemisc_featurePrimer 6cgtcgtcacc gcta
14713DNAArtificial
Sequencemisc_featurePrimer 7cgtggtaccc ctt
13830DNAArtificial SequencePCR Primer 8gtgagatgtt
gggttaagtc ccgtaacgag
30921DNAArtificial SequencePCR Primer 9gacgtcatcc ccaccttcct c
211025DNAArtificial SequencePCR Primer
10atgttgggtt aagtcccgca acgag
251123DNAArtificial SequencePCR Primer 11ttgacgtcat ccccaccttc ctc
231222DNAArtificial SequencePCR
Primer 12ttaagtcccg caacgatcgc aa
221322DNAArtificial SequencePCR Primer 13tgacgtcatc cccaccttcc tc
221420DNAArtificial SequencePCR
Primer 14gctacacacg tgctacaatg
201521DNAArtificial SequencePCR Primer 15cgagttgcag actgcgatcc g
211622DNAArtificial SequencePCR
Primer 16aagtcggaat cgctagtaat cg
221719DNAArtificial SequencePCR Primer 17gacgggcggt gtgtacaag
191825DNAArtificial SequencePCR
Primer 18tgaacgctgg tggcatgctt aacac
251922DNAArtificial SequencePCR Primer 19tacgcattac tcacccgtcc gc
222026DNAArtificial SequencePCR
Primer 20gtggcatgcc taatacatgc aagtcg
262120DNAArtificial SequencePCR Primer 21ttactcaccc gtccgccgct
202220DNAArtificial SequencePCR
Primer 22taacacatgc aagtcgaacg
202317DNAArtificial SequencePCR Primer 23ttactcaccc gtccgcc
172418DNAArtificial SequencePCR
Primer 24gtgtagcggt gaaatgcg
182522DNAArtificial SequencePCR Primer 25gtatctaatc ctgtttgctc cc
222620DNAArtificial SequencePCR
Primer 26agaacaccga tggcgaaggc
202721DNAArtificial SequencePCR Primer 27cgtggactac cagggtatct a
212822DNAArtificial SequencePCR
Primer 28ggattagaga ccctggtagt cc
222918DNAArtificial SequencePCR Primer 29ggccgtactc cccaggcg
183026DNAArtificial SequencePCR
Primer 30ggattagata ccctggtagt ccacgc
263118DNAArtificial SequencePCR Primer 31ggccgtactc cccaggcg
183222DNAArtificial SequencePCR
Primer 32tagataccct ggtagtccac gc
223315DNAArtificial SequencePCR Primer 33cgtactcccc aggcg
153422DNAArtificial SequencePCR
Primer 34ttcgatgcaa cgcgaagaac ct
223520DNAArtificial SequencePCR Primer 35acgagctgac gacagccatg
203617DNAArtificial SequencePCR
Primer 36acgcgaagaa ccttacc
173718DNAArtificial SequencePCR Primer 37acgacacgag ctgacgac
183818DNAArtificial SequencePCR
Primer 38ctgacacctg cccggtgc
183919DNAArtificial SequencePCR Primer 39gaccgttata gttacggcc
194025DNAArtificial SequencePCR
Primer 40tctgtcccta gtacgagagg accgg
254118DNAArtificial SequencePCR Primer 41tgcttagatg ctttcagc
184224DNAArtificial SequencePCR
Primer 42ctgtccctag tacgagagga ccgg
244324DNAArtificial SequencePCR Primer 43gtttcatgct tagatgcttt cagc
244426DNAArtificial SequencePCR
Primer 44ggggagtgaa agagatcctg aaaccg
264521DNAArtificial SequencePCR Primer 45acaaaaggta cgccgtcacc c
214626DNAArtificial SequencePCR
Primer 46ggggagtgaa agagatcctg aaaccg
264721DNAArtificial SequencePCR Primer 47acaaaaggca cgccatcacc c
214822DNAArtificial SequencePCR
Primer 48cgagagggaa acaacccaga cc
224919DNAArtificial SequencePCR Primer 49tggctgcttc taagccaac
195029DNAArtificial SequencePCR
Primer 50tgctcgtggt gcacaagtaa cggatatta
295129DNAArtificial SequencePCR Primer 51tgctgctttc gcatggttaa
ttgcttcaa 295228DNAArtificial
SequencePCR Primer 52caaaacttat taggtaagcg tgttgact
285330DNAArtificial SequencePCR Primer 53tcaagcgcca
tttcttttgg taaaccacat
305428DNAArtificial SequencePCR Primer 54caaaacttat taggtaagcg tgttgact
285530DNAArtificial SequencePCR
Primer 55tcaagcgcca tctctttcgg taatccacat
305627DNAArtificial SequencePCR Primer 56taagaagccg gaaaccatca
actaccg 275720DNAArtificial
SequencePCR Primer 57ggcgcttgta cttaccgcac
205819DNAArtificial SequencePCR Primer 58tgattctggt
gcccgtggt
195922DNAArtificial SequencePCR Primer 59ttggccatca ggccacgcat ac
226019DNAArtificial SequencePCR
Primer 60tgattccggt gcccgtggt
196122DNAArtificial SequencePCR Primer 61ttggccatca gaccacgcat ac
226224DNAArtificial SequencePCR
Primer 62ctggcaggta tgcgtggtct gatg
246325DNAArtificial SequencePCR Primer 63cgcaccgtgg gttgagatga agtac
256424DNAArtificial SequencePCR
Primer 64cttgctggta tgcgtggtct gatg
246525DNAArtificial SequencePCR Primer 65cgcaccatgc gtagagatga agtac
256626DNAArtificial SequencePCR
Primer 66cgtcgggtga ttaaccgtaa caaccg
266725DNAArtificial SequencePCR Primer 67gtttttcgtt gcgtacgatg atgtc
256826DNAArtificial SequencePCR
Primer 68cgtcgtgtaa ttaaccgtaa caaccg
266927DNAArtificial SequencePCR Primer 69acgtttttcg ttttgaacga
taatgct 277027DNAArtificial
SequencePCR Primer 70caaaggtaag caaggtcgtt tccgtca
277124DNAArtificial SequencePCR Primer 71cgaacggcct
gagtagtcaa cacg
247227DNAArtificial SequencePCR Primer 72caaaggtaag caaggacgtt tccgtca
277324DNAArtificial SequencePCR
Primer 73cgaacggcca gagtagtcaa cacg
247421DNAArtificial SequencePCR Primer 74tagactgccc aggacacgct g
217521DNAArtificial SequencePCR
Primer 75gccgtccatc tgagcagcac c
217621DNAArtificial SequencePCR Primer 76ttgactgccc aggtcacgct g
217721DNAArtificial SequencePCR
Primer 77gccgtccatt tgagcagcac c
217825DNAArtificial SequencePCR Primer 78aactaccgtc cgcagttcta cttcc
257924DNAArtificial SequencePCR
Primer 79gttgtcgcca ggcataacca tttc
248025DNAArtificial SequencePCR Primer 80aactaccgtc ctcagttcta cttcc
258124DNAArtificial SequencePCR
Primer 81gttgtcacca ggcattacca tttc
248228DNAArtificial SequencePCR Primer 82ccacagttct acttccgtac
tactgacg 288330DNAArtificial
SequencePCR Primer 83tccaggcatt accatttcta ctccttctgg
308430DNAArtificial SequencePCR Primer 84gacctacagt
aagaggttct gtaatgaacc
308524DNAArtificial SequencePCR Primer 85tccaagtgct ggtttacccc atgg
248623DNAArtificial SequencePCR
Primer 86catccacacg gtggtggtga agg
238722DNAArtificial SequencePCR Primer 87gtgctggttt accccatgga gt
228825DNAArtificial SequencePCR
Primer 88cgtgttgact attcggggcg ttcag
258930DNAArtificial SequencePCR Primer 89attcaagagc catttctttt
ggtaaaccac 309029DNAArtificial
SequencePCR Primer 90tcaacaacct cttggaggta aagctcagt
299130DNAArtificial SequencePCR Primer 91tttcttgaag
agtatgagct gctccgtaag
309223DNAArtificial SequencePCR Primer 92catccacacg gtggtggtga agg
239329DNAArtificial SequencePCR
Primer 93tgttttgtat ccaagtgctg gtttacccc
299420DNAArtificial SequencePCR Primer 94cgtggcggcg tggttatcga
209524DNAArtificial SequencePCR
Primer 95cggtacgaac tggatgtcgc cgtt
249622DNAArtificial SequencePCR Primer 96tatcgctcag gcgaactcca ac
229721DNAArtificial SequencePCR
Primer 97gctggattcg cctttgctac g
219830DNAArtificial SequencePCR Primer 98tgtaatgaac cctaatgacc
atccacacgg 309927DNAArtificial
SequencePCR Primer 99ccaagtgctg gtttacccca tggagta
2710030DNAArtificial SequencePCR Primer 100taatgaaccc
taatgaccat ccacacggtg
3010126DNAArtificial SequencePCR Primer 101tccaagtgct ggtttacccc atggag
2610229DNAArtificial SequencePCR
Primer 102cttggaggta agtctcattt tggtgggca
2910330DNAArtificial SequencePCR Primer 103cgtataagct gcaccataag
cttgtaatgc 3010418DNAArtificial
SequencePCR Primer 104cgacgcgctg cgcttcac
1810524DNAArtificial SequencePCR Primer 105gcgttccaca
gcttgttgca gaag
2410618DNAArtificial SequencePCR Primer 106gaccacctcg gcaaccgt
1810718DNAArtificial SequencePCR
Primer 107ttcgctctcg gcctggcc
1810827DNAArtificial SequencePCR Primer 108gcactatgca cacgtagatt
gtcctgg 2710926DNAArtificial
SequencePCR Primer 109tatagcacca tccatctgag cggcac
2611022DNAArtificial SequencePCR Primer 110cggcgtactt
caacgacagc ca
2211120DNAArtificial SequencePCR Primer 111cgcggtcggc tcgttgatga
2011224DNAArtificial SequencePCR
Primer 112cttctgcaac aagctgtgga acgc
2411323DNAArtificial SequencePCR Primer 113tcgcagttca tcagcacgaa
gcg 2311418DNAArtificial
SequencePCR Primer 114aagacgacct gcacgggc
1811519DNAArtificial SequencePCR Primer 115gcgctccacg
tcttcacgc
1911622DNAArtificial SequencePCR Primer 116ctgttcttag tacgagagga cc
2211721DNAArtificial SequencePCR
Primer 117ttcgtgctta gatgctttca g
2111817DNAArtificial SequencePCR Primer 118acgcgaagaa ccttacc
1711918DNAArtificial
SequencePCR Primer 119acgacacgag ctgacgac
1812014DNAArtificial SequencePCR Primer 120cgaagaacct
tacc
1412112DNAArtificial SequencePCR Primer 121acacgagctg ac
1212214DNAArtificial SequencePCR
Primer 122cgaagaacct tacc
1412312DNAArtificial SequencePCR Primer 123acacgagctg ac
1212419DNAArtificial
SequencePCR Primer 124cctgataagg gtgaggtcg
1912520DNAArtificial SequencePCR Primer 125acgtccttca
tcgcctctga
2012622DNAArtificial SequencePCR Primer 126gttgtgaggt taagcgacta ag
2212721DNAArtificial SequencePCR
Primer 127ctatcggtca gtcaggagta t
2112822DNAArtificial SequencePCR Primer 128gttgtgaggt taagcgacta ag
2212920DNAArtificial
SequencePCR Primer 129ttgcatcggg ttggtaagtc
2013021DNAArtificial SequencePCR Primer 130atactcctga
ctgaccgata g
2113119DNAArtificial SequencePCR Primer 131aacatagcct tctccgtcc
1913220DNAArtificial SequencePCR
Primer 132gacttaccaa cccgatgcaa
2013324DNAArtificial SequencePCR Primer 133taccttagga ccgttatagt
tacg 2413419DNAArtificial
SequencePCR Primer 134ggacggagaa ggctatgtt
1913520DNAArtificial SequencePCR Primer 135ccaaacaccg
ccgtcgatat
2013624DNAArtificial SequencePCR Primer 136cgtaactata acggtcctaa ggta
2413720DNAArtificial SequencePCR
Primer 137gcttacacac ccggcctatc
2013820DNAArtificial SequencePCR Primer 138atatcgacgg cggtgtttgg
2013919DNAArtificial
SequencePCR Primer 139gcgtgacagg caggtattc
1914022DNAArtificial SequencePCR Primer 140agtctcaaga
gtgaacacgt aa
2214118DNAArtificial SequencePCR Primer 141gctgctggca cggagtta
1814220DNAArtificial SequencePCR
Primer 142gacacggtcc agactcctac
2014318DNAArtificial SequencePCR Primer 143ccatgcagca cctgtctc
1814420DNAArtificial
SequencePCR Primer 144gatctggagg aataccggtg
2014520DNAArtificial SequencePCR Primer 145acggttacct
tgttacgact
2014620DNAArtificial SequencePCR Primer 146gagagcaagc ggacctcata
2014717DNAArtificial SequencePCR
Primer 147cctcctgcgt gcaaagc
1714819DNAArtificial SequencePCR Primer 148tggaagatct gggtcaggc
1914922DNAArtificial
SequencePCR Primer 149caatctgctg acggatctga gc
2215022DNAArtificial SequencePCR Primer 150gtcgtgaaaa
cgagctggaa ga
2215118DNAArtificial SequencePCR Primer 151catgatggtc acaaccgg
1815222DNAArtificial SequencePCR
Primer 152tggcgaacct ggtgaacgaa gc
2215325DNAArtificial SequencePCR Primer 153ctttcgcttt ctcgaactca
accat 2515426DNAArtificial
SequencePCR Primer 154cgtcagggta aattccgtga agttaa
2615521DNAArtificial SequencePCR Primer 155aacttcgcct
tcggtcatgt t
2115624DNAArtificial SequencePCR Primer 156ggtgaaagaa gttgcctcta aagc
2415723DNAArtificial SequencePCR
Primer 157ttcaggtcca tcgggttcat gcc
2315820DNAArtificial SequencePCR Primer 158cgtggcggcg tggttatcga
2015920DNAArtificial
SequencePCR Primer 159acgaactgga tgtcgccgtt
2016020DNAArtificial SequencePCR Primer 160cggaattact
gggcgtaaag
2016118DNAArtificial SequencePCR Primer 161cgcatttcac cgctacac
1816222DNAArtificial SequencePCR
Primer 162acccagtgct gctgaaccgt gc
2216321DNAArtificial SequencePCR Primer 163gttcaaatgc ctggataccc a
2116422DNAArtificial
SequencePCR Primer 164gggagcaaac aggattagat ac
2216515DNAArtificial SequencePCR Primer 165cgtactcccc
aggcg
1516621DNAArtificial SequencePCR Primer 166tggcccgaaa gaagctgagc g
2116721DNAArtificial SequencePCR
Primer 167acgcgggcat gcagagatgc c
2116819DNAArtificial SequencePCR Primer 168atgttgggtt aagtcccgc
1916920DNAArtificial
SequencePCR Primer 169tgacgtcatc cccaccttcc
2017019DNAArtificial SequencePCR Primer 170cttgtacaca
ccgcccgtc
1917117DNAArtificial SequencePCR Primer 171aaggaggtga tccagcc
1717221DNAArtificial SequencePCR
Primer 172cggattggag tctgcaactc g
2117319DNAArtificial SequencePCR Primer 173gacgggcggt gtgtacaag
1917415DNAArtificial
SequencePCR Primer 174ggtggatgcc ttggc
1517516DNAArtificial SequencePCR Primer 175gggtttcccc
attcgg
1617621DNAArtificial SequencePCR Primer 176gggaactgaa acatctaagt a
2117715DNAArtificial SequencePCR
Primer 177ttcgctcgcc gctac
1517819DNAArtificial SequencePCR Primer 178taccccaaac cgacacagg
1917918DNAArtificial
SequencePCR Primer 179ccttctcccg aagttacg
1818019DNAArtificial SequencePCR Primer 180ccgtaacttc
gggagaagg
1918115DNAArtificial SequencePCR Primer 181caccgggcag gcgtc
1518216DNAArtificial SequencePCR
Primer 182gacgcctgcc cggtgc
1618321DNAArtificial SequencePCR Primer 183ccgacaagga atttcgctac c
2118423DNAArtificial
SequencePCR Primer 184aaggtactcc ggggataaca ggc
2318522DNAArtificial SequencePCR Primer 185agccgacatc
gaggtgccaa ac
2218618DNAArtificial SequencePCR Primer 186gacagttcgg tccctatc
1818717DNAArtificial SequencePCR
Primer 187ccggtcctct cgtacta
1718817DNAArtificial SequencePCR Primer 188tagtacgaga ggaccgg
1718922DNAArtificial
SequencePCR Primer 189ttagatgctt tcagcactta tc
2219024DNAArtificial SequencePCR Primer 190aaactagata
acagtagaca tcac
2419118DNAArtificial SequencePCR Primer 191gtgcgccctt tctaactt
1819220DNAArtificial SequencePCR
Primer 192agagtttgat catggctcag
2019317DNAArtificial SequencePCR Primer 193actgctgcct cccgtag
1719419DNAArtificial
SequencePCR Primer 194cactggaact gagacacgg
1919520DNAArtificial SequencePCR Primer 195ctttacgccc
agtaattccg
2019619DNAArtificial SequencePCR Primer 196ccagcagccg cggtaatac
1919722DNAArtificial SequencePCR
Primer 197gtatctaatc ctgtttgctc cc
2219818DNAArtificial SequencePCR Primer 198gtgtagcggt gaaatgcg
1819919DNAArtificial
SequencePCR Primer 199ggtaaggttc ttcgcgttg
1920018DNAArtificial SequencePCR Primer 200aagcggtgga
gcatgtgg
1820121DNAArtificial SequencePCR Primer 201attgtagcac gtgtgtagcc c
2120219DNAArtificial SequencePCR
Primer 202caagtcatca tggccctta
1920317DNAArtificial SequencePCR Primer 203aaggaggtga tccagcc
1720420DNAArtificial
SequencePCR Primer 204agagtttgat catggctcag
2020517DNAArtificial SequencePCR Primer 205aaggaggtga
tccagcc
1720619DNAArtificial SequencePCR Primer 206acctgcccag tgctggaag
1920718DNAArtificial SequencePCR
Primer 207tcgctacctt aggaccgt
1820821DNAArtificial SequencePCR Primer 208gccttgtaca cacctcccgt c
2120920DNAArtificial
SequencePCR Primer 209cacggctacc ttgttacgac
2021022DNAArtificial SequencePCR Primer 210ttgtacacac
cgcccgtcat ac
2221120DNAArtificial SequencePCR Primer 211ccttgttacg acttcacccc
2021221DNAArtificial SequencePCR
Primer 212tacggtgaat acgttcccgg g
2121322DNAArtificial SequencePCR Primer 213accttgttac gacttcaccc ca
2221419DNAArtificial
SequencePCR Primer 214accacgccgt aaacgatga
1921521DNAArtificial SequencePCR Primer 215cccccgtcaa
ttcctttgag t
2121622DNAArtificial SequencePCR Primer 216gataccctgg tagtccacac cg
2221719DNAArtificial SequencePCR
Primer 217gccttgcgac cgtactccc
1921822DNAArtificial SequencePCR Primer 218tagataccct ggtagtccac gc
2221918DNAArtificial
SequencePCR Primer 219gcgaccgtac tccccagg
1822018DNAArtificial SequencePCR Primer 220tagtcccgca
acgagcgc
1822122DNAArtificial SequencePCR Primer 221gacgtcatcc ccaccttcct cc
2222222DNAArtificial SequencePCR
Primer 222tagaacgtcg cgagacagtt cg
2222320DNAArtificial SequencePCR Primer 223agtccatccc ggtcctctcg
2022417DNAArtificial
SequencePCR Primer 224gaggaaagtc cgggctc
1722518DNAArtificial SequencePCR Primer 225ataagccggg
ttctgtcg
1822619DNAArtificial SequencePCR Primer 226gaggaaagtc catgctcgc
1922722DNAArtificial SequencePCR
Primer 227gtaagccatg ttttgttcca tc
2222817DNAArtificial SequencePCR Primer 228gaggaaagtc cgggctc
1722918DNAArtificial
SequencePCR Primer 229ataagccggg ttctgtcg
1823035DNAArtificial SequencePCR Primer 230gcgggatcct
ctagaggtgt taaatagcct ggcag
3523135DNAArtificial SequencePCR Primer 231gcgggatcct ctagaagacc
tcctgcgtgc aaagc 3523219DNAArtificial
SequencePCR Primer 232gaggaaagtc catgctcac
1923322DNAArtificial SequencePCR Primer 233ataagccatg
ttctgttcca tc
2223419DNAArtificial SequencePCR Primer 234atgttgggtt aagtcccgc
1923517DNAArtificial SequencePCR
Primer 235aaggaggtga tccagcc
1723620DNAArtificial SequencePCR Primer 236cggaattact gggcgtaaag
2023722DNAArtificial
SequencePCR Primer 237gtatctaatc ctgtttgctc cc
2223819DNAArtificial SequencePCR Primer 238atgttgggtt
aagtcccgc
1923920DNAArtificial SequencePCR Primer 239tgacgtcatg cccaccttcc
2024019DNAArtificial SequencePCR
Primer 240atgttgggtt aagtcccgc
1924120DNAArtificial SequencePCR Primer 241tgacgtcatg gccaccttcc
2024234DNAArtificial
SequencePCR Primer 242gcgggatcct ctagacctga taagggtgag gtcg
3424334DNAArtificial SequencePCR Primer 243gcgggatcct
ctagagcgtg acaggcaggt attc
3424417DNAArtificial SequencePCR Primer 244acgcgaagaa ccttacc
1724519DNAArtificial SequencePCR
Primer 245gacgggcggt gtgtacaag
1924618DNAArtificial SequencePCR Primer 246gtgtagcggt gaaatgcg
1824721DNAArtificial
SequencePCR Primer 247cgagttgcag actgcgatcc g
2124820DNAArtificial SequencePCR Primer 248taacacatgc
aagtcgaacg
2024915DNAArtificial SequencePCR Primer 249cgtactcccc aggcg
1525020DNAArtificial SequencePCR
Primer 250taacacatgc aagtcgaacg
2025118DNAArtificial SequencePCR Primer 251acgacacgag ctgacgac
1825222DNAArtificial
SequencePCR Primer 252acaacgaagt acaatacaag ac
2225322DNAArtificial SequencePCR Primer 253cttctacatt
tttagccatc ac
2225422DNAArtificial SequencePCR Primer 254ttaagtcccg caacgagcgc aa
2225522DNAArtificial SequencePCR
Primer 255tgacgtcatc cccaccttcc tc
2225628DNAArtificial SequencePCR Primer 256tgagtgatga aggccttagg
gttgtaaa 2825721DNAArtificial
SequencePCR Primer 257cggctgctgg cacgaagtta g
2125823DNAArtificial SequencePCR Primer 258atggacaagg
ttggcaagga agg
2325920DNAArtificial SequencePCR Primer 259tagccgcggt cgaattgcat
2026026DNAArtificial SequencePCR
Primer 260aaggaaggcg tgatcaccgt tgaaga
2626123DNAArtificial SequencePCR Primer 261ccgcggtcga attgcatgcc
ttc 2326216DNAArtificial
SequencePCR Primer 262acgcgctgcg cttcac
1626320DNAArtificial SequencePCR Primer 263ttgcagaagt
tgcggtagcc
2026418DNAArtificial SequencePCR Primer 264tcgaccacct gggcaacc
1826519DNAArtificial SequencePCR
Primer 265atcaggtcgt gcggcatca
1926617DNAArtificial SequencePCR Primer 266cacggtgccg gcgtact
1726720DNAArtificial
SequencePCR Primer 267gcggtcggct cgttgatgat
2026825DNAArtificial SequencePCR Primer 268ttggaggtaa
gtctcatttt ggtgg
2526925DNAArtificial SequencePCR Primer 269aagctgcacc ataagcttgt aatgc
2527020DNAArtificial SequencePCR
Primer 270cagcgtttcg gcgaaatgga
2027124DNAArtificial SequencePCR Primer 271cgacttgacg gttaacattt
cctg 2427223DNAArtificial
SequencePCR Primer 272gggcagcgtt tcggcgaaat gga
2327327DNAArtificial SequencePCR Primer 273gtccgacttg
acggtcaaca tttcctg
2727429DNAArtificial SequencePCR Primer 274caggagtcgt tcaactcgat
ctacatgat 2927519DNAArtificial
SequencePCR Primer 275acgccatcag gccacgcat
1927618DNAArtificial SequencePCR Primer 276gcacaacctg
cggctgcg
1827718DNAArtificial SequencePCR Primer 277acggcacgag gtagtcgc
1827820DNAArtificial SequencePCR
Primer 278cgccgacttc gacggtgacc
2027919DNAArtificial SequencePCR Primer 279gagcatcagc gtgcgtgct
1928023DNAArtificial
SequencePCR Primer 280ccacacgccg ttcttcaaca act
2328125DNAArtificial SequencePCR Primer 281ggcatcacca
tttccttgtc cttcg
2528227DNAArtificial SequencePCR Primer 282gagagtttga tcctggctca gaacgaa
2728322DNAArtificial SequencePCR
Primer 283tgttactcac ccgtctgcca ct
2228419DNAArtificial SequencePCR Primer 284accgagcaag gagaccagc
1928523DNAArtificial
SequencePCR Primer 285tataacgcac atcgtcaggg tga
2328620DNAArtificial SequencePCR Primer 286agacccaatt
acattggctt
2028720DNAArtificial SequencePCR Primer 287ccagtgctgt tgtagtacat
2028820DNAArtificial SequencePCR
Primer 288atgtactaca acagtactgg
2028920DNAArtificial SequencePCR Primer 289caagtcaacc acagcattca
2029020DNAArtificial
SequencePCR Primer 290gggcttatgt actacaacag
2029121DNAArtificial SequencePCR Primer 291tctgtcttgc
aagtcaacca c
2129221DNAArtificial SequencePCR Primer 292ggaatttttt gatggtagag a
2129319DNAArtificial SequencePCR
Primer 293taaagcacaa tttcaggcg
1929420DNAArtificial SequencePCR Primer 294tagatctggc tttctttgac
2029521DNAArtificial
SequencePCR Primer 295atatgagtat ctggagtctg c
2129621DNAArtificial SequencePCR Primer 296ggaaagacat
tactgcagac a
2129720DNAArtificial SequencePCR Primer 297ccaacttgag gctctggctg
2029819DNAArtificial SequencePCR
Primer 298acagacactt accagggtg
1929920DNAArtificial SequencePCR Primer 299actgtggtgt catctttgtc
2030022DNAArtificial
SequencePCR Primer 300tcactaaaga caaaggtctt cc
2230121DNAArtificial SequencePCR Primer 301ggcttcgccg
tctgtaattt c
2130221DNAArtificial SequencePCR Primer 302cggatccaag ctaatctttg g
2130324DNAArtificial SequencePCR
Primer 303ggtatgtact cataggtgtt ggtg
2430420DNAArtificial SequencePCR Primer 304agacccaatt acattggctt
2030520DNAArtificial
SequencePCR Primer 305ccagtgctgt tgtagtacat
2030620DNAArtificial SequencePCR Primer 306atgtactaca
acagtactgg
2030720DNAArtificial SequencePCR Primer 307caagtcaacc acagcattca
2030820DNAArtificial SequencePCR
Primer 308gggcttatgt actacaacag
2030921DNAArtificial SequencePCR Primer 309tctgtcttgc aagtcaacca c
2131021DNAArtificial
SequencePCR Primer 310ggaatttttt gatggtagag a
2131119DNAArtificial SequencePCR Primer 311taaagcacaa
tttcaggcg
1931220DNAArtificial SequencePCR Primer 312tagatctggc tttctttgac
2031321DNAArtificial SequencePCR
Primer 313atatgagtat ctggagtctg c
2131421DNAArtificial SequencePCR Primer 314cggatccaag ctaatctttg g
2131524DNAArtificial
SequencePCR Primer 315ggtatgtact cataggtgtt ggtg
2431623DNAArtificial SequencePCR Primer 316aacagaccca
attacattgg ctt
2331722DNAArtificial SequencePCR Primer 317gaggcacttg tatgtggaaa gg
2231823DNAArtificial SequencePCR
Primer 318atgcctaaca gacccaatta cat
2331922DNAArtificial SequencePCR Primer 319ttcatgtagt cgtaggtgtt gg
2232024DNAArtificial
SequencePCR Primer 320cgcgcctaat acatctcagt ggat
2432123DNAArtificial SequencePCR Primer 321aagccaatgt
aattgggtct gtt
2332222DNAArtificial SequencePCR Primer 322ctactctggc actgcctaca ac
2232323DNAArtificial SequencePCR
Primer 323atgtaattgg gtctgttagg cat
2332424DNAArtificial SequencePCR Primer 324caatccgttc tggttccgga
tgaa 2432524DNAArtificial
SequencePCR Primer 325cttgccggtc gttcaaagag gtag
2432618DNAArtificial SequencePCR Primer 326agtccgggtc
tggtgcag
1832717DNAArtificial SequencePCR Primer 327cggtcggtgg tcacatc
1732818DNAArtificial SequencePCR
Primer 328atggccaccc catcgatg
1832919DNAArtificial SequencePCR Primer 329ctgtccggcg atgtgcatg
1933023DNAArtificial
SequencePCR Primer 330ggtcgttatg tgcctttcca cat
2333125DNAArtificial SequencePCR Primer 331tcctttctga
agttccactc atagg
2533222DNAArtificial SequencePCR Primer 332acaacattgg ctaccagggc tt
2233325DNAArtificial SequencePCR
Primer 333cctgcctgct cataggctgg aagtt
2533422DNAArtificial SequencePCR Primer 334ggattagaga ccctggtagt cc
2233518DNAArtificial
SequencePCR Primer 335ggccgtactc cccaggcg
1833622DNAArtificial SequencePCR Primer 336ttcgatgcaa
cgcgaagaac ct
2233720DNAArtificial SequencePCR Primer 337acgagctgac gacagccatg
2033825DNAArtificial SequencePCR
Primer 338tctgtcccta gtacgagagg accgg
2533918DNAArtificial SequencePCR Primer 339tgcttagatg ctttcagc
1834024DNAArtificial
SequencePCR Primer 340ctggcaggta tgcgtggtct gatg
2434125DNAArtificial SequencePCR Primer 341cgcaccgtgg
gttgagatga agtac
2534230DNAArtificial SequencePCR Primer 342ggggattcag ccatcaaagc
agctattgac 3034326DNAArtificial
SequencePCR Primer 343ccaacctttt ccacaacaga atcagc
2634430DNAArtificial SequencePCR Primer 344ccttacttcg
aactatgaat cttttggaag
3034529DNAArtificial SequencePCR Primer 345cccatttttt cacgcatgct
gaaaatatc 2934623DNAArtificial
SequencePCR Primer 346cgcaaaaaaa tccagctatt agc
2334729DNAArtificial SequencePCR Primer 347aaactatttt
tttagctata ctcgaacac
2934829DNAArtificial SequencePCR Primer 348atgattacaa ttcaagaagg
tcgtcacgc 2934927DNAArtificial
SequencePCR Primer 349ttggacctgt aatcagctga atactgg
2735029DNAArtificial SequencePCR Primer 350gatgactttt
tagctaatgg tcaggcagc
2935129DNAArtificial SequencePCR Primer 351aatcgacgac catcttggaa
agatttctc 2935225DNAArtificial
SequencePCR Primer 352gcttcaggaa tcaatgatgg agcag
2535325DNAArtificial SequencePCR Primer 353gggtctacac
ctgcacttgc ataac
2535418DNAArtificial SequencePCR Primer 354gtactgaatc cgcctaag
1835523DNAArtificial SequencePCR
Primer 355gtgaataaag tatcgcccta ata
2335618DNAArtificial SequencePCR Primer 356gaagttgaac cgggatca
1835721DNAArtificial
SequencePCR Primer 357attatcggtc gttgttaatg t
2135823DNAArtificial SequencePCR Primer 358ctgtctgtag
ataaactagg att
2335918DNAArtificial SequencePCR Primer 359cgttcttctc tggaggat
1836015DNAArtificial SequencePCR
Primer 360cgatactacg gacgc
1536122DNAArtificial SequencePCR Primer 361ctttatgaat tactttacat at
2236218DNAArtificial
SequencePCR Primer 362ctcctccatc actaggaa
1836322DNAArtificial SequencePCR Primer 363ctataacatt
caaagcttat tg
2236423DNAArtificial SequencePCR Primer 364cgcgataata gatagtgcta aac
2336519DNAArtificial SequencePCR
Primer 365gcttccacca ggtcattaa
1936618DNAArtificial SequencePCR Primer 366gtactgaatc cgcctaag
1836723DNAArtificial
SequencePCR Primer 367gtgaataaag tatcgcccta ata
2336818DNAArtificial SequencePCR Primer 368gaagttgaac
cgggatca
1836921DNAArtificial SequencePCR Primer 369attatcggtc gttgttaatg t
2137023DNAArtificial SequencePCR
Primer 370ctgtctgtag ataaactagg att
2337118DNAArtificial SequencePCR Primer 371cgttcttctc tggaggat
1837215DNAArtificial
SequencePCR Primer 372cgatactacg gacgc
1537322DNAArtificial SequencePCR Primer 373ctttatgaat
tactttacat at
2237418DNAArtificial SequencePCR Primer 374ctcctccatc actaggaa
1837522DNAArtificial SequencePCR
Primer 375ctataacatt caaagcttat tg
2237623DNAArtificial SequencePCR Primer 376cgcgataata gatagtgcta
aac 2337719DNAArtificial
SequencePCR Primer 377gcttccacca ggtcattaa
19378375RNAArtificial SequenceBacterial Ribosomal RNA
378nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnng
60aggaaagucc ggncucnnna nncannnugn nngnuaannn cnnnnnnngn nannnnngac
120naguncnaca gagngnnnnc cgccnnnnnn nnnnnnnnnn nnnnnnnggn aagggugnaa
180nggnnnngua agagnncacc gnnnnnnnng nnannnnnnn nnncnnggna aacuccnnnn
240gnagcaagnn nnnnnnngnn nnnnnnnnnn nnngnncnnn nnnnnnnnnn nnannnngcu
300ngagnnnnnn ngnnannnnn nnccnagann naugnnnnnn cnnnacagaa nccggcnuan
360nnnnnnnnnn nnnnn
3753799DNAArtificial SequenceDouble stranded nucleic acid 379tacgtacgt
93809DNAArtificial SequenceDouble stranded nucleic acid 380atgcatgca
938165RNAArtificial SequenceSynthetic oligonucleotide 381gacccauggu
cgcucgcucc ucuccuacuu ggauaacugu gguaauucua gagcuaauac 60augcc
6538248RNAArtificial SequenceSynthetic oligonucleotide 382gccauaugga
gggggauaac uacuggaaac gguagcuaau accgcaua 48
User Contributions:
comments("1"); ?> comment_form("1"); ?>Inventors list |
Agents list |
Assignees list |
List by place |
Classification tree browser |
Top 100 Inventors |
Top 100 Agents |
Top 100 Assignees |
Usenet FAQ Index |
Documents |
Other FAQs |
User Contributions:
Comment about this patent or add new information about this topic:
 |  |
 |  |
 |  |
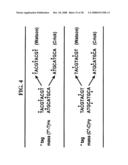 |  |
 | 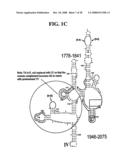 |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 | 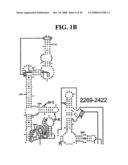 |
 |  |
 |  |
 |  |
 | 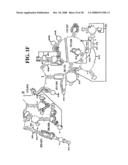 |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 | 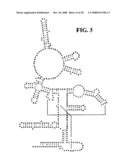 |
 |  |
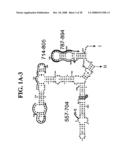 |  |
 |  |
 |  |
 | 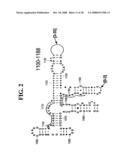 |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
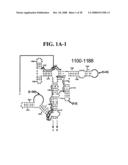 |  |
 |  |
 | 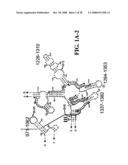 |
 |  |
 |  |
 |  |
 | 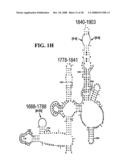 |
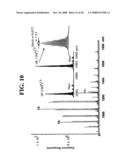 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 | 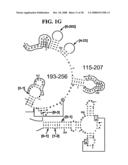 |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
 |
| New patent applications in this class: | |
| Date | Title |
|---|---|
| 2022-05-05 | Method for diagnosing human t-cell leukemia virus type 1 (htlv-1) associated diseases |
| 2022-05-05 | Systems and methods for assay processing |
| 2022-05-05 | Rapid pathology/cytology without wash |
| 2019-05-16 | Biofluidic triggering system and method |
| 2019-05-16 | Method and system for detection of disease agents in blood |
| New patent applications from these inventors: | |
| Date | Title |
|---|---|
| 2022-09-15 | 2'-methoxy substituted oligomeric compounds and compositions for use in gene modulations |
| 2021-12-09 | Chirally enriched oligomeric compounds |
| 2021-11-18 | Oligonucleotide mediated no-go decay |
| 2020-12-31 | Methods of modulating antisense activity |
| 2018-04-19 | 2'-methoxy substituted oligomeric compounds and compositions for use in gene modulations |
| Top Inventors for class "Chemistry: molecular biology and microbiology" | |
| Rank | Inventor's name |
|---|---|
| 1 | Marshall Medoff |
| 2 | Anthony P. Burgard |
| 3 | Mark J. Burk |
| 4 | Robin E. Osterhout |
| 5 | Rangarajan Sampath |